
Abstract
The increase in salinity of aquatic and terrestrial ecosystems is a major consequence of current climate changes. It is therefore essential to be able to propose more adapted genotypes to farmers whether it concerns farmed fish or annual and perennial crops. To address these challenges, research is focused on acquiring better knowledge on mechanisms of adaptation to salt stress in order to effectively guide genetic improvement programmes and optimise breeding. We present various cases of adaptation to salinity in fish, rice and citrus. The selection of a tilapia strain adapted to salinity, especially via successive hybridizations and backcrosses, broadens the prospects for these farmed fish. Breeding rice to enhance salt tolerance has long been based on ‘conventional’ selection methods and is now oriented towards molecular marker assisted selection, while citrus adaptation to salt stress is based on rootstock choices and exploitation of polyploidy.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Background
The growing presence of salt in soils and changes in salinity in aquatic environments are major ecological disturbances related to climate change. In tropical agriculture, the severity of the phenomenon is especially high in irrigated areas. In this chapter we discuss the effects of increased salt water content on Nile tilapia, a major species for tropical fish farming. In agriculture, soil salinization can have a similar impact on annual crops such as rice, or tree crops like orange. Although tolerance mechanisms may be similar at the cellular level, research strategies to develop genotypes better adapted to these increasingly restrictive conditions often differ.
2 Tilapia Adaptation to Salinity
Rising sea levels and higher typhoon frequencies and intensities (e.g. in the Philippines) are causing saltwater intrusion in estuaries, salinization of rivers, groundwater, aquaculture ponds and saltwater flooding of mangroves (SEAFDEC-AQD 2013–2014). In West Africa, hypersalinity has been observed in Sahelian estuaries with a low flow rate, because the high water evaporation associated with successive droughts is no longer compensated by rainfall. The Sine-Saloum Estuary (Senegal, Gambia) has thus not only become hypersaline, but its salinity gradient has been reversed (with higher salinity upstream than downstream) (Savenije and Pagès 1992). In these estuaries, the lagoon tilapia Sarotherodon melanotheron heudelotii is tolerant to hypersalinity, but the energetic cost and the chronic stress of adapting to these waters have negative impacts on some of its life traits, such as growth and fertility, to the extent that the future of wild populations is threatened (Panfili et al. 2006).
Tilapia is the second major farmed fish group after carp (mainly produced and consumed in China), and all forecasts indicate that it will most likely exceed carp by 2025. Tilapia, of African origin, are farmed in over 100 countries. Within this species group, the Nile tilapia (Oreochromis niloticus) grows extremely well in freshwater environments and represents 90 % of the total tilapia production. However, it does not tolerate salinity variations.
Three main approaches can promote better tilapia adaptation to salinity. These include using the strong euryhalinity of O. mossambicus or S. melanotheron and improving their low growth through a selection programme in a saline environment; benefiting from the high growth-rate of Nile tilapia and selecting its salinity tolerance; and hybridizing a species with high salinity tolerance, such as O. mossambicus or S. melanotheron heudelotii with a fast growing species such as O. niloticus.
2.1 Osmoregulation of Fish in Saltwater Environments
Adaptation to a saline environment involves strict control of an organism’s salt and water balances—this is osmoregulation which has an energetic cost that represents 25–50 % of the metabolic expenditure. Since the 1960s, O. mossambicus and S. melanotheron heudelotii have become models for studying osmoregulation for tilapia adaptation to salinity.
Fish are hyperosmotic in freshwater environments. They balance their water input by producing highly diluted urine, while limiting Na+ (sodium) and Cl− (chloride) ion loss, thus enabling them to maintain stable plasma levels. In saltwater environments, however, they are hypoosmotic and tend to drink. Ions are then filtered in the gut and the excess salt subsequently eliminated via their gills.
An osmoregulation study performed in four S. melanotheron heudelotii populations showed that the strategies of adaptation to hypersaline waters were partly based on high branchial activity of the Na+/K+-ATPase enzyme, which helps maintain plasma osmolality at around 400 mOsm/kg (Fig. 4.1a, b). Morphological and functional modifications were noted in gill chloride cells. The number of these cells—which have a high mitochondria content and are specialized in ion transport—increased with the salinity, and in hypersaline conditions were observed along multiple layers with an increase in cell volume (Fig. 4.1c) (Ouattara et al. 2009).

