
Abstract
Many patterns observed in ecology, such as species richness, life history variation, habitat use, and distribution, have physiological underpinnings. For many ectothermic organisms, temperature relationships shape these patterns, but for terrestrial amphibians, water balance may supersede temperature as the most critical physiologically limiting factor. Many amphibian species have little resistance to water loss, which restricts them to moist microhabitats, and may significantly affect foraging, dispersal, and courtship. Using plaster models as surrogates for terrestrial plethodontid salamanders (Plethodon albagula), we measured water loss under ecologically relevant field conditions to estimate the duration of surface activity time across the landscape. Surface activity time was significantly affected by topography, solar exposure, canopy cover, maximum air temperature, and time since rain. Spatially, surface activity times were highest in ravine habitats and lowest on ridges. Surface activity time was a significant predictor of salamander abundance, as well as a predictor of successful recruitment; the probability of a juvenile salamander occupying an area with high surface activity time was two times greater than an area with limited predicted surface activity. Our results suggest that survival, recruitment, or both are demographic processes that are affected by water loss and the ability of salamanders to be surface-active. Results from our study extend our understanding of plethodontid salamander ecology, emphasize the limitations imposed by their unique physiology, and highlight the importance of water loss to spatial population dynamics. These findings are timely for understanding the effects that fluctuating temperature and moisture conditions predicted for future climates will have on plethodontid salamanders.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
An organism’s physiology is dynamically related to its environment. Physiology dictates the habitats that are occupied and behavior within these habitats, while the environment can affect physiological performance and subsequently, ecological performance (Huey 1991). In concert with the environment, physiology can affect an organism’s performance at the local scale (Brewster et al. 2013), life history at a regional scale (Kearney 2012), and dictate limits on distribution (Buckley et al. 2010; Gifford and Kozak 2012; Kearney and Porter 2009). Further, potential responses to habitat or climate change can be modeled mechanistically by incorporating physiology (Kearney et al. 2008; Keith et al. 2008; Sinervo et al. 2010). The role of physiology is especially evident in ectothermic organisms, with the preponderance of emphasis being placed on thermal aspects of behavior, physiology, and life history evolution (Angilletta 2009; Angilletta et al. 2004).
Although not independent of temperature and metabolic processes, water balance is another critical physiological characteristic that weighs heavily on the behavior, distribution, and ecology of terrestrial taxa. Many animals such as insects, birds, small mammals and reptiles often exhibit differential success in relation to microclimate, and actively select favorable microclimatic conditions (Adolph 1990; Getz 1968; Wachob 1996; Weiss et al. 1988). Less vagile animals such as terrestrial gastropods and arthropods are even more dependent upon suitable temperature and moisture microclimates (Cameron 1970; Cloudsley-Thompson 1962). Due to their extremely large surface area relative to mass, Cloudsley-Thompson (1962) went so far as to say that water conservation in terrestrial arthropods is the most critical physiological limitation to their existence. To minimize water loss, many terrestrial organisms inhabit cool, moist microclimates, spend all or most of their time under cover, and are primarily active at night when temperatures are cooler and relative humidity is higher (Boag 1985; Cloudsley-Thompson 1962).
Among terrestrial vertebrates, amphibians are especially susceptible to water loss (Tracy et al. 2010; Wells 2007). The skin of most amphibians provides little to no resistance to water loss (Spight 1968; Spotila and Berman 1976), even when the atmosphere is near saturation (Adolph 1932). All terrestrial amphibians must manage their hydric relationships, but it is particularly critical for woodland salamanders of the genus Plethodon. These salamanders are unique among terrestrial vertebrates in that they are lungless and respire cutaneously by diffusion (Whitford and Hutchison 1967). As a consequence, plethodontid skin must remain moist and permeable to facilitate gas exchange, but these requirements impose physiological and ecological limitations. Because of its permeability, the skin of plethodontid salamanders loses water at a rate that is nearly identical to a free water surface of equivalent surface area (Peterman et al. 2013; Spotila and Berman 1976). Uninhibited water loss impinges upon salamander activity, potentially limiting foraging, dispersal, and reproductive efforts. Terrestrial plethodontid salamanders spend the majority of their life underground or sheltered by cover objects such as rocks and logs on the ground surface (Petranka 1998). Surface activity and foraging of salamanders is greatest under moist conditions (Grover 1998; Keen 1979, 1984), and the duration of time spent foraging is directly tied to water balance (Feder and Londos 1984). To minimize water loss, salamanders are predominantly nocturnal, and are generally associated with cool, moist microhabitats (Heatwole 1962; Peterman and Semlitsch 2013; Spotila 1972).
Numerous proximate factors are related to salamander surface activity, such as time since last foraging, breeding season and reproductive condition, and local weather conditions. From a physiological perspective, four measurements are needed to predict the duration of surface activity that can be tolerated: salamander mass (used to calculate surface area; Whitford and Hutchison 1967), air temperature, relative humidity, and wind speed (Feder 1983). These factors can be used to predict that surface activity will be greatest for large salamanders when humidity is high, temperatures are cool, and there is no wind. Ecologically, this means that adults may have an advantage over juveniles in being able to sustain prolonged surface activity due to their lower surface area to volume ratio, and microclimate variation produced by landscape features such as topography may profoundly affect surface activity times by modulating temperature, wind, and humidity. Limited surface activity may limit foraging time, and consequently affect individual growth and reproduction. Dispersal may also be curtailed, reducing gene flow among local populations.
Despite the intuitive effects that hydric constraints impose on terrestrial plethodontid salamander activity time, habitat use, and population dynamics, direct tests of these processes have been limited. Within a controlled laboratory setting, Feder and Londos (1984) found that a stream salamander (Desmognathus ochrophaeus, Cope) would abandon foraging in dry air twice as quickly as in moist air (3.8 vs. 7.5 % loss of body mass, respectively). Grover (1998) experimentally demonstrated that surface activity of salamanders, especially juveniles, increased with increased soil moisture. Peterman and Semlitsch (2013) found that terrestrial salamander abundance was greatest in dense-canopy ravines with low solar exposure and high moisture, and found evidence of differential reproductive success related to these landscape features. Effects of water loss on population dynamics have indirectly been observed through variation in egg production. Grover and Wilbur (2002) found that salamanders in high moisture conditions produced more eggs, and both Milanovich et al. (2006) and Maiorana (1977) found annual fecundity to increase with precipitation. These findings suggest that wetter conditions may accommodate increased surface activity and foraging, increasing the energy available to be allocated to reproduction.
