
Abstract
The first cryptodiran turtle with a pitted palate, Brachyopsemys tingitana gen. et sp. nov., is described on the basis of skulls and a lower jaw from the early Paleocene (Danian) of the Ouled Abdoun Basin, Morocco. The new taxon shares a number of apomorphic features with Sandownia harrisi from the Aptian of the Isle of White, England, Angolachelys mbaxi from the Turonian of Angola, and an unnamed turtle from the Albian Glen Rose Formation in Texas. A phylogenetic analysis provides evidence that they form a monophyletic clade which we name the Sandownidae fam. nov. The phylogenetic relationships of Sandownidae are problematic. Their affinities among the Eucryptodira remain uncertain; a well-preserved shell of a member of the family may be useful in determining relationships. The family appears to be a bottom-dwelling group that occurred in near-shore marine environments along the coast of the Atlantic across the Cretaceous-Tertiary boundary. Its evolutionary history is connected to the development of the Atlantic Ocean.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
Among recently described or discovered Cretaceous and early Tertiary turtles are four cryptodire skulls that all exhibit a remarkable suite of characters including an extensive skull roof, an extensive secondary palate with a large contribution by the jugal, and a reduced or absent ventral exposure of the basisphenoid. This paper is an attempt to determine if these highly functional characters (broad palate and complete skull roof) result from shared common ancestry or if they might be the result of convergent evolution. The four taxa that show these characters are Sandownia harrisi from the Aptian of the Isle of Wight, England (Meylan et al. 2000), Angolachelys mbaxi from the Turonian of Angola (Mateus et al. 2009), an unnamed turtle (“the Glen Rose turtle”) from the Albian Glen Rose Formation of Texas (Barck 1992; Vineyard and Jacobs 2009), and the turtle described as a new genus and species in this paper.
Sandownia harrisi is a remarkably well preserved skull and partial lower jaw that has been described in detail (Meylan et al. 2000). Its phylogenetic position was proposed on the basis of a set of analyses that were relatively limited in scope and it was considered to be a member of the Trionychoidea.
Angolachelys mbaxi is known from a single, poorly preserved skull that is badly broken dorsally. The type description includes a brief treatment of the morphology of the skull, a dentary, two procoelous cervicals, and an ungual phalanx (Mateus et al. 2009). There is no data set in the type description (a data set was made available to us by the authors). A single tree selected by the authors from among 60 MPT was used to argue for recognition of the “Angolachelonia” including A. mbexi, S. harrisi, Solnhofia parsonsi, and the Glen Rose turtle.
The Glen Rose turtle is yet to be named and fully described but it has been presented at several conferences (Vineyard 1999; Vineyard and Jacobs 2009) and is highly relevant to the problem at hand. Derived characters were listed by Vineyard and Jacobs (2009) and several of these are shared with the other three taxa of interest here.
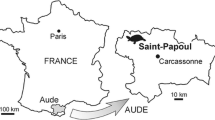
The fourth taxon is named below. It is known from two complete skulls, and a third skull that is missing the skull roof but has associated lower jaws. These specimens were collected by local people in the Ouled Abdoun Phosphate Basin, Morocco. During the last decade, abundant fossil turtles have been discovered in the Maastrichtian (Late Cretaceous) to Ypresian (Early Eocene) phosphate deposits of the Ouled Abdoun Basin. They include pleurodiran bothremydids and cryptodiran chelonioid sea turtles (Karl et al. 1998; Tong and Hirayama 2002, 2004, 2008; Gaffney and Tong 2003; Hirayama and Tong 2003; Gaffney et al. 2006; Nour-Eddine et al. 2009; Tong et al. in press). According to the local collectors, one of the skulls described here, American Musuem of Natural History (AMNH) 30001, was collected in the ‘dalle couche 2’ (Slab of layer 2) of the ‘Recette 4’ (Quarry 4), in the Ouled Abdoun Basin. Shark teeth found in the matrix surrounding the specimens during preparation included Palaeogaleus brivesi, Lamniformes indet., Ixobatis mucronata, Plicatosyllium sp., Dasiatis hexagonalis, Danogaleus gueriri, Rhinobatoid indet., Youssoubatis ganntourensis, Ctenopristis nougareti and Squalus sp. This composition of Danian selachians mixed with some Maastrichtian elements indicates a Danian age (Cappetta, pers. com. 2001). These turtles were mentioned as a ‘pitted cryptodire’ in Gaffney et al. (2006). Two of three skulls and the lower jaw have been fully prepared and are described herein. The specimens are housed in the American Museum of Natural History (AMNH). It is certain that this material represents an undescribed form and the occasion of its description provides an opportunity to reconsider this bizarre group of turtles. Thus, this paper has three purposes: (1) to describe a remarkable new cryptodire from the Paleocene of Morocco; (2) to examine the question of monophyly of a set of possibly related Cretaceous and early Tertiary cryptodires with extensive secondary palates and extensive skull roofs; and (3) to determine the best hypothesis of relationship among these turtles and other cryptodires.
Systematic Palaeontology
-
Order Testudines Linnaeus 1758
-
Suborder Cryptodira Cope 1868
-
Parvorder Eucryptodira Gaffney 1975
-
Superfamily Incertae Sedis
-
Family Sandownidae nov.
Type genus: Sandownia Meylan, Moody, Walker and Chapman 2000
Included genera: Sandownia Meylan, Moody, Walker and Chapman 2000; Angolachelys Mateus, Jacobs, Polcyn, Schulp, Vineyard, Neto and Antunes 2009; Brachyopsemys gen. nov.
Diagnosis: Centrocryptodiran turtles (otic trochlea present and internal carotid artery located deep within pterygoid) with the jugal making a significant contribution to the triturating surface; pterygoids partly or completely cover basisphenoid; premaxillae partly or completely fused and slightly protuberant anteriorly; prefrontal L-shaped, with a posteriolateral process along the ventral margin of the orbit; external process of the pterygoid absent and occipital condyle with anterior location in between the articular surfaces of the quadrate. Long symphysis of dentary; broad triangular lower triturating surface that slopes ventrally from the coronoid process and lacks labial and lingual ridges (known in Sandownia harrisi and the taxon described below).
Remarks: Mateus et al. (2009) named the unranked taxon “Angolachelonia” and defined it as the clade originating from the most recent common ancestor of Angolachelys mbaxi and Solnhofia parsonsi. Our phylogenetic studies (see below) suggest that, as defined, Angolachelonia is a junior synonym of Parvorder Eucryptodira (sensu Gaffney and Meylan 1988). We have found strong support for monophyly of the clade consisting of Angolachelys mbexi, Sandownia harrisi and the taxon described below and chose to recognize this relationship through the use of the family group name, Sandownidae, with the well-preserved type genus Sandownia. Given that the name “Angolachelonia” is non-Linnaean, it cannot be construed as a family-group name and as defined may be a synonym of an ordinal group name. We find it more useful to construct this new family-group name based on character diagnoses rather than on a name based on a phylogeny that was selected from among 60 MPT (Mateus et al. 2009). It is clear that the Glen Rose turtle is a member of the family Sandownidae.
-
Brachyopsemys gen. nov.
Type species: Brachyopsemys tingitana sp. nov.
Etymology: Brachyops: Greek, short face; emys: Greek, turtle.
Diagnosis: Skull broad and low, wider than long, rounded anteriorly in dorsal view, with a very extensive skull roof (more extensive than other sandownids), very shallow temporal emargination with parietal-squamosal contact; orbits facing anterolaterally, placed forward and closely spaced; large triturating surface with a pair of deep pits in the jugal (unique among cryptodires, seen also in the pleurodire family Bothremydidae); unique among turtles in having prefrontal-jugal contact present below orbit excluding maxilla from orbit margin and a medial process of the jugal that forms a large, flat, horizontal, plate-like flange within the fossa temporalis inferior; unique among described members of the family Sandownidae in having the incisura columellae auris open.

Brachyopsemys tingitana gen. et sp. nov., from the Danian (early Palaeocene) of Morocco. AMNH 30001, holotype skull, photographs in: a dorsal, b ventral, c anterior, d left lateral, e posterior, and f right lateral views. Scale bar = 50 mm

Brachyopsemys tingitana gen. et sp. nov., from the Danian (early Palaeocene) of Morocco. AMNH 30001, holotype skull, interpretive drawings in: a dorsal, b ventral, c anterior, d left lateral, e posterior, and f right lateral views. Scale bar = 50 mm. Abbreviations: bo basioccipital; ex exoccipital; FRPA frontoparietal scale; fr frontal; FR frontal scale; ju jugal; mx maxilla; MX maxillary scale; op opisthotic; pa parietal; PA parietal scale; pal palatine; pf prefrontal; pm premaxilla; po postorbital; pr prootic; pt pterygoid; qj quadratojugal; qu quadrate; so supraoccipital; sq squamosal; TE temporal scale; vo vomer

Brachyopsemys tingitana gen. et sp. nov., from the Danian (early Palaeocene) of Morocco. AMNH 30554, referred skull, photographs (left) and interpretive drawings (right) in: a–b dorsal, and c–d ventral views. Abbreviations see Fig. 13.2

Brachyopsemys tingitana gen. et sp. nov., from the Danian (early Palaeocene) of Morocco. AMNH 30558, lower jaw, photographs (left) and interpretive drawings (right) in: a–b dorsal, c–d ventral, e–f right lateral, and g–h lingual views. Scale bar = 50 mm. Abbreviations: ang angular; art articular; cor coronoid; den dentary; fdm foramen dentofaciale majus; pra prearticular; spl splenial; sur surangular
Holotype: A complete skull (AMNH 30001).
Holotype locality: Recette 4, Ouled Abdoun Basin, Morocco.
Holotype unit and age: Dalle Couche 2, early Paleocene (Danian).
Etymology: From Moretania Tingitana, an ancient Roman province corresponding to northern Morocco.
Referred specimens: An incomplete skull, lacking the skull roof, with associated lower jaw (AMNH 30558); a complete skull (AMNH 30554).
Occurrence: Early Paleocene (Danian) of Morocco.
Diagnosis: Same as for genus.
Measurements: see Table 13.1.
Description
Preservation: AMNH 30001 is a well preserved complete skull without deformation. Some areas within the fossa temporalis remain filled with matrix. AMNH 30558 is a nearly complete skull, lacking only the skull roof, with an associated nearly complete lower jaw. This specimen has been prepared to reveal much of the braincase, orbit floor and otic region. AMNH 30554 is a complete skull that remains to be prepared.
General aspect: The skull is low and broad, with a rough external surface and scute sulci on the skull roof but without obvious ornamentation. It has a nearly rounded shape as seen from above but the lateral margins are parallel. The skull roof is complete, with very shallow temporal emargination, which is even shallower than that of Sandownia harrisi (Meylan et al. 2000), but comparable to that of Erquelinnesia gosseleti (Zangerl 1971). The crista supraoccipitalis does not extend beyond the posterior margin of the skull roof so is not visible dorsally, but it does extend significantly beyond the condylus occipitalis. The apertura narium externa is oval-shaped and wider than high; facing forward. The orbits are oval and quite small when compared to those of extant chelonioids. They are placed very anteriorly, facing mainly forward and are very close to each other. The interorbital bar is very narrow as in S. harrisi. In lateral view, the skull roof slopes gently from the interorbital bar posteriorly. The cheek region, below the orbit, is convex laterally to form a rounded swelling, giving the skull a ‘chubby-cheeked’ aspect. The cheek emargination is very shallow, shallower than in Sandownia, and is much lower than the lower rim of the orbit. In ventral view, the triturating surface is very large and wide, with a pair of deep pits located laterally. The condylus occipitalis lies anteriorly, in line between the condylus mandibularis of the two quadrates.
Skull roof scales: Some weak sulci are visible in AMNH 30001. On the midline, there is a pair of frontal scales between the orbits, followed by a pair of frontoparietals and a single parietal scale. The supraorbital scale lies posterior to the orbit and lateral to the frontoparietal scale. Posterior to it and lateral to the parietal scale is a temporal scale. One suborbital scale can be distinguished on each side under the orbit. Below it lies the maxillary scale (Figs. 13.1, 13.2). Skull scale terminology follows Tong and Hirayama (2002).
Prefrontal: Both prefrontals are complete in AMNH 30001. The prefrontal is L-shaped with the vertical part contributing to the narrow interorbital bar and a horizontal part that forms a significant portion of the anterior and ventral orbit margin. This condition is as in Sandownia harrisi, in contrast to cheloniids, trionychids and most other turtles in which the prefrontal has no laterally directed horizontal branch. The vertical ramus contacts the frontal dorsally between the orbits; ventrally, it forms the upper rim of the apertura narium externa. The surface between the orbits and the apertura narium externa is flat and nearly vertical. The horizontal process extends from the vertical portion laterally to form the anteroventral margin of the orbit and the anterior margin of the foramen orbito-nasale. It is divided into two branches by the foramen orbito-nasale: a smaller medial branch turns posteriorly to contact the vomer medially and the palatine posteriorly, and a larger lateral branch that forms the ventral orbit margin and contacts the jugal laterally, palatine posteriorly, and maxilla anteroventrally on the skull surface. In addition, there is a very short prefrontal-maxilla contact posterolaterally on the anterior edge of the orbital floor between the palatine and jugal. The prefrontal of S. harrisi differs in the longer dorsal branch, the smaller contribution to the external nares and the absence of prefrontal-jugal contact. The foramen orbito-nasale in Brachyopsemys is a large oval-shaped opening (6.5 × 4 mm) lying on the anteromedial corner of the orbit floor as in Chelonia mydas. In trionychids such as Amyda cartilaginea and Lissemys punctata, this foramen is much larger and more posteriorly placed.