a Plasma osmolality of natural S. melanotheron heudelotii tilapia populations living in very different saline environments: freshwater (FW), brackish water (BW), seawater (SW) and hypersaline water (HSW). b Gill Na+/K+-ATPase enzyme activity. c Na+/K+-ATPase pump immunolocalization, 72 h after transfer from FW to HSW at 70 g/l; Fi filament; La lamellae (from Ouattara et al. 2009)
The expression of candidate genes such as growth hormone and growth factors is stimulated in gills during transfer from freshwater to seawater, whereas it is inhibited in kidneys (Link et al. 2010). In fish adapted to hypersalinity, the genes identified were found to be mainly involved in transport activities, biological regulation and metabolic processes, thus confirming a higher metabolic activity in these fish (Tine et al. 2008).
2.2 Selection of a Saltwater-Adapted Tilapia Strain
The Molobicus project (CIRAD and the Bureau of Fisheries and Aquaculture Research, Philippines) focuses on the production of a fast-growing tilapia strain that is resistant to brackish water and seawater. The first phase carried out in freshwater, was based on hybridization and successive backcrosses between O. niloticus (high growth/low salt tolerance) and O. mossambicus (low growth/high salt tolerance). A rotational crossing design ensured high genetic variability in the hybrid population. As of the 2nd generation (H2), the strain bearing 25 % O. niloticus genes had high salt tolerance, thus enabling its rearing in brackish water. Growth of the hybrid population was then selected according to an intrafamilial design on the basis of 50 unrelated hybrid H2 families. This step was conducted in a saltwater environment and combined passive selection of salt-tolerant individuals and active selection for growth. An ‘extensive’ line (low-density, fertilized ponds, no feed provided) and an ‘intensive’ line (high-density ponds, protein-rich feed supply) were selected in parallel. After four generations, 7.3 % (12.5 g) genetic gain per generation was obtained (from Verdal et al. 2014; Fig. 4.2).

Testing the salinity tolerance of Molobicus hybrids (H2-2nd and H3-3rd generations) and of the parental species O. niloticus and O. mossambicus during a gradual increase in salinity of 3 g/l a day
2.3 Future Research Opportunities
Selection programmes aimed at obtaining tilapia strains that are salinity-tolerant should, despite the cost, be based on selection approaches assisted by molecular markers. The sequencing of the Nile tilapia genome (Brawand et al. 2014) will allow to increase the number of genes and markers that can be used in association studies with the salinity-adaptation trait. However, this genomic selection is not yet possible due to the low tilapia prices. The presence of females that reproduce on a monthly basis while having a much lower growth-rate than males is an obstacle to tilapia production. This growth dimorphism is also found in salinity-tolerant tilapia strains. The high estrogen level in females also seems to reduce salinity tolerance. Temperature-dependent sex determination (as a substitute for current hormone treatments) should be an effective way to produce only males (more resistant to salinity with a higher growth-rate). A further option involves the production of sterile individuals (no energy expended in reproduction) via triploidization.
3 Salinity and Rice Growing
Around 10 % of the 160 million ha of rice fields worldwide are affected to various extents by permanent or temporary salinity due to the presence of sodium chloride (NaCl), especially in coastal and delta regions. Climate change will increase the incidence and severity of this problem by causing a rise in sea level and temperature, leading to greater evapotranspiration. Rice (Oryza sativa L.) is moderately susceptible to salinity. The growth and development of this species are affected when the soil electrical conductivity associated with the presence of Na+ ions exceeds the threshold of 3 decisiemens per metre (dS/m). Sensitivity is especially high at the seedling stage and during the reproductive phase. The yield components most affected are the number of grains per panicle, spikelet fertility and grain weight (Fig. 4.3).

