
Abstract
People in developing countries mostly depend for their diet on special staple crops, so called orphan crops. These crops play a key role in food security since they are grown by many resource-poor farmers and consumed locally. Despite their huge importance in the economy and livelihood of the developing world, orphan crops have received little attention in terms of scientific improvement. Although conventional breeding is widely implemented to improve crop plants, alternative methods such as marker-assisted breeding and reverse genetics approaches have proved to be efficient in developing crop cultivars. In this review, we present detailed description of a non-transgenic and reverse genetics technique called TILLING (Targeting Induced Local Lesion IN Genomes). The method was originally optimized in the model plant Arabidposis thaliana and subsequently applied to crops such as maize, wheat, and rice. We also present detailed procedures for several TILLING strategies and discuss their benefits and drawbacks. The application of the technique for orphan crop improvement is also discussed based on several TILLING platforms currently carried-out on these understudied crops of the world.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Most people in developing countries depend on special crops as their food source and/or income generation. These crops are mostly known as orphan crops but also as understudied-, lost-, disadvantaged- crops, or NUS (neglected and underutilized crops). According to the Global Facilitation Unit for Underutilized Species (GFU), several criteria must be met for a species to be considered as an orphan crop, including proven food or energy value and that the plant has been widely cultivated in the past, or is currently cultivated in a limited geographical region (http://www.underutilized-species.org/species/about_species.asp; accessed August 2011). Orphan crops are annually grown on more than 250 million ha in developing countries (Naylor et al. 2004). In general, these crops play a key role in the livelihood of resource-poor farmers and consumers in developing countries where they perform better than major crops under extreme soil and climate conditions prevalent in the region.
1.1 Types and Diversity of Orphan Crops
Orphan crops belong to the major groups of crops including cereals [e.g., finger millet (Eleusine coracana), tef (Eragrostis tef), fonio (Digitaria spp)], legumes [e.g., cowpea (Vigna unguiculata), bambara groundnut (Vigna subterranea), grass pea (Lathyrus sativus)], and root crops [e.g., cassava (Manihot esculenta), yam (Dioscorea spp), enset (Ensete ventricosum)]. Although orphan crops are many in numbers, brief descriptions are provided below for the most important ones in terms of area they are grown or population they feed.
Finger millet is the most important millet cultivated in Africa and Asia predominantly as a staple food grain (http://test1.icrisat.org/SmallMillets/SmallMillets.htm; accessed July 2011). The plant is tolerant to drought and the seeds contain methionine (NRC 1996), a valuable amino acid lacking in the diets of millions of the poor people who live on starchy staples such as cassava. Finger millet is also a popular food among diabetic patients because of its low glycemic index and slow digestion (Chandrashekar 2010).
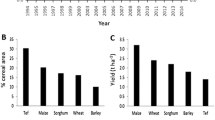
Tef is a cereal crop mainly grown in the Horn of Africa, and its annual cultivation in Ethiopia alone accounts for over 2.8 million ha of land (CSA 2011). The crop is tolerant to abiotic stresses especially to poorly drained soils where other crops such as maize and wheat do not withstand. In addition, tef is considered a healthy food, since the seeds do not contain gluten, the cause for celiac disease (Spaenij-Dekking et al. 2005; Hopman et al. 2008). Despite many benefits associated with the cultivation of tef, the grain yield obtained from the crop is extremely low. The main cause for this extremely low productivity is the susceptibility of the tef plant to lodging (Assefa et al. 2009). Tef plants possess tall and tender stems which are susceptible to damage by wind and rain, therefore lodging (the permanent displacement of the stem from the up-right position) contributes to significant loss in production.
Fonio (acha) is an indigenous West African crop mainly grown by small farmers for home consumption. The crop is not only tolerant to drought, but also a very fast maturing crop. It is also considered as highly nutritious, since it is rich in methionine and cysteine, the two amino acids essential for human health but deficient in major cereals such as wheat, rice, and maize (IPGRI 2004).
Bambara groundnut is an annual legume crop grown for human consumption and is the third most important grain legume in Africa after cowpea and groundnut (Asiwe 2009). The seeds of bambara groundnut are known as a complete food, because they contain adequate quantities of protein (19 %), carbohydrate (63 %), and fat (6.5 %; NRC 2006).
Grass pea is another legume commonly grown for human consumption in Asia and Africa. The plant is extremely tolerant to drought and is considered as an insurance crop, since it produces reliable yields when all other crops fail due to moisture scarcity. Like other legumes, grass pea is a source of protein, particularly for resource poor farmers and consumers. However, the seeds of grass pea contain a neuron-toxic substance called ODAP (β-N-Oxalyl-L-α, β-diaminopropanoic acid; Yan et al. 2006). ODAP is the cause of a disease called neurolathyrism, a neurodegenerative disease that results in paralysis of the lower body. Serious neurolathyrism epidemics were reported during famines when grass pea was the only food source (Getahun et al. 2003).
Cassava (manioc) is a woody shrub, native in South America and intensively cultivated in the tropic and sub-tropic areas of the world (Nassar 2009). It is tolerant to drought and also performs better than other crops on soils poor in nutrients. The major problems related to cassava are low protein content and presence of poisonous compounds called cyanogenic glycosides (CG) in the edible part which release cyanide (Ceballos et al. 2004). Konzo is the paralytic disease associated with consumption of insufficiently processed cassava.
Yam combines different species under the genus Dioscorea. It is grown on about 5 million hectares of land worldwide (IITA, http://www.iita.org/; accessed July 2011) and is regarded as a staple food in West Africa. The edible roots look like sweet potato (Ipomoea batatas), although these species are not taxonomically related.
Enset is commonly known as ‘false banana’ for its close resemblance to the domesticated banana plant. Unlike banana, where the fruit is consumed, in enset the pseudostem and the underground corm are the edible parts. Enset is the major food for over 10 million people in densely populated regions of Ethiopia. The plant is considered as extremely drought tolerant and adapts to different soil types (Brandt et al. 1997). Although food products from enset are rich in starch, they are deficient in other essential nutrients; hence enset-based diets need supplementation with proteins.
1.2 Role of Orphan Crops in Developing Countries
Feeding an increasing population is a major global challenge, especially in the face of climate change. Understudied- or orphan crops play a key role in food security, nutrition, and income generation to resource-poor farmers and consumers in developing countries and are compatible with agro-ecology and socio-economic conditions. Adaptation or tolerance of orphan crops to abiotic stresses such as moisture scarcity also increases their importance in food security. For example, orphan crops such as finger millet and bambara groundnut are extremely tolerant to prolonged periods of drought or to moisture scarcity (Padulosi et al. 2009; Stadler et al. 2009).
According to Naylor et al. (2004) food security can only be achieved if emphasis is given to the improvement of crops widely cultivated by the poor. Among these, sorghum, millet, and cassava are considered critical crops in African agriculture (Ejeta 2010). In Ethiopia, tef is the main staple cereal grown on the largest area allocated for cereal crops (CSA 2011). In Uganda, sweet potato is a dominant staple crop which ranks third next to cassava and banana in providing the diet for poor rural households (Naylor et al. 2004). Furthermore, cowpea is a major source of dietary protein for the resource poor people in the Sub-Saharan Africa, South Asia, and parts of Latin America (Boukar and Fatokun 2009). Minor millets are also grown primarily by poor farmers in Africa and South Asia (Naylor et al. 2004). In India, for instance, minor millets are annually cultivated on about three million hectares of land, equivalent to 12 % of the whole area under coarse cereals (Paludosi et al. 2009).
1.3 Major Constraints of Orphan Crops Cultivation
Despite many beneficial aspects, orphan crops are also bound by a number of challenges especially related to poor productivity. As indicated above, the major bottlenecks affecting these crops are traits such as low yield (e.g., tef and millet), poor nutritional value (e.g., cassava and enset), and the production of toxic substances (e.g., cassava and grass pea).
In addition, crop productivity is affected by a variety of abiotic and biotic stresses. Major abiotic stresses are drought, soil salinity, and acidity. There is some evidence that in recent decades agricultural land has been lost to desertification, salinization, soil erosion, and other consequences of unsustainable land use (Godfray et al. 2010). Furthermore, due to the presence of high density and diversity of pests, diseases, and weeds in tropical regions, productivity of crops significantly decreases. Other factors that affect food production are the use of agricultural land for urbanization and biofuel production, and environmental challenges such as global warming (Godfray et al. 2010).
1.4 Need for Orphan Crop Improvement
Crop production could be increased by either expanding the arable area or through intensification, i.e., using improved seeds, fertilizer, fungicides, herbicides, irrigation, etc. According to the Food and Agriculture Organization (FAO), agricultural intensification represents about 80 % of future increases in crop production in developing countries (FAO 2002). Based on this goal, crop breeders and scientists are focusing towards achieving improved cultivars that produce higher yields and at the same time tolerate the sub-optimal soil and climatic conditions.
Among traits that contributed to higher crop productivity in the last century, those which alter the architecture of plants rank first. Architectural changes include alteration in branching pattern and reduction in plant height. The major achievement of the Green Revolution in the 1960s was due to the development and introduction of semi-dwarf crop varieties of wheat and rice along with optimum level of inputs for crop production. These broadly adapted semi-dwarf cultivars were responding to fertilizer application and this led to tremendous increases in productivity. Currently, a number of genes affecting plant height are known in major cereal crops including wheat, rice, and maize (for review, Wang and Li 2006). According to the International Food Policy Research Institute (IFPRI), the Green Revolution represented the successful adaptation and transfer of scientific revolution in agriculture (IFPRI 2002). However, this agricultural revolution which boosted crop production in Asia and Latin America did not occur in Africa. This is mainly due to the fact that the Green Revolution focused on rice and wheat, but not on African crops such as sorghum and millet (Ejeta 2010). Therefore, new strategies and approaches have to be implemented to boost crop productivity in African orphan crops.
