
Abstract
The mineral in bone is located primarily within the collagen fibril, and during mineralization the fibril is formed first and then water within the fibril is replaced with mineral. Our goal is to understand the mechanism of fibril mineralization, and as a first step we recently determined the size exclusion characteristics of the fibril. This study indicates that apatite crystals up to 12 unit cells in size can access the water within the fibril while molecules larger than a 40-kDa protein are excluded. We proposed a novel mechanism for fibril mineralization based on these observations, one that relies exclusively on agents excluded from the fibril. One agent generates crystals outside the fibril, some of which diffuse into the fibril and grow, and the other selectively inhibits crystal growth outside of the fibril. We have tested this mechanism by examining the impact of removing the major serum inhibitor of apatite growth, fetuin, on the serum-induced calcification of collagen. The results of this test show that fetuin determines the location of serum-driven mineralization: in fetuin’s presence, mineral forms only within collagen fibrils; in fetuin’s absence, mineral forms only in solution outside the fibrils. The X-ray diffraction spectrum of serum-induced mineral is comparable to the spectrum of bone crystals. These observations show that serum calcification activity consists of an as yet unidentified agent that generates crystal nuclei, some of which diffuse into the fibril, and fetuin, which favors fibril mineralization by selectively inhibiting the growth of crystals outside the fibril.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
The type I collagen fibril plays several critical roles in bone mineralization. The mineral in bone is located primarily within the fibril [1–6], and during mineralization the fibril is formed first and then water within the fibril is replaced with mineral [7, 8]. The collagen fibril therefore provides the aqueous compartment in which mineral grows. We have recently shown that the physical structure of the collagen fibril plays an important additional role in mineralization: the role of a gatekeeper that allows molecules smaller than a 6-kDa protein to freely access the water within the fibril while preventing molecules larger than a 40-kDa protein from entering the fibril [9]. Molecules smaller than a 6-kDa protein can therefore enter the fibril and interact directly with mineral to influence crystal growth. Molecules larger than a 40-kDa protein cannot enter the fibril and so have no ability to act directly on the apatite crystals growing within the fibril.
Molecules too large to enter the collagen fibril can still have important effects on mineralization within the fibril. We have suggested that large inhibitors of apatite growth can paradoxically favor mineralization within the fibril by selectively preventing apatite growth outside of the fibril [9]. We have also proposed that large nucleators of apatite formation may generate small crystal nuclei outside of the collagen fibril and that some of these nuclei subsequently diffuse into the fibril and grow [9]. Because the size exclusion characteristics of the fibril allow rapid penetration of molecules the size of a 6-kDa protein, apatite crystals up to 12 unit cells in size should in principle be able to freely access all of the water within the fibril [9]. The present study tests these hypotheses for the possible function of large molecules in mineralization.
The calcification assay we employed to test the function of large proteins in collagen mineralization is based on our discovery that the type I collagen fibrils of tendon and demineralized bone calcify when incubated in serum (or plasma) for 6 days at 37°C and pH 7.4 [10, 11]. The calcification activity responsible for collagen mineralization in serum consists of one or more proteins that are 50–150 kDa in size [11]. Because these molecules are too large to penetrate the collagen fibril, they must be able to act outside the fibril to cause calcification within the fibril. The serum-driven calcification of a collagen fibril is therefore an excellent model system to explore the mechanisms by which molecules too large to penetrate the collagen fibril can nonetheless cause the fibril to calcify.
Although serum-driven collagen calcification is an in vitro, cell-free assay, there are several reasons to believe that it could be relevant to understanding mechanisms by which collagen fibrils are mineralized in normal bone formation. (1) The assay conditions are physiologically relevant: collagen added to serum calcifies when incubated at the temperature and pH of mammalian blood, without the need to add anything to serum to promote mineralization, such as β-glycerophosphate or phosphate [see 10 and references therein]. (2) Serum is relevant to bone mineralization: osteoblasts form bone in a vascular compartment [12], and proteins in serum have direct access to the site of collagen fibril formation and mineralization while proteins secreted by the osteoblast appear rapidly in serum. (3) Serum-driven calcification is evolutionarily conserved: serum calcification activity appeared in animals at the time vertebrates acquired the ability to form calcium phosphate mineral structures, with no evidence for a similar activity in the serum of invertebrates [13]. (4) Serum-driven calcification is specific: calcification is restricted to those structures that were calcified in bone prior to demineralization, with no evidence of calcification in cartilage at the bone ends or in cell debris [10, 11]. (5) Serum-driven calcification can achieve total recalcification of demineralized bone: serum-driven calcification progresses until the recalcified bone is comparable to the original bone prior to demineralization in mineral content and composition, radiographic density, and powder X-ray diffraction spectrum [11].
The initial goal of the present experiments was to examine the possible function of the 48-kDa protein fetuin in the serum-driven calcification of collagen matrices. Our working hypothesis was that fetuin promotes calcification within the collagen fibril by selectively inhibiting apatite growth outside of the fibril. This hypothesis is supported by the observation that fetuin is the most abundant serum inhibitor of apatite crystal growth [14, 15] and by the observation that fetuin is too large to penetrate the interior of the collagen fibril [9], where serum-induced collagen calcification occurs [11]. The present study tests this hypothesis by examining the impact of removing fetuin from serum on the ability of serum to mineralize the collagen fibril. The results of this test show that the presence of fetuin in serum determines the location of serum-driven mineralization: in the presence of fetuin, mineral forms only within the collagen fibril; in the absence of fetuin, mineral forms only in the solution outside the fibril.