A rice field affected by salinity. In the absence of sufficient rainfall or irrigation, intense evaporation is accompanied by a rise in the groundwater salt content, northern India (© IRRI)
Control of the salinity impact could be based on three non-exclusive approaches: overcoming the problem by changing the cropping environment, avoiding rice cropping during periods of the year when there is a high salt stress risk, and mitigating the effects of salinity on the crop. The first approach requires expensive irrigation schemes and is often unaffordable for rice smallholders. The second approach, which requires rice varieties with an adapted crop cycle, remains uncertain because the onset of water salinization episodes is not always predictable. The third approach involves the use of tolerant varieties and is thus the main way to cope with salinity in rice cropping.
3.1 Rice × Salinity Interaction—Adaptation Mechanisms
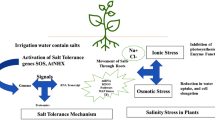
In saline environments, Na+ ions are taken up by rice roots and enter the transpiration stream via the apoplastic pathway or move toward aerial parts via symplastic pathways involving the activity of specific transmembrane transporters. This results in two types of tissue stress: osmotic, associated with the higher concentration of solutes in the soil, and ionic, due to alteration of the K+/Na+ ion ratio. The almost immediate short-term response is the reduction in transpiration and photosynthesis to avoid dehydration, and the activation of perception and signalling mechanisms. The medium-term response, i.e. after a few hours, includes the activation of osmotic adjustment mechanisms, the restoration of K+/Na+ homeostasis and oxygen-free radical (OFR) control. Adjustment of the osmotic potential between the cytoplasmic matrix (cytosol) and the apoplastic solution to prevent cell dehydration and the resulting protein denaturation is achieved by the accumulation of nontoxic metabolites, or so-called ‘compatible solutes’, particularly the proline amino acid. The restoration of K+/Na+ homeostasis throughout the plant requires selective exclusion of Na+ ions by epidermal root cells, preferential K+ uptake in the xylem, Na+ retention in the upper root parts and stem base, K+ to Na+ exchange and intracellular sequestration of Na+ ions in vacuoles. K+/Na+ homeostasis avoids inhibition of the activity of cellular enzymes, allowing them to function normally despite the high Na+ concentration. The plant increases antioxidant synthesis to control OFRs, which will otherwise destabilize the cell structure. In the longer term, i.e. a few days, the change in cellular osmotic and ionic balances affects the entire plant in its growth, development and grain production.
Finally, if salinity intensity is too high or if the expression of salt-tolerance mechanisms is not sufficient to exclude salt from the transpiration system, the accumulation of salt to toxic levels in the oldest leaves, that have transpired the longest, will lead to their death. As the growth of new leaves depends on carbohydrates from older leaves, the plant’s fate depends on the balance between the rates of old leaf death and new leaf emergence. Salt stress also delays panicle initiation and flowering and reduces spikelet fertility because of the lower pollen viability. The plant will die before flowering or maturity if the salinity intensity is very high and if salt exposure exceeds 2–3 weeks.
3.2 Genetic Basis of Salinity Tolerance in Rice
There is substantial genetic variability in salinity tolerance in rice. The Indian landrace variety Nona Bokra, belonging to the indica group, is a reference in this respect. Genetic variability has also been noted for most tissue and cellular mechanisms of rice responses to salinity. Correlations are loose between individual tolerance mechanisms and between those mechanisms and whole plant survival or grain production under salinity stress. Regardless of the mechanisms involved, the plant’s ability to maintain a high tissue K+/Na+ ratio is a key tolerance factor. The leaf K+/Na+ ratio is a good predictor of production loss under salt stress (Fig. 4.4).

Scatterplot of 200 rice varieties projected on the two first axes of a principal component analysis using seven developmental response variables and an ionic response variable (leaf K+/Na+ ratio) under moderate salt stress in the vegetative phase. The leaf K+/Na+ ratio, which highly contributed to defining axis 1, clearly differentiated tolerant varieties such as Nona Bokra from sensitive varieties such as IR29 (from Ahmadi et al. 2011)
An analysis of phenotypic diversity regarding the impact of salinity on whole plant growth and development and on the leaf K+/Na+ ratio led to the identification of tolerant varieties in O. sativa indica and temperate japonica groups (Fig. 4.4). Tolerance seems infrequent and low in O. glaberrima, the African cultivated rice species. However, high tolerance exists in O. coarctata, a distant wild relative of cultivated rice. A viable plant was recently obtained from an O. sativa x O. coarctata cross, thus enhancing the prospects for developing a ‘supertolerant’ rice variety.