By incorporating physiology with spatial and temporal climate variation, mechanistic biophysical models are capable of accurately predicting the distribution (Kearney and Porter 2009), biotic interactions (Buckley and Roughgarden 2005; Gifford and Kozak 2012), and life history variation (Kearney 2012; Tracy et al. 2010) of species. To encompass spatial heterogeneity, most of these studies cover broad geographical or elevational ranges. However, environmental gradients can occur over significantly smaller spatial scales in topographically complex landscapes (Bennie et al. 2008; Chen et al. 1999). Further, fine-scale variation in microclimate can affect species occurrence, population dynamics, and resilience to changing climatic conditions, especially in species with low vagility (Antvogel and Bonn 2001; Peterman and Semlitsch 2013; Scherrer and Körner 2011; Weiss et al. 1988). Although the importance of fine-scale microclimatic variation is well understood (Huey 1991), most analyses of physiological processes have not been spatially explicit.
The objective of our study was to explicitly test, for the first time, the hypothesis that water balance is a limiting factor for terrestrial salamanders (Feder 1983). Specifically, we conducted field experiments to determine if spatial variation in water loss and surface activity time affects the distribution of salamanders as well as population dynamics across the landscape. We model physiological landscapes describing fine-scale spatial variation in water loss rates for a terrestrial plethodontid salamander, Plethodon albagula (western slimy salamander, Grobman 1944), and then convert these rates to potential surface activity times. In calculating rates of water loss and surface activity time, we seek to (1) determine the landscape and environmental factors influencing spatial variation in water loss on a topographically complex landscape, (2) determine whether salamander distribution on the landscape can be predicted by the physiological limitations imposed by water loss and activity time, and (3) assess the effects of surface activity time on spatial population dynamics. We hypothesized that rates of water loss would be dependent upon both topographical landscape features as well as climatic conditions. Specifically, we predicted that topographic complexity would result in heterogeneous water loss rates across the landscape and that ravine habitat with low solar exposure would have the lowest rates of water loss. Temporally, we predicted that abiotic factors such as time since rain, air temperature, and relative humidity would significantly affect daily and seasonal patterns of water loss. Because Peterman and Semlitsch (2013) found salamander abundance to be greatest in sheltered ravine habitats and lowest on ridges, we hypothesized that spatial patterns of water loss would corroborate these patterns, with ravines exhibiting low rates of water loss and ridges high rates of water loss. We also hypothesized that water loss, as an integrated measure of the landscape and climate, would significantly predict the spatial distribution of salamander abundance. Lastly, as a mechanism limiting population growth, we hypothesized that evidence of successful reproduction would be greatest in ravines with lower rates of water loss.
Materials and methods
Study species
Plethodon albagula (western slimy salamander) is a large plethodontid salamander of the P. glutinosus species complex that lives in forested habitats throughout the Ozark and Ouchita mountains of Missouri, Arkansas, eastern Oklahoma, and northeastern Texas, USA (Highton 1989). Within these forested habitats, salamander abundance is greatest in moist, forested ravines (Peterman and Semlitsch 2013). Surface activity varies seasonally, with peak activity occurring in spring and to a lesser extent during autumn (Milanovich et al. 2011); terrestrial plethodontid salamanders generally seek subterranean refuge during hot, dry summer conditions (Taub 1961). While detection of plethodontid salamanders during the day is generally associated with the availability of surface cover objects that can be searched, the actual distribution of salamanders across the landscape is independent of the amount of surface cover available (Peterman and Semlitsch 2013).
Plaster models
We assayed water loss across the landscape using cylindrical plaster of Paris models (hereafter “replicas”) as analogues for live salamanders. Plaster replicas were made following methods described by Peterman et al. (2013), and had surface areas equivalent to adult- and juvenile-sized salamanders that were 7.25 and 2.25 g, respectively. Previous research has shown these replicas lose water linearly and at rates equivalent to similarly sized salamanders (Peterman et al. 2013). Models were cured in a drying oven for 24 h at 70 °C, and then weighed to the nearest 0.01 g on a portable digital balance (Durascale; My Weigh, Vancouver, BC, Canada). Prior to deployment, all replicas were soaked in water for at least 4 h; replicas were deployed within 1 h of sunset, and retrieved within 1 h of sunrise.
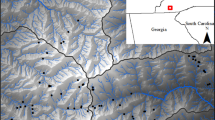
Replicas were deployed at Daniel Boone Conservation Area (DBCA; Fig. 1) along 250-m-long transects, spaced at approximately 50-m intervals (n = 18 transects; 108 locations). Deployment occurred along transects, rather than randomly, due to logistical constraints related to the time sensitivity of deployment and collection. Locations of replica deployment were marked in the field using a handheld GPS (Garmin 62sc; Olathe, KS, USA) with multiple locations being taken until the estimated precision was ≤3 m. Replicas were deployed in both spring (8 April–8 May 2012) and summer (15 August–28 August 2012). At each location, adult- and juvenile-sized replicas were deployed under the leaf litter, and another pair was deployed on top of the leaf litter. Because the focus of this study was the effects of landscape and climate features on water loss, all replicas were housed within cylindrical cages made of 3-mm hardware cloth to prevent replicas from coming in direct contact with leaf litter or soil, which could have confounding effects on water loss rates. These same hardware cloth cages were used during the initial calibration of plaster replicas with live salamanders (Peterman et al. 2013). Each replica was weighed with the portable digital balance upon deployment and retrieval.