Nasal: The nasal is clearly absent.
Frontal: Both frontals are complete in AMNH 30001. It is a rather small and trapezoidal-shaped bone forming the dorsal rim of the orbit. On the skull roof, the frontal contacts the prefrontal anteriorly, the parietal posteriorly with a long and transversal suture, and the postorbital posteriolaterally by a short suture. The ventral surface of the frontal bears a thick, blunt parasagittal ridge which separates the fossa orbitalis laterally from the sulcus olfactorius medially. The sulcus olfactorius is narrow anteriorly and widens posteriorly. The morphology of the frontal in AMNH 30001 is similar to Sandownia harrisi. Trionychids also have a very small frontal that contributes to the dorsal rim of the orbit, while in cheloniids the frontal is larger.
Parietal: The parietals are complete in AMNH 30001 but are not fully prepared ventrally. Only small ventral portions of the processus inferior parietalis are preserved in AMNH 30558. The parietal is a large and anteroposteriorly elongated bone as seen from above, producing a large part of the skull roof. On the skull roof, it forms the medial portion of the very shallow temporal emargination and contacts the frontal anteriorly, the postorbital anterolaterally, the squamosal posterolaterally and the supraoccipital posteromedially. Contact with the squamosal is extensive. The very complete skull roof with parietal-squamosal contact is similar to Sandownia harrisi, but the temporal emargination in the latter is deeper. A parietal-squamosal contact is also present in cheloniids and other turtles with an extensive skull roof (see Discussion).
The anterior part of processus inferior parietalis in AMNH 30001 is free of matrix; it is low and contacts the palatine ventrally. The small pieces of the processus inferior parietalis preserved in AMNH 30558 show the contact with the palatine anteroventrally by a rather long suture, the pterygoid ventrally, and the prootic posteroventrally. Ventrally, between the pterygoid and the prootic, the parietal forms the upper margin of the foramen nervi trigemini.
Postorbital: Both postorbitals are slightly damaged on the skull roof of AMNH 30001, but the outline can be easily made out. Only a very small portion of the postorbital is preserved on the left side of AMNH 30558. The postorbital is a large anteroposteriorly elongate sheet of bone forming the posterior margin of the orbit. The postorbital has typical contacts with the frontal anteromedially, the parietal posteromedially, the jugal anterolaterally, the quadratojugal laterally and the squamosal posteriorly. In addition, there is a short postorbital-quadrate contact between the quadratojugal and the squamosal only on the left side of AMNH 30001. This is considered as an anomaly, since there is usually a quadratojugal-squamosal contact preventing postorbital-quadrate contact among turtles. The fossa orbitalis is completely open posteriorly.
Jugal: The right jugal is complete in AMNH 30001, while the left one is slightly damaged on its surface. In AMNH 30558, both jugals are nearly complete, with the dorsal part damaged on both sides.
The jugal is unusually large and strongly convex dorsolaterally; it makes a large contribution to the lateral skull surface as in Sandownia harrisi (Meylan et al. 2000). But where the jugal and the maxilla have an equal contribution to the lateral skull surface in S. harrisi, the jugal of Brachyopsemys is much larger than the maxilla due to its anterior enlargement. The jugal has the usual contacts with the postorbital posterodorsally, the quadratojugal posteriorly, and the maxilla anteroventrally. A remarkable contact occurs medially, along the lower orbit margin, where the jugal meets the lateral process of the prefrontal excluding the maxilla from the orbit. This condition is unique among turtles. The jugal makes a small contribution to the anterolateral part of the orbit floor, where it contacts the maxilla medially and the pterygoid posteriorly. Laterally in the fossa temporalis inferior, there is a small foramen on the lateral wall of the triturating surface lateral to the palatal pit. This foramen is visible on both sides in AMNH 30558.
Ventrolaterally the jugal makes a large contribution to the lower cheek margin where it turns medially to form a large process, which is a flat, horizontal plate-like flange within the fossa temporalis inferior. Posterior to the flange, there is a well-developed lateral knob on the lower margin of the jugal above the anterior part of the very shallow cheek emargination. This knob is well preserved on both sides of AMNH 30558. In AMNH 30001, the knob is clearly visible on the right side, but less well-developed than in AMNH 30558; while it is damaged on the left side. A similar structure appears to be present in the Glen Rose turtle (Barck 1992). This structure may be unique among turtles, however, a similar knob is developed in some species of Palatobaena (Lyson and Joyce 2009). This structure may be associated with an M. zygomaticomandibularis, a muscle that has only been identified in trionychids (Schumacher 1973). This muscle extends from the medial surface of the jugal, quadratojugal and quadrate to the lateral surface of the dentary and surangular. The lower jaw of Brachyopsemys has a very large area for muscle insertion ventral to and posterior to the coronoid process that seems a very likely candidate for insertion of this muscle. The medial process of the jugal decribed above would provide additional area for origin of a large M. zygomaticomandibularis. Small knobs in a smilar location on the jugal of Apalone ferox have been illustrated by Dalrymple (1977, Fig. 5).
The ventral surface of the jugal forms the posterolateral portion of the triturating surface lateral to the maxilla and pterygoid. Within the ventral surface of the jugal is a deep palatal pit. This conical and dorsolaterally directed pit is deep; it makes the jugal convex dorsolaterally on the skull roof surface to give the skull a ‘chubby-cheeked’ aspect. Laterally the pit is bordered by a vertical, thin ridge with a sharp margin. On the ventral surface, the jugal contacts the maxilla anteromedially and the pterygoid posteromedially. A jugal-palatine contact is absent as in S. harrisi and trionychids, in contrast to most cheloniids in which this contact is present. The morphology and contacts of the jugal, maxilla, palatine and pterygoid on the triturating surface in Brachyopsemys are closely comparable to S. harrisi, however, the palatal pit is absent in the latter. Similar palatal pits are known in some bothremydids (Gaffney et al. 2006), but have never been reported in the Cryptodira.
Quadratojugal: Both quadratojugals are complete in AMNH 30001. In AMNH 30558, the left quadratojugal is nearly complete, but the right one is damaged.
The quadratojugal is a vertical, flat bone covering the lateral side of the skull between the jugal and quadrate. It is large as in Sandownia harrisi and cheloniids, in contrast to the small quadratojugal of trionychids. Ventrally, it forms the posterior portion of the very shallow cheek emargination. It contacts the jugal anteriorly, the postorbital dorsally and the quadrate posteriorly. It contacts the squamosal on the right side of AMNH 30001, but this contact is absent on the left side of AMNH 30001.
Squamosal: Both squamosals are complete in AMNH 30001. In AMNH 30558, a small portion of squamosal is preserved on the left side, the right one is missing.
The squamosal is a roughly rectangular flat bone, longer than wide, covering the posterolateral corner of the skull roof and contributing to the lateral portion of the temporal margin. It contacts the postorbital anteriorly, the parietal medially by a long suture and the quadrate ventrally. In addition, the squamosal contacts the quadratojugal on the right side of AMNH 30001. The squamosal forms the roof of the antrum postoticum, but the sutures on the ventral surface, posterior to the cavum tympani are somewhat confusing, the antrum postoticum seems to be formed ventrally by the quadrate. If this interpretation is correct, there is no squamosal-opisthotic contact. The large size and the contact of the squamosal on the skull roof are comparable to Sandownia and cheloniids, and other turtles with an extensive skull roof; but different from the cone-shaped squamosal of trionychids.
Premaxilla: The premaxilla is complete in AMNH 30001. AMNH 30558 has nearly complete premaxillae, with the dorsal surface only slightly damaged.
In both AMNH 30001 and AMNH 30558, the right and left premaxillae are completely fused as in Trionychia (Meylan and Gaffney 1989) and Erquelinnesia (Zangerl 1971); there is no midline suture visible on dorsal nor on ventral surface. The premaxillae are partially fused in Sandownia (Meylan et al. 2000). Dorsally the premaxilla forms the lower rim of the aperture narium externa and contributes to the floor of the fossa nasalis where it contacts the maxilla posterolaterally. The dorsal exposure of the premaxilla is wide anteriorly and sends a median posterior process between the maxillae. The dorsal surface of the premaxilla bears a low and blunt parasagittal ridge. The ridges of the right and left premaxillae together produce a midline grove. This structure is clearly visible in AMNH 30001, in the prepared part of the fossa nasalis, but its extent cannot be determined at the present state of preparation. This surface is damaged in AMNH 30558.
In ventral view, the premaxilla contributes to the anteromedial portion of the triturating surface and its labial ridge. It contacts the maxilla posterolaterally. There is no premaxilla-vomer contact on either the dorsal or ventral surface. A premaxilla-vomer contact is present in most cryptodiran turtles including Sandownia harrisi, but trionychids usually have no vomer-premaxilla contact due to the reduction of the ventral portion of the vomer and/or the development of the foramen intermaxillaris. Cheloniids may not have a vomer-premaxilla contact in ventral view due to medial expansion of the palatine process of the maxillae, but it may be retained dorsally in the floor of the fossa nasalis (Gaffney 1979, p. 87). Thus the condition in our specimens is more similar to that of trionychids.
Maxilla: Both maxillae are complete in AMNH 30001. In AMNH 30558, they are nearly complete, but the dorsal process is broken off on both sides.
Dorsally, the maxilla makes up the lateral margin of the apertura narium externa and most of the fossa nasalis floor, but it does not reach the orbital margin in contrast to all other turtles. The maxilla has a reduced exposure on the skull surface, where it contacts the premaxilla medially, the prefrontal dorsomedially and the jugal posterolaterally. The lateral and ventral surfaces of the maxilla and the premaxilla bear numerous nutritive foramina, suggesting that they were covered by a large, thick rhamphotheca. However, the nutritive foramina are larger, but sparser on the triturating surface than on the lateral surface, which suggests that the rhamphotheca was thicker on the feeding surface than on the lateral surface. The extension of this epidermal jaw covering on the skull surface is marked by a wide and shallow sulcus on the lateral surface of the maxilla, along the maxilla-jugal suture. Such a sulcus is also present in Sandownia harrisi (Meylan et al. 2000).
In the fossa nasalis, the maxilla contacts the premaxilla anteromedially, the vomer posteromedially and the palatine posterolaterally. The maxillae share a common midline contact between the premaxilla and vomer on the fossa nasalis floor. Although the maxilla does not contribute to the orbit margin, it is exposed on the orbit floor as in other turtles, but posterior to the jugal-prefrontal contact. The exposure is triangular, narrowed posteriorly. There are two foramina lying on its anterior end: the large lateral one (about 2 mm in diameter) is the foramen supramaxillare, whereas the very small medial one is interpreted here as the foramen supraorbitale. The maxilla does not reach the foramen orbito-nasale, in contrast to Chelonia mydas. Maxillary contacts on the orbit floor include the jugal anterolaterally and laterally, the prefrontal anteromedially, the palatine medially and the pterygoid posteriorly.
In ventral view, the maxilla, premaxilla, vomer, palatine, jugal and pterygoid, all contribute to the very large, concave triturating surface. This surface is bordered by a very thick labial ridge. The labial ridge made by the premaxilla in the middle is very low; it increases in height posterolaterally in the maxilla. The triturating surface has no medial ridge and no obvious posterior limit. Laterally, the maxilla slopes into a very deep pit on the triturating surface. The pit lies near the lateral border of the triturating surface and is formed mainly by the jugal. The maxilla contributes to its medial portion. On the triturating surface, the maxilla contacts the premaxilla anteromedially, the vomer posteromedially, the palatine and the pterygoid posteriorly, and the jugal laterally. In addition, the maxillae share a long common midline contact between the premaxilla and the vomer. It has a very small contribution to the apertura narium interna margin between the vomer and the palatine. A maxillary midline contact is commonly seen in trionychids because of the reduction of the vomer. It may also occur in some living cheloniids (Caretta caretta) or as an individual variation (Gaffney 1979). This contact is absent in Sandownia harrisi.
Vomer: The vomer is complete in AMNH 30001. In AMNH 30558, the dorsal surface of the vomer is damaged; the remaining part is totally free of matrix.
The vomer is a small hourglass-shaped element. Ventrally, it has a very small exposure on the triturating surface behind the long maxilla midline suture. It forms the anteromedial margin of the apertura narium interna. It contacts the maxilla anteriorly and laterally on the triturating surface and on the floor of the fossa nasalis, but apparently a vomer-palatine contact is prevented by the maxilla. A reduced vomer with a long midline suture between the maxillae is found in trionychids and carettochelyids, whereas ventral exposure of the vomer is large in cheloniids, and also in Sandownia harrisi. Dorsally, the vomer sends a sagital process (vomerine pillar) to meet the dorsal portion of the vomer between the palatines. The vomerine pillar is short, its posterior end being anterior to the anterior edge of the apertura narium interna. This structure is visible in AMNH 30558, as well as in AMNH 30001 by CT scan.