2 Major Crop Improvement Techniques
Improvement of existing crop varieties and cultivation needs integrative research strategies. However, due to the lack of attention and funds, orphan crop researchers had little exposure to advanced scientific technologies. Crop improvement techniques are broadly grouped into (1) conventional approaches that include various types of selection methods, introgression (hybridization), and mutation breeding, and (2) biotechnological or molecular approaches that include transgenic and non-transgenic methods such as marker-assisted breeding (MAB) and TILLING (Targeting Induced Local Lesions IN Genomes).
2.1 Selection
Selection is an ancient breeding method that is still implemented at large-scale to improve crop plants particularly orphan crops. The technique relies mainly on the selection of plants according to their phenotype and performance. Diverse types of selection techniques have been developed for a variety of crops depending on the pollination behaviour and other factors. For example, mass selection is applied to a certain level in self-fertilizing plants and is an effective method for improving landraces, especially for transferring highly heritable traits (Ghosh 2011). Mass selection refers to the technique whereby individual plants are selected based on their phenotypic performance, and bulk seeds from selection are used to produce the next generation.
2.2 Introgression
Introgression or artificial hybridization refers to crossing closely related species in order to create genetic variation which can be utilized for improving traits of choice. According to Baenziger et al. (2006) the success in hybridization depends mainly on the selection of parents. Hybridization can be broadly grouped into intra-specific (crossing within the species) or inter-specific (crossing between species). Successes in intra-specific crosses resulted in semi-dwarf cultivars of wheat and rice which boosted the productivity of both crops during and after the Green Revolution (Borlaug 2007). The major breakthrough from the inter-specific crossing was the development of an artificial cereal called Triticale. Triticale is a cross between wheat and rye and proved to be tolerant to abiotic stresses such as soil acidity (Kim et al. 2001).
2.3 Mutation Breeding
Mutation breeding relies on the implementation of either physical or chemical agents in order to create variability in the population of interest. Mutagenesis created by these agents is widely used for crop improvement, and foods derived from the process are readily accepted by the public for consumption (Jain 2007). During the last seventy years, more than two thousand varieties of crop plants derived from mutation breeding have been released either as direct mutants or as parents to derive new varieties. Most mutation breeding programs aimed at altering traits such as plant height and disease resistance in well-adapted plant varieties. The technique of mutation breeding has been widely implemented in crops such as rice, barley, wheat, chickpea, and soybean (Ahloowalia et al. 2004). In rice alone, over 400 improved varieties have been derived from this technique (Ahloowalia et al. 2004). Among these, mutation in the semi-dwarf cultivar ‘Calrose 76’ significantly contributed to rice improvement. In this particular cultivar the Sd 1 (semi-dwarf 1) gene is down-regulated using gamma ray mutagenesis. Two high-yielding and semi-dwarf cultivars of barely, namely ‘Golden Promise’ and ‘Diamant’ which were also obtained through induced mutation played key role in the development of the European brewing industry. Orphan crops have also benefited from mutation breeding. For example, the release of early maturing and virus resistant mung bean, and high-yielding and blight resistant chick pea were the major contributions of mutation breeding (Ahloowalia et al. 2004; Jain 2007). In addition, the FAO-IAEA project “Genetic improvement of underutilized and neglected crops in low income food deficit countries” (LIFDCs) implemented irradiation and related techniques in order to improve orphan crops (Jain 2009).
2.4 In Vitro Culture
Plant tissue culture is the aseptic culture of cells, tissues, organs, and their components under defined physical and chemical conditions in vitro (Thorpe 2007). Developing an efficient regeneration system requires optimization for various types of explants and media components. Hormones and growth regulators play key role in determining the conversion of somatic cells to embryogenic tissues (Jimenez 2005). The tissue culture techniques have been successfully implemented in diverse types of plants including cereals (Maqbool et al. 2001; Vasil 2005), legumes (Lakshmanan and Taji 2000), vegetables (Bal and Abak 2007; Ochoa-Alejo and Ramirez-Malagon 2001), oil plants (Wahid et al. 2005), trees (Giri et al. 2004), and forestry (Golle et al. 2009). Tissue culture also enables to rescue and utilize desirable properties of endangered plant species in crop improvement (Bapat et al. 2008). Among diverse tissue culture techniques, the doubled haploids are becoming a popular method in crop improvement (Germana 2011).
2.5 Marker-Assisted Breeding
Marker-assisted breeding (MAB or MAS for marker-assisted selection) is the utilization of molecular markers located near genes, which can be traced to breed for traits that are difficult to observe. Tester and Langridge (2010) indicated the benefits of applying new technologies and molecular markers in crop improvement. These molecular markers are utilized to effectively assemble favorable alleles in phenotypic selection (Charcosset and Moreau 2004). Commonly applied markers are Amplified Fragment Length Polymorphisms (AFLPs), and microsatellites (or Simple Sequence Repeats; SSRs), while markers based on single nucleotide polymorphism (SNPs) are rapidly emerging. According to Collard and Mackill (2008) the following factors should be considered before selecting the type of DNA marker to be used in MAS: reliability, quantity and quality of DNA required, technical procedure for marker assay, level of polymorphism, and cost.
2.6 Transgenic Approach
Transgenic technology is also considered as another approach to improve the productivity of crops. To date, only a few crops have benefited from transgenic improvements, but due to high adoption rate, the global area under transgenic crops has increased tremendously in the last decade (James 2010). Although the technology has shown significant impact in increasing crop productivity, due to extensive regulatory procedures and negative public perception, its expansion to other crops and geographical regions remains restricted (Tester and Langridge 2010). Some of the recent investigations on transgenics dealt with solving the major concerns related to the technology to promote the acceptance by the public. Among the concerns, the presence of antibiotic resistance and non-plant promoters were the major ones. However, Ayalew and Stewart (2005) enabled to substitute antibiotic resistant markers with those without any adverse effects. In addition, Jacobsen and Schouten (2007) introduced the modified cisgenesis method whereby plant specific promoters are used instead of foreign promoters from bacteria or other organisms. Advances in increasing the efficiency of gene targeting as demonstrated by Shukla et al. (2009) and Townsend et al. (2009) using zinc-finger proteins will also promote specific or targeted gene transfer, and avoids unwanted or unnecessary pieces of DNA movement to the final crop of interest.
2.7 Orphan Crop Improvement
Orphan crop improvement largely depends on conventional techniques such as selection and hybridization as molecular techniques have not yet been implemented. Only a limited number of orphan crops breeding programs use modern techniques such as marker-assisted breeding (for details, Tadele 2009). In addition, genomic information such as whole-genome sequences is not yet available for the majority of orphan crops. This situation is rapidly changing with the increased interest in orphan crops. One example is the draft cassava genome sequence which was recently released (Cassava Genome Project 2010, http:://www.phytozome.net/cassava). In addition, comparative mapping studies have revealed that the genomes of plant species within families are highly conserved. Therefore, orthologous genes from orphan crops can be identified and isolated based on information from sequenced major crops (Moore et al. 1995; Schmidt 2000; Choi et al. 2004; Devos 2005; Jung et al. 2009).
In summary, research on orphan crops needs to focus not only on conventional approaches but also on modern and high-throughput techniques. One of the most promising modern techniques is called TILLING (Targeting Induced Local Lesion IN Genomes) which will be described in detail in the following sections.
3 TILLING: An Efficient and Rapid Method of Mutation Discovery
TILLING is a non-transgenic and relatively low-cost reverse genetics method which uses traditional mutagenesis followed by high-throughput mutation detection. In contrast to the forward genetics approaches where mutants are first identified based on phenotypes, in reverse genetics the candidate genes responsible for the expected phenotype are first discovered. TILLING identifies single base pair changes, or small deletions, in specific targeted genes (McCallum et al. 2000; Till et al. 2003; Henikoff et al. 2004; Comai and Henikoff 2006).
3.1 History of TILLING and its Applications
TILLING was developed in the year 2000 in the model plant Arabidopsis thaliana (McCallum et al. 2000). Nowadays, the technique is successfully adapted to numerous animal and plant species, since it can be applied to all organisms independent of the genome size, reproductive system, generation time, and ploidy level (McCallum et al. 2000; Till et al. 2003; Henikoff et al. 2004; Comai and Henikoff 2006). While TILLING identifies induced mutations in mutagenized populations, the modified form known as EcoTILLING detects naturally occurring SNPs, especially in landraces and wild accessions (Comai et al. 2004; Haughn and Gilchrist 2006). The latter has additional applications in genetic mapping, breeding and genotyping, and also provides information concerning gene structure, linkage disequilibrium, population structure, or adaptation (Haughn and Gilchrist 2006).
TILLING has proved to have additional benefits in addition to identifying polymorphism in genomes. The first part of TILLING requires the development of large number of mutagenized populations. Mutagenesis has been widely applied by breeders for many decades as a conventional improvement technique, and it has enabled the release of many crop cultivars (Ahloowalia et al. 2004; Wang et al. 2006). However, through TILLING, this random mutagenesis is better exploited by screening for mutations in defined genes controlling the trait of interest (Wang et al. 2006). Since TILLING directly introduces genetic variation on improved or elite germplasm, it avoids the need for introgression of a mutant allele in a non-adapted background into current high-yielding varieties and avoids the problem of linkage drag. Therefore, the introduction of agriculturally undesirable traits is avoided (Slade and Knauf 2005; Uauy et al. 2009; Sestili et al. 2010).
