
Abstract
Circulating γδ T cells are cytotoxic lymphocytes that are unique to primates. Recent studies have shown that amino-bisphosphonates (nBP) activate γδ T cells to kill tumor cells in an indirect mechanism, which requires antigen presenting cells (APC). We hypothesized that selective targeting of nBP to monocytes would result in a more potent γδ T cells activation in circulation, and in tissue associated macrophages (TAM) following monocytes-laden drug extravasation and liposomes accumulation at the tumor site. In addition, inhibition of TAM by alendronate liposomes (ALN-L) is expected. ALN was targeted exclusively to monocytes, but not to lymphocytes, by encapsulating it in negatively-charged liposomes. The proportion of human γd-T cells in the CD3+ population following treatment with ALN-L or the free drug was increased, from 5.6 ± 0.4% to 50.9 ;± 12.2% and 49.5 ± 12.9%, respectively. ALN solution and liposomes treatments resulted in an increased, and in a dose dependent manner, TNFα secretion from h-PBMC. Preliminary results showed that ALN-L inhibited tumor growth in a nude mouse breast tumor model. It is suggested that enhanced activation of γδ T cells could be obtained due to interaction with circulating monocytes as well as by TAM endocytosing liposomal nBP leading to a potentiated anti-tumor effect of nBP. It should be noted that this could be validated only in primates/humans since γδ T cells are unique in these species.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
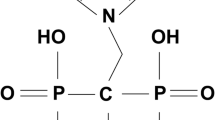
Keywords
16.1 Introduction
Human circulating γδ T cells lymphocytes constitute of 1–5% of peripheral blood T cells (Hayday 2000) and exhibit major histocompatibility complex (MHC) unrestricted cytotoxicity against a large number of tumor types (Ferrarini et al. 2002). Most of the circulating γδ T cells belong to the Vγ9Vδ2 subset that are unique to primates (Hinz et al. 1997). Recent studies have shown that amino-bisphosphonates (nBP), antiresorptive drugs utilized clinically in bone-related disorders (Rodan 1998a), activate Vγ9Vδ2 T cells (Kunzmann et al. 2000). There has thus been interest in using nBPs in cancer immunotherapy, with promising results against B-cell malignancies (Wilhelm et al. 2003) and hormone refractory prostate cancer (Dieli et al. 2007). And in a very recent clinical trial, it was shown that a nBP (zolendronate) exerts a significant anticancer benefit when added to hormone therapy, reducing the risk of breast cancer returning by 36% (Gnant et al. 2009). Tumor cells are killed in an indirect mechanism, which requires antigen presenting cells (APC) (Caccamo et al. 2008; Kunzmann et al. 2000). nBP are internalized to some extent by mononuclear cells such as monocytes and dendritic cells (DCs) leading to intracellular accumulation of the isopentenyl pyrophosphate (IPP) metabolite (Roelofs et al. 2009). Consequently, this endogenous phosphoantigen is ultimately recognized by γδ T cells with subsequent cell activation, proliferation, and the release of TNFα, IL6 and IFNγ (Mariani et al. 2005). However, the family of nBP is both highly hydrophilic and charged, and is rapidly eliminated from the circulation by binding to bone and via urine excretion (Rodan 1998a, b; Fleisch 1998). Therefore, peripheral monocytes and tumors are exposed only briefly and to a relatively low concentration of nBP. Thus, in order to potentiate the effect of nBP selective targeting of the drug to APCs is necessary.
Encapsulating a BP in a particulate delivery system, such as liposomes, deviates these bone-seeking molecules to circulating monocytes and macrophages of the mononuclear phagocytic system (MPS) (van Rooijen and van Kesteren-Hendrikx 2003; Danenberg et al. 2002). The anti-inflammatory effect resulting from macrophage depletion by clodronate liposomes (CLOD-L; a non nBP) has been documented in experimental arthritis (Richards et al. 2001), delayed graft rejection (Slegers et al. 2000), CNS inflammation (Zito et al. 2001), and tumor angiogenesis (Zeisberger et al. 2006), and restenosis (Danenberg et al. 2002, 2003a; Epstein-Barash et al. 2010). Previous studies in our group demonstrated the high efficacy of alendronate liposomes (ALN-L; a nBP) in inflammatory-related disorders such as restenosis (Danenberg et al. 2002, 2003b; Epstein et al. 2007, 2008; Epstein-Barash et al. 2010) and endometriosis (Haber et al.. 2009, 2010).
Macrophages populate the microenvironment of most if not all solid tumors, representing >50% of the tumor mass in certain breast cancers (Lewis and Pollard 2006). Monocytes are recruited into solid tumor stroma where they differentiate into tumor-associated macrophages (TAM). Depletion of TAM by long-circulating CLOD-L, exploiting the enhanced permeability of the tumor microcirculation, has been reported (Banciu et al. 2008). A more potent activation of γδ T cells could be achieved by a liposomal delivery system due to the preferential uptake by circulating monocytes (Monkkonen et al. 1994; Monkkonen and Heath 1993; Epstein et al. 2008). The indirect therapeutic effect of BP in tumor is mediated by two different mechanisms, stimulatory effect on γδ T cells and the inhibition of TAM. We hypothesized that a potent therapeutic effect could be achieved by a liposomal delivery system of nBP, mediated by the two mechanisms. Selective targeting of nBP to monocytes by liposomes would result in a more potent γδ T cells activation in circulation, and at the tumor tissue by TAM following both, monocytes-laden drug extravasation and passive liposomes accumulation at the tumor site. In addition, inhibition of TAM by liposomal ALN is expected. Liposomal ALN is expected to be more potent than liposomal CLOD since this delivery system is more potent in depleting monocytes and macrophages (Danenberg et al. 2003a; Epstein-Barash et al. 2010). It should be noted that since ALN-L depleting effect on circulating monocytes is noted 36–48 h following treatment (Afergan et al. 2008; Epstein et al. 2007, 2008; Epstein-Barash et al. 2010), activation of γδ T cells can be achieved during this period.