Dorsally, on the orbit floor, the vomer contacts the prefrontal anterolaterally and the palatine laterally. Posteriorly, there is a short vomer-pterygoid contact preventing palatine midline contact.
Palatine: Both palatines are complete in both AMNH 30001 and AMNH 30558. In AMNH 30001, the ventral surface is fully visible; but the posterior end of the dorsal side is still covered by matrix. The palatine is free of matrix on both dorsal and ventral sides in AMNH 30558.
The palatine is a C-shaped bone in cross section. The ventral exposure of the palatine, on the triturating surface is roughly diamond-shaped, lying posteromedial to the maxilla. It contributes to the posteromedial portion of the triturating surface and most of the apertura narium interna margin. The apertura narium interna is located at about the middle of the palatal surface of the skull, the position being comparable to that of Sandownia harrisi. On the palatal surface, the palatine contacts the maxilla anteriorly and the pterygoid posteriorly. There is no palatine-jugal contact on the palatal surface as in S. harrisi and trionychids. The palatines lie very closely together on the triturating surface; however, the vomer and maxilla prevent a palatine midline contact. On the floor of the fossa nasalis, the palatines are separated from one another by the vomer anteriorly and by the pterygoid posteriorly.
The dorsal shelf of the palatine contributes to the medial portion of the orbit floor, where it contacts the prefrontal anteriorly, both medial and lateral to the foramen orbito-nasale. It meets the vomer medially, the maxilla anterolaterally, and the pterygoid posteriorly. It seems that there is no palatine midline contact on the dorsal surface. Anteriorly, the palatine makes up the posterior margin of the foramen orbito-nasale; and posteriorly, the medial rim of the foramen palatinum posterius. The foramen palatinum posterius is a large and anteroposteriorly elongate opening lying on the posterior end of the palatine, on the palatine-pterygoid suture as in Sandownia harrisi. This foramen is usually located on the palatine-maxilla or palatine-pterygoid suture in trionychids, but lateral to the palatine; it is absent in cheloniids.
The dorsal process of the palatine appears to be a thin sheet that covers the anterior part of the lateral wall of the braincase. However, the pterygoid contacts the vomer dorsally and ventrally and appears to make up most of the thickness of the side wall in AMNH 30558. The palatine appears only to contribute to the thickness of the anterior part of the side wall. A pterygoid-vomer contact excludes palatine contact of the descending process of the parietal.
Epipterygoid: In AMNH 30558, there is a fragment of bone on the left side wall of the braincase between the foramen palatinum posterius and the foramen nervi trigemini that is located in the approximate position of the epipterygoid. However, this could be a fragment of the descending process of the parietal. Thus, at this time it is not possible to say if the epipterygoid was present or absent.
Pterygoid: Both pterygoids are complete in AMNH 30001, but only the ventral side is free of matrix. They are very slightly damaged in AMNH 30558 and prepared on both ventral and dorsal sides.
In ventral view, the pterygoid is anteroposteriorly elongate, forming the posterior part of the secondary palate between the jugal and palatine where it has a short contact with the maxilla anteriorly, as in Sandownia harrisi. Medially the pterygoid forms the roof of the apertura narium interna where they share a long, common, midline suture between the vomer and the basioccipital. The pterygoid completely covers the basisphenoid ventrally, so the latter is not visible in palatal view. Anteriorly, in the apertura narium interna, the pterygoid contacts the vomer anteromedially and the palatine anterolaterally in AMNH 30558. These contacts are covered by matrix in AMNH 30001. The processus pterygoideus externus seen in most cryptodires is absent. The lateral margin of the pterygoid is a high and thin vertical ridge extending from the jugal anteriorly to the quadrate posteriorly. Just anterior to the processus articularis of the quadrate along this ridge is a small protuberance. This structure is well preserved in AMNH 30001, but slightly damaged in AMNH 30558. Medial to this protuberance is a deep anteromedially-posterolaterally directed groove, probably for pterygoideus musculature. It is bordered laterally by a ridge that forms the lateral margin of the pterygoid, which is continuous with the processus articularis of the quadrate. Posteromedially the groove is bordered by a low and thin ridge formed by the quadrate and pterygoid. This groove extends posterolaterally to the posterior surface of the processus articularis. The pterygoid contacts the basioccipital posteromedially, the exoccipital posteriorly and the quadrate posterolaterally. A small foramen posterius canalis carotici interni lies under the posteromedial ridge of the pterygoid-quadrate groove, on the pterygoid-exoccipital suture. The foramen is well preserved and visible in AMNH 30558, but remains covered by matrix in AMNH 30001. It is formed mainly by the pterygoid; the exoccipital forms the dorsomedial margin. A foramen carotico-pharyngeale as seen in Sandownia harrisi is absent.
On the dorsal surface, the pterygoid contributes to the posteriolateral portion of the orbital floor where it contacts the jugal anterolaterally, the maxilla anteriorly, and the palatine anteromedially. The foramen palatinum posterius is visible dorsally in AMNH 30558, lying on the posteriomedial corner of the orbit floor, on the pterygoid-palatine suture. On both sides a clear, round depression is visible posteriolateral to the foramen palatinum posterius. A midline contact of the pterygoids is visible on the dorsal surface, between the vomer and the basisphenoid. Dorsally, the pterygoid contacts the parietal, and posterodorsally it contacts the prootic and quadrate. The crista pterygoidea is very thick and seems to form the entire lower portion of the sidewall of the braincase. The foramen cavernosum is a small slit-like opening. The sulcus cavernosus is narrow.
Quadrate: The quadrates are complete in AMNH 30001; they are damaged on both sides in AMNH 30558.
Laterally, the quadrate forms the C-shaped cavum tympani. The cavum is small and deep, with deep dorsal and anterior walls. The incisura columellae auris is widely open posteroventrally as in cheloniids such as Chelonia mydas, in contrast to the closed incisura of Sandownia harrisi and trionychids. A small tubercle is present on the lower margin of the incisura columellae auris, which is more developed in AMNH 30558 than in AMNH 30001. A rounded and shallow depression is present anterior to the incisura columellae auris. The antrum postoticum is preserved in both sides of AMNH 30001, but it is full of matrix; it is also partly preserved on the left side of AMNH 30558. It is moderate in size. In lateral view, in addition to the usual contacts with the quadratojugal anteriorly and the squamosal dorsally, the quadrate contacts the postorbital anterodorsally by a short suture on the left side of AMNH 30001.
Dorsally, the quadrate forms the lateral portion of the very large processus trochlearis oticum and contributes to the lateral margin of the foramen stapedio-temporale (see prootic). Ventrally, the quadrate forms the low processus articularis. The posteromedial surface of the processus contains a concavity that is continuous with the anteromedial-posterolaterally directed groove of the pterygoid. A small foramen chorda tympani inferius lies within the posterior end of this groove, posterodorsal to the condylus mandibularis. The condylus mandibularis is roughly triangular; being wider than long; it is divided by a parasagittal furrow into two facets: a larger lateral one and a smaller medial one. In AMNH 30558, both facets are slightly convex. In AMNH 30001, the lateral facet is convex while the medial facet is flat. The quadrate contacts the pterygoid anteromedially, the prootic anterodorsomedially, and the opisthotic posteromedially. The fenestra postotica is a relatively small and oval opening formed by the pterygoid ventromedially, the quadrate laterally, the exoccipital medially and the opisthotic dorsally. It is closed medially and separated from the foramen jugulare posterius.
Basisphenoid: The basisphenoid is not visible in AMNH 30001; it is nearly complete in AMNH 30558 and exposed only on the dorsal surface. In ventral view, in both specimens it is completely covered by the pterygoid and basioccipital, apparently due to the shortening of the posterior part of the skull and extreme anterior location of the occipital condyle. Sandownia also has reduced ventral exposure of the basisphenoid, but a small basisphenoid exposure is visible in ventral view, surrounded by the pterygoid. The basisphenoid completely covered by pterygoid ventrally is also present in a solemydid referred to Helochelydra nopcsai by Joyce et al. (2011).
In dorsal view, the basisphenoid is anteroposteriorly elongate, forming the floor of the cavum cranii. The trabeculae meet in the midline to form the rostrum basisphenoidale. The sella turcica is a rather deep and anteroposteriorly elongate depression which is open anteriorly between converging lateral margins. Some bone is missing at the anterior end of the rostrum. The foramen anterius canalis carotici internae lie at the posterior end of the sella turcica. They are small and more widely spaced than in cheloniids [C. mydas, Euclastes wielandi (AMNH 30022) from the Danian of Morocco phosphates, Hirayama and Tong 2003]. The rostrum basisphenoidale is slightly damaged at its anterior end; it is relatively long, although not as rod-like as seen in cheloniids. The general morphology of the rostrum and sella turcica is comparable to that of Platysternon megacephalum (Gaffney 1979, Fig. 63), but different from the short and widely separated trabeculae of Sandownia harrisi, Ferganemys verzilini (Nessov 1977), and trionychids. The rostrum is slightly raised from the pterygoids allowing the passage of a rather large canal between the basisphenoid and pterygoids. These canals converge on the midline from each side. The anterior opening of the canals, which lies under the rostrum, is interpreted as the foramen anterius canalis caroticus laterale. The anterior location of this foramen is similar to the condition in S. harrisi, and to some extent to Kallokibotion bajazidi (Gaffney and Meylan 1992); but different from Ferganemys, Amyda cartilagineus and Dermatemys mawii in which this foramen is located more posteriorly and lateral to the dorsum sellae (Gaffney 1979, Figs. 58, 59). The foramen anterius canalis caroticus laterale is larger than the foramen anterius canalis carotici interni, as in S. harrisi.
The dorsum sellae is high and does not conceal the sella turcica, similar to Moroccan Euclastes wielandi (AMNH 30022). The processus clinoideus is broken off on both sides, but the remaining base indicates their position at the dorsolateral edge of the dorsum sellae. The posterior foramen nervi abducentis lies posterior to the processus clinoideus; the anterior foramen nervi abducentis is visible on the lateral surface of the basisphenoid, under the processsus clinoideus. The basis tuberculi basalis is a small tubercule lying on the basisphenoid/basioccipital suture. The basisphenoid contacts the pterygoid ventrally, the prootic dorsolaterally and the basioccipital posteriorly.
Basioccipital: The basioccipital is complete in both specimens, but only in AMNH 30558 are both the dorsal and ventral surfaces prepared. The basioccipital is an arrow-shaped bone in ventral view. There are some differences between AMNH 30001 and AMNH 30558. In 30001, the ventral surface of the basioccipital is shorter and concave with a rounded depression in the middle. While in AMNH 30558, the basioccipital is longer anteroposteriorly; a slight transverse crest divides the basioccipital into an anterior triangular and flat part, and a slightly concave posterior part; there is no rounded depression on it. Posteriorly, the basioccipital forms the ventral portion of condylus occipitalis. The condylus is well ossified, differing from the cartilaginous condyle seen in the holotype of Sandownia harrisi. The tuberculum basioccipitale is absent. The condylus occipitalis lies in line with the condylus mandibularis of the quadrate, as in S. harrisi. On the ventral surface, the basioccipital contacts the pterygoid anteriorly by an inverse V-shaped suture and the exoccipital laterally.
The dorsal exposure of the basioccipital on the floor of the cavum cranii is a small triangle. It bears most of the basis tuberculi basalis which is an anteroposteriorly elongated tubercle on the basioccipital-basisphenoid suture. In dorsal view, the basioccipital contacts the basisphenoid anteriorly, the opisthotic anterolaterally and the exoccipital posterolaterally.
Exoccipital: Both exoccipitals are complete in AMNH 30001, but damaged in AMNH 30558. The exoccipital forms the dorsal portion of the condylus occipitalis, the lateral margin of the foramen magnum and extends laterally to the medial margin of the fenestra postotica. It contacts the pterygoid ventrolaterally, the basioccipital ventromedially, the opisthotic dorsolaterally and the supraoccipital dorsally. The foramen jugulare posterius is a small oval opening on the exoccipital/opisthotic suture but is mostly formed by exoccipital. It is completely enclosed and separated from the fenestra postotica by exoccipital and opisthotic as in Chelydra serpentina (Gaffney 1979, Fig. 95), but differs from the condition in Sandownia harrisi. Two foramina nervi hypoglossi of equal size, lie on the posterior surface of the exoccipital. They are also visible on the dorsal surface of exoccipital in the floor of the cavum cranii of AMNH 30558.
Supraoccipital: The supraoccipital is complete but not fully prepared in AMNH 30001; the anteromedial part is still covered by matrix. In AMNH 30558, only a small portion of the supraoccipital is preserved on the left side. As in Sandownia harrisi, the dorsal exposure of the supraoccipital on the skull roof is very small. It is diamond-shaped and posterior to the parietals. The crista supraoccipitalis is complete on AMNH 30001. It is long but completely covered by parietals dorsally, as in Erquelinnesia gosseleti. The supraoccipital forms the upper rim of the foramen magnum and contacts the parietal anterodorsally, the prootic and opisthotic laterally and the exoccipital ventrally.