Locations of plaster replica Plethodon albagula (western slimy salamander) deployment at Daniel Boone Conservation Area, MI, USA. Replicas were deployed at six locations separated by 50 m along each transect, and there were 18 transects (n = 108 locations)
Spatial and temporal covariates
Spatial covariates used in this analysis were calculated in ArcGIS 9.3 (ESRI, Redlands, CA, USA) and are described in detail by Peterman and Semlitsch (2013). Previously, these covariates were used to predict the spatial distribution of abundance of P. albagula (see details below). In the current study, we assessed the effects of topographic position (TPI), topographic wetness index (TWI), potential relative radiation (PRR), and distance from stream. These variables have a resolution of 3 m, and were derived from 1/9 arc second National Elevation Dataset (http://seamless.usgs.gov/products/9arc.php). Canopy cover was also estimated at DBCA using the normalized difference vegetation index (NDVI), which was calculated from cloud-free Landsat 7 satellite images of our study area taken on 15 June, 20 July, and 9 August 2012 (http://glovis.usgs.gov/). A mean NDVI was calculated by averaging these days together. The resolution of the NDVI layer was 30 m, so it was resampled to a resolution of 3 m. Because the majority of our spring trials were conducted prior to full leaf-out, NDVI was not included in the spring models. For this analysis, we used time since rain, maximum overnight humidity, and maximum temperature of the previous day as temporal climatological covariates. These data were collected from the Big Spring weather station (http://www.wunderground.com), which is located 8 km west of DBCA. For extrapolating our model to the entire DBCA landscape, we determined averages for these measures in spring (1 April–31 May) and summer (1 June–31 August) from data collected in 2005–2012.
Statistical analyses
For each replica, we calculated the proportion of water lost per hour [proportion loss = (deployed mass − retrieved mass)/(deployed mass − dry mass)/time deployed], which became our dependent variable. For this analysis, we did not have competing a priori hypotheses concerning the factors that would affect water loss, but rather, we were interested in fitting the best model possible to explain the spatial and temporal patterns of water loss in our plaster replicas. As such, we did not conduct model selection to detemine parameters to include or exclude from each model, but instead fit a small number of meaningful parameters to each model. Our modeling work flow proceeded as follows. We first divided our data by replica size and season (size-season) to create four independent data sets (juvenile-spring, juvenile-summer, adult-spring, and adult-summer). We then assessed the correlation of each of our independent variables with each other, as well as their correlation with the dependent variable. If two variables had a Pearson’s correlation r ≥ 0.70, we excluded the variable that had the lowest correlation with the dependent variable. Lastly, to limit complexity, we did not include interactions of independent variables, and excluded variables that had r < 0.10 correlation with the dependent variable. Correlations among independent variables revealed that TPI and distance from stream were highly correlated (r = 0.74), but TPI had a greater correlation with rate of water loss in the spring datasets, and distance to stream had a greater correlation in summer datasets. We also found TWI and maximum overnight humidity to have low correlation with water loss across all size-season combinations (r ≤ 0.07), so these variables were not included in the mixed effects models. To account for heterogeneous variance in our data, we fit different variance structures to our data using ‘nlme’ in R (Pinheiro et al. 2013; R Core Team 2014; Zuur et al. 2009), and the best-fit variance structure was determined using AIC (Akaike 1974). Using the model with the best-fit variance structure, we then tested different random effects parameterizations to account for the nested nature of our data (i.e. models within location, locations within transect, transects within date). The percent variance explained by our top model for each size-season combination was assessed using the marginal R 2 measure of Nakagawa and Schielzeth (2013) and calculated with ‘MuMIn’ (Barton 2013). The marginal R 2 describes the percent variation explained in the fixed effects model alone. The full list of variance structures and random effects parameterizations tested in model selection can be found in Electronic Appendix S1.
The fixed effects parameter estimates for the top size-season models were then used to predict water loss rates across the DBCA landscape. Replica position (under leaves or on the surface) was a factor in each model, so for each size-season combination, we calculated a surface and a leaf water loss estimate. For the remainder of this paper, we consider salamander surface activity to be evenly divided between these two states (i.e. 50 % surface, 50 % under leaves). Biologically, this simplification is likely to represent a salamander that is actively moving in search of a mate, dispersing or partially exposed while foraging. Therefore, to calculate a single size-season water loss rate, we averaged the model predictions from surface and leaf models. Because the main objective in this study was to demonstrate water loss as a limiting factor for terrestrial salamanders, we converted water loss rates to surface activity times (SAT). There is no empirical data describing the threshold of water loss when terrestrial plethodontid salamanders cease surface activity and seek refuge, and only one study has experimentally assessed this in a stream-associated salamander (Feder and Londos 1984). Previous studies have used 10 % of body mass lost as the point at which salamanders stop foraging (Feder 1983; Gifford and Kozak 2012), and this threshold has been supported through controlled experimentation (Feder and Londos 1984). For our study, we used 10 % of total water lost as the threshold; SAT was calculated as the time (h) to 10 % water loss. It should be noted that the proportion of a salamander’s body mass comprised of water decreases as mass increases (Peterman et al. 2013):
Ten percent mass loss for juvenile and adult salamanders of sizes equivalent to our replicas would result in 11.9 and 13.3 % loss of water, respectively.
One of our objectives in this study was to determine how predicted SAT relates to the predicted spatial distribution of abundance. The methods and model used to predict salamander abundance across the landscape are described in detail by Peterman and Semlitsch (2013). Briefly, we surveyed 135 plots (3 m × 3 m) at DBCA that were spaced ≥75 m apart seven times in the spring of 2011. During each survey, all moveable cover objects, including rocks, logs, and bark, were carefully lifted and all salamanders were captured by hand. Cover objects were returned to their original position and salamanders were released following data collection. We fit binomial mixture models to our repeated count data using a Bayesian framework (Royle 2004). To account for imperfect observation of salamanders in space and time, we modeled salamander detection probability as a function of survey date, the number of days since a soaking rain event (rain ≥5 mm), and temperature during each survey. After correcting for imperfect detection, abundance was modeled as a function of NDVI, TPI, TWI, and PRR. We then projected the fitted abundance model across the landscape to spatially represent the distribution of salamanders at DBCA.
