
Summary
An electrophysiological method for investigating a dorsally exposed fish brain was worked out and described.
Electrical response to acoustic stimuli were picked up from 13 points in the vicinity of the midbrain and medulla. The points were brought into histological relationship with nervous structures (Figs. 3, 4). No acoustic responses were found in the diencephalon or telencephalon.
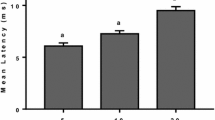
The various acoustic areas differ in their latencies. At optimal stimulus intensity the shortest latency observed was 0.9–1.2 msec (acoustic nerve) and the longest, 3.5–4.3 msec (midbrain).
Common to all areas with auditory activity is the dependence of the excitation pattern upon stimulus frequency. The pattern shows synchronization, constant phase relationship, and doubled frequency. Switching off a loudspeaker at frequencies between 900 and 1700 cps is answered with an off-effect in all areas except “n” of the midbrain. A decrease in excitation is found in the acoustic nerve from 900 cps upwards, whereas the decrease in the midbrain and medulla appears at frequencies as low as 500 cps (Figs. 5, 7, 8).
Excitation is triggered at the same phase angle for all frequencies. Nothing can be said concerning the size of the phase angle (Fig. 10).
All acoustically active areas except for “d” in the medulla show the same or very similar sensitivity to the frequencies within the range (Figs. 11, 12). There was no indication of a topographical separation of frequencies in the brain of the tench.
The latency and amplitude of the potentials is a function of the stimulus intensity. The latency shows the same dependence on intensity in all areas, whereas the dependence of amplitude on intensity is less pronounced in the midbrain than in the medulla or acoustic nerve (Figs. 13, 14).
The synchronization, constant phase relationship and correlation of amplitude of excitation potentials to differences in intensity diminish with progress upwards along the auditory pathway. These results suggest that processing of auditory input is not so advanced in the fish brain as in the brains of higher vertebrates.
Zusammenfassung
-
1.
Zur elektrophysiologischen Untersuchung des von dorsal eröffneten Fischgehirns wurde eine Methode erarbeitet und beschrieben.
-
2.
Im Bereich der Medulla und des Mittelhirns wurden an 13 Orten elektrische Antworten auf akustische Reize abgeleitet und histologisch auf nervöse Strukturen bezogen. Im Zwischen- und Vorderhirn konnten keine akustischen Antworten festgestellt werden.
-
3.
Die verschiedenen Bereiche unterscheiden sich durch die Latenzzeiten. Bei optimaler Reizstärke beträgt die kürzeste beobachtete Latenzzeit 0,9–1,2 msec (Hörnerv), die längste 3,5–4,3 msec (Mittelhirn).
-
4.
Allen akustisch aktivierbaren Bereichen ist gemeinsam, daß das abgeleitete Erregungsmuster reizsynchron, phasengebunden und mit der doppelten Reizfrequenz verläuft. Das Ausschalten des Lautsprechers wird für die Frequenzen 900–1700 Hz mit einem Schlußeffekt beantwortet (mit Ausnahme von Bereich „n“ des Mittelhirns). Während im Hörnerv erst ab 900 Hz ein Erregungsabfall gefunden wird, tritt er in Medulla und Mittelhirn schon ab 500 Hz auf.
-
5.
Es wird gezeigt, daß die Erregung für alle Frequenzen beim selben Phasenwinkel des Reizes ausgelöst wird. Über die Größe dieses Phasenwinkels kann keine Aussage gemacht werden.
-
6.
Mit Ausnahme von Bereich „d“ der Medulla, zeigen alle gefundenen akustisch aktivierbaren Bereiche die gleiche oder sehr ähnliche Frequenzempfindlichkeit. Hinweise auf eine topographische Trennung der Frequenzen im Gehirn der Schleie wurden nicht gefunden.
-
7.
Latenzzeit und Potentialhöhe sind Funktionen der Intensität. Die Intensitätsfunktion der Latenzzeit zeigt in allen Bereichen gleichartiges Verhalten; die Intensitätsfunktion der Erregungshöhe hingegen läßt erkennen, daß die Abhängigkeit der Amplitudenhöhe von der Reizstärke im Mittelhirn nicht so ausgeprägt ist wie in Medulla und Hörnerv.
-
8.
Synchronisation, Phasenbindung und das Ansprechen der Potentialhöhe auf Intensitätsunterschiede werden mit aufsteigender Hörbahn schlechter. Die Diskussion dieser Erscheinungen führt zu der Annahme, daß die Erregungsverarbeitung im Fischgehirn hinter derjenigen bei höheren Wirbeltieren zurücksteht.
Article PDF
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Literatur
Burkhardt, D.: Die Eigenschaften und Funktionstypen der Sinnesorgane. Ergebn. Biol. 22, 226–267 (1960).
—: Allgemeine Sinnesphysiologie und Elektrophysiologie der Receptoren. Fortsohr. Zool. 13, 146–189 (1961).
Dijkgraaf, S., u. F. J. Verheijen: Neue Versuche über das Tonunterscheidungsvermögen der Elritze. Z. vergl. Physiol. 32, 248–256 (1950).
Enger, P. S.: Single unit activity in the peripheral system of a teleost fish. Acta physiol. scand. 58, Suppl. 210 1–48 (1963).