Prootic: Both prootics are complete in AMNH 30001, but due to the very complete skull roof, its dorsal surface is not prepared. Only the ventral side of the right prootic is visible. In AMNH 30558, both prootics are prepared on ventral and dorsal sides; the left prootic is nearly complete; the right one is damaged.
The prootic forms the medial part of an obvious processus trochlearis oticum. The processus is very large and curved. It is convex dorsally and concave ventrally with a sharp anteroventral edge. The processus is formed by the prootic medially and the quadrate laterally, the prootic contribution to the process being less than that of the quadrate. The size of the process is comparable to that of Euclastes wielandi from Morocco, but smaller than that of Tasbacka ouledabdounensis (Tong and Hirayama 2002) and Erquelinnesia gosseleti (Zangerl 1971). In E. wielandi the processus has a nearly flat ventral surface and it is formed equally by prootic and quadrate. In this new taxon, the ventral surface of the processus is more concave and mostly formed by the quadrate. In the Trionychoidae, there is “large portion of processus trochlearis oticum made up by parietal” (Meylan and Gaffney 1989, p. 46). Adocus sp. also has an unusually large and developed processus trochlearis oticum (Meylan and Gaffney 1989) that is formed by the parietal, prootic and quadrate, as in trionychids, and it is dorsally concave, unlike the case in our specimens in which the process is dorsally convex.
The foramen stapedio-temporale is complete on the left side of AMNH 30558, and damaged on the right side. This area is not prepared in AMNH 30001. The foramen lies on the prootic-opisthotic-quadrate suture. It is a large opening (3 mm in diameter anteroposteriorly) and extending medially from it is a large and deep groove. It is much larger than the foramen anterius canalis carotici interni. The difference in size between these two foramina seems to be comparable to Plesiochelys spp. and Portlandemys mcdowelli (Gaffney 1976). The foramen and the groove are formed mainly in the prootic. The quadrate contribution is limited to the anterolateral margin of the foramen and the opisthotic contributes to the posterolateral margin. On the dorsal surface of the otic chamber, the prootic contacts the supraoccipital posteromedially, the opisthotic posteriorly and the quadrate laterally. A prootic-parietal contact is not preserved in AMNH 30558, and not visible in AMNH 30001. In AMNH 30558, the supraoccipital and the medial part of the prootic are missing on the right side, exposing the cavum labyrinthicum. The fenestra ovalis and the recessus labyrinthicus prooticus are visible within.
Anteroventrally, the prootic forms the dorsal margin of the foramen nervi trigemini, which is well preserved on the left side of AMNH 30558. This foramen is a small rounded opening. The prootic contacts the pterygoid ventromedially.
Opisthotic: Both opisthotics are complete in AMNH 30001 and on the left side of AMNH 30558. The right opisthotic is damaged in AMNH 30558. Only the posterior surface is exposed between the exoccipital and quadrate in AMNH 30001, it is fully prepared in AMNH 30558.
The opisthotic is a blade-like bone, forming the dorsal margin of the fenestra postotica, part of the dorsal roof of the cavum acustico-jugulare, and dorsolateral margin of the foramen jugulare posterius. On the dorsal surface, it contributes also to the posterolateral margin of the foramen stapedio-temporale. The opisthotic contacts the prootic anteriorly, the quadrate laterally, the supraoccipital dorsomedially and the exoccipital ventromedially.
In the right cavum labyrinthicum of AMNH 30558, the processus interfenestralis, the recessus labyrinthicus opisthoticus, and the canalis semicircularis posterius are exposed. The hiatus acousticus and the foramen jugulare anterius are visible on both sides.
Lower jaw (Fig. 13.4): The lower jaw of AMNH 30558 is nearly complete; with the left ramus slightly damaged.
Dentary: The right dentary is complete; the left one is slightly damaged. The left and right dentaries are fused as in most turtles. The dentary forms a large triturating surface and most of lateral surface of the ramus. It contacts the coronoid dorsally, the surangular posteriodorsolaterally and the angular posteroventrally. The triturating surface is large, wide and almost flat. The labial margin forms an obtuse angle; the lingual margin is parallel to the anterior portion of the labial margin. The symphysis is long and forms the highest point of the triturating surface. The surface slopes downward from the back to the front and from the symphysis laterally. There is neither lingual nor labial ridge on the lower triturating surface, but a median notch is present at the anterior margin. However, the posterolateral border of the triturating surface bears a small ridge which delimits a small depression posteromedial to it, anterolateral to the processus coronoideus. This small depression faces the deep pit in the jugal on the upper triturating surface when the lower jaw is attached to the skull. Very large nutrient foramina on the anterior part of the lower triturating surface suggest that the rhamphotheca was thicker anteriorly than posteriorly. The morphology of the triturating surface is very different from cheloniids which have an extensive secondary palate such as Erquelinnesia gosseleti. (Zangerl 1971), Euclastes spp. (Zangerl 1953; Hirayama and Tong 2003), Tasbacka spp. (Nessov 1987) and pitted pleurodires such as Bothremys spp. (Gaffney and Zangerl 1968; Gaffney et al. 2006). The lower jaw is quite comparable to that of Sandownia harrisi in that (1) the symphysis of the dentary is long, (2) the lower triturating surface is broad and triangular and slopes ventrally from the coronoid process, (3) labial and lingual ridges are absent, and (4) there is a clear constriction anterior to the coronoid. However, Sandownia differs in that is has a longer symphysis and stronger constriction.
The ventral surface of the lower jaw is perfectly flat and horizontal with a midline groove under the triturating surface. The groove is narrow posteriorly and widens anteriorly resulting in a median notch that is seen in dorsal view. The lingual surface of the dentary bears a deep sulcus cartilaginis Meckelii. The foramen alveolare inferius is a small oval opening lying on the bottom of the sulcus. At the posterior end of the sulcus, there is a small sutural area along the lower margin just opposite of another larger sutural area on the coronoid. These suture surfaces are presumably for contact with the splenial which appears to have been lost on both sides.
The lateral surface of the lower jaw posterior to the triturating surface bears a deep longitudinal groove along its lower margin, which is bordered by a sharp horizontal ridge inferiorly. The lateral surface of the dentary above the groove is generally convex laterally, with a shallow depression just below the small lateral ridge of the lower triturating surface mentioned above. In this depression is a small, rounded foramen dentofaciale majus. A blunt and very low anterodorsally–posteroventrally directed swelling lies under the coronoid process. A more developed ridge is present along the dentary-surangular suture (see surangular).
The lateral depression of the dentary extends onto the surangular and all the way to the posterior end of the retroarticular process. It would appear to form an enlarged area for insertion of a very well developed zygomaticomandibularis muscle as in trionychids. In particular, the sharp horizontal ridge along the ventral margin of the dentary is in the same location as a ridge that marks the ventral limit of insertion of this muscle in a jaw of Apalone ferox.
Coronoid: The left coronoid is complete, while the right one is slightly damaged. The coronoid alone forms the rather tall processus coronoideus which lies at about mid-length of the ramus. The anterior surface of the process bears a deep pit in the posterior part of the lower triturating surface. There is a small foramen on the bottom of the pit on the left side, while three foramina are present in the right pit. The tall coronoid process lying at the midpoint of the jaw ramus length is present in trionychids, Adocus sp. (Meylan and Gaffney 1989) and carettochelyids [Carettochelys insculpta (Gaffney 1979), Anosteira maomingensis (Tong et al. 2010)]. The process of AMNH 30558 is more developed than that of cheloniids, such as Chelonia mydas and Lepidochelys kempii, but not as tall as in members of the Trionychoidae, such as Adocus sp., Amyda cartilaginea, and carettochelyids. The coronoid process is formed by the coronoid only in AMNH 30558, like that in carettochelyids, but unlike in Adocus sp. and A. cartilaginea (Meylan and Gaffney 1989, Fig. 9) and Lissemys punctata, in which the dentary makes up part of the process. Sandownia harrisi also has a tall coronoid process formed only by the coronoid, which is similar to AMNH 30558.
Posteriorly, the coronoid contributes to the margin of the fossa Meckelii; ventromedially it forms part of the upper margin of the sulcus cartilaginis Meckelii. On the lateral margin of the fossa Meckelii, the coronoid forms, together with the surangular posteriorly, a dorsal process. The coronoid contacts the dentary anteriorly and ventrolaterally, the surangular posterolaterally and the prearticular posteromedially. A sutural surface can be seen on the lower margin of the medial surface of the coronoid, which is presumably for the splenial contact.
Surangular: Both surangulars are complete. The surangular forms the posterolateral part of the jaw ramus. A well developed ridge along the dentary-surangular suture extends posteriorly onto the surangular and roughly parallel to the lateral margin of the area articularis mandibularis. There is a small, apparently unnamed foramen under the ridge, and above the dentary-surangular suture. Anterodorsally, the surangular contributes to the posterolateral rim of the fossa Meckelii and the small process on the lateral margin of the fossa (see coronoid). The surangular contacts the coronoid anterodorsally, the dentary anteroventrally, the articular posterodorsally and the angular posteroventrally. The foramen nervi articulotemporalis is a small, anterior-facing opening that lies under the anterolateral part of the area articularis mandibularis.
Prearticular : The right prearticular is complete; the left is a little damaged. The prearticular is a sheet of bone contributing to the medial rim of the fossa Meckelii. It contacts the coronoid anteriorly, the articular posteriodorsally and the angular ventrally. There is a suture surface on the anteroventral margin of the prearticular, which is a continuation of a similar surface on the coronoid and is opposite to a sutured surface on the dentary. These suture surfaces are presumably for the splenial contact, although the splenial is not preserved.
Articular: Both articulars are slightly damaged at their posteromedial surface. The articular forms the large area articularis mandibularis which is made up by two facets: the larger lateral facet and smaller median one. Both medial and lateral facets are slightly concave and separated by an anteroposteriorly elongated swelling. Posterior to the articular area, the processus retroarticularis is damaged, so the whole shape and length cannot be determined. The articular contacts the surangular ventrolaterally, the prearticular ventromedially. The foramen posterius chorda tympani is not preserved on either side.
Angular: Both angulars are almost complete. The angular makes up the posteroventral part of the lower jaw. It contacts the dentary anteriorly, the prearticular dorsomedially and the surangular dorsolaterally. Ventrolaterally, the angular makes up, with the dentary, a sharp and horizontal ridge at the lower margin of the ramus, above and along this ridge lies a long and deep groove.
Splenial: The splenial itself is not preserved on either side, but sutured surfaces on the coronoid, prearticular and dentary posterior to the sulcus cartilaginis Meckelii indicate its presence. These sutured surfaces can be seen on both sides of the lower jaw, but are better preserved on the right side. The presence of the splenial is primitive since it is absent in most recent turtles but it occurs commonly in Mesozoic forms (Gaffney 1979).
Discussion
Morphology of Brachyopsemys
This is the first report of palatal pits in a member of the Cryptodira. Palatal pits were previously known only in one group of pleurodires; they characterize the subtribe Bothremydina in the family Bothremydidae (Gaffney et al. 2006). This subtribe includes five genera and nine species, and has a wide geographic distribution (North America, North Africa, Southern Europe and Middle East) and a long geological range (Late Cretaceous to Eocene). Within the Bothremydina is one genus (two species) that lack palatal pits, and they are considered to be secondarily lost. Both Brachyopsemys and members of the Bothremydini have a low, broad skull and extensive secondary palate. The general morphology and position of the palatal pits in Brachyopsemys is comparable to those of Bothremydina, such as Bothremys maghrebiana (Fig. 13.5b). However, when compared in detail, some differences can be noted in the morphology and composition of the secondary palate. Brachyopsemys has a complete secondary palate without midline separation. A similar condition is also known in cheloniids (Erquelinnesia, Tasbacka, Euclastes, Puppigerus) and the eurysternid, Solnhofia parsonsi. The secondary palate of Bothremydina is divided by a midline groove. In addition, the palate of Brachyopsemys has a significant contribution by the pterygoid, in contrast to Bothremydini. The exact composition of the pits also differs. In Bothremydina the palatal pits are formed by the maxilla, jugal and palatine; while in Brachyopsemys, the pits are formed by the maxilla and jugal only, without a contribution from the palatine. The jugal contributes to the palatal pit laterally in Brachyopsemys rather than posteriorly or only on the bottom of the pit, as seen in Bothremydina.

Comparison of a pitted palate cryptodire (Brachyopsemys) and pleurodire (Bothremys). a Reconstruction of palate of Brachyopsemys. b Reconstruction of palate of Bothremys maghrebiana (after Gaffney et al. 2006). Not to scale
Among turtles with pitted palates, the shape and the depth of the pit varies, from a wide and relatively shallow depression to a well-defined and deep rounded pit. The appearance of a secondary palate in turtles appears to be a trophic modification with the development of the triturating area into a broad crushing surface (Meylan et al. 2000). Turtles with enlarged triturating surface usually have durophagous diet. The function of such a pit is likely to help to hold slippery shells (brachiopods, mollusks) to facilitate crushing. Variation in the shape and size of the pits probably limits the size range of the prey.