Insight has been gained on the genetic and molecular bases of a high number of biological processes described above through techniques involving genetic mapping, functional genomics, as well as analysis of the differential expression of genes in different organs at various rice development stages under salt stress.
Genetic mapping in the offspring of crosses between a susceptible variety and a tolerant variety, which were assessed under hydroponic or field cropping conditions, led to the identification of a high number of quantitative trait loci (QTLs), especially on chromosomes 1, 4, 6 and 7.Footnote 1 One of these QTLs located on the short arm of chromosome 1 and having a marked effect on plant survival and grain production was the focus of positional cloning. The identified gene, i.e. SKC1 (OsHKT8), codes for a Na+ transporter and promotes Na+/K+ homeostasis. The same zone on chromosome 1 seems to host a group of genes involved in the rice salt stress response.
Differential expression and functional and comparative genomics approaches have revealed a very high number of candidate genes involved in the rice salt stress response (Negrão et al. 2011). All of these approaches are based on genetic engineering techniques, which are well established in rice.
An association study focused on a temperate japonica group led to the detection of 19 loci significantly associated with one or several phenotypic salinity response variables (Ahmadi et al. 2011). The analysis of the function of these loci showed that all major salinity adaptation mechanisms were present in the temperate japonica group, but that no varieties in the panel had a sufficient level of expression for all of the mechanisms. Under moderate salt stress, some varieties achieved the same level of tolerance as the tolerant Nona Bokra reference variety, but without having the same allele at several tolerance-related loci. This suggests that there are differences between indica and japonica varieties regarding QTLs and genes involved in salinity tolerance and that it would therefore be possible to enhance rice salinity tolerance beyond that of the reference variety.
3.3 Breeding Salt-Tolerant Rice
Rice breeding programmes geared towards obtaining salt-tolerant varieties were for a long time based on selection for yield under salt stress in progeny of crosses between a salinity susceptible yet popular variety and a tolerant variety. The efficiency of this ‘conventional’ selection method was hampered by the lack of control of the period of salt stress and its intensity and by the difficulty of distinguishing and cumulating the effects of different tolerance mechanisms.
The advent of molecular marker-assisted selection (MAS) has overcome some of these difficulties. For instance, the favourable allele of the SKC1 tolerance gene in the Nona Bokra variety was transferred to many modern varieties. Salinity tolerance is, however, a complex trait involving a high number of genes, each explaining a small share of the observed phenotypic diversity. The SKC1 gene has the greatest known effect but it only explains 40 % of the observed variation in its segregating progeny. We thus recommend recurrent MAS schemes in biparental or (even better) multi-parental crosses, to achieve pyramiding of favourable alleles from the many loci involved in tolerance.
Future progress towards gaining insight into the functioning of one or several gene networks involved in the rice response to salinity should enable better targeting of genes to be included in conventional MAS approaches based on associations between molecular markers and the targeted phenotypic trait. But as tolerance to salinity is a complex trait, the new so-called genome-wide MAS is probably the most suitable breeding approach. This approach does not require any prior knowledge of associations between the genotype (marker) and phenotype, but it does require an ability to accurately assess the tolerance of a large number of plants in order to formulate prediction equations.
Another way of improving rice salinity tolerance involves modification of the expression of genes involved in plants’ response to salinity using genetic engineering techniques. Over the last 15 years, these techniques have been applied to a large number of mechanisms and genes of rice salinity tolerance. The results are hard to assess in the current societal and regulatory setting with regard to field testing of genetically modified plants. Future progress on the accuracy of genetic engineering and of spatiotemporal control of expression of target genes should enhance the efficiency of these techniques within the limits imposed by global balances that govern plant growth and development. Genetic engineering techniques can also be used to insert genes from wild relatives or other salt-tolerant species into rice.
