
Summary
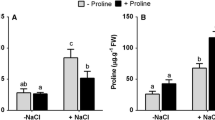
The interaction of salinity and nitrogen availability on the growth and accumulation of proline and glycinebetaine by Spartina alterniflora Loisel. grown in hydroponic solution in controlled environements was investigated. Growth of shoots was reduced by increasing salinity and enhanced at higher nitrogen levels. Proline was accumulated when a threshold salinity was reached. At higher nitrogen levels, the threshold salinity to induce proline accumulation was lowered and the amount of proline accumulated was enhanced. Glycinebetaine content was higher in plants grown at higher nitrogen levels, but was less affected by salinity and nitrogen levels than proline. Glycinebetaine contents were approximately 10 times higher than proline contents. When plants that were non-stressed for 1 month were exposed to salinity stress, proline and glycinebetaine increased immediately and reached a plateau in 1–2 days. After the removal of salinity stress, proline disappeared rapidly but glycinebetaine content remained unchanged. Unlike NaCl, neither proline nor glycinebetaine had a significant effect on the in vitro activity of malate dehydrogenase activity from leaves.
Article PDF
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
References
Barbour MG (1978) The effect of competition and salinity on the growth of a salt marsh plant species. Oecologia (Berl) 37:93–99
Cavalieri AJ, Huang AHC (1977) Effect of NaCl on the in vitro activity of malate dehydrogenase in salt marsh halophytes of the U.S. Physiol Plant 41:79–84
Cavalieri AJ, Huang AHC (1979) Evaluation of proline accumulation in the adaptation of diverse species of marsh halophytes to the saline environment. Am J Bot 66:307–312
Cavalieri AJ, Huang AHC (1981) Accumulation of proline and glycinebetaine in Spartina alterniflora Loisel. in response to NaCl and nitrogen in the marsh. Oecologia 49:224–228
Flowers TJ, Troke BJ, Yeo AR (1977) The mechanism of salt tolerance in halophytes. Ann Rev Plant Physiol 28:89–121
Gallagher JL (1975) Effect of an ammonium nitrate pulse on the growth and elemental composition of natural stands of Spartina alterniflora and Juncus roemerianus. Am J Bot 62:644–648
Gorham J, Hughes LL, Wyn Jones RG (1980) Chemical composition of salt-marsh plants from Ynys Mon (Anglesey): the concept of physiotypes. Plant. Cell and Environ 3:309–318
Haines BL, Dunn EL (1976) Growth and resource allocation responses of Spartina alterniflora Loisel. to three levels of NH4−N, Fe and NaCl in solution culture. Bot Gaz 137:224–230
Hall JL, Harvey DMR, Flowers TJ (1978) Evidence for the cytoplasmic localization of betaine in leaf cells of Suaeda maritima. Planta 140:59–62
Hanson AD, Nelsen CE (1978) Betaine accumulation and 14C-formate metabolism in water stressed barley leaves. Plant Physiol 62:305–312
Jefferies RL, Davy AJ, Rudmik T (1979) The growth strategies of coastal halophytes. In: Jefferies RL and Davy AJ (eds), Ecological Processes in Coastal Environments. Blackwell Scientific Publications. London. 232–268
Keefe CW (1972) Marsh Production: A summary of the literature. Contributions in Mar Sci 16:163–181
Longstreth DJ, Strain BR (1977) Effects of salinity and illumination on photosynthesis and water balance of Spartina alterniflora Loisel. Oecologia (Berl.) 31:191–199
Mendelssohn IA (1979) Nitrogen metabolism in the height forms of Spartina alterniflora in North Carolina. Ecology 60:574–584
Parrondo RT, Gosselink JG, Hopkinson CS (1978) Effects of salinity and drainage on the growth of three salt marsh grasses. Bot Gaz 139:120–127
Pearce RB, Strange RN, Smith H (1976) Glycinebetaine and choline in wheat: Distribution in relation to infection by Fusarium graminearum. Phytochem 15:953–954
Pollard A, Wyn Jones RB (1979) Enzyme activities in concentrated solutions of glycinebetaine and other solutes. Planta 144:291–298
Rozema J, Buizer DAG, Fabritius HE (1978) Population dynamics of Glaux maritima and ecophysiological adaptations to salinity and inundation. Oikos 30:539–548
Singh TN, Paleg LG, Aspinall D (1973) Stress metabolism. 1. Nitrogen metabolism and growth in the barley plant during water stress. Aust J Biol Sci 26:45–56
Stewart GR, Lee JS (1974) The role of proline accumulation in halophytes. Planta 120:279–289
Stewart GR, Larher F, Ahmad I, Lee JA (1979) Nitrogen metabolism and salt-tolerance in higher plant halophytes. In: Jefferies RJ, Davy AJ (eds) Ecological Processes in Coastal Environments. Blackwell Scientific Publications. London 211–227
Storey R, Ahmad N, Wyn Jones RG (1977) Taxonomic and ecological aspects of the distribution of glycinebetaine and related compounds in plants. Oecologia (Berl.) 27:319–332
Szwarebaum I, Waisel Y (1973) Inter-relationships between halophytes and glycophytes grown on saline and non-saline media. J Ecol 61:775–786
Turner RE (1976) Geographic variations in salt marsh macrophyte production: A review. Contributions in Mar Sci 20:47–68
Valiela I, Teal JM, Deuser WF (1978) The nature of growth forms in the salt marsh grass Spartina alterniflora. Am Natl 112:461–470
Wyn Jones RG, Storey R, Leigh RA, Ahmad N, Pollard A (1977) A hypothesis on cytoplasmic osmoregulation. In: Marre R, Cifferi O (eds) Regulation of Cell Membrane Activities in Plants. North Holland, Amsterdam, 122–136
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Cavalieri, A.J. Proline and glycinebetaine accumulation by Spartina alterniflora Loisel. in response to NaCl and nitrogen in a controlled environment. Oecologia 57, 20–24 (1983). https://doi.org/10.1007/BF00379556
Received:
Issue Date:
DOI: https://doi.org/10.1007/BF00379556