Because fetuin is the subject of this study, it is useful to review briefly its structure, occurrence, and calcification-inhibitory activity. Fetuin is a 48-kDa glycoprotein that consists of two N-terminal cystatin domains and a smaller C-terminal domain. The five oligosaccharide moieties of the protein account for ∼25% of fetuin’s mass and, because of their disordered structures, give fetuin an apparent size in sodium dodecyl sulfate (SDS) gel electrophoresis and Sephacryl gel filtration of about 59 kDa. Fetuin is synthesized in the liver and found at high concentrations in mammalian serum [16, 17] and bone [18–23]. The serum fetuin concentration in adult mammals ranges 0.5–1.5 mg/mL, while that in the fetus and neonate is typically far higher [17]. Fetuin is also one of the most abundant noncollagenous proteins found in bone [18–23], with a concentration of about 1 mg/g bone in rat [22], bovine [18], and human [20, 24] bone. In spite of the abundance of fetuin in bone, however, it has not been possible to demonstrate the synthesis of fetuin in calcified tissues; it is therefore presently thought that the fetuin found in bone arises from hepatic synthesis via serum [21, 23]. This view is supported by the observation that fetuin binds strongly to apatite, the mineral phase of bone, and is selectively concentrated from serum onto apatite in vitro [19].
In vitro studies have demonstrated that fetuin is an important inhibitor of apatite growth and precipitation in serum containing increased levels of calcium and phosphate [15] and that targeted deletion of the fetuin gene reduces the ability of serum to arrest apatite formation by over 70% [14]. More recent studies have shown that a fetuin–mineral complex is formed in the course of the fetuin-mediated inhibition of apatite growth and precipitation in serum containing increased calcium and phosphate [25]. Purified bovine fetuin has also been shown to be a potent inhibitor of the growth and precipitation of a calcium phosphate mineral phase from supersaturated solutions of calcium phosphate [15], and recent studies have shown that a fetuin–mineral complex is formed in the course of this inhibition [25].
Experimental Procedures
Materials
Forty-day-old and newborn albino rats (Sprague-Dawley) were purchased from Harlan Labs (Indianapolis, IN). Adult bovine serum was purchased from Invitrogen (Carlsbad, CA). Each 500 mL volume of Dulbecco’s modified eagle medium (DMEM; GIBCO, Grand Island, NY) was supplemented with 5 mL of penicillin-streptomycin (GIBCO) and 1 mL of 10% sodium azide to prevent bacterial growth. Unless otherwise stated, the concentration of phosphate in DMEM was increased from the basal 0.9 mM to a final 2 mM by addition of 0.5 M sodium phosphate buffer (pH 7.4). When prepared as described [26], DMEM containing 2 mM phosphate is stable for at least 3 weeks at 37°C, with no evidence for loss of calcium or phosphate from the medium or formation of a mineral phase. Bovine fetuin, purified type I collagen from bovine Achilles tendon, and alizarin red S were purchased from Sigma (St. Louis, MO).
Rats were killed by exsanguination while under isoflurane anesthetic; the UCSD Animal Subjects Committee approved all animal experiments. Tail tendons were dissected from 40-day-old rats, and tibias were dissected from newborn rats. Both tissues were extracted with a 1,000-fold excess (v/w) of 0.5 M ethylenediaminetetraacetic acid (EDTA, pH 7.5) for 72 hours at room temperature, to kill cells and remove any mineral that might be present; the tissues were then washed exhaustively with ultrapure water to remove all traces of EDTA and stored at −20°C until use.
Calcification Procedures
Experiments to examine the calcification of collagen matrices were carried out using 24-well cell culture clusters (Costar 3524; Corning, Corning, NY) in a humidified incubator at 37°C and 5% CO2. Each well contained a 1 mL volume of DMEM alone or of DMEM containing 10% bovine serum or fetuin-depleted bovine serum. The amount of matrix added to each 1 ml volume was as follows: a single hydrated, demineralized newborn rat tibia, a portion of tail tendon (3 mg dry weight, hydrated before use), or a portion of type I collagen (3 mg dry weight, hydrated before use). Each tissue was then incubated for 6 days.
Biochemical Analyses
The procedures used for alizarin red staining have been described [27]. For histological analyses, tibias were fixed in 100% ethanol for at least 1 day at room temperature; San Diego Pathology (San Diego, CA) sectioned and von Kossa–stained the tibias. For quantitative assessment of the extent of calcification, alizarin red–stained matrices and precipitates formed outside the matrix were extracted for 24 hours at room temperature with 1 mL of 0.15 M HCl, as described [27]. Calcium levels in culture media and in the acid extracts of tissues and precipitates were determined colorimetrically using cresolphthalein complexone (JAS Diagnostics, Miami FL), and phosphate levels were determined colorimetrically as described [28].
Powder X-ray diffraction was used to compare the mineral phase formed in fetuin-depleted serum with the crystals isolated from rat bone [29]. The mineral was generated by incubating 2 mL DMEM containing 10% fetuin-depleted bovine serum for 48 hours at 37°C. The mineral suspension was diluted to 20 mL with fresh DMEM and incubated for another 48 hours, and the resulting 20 mL of mineral suspension was subsequently diluted to 200 mL with fresh DMEM and incubated for a final 48 hours. The mineral was collected by centrifugation, washed with ethanol, and dried to give 23 mg of mineral. The X-ray diffraction spectrum of this mineral was measured with Cu Kα X-rays (λ = 1.54 Å) using a Rigaku (Tokyo, Japan) Miniflex diffractometer.