Herein, we examined the effect of monocytes targeted ALN-L on γδ T cells activation and on tumor growth. To the best of our knowledge there are no reports in the literature exploiting circulating monocytes for the activation of γδ T cells by liposomal nBPs. Furthermore, the inhibitory effect of ALN-L on tumor growth has not yet been studied. It should be noted that the indirect mechanism of tumor inhibition mediated by γδ T cells activation couldn’t be elucidated in the current study, because γδ T cells are unique to primates (Hinz et al. 1997).
16.2 Materials and Methods
16.2.1 Liposome Preparation
Liposomes were prepared by the modified thin-film hydration method (Danenberg et al. 2002; Epstein et al. 2008; Epstein-Barash et al. 2010) with 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC, Lipoid, Ludwigshafen, Germany), the negatively charged distearoyl-phosphatidylglycerol (DSPG, Lipoid), and cholesterol (Sigma-Aldrich, Israel). Liposomes were prepared at a molar ratio of 3:1:2. The lipids were dissolved in t-butanol and lyophilized overnight. The lyophilized cake was hydrated with an aqueous solution containing 200 mM ALN or CLOD at 55–60°C, and left to stand for 1 h. Blank liposomes were prepared by the same technique, with buffer instead of the drug. Fluorescent liposomes were prepared similarly with 0.025% w/w DSPE-rhodamine (Avanti Polar Lipids, Alabaster, AL) and 15.3% w/w dextran-FITC (Sigma-Aldrich, Israel), membrane and core markers, respectively (Afergan et al. 2008; Epstein-Barash et al. 2010). For small-sized liposomes, the ethanol injection method was used (Epstein et al. 2008; Epstein-Barash et al. 2010). The obtained liposomes were homogenized to a desired size by means of an extruder. To remove un-encapsulated drug, the liposomes were passed through a Sephadex G-50 column and were eluted with buffer.
16.2.2 Liposome Characterization
Liposome size and zeta potential were determined at room temperature after appropriate dilution with MES/HEPES buffer by photon correlation spectroscopy (ALV-GmBH, Langen, Germany) and NanoZ (Malvern Instruments, Malvern, UK), respectively. The phospholipid content was determined colorimetrically with the Bartlett assay (Bartlett 1959; Danenberg et al. 2002; Epstein et al. 2007), cholesterol content was measured by means of HPLC (Lang 1990), and ALN and CLOD content was determined by spectrophotometric assay of their complex with copper (II) ions at λ = 240 nm (Danenberg et al. 2003a; Epstein-Barash et al. 2010).
16.2.3 Ex Vivo Cell Culture
16.2.3.1 Human Peripheral Blood Mononuclear Cells (h-PBMCs)
h-PBMCs were isolated from human buffy coat using Ficoll-Hypaque (Amersham-Pharmacia, Uppsala, Sweden) density gradient centrifugation. h-PBMCs were cultured in RPMI media supplemented with 100 U/ml penicillin, 100 μg/ml glutamine, 10% fetal calf serum (Biological Industries, Beit Haemek, Israel), and 10 U/ml rhIL-2 (Chiron B.V., Amsterdam, The Netherlands).
16.2.3.2 Generation of Monocyte-Derived Immature DC (imDC)
ImDC were generated from monocytes, separated from adhered h-PBMC, by incubation in RPMI 1640 with 10% heat-inactivated human AB serum, supplemented with 1,000 U/ml IL-4 and 1,000 U/ml GM-CSF (R&D, Minneapolis, MN, USA) on days 1 and 4. ImDC were harvested on day five and phenotypically defined by FACS by the following monoclonal antibodies (mAbs): anti-CD14-PE (Dako, Glostrup, Denmark), anti-CD11C-FITC, anti-CD83-FITC and anti-HLA-DR-FITC (Beckman Coulter, Fullerton, CA, USA); and anti-86-PE (BD bioscience, San Jose, CA, USA). Isotype matched mAb were utilized as control (Beckman Coulter and BD bioscience).
16.2.4 Liposome Uptake by h-PBMC and imDC
Freshly isolated h-PBMCs and monocyte-derived imDC were incubated with fluorescent liposomes. Following 4 h of incubation, cells were harvested for assessment of liposomes cellular uptake by confocal microscopy (Zeiss LSM 410) and quantitative uptake by means of FACS (FACScan, Becton Dickinson, USA).
16.2.5 Quantification of TNFα Release
h-PBMCs were cultured at a density of 1 * 106 cells/ml in 96-well u-shaped plates and treated with ALN solution or liposomes for 48 h in the presence of 10 U/ml rhIL-2. The medium was removed after centrifugation of the plate at 1,500 g, and levels of TNFα were quantified with ELISA kit (Peprotech EC Ltd, London, UK) according to the manufacturer’s instructions.
16.2.6 Proliferation of γ d T Cells
h-PBMCs were cultured at a concentration of 1 * 106 cell/ml in 24/well plates. Cells were treated for 9 days with BP solution or liposomes with or without 0.5 μg/ml LPS (LPS, E. coli, serotype 0127:B8, Sigma, Saint Louis, Missouri, USA) in the presence of 10 U/ml rIL-2. On day 4 and 7 of the culture period, one half of the medium was replaced with fresh medium containing rIL-2 and treatments. On day 9 non-adherent cells were harvested and aliquots of 1 * 106 cells were washed with FACS buffer (1% w/v bovine serum albumin (BSA) and 0.02% w/v sodium azide in PBS pH 7.4) and stained with 10 μl anti-CD3-FITC and 5 μl anti-pan-γδ-TCR-PC5 Abs, or the respective isotype-matched controls (Beckman Coulter, Fullerton, CA, USA) for 30′ at 4°C in a final volume of 100 μl FACS buffer. Adherent cells were detached using 0.25% trypsin/EDTA solution (Biological Industries, Beit Haemek, Israel) and aliquots of 1 * 106 cells were washed with FACS buffer and stained with anti-CD14 Ab (Dako, Glostrup, Denmark) for 30′ at 4°C in a final volume of 100 μl FACS buffer. Dead cells exclusion was done by adding 1 μg/ml topro3 (Invitrogen, California, USA) to a sample tube of both non-stained adherent and non-adherent cells. Cells were analyzed using BD FACS DIVA LSR II System (Becton Dickinson, USA) and FCS express (Denovo software) was used for quantitative analysis.