There is hence considerable room for progress in improving rice tolerance to salinity.
4 Citrus Adaptation to Salinity
Citrus represents the top tree fruit crop on the world market. Citrus fruits—native to tropical and subtropical Asian regions—are grown in all hot regions, usually under irrigation, as is the case throughout the Mediterranean Basin. Water resources are increasingly limited on the southern shores of the Mediterranean Sea, for instance, where the water demand is so substantial that the groundwater is lowering, with a concomitant increase in salt content. This is already having an impact on agriculture. Citrus trees are amongst the most susceptible of all trees to salt stress. The critical electrical conductivity level of water is 3 dS/m. For grapefruit and orange trees, the minimum tolerable electrical conductivity in soil is around 1.8 dS/m. The sensitivity can, however, vary depending on different parameters, such as tree age, rootstock, grafted variety, irrigation system, soil type and climatic conditions.
To ensure better citrus adaptation to salinity, it is essential to effectively manage irrigation practices in order to hamper salt seepage in the soil, while having access to rootstocks capable of limiting Na+ and Cl− uptake in the root system and to varieties that foster physiological and molecular mechanisms that will reduce the impact of toxic ions.
4.1 Citrus Propagation Strategies
Citrus are classified in three main sexually compatible genera: Citrus, Poncirus and Fortunella. Most currently used rootstocks belong to Citrus and Poncirus, or are hybrids obtained by crosses between these two genera. Rootstocks are seed propagated. Rootstock seeds that are generally used are polyembryonic. Supernumerary embryos are from nucellar cells (somatic tissue) and are genetically identical to the parent tree. The most commonly cultivated fruit varieties (orange, mandarin, lemon, grapefruit and lime) all belong to the Citrus genus. Varieties are clonally propagated by grafting.
Most citrus varieties are diploid (2x). Other polyploidy levels have also been identified such as tetraploidy (4x) resulting from incomplete meiosis, which produces nucellar cells with a double set of chromosomes. Around twenty 4x rootstocks have been selected in the San Giuliano collection in Corsica, jointly managed by INRA and CIRAD. These trees produce polyembryonic seeds that give rise to 4x seedlings after they are sown.
4.2 Citrus × Salinity Interactions—Adaptation Mechanisms
Unlike most plants, the susceptibility of citrus to salt stress is due to the presence of Cl− ions but the molecular mechanisms involved have yet to be specifically identified. The negative impacts of salinity on citrus trees result in symptoms such as leaf burn, stunting and reduced fruit production. The impact of salinity on plant growth and development is associated with serious physiological disorders, leading to a reduction in the cellular osmotic potential, the accumulation of Cl− and Na+ ions to toxic levels, and finally to nutritional imbalances that can ultimately induce a reduction in growth and fruit yield. Several adaptation processes are induced, such as cellular K+ uptake, the compartmentalization of Na+ and Cl− ions out of the cytoplasm and solute synthesis, thus enhancing osmotic adjustment. Finally, oxidative stress triggering is a side effect of the presence of toxic ions in the cytoplasm, which can cause major damage to the photosynthesis machinery. As noted in rice, the presence of cellular OFRs triggers detoxification systems to limit the induced oxidative stress.
4.3 Rootstock, Diversity and Salt Tolerance
Oppenheimer (1937) was the first to demonstrate the impact of rootstock on the behaviour of citrus trees exposed to salt stress. Three main groups were identified: a tolerant group, including ‘Sour’ orange and ‘Cleopatra’ mandarin which are considered to be well adapted to abiotic constraints, a susceptible group with ‘Rough’ lemon and ‘Carrizo’ citrange, and finally a highly susceptible group represented by Poncirus accessions (Poncirus trifoliata). Despite the good behaviour of some rootstocks in response to salt stress, their use is very limited due to their susceptibility to certain diseases, such as Phytophthora spp. and tristeza. Poncirus accessions are tristeza tolerant but do not limit root Cl− uptake. Once in the roots, these ions transit through the xylem via the transpiration stream to the upper part of the tree, often causing leaf necrosis. The leaf Cl− content can thus be considered a good criterion for assessing the salt tolerance/susceptibility properties of rootstock seedlings (Hussain et al. 2012a). ‘Cleopatra’ mandarin x Poncirus hybrids are now widely used in rootstock selection programmes to transmit biotic and abiotic stress traits to progeny.