We conducted Pearson product-moment correlation tests between the abundance estimates at the 135 survey plots from Peterman and Semlitsch (2013) and the spatial SAT predictions made in this study to get a point estimate correlation. We also assessed spatial patterns of correlation between SAT and abundance within ArcGIS using a moving window correlation (Dilts 2010) with a window size of 51 m (17 × 17 pixels). SAT is a physiological measure estimated from several of the same landscape covariates included in the abundance model of Peterman and Semlitsch (2013). To estimate the strength of SAT as a predictor of abundance, we re-ran the binomial mixture model of Peterman and Semlitsch (2013) in this study, but modeled abundance at each of the 135 survey plots solely as a function of SAT. Details of the model parameterization and settings can be found in Electronic Appendix S2.
Peterman and Semlitsch (2013) also used multistate models to identify a potential disconnect between reproductive effort (presence of gravid females) and realized recruitment (presence of juveniles). We generalize that analysis for this study to estimate the probability of juvenile and adult occurrence at each of the 135 plots surveyed by Peterman and Semlitsch (2013). We use these occurrence probabilities as an indirect assessment of reproductive effort. We constructed multistate models using a conditional binomial parameterization in program PRESENCE v.3.1 (MacKenzie et al. 2009). Models were fit separately for adult and juvenile salamanders, with three states being present in each model: (1) no salamanders present (site unoccupied); (2) salamanders present, but focal size class absent; and (3) focal size class present, where the focal size class is either adult [snout-vent length (SVL) ≥ 55 mm; Milanovich et al. 2006] or juvenile (SVL < 55 mm), respectively. As in the abundance model described above, we replaced the individual landscape covariates used by Peterman and Semlitsch (2013) with our integrated SAT measure. From this model, we estimated the conditional probability of occurrence, which is the probability of a focal demographic group occurring at a site, given that a site is suitable to be occupied. Extended details of this analysis and model parameterization are in Peterman and Semlitsch (2013) and Electronic Appendix S2.
Lastly, we determined the mean SVL of salamanders observed at each of the 135 survey plots, and used a linear regression model to assess the relationship between SVL and SAT. Our objectives in re-analyzing the data of Peterman and Semlitsch (2013) are to determine if SAT, as an integrated multivariate parameter, predicts abundance and occupancy of demographic groups, thereby providing a physiological mechanism for the effects of environmental gradients.
Results
To account for heterogeneity within our data, an exponential variance structure was fit to both the juvenile and adult spring data, a combined identity–exponential variance structure was fit to the juvenile summer data, and an identity variance structure was fit to the adult summer data (Table 1). Random-effects fit to each model had both slopes and intercepts varying by covariates (Table 1). The average interval between rainfall events, as determined from the 7 years of climate data, is 1.5 days (±1.98 SD) and 2.2 days (±2.85) and the average daily maximum temperature is 22.5 °C (±6.22) and 31.2 °C (±4.00) for spring and summer seasons, respectively. These 7-year mean estimates were used to make spatial predictions of water loss.
Our final mixed effects models explained the majority of the variance in our data (\(R_{{_{{{\text{GLMM}}\left( m \right)}} }}^{{^{ 2} }}\) = 82.90–98.69 %; Table 1). Notably, simple linear regression models that do not properly account for heterogeneity in variance or the nestedness of our sampling design described 67.15–81.60 % of variation in our data (Table 1). Plaster replica position was a significant predictor of water loss rate for both replica sizes in both seasons, with replicas on the surface losing 1.26–2.64 % more water per hour than adjacent replicas placed under leaves (Table 1). In the spring, water loss in juvenile replicas increased significantly with topographic position (TPI), meaning that water loss was greatest in ridge-like habitat and least in ravine-like habitats. In contrast, topographic position had no effect on adult replicas. Distance from stream had a significant effect on both juvenile and adult replica water loss in the summer, with water loss rates increasing with distance from streams. Solar exposure (PRR) had no effect in the spring, but significantly increased rates of water loss in the summer (Table 1). The number of days since rainfall also significantly increased the rate of water loss in all size-season replicas. As anticipated, water loss increased with maximum temperature in the spring for both juvenile and adult replicas. Surprisingly, temperature had no effect on adult replica water loss in the summer, and had a negative effect on juvenile replica water loss (Table 1). Lastly, canopy cover, as measured by NDVI, was found to have no effect on juvenile replica water loss, but had a significant effect on adult replica water loss; as canopy cover increased, adult replica water loss decreased.
Spatially, there is extensive congruence among each size-season SAT map (Fig. S1), and correlations among these ranged from 0.62 to 0.95 (Table S1). The highest SAT are concentrated within ravine habitats, which are separated by ridges with lower SAT. Mean SAT on the landscape ranged from 1.94 h for juveniles in the summer to 9.90 h for adults in the spring (Table 2). Paired t tests revealed that juvenile SAT is significantly less than adult SAT in spring and summer, and that all SAT are significantly less in the summer (all tests P < 0.0001). In general, the estimated SAT is 3 times longer in spring than summer, and is about 1.5 times longer for adults than juveniles, regardless of season (Table 2; Fig. S1). Correlations of predicted salamander abundance with size-season SAT at the 135 survey plots were also high (r = 0.35–0.63; Table 2). Adult summer SAT had the highest correlation with predicted abundance (r = 0.63), largely because of the significance of canopy cover in mitigating water loss (Table 1). Spatial similarities between predicted salamander abundance and adult summer SAT are evident (Fig. 2a, b); the correlation between abundance and SAT is generally highest in areas of low predicted abundance and low SAT (Fig. 2c).

a Predicted salamander abundance, b summer surface activity time (h) estimated from adult-sized plaster replicas, and c spatial Pearson’s r correlation values. There is generally very high, positive correlation between estimated abundance and surface activity time; as activity time increases, so does the predicted abundance of salamanders
Because adult summer SAT had the highest correlation with predicted abundance, we explored in more detail its relationships with predicted abundance, salamander size distribution, and probability of occurrence. We do note, however, that the other size-season models also had significant correlations with predicted abundance (r = 0.35–0.48; Table 2). The binomial mixture model fit with predicted adult summer SAT as the sole independent variable in the abundance model fit the data well, and SAT had a significant effect on abundance, with abundance increasing as predicted SAT increased (Electronic Appendix S2; Fig. 3a). Further, we found that the mean SVL of salamanders observed at 88 of the 135 surveyed plots (n = 407 unique salamanders measured; Peterman and Semlitsch 2013) significantly increased as predicted SAT decreased (F 1, 86 = 8.38; P = 0.005; R 2 = 0.089; Fig. 3b), suggesting that, on average, larger salamanders are found in areas with limited SAT. Similarly, we found that the conditional probability of juvenile salamander occupancy at the 135 surveyed plots, correcting for imperfect detection, significantly increased as predicted SAT increased (Electronic Appendix S2; Fig. 3c). In contrast, the conditional probability of adult occupancy was not significantly related to adult summer SAT (Fig. 3c), and there was little variation in predicted adult occupancy probability across the range of predicted SAT (adult conditional occupancy probability = 0.91–0.95). Predicted conditional occupancy probabilities of juveniles at the same 135 sites ranged from 0.35 to 0.92; (Fig. 3c).