16.2.7 Murine Breast Cancer Model
Animal care and procedures were in accordance with the standards for care and use of laboratory animals of The Hebrew University of Jerusalem. Animals were fed with standard laboratory chow and tap water ad libitum. Female athymic nude mice (Harlan Laboratories, Jerusalem, Israel), 6 weeks old, were anaesthetized by isoflurane (Minrad International, USA), and were inoculated subcutaneously in the right flank with human MDA-231 (ATCC, Rockville, MD, USA) breast cancer cells (1 * 106 cells in 200 μl PBS). The injected cancer cells were allowed to grow for 14 days into tumors in the host animals before treatment. The animals were randomly assigned to a treatment of 20 mg/kg free ALN, 20 mg/kg ALN-L or empty liposomes (mean size of 85 ± 20 nm), by 3 lateral tail vain injections, on day 14, 16 and 18 (n = 4/group). The maximum and minimum diameters of the tumors were measured using a sliding caliper on day 14, 16 and 18. The volume of the tumors was calculated using the formula: 0.5(short axis)2(long axis) (Geran et al. 1972).
16.2.8 Data Analysis
All data are expressed as the mean ± standard deviation. Comparisons among treatment groups were made by 2-way analysis of variance (ANOVA) followed by Tukey test, and unpaired two-tailed t-test when necessary. Differences were termed statistically significant at p < 0.05.
16.3 Results
16.3.1 Liposomes Characteristics
The physicochemical characteristics of ALN-L, used for γδ T cells activation and tumor growth inhibition, are summarized in Table 16.1. The liposomal formulations obtained were negatively charged and in two nano-sizes of 190 ± 24 and 85 ± 20 nm.
16.3.2 Internalization of Liposomes by h-PBMCs Ex Vivo
In order to validate that the liposomes target specifically phagocytic cells, h-PBMCs, were incubated with fluorescent liposomes. The fluorescent liposomes were double-labeled with DSPE-rhodamine (red, membrane marker) and dextran-FITC (green, hydrophilic core marker). Following incubation with h-PBMCs, which comprise of monocytes and lymphocytes, the cells were analyzed by means of FACS. Treatment of h-PBMCs with blank liposomes resulted in internalization by 83 ± 1.8% of the monocyte cell population, whereas only 6 ± 0.6% of the lymphocytes internalized the liposomes (Fig. 16.1b). In agreement with the FACS results, confocal microscopy showed specific internalization of liposomes into monocytes (Fig. 16.1c, lower panel), and no liposomes were traced in association with lymphocytes (Fig. 16.1c, upper panel). Co-localization of the lipophilic membrane marker with the hydrophilic core marker (Fig. 16.1c right column, orange color) indicated the uptake of intact liposomes.

Uptake of blank liposomes by h-PBMCs. Representative FACS images (a), calculated mean ± SD, N = 3 (b), and confocal images (c). h-PBMCs were incubated with fluorescent liposomes labeled with DSPE-rhodamine (membrane marker) and dextran-FITC (hydrophilic core marker). Following 4 h of incubation cells were harvested for assessment of liposomes internalization by confocal microscopy and FACS analysis (**p < 0.01)
16.3.3 ALN Loaded Liposomes Bioactivity Ex Vivo
16.3.3.1 Proliferation of γ d T Cells
In cultures of h-PBMCs, the proportion of γδ T cells in the CD3+ cell population was determined following various treatments in the presence of IL2 (Fig. 16.2). The control group, non-treated h-PBMCs, had 5.6 ± 0.4% γδ T cells in the CD3+cell population. Treatment with ALN in solution or encapsulated in liposomes (1 μM) significantly increased the proportion of γδ T cells, 50.9 ± 12.2% and 49.5 ± 12.9% of the CD3+cell population, respectively. Empty liposomes, CLOD (a non nBP) in solution and encapsulated in liposomes did not affect γδ T cells proliferation. The similar increase of the γδ T cell population following treatment with ALN as a free drug and in liposomes was associated with no changes in both cells’ viability (Fig. 16.2c) and CD3 expression (Fig. 16.2d).

The effect of bisphosphonates (clodronate, CLOD) and amino BP (alendronate, ALN) and liposomes (liposomal CLOD and ALN, CLOD-L and ALN-L, respectively) on γδ T cells proliferation. (a) FACS profiles of the T cells gated population. (b) Proportion of CD3+ γδ-T cells (%). h-PBMCs were double stained with anti-CD3 and anti γδ TCR Abs before FACS analysis of the T-cell–gated population. (c) h-PBMCs viability by TO-PRO-3 staining and (d) CD3+ counts. h-PBMCs were cultured for 9 days with 1 μM of various formulations in the presence of IL-2. Data shown is the mean ± SD of experiments with h-PBMCs from 4 to 7 independent donors (**p < 0.01)
16.3.3.2 TNFα Secretion
After verifying the selective targeting of liposomes to monocytes in h-PBMCs cultures, the potential of the delivered ALN to exert its bioactivity on γδ T cell was studied by examining TNFα activation. Treatment with ALN in solution or in liposomes resulted in an increased, dose-dependent, TNFα secretion from h-PBMCs (Fig. 16.3).

The effect of ALN on TNFα secretion as a surrogate for γδ T activation. (a) h-PBMCs were incubated for 48 h with ALN in solution (ALN) or in liposomes (ALN-L) in the presence of IL-2. Data are values of h-PBMCs from two independent donors (mean ± SD)
16.3.3.3 Proliferation of γ d T Cells in LPS-stimulated Monocytes
Proliferation of γδ-T cells was evaluated in the presence of LPS-induced activation of monocytes. Elevated proportions of γδ-T cells were found in LPS treated h-PBMCs with or without ALN solution, in comparison with to no LPS, 56.4 ± 4.5%, 15.2 ± 3.2% and 44.6 ± 12.2%, 6.1 ± 3.4% of the CD3+cell population, respectively. The stimulatory effect of ALN-L on γδ T cells proliferation was diminished when the h-PBMCs were co-treated with LPS, 52.5 ± 12% and 24.3 ± 4.8% of the CD3+cell population, respectively (Fig. 16.4).