Phylogenetic Analysis
In an attempt to understand the phylogenetic position of this new taxon, we scored it for the data set developed by Gaffney et al. (1991) and expanded by Shaffer et al. (1997) and Near et al. (2005). This data set is further expanded here to include additional taxa and new characters. Characters 1–41 are those of Gaffney et al. (1991) and are described in that paper. Characters 42–115 are described in Shaffer et al. (1997). Characters 116–142 were used in analyses to place fossils in Near et al. (2005) but were not described in that paper. They are included here (Appendices 13.1 and 13.2) along with descriptions and scores of seven additional characters that were added to help resolve the position of Brachyopsemys. Thus, scores for a total of 149 characters are given in Appendix 13.2. During the course of this study, it was noted that character 23 of Gaffney et al. (1991) is the same as character 109 of Shaffer et al. (1997). The description of character 109 is more complete so character 23 was effectively deleted from the data set by scoring all taxa as “?”. This method for eliminating character 23 means that characters were not renumbered which we expect will facilitate comparisons among studies.
In order to examine the possiblity that Brachyopsemys might be related to Sandownia and Angolachelys and to test the hypothesis that these taxa may be more closely related to Solnhofia than to other cryptodires [i.e., members of the “Angolachelonia” of Mateus et al. (2009)], we have included Solnhofia on the basis of Gaffney (1975) and Joyce (2000), Thalassemys moseri on the basis of Rieppel (1980), Erquelinnesia on the basis of Zangerl (1971), Angolachelys on the basis of Mateus et al. (2009), Euclastes on the basis of Hirayama and Tong (2003); Tasbacka on the basis of Tong and Hirayama (2002) and Nessov (1987), and Brachyopsemys based on the current study. The ‘Glen Rose turtle’ is not included in our cladistic analysis.
Danilov and Parham (2005) have noted that a skull previously interpreted as Lindholmemys (Nessov 1986; Shaffer et al. 1997) is an adocid, necessitating removal of this genus from the data set.
Results of Phylogenetic Analysis
We are able to code 58 of 149 characters for Brachyopsemys. For this analysis five shell taxa that seemed to be of little relevance to this study (Peltochelys, Aspideretes maortuensis, Hadrianus, Hoplochelys and Pelusios rusingae) were excluded. A heuristic search using PAUP* 4.0b of the data set given in Appendix 13.2 produced 65 equally parsimonious trees of 366 steps each. A majority-rule consensus tree is given in Fig. 13.6. The percent of 366 trees supporting each node is given above the preceding stem, values for 100 bootstrap replicates is indicated below each stem.

Majority-rule consensus tree of 65 trees of 366 steps each, resulting from a heuristic search of the data set given in Appendix 2 (excluding the shell taxa Peltochelys, Aspideretes maortuensis, Hadrianus, Hoplochelys, and Pelusios rusingae), showing inferred relationships of the new genus Brachyopsemys (bolded) and the new family Sandownidae. Majority-rule values are given to left of each branch; bootstrap values greater than 50 (for 100 replicates) are given to right of each branch
Monophyly of Sandownidae
The monophyly of Sandownidae is supported by a series of characters including the significant contribution made by the jugal to the upper triturating surface, the posterior extension of the pterygoids to partly or completely cover the basisphenoid, the partial or complete fusion of the premaxillae that are slightly protuberant anteriorly, the absence of the external process of the pterygoid, the absence of the foramen praepalatinum, and the L-shaped prefrontal that sends a posteriolateral process along the ventral margin of the orbit (this reaches the jugal in Brachyopsemys). In addition, all three taxa show a complete skull roof in which parietal-squamosal contact excludes the postorbital from temporal exposure (Table 13.2).
The Phylogenetic Position of Sandownidae
Brachyopsemys and the other members of the Sandownidae can be referred to the Cryptodira on the basis of a very well-developed processus trochlearis oticum for the main adductor tendon of the jaw adductor musculature (Gaffney and Meylan 1988). The morphology of the pterygoid and in particular the condition of the palatine artery and internal carotid artery suggest referral to the Polycryptodira (Gaffney 1996). Although monophyly of the family seems firmly supported, the position of the Sandownidae among the Polycryptodira is problematic. Meylan et al. (2000) focused on the trionychoid-like features of Sandownia harrisi and employed an analysis of limited scope to suggest that this form was a trionychoid related to Carettochelys and the Trionychidae. However, those authors acknowledged that Sandownia has features of the Chelonioidea. Joyce (2007) included Sandownia in his cladistic analysis of Mesozoic turtles, and suggested that Sandownia has “no firm affinities with any group of Cryptodires”. He also stated that “placement of Sandownia harrisi as sister to ‘Talassemys marina’ within Clade 20 only requires one step more than its placement as a basal representative of Trionychoidea” (Joyce 2007, p. 66). His choice among alternative options for placement of Sandownia was based in part on stratigraphic considerations. In spite of the fact that three species of sandownid are now known from skulls, we have to agree with the conclusion of Joyce (2007) stated above. Although the phylogenetic utility of the skull of turtles has been firmly established over the last four decades, the observation that no firm conclusion can yet be made about the phylogenetic position of the Sandownidae suggests that there may be occasions in which it will be availability of well preserved shells that will result in resolution of turtle phylogeny. The trionychoid and chelonioid features of the Sandownidae are discussed below.
Two important characters bearing on the phylogenetic position of the Sandownidae merit further discussion. Our assessment of the material described by Mateus et al. (2009) suggests that nasals are probably absent from Angolachelys mbexi. However, the authors interpreted the material to include nasals. Nasals are also reported as present in the Glen Rose turtle and “not expanded laterally and displaced anteriorly by the prefrontals” (Barck 1992, p. 18). If the Sandownidae were derived from a primitive chelonioid stem, one might expect nasals to be present basally. Similarly, Mateus et al. (2009) have interpreted the foramen palatinum posterius to be absent in Angolachelys as in chelonioids (they are present in Sandownia and Brachyopsemys). We consider the preservation in this area of the palate of Angolachelys to be questionable. Absence of the foramen palatinum posterius would be a chelonioid feature but presence in Sandownia and Brachyopsemys would require a reversal of this character.
Trionychian Features of Sandownidae
Although Brachyopsemys has some contribution by the palatine to the sidewall of the braincase, an important character of Trionychoidae and Kinosternidae, the contribution cannot be considered to be as significant as in Adocus sp. (Meylan and Gaffney 1989) or other taxa that have been referred to these two groups. Rather the dorsal plate of the palatine overlaps the lateral surface of the anterior part of a greatly thickened crista pterygoidea forming a thin veneer on the lateral surface. There is no contact to the descending process of the parietal as is seen in Trionychia and kinosternids. This condition differs from both chelonioids and trionychoids, but is somewhat closer to trionychoids. In fact, the contribution of the palatine to the sidewall of the braincase is related to the length of the sidewall itself and the position of the palatine. In trionychoids, the sidewall of the braincase is long, the ventral portion being made by the palatine anteriorly and pterygoid posteriorly. While the chelonioids have a very short and tall sidewall of the braincase, in which the ventral portion is made up entirely by pterygoid. The relative position of the palatine to the braincase in Brachyopsemys specimens thus more closely resembles the condition in trionychoids. The fact that the palatine does not contribute to the whole thickness of the sidewall of the braincase is due to the anterior position of the pterygoids. The palatine midline contact can be thus considered as secondarily lost, probably due to the forward shift of the basicranium. The type description of Angolachelys indicates that the palatine does not reach the sidewall of the braincase.
Members of Sandownidae also resemble the Trionychia in the absence of the external process of the pterygoid (character 77), the reduced vomer (not reduced in Sandownia harrisi), the maxillae meeting on the midline between the premaxilla and the vomer, and partial or complete fusion of the premaxillae (character 81). Fused premaxillae are also present in the cheloniid Erquelinnesia gosseleti (Zangerl 1971). The completely fused premaxillae in Brachyopsemys and Angolachelys are more derived than partially fused premaxilla in Sandownia (Meylan et al. 2000). The tall coronoid process located near the middle of the mandibular ramus is also present in Trionychoidae, such as the adocid Adocus (Meylan and Gaffney 1989), the nanhsiungchelyid Zangerlia neimongolensis (Brinkman and Peng 1996) and the carettochelyid Anosteira maomingensis (Tong et al. 2010). There is vomer to pterygoid contact (character 75) in Sandownia, Angolachelys and Brachyopsemys as in Trionychia.
The zygomaticomandibularis muscle is only reported from the Trionychidae (Schumacher 1973). However, surveys of the jaw musculature of living turtles are still incomplete and careful study of the homology of this muscle and the identification of criteria for its detection in fossil skulls remain to be established. However, the possibility that further study of this muscle might help to elucidate relationships of the Sandownidae should not be overlooked. Sandownia harrisi would appear to have the same depressed area below the coronoid on the lateral surface of the jaw (Meylan et al. 2000, Fig. 4c).
Other important characteristics of the Trionychia are clearly absent from members of Sandownidae including parietal contribution to the processus trochlearis oticum (character 74) and the enlarged basisphenoid (character 79). In fact the basisphenoid is reduced (at least in ventral exposure) in sandownids. Furthermore, Brachyopsemys has a large foramen stapedio-temporale which is much larger than the foramen anterius canalis carotici interni. This suggests that it lacks ‘decrease of stapedial circulation and increase of carotid circulation’, a feature previously considered a synapomorphy of Trionychoidea (Meylan and Gaffney 1989). Recent studies on the cranial circulation of turtles do not support the monophyly of Trionychoidea (Trionychoidae plus Kinosternoidae) (Jamniczky and Russell 2007; Jamniczky 2008), as is also the case for recent molecular phylogenies (Shaffer et al. 1997; Fujita et al. 2004). The enlarged foramen stapedio-temporale in Brachyopsemys may represent a primitive condition as seen in Testudinoidea, primitive cheloniids and also plesiochelyids.
Chelonioid Features of Sandownidae
The most apparent suite of chelonioid characters of Sandownidae is a set of correlated characters of the skull roof. Because of strong parietal-squamosal contact (character 15), temporal exposure of the postorbital (character 111) is absent as is the presence of a short postorbital exposed by temporal emargination (character 142). Although contact of the parietal and squamosal is present in Proganochelys quenstedti (Gaffney 1990) and primitive cryptodires such as Dorsetochelys delairi (Evans and Kemp 1976), Mesochelys durlstonensis (Evans and Kemp 1975), and some baenids (Gaffney 1979), posterior exposure of a long postorbital in chelydrids and some potentially basal chelonioids (i.e., Toxochelys latiremis) suggests that the extensive skull roof, that includes parietal-squamosal contact in some members of the Chelonioidea, may be a reversal to the primitive condition.
The antrum postoticum (character 140) is small in all members of the Sandownidae for which it is known as is the case in chelonioids. Similarly the foramen praepalatinum is absent from the palate (character 145) in both groups. This foramen is also absent in Trionychia but in this case absence is due to the presence of a large intermaxillary foramen.
The fortuitous preservation of Brachyopsemys specimen AMNH 30558 allows study of the floor of the braincase and reveals what may be the most important chelonioid-like characters. The trabeculae of the basisphenoid clearly converge and join anterior to a small sella turcica. Although not as close as in Chelonia mydas, the foramen anterius canalis carotici interni are relatively close together (character 64) and furthermore the dorsum sellae does not overhang or conceal the sella turcica (character 65) either in Sandownia harrisi or in Brachyopsemys. However, it is also clear from this specimen that the foramen caroticum laterale in Brachyopsemys is not enlarged as it is in chelonioids (character 91); see also Hirayama (1994, character 29).
Brachyopsemys and Sandownia harrisi are also chelonioid-like in have clearly defined scale areas on the skull roof. The presence-absence of skull roof scales was not included in the data matrix, but it should be noted that within the Trionychia scales are absent from the skull roof and some clades have a distinctive sculpture pattern (carettochelyids) that is not seen in members of the Sandownidae. Some nanhsiungchelyids [Nanhsiungchelys wuchingensis (Ye 1966)] have both skull roof scales and a sculptured skull roofing surface.
The chelonioid affinity of sandownids is further supported by the lack of shell ornamentation. Although no shell material has been associated with Brachyopsemys, and Sandownia (Meylan et al. 2000), shell fragments collected along with Angolachelys (Mateus et al. 2009) and the Glen Rose turtle (Barck 1992) have no ornamentation on the surface. All Trionychoidae have a sculptured shell surface.
Relationships Within Sandownidae
Within the Sandownidae, Sandownia and Angolachelys appear more closely related to each other than to Brachyopsemys. Sandownia and Angolachelys share the derived feature, closure of the incisura columellae auris. Meylan et al. (2000) suggested that this feature in Sandownia harrisi supported its referal to the Trionychia. The presence of an open incisura in Brachyopsemys suggests that closure has occurred within the Sandownidae.
Barck (1992) reported an undetermined eucryptodiran turtle (the Glen Rose turtle) from the Early Cretaceous of Texas. The wide and low skull, extensive skull roof, forward position and closely spaced orbits, large triturating surface and condylus occipitalis located between the condylus mandibularis of the quadrate resemble Brachyopsemys and other members of the Sandownidae. The skull from Texas is not described in detail, however, it was included in the Mateus et al. (2009) data set and was resolved as the sister group of Sandownia plus Angolachelys. In a separate study, it is resolved as the sister taxon of (Euclastes [Sandownia (Angolachelys+Erquelinnesia)]) (Vineyard and Jacobs 2009). When this species is fully described, inclusion in a phylogenetic analysis will almost certainly place it within Sandownidae.