Flock, A.: Electron microscopic and electrophysiological studies on the lateral line canal organ. Acta oto-laryng. (Stockh.) Suppl. 199, 1–90 (1965).
—, and J. Wersäll: A study of the orientation of the sensory hairs of the receptor cells in the lateral line organ of fish, with special reference to the function of the receptors. J. Cell Biol. 15, 19–27 (1962).
Furukawa, T.: Excitation and inhibition in the Mauthner cell. The US-Japan Scientific Cooperation Program, Neurophysiol. Symp. (1964).
Galambos, R.: Microelectrode studies on medial geniculate body of cat. III. Response to pure tones. J. Neurophysiol. 15, 381–400 (1952).
Hubel, D. H.: Tungsten microelectrode for recording from single units. Sci. 125, 549–550 (1957).
Jielof, R., A. Spoor, and Hl. de Vries: The microphonic activity of the lateral line. J. Physiol. (Lond.) 116, 137–157 (1952).
Kappers, A.: The comparative anatomy of the nervous system of vertebrates, including man (1936); Nachdruck. New York: Hafner 1960.
Katsuki, Y.: Comparative neurophysiology of hearing. Physiol. Rev. 45, 380–424 (1965).
Kuiper, J. W.: The microphonic effect of the lateral line organ, 159 p. Groningen: Publ. Natuurk. Lab. 1956.
Liklider, J. C. R.: Three auditory theories; in: Psychology: a study of science (ed. Koch), p. 41–144. New York: McGraw-Hill Book Co. 1959.
Lowenstein, O., M. P. Osborne, and J. Wersäll: Structure and innervation of the sensory epithelia of the labyrinth in the Thornback ray (Raja clavata). Proc. roy. Soc. B 160, 1–12 (1964).
Mayser, P.: Vergleichend anatomische Studien über das Gehirn der Knochenfische mit besonderer Berücksichtigung der Cyprinoiden. Z. wiss. Zool. 36, 259–364 (1882).
Pearson, A.: The acustico-lateral nervous system in fishes. J. comp. Neurol. 64, 235–273 (1936).
Poggendorf, D.: Die absoluten Hörschwellen des Zwergwelses (Amiurus nebulosus) und Beiträge zur Physik des Weberschen Apparates der Ostariophysen. Z. vergl. Physiol. 34, 222–257 (1952).
Potter, H. D.: Mesencephalic auditory region of the bullfrog. J. Neurophysiol. 28, 1132–1154 (1965).
—: Patterns of acoustically evoked discharges of neurons in the mesencephalon of the bullfrog. J. Neurophysiol. 28, 1155–1184 (1965).
Schwartzkopff, J.: Die Übertragung akustischer Information durch Nerventätigkeit nach dem Salvenprinzip; in: Aufnahme und Verarbeitung von Nachrichten durch Organismen (Hrsg. Nachrichtentechn. Ges. im VDE), S. 87–99. Stuttgart: S. Hirzel 1961.
Suzuki, N.: A contribution to the neurobiological study on the acustico-lateral area in the teleostean brain (Mugil cephalus). J. med. Sci. 11, 1–9 (1962).
Takeuchi, A.: Neuromuscular transmission of fish skeletal muscles investigated with intracellular microelectrode. J. cell. comp. Physiol. 54, 211–220 (1959).
Tavolga, W. N., and J. Wodinsky: Auditory capacities in fishes. Pure tone thresholds in nine species of marine teleosts. Bull. Amer. Mus. Nat. Hist. New York 126, Art. 2 (1963).
Wallenberg, A.: Beiträge zur Kenntnis des Gehirns der Teleostier und Selachier. Anat. Anz. 31, 369–399 (1907).
Wever, E. G.: Theory of hearing; 484 p. New York and London: Wiley, Chapman & Hall 1949.
Whitfield, I. C., and E. F. Evans: Responses of auditory cortical neurons to stimuli of changing frequency. J. Neurophysiol. 28, 655–672 (1965).
Wodinsky, J., and W. N. Tavolga: Sound detection in teleost fishes. Marine Bio-Acoust. Proc. Symp. Bimini, Bahamas p. 269–280 (1964).
Wohlfahrt, Th. A.: Untersuchungen über das Tonunterscheidungsvermögen der Elritze. Z. vergl. Physiol. 26, 570–604 (1939).
Zotterman, Y.: The microphonic effect of teleost labyrinths and its biological significance. J. Physiol. (Lond.) 102, 313–318 (1943).
Author information
Authors and Affiliations
Additional information
Dissertation der Naturwissenschaftlichen Fakultät der Universität München.
Herrn Prof. Schwartzkopff sage ich verbindlichen Dank für die ständige Unterstützung dieser Arbeit. Die Versuche wurden zum Teil mit Geräten durchgeführt, die die Deutsche Forschungsgemeinschaft zur Verfügung stellte.
Rights and permissions
About this article
Cite this article
Grözinger, B. Elektro-physiologische Untersuchungen an der Hörbahn der Schleie (Tinca tinca [L.]). Z. Vergl. Physiol. 57, 44–76 (1967). https://doi.org/10.1007/BF00298222
Received:
Issue Date:
DOI: https://doi.org/10.1007/BF00298222