The effect of alendronate (ALN) and liposomal Aln (ALN-L) on γδ T cells proliferation in the presence of LPS. The proportion of CD3+ γδ T cells was determined 9 days following h-PBMCs incubation with 1 μM ALN solution or liposomes in the presence of IL-2 with or without 0.5 μg/ml LPS. h-PBMCs were double stained with anti-CD3 and anti γδ TCR Abs before FACS analysis of the T-cell-gated population. Data shown is the mean ± SD of experiments with h-PBMCs from two independent donors
16.3.3.4 Interplay Between γ d T Cells and APCs
After assessing the bioactivity of ALN-L on γδ T cells, its effect on monocytes was further examined. Following treatment of h-PBMCs with ALN solution or liposomes, the proliferation of γδ T cells and the proportion of monocytes in the h-PBMCs culture were quantified (Fig. 16.5). In order to avoid monocytes depletion, a low concentration of ALN was used (<1 μM). Treatment with both ALN solution and liposomes resulted in elevated counts of γδ T cells in the CD3+cell population, and in a dose dependent manner (Fig. 16.5a). The proliferation of γδ T cell was inversely correlated with CD14+ expression in the adherent cells (Fig. 16.5b). It should be noted that lymphocytes and monocytes viability was similar in all treatments (data not shown).

The effect of alendronate (ALN) and liposomal Aln (ALN-L) on γδ T cells proliferation (a) and CD14+ expression (b). h-PBMCs were cultured for 9 days with 0.1 and 1 μM ALN solution or liposomes in the presence of IL-2. Non-adherent cells were double stained with anti-CD3 and anti γδ TCR Abs and adherent cells were stained with CD14 Ab before FACS analysis. Data shown is the mean ± SD of experiments with h-PBMCs from two independent donors
Down regulation of the CD14 receptor could suggest differentiation of monocytes to dendritic cells (DC) following γδ T cell stimulation. In order to validate that liposomes also target DC, human imDC were incubated with fluorescently labelled liposomes. Human imDC were phenotypically defined as CD14−, CD11c+, CD83−, CD86+ and HLA-DR+ (Fig. 16.6 left). Following 4 h incubation with fluorescent liposomes, 48 ± 3% of the imDC, gated as Cd11C+, were fluorescently stained (Fig. 16.6 right).

Human derived imDC phenotype characterization and uptake of liposomes. FACS images of imDC phenotypes (left) and quantitative uptake of fluorescently labeled liposomes (right). imDC were generated from CD14+ h-monocytes treated with GMCSF and IL4 for 5 days. imDC were incubated with fluorescent liposomes labeled with DSPE-rhodamine (membrane marker). Following 4 h of incubation cells were harvested for assessment of liposomes cellular uptake of CD11C gated cells (mean ± SD)
16.3.4 Effect of ALN-L on Tumor Growth In Vivo
The effect of ALN-L on tumor growth was studied in vivo (Fig. 16.7). Athymic nude mice were inoculated with human MDA-231 breast cancer cells, and blank liposomes, ALN and ALN-L were IV administered at day 14, 16 and 18. Animals treated with ALN solution exhibited necrosis at the injection area, and the animals were euthanized before the end of the experiment. This phenomenon was not observed in liposome treated animals, either empty or ALN loaded. Tumor growth inhibition was observed following treatment with ALN-L, but the results did not reach statistical significance due to the small number of animals.

The anti-tumor effect of liposomal alendronate (ALN-L) in athymic mouse model of human breast cancer. Shown is the tumor volume following treatment with ALN-L (20 mg/kg) and control (blank liposomes) by 3 IV injections (on day 14, 16, and 18) 14 days post tumor cells inoculation. Results are presented as the mean ± SD (n = 4)
16.4 Discussion
We demonstrate here that selective targeting of ALN to monocytes by conventional liposomes resulted in γδ T cells proliferation ex vivo. Furthermore, we show preliminary results of tumor growth inhibition in vivo by ALN-L in a mechanism not involving γδ T cells activation.
Activated γδ T cells display distinct natural killer functions and directly eliminate transformed cells, a feature that is successfully being exploited in immunotherapy trails in cancer patients (Dieli et al. 2007; Wilhelm et al. 2003). Intravenous stimulation by nBP of γδ T cells in patients for cancer immunotherapy is thought to involve accumulation of IPP in APCs (Eberl et al. 2009; Miyagawa et al. 2001; Roelofs et al. 2009). The full clinical impact of these drugs efficacy is impeded due to rapid urine elimination and bone accumulation (Rodan 1998a; Fleisch 1998). Moreover, due to their high charge and hydrophilicity, free BP do not easily cross cell membranes. We hypothesized that selective targeting of nBP to monocytes would result in a more potent γδ T cells activation. We utilized liposomes to deliver encapsulated ALN to APCs for optimizing ALN efficacy due to increased uptake. Since ALN-L depleting effect on circulating monocytes is noted 36–48 h following treatment (Afergan et al. 2008; Epstein et al. 2007, 2008; Epstein-Barash et al. 2010), activation of γδ T cells can be achieved during this period. In this study we have used ‘conventional’ liposomes endowed with preferable physicochemical properties for monocyte-targeting; negatively charged membrane, that enhances the internalization of liposomes into phagocytic cells (Epstein-Barash et al. 2010; Patel 1992; Patel and Moghimi 1998), and in a mean diameter size of <200 nm, for maximizing both efficacy and safety (Allen and Hansen 1991; Epstein-Barash et al. 2010; Rodrigueza et al. 1993; Torchilin 2005).