Relationship of estimated adult summer active time with a predicted abundance; b mean SVL of salamanders; and c conditional probability of occurrence of adult and juvenile size classes based on data collected during seven surveys of 135 plots. Dashed lines around the estimates represent 95 % prediction intervals. Increased surface activity time resulted in more salamanders being present, and juvenile salamanders were more likely to be found in areas with higher surface activity times
Discussion
Our study assessed patterns of water loss as a process that varies spatially and temporally as a function of fine-scale environmental gradients and temporal climatic conditions. We found that spatial estimates of SAT derived from rates of water loss were significantly correlated with predicted salamander abundance and that SAT was a significant predictor of abundance as well as population demographic characteristics. Importantly, our SAT estimates were independently derived from plaster replicas deployed under field conditions, and were in no way contingent upon actual salamander distributions. Results from our study extend our understanding of terrestrial plethodontid ecology. Previous research has only logically conjectured the importance of hydric relationships and surface activity as mechanisms underlying local distribution and population dynamics by extrapolating results from controlled laboratory experiments or indirectly through field observations (Feder 1983; Feder and Londos 1984; Spotila 1972). As an integrated measure of the local environment and climate, SAT was a significant predictor of abundance as well as the occurrence of juvenile salamanders (a potential indicator of successful recruitment). Combined with our findings that SAT and abundance are spatially correlated, we have compelling evidence that water loss is a physiologically limiting factor underlying the abundance–habitat and population dynamic relationships described by Peterman and Semlitsch (2013).
Smaller organisms and individuals are often most susceptible to water loss (Spight 1968), and we found that juvenile-sized plaster replicas lost water at 1.5–3 times greater rate than adult-sized replicas. Such differences significantly curtail surface activity, and could lead to differential survival across the landscape. In support of this, we found that the mean body size of salamanders was smaller in plots with lower rates of water loss and high SAT (Fig. 3b). Further, we found that the probability of encountering a juvenile salamander in areas of high SAT was significantly greater than areas of low SAT. In contrast, we found that adults were more uniformly distributed across the landscape, regardless of SAT (Fig. 3c). These patterns suggest that reproductive rates may be greater in high SAT regions of the landscape, or that survival of juvenile salamanders is higher in high SAT areas. Either or both of these processes would contribute to the increased abundance of salamanders in high SAT regions (Fig. 3a). Differentiating these processes as the mechanisms underlying the spatial variation in size distribution will likely only be possible through long-term, detailed studies of local demographic processes.
In corroboration with seasonal patterns of surface activity of salamanders in the field (Milanovich et al. 2006), estimated SAT differed significantly among spring and summer seasons (Table 2; Fig. S1). Although SAT was three times greater in the spring, there is still pronounced spatial heterogeneity in SAT due to the influence of topographic position in affecting water loss. The mixed effects models describing the spatial patterns of water loss for adult- and juvenile-sized replicas in the spring were nearly identical (Table 1). In the summer, there was no relationship between juvenile replicas and canopy cover, while adult replicas lost significantly less water as canopy cover increased. We speculate that the rate of water loss was so rapid in the high surface area juvenile models that canopy cover did little to attenuate losses. Although Peterman et al. (2013) found water loss rates of plaster replicas to be linear over an 8-h laboratory test with up to 35 % water loss, we note the possibility that rates of water loss could become non-linear as dehydration deficits approaches 100 % (summer dehydration deficit for juvenile replicas: mean = 60.3 %, max. = 98.5 %; adult replicas: mean = 39.4 %, max. = 82.1 %). Such non-linearity could contribute to the observed differences in parameter estimates for adult and juvenile models.
If reproductive success differs across the landscape, then P. albagula may be described as existing as a spatially-structured population (Harrison 1991; Thomas and Kunin 1999). Specifically, reproductive rates and success may be greatest within forested ravines with high SAT, and be negligible or non-existent where SAT is low. As such, the presence of salamanders in low SAT areas of the landscape would predominantly depend upon salamanders dispersing from high SAT regions, implying fine-scale source–sink dynamics (Pulliam 1988). Little is known concerning dispersal in plethodontid salamanders, but as adults they are generally considered to be highly philopatric with small home ranges (Kleeberger and Werner 1982; Liebgold et al. 2011; Ousterhout and Liebgold 2010). A landscape genetic analysis of P. albagula at DBCA, found that the rate of water loss was unequivocally the best predictor of spatial genetic differentiation (Peterman et al. 2014). Surprisingly, movement rates were predicted to increase through areas of the landscape with higher rates of water loss, suggestive of compensatory movement behaviors. Salamander increase their movement rate to avoid high cost regions of the landscape, but have minimal incentive to disperse from low cost areas such as those with low rates of water loss and high activity times.
Our findings suggest that water relationships temporally and seasonally shape activity times, locally dictate habitat use, and regionally delineate distributions. Nonetheless, water loss is not a physiological process working in isolation. Metabolic rates of ectotherms are temperature-dependent, increasing with environmental temperature. Because evaporative water loss also increases with temperature (Spotila 1972; Tracy et al. 2010), plethodontid salamanders are doubly challenged under hot, dry conditions. As metabolic demands increase with temperature there is a greater need for energy intake, but surface activity will likely be curtailed at higher temperatures due to increased rates of water loss. This limitation on activity is akin to the hours of restriction described by Sinervo et al. (2010), which has been implicated in the local extinction of lizard populations. The relationship of energy expenditure and intake, as a function of temperature and foraging time (limited by water loss), was incorporated into a mechanistic energy budget model and used to accurately predict the elevational distribution of a montane woodland salamander (Gifford and Kozak 2012). Although temperature variation exists across our landscape and correlates with predicted abundance (Peterman and Semlitsch 2013), the independent (or interactive) role that spatial variation in temperature has on salamander metabolic rate, and subsequently on abundance and population dynamics, is unclear. Mechanistic modeling approaches, as used by Gifford and Kozak (2012), may be able to provide insight into these questions.