Immunological Procedures
Rabbits were immunized against purified bovine fetuin. The procedures employed for the bovine fetuin radioimmunoassay used this antiserum at a final 1:2,000 dilution. The radioimmunoassay diluent, sample volumes, and procedures are identical to those used in the rat fetuin radioimmunoassay [30]. For affinity purification of antifetuin antibody, 16 mg of purified bovine fetuin were covalently attached to 5 mL of cyanogen bromide (CNBr)–activated Sepharose 4B (Amersham Biosciences, Chalfont St. Giles, UK) and packed into a column. Antifetuin antiserum (10 mL) was then passed over this fetuin affinity column, and the bound antibody was eluted with 100 mM glycine (pH 2.5). An antifetuin antibody column was subsequently prepared by covalently attaching 7 mg of purified antifetuin antibody to 5 mL of CNBr-activated Sepharose 4B. The antifetuin antibody column was then equilibrated with the DMEM calcification buffer, and bovine serum was dialyzed against the same buffer. Adult bovine serum was freed of fetuin by passing 0.85-mL aliquots of dialyzed serum over the column at room temperature. The absorbance at 280 nm of each 0.8-mL fraction was then determined, and the fetuin content of the fractions was measured by radioimmunoassay. The four fractions with the highest absorbance were pooled and then diluted with DMEM until the absorbance at 280 nm equaled that of 10% bovine serum. Protein bound to the column was removed by washing the column with 100 mM glycine (pH 2.5) and collecting 1-mL fractions in tubes that contained 0.1 mL of 0.1 M Tris (pH 8). The desorbed protein was dialyzed against 5 mM ammonium bicarbonate and dried; a portion of the desorbed protein was electrophoresed using a 4–12% polyacrylamide gel, as described [30].
The 10% control serum used in these studies was prepared by the same procedures, with the sole exception being that the control column was prepared by covalently attaching 7 mg of purified rabbit immunoglobulin G (IgG, Sigma) to 5 mL of CNBr-activated Sepharose 4B rather than 7 mg of rabbit anti-bovine fetuin antibody. Aliquots (0.85 mL) of dialyzed adult bovine serum were passed over the control column at room temperature, and the four fractions with the highest absorbance were pooled and diluted with DMEM until the absorbance at 280 nm equaled that of 10% bovine serum.
Results
Removal of Fetuin from Bovine Serum by Antibody Affinity Chromatography
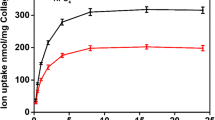
We developed procedures to remove fetuin from bovine serum by antibody affinity chromatography in order to evaluate the possible role of the protein in serum-induced calcification. Rabbits were immunized with purified bovine fetuin, and the resulting antisera were used to construct a radioimmunoassay for bovine fetuin that could be used to monitor the effectiveness of fetuin depletion procedures (Fig. 1). Polyclonal antifetuin antibodies were purified from rabbit antiserum using Sepharose 4B with covalently attached bovine fetuin, and the resulting purified antifetuin antibodies were then attached covalently to Sepharose 4B and packed into a column.

Radioimmunoassay of bovine fetuin and detection of bovine fetuin antigen in adult bovine serum. Relative fraction of 125I-labeled bovine fetuin bound to antibody (B/Bo) at increasing amounts of purified bovine fetuin and at increasing volumes of adult bovine serum
Because the goal of fetuin removal from serum was to test its role in serum-induced calcification, we used a suitable buffer for study of serum-induced calcification [10, 11], DMEM culture medium, to equilibrate the antifetuin antibody column. Adult bovine serum was then dialyzed against DMEM and passed over this column to remove fetuin. The results of a typical experiment are shown in Figure 2. As can be seen, there is a massive peak of unbound serum protein absorbance that elutes at the column volume; this unbound protein peak accounts for about 98% of the A280 applied to the column and is devoid of fetuin. The four fractions with the highest absorbance were pooled; calcification solutions containing fetuin-depleted bovine serum were then prepared by diluting these pooled fractions with DMEM to yield a final serum concentration of 10% by absorbance. The 10% control bovine serum used in these studies was prepared by a similar procedure, with the sole difference being that the control column was prepared by covalently attaching purified normal rabbit IgG to Sepharose rather than rabbit anti-bovine fetuin antibody. Table 1 shows that the fetuin content of the resulting fetuin-depleted 10% bovine serum is over 1,000-fold lower than that of the 10% control bovine serum.

Removal of fetuin from bovine serum by antibody affinity chromatography. In order to prepare fetuin-depleted bovine serum for tests of the role of fetuin in serum-induced mineralization, adult bovine serum was dialyzed against a buffer suitable for calcification (DMEM) and then passed over a column that contained 7 mg of affinity-purified rabbit anti-bovine fetuin antibody attached covalently to 5 mL of Sepharose 4B (see “Experimental Procedures”). Elution buffer, DMEM; fraction volume, ∼0.8 mL; fetuin concentration was determined by radioimmunoassay (Fig. 1). Inset: Fractions 19–24 were pooled, and 10 μg protein from this pool were electrophoresed on a 4–12% SDS polyacrylamide gel and stained with Coomassie brilliant blue. Note that the major band is in the 59 kDa position expected for bovine fetuin [30]
After elution of those proteins that did not bind to the column, the antifetuin antibody column was washed with DMEM until the absorbance at 280 nm was less than 0.01, and bound fetuin was then eluted from the column by washing with acid (Fig. 2). The resulting small peak of A280nm absorbance (not evident in the scale used for Fig. 2) accounted for about 1% of the initial serum absorbance. The amount of fetuin immunoreactivity in this peak corresponded to the fetuin content of the serum applied to the column, and the SDS gel of the bound protein fraction revealed a single major component in the 59-kDa position expected for fetuin [30].
Evidence that Fetuin Is Required for the Serum-Induced Recalcification of Demineralized Bone
In the initial study, the impact of fetuin depletion on serum-induced calcification was evaluated by incubating demineralized newborn rat tibias for 6 days at 37°C in DMEM alone, in DMEM containing 10% control bovine serum, or in DMEM containing 10% fetuin-depleted bovine serum. In agreement with earlier studies [10, 11], demineralized tibias calcified after incubation in DMEM containing 10% serum but did not calcify after incubation in DMEM alone (Figs. 3 and 4). The pattern of alizarin red staining in the tibias incubated in DMEM containing 10% control serum matched that seen in the original tibia prior to demineralization (not shown; see [10, 11] for examples).