Since γδ T cells are unique to primates (Hinz et al. 1997) our study on the effect of liposomal formulations was limited to ex vivo experiments. We first assessed the selective targeting of conventional liposomes to monocytes in ex vivo cultures of h-PBMCs, which comprise of monocytes and lymphocytes. It should be noted that the specific uptake by DC in the PBMC culture was not determined. Liposomes were selectively internalized intact by 83 ± 1.8% of h-monocytes, whereas only 6 ± 0.6% of the lymphocytes internalized liposomes following 4 h of incubation (Fig. 16.1). The scant uptake of liposomes by lymphocytes is attributed to B cells, and not T cells, as previously demonstrated for nanoparticles (Sela et al. 2010). The intracellular accumulation of IPP in monocytes, caused by nBP inhibition of FPP synthase of the mevalonate pathway, subsequently activates cytotoxic γδ T cells proliferation and secretion of pro-inflammatory cytokines (Hewitt et al. 2005). As expected from a non nBP (Kunzmann et al. 2000), treatment with CLOD-L or the free drug had no effect on γδ T cells proliferation (Fig. 16.2b). The similar stimulatory effect of liposomal and free ALN on γδ T cells proliferation (Fig. 16.2b) and TNFa secretion (Fig. 16.3) is most likely due to a similar uptake of free and ALN-L by monocytes in culture. The significantly increased cellular uptake, in tissue cultures, of particulated dosage forms of BPs in comparison to free BPs is not observed when the free BP concentration is low (<1 μM) (Monkkonen et al. 1994). This is because the cellular uptake of free BPs by monocytes/macrophages in vitro is mediated probably by a calcium complex formed in the culture media (Monkkonen and Heath 1993; Monkkonen et al. 1994). At low BPs concentration the calcium to BP ratio is high, enabling enhanced complexation of free ALN and internalization.
Human γδ T cells recognition of LPS is predominately presented by CD1 on APCs resulting in increased γδ T cells proliferation (Hava et al. 2005; Cui et al. 2009). ALN in liposomes and as a free drug increases the secretion of inflammatory cytokine by LPS-stimulated monocytes (Epstein-Barash et al. 2010; Makkonen et al. 1999). However, treatment by LPS reduced the stimulatory effect of ALN-L (Fig. 16.4), which is in contrast to the expected synergistic stimulation of γδ T cells by LPS and ALN co-treatment. The molecule through which IPP is presented to the γδ T cell receptor is still unknown (Clezardin 2010). It could be suggested that the presentation of LPS and phospholipids on monocytes hindered the presentation of IPP resulting in the reduced stimulatory effect of γδ T cells proliferation.
Down regulation of CD14 in monocytes (adherent cells), by both ALN as a free drug and in liposomes, correlated conversely with the proliferation of γδ T cells (Fig. 16.5), suggesting that activated γδ T cells may have induced this phenotypic change. This notion is supported by the pronounced reciprocal effect of γδ T cells stimulation on APCs (Devilder et al. 2006). Activated γδ T cells secrete cytokines such as TNF-α, which in turn down regulate CD14 expression, and cause differentiation of monocytes into DC (Eberl et al. 2009). Recently, it has been shown that amplified activation of γδ T cells can be achieved by another nBP (zolendronate) treated DC (Cabillic et al. 2010). The liposomes ability to target human imDC (Fig. 16.6 right) suggests that a positive feedback could be obtained for the amplification of γδ T cells. ALN-L will activate γδ T via monocytes, which in turn will stimulate monocytes differentiation into DC, an the latter will engulf liposomes further activating γδ T cells (Takahara et al. 2008).
We observed an anti-tumor effect of liposomal ALN in mice (Fig. 16.7), with no overt infection or side effects. A growing body of evidence suggests that the anti-tumor effect of nBP is mediated by both direct and in-direct mechanisms (Caraglia et al. 2010; Clezardin 2010; Clezardin and Massaia 2010). Since activation of γδ T cell is excluded in mice, the anti-tumor effect in our study should be attributed to passive tumor targeting as has been demonstrated for <200 nm liposomes in a murine bearing human breast cancer (Mayer et al. 1990). A direct cytotoxic effect on tumor cells is also unlikely since no significant accumulation of non-targeted liposomal formulations is expected in tumor cells (Kirpotin et al. 2006; Shmeeda et al. 2010). Thus, it is reasonable to assume that the tumor growth inhibition by ALN-L was mediated by TAM depletion, and possibly the induced differentiation of M2 into M1 (Veltman et al. 2010). The depletion of TAM and blood monocytes by liposomal formulations of CLOD and tumor growth inhibition has been reported (Banciu et al. 2008; Hiraoka et al. 2008; Zeisberger et al. 2006). ALN-L may possess a stronger anti-tumor effect than CLOD-L since it is more potent in depleting monocyte/macrophages (Danenberg et al. 2003a; Epstein-Barash et al. 2010), and at the same time, can stimulate γδ T cells activation since it is an nBP.
In attempts to increase the circulation time of nBP, Zhang et al. synthesized lipophilic pyridinium BP, which exhibit a pronounced γδ T cells stimulatory effect following incubation with h-PBMCs ex vivo (Zhang et al. 2010). However, systemic side effects were noted following the activation of γδ T cells (Adami et al. 1987; Hewitt et al. 2005), and the uptake of these compounds by APCs in vivo has not been determined. Recently, Shmeeda et al. reported that folate-targeted liposomes containing zolendronate has a potent in vitro cytotoxic activity on tumor cells (Shmeeda et al. 2010). In another study it was found that zolendronate activates γδ T cells to lyse tumor cells in cell culture (Li et al. 2009). Thus, it can be assumed that treatment with a combined formulation of targeted and conventional liposomes encapsulating nBP may result in a synergistic anti-tumor effect mediated by activation of γδ T cells as well as by depletion of tumor cells and monocytes/TAM. In light of our results that ALN-L activate γδ T cells ex vivo, we can envisage the following picture in vivo: Enhanced activation of γδ T cells would be obtained due to interaction with circulating monocytes as well as by TAM endocytosing liposomal nBP, leading to a potentiated anti-tumor effect of nBP. It should be noted that this scenario is speculative since γδ T cells are unique to primates (Hinz et al. 1997). The potential anti-tumor activity of ALN-L by activating cytotoxic γδ T cells and the resultant inhibition of tumor growth, presented in this study, could be assessed by investigations in primates correlating depletion of TAM and γδ T cells activation.