Although we observed significant spatial correlation between SAT and predicted salamander abundance, correlations were not perfect. Included in the original abundance model of Peterman and Semlitsch (2013) were topographic wetness and an interaction between topographic wetness and solar exposure. These terms were not included in our mixed effects models to limit model complexity and because there was minimal correlation with measured rates of water loss. Exclusion of these factors could explain some of the SAT–abundance discrepancies, although our mixed effects models were able to explain the majority of the variation in our data, leaving little unexplained variance to be accounted for by other factors.
Plaster replicas effectively mimicked water loss rates of living salamanders (Peterman et al. 2013), but we nonetheless made several simplifying assumptions. First, evaporative water loss in wet-skinned amphibians is determined by the moisture content of the air and the difference in the water vapor density at the surface of the animal (Spotila et al. 1992), but atmospheric moisture can vary over small spatial scales and as a function of topography and vegetation (Campbell and Norman 1998). While we attempted to account for humidity variation by using synoptic meteorological measurements, relative humidity did not correlate with water loss and was omitted from our mixed effect models. Fine-scale estimation of variation of relative humidity is likely necessary to more accurately estimate evaporative water loss in salamanders, but we note that TPI and distance from stream in our study are likely to be highly correlated with fine-scale humidity variation (Holden and Jolly 2011). Second, under wind-free conditions, a boundary layer will form around a stationary object (Tracy 1976), which reduces the rate of evaporative water loss. Our estimates of water loss from plaster replicas are therefore likely conservative as foraging or dispersal movements of surface-active salamanders would disrupt the boundary layer and increase rates of water loss. Third, a critical aspect of terrestrial salamander water balance is their ability to rehydrate by absorbing water across their skin (Spotila 1972), but we sought to avoid contact of our replicas with the leaf litter and soil to minimize the potentially confounding effects of these factors on evaporative water loss. Fourth, we calculated SAT based on the assumption that surface-active salamanders spend equal amounts of time above and below leaves. Depending upon whether salamanders spend more time under or on top of the leaf litter, the rate of water loss will be reduced or increased by as much as 0.63 %/h, respectively (Table 1). Finally, plethodontid salamanders, like most amphibians, can assume water conservation postures to reduce rates of water loss (Ray 1958; Spotila 1972). However, such behaviors appear relatively ineffective and are likely a last ditch effort only sought if retreat to a moist refuge is not possible (Wells 2007).
Our study is the first to estimate spatially explicit rates of water loss for a terrestrial amphibian under relevant ecological field conditions. Previous research has carefully detailed the physiological relationships of amphibians with their environment (reviewed by Feder 1983; Shoemaker et al. 1992; Spotila et al. 1992; Wells 2007), but only superficial attempts have been made to relate physiology with patterns observed in nature (Spotila 1972). Water loss is unlikely to be the only factor limiting terrestrial salamander activity and spatial distributions. However, with results presented in this study, as well as the findings of Peterman et al. (2014) that water loss rates predict spatial genetic differentiation, there is strong support that water balance is critical. Future work should explore how temperature, metabolic rate, and spatial energy budgets (Gifford and Kozak 2012) relate to patterns of abundance and population processes. Additionally, spatial genetic processes of terrestrial salamanders in relation to landscape features are largely unknown, but understanding how fine-scale environmental gradients relate to population and landscape genetics may provide critical insight into how physiology affects local population dynamics and dispersal (Peterman et al. 2014).
Finally, our findings that abundance and spatial demographic patterns can be predicted by SAT have implications for the future persistence of terrestrial salamanders. Beyond plethodontid salamanders specifically, and amphibians generally, our results may be extensible to other small organisms such as terrestrial gastropods and arthropods that have close ties with temperature, moisture and relative humidity (Cameron 1970; Cloudsley-Thompson 1962). Two of the greatest threats facing all organisms are loss of habitat and changing climate (Mantyka-Pringle et al. 2012). Current climate change scenarios are forecasting more extreme temperatures and increased variability in the interval and amount of rainfall (Field et al. 2012), and changes in these climatological parameters have already, or are predicted to, profoundly affect terrestrial organisms (Milanovich et al. 2010; Parmesan and Yohe 2003). By incorporating water loss and surface activity time into biophysical or dynamic population models, it may be possible to gain a better understanding of the effects that changing environmental and climatological conditions will have on terrestrial animals that are sensitive to water loss.