Evidence that fetuin is required for the serum-induced recalcification of demineralized bone: analysis for Ca and P. In order to evaluate the possible role of fetuin in the serum-induced recalcification of bone, demineralized newborn rat tibias were separately incubated for 6 days at 37°C in 1 mL DMEM containing 2 mM Pi and no serum, 10% control bovine serum, 10% fetuin-depleted bovine serum, or 10% fetuin depleted bovine serum plus 130 μg/mL of purified bovine fetuin. Tibias were removed, stained with alizarin red, photographed, and then analyzed for calcium and phosphate. The medium and any precipitate were removed from the well and centrifuged to pellet any precipitate, and the pellet fraction was analyzed for calcium and phosphate (see “Experimental Procedures”). This experiment was performed in triplicate. The data show the average calcium and phosphate in the tibia and the pellet fraction from each well; error bars show standard deviations. *Ca or Pi in the extract is < 0.01 μmol

Evidence that fetuin is required for the serum-induced calcification of demineralized bone: alizarin red and von Kossa staining. Demineralized newborn rat tibias were separately incubated for 6 days at 37°C in 1 mL DMEM containing 2 mM Pi and 10% control bovine serum, 10% fetuin-depleted bovine serum, or 10% fetuin-depleted bovine serum containing 130 μg/mL of purified bovine fetuin. After incubation, the tibias were either stained for calcification with alizarin red or fixed in ethanol, cut in 5-μm-thick sections, stained for calcification with von Kossa (stains calcification black), and counterstained with nuclear-fast red
In contrast to tibias incubated in 10% control serum, tibias incubated in 10% fetuin-depleted serum did not have significant incorporation of calcium and phosphate (Fig. 3) and did not stain for calcification by alizarin red (Fig. 4); histological sections of these tibias also revealed no von Kossa staining for calcification (Fig. 4). Removal of fetuin from serum therefore eliminates the serum-induced recalcification of demineralized bone.
To confirm the essential role of fetuin in serum-induced calcification, we added sufficient purified bovine fetuin to the fetuin-depleted bovine serum in order to attain a final fetuin concentration comparable to that found in the original serum prior to fetuin depletion and in the 10% bovine serum control (see Table 1). The calcification of tibias incubated in this fetuin-repleted serum was indistinguishable from the calcification of tibias incubated in the 10% bovine serum control: the pattern of alizarin red staining was identical (Fig. 4), the amount of calcium and phosphate incorporated was comparable (Fig. 3), and the von Kossa staining was restricted to the collagen matrix (Fig. 4). Comparable results were obtained when fetuin purified during the course of the preparation of fetuin-depleted serum (see Fig. 2 inset) was substituted for commercial fetuin (data not shown). The addition of purified fetuin therefore fully restores the ability of fetuin-depleted serum to induce the recalcification of a demineralized tibia.
In the course of these experiments, we noticed the presence of a fine precipitate coating the entire bottom of each culture well that contained a tibia incubated in DMEM plus 10% fetuin-depleted serum (not shown); no precipitate could be detected in wells that contained a tibia incubated in DMEM alone, in wells that contained a tibia incubated in DMEM plus 10% control bovine serum, or in wells that contained a tibia incubated in DMEM plus 10% fetuin-depleted serum supplemented with purified bovine fetuin. To assess the nature of this precipitate, the precipitate was collected, stained with alizarin red, and analyzed for calcium and phosphate. This analysis showed that the precipitate isolated from the wells containing 10% fetuin-depleted serum stained intensely with alizarin red and that the amounts of calcium and phosphate recovered from the precipitate were comparable to the amounts incorporated into tibias that had been incubated in DMEM containing 10% serum or 10% fetuin-repleted serum (Fig. 3). This result suggests that the role of fetuin in the serum-induced recalcification of demineralized bone is to direct mineral formation into the collagen matrix of bone.
In order to determine the dependence of collagen calcification on fetuin dose, we repeated the above experiments using fetuin-depleted serum containing different added fetuin concentrations (data not shown). The results of this experiment showed that tibias incubated with 130 and 100 μg/mL fetuin stained with alizarin red and contained amounts of calcium and phosphate comparable to the values shown in Figure 3, while there was no detectable mineral precipitate outside the tibia. In contrast, tibias incubated with 0, 10, and 40 μg/mL fetuin did not stain with alizarin red and did not contain detectable calcium or phosphate, and there was a mineral precipitate outside the tibia that contained calcium and phosphate comparable to the values shown in Figure 3. The tibia incubated with 70 μg/mL fetuin was stained with alizarin red, and there was also a detectable mineral precipitate outside the tibia; chemical analysis of the tibia and precipitate showed that 73% of the mineral was in the tibia and 27% of the mineral was in the precipitate.
A final experiment was carried out to evaluate the effect of reducing the phosphate concentration of the DMEM from 2 to 0.9 mM [10]. This experiment showed that tibias do not calcify when incubated in DMEM (0.9 mM Pi) containing 10% control bovine serum, 10% fetuin-depleted bovine serum, or 10% fetuin-depleted serum plus added fetuin (not shown). There was also no evidence for a mineral precipitate in any condition. These results demonstrate that the serum-induced formation of a mineral phase in DMEM will not occur unless the phosphate content of the DMEM is at the 2 mM concentration found in bovine serum.
Evidence that Fetuin Is Required for the Serum-Induced Calcification of Tendons and Purified Collagen
Additional experiments were carried out to further explore the role of fetuin in the serum-induced calcification of collagenous matrices. One test examined the role of fetuin in the serum-induced calcification of rat tail tendon, a type I collagen matrix that is chemically identical to the type I collagen matrix of bone but does not normally calcify in rats. Tendons incubated in 10% control bovine serum calcified, tendons incubated in 10% fetuin-depleted serum did not calcify, and tendons incubated in 10% fetuin-depleted serum containing purified fetuin calcified (Fig. 5). There was again a fine precipitate coating the bottom of all wells containing fetuin-depleted serum, and the amount of calcium and phosphate in this precipitate was comparable to that found in tendons incubated in 10% serum that contained fetuin (Fig. 5).