So far, TILLING and EcoTILLING have been implemented in the following crops: maize (Till et al. 2004a); wheat (Slade et al. 2005; Dong et al. 2009a, b; Uauy et al. 2009; Tsai et al. 2011); rice (Sato et al. 2006; Raghavan et al. 2007; Till et al. 2007a; Suzuki et al. 2008); barley (Caldwell et al. 2004; Talamè et al. 2008; Lababidi et al. 2009); sorghum (Xin et al. 2008); soybean (Cooper et al. 2008); pea (Triques et al. 2007); potato (Elias et al. 2009); and orphan crops such as banana (Till et al. 2010); and tef (Tadele 2009; Esfeld et al. prep., Table 6.1).
3.2 The TILLING Method
The TILLING procedure includes the following steps: mutagenesis, development of a non-chimeric population, DNA isolation, and mutation detection (Comai and Henikoff 2006; Tadele et al. 2009; Wang et al. 2010). EcoTILLING follows a similar set of procedures except for using natural populations as the experimental samples instead of induced mutagenized populations. The standard TILLING procedures are presented in the following sections and are summarized in Fig. 6.1 (see also Wang et al. 2010).

Overview of a standard TILLING protocol. a The initial step is establishing a mutant population and assembling the DNA of individual M2 plants into pools and plates for efficient screening. b The mutation detection step involves cleaving of PCR products followed by resolution on gel based systems, such as LI-COR. c The last step involves the confirmation of the putative SNP through direct sequencing, characterizing SNPs based on their predicted effect on protein function (using PARSESNP and SIFT), and the subsequent phenotypic characterization and deployment into breeding programs
3.2.1 Mutagenesis: The Critical Step in Generating Experimental Material
As already mentioned, conventional mutagenesis has a long history in crop breeding and thus the broad experience simplifies its application (Henikoff et al. 2004; Comai and Henikoff 2006; Tadele et al. 2009). Mutagenesis can be applied to all species including orphan crops, even if they lack advanced genetic tools (Cooper et al. 2008). Since no exogenous DNA is introduced into the plant, mutagenesis is considered as non-transgenic. The products are, therefore, exempted from regulatory restrictions or procedures imposed on transgenic products (Slade and Knauf 2005; Till et al. 2007b; Cooper et al. 2008).
3.2.1.1 Selection of the Mutagen
Broadly, mutagens are grouped under chemical and physical agents. Commonly used chemical mutagens include ethyl methanesulfonate (EMS), sodium azide (NaN3), N-methyl-N-nitrosourea (MNU), methyl methanesulfonate (MMS), hydrogen fluoride (HF), and hydroxylamine (NH2OH). Physical mutagens such as gamma-ray, x-ray, and fast neutrons are also widely used. In general, mutagens are known to create diverse types of mutations in the genome which range from point mutations to double-strand breaks. Since TILLING detects nucleotide polymorphisms, mutagens which create point mutations are preferentially selected to generate populations for screening. A mutagen proved to induce point mutations and widely used in developing TILLING populations is EMS (Caldwell et al. 2004; Slade et al. 2005; Xin et al. 2008). EMS normally creates G:C to A:T transitions in the genome, due to the alkylation of G nucleotide residues which then pairs with T instead of C (Comai and Henikoff 2006; Parry et al. 2009). In Arabidopsis, wheat, and maize about 99 % of EMS induced point mutations are transitions, however, in rice and barley these figures range between 70–84 % (see Cooper et al. 2008). Some studies showed that in rice and barley mutagens such as sodium azide and MNU were more efficient in introducing mutations (Cooper et al. 2008; Suzuki et al. 2008; Talamè et al. 2008).
Mutagenesis is the critical step, since the balance between optimum mutation density and a feasible germination rate contribute to the success of the TILLING population (Haughn and Gilchrist 2006; Parry et al. 2009). Before embarking on large-scale mutagenesis, pilot studies should be made to determine the right mutagen, optimum concentration, and handling procedures (Tadele et al. 2009). While working with mutagens, it is also important to consider the safety precaution measures, since mutagens are carcinogenic and extremely toxic to living organisms, including humans.
In general, the optimum concentration of the mutagen is lower in diploid species than in polyploids, as the former have a lower tolerance towards mutagens than the latter. The high tolerance of polyploids to mutagen is mainly due to complementation of essential genes by homoeologous copies. As a consequence, to obtain the same number of mutants, larger sized populations need to be screened in diploid species than in polyploid species (Parry et al. 2009).
3.2.1.2 Developing Mutagenized Population
Once the type and optimum concentration of the mutagen is identified, large-scale mutagenesis is made using the explant of choice. In most crop plants, the seeds are used for mutagenesis except for maize, where the pollen grains are mutagenized using EMS followed by introgression to the un-mutagenized female parent (Comai and Henikoff 2006; Till et al. 2007b). The first generation of mutagenized seeds (defined as M1 population) is typically chimeric; i.e., different cells make different genotypes, due to the multicellular stage of embryos in seeds (Tadele et al. 2009). Hence, M1 plants are self-pollinated to establish the subsequent M2 populations. At this point, the mutations do not segregate within cells of an individual plant, and therefore the induced changes are considered stable and heritable (Till et al. 2007b; Cooper et al. 2008). Tissue samples are collected from individual M2 plants for DNA isolation, and M3 seeds are harvested and stored for subsequent studies (Henikoff et al. 2004).
3.2.2 DNA Sampling and Pooling
Tissue from individual M2 lines is used for genomic DNA isolation using either high-throughput 96-well plate procedures or small scale methods. After DNA isolation, the quality and quantity of the DNA needs to be checked. The easiest and best way to do this is by loading an aliquot of DNA on electrophoresis gels, as the quality of DNA can be easily observed on an agarose gel. The DNA concentration can also be estimated by comparing it to DNA markers of known concentration. Once the concentration of each sample is estimated, the DNA samples are normalized to the same concentration level to ensure equal representation of each sample within a DNA pool and also later as template in the PCR reaction. After achieving identical DNA concentration for all samples, pooling takes place as this is a pre-requisite to detect mutations. DNA samples are pooled from 2-fold up to 8-fold in a one- or two-dimensional range to reduce screening costs and time, (Till et al. 2007b; Tadele et al. 2009). One can either pool one wild-type (wt) and one mutant DNA, however, most often individual mutant plants are pooled together. Since only one plant of the pool is likely to carry a mutation in the target sequence it ensures that wt and mutant sequences are represented in a pool, while simultaneously screening in a time and cost efficient manner (Haughn and Gilchrist 2006). The dimension refers to the way of pooling as each sample is either represented once on a plate or at two different unique positions. Two-dimensional pooling has the additional advantage of reducing false positives, since candidate mutants are visualized at two independent positions (Rigola et al. 2009; Tadele et al. 2009). Furthermore, individual mutants can be directly targeted in a one-step screening, as the pools do not have to be deconvoluted and re-analysed to identify the actual mutant individual within the pool. On the other hand, two-dimensional pooling decreases the high-throughput, since more pools have to be analysed. Still, the advantage of direct single candidate detection and avoidance of false positives often justifies the additional effort of that particular pooling scheme.
For EcoTILLING, the pooling strategy is different. Here, DNAs from two genotypes are combined in a ratio of 1:1, and in general a single genotype is used as the reference for all lines. This different pooling strategy reflects the fact that more SNPs are expected between distantly related genotypes (Raghavan et al. 2007). Therefore, pooling of individuals with too many nucleotide differences, as expected if genotypes were pooled, would reduce the detection efficiency. In addition, the use of a common reference genotype for all the collection allows the immediate identification of haplotypes within the diverse germplasm and a targeted re-sequencing of only the unique haplotypes.
3.2.3 PCR Amplification
TILLING is used to screen for mutations in specific genes that are expected or known to correspond to the trait of interest. With an established TILLING population, virtually any gene can be targeted, however, some prerequisites have to be met.
3.2.3.1 Primer Design
Designing the right primer pair is crucial and sometimes a challenge in TILLING projects. Primers need to be specific to amplify only the gene of interest and the appropriate gene region. Multiple amplifications reduce the detection efficiency. Primer specificity is especially important for members of multi-gene families, or in polyploid species where multiple homoeologues are present (Slade and Knauf 2005; Tadele et al. 2009; Uauy et al. 2009). This is commonly achieved by using copy-specific primers. In polyploid species, specificity can be achieved by designing primers in more divergent regions, particularly in the introns or in the 5′ and 3′ UTR region (Slade et al. 2005; Uauy et al. 2009; Sestili et al. 2010). In addition, in cases where only a few differences between homologues or regions exist, appropriate primers can be selected based on the following two criteria: (1) aligning the first nucleotide from the 3′ end of the primer to the genome-specific SNP, and (2) introducing a mismatch in the primer at the third or fourth position from the 3′ end (Uauy et al. 2009).
The problem of multiple target PCR amplification was also solved using a High Resolution Melting (HRM) analysis of mixed PCR amplicons containing homoeologous gene fragments and sequence analysis using Mutation Surveyor® software that simultaneously detects mutations in homoeologous genes (Dong et al. 2009b; Ishikawa et al. 2010). Although not widely implemented at present, the third option of avoiding unspecific PCR amplification is made by pre-treating the DNA with a restriction enzyme that specifically cuts only a single copy of the genome (Cooper et al. 2008).
Although prior information of the genome sequence is not required to perform TILLING, the presence of full-length genomic sequence for the gene of interest improves the chance of success. The knowledge or presence of the genome sequence accelerates the development of suitable targets, especially the designing of effective primers (Henikoff et al. 2004; Parry et al. 2009).