References
Adolph EF (1932) The vapor tension relations of frogs. Biol Bull 62:112–125. doi:10.2307/1537147
Adolph SC (1990) Influence of behavioral thermoregulation on microhabitat use by two Sceloporus lizards. Ecology 71:315–327. doi:10.2307/1940271
Akaike H (1974) New look at statistical-model identification. IEEE Trans Automatic Control 19:716–723
Angilletta MJ (2009) Thermal adaptation: a theoretical and empirical synthesis. Oxford University Press, New York
Angilletta MJ, Steury TD, Sears MW (2004) Temperature, growth rate, and body size in ectotherms: fitting pieces of a life-history puzzle. Integr Comp Biol 44:498–509. doi:10.1093/icb/44.6.498
Antvogel H, Bonn A (2001) Environmental parameters and microspatial distribution of insects: a case study of carabids in an alluvial forest. Ecography 24:470–482. doi:10.1111/j.1600-0587.2001.tb00482.x
Barton K (2013) MuMIn: multi-model inference. R package version 1.9.5. http://CRAN.R-project.org/package=MuMIn
Bennie J, Huntley B, Wiltshire A, Hill MO, Baxter R (2008) Slope, aspect and climate: spatially explicit and implicit models of topographic microclimate in chalk grassland. Ecol Model 216:47–59. doi:10.1016/j.ecolmodel.2008.04.010
Boag DA (1985) Microdistribution of three genera of small terrestrial snails (Stylommatophora: Pulmonata). Can J Zool 63:1089–1095. doi:10.1139/z85-163
Brewster CL, Sikes RS, Gifford ME (2013) Quantifying the cost of thermoregulation: thermal and energetic constraints on growth rates in hatchling lizards. Funct Ecol 27:490–497. doi:10.1111/1365-2435.12066
Buckley LB, Roughgarden J (2005) Effect of species interactions on landscape abundance patterns. J Anim Ecol 74:1182–1194. doi:10.1111/j.1365-2656.2005.01012.x
Buckley LB, Urban MC, Angilletta MJ, Crozier LG, Rissler LJ, Sears MW (2010) Can mechanism inform species’ distribution models? Ecol Lett 13:1041–1054. doi:10.1111/j.1461-0248.2010.01479.x
Cameron RAD (1970) Differences in the distributions of three species of Helicid snail in the limestone district of Derbyshire. Proc R Soc Lond B 176:131–159. doi:10.1098/rspb.1970.0039
Campbell GS, Norman JM (1998) Introduction to environmental biophysics, 2nd edn. Springer, New York
Chen J et al (1999) Microclimate in forest ecosystem and landscape ecology. Bioscience 49:288–297
Cloudsley-Thompson J (1962) Microclimates and the distribution of terrestrial arthropods. Annu Rev Entomol 7:199–222
Dilts T (2010) Topography tools for ArcGIS v.9.3. http://arcscripts.esri.com/details.asp?dbid=15996
Feder ME (1983) Integrating the ecology and physiology of plethodontid salamanders. Herpetologica 39:291–310. doi:10.2307/3892572
Feder ME, Londos PL (1984) Hydric constraints upon foraging in a terrestrial salamander, Desmognathus ochrophaeus; (Amphibia: Plethodontidae). Oecologia 64:413–418. doi:10.1007/bf00379141
Field CB et al (2012) Managing the risks of extreme events and disasters to advance climate change adaptation. Cambridge University Press, Cambridge
Getz LL (1968) Influence of water balance and microclimate on the local distribution of the redback vole and white-footed mouse. Ecology 49:276–286. doi:10.2307/1934456
Gifford ME, Kozak KH (2012) Islands in the sky or squeezed at the top? Ecological causes of elevational range limits in montane salamanders. Ecography 35:193–203. doi:10.1111/j.1600-0587.2011.06866.x
Grover MC (1998) Influence of cover and moisture on abundances of the terrestrial salamanders Plethodon cinereus and Plethodon glutinosus. J Herpetol 32:489–497
Grover MC, Wilbur HM (2002) Ecology of ecotones: interactions between salamanders on a complex environmental gradient. Ecology 83:2112–2123
Harrison S (1991) Local extinction in a metapopulation context: an empirical evaluation. Biol J Linn Soc 42:73–88. doi:10.1111/j.1095-8312.1991.tb00552.x
Heatwole H (1962) Environmental factors influencing local distribution and activity of the salamander, Plethodon cinereus. Ecology 43:460–472
Highton R (1989) Biochemical evolution in the slimy salamanders of the Plethodon glutinosus complex in the eastern United States. Part 1. Geographic protein variation. Illinois Biol Monogr 57:1–78
Holden ZA, Jolly WM (2011) Modeling topographic influences on fuel moisture and fire danger in complex terrain to improve wildland fire management decision support. For Ecol Manag 262:2133–2141. doi:10.1016/j.foreco.2011.08.002
Huey RB (1991) Physiological consequences of habitat selection. Am Nat 137:S91–S115. doi:10.2307/2462290
Kearney M (2012) Metabolic theory, life history and the distribution of a terrestrial ectotherm. Funct Ecol 26:167–179. doi:10.1111/j.1365-2435.2011.01917.x
Kearney M, Porter W (2009) Mechanistic niche modelling: combining physiological and spatial data to predict species’ ranges. Ecol Lett 12:334–350. doi:10.1111/j.1461-0248.2008.01277.x
Kearney M, Phillips BL, Tracy CR, Christian KA, Betts G, Porter WP (2008) Modelling species distributions without using species distributions: the cane toad in Australia under current and future climates. Ecography 31:423–434. doi:10.1111/j.0906-7590.2008.05457.x
Keen WH (1979) Feeding and activity patterns in the salamander Desmognathus ochrophaeus (Amphibia, Urodela, Plethodontidae). J Herpetol 13:461–467. doi:10.2307/1563483
Keen WH (1984) Influence of moisture on the activity of a plethodontid salamander. Copeia 1984:684–688
Keith DA et al (2008) Predicting extinction risks under climate change: coupling stochastic population models with dynamic bioclimatic habitat models. Biol Lett 4:560–563. doi:10.1098/rsbl.2008.0049
Kleeberger SR, Werner JK (1982) Home range and homing behavior of Plethodon cinereus in northern Michigan. Copeia 1982:409–415. doi:10.2307/1444622
Liebgold EB, Brodie ED, Cabe PR (2011) Female philopatry and male-biased dispersal in a direct-developing salamander, Plethodon cinereus. Mol Ecol 20:249–257. doi:10.1111/j.1365-294X.2010.04946.x
MacKenzie DI, Nichols JD, Seamans ME, Gutiérrez RJ (2009) Modeling species occurrence dynamics with multiple states and imperfect detection. Ecology 90:823–835. doi:10.1890/08-0141.1
Maiorana VC (1977) Tail autotomy, functional conflicts and their resolution by a salamander. Nature 265:533–535
Mantyka-Pringle CS, Martin TG, Rhodes JR (2012) Interactions between climate and habitat loss effects on biodiversity: a systematic review and meta-analysis. Glob Change Biol 18:1239–1252. doi:10.1111/j.1365-2486.2011.02593.x
