
Abstract
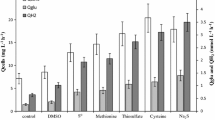
Thauera selenatis was grown anaerobically in minimal medium with either selenate or nitrate as the terminal electron acceptor and acetate as the carbon source and electron donor. The molar cell protein yields, YM-protein (selenate) and YM-protein (nitrate), were found to be 7.8 g cell protein/mol selenite formed and 7.5 g cell protein/mol nitrite formed, respectively. These values represent YM values of 57 and 55 g (dry weight)/mol acetate when selenate or nitrate was the electron acceptor, respectively. Based upon a calculated YATP value of 10.0 g (dry weight) cells/mol ATP, for growth on acetate in inorganic salts, growth with selenate as the terminal electron acceptor theoretically yielded 5.7 ATP/acetate oxidized, and 5.5 ATP when nitrate was the terminal electron acceptor. The results support the conclusion that energy is conserved via electron transport phosphorylation when selenate or nitrate reduction are the terminal electron acceptors during anaerobic growth with acetate.
Article PDF
Similar content being viewed by others

Avoid common mistakes on your manuscript.
References
Brown NW, Watkinson JH (1977) An automated method for the determination of nanogram quantities of selenium. Anal Chim Acta 89: 29–35
Clesceri LS (ed. Chairman) (1989) Standard Methods for the Examination of Water and Wastewater, 17th edn. American Public Health Association, Washington DC
DeMoll-Decker H, Macy JM (1993) The periplasmic nitrite reductase of Thauera selenatis may catalyze the reduction of selenite to elemental selenium. Arch Microbiol 160: 241–247
Latimer WM (1938) The oxidation states of the elements and their potentials in aqueous solution. Prentice-Hall, Englewood Cliffs NJ
Macy JM, Michel TA, Kirsch DG (1989) Selenate reduction by a Pseudomonas species: a new mode of anaerobic respiration. FEMS Microbiol Lett 61: 195–198
Macy JM, Rech S, Auling G, Dorsch M, Stackebrandt E, Sly L (1993) Thauera selenatis gen. nov. sp. nov., a member of the beta-subclass of Proteobacteria with a novel type of anaerobic respiration. Int J Syst Bacteriol 43: 135–142
Neidhardt FC, Ingraham JL, Schaechter M (1990) Physiology of the bacterial cell. Sinauer, Sunderland Mass.
Rech S, Macy JM (1992) The terminal reductases for selenate and nitrate respiration in Thauera selenatis are two distinct enzymes. J Bacteriol 174: 7316–7320
Schmidt K, Jensen S, Schlegel HG (1963) Die Carotinoide der Thiorhodaceae. Arch Mikrobiol 46: 117–126
Stouthamer, AH (1973) A theoretical study on the amount of ATP reguired for synthesis of microbial cell material. Antonie van Leeuwenhoek 39: 545–565
Stouthamer AH, Bettenhausen C (1973) Utilization of energy for growth and maintenance in continuous and batch cultures of microorganisms. Biochim Biophys Acta 301: 53–70
Thauer RK, Morris JG (1984) Metabolism of chemotrophic anaerobes. Old views and new aspects. In: Kelly DP, Carr NG (eds) The microbe, part II: Prokaryotes and eukaryotes. Cambridge University Press, Cambridge, pp 123–168
Thauer RK, Jungermann K, Decker K, (1977) Energy conservation in chemotrophic anaerobic bacteria. Bacteriol Rev 41: 100–180
Thauer RK, Moeller-Zinkhan D, Spormann AM (1989) Biochemistry of acetate catabolism in anaerobic chemotrophic bacteria. Annu Rev Microbiol 43: 43–67
Whetter PA, Ullray DE (1978) Improved method for determining selenium. J Assoc Off Anal Chem 61: 927–930
Zumft WG, Kroneck PMH (1990) Metabolism of nitrous oxide. In: Revsbech NP, Sorensen J (eds) Denitrification in soil and sediment. Plenum Press, New York London, pp 37–55
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Macy, J.M., Lawson, S. Cell yield (YM) of Thauera selenatis grown anaerobically with acetate plus selenate or nitrate. Arch. Microbiol. 160, 295–298 (1993). https://doi.org/10.1007/BF00292080
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF00292080