Distribution of Sandownidae
The four members of the family (Sandownia, the Glen Rose turtle, Angolachelys and Brachyopsemys) have a known geological range from the Aptian to Paleocene. The oldest representative, Sandownia is from the early Aptian Lower Lobster beds within the Lower Greensand Formation, Isle of Wight, England. The rich microflora dominated by non-marine taxa and low diversity of marine components indicates a restricted, near-shore marine environment, with reduced salinity (Meylan et al. 2000). The second taxon, the Glen Rose turtle, comes from the Glen Rose Formation of Hood County, in central Texas. The Glen Rose Formation consists of limestones and marls deposited in marginal marine environments. It is of early Albian age and has yielded abundant invertebrates, vertebrates and plant remains (Barck 1992; Vineyard and Jacobs 2009). Angolachelys is the only member of the group from the south Atlantic. It is from the Turonian of Iembe, in the Kawanza Basin, Angola. The shallow marine sediments of the Tadi Beds of the Itombe Formation at the Iembe site and nearby localities have yielded ammonites, sharks, bony fishes, mosasaurs, plesiosaurs and sauropod dinosaurs (Jacobs et al. 2006; Mateus et al. 2009). Brachyopsemys is the youngest member of the group. The phosphate sediments of the Ouled Abdoun Basin were deposited in the long, narrow gulf opening to the Atlantic margin during the latest Cretaceous and early Tertiary. Abundant vertebrate remains, consisting mostly of marine taxa, have been collected from that basin (Gaffney et al. 2006). From a paleogeographical and paleoecological point of view, this family appears to be a bottom-dwelling group that occurred in near-shore, shallow marine environments along the coast of the Atlantic (Fig. 13.7). Its evolutionary history is likely to be connected to the development of the Atlantic Ocean. The addition of the Paleocene Brachopsemys to the group indicates that this is another family of turtles that survived the Cretaceous-Tertiary boundary.

Paleogeographical reconstruction of Maastrichtian stage (Smith et al. 1994), showing the position of the members of the family Sandownidae. 1 Glen Rose turtle; 2 Sandownia; 3 Brachyopsemys; 4 Angolachelys
References
Barck, A. (1992). Paleontology of the Glen Rose Formation (Lower Cretaceous), Hood Country, Texas. The Texas Journal of Science, 44, 3–24.
Brinkman, D. B., & Peng, J.-H. (1996). A new species of Zangerlia (Testudines: Nanhsiungchelyidae) from the Upper Cretaceous redbeds at Bayan Mandahu, Inner Mongolia, and the relationships of the genus. Canadian Journal of Earth Sciences, 33, 526–540.
Dalrymple, G. H. (1977). Intraspecific variation in the cranial feeding mechanism of turtles of the genus Trionyx (Reptilia, Testudines, Trionychidae). Journal of Herpetology, 11, 255–285.
Danilov, I. G., & Parham, J. F. (2005). A reassessment of the referral of an isolated skull from the Late Cretaceous of Uzbekistan to the stem-testudinoid turtle genus Lindholmemys. Journal of Vertebrate Paleontology, 25, 784–791.
Evans, J., & Kemp, T. S. (1975). The cranial morphology of a new Lower Cretaceous turtle from southern England. Palaeontology, 18, 25–40.
Evans, J., & Kemp, T. S. (1976). A new turtle skull from the Purbeckian of England and a note on the early dichotomies of cryptodiran turtles. Palaeontology, 19, 317–324.
Fujita, M. K., Engstrom, T. N., Starkey, D. E., & Shaffer, H. B. (2004). Turtle phylogeny: Insights from a novel nuclear intron. Molecular Phylogenetics and Evolution, 31, 1031–1040.
Gaffney, E. S. (1975). Solnhofia parsonsi, a new Cryptodiran turtle from the Late Jurassic of Europe. American Museum Novitates, 2576, 1–25.
Gaffney, E. S. (1976). Cranial morphology of the European Jurassic turtles Portlandemys and Plesiochelys. Bulletin of the American Museum of Natural History, 157, 489–543.
Gaffney, E. S. (1979). Comparative cranial morphology of recent and fossil turtles. Bulletin of the American Museum of Natural History, 164, 67–376.
Gaffney, E. S. (1990). The comparative osteology of the Triassic turtle Proganochelys. Bulletin of the American Museum of Natural History, 194, 1–263.
Gaffney, E. S. (1996). The postcranial morphology of Meiolania platyceps and a review of the Meiolaniidae. Bulletin of the American Museum of Natural History, 229, 1–166.
Gaffney, E. S., & Meylan, P. A. (1988). A phylogeny of turtles. In M. J. Benton (Ed.), The phylogeny and classification of the tetrapods (pp. 157–219). Oxford: Clarendon Press.
Gaffney, E. S., & Meylan, P. A. (1992). The Transylvanian turtle, Kallokibotion, a primitive Cryptodire of Cretaceous age. American Museum Novitates, 3040, 1–37.
Gaffney, E. S., Meylan, P. A., & Wyss, A. R. (1991). A computer assisted analysis of the relationships of the higher categories of turtles. Cladistics, 7, 313–335.
Gaffney, E. S., & Tong, H. (2003). Phosphatochelys, a new side-necked turtle (Pelomedusoides: Bothremydidae) from the Paleocene of Morocco. Bulletin of the American Museum of Natural History, 13, 644–659.
Gaffney, E. S., & Zangerl, R. (1968). A revision of the chelonian genus Bothremys (Pleurodira: Pelomedusidae). Fieldiana Geology, 16, 193–239.
Gaffney, E. S., Tong, H., & Meylan, P. A. (2006). Evolution of the side-necked turtles: The families Bothremydidae, Euraxemydidae, and Araripemydidae. Bulletin of the American Museum of Natural History, 300, 1–700.
Hirayama, R. (1994). Phylogenetic systematics of chelonioid sea turtles. The Island Arc, 3, 270–284.
Hirayama, R., & Tong, H. (2003). Osteopygis (Testudines: Cheloniidae) from the Lower Tertiary of the Ouled Abdoun phosphate basin, Morocco. Palaeontology, 46, 845–856.
Hutchison, J. H., & Bramble, D. M. (1981). Homology of the plastral scales of the Kinosternidae and related turtles. Herpetologica, 37, 73–85.
Jacobs, L. L., Mateus, O., Polcyn, M. J., Schulp, A. S., Antunes, M. T., Morais, M. L., et al. (2006). The occurence and geological setting of Cretaceous dinosaurs, mosasaurs, plesiosaurs, and turtles from Angola. Journal of Paleontological Society of Korea, 22, 91–110.
Jamniczky, H. A. (2008). Turtle carotid circulation: A character analysis case study. Biological Journal of the Linnean Society, 93, 239–256.
Jamniczky, H. A., & Russell, A. P. (2007). Reappraisal of patterns of nonmarine Cryptodiran turtle carotid circulation: Evidence from osteological correlates and soft tissues. Journal of Morphology, 268, 571–587.
Joyce, W. G. (2000). The first complete skeleton of Solnhofia parsonsi (Cryptodira, Eurysternidae) from the Upper Jurassic of Germany and its taxonomic implications. Journal of Paleontology, 74, 684–700.
Joyce, W. G. (2007). Phylogenetic relationships of Mesozoic turtles. Bulletin of the Peabody Museum of Natural History, 48, 3–102.
Joyce, W. G., Chapman, S. D., Moody, R. T. J., & Walker, C. A. (2011). The skull of the solemydid turtle Helochelydra nopcsai from the Early Cretaceous of the Isle of Wight (UK) and a review of Solemydidae. Special Papers in Palaeontology, 86, 75–97
Karl, H.-V., Tichy, G., & Ruschark, H. (1998). Osteopygoides priscus n. gen. n. sp. und die Taxonomie und Evolution der Osteopygidae (Testudines: Chelonioidea). Mitteilungen der Geologischen und Palaontologischen Landesmuseum Joanneum, 56, 329–350.
Lyson, T. R., & Joyce, W. G. (2009). A revision of Plesiobaena (Testudines: Baenidae) and an assessment of baenid ecology across the K/T boundary. Journal of Paleontology, 83, 833–853.
Mateus, O., Jacobs, L., Polcyn, M., Schulp, A. S., Vineyard, D., Neto, A. B., et al. (2009). The oldest African Eucryptodiran turtle from the Cretaceous of Angola. Acta Palaeontologica Polonica, 54, 581–588.
Meylan, P. A. (1987). The phylogenetic relationships of soft-shelled turtles (Family Trionychidae). Bulletin of the American Museum of Natural History, 186, 1–101.
Meylan, P. A., & Gaffney, E. S. (1989). The skeletal morphology of the Cretaceous Cryptodiran turtle, Adocus, and the relationships of the Trionychoidea. American Museum Novitates, 2941, 1–60.
Meylan, P. A., Moody, R. T. J., Walker, C. A., & Chapman, S. D. (2000). Sandownia harrisi, a highly derived trionychoid turtle (Testudines: Cryptodira) from the Early Cretaceous of the Isle of Wight, England. Journal of Vertebrate Paleontology, 20, 522–532.
Near, T. J., Meylan, P. A., & Shaffer, H. B. (2005). Assessing concordance of fossil calibration points in molecular clock studies: An example using turtles. The American Naturalist, 165, 137–146.
Nessov, L. A. (1977). Skull morphology of the Early Cretaceous turtle belonging to the family Adocidae. Trudy Zoologicheskogo Instituta AN SSSR, 74, 75–79.
Nessov, L. A. (1986). Some Late Mesozoic and Paleocene turtles of Soviet Middle Asia. Studia Geologica Salamanticensia, volumen Especial 2. Studia Palaeocheloniologica, 2, 7–22.
Nessov, L. A. (1987). The Paleogene sea turtles of southern Kazakhstan and the phylogenetic relationships between the Toxochelyidae and the Cheloniidae. Palaeontological Journal, 4, 76–87.
Nour-Eddine, J., de Lapparent de Broin, F., Bardet, N., Vacant, R., Bouya, B., Amaghzazc, M., & Meslouh, S. (2009). Euclastes acutirostris, a new species of littoral turtle (Cryptodira, Cheloniidae) from the Palaeocene phosphates of Morocco (Oulad Abdoun Basin, Danian-Thanetian). Compte Rendu Palevol, 8, 447–459.
Rieppel, O. (1980). The skull of the Upper Jurassic cryptodire turtle Thalassemys, with a reconsideration of the chelonian braincase. Palaeontographica, 171, 105–140.
Schumacher, G.-H. (1973). The head muscles and hyolaryngeal skeleton of turtles and crocodilians. In: C. Gans & T. S. Parsons (Eds.), Biology of the Reptilia (pp. 101–199). London, New York: Academic Press.
Shaffer, H. B., Meylan, P. A., & McKnight, M. L. (1997). Tests of turtle phylogeny: Molecular, morphological, and paleontological approches. Systematic Biology, 46, 235–268.
Smith, A. G., Smith, D. G., & Funnell, B. M. (1994). Atlas of Mesozoic and Cenozoic coastlines. Cambridge: Cambridge University Press.
Tong, H., & Hirayama, R. (2002). A new species of Tasbacka (Testudines: Cryptodira: Cheloniidae) from the Paleocene of the Ouled Abdoun phosphate basin, Morocco. Neues Jarrbuch fur Geologie und Palaeontologie, Monatsheft, 2002, 277–294.
Tong, H., & Hirayama, R. (2004). First Cretaceous dermochelyid turtle from Africa. Revue de Paléobiologie, spécial, 9, 55–59.
Tong, H., & Hirayama, R. (2008). A new species of Argillochelys (Testudines: Cryptodira: Cheloniidae) from the Ouled Abdoun phosphate basin, Morocco. Bulletin de la Société Géologique de France, 179, 623–630.
Tong, H., Hirayama, R., & Tabouelle, J. (in press). Puppigerus camperi (Testudines: Cryptodira: Cheloniidae) from the Ypresian (Early Eocene) of Ouled Abdoun Basin, Morocco. Bulletin de la Société Géologique de France.
Tong, H., Zhang, J.-Y., & Li, J.-J. (2010). Anosteira maomingensis (Testudines: Carettochelyidae) from the Late Eocene of Maoming, Guangdong, southern China: New material and redescription. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 256, 279–290.
Vineyard, D. (1999). Turtle skulls from the Early Cretaceous of Texas. Journal of Vertebrate Paleontology, 19(Suppl. 3), 82A.
Vineyard, D. P., & Jacobs, L. L. (2009). A new eucryptodiran turtle from the Mid-Cretaceous (Albian) Glen Rose Formation of Texas. Gaffney Turtle Symposium (October 17–18, 2009, Royal Tyrrell Museum, Drumheller, Canada), Abstract Volume, pp. 181–186.
Ye, X.-K. (1966). A new Cretaceous turtle of Nanhsiung, northern Kwangtung. Vertebrata Palasiatica, 10, 191–200.
Zangerl, R. (1953). The vertebrate fauna of the Selma Formation of Alabama. Part VI: The turtles of the Family Toxochelyidae. Fieldiana: Geology Memoirs, 3, 137–277.