An analysis of physiological salt stress tolerance behaviour as a function of citrus species was recently conducted (Fig. 4.5; Hussain et al. 2012a). Citron were the most susceptible whereas mandarin and pummelo were much more tolerant. Susceptible genotypes exhibited chlorosis symptoms combined with high leaf Cl− and Na+ ion uptake. Photosynthesis and growth were maintained in the tolerant genotypes and had low Cl− and Na+ ion uptake. Rapid leaf loss was observed in some species such as grapefruit. This phenomenon was followed by growth of new leafy shoots, which could be interpreted as an adaptation response. New sources of tolerance could thus likely be tapped within citrus diversity in order to enhance salt tolerance in citrus trees.

Illustration of the impact of salt stress on major citrus fruit varieties (Hussain et al. 2012a)
4.4 Polyploidy and Adaptation to Salt Stress
The morphology and anatomy of 4x citrus genotypes differ markedly from those of their 2x counterparts. The growth of 4x plants is usually lower than that of 2x plants. Stems and roots of 4x plants are generally thicker and more succulent than those of 2x plants (Allario et al. 2011; Hussain et al. 2012b) since 4x cells are larger than 2x cells.
In salt stress conditions, 4x rootstocks were found to be more tolerant than 2x rootstocks (Saleh et al. 2008; Mouhaya et al. 2010), although leaf Cl− and Na+ ion accumulation was not significantly different. These latter results suggest that 4x genotypes have better compartmentalization and detoxification capacities than 2x, as documented in other 4x plants (Zhang et al. 2010). However, the impact of salt stress on grafted 4x rootstocks in orchard conditions is not yet known.
4.5 Breeding Salt-Tolerant Varieties
Germplasm and polyploidization (tetraploidization) will be exploited in the coming years to select new rootstocks in order to hamper toxic ion uptake in roots and thereby enhance salt stress tolerance. Several rootstocks developed by CIRAD are currently being evaluated, which should broaden the range of rootstocks available for citrus industry. Another way to deal with this issue is to assess the salt tolerance properties of newly created fruit varieties (scions) rather than those of rootstocks. Several thousands of 2x and 3x citrus varieties are being assessed in collaboration with private partners as part of the CIRAD citrus breeding programme. An assessment was recently conducted regarding the salt tolerance properties (including compartmentalization of toxic ions and improved OFR detoxification capacity) of varieties preselected for their taste qualities. It should thus be possible to strengthen the tolerance properties of orchard trees through the association of more salt-tolerant varieties with rootstocks adapted to saline soil.
5 Conclusion
There have been major efforts to develop more salt-tolerant genotypes. However, it is essential that not only the biological aspects but also the economic and social factors be taken into account to ensure that human activities will mitigate the impact of this stress on ecosystems as much as possible. Research innovations can prompt rethinking on livestock and crop farming practices, but users must also participate closely in the assessment of these new genotypes adapted to growing in salt stress conditions.
Notes
- 1.
These have been compiled in the following database: <http://tropgenedb.cirad.fr>.