Depending on the detection method, the length of the amplified products can range from 0.3 to a maximum of ~3 kb (Slade and Knauf 2005; Raghavan et al. 2007). To ensure that the appropriate gene region is amplified by PCR, the CODDLE (Codons Optimized to Detect Deleterious LEsions; http://www.proweb.org/coddle/; accessed August 2011) software can be used. CODDLE defines the best region to be screened for mutations by detecting functional domains or domains which are likely to be the most sensitive to amino-acid substitutions. The use of CODDLE increases the probability of detecting deleterious mutations in the gene of interest, obtaining regions with high frequency of stop codons, and those which are evolutionary conserved. Hence it is useful for providing an allelic series of mutations (Colbert et al. 2001).
3.2.3.2 PCR Amplification and Hetero-duplex Formation
To assure the specificity and to maximise the yield of PCR products for the subsequent steps, most TILLING projects use touch-down PCR protocols. Non-specific amplification can lead to additional heteroduplex formation, reducing the detection frequency and resulting in cleaved products even in control samples (Raghavan et al. 2007). Copy-specific primers deduced from unique gene regions overcome this problem and allow a normal PCR protocol to be used, resulting in an appropriate amount of the specific PCR product for mutation detection. Proof-reading polymerases can be used (although the costs are high), but are not necessary in our experience.
The PCR amplification is followed by the heteroduplex formation step where the PCR amplified products are first denatured and then allowed to slowly cool, which facilitates the formation of heteroduplex molecules (see Fig. 6.1; Till et al. 2003; Gilchrist and Haughn 2005). This is usually introduced as an additional step at the end of the PCR program.
3.2.4 Mutation Detection
Mutations can be detected using different techniques (Fig. 6.2). Initially, the detection of single base pair differences was done using denaturing HPLC (DHPLC; McCallum et al. 2000), whereas nowadays single-strand cleaving endonucleases are used to detect mismatches in heteroduplexes (Henikoff et al. 2004; Gilchrist and Haughn 2005). Several single-strand specific nucleases, members of the S1 nuclease family (e.g. CEL I or mung bean nuclease), recognize and cleave the mismatches formed in heteroduplexes (Till et al. 2004b; Comai and Henikoff 2006).

Three options of gel-based mutation detection methods in TILLING: (a) LI-COR DNA analyzer, (b) agarose gel, and (c) non-denaturing PAGE gel. In the LI-COR method, products are PCR amplified using infra-red dye labelled primers, and digested using endonucleases. They are then loaded on a denaturing polyacrylamide gel for mutation detection and visualized as complementary bands in the gel images produced for each of the dyes (as shown above for channels 700 and 800). The agarose (b) and non-denaturing polyacrylamide gels (c), however, do not require labelled primers, since mutations can be visualized on gels stained with ethidium bromide. Yellow arrows indicate the presence of complementary bands corresponding to the two digested products originating from a mutant individual within the pooled DNA samples
The comparison among different nucleases in terms of efficiency has been studied (Till et al. 2004b). CEL I is the most commonly used and preferred enzyme for mutation detection in TILLING projects (Till et al. 2004b, 2006), despite a slight preference of CEL I in cleaving specific types of mismatches (see Triques et al. 2007). CEL I cleaves to the 3′ side of mismatches in heteroduplexes while leaving homoduplexes intact (Henikoff et al. 2004). Thereby, CEL I cleaves each strand of the heteroduplex at the site of the mismatch that produces two complementary fragments (Haughn and Gilchrist 2006). Although CEL I can be purchased through commercial suppliers (Surveyor Mutation Detection Kit; Transgenomic®), the majority of TILLING platforms isolate the enzyme from celery stalks (celery juice extract; CJE) through an easy, cheap, and very robust extraction method established by Till et al. (2006). Recently, other endonucleases such as Brassica petiole extract (BPE; Sato et al. 2006) and ENDO1, which is extracted from A. thaliana have been proven to be as efficient, or even more efficient than CEL I in cleaving heteroduplexes (Triques et al. 2007).
Mutation detection after cleavage of heteroduplexes can be done using different methods. The most commonly used method is through a denaturing polyacrylamide gel run on a LI-COR DNA analyser (referred as LI-COR, Fig. 6.2a). For this approach, PCR products are amplified using infra-red dye (IRD) labelled primers. Both, the forward- and reverse- primers are labelled at the 5′ end with a specific dye that can be detected in one of the two channels of the LI-COR. Since the amplification efficiency of labelled primers is lower than that of unlabelled primers, the addition of both labelled and unlabelled primers in every PCR reaction is required to obtain a high-yielding PCR product. After PCR amplification and endonuclease digestion, products are purified using either Sephadex® purification (Till et al. 2006) or ethanol precipitation.
The purified samples are loaded on 5–6 % denaturing polyacrylamide gels. The common loading is done by pipetting the samples first in a comb loading tray (Gel Company Inc, San Francisco, USA) before they are loaded to a membrane comb (Gel Company Inc) by immersing the comb into the samples in a tray. A labelled fragment of defined size could be applied at specified distance in order to facilitate the counting of the lanes. Before loading the samples onto the gel, a pre-run of the LI-COR is made for about 20 min. After the successful pre-run, samples are loaded onto the membrane comb and then the comb is inserted on the top of the gel. The run-time for the gel is dependent on the product size. Since forward- and reverse primers are labelled with different infra-red dyes, cleaved products are visible in both channels of the LI-COR (Fig. 6.1). The sum of the two cleaved products gives the size of the original PCR product. The GelBuddy program (http://www.proweb.org/gelbuddy; accessed April 2011) is used to automate band calling in polyacrylamide gels (Till et al. 2006).
Modified mutation detection methods have been developed by various TILLING platforms. These methods include detection by agarose (Fig. 6.2b) and non-denaturing polyacrylamide gels (Fig. 6.2c; Sato et al. 2006; Raghavan et al. 2007; Dong et al. 2009a; Uauy et al. 2009). These alternative methods have applications especially in developing countries, where resources are limited and frequent power failures result in complete loss of data when using a LI-COR system. The information or data from the electrophoresis systems can easily be recovered, even after multiple power interruptions. In addition, the LI-COR system is not affordable in many individual labs as it uses expensive primers and equipment.
Alternative detection methods which do not require labelled primers can be easily adopted in developing countries laboratories as well as for orphan crop improvement. Raghavan et al. (2007) and Dong et al. (2009a) used conventional agarose gels for mutation detection, a method appropriate for low-budget and ill-equipped laboratories. Another modification by Uauy et al. (2009) used non-denaturing polyacrylamide gels stained with ethidium bromide to detect mutations. Advantages of these alternative TILLING methods include the use of only unlabelled primers which are easier to obtain and handle. In addition, the size of the PCR product can be increased to 2–3 kb (instead of a maximum of 1.5 kb in the case of the LI-COR). Furthermore, no prior purification of the digested samples is necessary, which further reduces the cost and time (Raghavan et al. 2007; Dong et al. 2009a). The lower costs and simplicity of the technique allows further applications in germplasm characterization and mapping studies (Raghavan et al. 2007). Despite all these benefits in terms of ease of use and relative simplicity, alternative methods are not as sensitive as the LI-COR and do not provide an exact location or site of the mutation within the target amplicon.
In addition to the above mentioned methods, Next Generation Sequencing (NGS) platforms have recently been implemented to detect mutations in TILLING populations (Rigola et al. 2009; Tsai et al. 2011). Rigola et al. (2009) used the 454-FLX platform (Roche Applied Science) to discover mutants in the tomato eIF4E gene, by screening more than 3,000 families in a single sequencing run. Tsai et al. (2011) applied the Illumina GA sequencing platform to identify mutations in populations of rice and wheat. Conventional endonuclease based TILLING projects are limited to eight samples per pool because of the detection efficiency. As a consequence, analyses with more than eight samples per pool might result in data noises (Rigola et al. 2009). Some of the benefits of sequencing based TILLING approaches are: (1) the sequence of each mutation and its impact on protein sequence is directly determined, (2) it does not rely on either labelled primers or endonucleases, (3) it is based on an objective statistical method and not on visual inspections, (4) it is flexible with respect to changing numbers of samples and amplicons, and (5) since it is based on highly redundant sequencing, the likelihood of identifying false positives is reduced (Rigola et al. 2009; Tsai et al. 2011). Despite these advantages, TILLING by sequencing requires access to expensive and technically challenging NGS equipment and experienced bioinformaticians for data analysis. Due to these reasons, TILLING by sequencing is currently implemented mainly by service providers. An overview summarizing the advantages and disadvantages of various TILLING strategies is shown in Table 6.2.
3.2.5 Confirmation by Sequencing
Mutations detected by gel-based TILLING methods need to be confirmed by sequencing. If the LI-COR is used, the detected mutation pinpoints directly to the location of the polymorphism and makes confirmation by sequencing quite efficient. The labelled primers on the LI-COR provide a directionality (5′ or 3′ end), which allows the sequencing reaction to target the specific site (Gilchrist and Haughn 2005; Till et al. 2007b). On the other hand, the alternative screening methods, which use unlabelled primers, do not provide an exact position of the mutation. Since the size of the fragments on the gels indicates only a putative but not exact location, sequencing should be carried out in order to pinpoint the mutation. In addition, the longer fragments (~2–3 kb) that can be amplified, imply that sometimes an additional sequencing primer within the amplicon might be needed to reveal the mutation.