Evidence that fetuin is required for the serum-induced calcification of rat tail tendon. To test the role of fetuin in the serum-induced calcification of tendon, a type I collagen matrix that does not normally calcify, rat tail tendons (dry weight 3 mg) were separately incubated for 6 days at 37°C in 1 mL DMEM containing 2 mM Pi and no serum, 10% control bovine serum, 10% fetuin-depleted bovine serum, or 10% fetuin-depleted bovine serum plus 130 μg/mL of purified bovine fetuin. Tendons were removed, stained with alizarin red, photographed, and then analyzed for calcium and phosphate. The medium and any precipitate were removed from the well and centrifuged to pellet any precipitate, and the pellet fraction was analyzed for calcium and phosphate (see “Experimental Procedures”). This experiment was performed in triplicate. The data show average calcium and phosphate in the tendons and the pellet fraction from each well; error bars show standard deviations. *Ca or Pi in the extract is < 0.01 μmol
Another test examined the role of fetuin in the serum-induced calcification of purified type I collagen fibers from bovine Achilles tendon. Purified collagen fibers incubated in 10% control bovine serum calcified, fibers incubated in 10% fetuin-depleted serum did not calcify, and fibers incubated in 10% fetuin-depleted serum containing purified fetuin calcified (Fig. 6). There was a fine precipitate coating the entire bottom of all wells containing fetuin-depleted serum, and the amount of calcium and phosphate in this precipitate was comparable to that found in collagen fibers incubated in 10% serum containing fetuin (Fig. 6).

Evidence that fetuin is required for the serum-induced calcification of purified bovine type I collagen. To assess the role of fetuin in the serum-induced calcification of collagen fibers, 3 mg of purified bovine type I collagen were separately incubated for 6 days at 37°C in 1 mL DMEM containing 2 mM Pi and no serum, 10% control bovine serum, 10% fetuin-depleted bovine serum, or 10% fetuin-depleted bovine serum plus 130 μg/mL of purified bovine fetuin. Collagen fibers were removed, stained with alizarin red, photographed, and then analyzed for calcium and phosphate. The medium and any precipitate were removed from the well and centrifuged to pellet any precipitate, and the pellet fraction was analyzed for calcium and phosphate (see “Experimental Procedures”). This experiment was performed in triplicate. The data show average calcium and phosphate in purified collagen and the pellet fraction from each well; error bars show standard deviations. *Ca or Pi in the extract is <0.01 μmol
The Ca/Pi ratio was calculated for the mineral phase formed in each of the above experiments. The Ca/Pi ratio of the mineral phase formed within a collagen matrix after incubation in 10% serum was 1.59 ± 0.15 (mean ± SD, n = 9; combined data for Figs. 3, 5, and 6), the Ca/Pi ratio for the mineral phase precipitated outside of a matrix after incubation in 10% fetuin-depleted serum was 1.56 ± 0.09 (n = 9), and the Ca/Pi ratio for the mineral phase formed within collagen after incubation in 10% fetuin-depleted serum with added fetuin was 1.58 ± 0.10 (n = 9). These Ca/Pi ratios are not significantly different from one another and are comparable to the Ca/Pi ratio previously found for the mineral phase deposited in collagen after incubation in serum [10, 13, 31] and the ratio found in bone [32, 33].
Taken together, these results show that fetuin plays a similar essential role in the serum-induced calcification of the type I collagen fibers in a tissue that was once calcified (demineralized bone), a tissue that does not normally calcify (tendon), and purified collagen. In each case, the essential role of fetuin in the serum-induced calcification is to direct mineral formation into the collagen matrix, and it appears to do this by preventing mineral precipitation outside of this matrix.
Evidence that Removal of Fetuin from Serum Unmasks a Potent Serum Nucleator of Mineral Formation
In each of the above experiments, removal of fetuin from serum prevented calcification of the collagen matrix but led to the formation of a fine precipitate of calcium phosphate mineral on the bottom of the well. In order to see if the formation of this precipitate is dependent on the presence of a matrix, this experiment was repeated using the same calcification solutions but no matrix. A fine precipitate coated the entire bottom of all wells that contained DMEM plus 10% fetuin-depleted serum, while no precipitate could be detected in wells that contained DMEM alone, DMEM plus 10% control bovine serum, or DMEM with 10% fetuin-depleted serum plus added purified fetuin. This precipitate stained intensely with alizarin red, and chemical analysis showed that it contained calcium and phosphate (Fig. 7) in amounts comparable to those previously seen in wells that contained fetuin-depleted serum and a collagen matrix. This result demonstrates that the formation of a precipitate in DMEM containing 10% fetuin-depleted serum is not dependent on the presence of a collagen matrix. The removal of fetuin from serum therefore appears to unmask a potent serum initiator of calcium phosphate mineral formation.