References
Ahmadi N, Negrão S, Katsantonis D, Frouin J, Ploux J, Letourmy P, Droc G, Babo P, Trindade H, Bruschi G, Greco R, Oliveira MM, Piffanelli P, Courtois B (2011) Targeted association analysis identified japonica rice varieties achieving Na+/K+ homeostasis without the allelic make-up of the salt tolerant indica variety Nona Bokra. Theor Appl Genet 123(6):881–895
Allario T, Brumos J, Colmenero-Flores JM, Tadeo F, Froelicher Y, Talon M, Navarro L, Ollitrault P, Morillon R (2011) Large changes in anatomy and physiology between diploid Rangpur lime (Citrus limonia) and its autotetraploid are not associated with large changes in leaf gene expression. J Exp Bot 62:2507–2519
Brawand D, Wagner C, Li YI, Malinsky M, Keller I et al (2014) The genomic substrate for adaptive radiation: genomes of five African cichlid fish. Nature 513:375–381
de Verdal H, Rosarioc W, Vandeputte M, Muyalde N, Morissens P, Baroiller JF, Chevassus B (2014) Response to selection for growth in an interspecific hybrid between Oreochromis mossambicus and O. niloticus in two distinct environments. Aquaculture 430:159–165
Hussain S, Luro F, Costantino G, Ollitrault P, Morillon R (2012a) Physiological analysis of salt stress behavior of citrus species and genera: low chloride accumulation is an indicator of salt tolerance. S Afr J Bot. doi:10.1016/j.sajb.2012.06.004
Hussain S, Curk F, Dhuique-Mayer C, Urban L, Ollitrault P, Luro F, Morillon R (2012b) Autotetraploid trifoliate orange (Poncirus trifoliata) rootstocks do not impact clementine quality but reduce fruit yields and highly modify rootstock/scion physiology. Sci Hortic 134:100–107
Link K, Berishvili G, Shved N, D’Cotta H, Baroiller JF, Eppler E, Reinecke M (2010) Seawater and freshwater challenges affect the insulin-like growth factors IGF-I and IGF-II in liver and osmoregulatory organs of the tilapia. Mol Cell Endocrinol 327:40–46
Mouhaya W, Allario T, Brumos J, Andrés F, Froelicher Y, Luro F, Talon M, Ollitrault P, Morillon R (2010) Sensitivity to high salinity in tetraploid citrus seedlings increases with water availability and correlates with expression of candidate genes. Funct Plant Biol 37:674–685
Negrão S, Courtois B, Ahmadi N, Abreu I, Saibo N, Oliveira MM (2011) Recent updates on salinity stress in rice: from physiological to molecular responses. Crit Rev Plant Sci 30:329–377
Oppenheimer HR (1937) Injurious salts and the ash composition of fruit trees. Hadar 10:316
Ouattara N, Bodinier C, Nègre-Sadargues G, D’Cotta H, Messad S, Charmantier G, Panfili J, Baroiller JF (2009) Changes in gill ionocyte morphology and function following transfer from fresh to hypersaline waters in the tilapia Sarotherodon melanotheron. Aquaculture 290:155–164
Panfili J, Thior D, Ecoutin JM, Ndiaye P, Albaret JJ (2006) Influence of salinity on the size at maturity for fish species reproducing in contrasting West African estuaries. J Fish Biol 69:95–113
Saleh B, Allario T, Dambier D, Ollitrault P, Morillon R (2008) Tetraploid citrus rootstocks are more tolerant to salt stress than diploid. Comptes rendus de biologie de l’Académie des sciences 331:703–710
Savenije HHG, Pagès J (1992) Hypersalinity: a dramatic change in the hydrology of Sahalian estuaries. J Hydrol 135:157–174
SEAFDEC-AQD (2013–2014) Southeast Asian Fisheries Development Center, Aquaculture Department, Adapting to Climate Change, R&D. <www.seafdec.org.ph>
Tine M, de Lorgeril J, D’Cotta H, Pepey E, Bonhomme F, Baroiller JF, Durand J-D (2008) Transcriptional responses of the black-chinned tilapia Sarotherodon melanotheron to extreme salinity. Marine Genomics 1:37–46
Zhang XY, Hu CG, Yao JL (2010) Tetraploidization of diploid Dioscorea results in activation of the antioxidant defense system and increased heat tolerance. J Plant Physiol 167:88–94
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Éditions Quæ
About this chapter
Cite this chapter
Ahmadi, N., Baroiller, JF., D’Cotta Carreras, H., Morillon, R. (2016). Adaptation to Salinity. In: Torquebiau, E. (eds) Climate Change and Agriculture Worldwide. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-7462-8_4
Download citation
DOI: https://doi.org/10.1007/978-94-017-7462-8_4
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-017-7460-4
Online ISBN: 978-94-017-7462-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)