Mutations are randomly induced and can target every gene (Colbert et al. 2001; Parry et al. 2009). The advantage of the chemical mutagenesis in TILLING is that it creates an allelic series of mutations. These include (1) nonsense mutations: a single base pair change converts an amino acid codon into a stop codon, (2) missense mutations: a single base pair change alters the amino acid encoded by a particular codon, (3) silent mutations: a single base pair change does not alter the amino acid encoded by a particular codon, and (4) splice junction mutations: a single base pair change alters the canonical GT/AG splice sites encoded within the introns and leads to mis-spliced messenger RNA, and in most cases, to a truncated protein (McCallum et al. 2000). The allelic series of induced mutations can potentially confer various phenotypes ranging from subtle to strong. Mutations in the coding region of the gene might alter plant metabolism or the effective level of a gene product that might be useful for breeding. TILLING is the only reverse genetics technique that can deliver such genetic diversity (Haughn and Gilchrist 2006) and several studies estimate the numbers or percentages of missense-, nonsense-, and silent mutations. The numbers vary slightly, but in general around 50 % of EMS-induced changes in the target are missense mutations, and ~5 % are truncations (either nonsense or splice junction mutations), whereas the remaining are either silent or located within non-coding regions (Haughn and Gilchrist 2006). Similar findings were reported for soybean (Cooper et al. 2008), Arabidopsis (Greene et al. 2003; Till et al. 2003; Till et al. 2004a), Brassica rapa (Stephenson et al. 2010), and durum- and bread-wheat (Slade et al. 2005; Dong et al. 2009a; Uauy et al. 2009).
To investigate whether the detected mutations have an influence on the protein, web-based software, namely PARSESNP (http://www.proweb.org/parsesnp/; accessed August 2011) and SIFT (http://sift.jcvi.org/; accessed August 2011) can be used. PARSESNP (Project Aligned Related Sequences and Evaluate SNPs) reveals the changes in the nucleotide and amino-acid sequences and documents any restriction endonuclease site that has been altered (Taylor et al. 2003; Gilchrist and Haughn 2005). SIFT (Sorting Intolerant From Tolerant) uses alignments between closely related sequences to predict whether the amino acid change is expected to have deleterious effects on the protein (Ng and Henikoff 2003; Henikoff et al. 2004).
3.2.6 Estimation of Mutation Frequency
The mutation frequency of a population can be calculated based on the confirmed mutations identified across several TILLING targets. This value is calculated as the total number of base pairs screened divided by the total number of mutations scored; whereby the total number of base pairs refers to amplicon size x screened individuals. The amplicon size needs to be adjusted by excluding the regions close to either primer (roughly 5–10 % of the total size), due to technical limitations in mutation detection.
The highest mutation density was obtained from the two polyploid wheat species, namely bread wheat and durum wheat. The mutation frequencies reported were one mutation per 25 kb in hexaploid bread wheat and one mutation per 40 kb in tetraploid durum wheat (Slade et al. 2005). The mutation density reported for other major crops were 1/374 to 1/1,000 kb for barley (Caldwell et al. 2004; Talamè et al. 2008), 1/300 to 1/600 kb for rice (Till et al. 2007a), and 1/526 kb for sorghum (Xin et al. 2008). According to Weil (2009), a mutation rate of 1/500 kb or less is considered feasible in TILLING. Higher mutation frequencies imply that the number of individuals to be screened in order to obtain a certain number of mutations is too large to make TILLING a practical endeavour. Table 6.1 shows mutation frequencies for different crop species. In general, mutation frequencies are determined by the type and concentration of the mutagen, genetic backgrounds as well as the gene region screened (Uauy et al. 2009). Hence, specific criteria should be applied to calculate the mutation frequency, so that the values obtained are comparable.
4 Beyond TILLING: What Follows Mutation Detection
Most TILLING and EcoTILLING projects on different crops so far are implemented as a proof-of-concept to test the applicability of the technique. Hence, the projects ceased once the intended goals were achieved. Several studies, however, implemented TILLING directly for crop improvement. Among these, granule-bound starch synthase genes responsible for producing waxy wheat were investigated in TILLING populations. Here, two mutation detection systems were used, namely LI-COR and thin (≤4 mm) agarose gels (Slade et al. 2005; Dong et al. 2009a). Although both detection methods enabled the development of complete waxy wheat, the latter method took only 18 months after crossing two truncation mutants obtained from the TILLING population (Dong et al. 2009a). Several other groups have targeted the starch branching enzyme gene (SBE-IIa) in wheat, as reduced transcript levels of this gene by RNA interference (RNAi) led to transgenic wheat with high amylose (Regina et al. 2006). The initial proof-of-concept by RNAi provides strong evidence that a TILLING based approach will work. Currently, many of these lines are being backcrossed and combined for evaluation (Uauy et al. 2009). The use of TILLING for crop improvement is also being pursued by companies such as Arcadia Biosciences (http://www.arcadiabio.com/), who are developing tomato lines that suffer less postharvest loss. The number of examples continue to increase with rapid advances in the knowledge of gene function (which should provide new targets for TILLING) and as several mutants make their way through breeding programs.
To apply TILLING in crop improvement, mutants discovered from screening need to be investigated further. Phenotypic screening will usually be the first option to link mutations in the candidate gene with the trait of interest. To save time and cost, only the most promising candidates should be screened, i.e. those that encode for nonsense-, splice- junctions- or radical missense- mutations as determined by PARSESNP and SIFT. Evidence of a causal effect between the target gene and the trait of interest is further advanced, if independent mutant alleles result in the same phenotype (Haughn and Gilchrist 2006). However, phenotypic screening is a challenge in polyploid species, since genetic buffering makes it less likely that recessive mutations reveal the expected phenotype. Therefore, it may be necessary to identify mutations in each homoeologous copy of the targeted gene and combine them by crossing (Parry et al. 2009). In addition to the use of the mutant alleles for breeding, backcrossing to the original line, or introgression to other elite cultivars is also necessary. The multiple backcrosses to the recurrent parent remove unlinked mutations that could lead to undesirable phenotypes (Slade and Knauf 2005; Sestili et al. 2010). Several methods exist to prove if the desired mutation is still in the subsequent generation. These include, (1) CAPS (Cleaved Amplified Polymorphic Sequence) or dCAPS (Derived Cleaved Amplified Polymorphic Sequences) markers that take advantage of restriction sites, (2) sequencing of the mutation region, (3) using TILLING with the parents as reference preferentially using the alternative TILLING strategy on agarose gels, and (4) designing SNP-specific assays such as KASPar (Allen et al. 2011). Marker-assisted backcrossing (MABC) can help to introduce the desired mutation in widely-adapted and high-yielding cultivars. In summary, the use of MABC to remove background mutations is needed to breed a stable line with good agronomic performance.
5 Application of TILLING and EcoTILLING to Orphan Crops
So far, TILLING and EcoTILLING are not widely implemented in orphan crops. However, there are some initiatives to apply these methods and we present some of these projects below.
5.1 Cassava (Manihot esculenta)
Conventional cassava breeding is challenging, due to limited genetic variation in elite germplasm. Therefore, radiation mutation of selected elite lines as well as wide crosses involving wild Manihot species and castor bean are done to broaden the genetic base of the germplasm pool, and expand the industrial uses of cassava. In collaboration with the International Atomic Energy Agency (IAEA) three different methods of irradiation have been used for inducing mutations. About 4,000 seeds from six different cassava clones were irradiated with gamma rays or with fast neutrons, which led to the harvest of about 5,000 M2 seeds (Ceballos et al. 2010). The International Institute of Tropical Agriculture (IITA; http://liferay.iita.org/web/mab/home/tilling;jsessionid=8166693C8CBE40D02B12544406EB36AF; accessed August 2011) is conducting a pilot study on EMS-induced mutagenesis to focus on a single trait. However, once the protocol is established, it shall be adapted to discover mutations for other important traits, and to other crops of the IITA. The aims of the project are (1) identification of mutants with desired traits that will be incorporated into the existing cassava improvement program, (2) marker development for MAS, and (3) to design large-scale reverse-genetics experiments to tag genes controlling desirable traits in cassava and other IITA mandate crops. The expected outputs are to (1) establish a catalogue of characterized mutant population, (2) identify DNA sequences of candidate genes as basis for marker development, and (3) to develop standard operating procedures (SOPs) for TILLING using IRD-labelled primers on a gene analyzer instrument.
5.2 Banana and Plantain (Musa spp)
Banana provides staple food for approximately 400 million people and is important for both export and local markets. The EcoTILLING project on banana and plantain utilized 80 accessions to discover and characterize nucleotide polymorphisms. From these accessions alone, over 800 novel alleles were obtained by tilling 14 gene targets (Till et al. 2010). Thus, EcoTILLING contributes to understanding genetic diversity, and improving Musa for food production. Till et al. (2010) used the technique for the simultaneous discovery of heterozygous and homozygous polymorphisms in diploid accessions, which permit fast evaluation of nucleotide diversity in accessions of the same genome type. The high base pair resolution allows the grouping of accessions based on shared banding patterns, and it is a fast method to compare and barcode a large number of accessions at relatively low cost. It can also be used to tag accessions in gene banks and for classification of newly acquired samples, and even phylogenetic approaches are possible. Additionally, EcoTILLING can be applied for functional genomic studies and mutation breeding (Nieto et al. 2007) and natural allele mining can serve as primary selection for material to be used in breeding programs. In addition to the intensive use of EcoTILLING, an EMS-mutagenized TILLING population of the triploid ‘Grande Naine’ was developed and large-scale efforts are currently being made (Till et al. 2010).
5.3 Tef (Eragrostis tef)
The Tef Improvement Project (TIP) focuses on tackling lodging, the major yield limiting factor in tef cultivation. Developing lodging resistant semi-dwarf tef cultivars is the main goal of the TIP next to developing drought tolerant cultivars, since moisture scarcity is becoming widespread in Ethiopia. So far around 4,500 M2 individuals were screened for several genes of interest (Esfeld et al. in prep). One of the major obstacles related to tef TILLING is the presence of two homoeologous gene copies due to the allotetraploid nature of tef. However, this particular problem is overcome by designing copy-specific primers for each gene of interest and amplifying one copy at a time (see Sect. 6.3.2.3.1. for further detail).