Evidence that fetuin depletion unmasks a potent serum initiator of mineral formation. To determine whether a calcium phosphate mineral phase forms spontaneously in fetuin-depleted serum even in the absence of a collagen matrix, the following solutions were prepared that contained 1 mL DMEM with 2 mM Pi and no serum, 10% control bovine serum, 10% fetuin-depleted bovine serum, or 10% fetuin-depleted bovine serum containing 130 μg/mL of purified bovine fetuin. The solutions were incubated for 6 days at 37°C in the absence of a collagen matrix. The medium and any precipitate were removed from the well and centrifuged to pellet any precipitate, and the pellet fraction was analyzed for calcium and phosphate (see “Experimental Procedures” for details). This experiment was performed in triplicate. The data show average calcium and phosphate in the pellet fraction from each well; error bars show standard deviations. *Ca or Pi in the extract is <0.01 μmol
Powder X-ray diffraction was used to characterize the mineral that forms during incubation of DMEM containing fetuin-depleted serum. As can be seen in Figure 8, the diffraction spectrum of this mineral is comparable to the spectrum of the apatite-like crystals isolated from rat bone. Both diffraction spectra are also comparable to the spectrum previously found for the mineral phase formed in a type I collagen matrix during incubation in DMEM containing fetuin-replete serum [10, 31]. The diffraction peaks seen in these spectra are in the positions expected for synthetic hydroxyapatite crystals, with no evidence for the presence of other calcium phosphate mineral phases [33]. The diffraction peaks are far broader than observed for synthetic hydroxyapatite crystals. For bone, this peak broadening has been attributed to smaller crystal size and/or reduced crystallinity [34, 35]. Because the diffraction peaks for the crystals generated in fetuin-depleted serum appear to be slightly broader than the peaks for bone crystals, it is possible that the crystals generated in serum may be smaller or less ordered than those found in bone.

The powder X-ray diffraction spectrum of the mineral formed in fetuin-depleted serum is comparable to the spectrum of bone mineral. Serum-induced mineral was generated by incubating DMEM containing 10% fetuin-depleted serum at 37°C (see “Experimental Procedures”), and bone crystals were prepared as described [29]. The X-ray diffraction spectrum of both powders was determined with a Rigaku Miniflex diffractometer
Discussion
The present investigation and our recently published study were both carried out with the goal of understanding the biochemical basis for the ability of serum to induce the calcification of a type I collagen fibril. The published study demonstrates that the physical structure of the collagen fibril is such that molecules smaller than a 6-kDa protein can freely access all of the water within the fibril, while molecules larger than a 40-kDa protein cannot enter the fibril. This study therefore shows that molecules smaller than a 6-kDa protein can enter the fibril and interact directly with mineral to influence crystal growth, while molecules larger than a 40-kDa protein cannot enter the fibril and so have no ability to act directly on the apatite crystals growing within the fibril.
The serum calcification activity that induces calcification of the collagen fibril consists of one or more proteins that are 50–150 kDa in molecular weight. Since these molecules are too large to penetrate the collagen fibril, there must be mechanisms by which proteins that act only outside the fibril can cause calcification to occur specifically within the fibril. One possibility is that large inhibitors of apatite growth favor mineralization within the fibril by selectively preventing apatite growth outside of the fibril. In addition, large nucleators of apatite formation may generate small crystal nuclei outside of the collagen fibril that subsequently diffuse into the fibril and grow. The present study tests these hypotheses for the possible function of large molecules in mineralization.
Our working hypothesis was that the serum protein fetuin promotes calcification within the collagen fibril by selectively inhibiting apatite growth outside of the fibril, and we tested this hypothesis by examining the impact of removing fetuin from serum on the ability of serum to mineralize the collagen fibril. The results of this study reveal that removing fetuin from serum completely prevents the serum-driven calcification of a type I collagen matrix. Removing fetuin from serum does not prevent the serum-driven formation of mineral, however, because a comparable amount of apatite-like mineral consistently forms on the bottom of all wells that contain fetuin-depleted serum (Fig. 8). The results of these experiments therefore support our working hypothesis, that large protein inhibitors of apatite growth such as fetuin can favor mineralization of the collagen fibril by selectively preventing apatite growth outside of the fibril. The net effect of this fetuin activity is extraordinary: all of the calcium and phosphate ions that, in the absence of fetuin, are incorporated into a mineral that forms throughout the ∼1-mLvolume that lies outside the fibril are, in the presence of fetuin, incorporated into a mineral that forms within the ∼5-μL volume of water that lies within the 3 mg collagen in the well.
Previous in vitro studies using pure fetuin in solutions containing high levels of calcium and phosphate provide insight into how fetuin may act to direct apatite growth within the collagen fiber. In these experiments, solutions were prepared that substantially exceed the calcium phosphate ion product required for homogeneous formation of an apatite-like mineral phase, and in the absence of fetuin a mineral phase forms in minutes [25]. When fetuin is added to these solutions, no mineral phase precipitates, no mineral phase can be sedimented by high-speed centrifugation, and the solution remains clear for about 24 hours. At this time the solution becomes opalescent and a fetuin–mineral complex can, for the first time, be sedimented from the solution by centrifugation [25]. Measurement of ionic calcium and phosphate levels during the first 24 hours further shows that small amounts of a mineral phase still form in the presence of fetuin and that the role of fetuin is to form a complex with these nascent mineral nuclei that retards their growth and prevents their precipitation (or sedimentation in a centrifuge) [25]. Purified fetuin therefore does not prevent mineral nuclei from forming in this homogeneous nucleation system. It traps the nascent mineral nuclei and dramatically retards their growth.
We believe that the role of fetuin in serum-driven calcification of a type I collagen matrix is similar to its action on a homogeneous apatite nucleation system: fetuin traps mineral nuclei and retards their growth. The major difference is that mineral nuclei are generated by the serum nucleator activity, not by a high calcium phosphate ion product. The serum nucleator elutes from a gel filtration column in the position expected for proteins 50–150 kDa in size and is therefore clearly too large to physically penetrate the collagen fibril. The products of nucleator action outside the fibril are presumably small crystal nuclei however, and even apatite crystals up to 12 unit cells in size should in principle be able to freely access all of the water within the fibril (see “Introduction”). Since fetuin can only trap those nuclei that it can access, the crystal nuclei that penetrate the fibril are free to grow far more rapidly than those nuclei trapped by fetuin outside of the fibril, and the collagen fibril therefore selectively calcifies. When fetuin is removed from serum, the same number of mineral nuclei still form, and some of these no doubt still penetrate the fibril. All crystal nuclei are now free to grow however. Because the vast majority of the nuclei are in the solution outside of the fiber, the only mineral formed in amounts that can be detected is the mineral precipitate found on the bottom of the well, not mineral within the fibril.