Abbreviations
- DSPC:
-
1,2-Distearoyl-sn-glycero-3-phosphocholine
- ALN-L:
-
Alendronate liposomes
- nBP:
-
Amino-bisphosphonate
- APC:
-
Antigen presenting cells
- BSA:
-
Bovine serum albumin
- CLOD-L:
-
Clodronate liposomes
- DCs:
-
Dendritic cells
- DSPG:
-
Distearoyl-phosphatidylglycerol
- MHC:
-
Histocompatibility complex
- h-PBMCs:
-
Human peripheral blood mononuclear cells
- imDC:
-
Immature DC
- IPP:
-
Isopentenyl pyrophosphate
- mAbs:
-
Monoclonal antibodies
- MPS:
-
Mononuclear phagocytic system
- TAM:
-
Tissue associated macrophages
References
Adami, S., Bhalla, A. K., Dorizzi, R., Montesanti, F., Rosini, S., Salvagno, G., & Locascio, V. (1987). The acute-phase response after bisphosphonate administration. Calcified Tissue International, 41, 326–331.
Afergan, E., Epstein, H., Dahan, R., Koroukhov, N., Rohekar, K., Danenberg, H. D., & Golomb, G. (2008). Delivery of serotonin to the brain by monocytes following phagocytosis of liposomes. Journal of Controlled Release, 132, 84–90.
Allen, T. M., & Hansen, C. (1991). Pharmacokinetics of stealth versus conventional liposomes: Effect of dose. Biochimica et Biophysica Acta, 1068, 133–141.
Banciu, M., Schiffelers, R. M., & Storm, G. (2008). Investigation into the role of tumor-associated macrophages in the antitumor activity of Doxil. Pharmaceutical Research, 25, 1948–1955.
Bartlett, G. R. (1959). Phosphorus assay in column chromatography. The Journal of Biological Chemistry, 234, 466–468.
Cabillic, F., Toutirais, O., Lavoue, V., de la Pintiere, C. T., Daniel, P., Rioux-Leclerc, N., Turlin, B., Monkkonen, H., Monkkonen, J., Boudjema, K., Catros, V., & Bouet-Toussaint, F. (2010). Aminobisphosphonate-pretreated dendritic cells trigger successful V gamma 9 V delta 2 T cell amplification for immunotherapy in advanced cancer patients. Cancer Immunology, Immunotherapy, 59, 1611–1619.
Caccamo, N., Meraviglia, S., Cicero, G., Gulotta, G., Moschella, F., Cordova, A., Gulotta, E., Salerno, A., & Dieli, F. (2008). Aminobisphosphonates as new weapons for gamma delta T cell-based immunotherapy of cancer. Current Medicinal Chemistry, 15, 1147–1153.
Caraglia, M., Marra, M., Naviglio, S., Botti, G., Addeo, R., & Abbruzzese, A. (2010). Zoledronic acid: An unending tale for an antiresorptive agent. Expert Opinion on Pharmacotherapy, 11, 141–154.
Clezardin, P. (2010). Bisphosphonates’ antitumor activity: An unravelled side of a multifaceted drug class. Bone, 48, 71–79.
Clezardin, P., & Massaia, M. (2010). Nitrogen-containing bisphosphonates and cancer immunotherapy. Current Pharmaceutical Design, 16, 3007–3014.
Cui, Y. C., Kang, L., Cui, L. X., & He, W. (2009). Human gamma delta T cell recognition of lipid A is predominately presented by CD1b or CD1c on dendritic cells. Biology Direct, 4, 47.
Danenberg, H. D., Fishbein, I., Gao, J., Monkkonen, J., Reich, R., Gati, I., Moerman, E., & Golomb, G. (2002). Macrophage depletion by clodronate-containing liposomes reduces neointimal formation after balloon injury in rats and rabbits. Circulation, 106, 599–605.
Danenberg, H. D., Fishbein, I., Epstein, H., Waltenberger, J., Moerman, E., Monkkonen, J., Gao, J., Gathi, I., Reichi, R., & Golomb, G. (2003a). Systemic depletion of macrophages by liposomal bisphosphonates reduces neointimal formation following balloon-injury in the rat carotid artery. Journal of Cardiovascular Pharmacology, 42, 671–679.
Danenberg, H. D., Golomb, G., Groothuis, A., Gao, J., Epstein, H., Swaminathan, R. V., Seifert, P., & Edelman, E. R. (2003b). Liposomal alendronate inhibits systemic innate immunity and reduces in-stent neointimal hyperplasia in rabbits. Circulation, 108, 2798–2804.
Devilder, M. C., Maillet, S., Bouyge-Moreau, I., Donnadieu, E., Bonneville, M., & Scotet, E. (2006). Potentiation of antigen-stimulated V gamma 9 V delta 2 T cell cytokine production by immature dendritic cells (DC) and recpirocal effect on DC maturation. The Journal of Immunology, 176, 1386–1393.
Dieli, F., Vermijlen, D., Fulfaro, F., Caccamo, N., Meraviglia, S., Cicero, G., Roberts, A., Buccheri, S., D’Asaro, M., Gebbia, N., Salerno, A., Eberl, M., & Hayday, A. C. (2007). Targeting human gamma delta T cells with zoledronate and interleukin-2 for immunotherapy of hormone-refractory prostate cancer. Cancer Research, 67, 7450–7457.
Eberl, M., Roberts, G. W., Meuter, S., Williams, J. D., Topley, N., & Moser, B. (2009). A rapid crosstalk of human gammadelta T cells and monocytes drives the acute inflammation in bacterial infections. PLoS Pathogens, 5, e1000308.
Epstein, H., Berger, V., Levi, I., Eisenberg, G., Koroukhov, N., Gao, J., & Golomb, G. (2007). Nanosuspensions of alendronate with gallium or gadolinium attenuate neointimal hyperplasia in rats. Journal of Controlled Release, 117, 322–332.
Epstein, H., Gutman, D., Cohen-Sela, E., Haber, E., Elmalak, O., Koroukhov, N., Danenberg, H., & Golomb, G. (2008). Preparation of alendronate liposomes for enhanced stability and bioactivity: In vitro and in vivo characterization. AAPSJ, 10, 505–515.