5.4 Pearl Millet (Pennisetum glaucum)
Pearl millet is one of the drought tolerant crops widely grown in Asia and Africa. The International Crop Research Institute for Semi-Arid Tropics (ICRISAT, http://www.icrisat.org/bt-gene-discovery.htm; accessed August 2011) produced a TILLING population using the inbred line “P1449-2-P1”. A total of 31,000 seeds were mutagenized in three different batches, using 5.0, 7.5, 9.0, and 10.0 mM EMS. DNA from at least 7,458 M2 TILLING lines is available, and this number is currently being increased to 10,000 M2 lines, as this was calculated to be the ideal population size for mining allelic variants in candidate genes. Eight-fold pooling of normalized genomic DNA from the 7,458 M2 lines has been completed. The pools of DNA will be made available for the international pearl millet community (http://www.icrisat.org/bt-gene-discovery.htm), however, the latest progress is not communicated on the web site.
5.5 Chickpea (Cicer arietinum)
Chickpea is an important food legume, extensively cultivated in Asia and Africa. Next to pearl millet, chickpea was chosen for TILLING at the ICRISAT (http://www.icrisat.org/bt-gene-discovery.htm; accessed August 2011). Over 20,000 seeds were treated with the EMS and roughly 9,000 germinated, of which only 6,000 set seeds.
5.6 Mung Bean (Vigna radiata)
Mung bean is an important economic crop and dietary staple in many developing countries. Barkley et al. (2008) performed an EcoTILLING study to discover and verify DNA polymorphisms in the mung bean collection USDA-ARS PGRCU. This collection has been evaluated for various morphological characters and some of the core accessions were evaluated with SSR markers. They showed both limited genetic and morphological diversity. Thus, EcoTILLING was used to further assess V. radiata for molecular diversity, and to evaluate the EcoTILLING method for rapid detection of SNPs in plant germplasm. The intron region was used to detect the polymorphism, as this region is ideal when assessing polymorphism in a species with high genetic identity. Thus, the overall aim of the study was to reveal variation among V. radiata var. radiata accessions, and its progenitor V. radiata var. sublobata, mine for SNPs within the V. radiata var. radiata accessions, and determine the number of different haplotypes in the collection. A total of 157 DNA polymorphisms were detected from ten primer sets, which resulted in 45 haplotypes. Numerous DNA polymorphisms between the two varieties were found, however, the core radiata collection is genetically similar. The results suggest, that either V. radiata var. radiata generally has a narrow genetic base or that the collection of the USDA-ARS PGRCU has to be expanded. Subsequent studies might use the obtained data to develop SNP markers that can be advantageous in MAS.
6 Conclusions
TILLING and EcoTILLING are high-throughput mutation detection systems that exploit non-transgenic allelic variation. The technique has proven highly efficient in obtaining desirable mutant lines and is also extremely versatile. This includes the possibility to develop many in-house protocols and reagents such as CJE, and the ability to implement detection methods using both basic technology platforms, such as agarose gels, as well as NGS equipment. The flexibility of the system has allowed the technology to become widely adopted in many crop plants, including maize, wheat, and rice since its first development in Arabidopsis a decade ago. Consequently, there is also growing interest in applying the technique in orphan crops that lack well developed genetic tools.
There are still several major challenges to overcome. The first is, the expansion of these approaches to orphan crops and the training of local expertise to implement these technologies. The development of TILLING requires support from local agencies, so that the populations generated, and downstream techniques and supplies, can be used efficiently by the research community. The second is, the development of NGS approaches and the associated technologies, such as RNAseq and genome capture which will greatly increase the possibility to rapidly access mutations in orphan crops. The possibility to rapidly sequence gene space by RNAseq and the ability to capture these sequences from mutant populations (via genome capture) now opens the possibility to re-sequence complete TILLING populations. Although these efforts require large financial and technical investments at the moment, we expect that in the near future the in silico cataloguing of mutations for a complete TILLING population will be a routine practice. Once mutations of interest have been obtained, the major challenge of shifting these mutations into breeding lines or varieties with advanced agronomic performance for resource poor farmers remains. It is only at this point, that TILLING will be delivering on the promise of improving food security.
Abbreviations
- AFLPs:
-
Amplified fragment length polymorphisms
- CAPS:
-
Cleaved amplified polymorphic sequence
- CJE:
-
Celery juice extract
- CODDLE:
-
Codons optimized to detect deleterious lesions
- dCAPS:
-
Derived cleaved amplified polymorphic sequences
- EMS:
-
Ethyl methanesulphonate
- HF:
-
Hydrogen fluoride
- IRD:
-
Infra-red dye
- MAB:
-
Marker-assisted breeding
- MAS:
-
Marker-assisted selection
- MNU:
-
N-methyl-N-nitrosourea
- ODAP:
-
β-N-Oxalyl-L-α, β-diaminopropanoic acid
- PARSESNP:
-
Project aligned related sequences and evaluate SNPs
- SIFT:
-
Sorting intolerant from tolerant
- SNPs:
-
Single nucleotide polymorphism
- SSR:
-
Simple sequence repeats
- TILLING:
-
Targeting induced local lesion IN genomes
References
Ahloowalia BS, Maluszynski M, Nichterlein K (2004) Global impact of mutation-derived varieties. Euphytica 135:187–204
Allen AM, Barker GLA, Berry ST, Coghill JA, Gwilliam R, Kirby S, Robinson P, Brenchley RC, D’Amore R, McKenzie N, Waite D, Hall A, Bevan M, Hall N, Edwards KJ (2011) Transcript-specific, single-nucleotide polymorphism discovery and linkage analysis in hexaploid bread wheat (Triticum aestivum L.). Plant Biotechnol J. doi:10.1111/j.1467-7652.2011.00628.x
Al-Safadi B, Arabi MIE (2007) In vitro induction, isolation and selection of potato mutants tolerant to salinity. Adv Hort Sci 20:127–132
Asiwe, JAN (2009) Field evaluation of bambara groundnut germplasm for important traits and resistance to root knot nematode infestation. In: Tadele Z (ed) New approaches to plant breeding of orphan crops in Africa: Proceedings of an International Conference, Bern, Switzerland. Stämpli AG, Bern, 19–21 Sept 2007
Assefa K, Belay G, Tefera H, Yu J-K, Sorrells ME (2009) Breeding tef: conventional and molecular approaches. In: Tadele Z (ed) New approaches to plant breeding of orphan crops in Africa: Proceedings of an International Conference, Bern, Switzerland. Stämpli AG, Bern, 19–21 Sept 2007
Ayalew M, Stewart N (2005) Overexpression of Arabidopsis thaliana ABC transporter confers kanamycin resistance to transgenic plants. Nat Biotechnol 23:1177–1180
Baenziger PS, Russell WK, Graef GL, Campbell BT (2006) Improving lives: 50 years of crop breeding, genetics and cytology. Crop Sci 46:2230–2244
Bal U, Abak K (2007) Haploidy in tomato (Lycopersicon esculentum Mill.): a critical review. Euphytica 158:1–9
Bapat VA, Yadav SR, Dixit GB (2008) Rescue of endangered plants through biotechnological applications. Natl Acad Sci Lett 31:201–210
Barkley NA, Wang ML, Gillaspie AG, Dean RE, Pederson GA, Jenkins TM (2008) Discovering and verifying DNA polymorphisms in a mung bean [V. radiata (L.) R. Wilczek] collection by EcoTILLING and sequencing. BMC research notes: 1:28. doi:10.1186/1756-0500-1-28
Borlaug NE (2007) Sixty-two years of fighting hunger: personal recollections. Euphytica 157:287–297
Boukar O, Fatokun C (2009) Strategies in cowpea breeding. In: Tadele Z (ed) New approaches to plant breeding of orphan crops in Africa. In: Proceedings of an International Conference, Bern, Switzerland. Stämpli AG, Bern, 19–21 Sept 2007
Brandt SA, Spring A, Hiebsch C, McCabe JT, Tabogie E, Diro M, Wolde-Michael G, Yntiso G, Shigeta M, Tesfaye S (1997) The tree against hunger: enset-based agricultural systems in Ethiopia. American Association for the advancement of science
Caldwell DG, McCallum N, Shaw P, Muehlbauer GJ, Marshall DF, Waugh R (2004) A structured mutant population for forward and reverse genetics in barley (Hordeum vulgare L.). Plant J 40:143–150
Ceballos H, Iglesias CA, Pérez JC, Dixon AG (2004) Cassava breeding: opportunities and challenges. PlantMol Biol 56:503–516