The phenotype of the fetuin-deficient mouse is consistent with the effects of fetuin depletion on serum found in the present study. Fetuin knockout mice have multiple calcium phosphate mineral deposits in a variety of soft tissues, particularly those involved in the transport or filtration of blood; these deposits are not within collagen fibrils [14, 36, 37]. Our results demonstrate that removal of fetuin from serum results in the formation of calcium phosphate crystals throughout serum and the absence of mineral formation within collagen. The close parallel between the effects of fetuin depletion in vivo and in vitro suggests that the serum nucleator of mineral formation unmasked by fetuin depletion in vitro may be responsible for the formation of the soft tissue mineral deposits seen in the fetuin knockout mouse.
Summary and Conclusion: A Hypothesis for the Mechanism of Normal Bone Mineralization
The present study was carried out to understand the mechanism by which a serum calcification factor activity consisting of proteins 50–150 kDa in size is able to drive the calcification of a collagen fibril. The results of this study show that serum calcification factor activity consists of at least two large proteins, neither of which can penetrate the collagen fibril. One as yet unidentified protein generates crystal nuclei outside of the fibril, some of which then diffuse into the fibril. The other protein, fetuin, inhibits the growth of crystal nuclei that remain in the solution outside of the fibril, thereby freeing calcium and phosphate ions for crystal growth within the fibril. We propose the term “shotgun mineralization” for this calcification mechanism: Crystals form throughout the solution, and only those that diffuse into a mineralizable matrix grow.
It is possible that mineralization of the collagen fibril occurs by a similar mechanism in vivo. Nucleators too large to penetrate the fibril may generate small crystals near the mineralization front, some of which penetrate the fibril; and large crystal growth inhibitors may bind to crystals that remain in the solution outside of the fibril, thereby ensuring that only crystals within the fibril can grow. As with many other critical processes in biochemical physiology, there are probably multiple layers of redundancy in the process of normal bone mineralization. Bone is known to contain a number of large inhibitors of apatite crystal growth in addition to fetuin, a redundancy in function that could account for the apparently normal calcification of the collagen fibril in the fetuin knockout mouse [14]. In addition to the serum nucleator activity, nucleators may include large proteins such as bone sialoprotein [38, 39] as well as large structures such as matrix vesicles [40].
Future studies will be needed to further assess the relationship between the mechanism of serum-induced collagen calcification in vitro and the mechanism of bone mineralization. The fetuin-depleted serum assay developed here can be used to search for other bone macromolecules that, when added to fetuin-deficient serum, restore the serum-driven calcification of the collagen fibril and prevent the growth and precipitation of mineral outside of the fibril. DMEM plus purified fetuin can be used as a test system to evaluate the ability of different bone macromolecules to generate crystal nuclei outside of the fibril that are small enough to penetrate the fibril and grow. Other studies will be needed to determine whether the initial serum-induced mineral forms within the hole region of the collagen fibril (the location of initial crystal formation in vivo), to compare the size and shape of the crystals within the fibril with the crystals found in normal bone, and to see if the mechanical properties of demineralized bone that has been fully recalcified by incubation in serum [31] are comparable to those of the original bone prior to demineralization.
References
Tong W, Glimcher MJ, Katz JL, Kuhn L, Eppell SJ (2003) Size and shape of mineralites in young bovine bone measured by atomic force microscopy. Calcif Tissue Int 72:592–598
Katz EP, Li S-T (1973) Structure and function of bone collagen fibrils. J Mol Biol 1973:1–15
Sasaki N, Sudoh Y (1996) X-ray pole analysis of apatite crystals and collagen molecules in bone. Calcif Tissue Int 60:361–367
Jager I, Fratzl P (2000) Mineralized Collagen fibrils: a mechanical model with a staggered arrangement of mineral particles. Biophys J 79:1737–1748
Landis WJ, Song MJ, Leith A, McEwen L, McEwen BF (1993) Mineral and organic matrix interaction in normally calcifying tendon visualized in three dimensions by high-voltage electron microscopic tomography and graphic image reconstruction. J Struct Biol 110:39–54
Rubin MA, Jasiuk I, Taylor J, Rubin J, Ganey T, Apkarian RP (2003) TEM analysis of the nanostructure of normal and osteoporotic human trabecular bone. Bone 33:270–282
Robinson RA, Elliott SR (1957) The water content of bone. The mass of water, inorganic crystals, organic matrix, and “CO2 space” components in a unit volume of dog bone. J Bone Joint Surg Am 39:167–188
Boivin G, Meunier P (2002) The degree of mineralization of bone tissue measured by computerized quantitative contact microtomography. Calcif Tissue Int 70:503–511
Toroian D, Lim JE, Price PA (2007) The size exclusion characteristics of type I collagen: implications for the role of non-collagenous bone constituents in mineralization. J Biol Chem 282:22437–22447