Zangerl, R. (1971). Two toxochelyid sea turtles from the Landenian sands of Erquelinnes (Hainaut) of Belgium. Institut Royal des Sciences Naturelles de Belgique, Mémoires, 169, 1–32.
Acknowledgments
We thank F. Escuillié (Gannat) and B. Segaoui (Erfoud) for providing the specimens,; O. Mateus (Lisbon) for the data set of Angolachelys; E. S. Gaffney (New York), W. Joyce (Tübingen) and R. Hirayama (Tokyo) for reviewing the manuscript; J. Pfaller (Gainesville) for discussions of the zygomaticomandibularis, and the Singer-Polygnac Fondation and Eckerd College for financial support. AMNH 30001 was prepared by J. Klausen, J. Shumsky and J. Kelly (New York). The reconstruction of the palate was produced by F. Ippolito after A. Montalvo (New York). Special thanks to Gene Gaffney for providing opportunities to forward our careers.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Appendices
Appendices
Appendix 13.1
Description of Characters: the matrix used in the study is a further amended matrix based on those of Gaffney et al. (1991) (characters 1–39) and Shaffer et al. (1997) (characters 40–115). Characters 116–142 were used to place fossils in Near et al. (2005), but were not described in that paper. The remainder have been added for the purposes of this study.
116. Number of bony dermal callosities in the plastron reduced. Absent = 0, present = 1. Primitively the turtle plastron is composed of one midline element (entoplastron) and at least five pairs of bones (epi-, hyo-, meso-, hypo- and xiphiplastra) although some forms may have more than one pair of mesoplastra (Odontochelys semitestacea and Proterochersis robusta). In some clades, only a deep dermal ossification represents these elements and an overlying superficial dermal callosity is absent (Meylan 1987, character 9). Three or fewer pairs of dermal callosities in the plastron, as seen in members of the Trionychinae, is considered derived.
117. Suprascapular fontanelles. Absent = 0, present = 1. Primitively the adult turtle carapace is closed at the junction of the nuchal, first costal and first neural bones. An opening in this region, as seen in certain members of the Trionychinae is considered derived.
118. Shape of epiplastron. Triangular = 0, I-shaped = 1, J-shaped = 2. Primitively the epiplastron of turtles is a more or less triangular element with a variable exterior margin. In certain trionychids it is reduced to an I-shaped (1) or J-shaped (2) deep dermal ossification.
119. Bridge length. Long bridge = 0, short bridge = 1. In most turtles, including most primitive forms, the bridge is long, about one-third of the total plastron length. In those forms in which it is shorter, one-quarter or less of plastron length as in Chelydra serpentina, Staurotypus or Apalone, this is considered a derived condition.
120. Medial process of jugal. Present = 0, absent = 1. Most turtles have a medially directed process of the jugal that articulates with the pterygoid or pterygoid and palatine (Gaffney 1979, p. 79). Where this medial process is absent, as in Dermochelys coriacea, this is considered a derived condition.
121. Coracoid length. Dorsal process of scapula longer than coracoids = 0, coracoids longer than dorsal process of scapula = 1. Turtles have a triradiate pectoral girdle in which the dorsal process of the scapula is normally the longest of the three processes. Those forms in which the coracoid is longer than the dorsal process of the scapula, as in Dermochelys, Lepidochelys kempi or Podocnemis, are considered to have a derived condition.
122. Wide angle between dorsal process and acromion process of scapula. About right angle = 0, angle > 105 = 1. The dorsal process of the scapula forms an approximate right angle in Proganochelys and most turtles. Where this wider (>105o as in most testudinids) this can be considered a derived condition.
123. Pectineal process of pubis expanded. No = 0, yes = 1. In most turtles, including Proganochelys, the pectineal process (lateral pubic process of Gaffney 1990, Fig. 144) is narrower than the inter-pubic contact. Those forms in which this process is as wide or wider than the interpubic contact, as in Lissemys, Apalone, and Dermochelys, can be considered to have a derived condition.
124. Humerus length. Femur longer than humerus = 0, femur shorter than humerus = 1. In most turtles including Proganochelys (Gaffney 1990, Fig. 151) the femur is longer than the humerus. In certain marine species (i.e., Dermochelys, Lepidochelys) and land tortoises (Geochelone, Gopherus) the femur is shorter than the humerus. The latter is considered as a derived condition.
125. Simple, flat carpals and tarsals. Multifaceted carpals and tarsals = 0, simple flat carpals and tarsals = 1. In most turtles including Proganochelys (Gaffney 1990, Figs. 161, 174) the carpals and tarsals are multifaceted, complex elements. In forms with paddle-like limbs, these elements are simple, thin discs of bone.
126. Epiplastral lip. Absent = 0, present = 1. Only the most primitive turtles possess a dorsal process of the epiplastron (see character 112). Where this process is absent, the epiplastra are normally only slightly thickened at the shell margin. Where the epiplastron is greatly thickened along its anterior margin, as in many members of the Testudinidae, this is considered as a derived condition.
127. Pleuro-marginal scute sulcus coincident with costoperipheral suture. No = 0, yes or on costals = 1. In most turtles including Proganochelys (Gaffney 1990, Fig. 86) there are scute sulci on the peripheral bones marking the medial limits of the marginal scutes. Where supramarginal scutes are absent, these are pleuro-marginal scute sulci. When these sulci lie on the costoperipheral suture (as in some members of the Testudinidae) or on the costals (posteriorly in Adocus), this can be considered a derived condition.
128. Bridge peripheral height. Bridge peripherals the same size as remaining peripherals = 0, bridge peripherals twice as tall as the remaining peripherals = 1. Although the bridge of Proganochelys is not well known (Gaffney 1990, p. 127), in most turtles, including primitive forms, the bridge peripherals are approximately the same size as the more anterior and posterior peripherals. Those forms in which the bridge peripherals are approximately twice as tall as the remaining peripherals (as in Gopherus and Geochelone), can be considered to have a derived condition.
129. Coracoid fan-shaped. No = 0, yes = 1. The coracoid of most turtles is longer than wide and in the Casichelyida is somewhat wider distally than proximally. Those forms that have a very short but distally very wide (i.e., fan-shaped) coracoid (as in Gopherus or Kinixys), can be considered to have a derived condition.
130. Palatine contributes to triturating surface of upper jaw. Absent = 0, present = 1. In most turtles including Proganochelys the palatine does not appear on the triturating surface. Those forms in which it makes a significant contribution to the triturating surface (as in Graptemys or Podocnemis) can be considered to have a derived condition.
131. Foramen caroticopharyngeale. Small = 0, large = 1. The foramen caroticopharyngeale is a small opening in the pterygoid through which the arteria carotico-pharyngeale exits the skull. In most turtles in is very small [i.e., Chelydra (Gaffney 1979, Fig. 9)] or not identifiable (Proganochelys). Those forms that have an enlarged foramen caroticopharyngeale [as in Adocus (Meylan and Gaffney 1989, Fig. 5, labeled as “foramen basisphenoidale”) and Clemmys (Gaffney 1979, Fig. 47)] can be considered to have a derived condition.
132. Carapacial contacts of inguinal buttress. Not reaching costals = 0, reaching costals = 1. In primitive turtles including Proganochelys the plastral buttresses do not reach the overlying costal bones of the carapace. When they do, there are two sets of contacts visible for the inguinal buttress. It may contact only the fifth costal (as in Podocnemis or Deirochelys) or it may contact both the fifth and sixth costals (as in Hardella). These are derived conditions.
133. Musk duct absent from axillary region. Absent = 0, present = 1. Musk ducts absent in Progaonchelys but are present in a variety of turtles and in many forms are visible in the shell as foramina (Podocnemis) or grooves (kinosternids) in the vicinity of the bridge. Since there are forms with only axillary or only inguinal musk ducts present, the occurrence of musk ducts is treated as two characters. This character will be problematic for fossils of those clades in which the living members have musk ducts that do not leave any trace in the skeleton (i.e., Trionychidae). Presence of axillary musk ducts can be considered derived.
134. Musk duct absent from inguinal region. See character 133. Absent = 0, present = 1. Presence of inguinal musk ducts can be considered derived.
135. Hinge at hyo-hypoplastral suture. Absent = 0, present = 1. Primitively the hyo- and hypoplastra are sutured along their contact. Forms in which this contact is kinetic (as in Terrapene) can be considered to have a derived condition.
136. Pectoral scales (set 4) reach entoplastron. No = 0, yes = 1. Primitively the turtle plastron is covered by seven sets of scales (Hutchison and Bramble 1981) as is the case in Proganochelys (Gaffney 1990, Fig. 92). The fourth pair, the pectorals, in Proganochelys and most turtles is located well posterior to the entoplastron. Those forms in which it reaches the entoplastron (as in Emys and Podocnemis) can be considered to have a derived condition.
137. Last pair of marginal scutes contacts suprapygal. No = 0, yes = 1. In most turtles the last pair of marginal scales remain unfused and in contact with only the peripheral elements. Those forms that have the last marginals (either paired or fused) that contact the suprapygal (as in Hardella) can be considered to have a derived condition.
138. Quadrate-basioccipital contact. No = 0, yes = 1. In Proganochelys and other primitive turtles, the quadrate is located well lateral to the basicranium, lacks a medial process, and is not in contact with the basioccipital. Those forms with a large medial process of the quadrate that contacts the basioccipital (as in Podocnemis) can be considered to have a derived condition (See also Gaffney et al. 2006, p. 600).
139. Jugal contributes to triturating surface of upper jaw. No = 0, yes = 1. In most turtles including Proganochelys the jugal does not appear on the triturating surface. Those forms in which it makes a significant contribution to the triturating surface (as in Bothremys or Sandownia) can be considered to have a derived condition.
140. Antrum postoticum. Absent = 0, small = 1, large = 2. The antrum postoticum is absent from Proganochelys because the incisura columellae auris is widely open. The presence of a small antrum postoticum (as in Podocnemis expansa or Chelonia) is considered derived and a moderate to large antrum (as in Pelusios or Gopherus) is considered further derived. (See also Gaffney et al. 2006, p. 592).
141. Quadrate contacts basisphenoid but not basioccipital. No = 0, yes = 1. In Proganochelys and other primitive turtles, the quadrate is located well lateral to the basicranium, lacks a medial process, and is not in contact with the basisphenoid or basioccipital. Those forms with a large medial process of the quadrate that contacts the basisphenoid but not the basioccipital (as in Pelusios) can be considered to have a derived condition that differs from that described in character 138 above.
142. Short postorbital exposed by temporal emargination. No = 0, yes = 1. Primitively the skull roof of turtles is not emarginate and the postorbital is not exposed by emargination. Those forms in which a small postorbital is exposed by temporal emargination (as in Pelusios or Trachemys) can be considered to have a derived condition.
143. Prefrontal-palatine contact. Present = 0, absent = 1. In Proganochelys the prefrontal apparently contacts the palatine medial to the foramen orbito-nasale (Gaffney 1990, Fig. 42b) as is the case in most cryptodires. Those forms in which this contact is absent can be considered to have a derived condition.
144. Jugal exposed on temporal margin. No = 0, yes = 1. Primitively the jugal is not exposed by temporal emargination, those forms in which it is exposed can be considered to have a derived condition.
145. Foramen praepalatinum. Present = 0, absent = 1, foramen intermaxillaris = 2. The presence of the F. praepalatinum in Proganochelys remains uncertain, however, as they are common to most Casichelydia their presence can be considered primitive. Two derived conditions are recognized. One in which the premaxilla is well-ossified and there is no foramen intermaxillaris and one in which a foramen intermaxillaris is well developed.
146. Crista supraoccipitalis projecting well posterior to occipital condyle. No = 0, yes = 1. In Proganochelys, the supraoccipital is very short and there is no posterior projection. In casichelyidians (Chelus, Pelusios) the supraoccipital is very short and does not extend well posterior to the occipital condyle. In others (Apalone, Chelydra, Chelonia), it extends well posterior to the occipital condyle.
147. Maxillae meet on the midline. No = 0, yes = 1. In Proganochelys and most turtles the premaxillae and vomers separate the maxilla on the midline of the palate. Where the maxillae do meet on them midline as in Caretta or Phrynops, this can be considered a derived condition.
148. Basisphenoid with reduced ventral exposure. No = 0, yes = 1. Primitively turtles have a broad and long basisphenoid. Those forms in which this exposure is much reduced or absent can be considered to have a derived condition.
149. L-shaped prefrontal as seen from anterior. No = 0, yes = 1. Absent primitive, present derived. The prefrontal normally forms the anteriomedial margin of the orbit. Those forms in which it has an L-shape and contributes to the ventromedial margin of the orbit are considered to have a derived condition.
Appendix 13.2
Data set used to estimate phylogenetic positions of Brachyopsemys and Sandownidae.