Ceballos H, Okogbenin E, Pérez JC, López-Valle LAB, Debouck D (2010) Cassava. In: Bradshaw JE (ed) Hand book of plant breeding: root and tuber crops. Springer, New York
Chandrashekar A (2010) Finger millet: Eleusine coracana. Adv Food Nutr Res 59:215–262
Charcosset A, Moreau L (2004) Use of molecular markers for the development of new cultivars and the evaluation of genetic diversity. Euphytica 137:81–94
Chawade A, Sikora P, Bräutigam M, Larsson M, Vivekanand V, Nakash MA, Chen T, Olsson O (2010) Development and characterization of an oat TILLING-population and identification of mutations in lignin and beta-glucan biosynthesis genes. BMC Plant Biol 10:86
Choi HK, Mun JH, Kim DJ, Zhu H, Baek JM, Mudge J, Roe B, Ellis N, Doyle J, Kiss GB, Young ND, Cook DR (2004) Estimating genome conservation between crop and model legume species. Proc Natl Acad Sci USA 101:15289–15294
Colbert T, Till BJ, Tompa R, Reynolds S, Steine MN, Yeung AT, McCallum CM, Comai L, Henikoff S (2001) High-throughput screening for induced point mutations. Plant Physiol 126:480–484
Collard BC, Mackill DJ (2008) Marker-assisted selection: an approach for precision plant breeding in the twenty-first century. Philos Trans R Soc Lond B Biol Sci 363:557–572
Comai L, Henikoff S (2006) TILLING: practical single-nucleotide mutation discovery. Plant J 45:684–694
Comai L, Young K, Till BJ, Reynolds SH, Greene EA, Codomo CA, Enns LC, Johnson JE, Burtner C, Odden AR, Henikoff S (2004) Efficient discovery of DNA polymorphisms in natural populations by Ecotilling. Plant J 37:778–786
Cooper JL, Till BJ, Laport RG, Darlow MC, Kleffner JM, Jamai A, El-Mellouki T, Liu S, Ritchie R, Nielsen N, Bilyeu KD, Meksem K, Comai L, Henikoff S (2008) TILLING to detect induced mutations in soybean. BMC Plant Biol 8:9
CSA (Central Statistical Agency; 2011) Agricultural sample survey 2010/11. Statistical Bulletin 505, Addis Ababa, Ethiopia
Devos KM (2005) Updating the “Crop Circle”. Curr Opin Plant Biol 8:155–162
Dong C, Dalton-Morgan J, Vincent K, Sharp P (2009a) A modified TILLING method for wheat breeding. Plant Genome 2:39–47
Dong C, Vincent K, Sharp P (2009b) Simultaneous mutation detection of three homoeologous genes in wheat by High Resolution Melting analysis and Mutation Surveyor. BMC Plant Biol 9:143
Ejeta G (2010) African Green Revolution needn’t be a mirage. Science 327:831–832
Elias R, Till BJ, Mba C, Al-Safadi B (2009) Optimizing TILLING and EcoTILLING techniques for potato (Solanum tuberosum L.). BMC research notes 2:141. doi:10.1186/1756-0500-2-141
FAO (Food and Agricultural Organization; 2002) World agriculture: towards 2015/2030. FAO, Rome
Germana MA (2011) Gametic embryogenesis and haploid technology as valuable support to plant breeding. Plant Cell Rep 30:839–857
Getahun H, Lambein F, Vanhoorne M, Stuyft PV (2003) Food-aid cereals to reduce neurolathyrism related to grasspea preparations during famine. Lancet 362:1808–1810
Ghosh A (2011) Modern methods for selection of plants with better characteristics. http://www.biotecharticles.com/Agriculture-Article/Modern-Methods-For-Selection-of-Plants-With-Better-Characterisitcs-860.html. Accessed 15 May 2012
Gilchrist EJ, Haughn GW (2005) TILLING without a plough: a new method with applications for reverse genetics. Curr Opin Plant Biol 8:211–215
Giri CC, Shyamkumar B, Anjaneyulu C (2004) Progress in tissue culture, genetic transformation and applications of biotechnology to trees: an overview. Trees-Struct Funct 18:115–135
Godfray HCF, Beddington JR, Crute IR, Haddad L, Lawrence D, Muir JF, Pretty J, Robinson S, Thomas SM, Toulmin C (2010) Food security: the challenge of feeding 9 billion people. Science 327:812–817
Golle DP, Reiniger LRS, Curti AR, Bevilacqua CB (2009) Forestry improvement: emphasis on biotechnology application. Cienc Rural 39:1606–1613
Greene EA, Codomo CA, Taylor NE, Henikoff JG, Till BJ, Reynolds SH, Enns LC, Burtner C, Johnson JE, Odden AR, Comai L, Henikoff S (2003) Spectrum of chemically induced mutations from a large-scale reverse-genetic screen in Arabidopsis. Genetics 164:731–740
Haughn GW, Gilchrist EJ (2006) TILLING in the Botanical Garden: a reverse genetic technique feasible for all plant species. Floriculture Ornamental Plant Biotechnol 1:476–482
Henikoff S, Till BJ, Comai L (2004) TILLING. Traditional mutagenesis meets functional genomics. Plant Physiol 135:630–636
Himelblau E, Gilchrist EJ, Buono K, Bizzell C, Mentzer L, Vogelzang R, Osborn T, Amasino RM, Parkin IAP, Haughn GW (2009) Forward and reverse genetics of rapid-cycling Brassica oleracea. Thero Appl Genet 118:953–961
Hopman E, Dekking L, Blokland ML, Wuisman M, Zuijderduin W, Koning F, Schweizer J (2008) Tef in the diet of celiac patients in The Netherlands. Scand J Gatroentero 43:277–282
IFPRI (International Food Policy Research Institute (2002) Green Revolution: curse or blessing?. IFPRI, Washington
IPGRI (International Plant Genetic Resources Institute, 2004) Promoting fonio production in West and Central Africa through germplasm management and improvement of post-harvest technology. Final report. (http://www.underutilized-species.org/%20Documents/PUBLICATIONS/fonio_rapport_final_GTZ_project1.pdf).Accessed Aug 2009)
Ishikawa T, Kamei Y, Otozai S, Kim J, Sato A, Kuwahara Y, Tanaka M, Deguchi T, Inohara H, Tsujimura T, Todo T (2010) High-resolution melting curve analysis for rapid detection of mutations in a Medaka TILLING library. BMC Mol Biol 11:70
Jacobsen E, Schouten HJ (2007) Cisgenesis strongly improves introgression breeding and induced translocation breeding of plants. Trends Biotechnol 25:219–223
Jain SM (2007) Biotechnology and mutagenesis in genetic improvement of cassava (Manihot esculenta). Gene Conserve 6:329–343
Jain SM (2009) Mutation induced genetic improvement of neglected crops. In: Tadele Z (ed) New approaches to plant breeding of orphan crops in Africa. In: Proceedings of an International Conference, Bern, Switzerland. Stämpli AG, Bern, 19–21 Sept 2007
James C (2010) Global status of commercialized biotech/GM Crops: 2010. International Service for the Acquisition of Agri-biotech Applications (ISAAA) Briefs No. 42. Ithaca, NY. ISBN: 978-1-892456-49-4. http://www.isaaa.org/. Accessed Aug 2011
Jimenez VM (2005) Involvement of plant hormones and plant growth regulators on in vitro somatic embryogenesis. Plant Growth Regul 47:91–110
Jung S, Jiwan D, Cho I, Lee T, Abbott A, Sosinski B, Main D (2009) Synteny of Prunus and other model plant species. BMC Genomics 10:76. doi:10.1186/1471-2164-10-76
Kim BY, Baier AC, Somers DJ, Gustafson JP (2001) Aluminum tolerance in triticale, wheat, and rye. Euphytica 120:329–337
Lababidi S, Mejlhede N, Rasmussen SK, Backes G, Al-Said W, Baum M, Jahoor A (2009) Identification of barley mutants in the cultivar ‘Lux’ at the Dhn loci through TILLING. Plant Breeding 128:332–336
Lakshmanan P, Taji A (2000) Somatic embryogenesis in leguminous plants. Plant Biol 2:136–148
Maqbool SB, Devi P, Sticklen MB (2001) Biotechnology: Genetic improvement of sorghum (Sorghum bicolor (L.) Moench). In Vitro Cell Dev Biol Plant 37:504–515
McCallum CM, Comai L, Greene EA, Henikoff S (2000) Targeted screening for induced mutations. Nat Biotechnol 18:455–457
Minoia S, Petrozza A, D’Onofrio O, Piron F, Mosca G, Sozio G, Cellini F, Bendahmane A, Carriero F (2010) A new mutant genetic resource for tomato crop improvement by TILLING technology. BMC Res Notes 3:69
Moore G, Devos KM, Wang Z, Gale MD (1995) Grasses, line up and form a circle. Curr Biol 5:737–739
Nassar NMA (2009) Wild species to improve orphan crops: the case of cassava. In: Tadele Z (ed) New approaches to plant breeding of orphan crops in Africa. In: Proceedings of an International Conference, Bern, Switzerland. Stämpli AG, Bern, 19–21 Sept 2007
Naylor RL, Falcon WP, Goodman RM, Jahn MM, Sengooba T, Tefera H, Nelson RJ (2004) Biotechnology in the developing world: a case for increased investments in orphan crops. Food Policy 29:15–44
Ng PC, Henikoff S (2003) SIFT: Predicting amino acid changes that affect protein function. Nucleic Acids Res 31:3812–3814
Nieto C, Piron F, Dalmais M, Marco CF, Moriones E, Gómez-Guillamón ML, Truniger V, Gómez P, Garcia-Mas J, Aranda MA, Bendahmane A (2007) EcoTILLING for the identification of allelic variants of melon eIF4E, a factor that controls virus susceptibility. BMC Plant Biol 7:34
NRC (National Research Council, 1996). Finger millet. In: Lost Crops of Africa. Volume I: Grains. National Academy of Press, Washington, DC
NRC (National Research Council, 2006) Bambara bean. In: Lost crops of Africa. Volume II: Vegetables. National Academy of Press, Washington, DC