Hamlin NJ, Price PA (2004) Mineralization of decalcified bone occurs under cell culture conditions and requires bovine serum but not cells. Calcif Tissue Int 75:231–242
Price PA, June HH, Hamlin NJ, Williamson MK (2004) Evidence for a serum factor that initiates the recalcification of demineralized bone. J Biol Chem 279:19169–19180
Parfitt AM (2000) The mechanism of coupling: a role for the vasculature. Bone 26:319–323
Hamlin NJ, Ong KG, Price PA (2006) A serum factor that recalcifies demineralized bone is conserved in bony fish and sharks but is not found in invertebrates. Calcif Tissue Int 76:326–334
Jahnen-Dechent W, Schinke T, Trindl A, Muller-Esterl W, Sablitzky F, Kaiser S, Blessing M (1997) Cloning and targeted deletion of the mouse fetuin gene. J Biol Chem 272:31496–31503
Schinke T, Amendt C, Trindl A, Poschke O, Muller-Esterl W, Jahnen-Dechent W (1996) The serum protein alpha 2-HS glycoprotein/fetuin inhibits apatite formation in vitro and in mineralizing calvaria cells. J Biol Chem 271:20789–20796
Pedersen KO (1944) Fetuin, a new globulin isolated from serum. Nature 154:575–580
Brown WM, Saunders NR, Mollgard K, Dziegielewska KM (1992) Fetuin—an old friend revisited. Bioessays 14:749–755
Ashton BA, Triffitt JT, Herring GM (1974) Isolation and partial characterization of a glycoprotein from bovine cortical bone. Eur J Biochem 45:525–533
Ashton BA, Hohling HJ, Triffitt JT (1976) Plasma proteins present in human cortical bone: enrichment of the alpha-2HS-glycoprotein. Calcif Tissue Res 22:27–33
Quelch KJ, Cole WG, Melick RA (1984) Noncollagenous proteins in normal and pathological human bone. Calcif Tissue Int 36:545–549
Mizuno M, Farach-Carson MC, Pinero GJ, Fujisawa R, Brunn JC, Seyer JM, Bousfield GR, Mark MP, Butler WT (1991) Identification of the rat bone 60K acidic glycoprotein as alpha 2HS-glycoprotein. Bone Miner 13:1–21
Ohnishi T, Arakaki N, Nakamura O, Hirono S, Daikuhara Y (1991) Purification, characterization, and studies on biosynthesis of a 59-kDa bone sialic acid-containing protein (BSP) from rat mandible using a monoclonal antibody. J Biol Chem 266:14636–14645
Wendel M, Heinegard D, Franzen A (1993) A major non-collagenous 62 kDa protein from rat bone mineralized matrix is identical to pp 63, a phosphorylated glycoprotein from liver. Matrix 13:331–339
Dickson IR, Poole AR, Veis A (1975) Localization of plasma alpha-2-HS glycoprotein in mineralizing human bone. Nature 256:430–432
Price PA, Lim JE (2003) The inhibition of calcium phosphate precipitation by fetuin is accompanied by the formation of a fetuin–mineral complex. J Biol Chem 278:22144–22152
Price PA, Chan WS, Jolson DM, Williamson MK (2006) The elastic lamellae of devitalized arteries calcify when incubated in serum. Evidence for a serum calcification factor. Arterioscler Thromb Vasc Biol 26:1079–1085
Price PA, Roublick AM, Williamson MK (2006) Artery calcification in uremic rats is increased by a low protein diet and prevented by treatment with ibandronate. Kidney Int doi:10.1038/sj.ki.5001841
Chen PS, Toribara TY, Warner H (1956) Microdetermination of phosphorus. Anal Chem 28:1756–1758
Weiner S, Price PA (1986) Disaggregation of bone into crystals. Calcif Tissue Int 39:365–375
Price PA, Nguyen TMT, Williamson MK (2003) Biochemical characterization of the serum fetuin–mineral complex. J Biol Chem 278:22153–22160
Price PA, June HH, Williamson MK (2003) Evidence for a serum nucleator that initiates the calcification of bone. J Biol Chem
Driessens FCM, Verbeeck RMH (1990) Biominerals. CRC Press, Boca Raton, FL
Elliott JC (1994) Structure and chemistry of the apatites and other calcium orthophosphates. Elsevier, Amsterdam
Bonar L, Roufosse A, WK S, Grynpas M, Glimcher M (1983) X-ray diffraction studies of the crystallinity of bone mineral in newly synthesized and density fractionated bone. Calcif Tissue Int 35:202–209
Meneghini C, Dalconi M, Nuzzo S, Mobilio S, Wenk R (2003) Rietveld refinement on X-ray diffraction patterns of bioapatite in human fetal bones. Biophys J 84:2021–2029
Schafer C, Heiss A, Schwarz A, Westenfeld R, Ketteler M, Floege J, Muller-Esterl W, Schinke T, Jahnen-Dechent W (2003) The serum protein alpha 2-Heremans-Schmid glycoprotein/fetuin-A is a systemically acting inhibitor of ectopic calcification. J Clin Invest 112:357–366
Westenfeld R, Schafer C, Smeets R, Brandeburg V, Floege J, Ketteler M, Jahnen-Dechent W (2007) Fetuin-A (AHSG) prevents extraosseous calcification induced by uraemia and phosphate challenge in mice. Nephrol Dial Transplant doi:10.1093/ndt/gfm094
Tye CE, Rattray KR, Warner KJ, Gordon JAR, Sodek J, Hunter GK, Goldberg HA (2003) Delineation of the hydroxyapatite-nucleating domains of bone sialoprotein. J Biol Chem 278:7949–7955
Midura R, Wang A, Lovitch D, Law D, Powell K, Gorski J (2004) Bone acidic glycoprotein-75 delineates the extracellular sites of future bone sialoprotein accumulation and apatite nucleation in osteoblastic cultures. J Biol Chem 279:25464–25473
Anderson HC (1995) Molecular biology of matrix vesicles. Clin Orthop Relat Res 314:266–280
Acknowledgement
We thank Gustaf Arrhenius for assistance with the X-ray diffraction analysis. This work was supported in part by grant HL58090 from the National Heart, Lung, and Blood Institute of the National Institutes of Health.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Toroian, D., Price, P.A. The Essential Role of Fetuin in the Serum-Induced Calcification of Collagen. Calcif Tissue Int 82, 116–126 (2008). https://doi.org/10.1007/s00223-007-9085-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00223-007-9085-2