Milanovich J, Trauth SE, Saugey DA, Jordan RR (2006) Fecundity, reproductive ecology, and influence of precipitation on clutch size in the western slimy salamander (Plethodon albagula). Herpetologica 62:292–301. doi:10.1655/0018-0831(2006)62[292:FREAIO]2.0.CO;2
Milanovich JR, Peterman WE, Nibbelink NP, Maerz JC (2010) Projected loss of a salamander diversity hotspot as a consequence of projected global climate change. PLoS ONE 5:e12189. doi:10.1371/journal.pone.0012189
Milanovich JR, Trauth SE, Saugey DA (2011) Reproduction and age composition of a population of woodland salamanders (Plethodon albagula) after a prescribed burn in southwestern arkansas. Southwest Nat 56:172–179
Nakagawa S, Schielzeth H (2013) A general and simple method for obtaining R 2 from generalized linear mixed-effects models. Methods Ecol Evol 4:133–142. doi:10.1111/j.2041-210x.2012.00261.x
Ousterhout BH, Liebgold EB (2010) Dispersal versus site tenacity of adult and juvenile red-backed salamanders (Plethodon cinereus). Herpetologica 66:269–275. doi:10.1655/09-023.1
Parmesan C, Yohe G (2003) A globally coherent fingerprint of climate change. Nature 421:37–42
Peterman WE, Semlitsch RD (2013) Fine-scale habitat associations of a terrestrial salamander: the role of environmental gradients and implications for population dynamics. PLoS ONE 8:e62184. doi:10.1371/journal.pone.0062184
Peterman WE, Locke JL, Semlitsch RD (2013) Spatial and temporal patterns of water loss in heterogeneous landscapes: using plaster models as amphibian analogues. Can J Zool 91:135–140. doi:10.1139/cjz-2012-0229
Peterman WE, Connette GM, Semlitsch RD, Eggert LS (2014) Ecological resistance surfaces predict fine-scale genetic differentiation in a terrestrial woodland salamander. Mol Ecol 23:2402–2413. doi:10.1111/mec.12747
Petranka JW (1998) Salamanders of the United States and Canada. Smithsonian Institution Press, Washington, DC
Pinheiro JC, Bates DM, DebRoy S, Sarkar D (2013) nlme: linear and nonlinear mixed effects models. R package version 3.1-108, vol. R package version 3.1-108
Pulliam HR (1988) Sources, sinks and population regulation. Am Nat 132:652–661
R Core Team (2014) R: A language and environment for statistical computing, R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/
Ray C (1958) Vital limits and rates of desiccation in salamanders. Ecology 39:75–83. doi:10.2307/1929968
Royle JA (2004) N-mixture models for estimating population size from spatially replicated counts. Biometrics 60:108–115
Scherrer D, Körner C (2011) Topographically controlled thermal-habitat differentiation buffers alpine plant diversity against climate warming. J Biogeogr 38:406–416. doi:10.1111/j.1365-2699.2010.02407.x
Shoemaker VH et al (1992) Exchange of water, ions, and respiratory gases in terrestrial amphibians. In: Feder ME, Burggren WW (eds) Environmental physiology of the amphibians. University of Chicago Press, Chicago, pp 125–150
Sinervo B et al (2010) Erosion of lizard diversity by climate change and altered thermal niches. Science 328:894–899
Spight TM (1968) The water economy of salamanders: evaporative water loss. Physiol Zool 41:195–203
Spotila JR (1972) Role of temperature and water in the ecology of lungless salamanders. Ecol Monogr 42:95–125
Spotila JR, Berman EN (1976) Determination of skin resistance and the role of the skin in controlling water loss in amphibians and reptiles. Comp Biochem Physiol A 55:407–411. doi:10.1016/0300-9629(76)90069-4
Spotila JA, O’Connor MP, Bakken GS (1992) Biophysics og heat and mass transfer. In: Feder ME, Burggren WW (eds) Environmental physiology of amphibians. University of Chicago Press, Chicago, pp 59–81
Taub FB (1961) The distribution of the red-backed salamander, Plethodon c. cinereus, within the soil. Ecology 42:681–698
Thomas CD, Kunin WE (1999) The spatial structure of populations. J Anim Ecol 68:647–657. doi:10.1046/j.1365-2656.1999.00330.x
Tracy CR (1976) A model of the dynamic exchanges of water and energy between a terrestrial amphibian and its environment. Ecol Monogr 46:293–326
Tracy CR, Christian KA, Tracy CR (2010) Not just small, wet, and cold: effects of body size and skin resistance on thermoregulation and arboreality of frogs. Ecology 91:1477–1484. doi:10.1890/09-0839.1
Wachob DG (1996) The effect of thermal microclimate on foraging site selection by wintering mountain chickadees. Condor 98:114–122. doi:10.2307/1369514
Weiss SB, Murphy DD, White RR (1988) Sun, slope, and butterflies: topographic determinants of habitat quality for Euphydryas editha. Ecology 69:1486–1496
Wells KD (2007) The ecology and behavior of amphibians. University of Chicago Press, Chicago
Whitford WG, Hutchison VH (1967) Body size and metabolic rate in salamanders. Physiol Zool 40:127–133. doi:10.2307/30152447
Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgments
We thank D. Hocking for discussions on statistical analysis, and G. Connette for helpful discussion and comments. This manuscript was improved by the insightful comments of two anonymous reviewers and H. Ylonen. Support was provided by the University of Missouri Research Board (CB000402), Trans World Airline Scholarship, and the Department of Defense Strategic Environmental Research and Development Program (RC2155). This research was done in accordance with the laws of the state of Missouri and the USA, approved the University of Missouri Animal Care and Use Committee (#7403), and conducted under Missouri Wildlife Collector’s Permit #15203.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Indrikis Krams.
Electronic supplementary material
Below is the link to the electronic supplementary material.
442_2014_3041_MOESM1_ESM.doc
Supplementary material 1 (DOC 20556 kb) Appendix S1. Detailed methods of how mixed effect models were made to estimate rates of water loss. Appendix S2. Details of field methods used to collect salamander abundance and size data, as well as detailed description of abundance and multistate modelling procedures
Rights and permissions
About this article

Cite this article
Peterman, W.E., Semlitsch, R.D. Spatial variation in water loss predicts terrestrial salamander distribution and population dynamics. Oecologia 176, 357–369 (2014). https://doi.org/10.1007/s00442-014-3041-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-014-3041-4