Proganochelys: 00000 00000 00000 00000 00?00 00000 00000 00000 00?00 00000 00000 0000? 00000 00000 ?0000 00000 00000 00?00 00000 00000 00000 ?0000 00000 00000 00000 0??00 00000 0?000 0000? 0000
Elseya: 11111 00001 10010 01001 11?10 11111 11101 11110 00400 11110 00001 0000? 00000 00000 ?0000 01000 00000 00000 00000 01000 11000 00010 011?1 00000 ?00?0 00011 01110 1000? 01001 0100
Phrynops: 11111 00001 10010 01001 11?10 11111 11101 11110 00400 11111 11001 0001? 00000 00000 ?0000 01000 00000 00000 00000 01000 11000 00010 011?1 00000 000?0 00010 01??0 ??00? 01101 0100
Chelus: 11111 00001 10010 01101 11?10 11111 11101 11110 00400 11111 11111 0001? 00000 00000 ?0000 01000 00000 00000 00000 01000 11000 00010 011?1 00000 ?00?0 00011 03??0 1000? 01000 0000
Chelodina: 11111 00001 10011 01001 11?10 11111 11101 11110 00600 11111 11111 0000? 00000 00000 ?0001 10000 ?0000 01000 00000 00000 00000 01000 11000 00010 111?1 00000 ?00?0 00010 01000 1000? 11101 0000
Podocnemis: 11?11 00001 10011 01111 11?10 11111 01101 11111 11100 00000 00001 0001? 00000 00000 ?0000 01000 00000 00000 00000 01000 10000 00010 01101 00000 100?0 00011 01110 10101 00101 1001
Pelusios: 11?11 00001 10011 01111 11?10 11110 01101 11111 11211 00000 00001 0001? 00000 00000 ?0000 01000 00000 00000 00000 01000 10000 00010 11101 00000 ?00?0 00011 01010 00002 11100 0000
Pelomedusa: 11?11 00001 10011 01111 11?10 11111 01101 11111 11311 00000 00001 0001? 00000 00000 ?0000 01000 00000 00000 00000 01000 10000 00010 11101 00000 ?00?0 00011 02110 00000 11100 0000
Platysternon: 11111 11111 11101 10110 00?11 00111 10111 00101 00600 00000 00000 11111 00000 00000 00000 01000 00000 00000 00000 00100 01100 01100 11101 00000 00000 00011 00??0 ??00? 00000 1010
Chelydra: 11111 11111 11101 10110 00?11 00111 10111 00101 00500 00000 00000 11111 00000 00010 00000 01000 00000 00000 00000 00000 00100 01100 11101 00010 00000 00010 00??0 ??002 00000 1000
Chelonia: 11111 11111 11100 10110 00?11 00111 10111 11101 00700 00000 00000 0010? 11110 00000 00000 00000 10000 01000 00100 01101 01100 00000 11011 01011 00??0 ??001 00001 1020
Dermochleys: 11111 11111 11100 10110 00?11 00111 10111 11101 00?00 00000 00000 0010? 11111 10000 ?0000 00000 0?000 000?? 100?? 0000? 00100 01101 01100 ?0?01 11011 0?010 00??0 ??000 00101 0000
Dermatemys: 11111 11111 11101 10110 00?11 01111 10111 11101 00700 00000 00000 00000 00000 01111 10100 01000 00000 00011 11000 01000 00110 01101 11101 00000 00000 00010 00??0 ?0000 01000 1000
Kinosternon: 11111 11111 11101 10110 00?11 01111 10111 11101 00700 00000 00000 00000 00000 01111 10000 01000 00000 00011 11111 10111 00110 01101 11101 00010 00000 00011 001?1 ?0000 01101 1000
Staurotypus: 11111 11111 11101 10110 00?11 01111 10111 11101 00600 00000 00000 10000 00000 01111 10000 01000 00000 00011 11111 10111 00110 01101 11101 00010 00000 00011 001?1 ?0000 01011 1000
Carettochelys: 11111 11111 11101 10110 00?11 01111 10111 11101 10900 00000 00000 0011? 02000 01111 ?1011 10111 1?000 000?? 000?1 01100 ?11?0 01101 11101 00000 100?0 00010 00??1 ??001 01002 1000
Lissemys: 11111 11111 11101 10110 00?11 01111 10111 11101 10800 00000 00000 00110 00000 01110 11011 10111 1?111 111?? 000?0 01000 ?01?0 01101 11101 00100 101?0 0?010 00??1 ??000 01112 1100
Apalone 11111 11111 11101 10110 00?11 01111 10111 11101 10800 00000 00100 00110 00000 01110 11011 10111 1?111 111?? 000?0 01000 ?01?0 01101 11101 11210 101?0 0?010 00??1 ??000 01112 1000
Clemmys 11111 11111 11101 10110 00?11 01111 10111 11101 00400 00000 00002 00000 00000 00000 00000 01000 00000 00010 00000 01000 11000 01101 11111 00000 00000 00010 12101 100?? 01000 1000
Graptemys 11111 11111 11101 10110 00?11 01111 10111 11101 00400 00000 00002 0000? 00000 00000 ?0000 01000 00000 00010 00000 01000 11101 11101 11111 00000 00000 00011 02000 000?? 01001 1000
Trachemys 11111 11111 11101 10110 00?11 01111 10111 11101 00400 00000 00002 00000 00000 00000 00000 01000 00000 00010 00000 01000 11001 11101 11111 00000 00000 00011 02000 000?? 01000 1000
Heosemys 11111 11111 11101 10110 00?11 01111 10111 11102 00500 00000 00001 0000? 00000 00000 ?0000 01000 00000 00010 00000 01000 11100 01101 11111 00000 00000 00010 ?1110 110?? 01??? ????
Chinemys 11111 11111 11101 10110 00?11 01111 10111 11102 00500 00000 00001 0000? 00000 00000 ?0000 01000 00000 00010 00000 01000 11100 01101 11111 00000 00000 00011 01110 1100? 0100? 1000
Geochelone 11111 11111 11101 10110 00?11 01111 10111 11102 00500 00000 00001 00010 00000 00000 00000 00000 00000 00010 00000 01000 11100 01101 11111 00001 01010 11120 01000 110?? 01000 1000
Adocus 11111 11111 11101 10110 00?11 01111 1011? ?1101 00?00 00000 00000 0000? 00000 0?101 ?1110 0100? 01000 00000 00000 00000 00100 ?1101 11111 00000 100?0 00011 10??0 11000 01000 1000
Emarginachelys 11111 11?11 11101 1?110 0??1? ?1111 1011? ??1?? ?0?00 0000? 00000 ??00? 00000 0?111 ?0100 0100? 00000 00?10 ?0000 0100? 00110 ?110? 11111 00000 00000 01000 00??0 ?0000 00000 1000
Meiolania 11111 11111 11100 10000 01?10 ?0111 10100 00000 00?00 00000 00000 ?000? 0?000 000?0 ?0000 0100? 00000 00?0? 00000 00000 ?0100 00000 0111? 00000 00000 00020 00??0 ??0?? 00000 0010
Sinemys 11?11 11111 11101 1?000 0??1? ?1111 10110 000?? 1??01 0??01 00000 ?111? ????0 00??? ?100? ?10?? ?0000 0???? 00000 ?10?? 0110? ?1000 111?? ????0 ?0?00 000?0 ?0110 000?? 0110? 0?00
Peltochelys ????? ????? ????? ????? ????? ??111 1???? ????? ????? ????? ????? ??0?? ?2??? ????? ????? ??0?? ?1000 ???00 ??001 ????? 00?0? ????? ??1?? 0000? ????? 0?0?? ???0? ????? ????? ????
Mongolemys 11111 11?11 11101 11110 0??1? ??111 101?? ????0 0???? 00?00 ?0001 0?00? 00??? 000?? ??000 0100? 00000 ?0?10 ?0000 ??0?0 0?10? 0??0? 111?? ????? ????? 0001? 11110 00001 01000 ?000\
Sandownia 11111 1?111 11100 11110 0??11 00??? ????? ????0 ???00 00?00 00?0? ???1? ??001 001?? ??001 00?0? 0???? ?0??? 10??? ?10?? ??1?? 0??0? 0???? ????0 ????? ????1 1???? ??01? 00001 0011
A. maortuensis ????? ????? ????? ????? ????? ??111 10111 ??1?? 1???? ????? ????0 0?1?? 0???? ????? ?1??? ??1?? ??111 ???1? ??010 ????? ????? ????? ??1?? 1121? 1?1?? 0??0? ?0??0 ????? ????? ????
Santanachelys 11111 1?111 11100 11110 0??11 00??? ????? ????1 ???00 00?00 00?0? ???0? 1?111 1?0?? ??001 01?0? 0???? ?0??? 1???? ?10?? ??1?? 0??0? 0???? 00001 11111 0?000 001?? ??0?? 00?0? 1?00
Hadrianus ????? ????? ????? ????? ????? ??111 10??? ????? 0???? ????? ????0 ??0?? ????? ????? ????? ??0?? ?0000 ???10 ??00? ???0? 2??0? ????? ?11?1 0000? ?10?? 1102? ?0??0 00??? ????? ????
Ptychogaster ?1111 11?11 1110? ????0 0???? ??111 10??? ????? 0???? ????? ????1 ??0?? ????? ????? ??0?? ?100? ?0000 ???10 ??00? ????0 2?00? ????? ?11?? 0000? ????? 000?? ???11 100?? 0???? ????
Baltemys ????? ????? ????? ????? ????? ??111 10??? ????? ????? ????? ????0 0?0?? ?0??? ????? ????? ??0?? ?0000 ???11 ??121 ????1 2??1? ????? ?11?? 0001? ????? 000?? ?01?0 00??? ????? ????
Hoplochelys ????? ????? ????? ????? ????? ??111 10??? ????? ????? ????? ????0 0?0?? ?0??? ????? ????? ??0?? ?0000 ???10 ??110 ????0 0??1? ????? ?11?? 0001? ????? 000?? ?01?0 10??? ????? ????
Araripemys 11111 00?01 10011 01111 0??10 ?0111 11101 10111 11?00 00000 00100 0010? 00000 00000 ?0000 01010 00001 00?10 00000 0100? 10000 00010 111?0 00000 ?0010 0?0?0 00??0 000?0 0011? ?000
P. rusingae ????? ????? ????? ????? ????? ??110 0???? ????? 11??? ????? ????? ??0?? ????? ????? ????? ??0?? ?0000 ????? ??0?0 ????? 1???? ????? ????? ?0??? ????? ??0?? ????0 ????? ????? ????
Cearachelys 11111 00001 1001? ?1111 11?10 ?1111 01101 ??1?? 1??10 00000 00000 0?01? 00110 000?? ??000 0100? 00000 ???00 00000 ?01?? 10000 00?10 111?? 00000 ????? 000?1 0?110 001?1 0010? 0000
Brachyopsemys 11111 11111 11100 10110 0??1? 00??? ????? ????0 ???00 00?00 00?0? ???0? ??000 001?? ??000 00?0? 1???? ?0??? 00??? ?10?? ??1?? 0??0? 0???? ????0 ????? ????1 0???? ??011 00101 1111
Erquilinnesia 11111 11111 11100 1?110 0??1? 00111 10??? ????? 0??00 00?00 0??00 0?10? ??1?1 1?0?0 ??00? 0000? 10000 ?0??? 10000 ?1000 ??10? ????? 0110? 0?000 ??0?1 01011 00??0 ?0001 00?01 1020
Thalassemys 11111 11111 11100 00000 00?1? 00?11 1?000 ????? ???00 00?00 00?00 0??0? ?000? 0?0?? ?00?1 0100? 000?0 00??0 0000? ?10?? ??10? 0110? 0??01 00?0? ????? ?0??? 0???0 0?001 00100 1000
Solnhofia 11111 11111 11101 10010 00?1? 0?111 1?0?? ?01?0 10?00 00000 00?00 0??0? ??01? 000?? ?0000 0100? 00000 00??0 00000 ?10?? ?0100 01100 0??0? 00000 00??0 ?00?1 00110 0?001 00100 1000
Tasbacka 11111 11111 11100 10110 00?1? 00111 101?? ??1?? ???00 00?00 00?00 ??10? ??11? 100?? ??000 0100? 0?000 ?0??? 00??? ?10?? ??1?0 0??0? 0?11? 00000 ??01? ?10?0 00??0 ??001 00101 1020
Euclastes 11111 11111 11100 10110 00?1? 00??? ????? ????? ????? 00?00 00?0? ???0? ??11? 1?0?? ??000 01?0? 0???? ?0??? 0???? ?10?? ??1?? 0??0? 0???? 0???0 ????? ????0 0???? ??001 00101 1020
Angolachelys 11111 11111 11100 10?10 0??1? ?0??? ??1?? ????? ????? 00?00 00?0? ???1? ????? ?00?? ??0?1 00?0? 1???? ?0??? ?0??? ?10?? ??1?? ???0? 0???? ????0 ????? ????1 0???? ??010 00100 ??11
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Tong, H., Meylan, P. (2013). Morphology and Relationships of Brachyopsemys tingitana gen. et sp. nov. from the Early Paleocene of Morocco and Recognition of the New Eucryptodiran Turtle Family: Sandownidae. In: Brinkman, D., Holroyd, P., Gardner, J. (eds) Morphology and Evolution of Turtles. Vertebrate Paleobiology and Paleoanthropology. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-4309-0_13
Download citation
DOI: https://doi.org/10.1007/978-94-007-4309-0_13
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-4308-3
Online ISBN: 978-94-007-4309-0
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)