Epstein-Barash, H., Gutman, D., Markovsky, E., Mishan-Eisenberg, G., Koroukhov, N., Szebeni, J., & Golomb, G. (2010). Physicochemical parameters affecting liposomal bisphosphonates bioactivity for restenosis therapy: Internalization, cell inhibition, activation of cytokines and complement, and mechanism of cell death. Journal of Controlled Release, 146, 182–195.
Ferrarini, M., Ferrero, E., Dagna, L., Poggi, A., & Zocchi, M. R. (2002). Human gamma delta T cells: A nonredundant system in the immune-surveillance against cancer. Trends in Immunology, 23, 14–18.
Fleisch, H. (1998). Bisphosphonates: Mechanisms of action. Endocrine Reviews, 19, 80–100.
Geran, R. I., Schumach, A. M., Abbott, B. J., Greenber, N. H., & Macdonal, M. M. (1972). Protocols for screening chemical agents and natural-products against animal tumors and other biological-systems. Cancer Chemotherapy Reports. Part 3, 3, 1–103.
Gnant, M., Mlineritsch, B., Schippinger, W., Luschin-Ebengreuth, G., Postlberger, S., Menzel, C., Jakesz, R., Seifert, M., Hubalek, M., Bjelic-Radisic, V., Samonigg, H., Tausch, C., Eidtmann, H., Steger, G., Kwasny, W., Dubsky, P., Fridrik, M., Fitzal, F., Stierer, M., Rucklinger, E., & Greil, R. (2009). Endocrine therapy plus zoledronic acid in premenopausal breast cancer. The New England Journal of Medicine, 360, 679–691.
Haber, E., Danenberg, H. D., Koroukhov, N., Ron-El, R., Golomb, G., & Schachter, M. (2009). Peritoneal macrophage depletion by liposomal bisphosphonate attenuates endometriosis in the rat model. Human Reproduction, 24, 398–407.
Haber, E., Afergan, E., Epstein, H., Gutman, D., Koroukhov, N., Ben-David, M., Schachter, M., & Golomb, G. (2010). Route of administration-dependent anti-inflammatory effect of liposomal alendronate. Journal of Controlled Release, 148, 226–233.
Hava, D. L., Brigl, M., van den Elzen, P., Zajonc, D. M., Wilson, I. A., & Brenner, M. B. (2005). CD1 assembly and the formation of CD1-antigen complexes. Current Opinion in Immunology, 17, 88–94.
Hayday, A. C. (2000). Gamma delta cells: A right time and a right place for a conserved third way of protection. Annual Review of Immunology, 18, 975–1026.
Hewitt, R. E., Lissina, A., Green, A. E., Slay, E. S., Price, D. A., & Sewell, A. K. (2005). The bisphosphonate acute phase response: Rapid and copious production of proinflammatory cytokines by peripheral blood gd T cells in response to aminobisphosphonates is inhibited by statins. Clinical and Experimental Immunology, 139, 101–111.
Hinz, T., Wesch, D., Halary, F., Marx, S., Choudhary, A., Arden, B., Janssen, O., Bonneville, M., & Kabelitz, D. (1997). Identification of the complete expressed human TCR V gamma repertoire by flow cytometry. International Immunology, 9, 1065–1072.
Hiraoka, K., Zenmyo, M., Watari, K., Iguchi, H., Fotovati, A., Kimura, Y. N., Hosoi, F., Shoda, T., Nagata, K., Osada, H., Ono, M., & Kuwano, M. (2008). Inhibition of bone and muscle metastases of lung cancer cells by a decrease in the number of monocytes/macrophages. Cancer Science, 99, 1595–1602.
Kirpotin, D. B., Drummond, D. C., Shao, Y., Shalaby, M. R., Hong, K. L., Nielsen, U. B., Marks, J. D., Benz, C. C., & Park, J. W. (2006). Antibody targeting of long-circulating lipidic nanoparticles does not increase tumor localization but does increase internalization in animal models. Cancer Research, 66, 6732–6740.
Kunzmann, V., Bauer, E., Feurle, J., Weissinger, F., Tony, H. P., & Wilhelm, M. (2000). Stimulation of gamma delta T cells by aminobisphosphonates and induction of antiplasma cell activity in multiple myeloma. Blood, 96, 384–392.
Lang, J. K. (1990). Quantitative-determination of cholesterol in liposome drug products and raw-materials by high-performance liquid-chromatography. Journal of Chromatography, 507, 157–163.
Lewis, C. E., & Pollard, J. W. (2006). Distinct role of macrophages in different tumor microenvironments. Cancer Research, 66, 605–612.
Li, J. Q., Herold, M. J., Kimmel, B., Muller, I., Rincon-Orozco, B., Kunzmann, V., & Herrmann, T. (2009). Reduced expression of the mevalonate pathway enzyme farnesyl pyrophosphate synthase unveils recognition of tumor cells by V gamma 9 V delta 2 T cells. The Journal of Immunology, 182, 8118–8124.
Makkonen, N., Salminen, A., Rogers, M. J., Frith, J. C., Urtti, A., Azhayeva, E., & Monkkonen, J. (1999). Contrasting effects of alendronate and clodronate on RAW 264 macrophages: The role of a bisphosphonate metabolite. European Journal of Pharmaceutical Sciences, 8, 109–118.
Mariani, S., Muraro, M., Pantaleoni, F., Fiore, F., Nuschak, B., Peola, S., Foglietta, M., Palumbo, A., Coscia, M., Castella, B., Bruno, B., Bertieri, R., Boano, L., Boccadoro, M., & Massaia, M. (2005). Effector gamma delta T cells and tumor cells as immune targets of zoledronic acid in multiple myeloma. Leukemia, 19, 664–670.
Mayer, L. D., Bally, M. B., Cullis, P. R., Wilson, S. L., & Emerman, J. T. (1990). Comparison of free and liposome encapsulated doxorubicin tumor drug uptake and antitumor efficacy in the SC115 murine mammary-tumor. Cancer Letters, 53, 183–190.