Ochoa-Alejo N, Ramirez-Malagon R (2001) In vitro chili pepper biotechnology. In Vitro Cell Dev Biol Plant 37:701–729
Padulosi S, Mal B, Ravi SB, Gowda J, Gowda KTK, Shanthakumar G, Yenagi N, Dutta M (2009) Food security and climate change: role of plant genetic resources of minor millets. Indian J Plant Genet Resour 22:1–16
Parry MA, Madgwick PJ, Bayon C, Tearall K, Hernandez-Lopez A, Baudo M, Rakszegi M, Hamada W, Al-Yassin A, Ouabbou H, Labhilili M, Phillips AL (2009) Mutation discovery for crop improvement. J Exp Bot 66:2817–2825
Porch TG, Blair MW, Lariguet P, Galeano C, Pankhurst CE, Broughton WJ (2009) Generation of a mutant population for TILLING common bean genotype BAT 93. J Amer Soc Horti Sci 134:348–355
Raghavan C, Naredo MEB, Wang HH, Atienza G, Liu B, Qiu FL, McNally KL, Leung H (2007) Rapid method for detecting SNPs on agarose gels and its application in candidate gene mapping. Mol Breed 19:87–101
Regina A, Bird A, Topping D, Bowden S, Freeman J, Barsby T, Kosar-Hashemi B, Li Z, Rahman S, Morell M (2006) High-amylose wheat generated by RNA interference improves indices of large-bowel health in rats. Proc Natl Acad Sci USA 103:3546–3551
Rigola D, van Oeveren J, Janssen A, Bonné A, Schneiders H, van der Poel HJ, van Orsouw NJ, Hogers RC, de Both MT, van Eijk MJ (2009) High-throughput detection of induced mutations and natural variation using KeyPoint technology. PLoS ONE 4:e4761
Sato Y, Shirasawa K, Takahashi Y, Nishimura M, Nishio T (2006) Mutant selection from progeny of gamma-ray-irradiated rice by DNA heteroduplex cleavage using Brassica Petiole extract. Breeding Sci 56:179–183
Schmidt R (2000) Synteny: recent advances and future prospects. Curr Opin Plant Biol 3:97–102
Sestili F, Botticella E, Bedo Z, Phillips A, Lafiandra D (2010) Production of novel allelic variation for genes involved in starch biosynthesis through mutagenesis. Mol Breeding 25:145–154
Shukla VK, Doyon Y, Miller JC, DeKelver RC, Moehle EA, Worden SE, Mitchell JC, Arnold NL, Gopalan S, Meng X, Choi VM, Rock JM, Wu YY, Katibah GE, Zhifang G, McCaskill D, Simpson MA, Blakeslee B, Greenwalt SA, Butler HJ, Hinkley SJ, Zhang L, Rebar EJ, Gregory PD, Urnov FD (2009) Precise genome modification in the crop species Zea mays using zinc-finger nucleases. Nature 459:437–441
Slade AJ, Knauf VC (2005) TILLING moves beyond functional genomics into crop improvement. Transgenic Res 14:109–115
Slade AJ, Fuerstenberg SI, Loeffler D, Steine MN, Facciotti D (2005) A reverse genetic, nontransgenic approach to wheat crop improvement by TILLING. Nat Biotechnol 23:75–81
Spaenij-Dekking L, Kooy-Winkelaar Y, Koning F (2005) The Ethiopian cereal tef in celiac disease. N Engl J Med 353:1748–1749
Stadler F, Mohler V, Kilian A, Bonin A, Thümmler F, Mayes S, Ros B, Wenzel G (2009) Initial results from applying diversity arrays technology (DArT) and massively parallel signature sequencing (MPSS) to bambara groundnut. In: Tadele Z (ed) New approaches to plant breeding of orphan crops in Africa. In: Proceedings of an International Conference, Bern, Switzerland. Stämpli AG, Bern, 19–21 Sept 2007
Stephenson P, Baker D, Girin T, Perez A, Amoah S, King GJ, Østergaard L (2010) A rich TILLING resource for studying gene function in Brassica rapa. BMC Plant Biol 10:62
Suzuki T, Eiguchi M, Kumamaru T, Satoh H, Matsusaka H, Moriguchi K, Nagato Y, Kurata N (2008) MNU-induced mutant pools and high performance TILLING enable finding of any gene mutation in rice. Mol Genet Genomics 279:213–223
Tadele Z (ed) (2009) New approaches to plant breeding of orphan crops in Africa: Proceedings of an International Conference, Bern, Switzerland. Stämpli AG, Bern, 19–21 Sept 2007
Tadele Z, Mba C, Till BJ (2009) TILLING for mutations in model plants and crops. In Mohan Jain S, Brar DS (eds) Molecular techniques in crop improvement. Springer, Netherlands
Talamè V, Bovina R, Sanguineti MC, Tuberosa R, Lundqvist U, Salvi S (2008) TILLMore, a resource for the discovery of chemically induced mutants in barley. Plant Biotechnol J 6:477–485
Taylor NE, Greene EA (2003) PARSESNP: A tool for the analysis of nucleotide polymorphisms. Nucleic Acids Res 31:3808–3811
Tester M, Langridge P (2010) Breeding technologies to increase crop production in a changing world. Science 327:818–822
Thorpe TA (2007) History of plant tissue culture. Mol Biotechnol 37:169–180
Till BJ, Reynolds SH, Greene EA, Codomo CA, Enns LC, Johnson JE, Burtner C, Odden AR, Young K, Taylor NE, Henikoff JG, Comai L, Henikoff S (2003) Large-scale discovery of induced point mutations with high-throughput TILLING. Genome Res 12:524–530
Till BJ, Reynolds SH, Weil C, Springer N, Burtner C, Young K, Bowers E, Codomo CA, Enns LC, Odden AR, Greene EA, Comai L, Henikoff S (2004a) Discovery of induced point mutations in maize genes by TILLING. BMC Plant Biol 4:12
Till BJ, Burtner C, Comai L, Henikoff S (2004b) Mismatch cleavage by single-strand specific nucleases. Nucleic Acids Res 32:2632–2641
Till BJ, Zerr T, Comai L, Henikoff S (2006) A protocol for TILLING and Ecotilling in plants and animals. Nat Protoc 1:2465–2477
Till BJ, Cooper J, Tai TH, Colowit P, Greene EA, Henikoff S, Comai L (2007a) Discovery of chemically induced mutations in rice by TILLING. BMC Plant Biol 7:19
Till BJ, Comai L, Henikoff S (2007b) TILLING and EcoTILLING for crop improvement. In Varshney RK, Tuberosa R (eds) Genomics-Assisted Crop Improvement, vol 1: genomics approaches and platforms. Springer, Netherlands
Till BJ, Jankowicz-Cieslak J, Sági L, Huynh OA, Utsushi H, Swennen R, Terauchi R, Mba C (2010) Discovery of nucleotide polymorphisms in the Musa gene pool by Ecotilling. Theor Appl Genet 121:1381–1389
Townsend JA, Wright DA, Winfrey RJ, Fu F, Maeder ML, Joung JK, Voytas DF (2009) High-frequency modification of plant genes using engineered zinc-finger nucleases. Nature 459:442–425
Triques K, Sturbois B, Gallais S, Dalmais M, Chauvin S, Clepet C, Aubourg S, Rameau C, Caboche M, Bendahmane A (2007) Characterization of Arabidopsis thaliana mismatch specific endonucleases: application to mutation discovery by TILLING in pea. Plant J 51:1116–1125
Tsai H, Howell T, Nitcher R, Missirian V, Watson B, Ngo KJ, Lieberman M, Fass J, Uauy C, Tran RK, Khan AA, Filkov V, Tai TH, Dubcovsky J, Comai L (2011) Discovery of rare mutations in populations: TILLING by sequencing. Plant Physiol 156:1257–1268
Uauy C, Paraiso F, Colasuonno P, Tran RK, Tsai H, Berardi S, Comai L, Dubcovsky J (2009) A modified TILLING approach to detect induced mutations in tetraploid and hexaploid wheat. BMC Plant Biol 9:115
Vasil IK (2005) Tissue cultures of maize. Maydica 50:361–365
Wahid MB, Abdullah SNA, Henson IE (2005) Oil palm—Achievements and potential. Plant Prod Sci 8:288–297
Wang D-K, Sun Z-X, Tao Y-Z (2006) Application of TILLING in plant improvement. Acta Genetica Sinica 33:957–964
Wang N, Wang Y, Tian F, King GJ, Zhang C, Long Y, Shi L, Meng J (2008) A functional genomics resource for Brassica napus: development of an EMS mutagenized population and discovery of FAE1 point mutations by TILLING. New Phytol 180:751–765
Wang T, Uauy C, Till B, Liu C-M (2010) Tilling and associated technologies. J Integr Plant Biol 52:1027–1030
Wang Y, Li J (2006) Genes controlling plant architecture. Curr Opin Biotech 17:123–129
Weil CF (2009) TILLING in grass species. Plant Physiol 149:158–164
Weil CF, Monde R-A (2007) Getting the point—mutations in maize. Crop Sci 47:S60–S67
Xin Z, Wang ML, Barkley NA, Burow G, Franks C, Pederson G, Burke J (2008) Applying genotyping (TILLING) and phenotyping analyses to elucidate gene function in a chemically induced sorghum mutant population. BMC Plant Biol 8:103–116
Yan ZY, Spencer PS, Li ZX, Liang YM, Wang YF, Wang CY, Li FM (2006) Lathyrus sativus (grass pea) and its neurotoxin ODAP. Phytochemistry 67:107–121
Acknowledgments
K. E. and Z. T. express their gratitude to Syngenta Foundation for Sustainable Agriculture and University of Bern for supporting the Tef Improvement Project. The TILLING research in the Uauy lab is funded by the UK Biotechnology and Biological Sciences Research Council (BBSRC) (BB/I000712/1).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Esfeld, K., Uauy, C., Tadele, Z. (2013). Application of TILLING for Orphan Crop Improvement. In: Jain, S., Dutta Gupta, S. (eds) Biotechnology of Neglected and Underutilized Crops. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-5500-0_6
Download citation
DOI: https://doi.org/10.1007/978-94-007-5500-0_6
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-5499-7
Online ISBN: 978-94-007-5500-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)