Miyagawa, F., Tanaka, Y., Yamashita, S., & Minato, N. (2001). Essential requirement of antigen presentation by monocyte lineage cells for the activation of primary human gamma delta T cells by aminobisphosphonate antigen. The Journal of Immunology, 166, 5508–5514.
Monkkonen, J., & Heath, T. D. (1993). The effects of liposome-encapsulated and free clodronate on the growth of macrophage-like cells in vitro: The role of calcium and iron. Calcified Tissue International, 53, 139–146.
Monkkonen, J., Taskinen, M., Auriola, S. O., & Urtti, A. (1994). Growth inhibition of macrophage-like and other cell types by liposome-encapsulated, calcium-bound, and free bisphosphonates in vitro. Journal of Drug Targeting, 2, 299–308.
Patel, H. M. (1992). Serum opsonins and liposomes: Their interaction and opsonophagocytosis. Critical Reviews in Therapeutic Drug Carrier Systems, 9, 39–90.
Patel, H. M., & Moghimi, S. M. (1998). Serum-mediated recognition of liposomes by phagocytic cells of the reticuloendothelial system – the concept of tissue specificity. Advanced Drug Delivery Reviews, 32, 45–60.
Richards, P. J., Williams, B. D., & Williams, A. S. (2001). Suppression of chronic streptococcal cell wall-induced arthritis in Lewis rats by liposomal clodronate. Rheumatology, 40, 978–987.
Rodan, G. A. (1998a). Control of bone formation and resorption: Biological and clinical perspective. Journal of Cellular Biochemistry. Supplement, 30–31, 55–61.
Rodan, G. A. (1998b). Mechanisms of action of bisphosphonates. Annual Review of Pharmacology and Toxicology, 38, 375–388.
Rodrigueza, W. V., Pritchard, P. H., & Hope, M. J. (1993). The influence of size and composition on the cholesterol mobilizing properties of liposomes in vivo. Biochimica et Biophysica Acta, 1153, 9–19.
Roelofs, A. J., Jauhiainen, M., Monkkonen, H., Rogers, M. J., Monkkonen, J., & Thompson, K. (2009). Peripheral blood monocytes are responsible for gamma delta T cell activation induced by zoledronic acid through accumulation of IPP/DMAPP. British Journal of Haematology, 144, 245–250.
Sela, E., Chorny, M., Gutman, D., Komemi, S., Koroukhov, N., & Golomb, G. (2010). Characterization of monocytes-targeted nanocarriers biodistribution in leukocytes in ex-vivo and in-vivo models. Nano Biomedicine and Engineering, 2, 1–10.
Shmeeda, H., Amitay, Y., Gorin, J., Tzemach, D., Mak, L., Ogorka, J., Kumar, S., Zhang, J. A., & Gabizon, A. (2010). Delivery of zoledronic acid encapsulated in folate-targeted liposome results in potent in vitro cytotoxic activity on tumor cells. Journal of Controlled Release, 146, 76–83.
Slegers, T., van Rooijen, N., van Rij, G., & van der Gaag, R. (2000). Delayed graft rejection in pre-vascularised corneas after subconjunctival injection of clodronate liposomes. Current Eye Research, 20, 322–324.
Takahara, M., Miyai, M., Tomiyama, M., Mutou, M., Nicol, A. J., & Nieda, M. (2008). Copulsing tumor antigen-pulsed dendritic cells with zoledronate efficiently enhance the expansion of tumor antigen-specific CD8+ T cells via V gamma 9 gamma delta T cell activation. Journal of Leukocyte Biology, 83, 742–754.
Torchilin, V. P. (2005). Recent advances with liposomes as pharmaceutical carriers. Nature Reviews. Drug Discovery, 4, 145–160.
van Rooijen, N., & Van Kesteren-Hendrikx, E. (2003). “In vivo” depletion of macrophages by liposome-mediated “suicide”. Methods in Enzymology, 373, 3–16.
Veltman, J. D., Lambers, M. E. H., van Nimwegen, M., Hendriks, R. W., Hoogsteden, H. C., Hegmans, J., & Aerts, J. (2010). Zoledronic acid impairs myeloid differentiation to tumour-associated macrophages in mesothelioma. British Journal of Cancer, 103, 629–641.
Wilhelm, M., Kunzmann, V., Eckstein, S., Reimer, P., Weissinger, F., Ruediger, T., & Tony, H. P. (2003). Gamma delta T cells for immune therapy of patients with lymphoid malignancies. Blood, 102, 200–206.
Zeisberger, S. M., Odermatt, B., Marty, C., Zehnder-Fjallman, A. H., Ballmer-Hofer, K., & Schwendener, R. A. (2006). Clodronate-liposome-mediated depletion of tumour-associated macrophages: A new and highly effective antiangiogenic therapy approach. British Journal of Cancer, 95, 272–281.
Zhang, Y. H., Cao, R., Yin, F. L., Lin, F. Y., Wang, H., Krysiak, K., No, J. H., Mukkamala, D., Houlihan, K., Li, J. K., Morita, C. T., & Oldfield, E. (2010). Lipophilic Pyridinium Bisphosphonates: Potent gamma delta T Cell Stimulators. Angewandte Chemie-International Edition, 49, 1136–1138.
Zito, M. A., Koennecke, L. A., McAuliffe, M. J., McNally, B., van Rooijen, N., & Heyes, M. P. (2001). Depletion of systemic macrophages by liposome-encapsulated clodronate attenuates striatal macrophage invasion and neurodegeneration following local endotoxin infusion in gerbils. Brain Research, 892, 13–26.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Gutman, D., Epstein-Barash, H., Tsuriel, M., Golomb, G. (2012). Alendronate Liposomes for Antitumor Therapy: Activation of γδ T Cells and Inhibition of Tumor Growth. In: Zahavy, E., Ordentlich, A., Yitzhaki, S., Shafferman, A. (eds) Nano-Biotechnology for Biomedical and Diagnostic Research. Advances in Experimental Medicine and Biology, vol 733. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-2555-3_16
Download citation
DOI: https://doi.org/10.1007/978-94-007-2555-3_16
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-2554-6
Online ISBN: 978-94-007-2555-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)