
Abstract
The current official silvicultural instructions of Finland recommend even-aged rotation forest management (RFM) combined with low thinning and artificial regeneration. However, the direction is gradually changing towards increasing freedom and flexibility in forest management. The first sign of a new silvicultural era was the gradual approval of high thinning, which is now accepted although it was strictly forbidden in the past. As a further step, uneven-aged management and other forms of continuous cover forestry (CCF) are now gaining popularity and acceptance. The harmful impacts of clear felling and plantation forestry on the recreational value and biodiversity of forests have been increasingly emphasized. In addition, encouraging results on the yield and profitability of CCF have been obtained recently. This chapter reviews those results. The review shows that CCF is often more profitable than even-aged management. The superiority of CCF increases if non-wood benefits are included in the analysis. The growth rates of most Finnish forests are too low to warrant the high stand establishment and management costs of even-aged plantation forestry. If the forest landowner wants to practice forestry that is profitable (without state subsidies) also with high discounting rates, on poor growing sites and in the northern parts of Finland, her only option is continuous cover management.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
In the past, Finnish landowners managed their forests using various types of selective cuttings. Only large trees were merchantable and therefore they were the only ones that were cut. Some smaller trees were used for household consumption. As a consequence, almost all forests were uneven-aged and uneven-sized (Lähde et~al. 1991, 1992b). Most of the mature stands of Finnish forests are still rather uneven-sized despite the fact that even-aged management with clear felling and low thinning has been recommended for several decades (Pukkala et al. 2011a). This is mainly because of the gradual regeneration and ingrowth processes, which tend to convert even-aged stands into two- and multi-layered stands.
The traditional uneven-aged management with selective cuttings did not provide enough cheap raw materials for the expanding pulp and paper industry in the middle of the 1900s. Therefore, a group of leading forestry authorities started to promote even-aged rotation forestry (also called rotation forest management, RFM) with low thinning, clear felling, and planting. This type of silviculture was preferred for the production of small-sized wood required by the pulp and paper mills (Pukkala et al. 2011a). One outcome of this mission was the Declaration Against Uneven-aged Management, issued in 1948. Together with other initiatives, it led to the gradual abandonment of uneven-aged forest management in Finland (Siiskonen 2007). Also natural regeneration with seed tree and shelter wood methods was discouraged. As a result, more and more forest areas were established by planting, using selected seed of mainly Scots pine at first and Norway spruce more recently.
Many landowners realized that RFM with clear felling and planting would not maximize their benefits, and they wanted to continue practicing selective cuttings. However, since the traditional uneven-aged management was now considered unacceptable and interpreted even illegal, these landowners were punished, and timber sales from their forests were barred. They had to switch to clear-felling, planting, and low thinning if they wanted to get any income from their forests (Pukkala et~al. 2011a).
There was no research available that would have proven the superiority of the new silvicultural paradigm. Not only the use but also the research of uneven-aged management was discouraged for several decades and no objective comparisons of alternative management systems were done. Until the 1980s, no experiments were established for comparing the yield and profitability of uneven-aged vs. even-aged management. It is evident that RFM provides more conifer pulpwood, but its profitability to the forest landowner was never analyzed. The harmful impacts of clear felling on visual landscapes, forest diversity, erosion, nutrient leaking, soil carbon balance, and various non-timber products and services were ignored when RFM was advertised.
At the moment, uneven-aged management is gaining popularity in Finland. The old practices are becoming acceptable again. They have been modernized and diversified, and are called as “continuous cover forestry” (CCF). All management systems that do not use clear-felling and planting are regarded as different variants of CCF. The increasing interest towards CCF is for several reasons. One is the decreasing importance of pulp and paper industries in the Finnish economy. Instead, nature tourism is a growing economy. Paper consumption is decreasing in countries that import Finnish paper. The high energy consumption of mechanical mass industry with harmful climatic impacts is another reason for the gradual decline of this industry, reducing the need for spruce pulpwood. The harmful impacts of RFM on the recreational value and biodiversity of forests have been increasingly emphasized. According to recent surveys (Valkeapää et~al. 2009), only 5% of Finns and half of forest landowners approve clear-felling without any reservations.

Uneven-aged spruce forest in South Finland (Photo: Timo Pukkala)
One reason for the incipient new era of increased freedom and variability in forest management are the encouraging results on the yield and profitability of CCF (Pukkala et~al. 2010; Tahvonen et~al. 2010). Although investigations on CCF were actively constrained for several decades, some researchers managed to set up silvicultural experiments in which various forms of CCF were included. These experiments were established during the 1980s with the aim of comparing alternative silvicultural systems (Lähde et~al. 1992a). The experiments have already provided useful empirical data for growth and yield analyses (e.g., Lähde et~al. 1999a). The measurements of the experiments have also been used for growth modeling. The growth models, in turn, have enabled detailed long-term analyses of the yield and profitability of alternative management systems. Several studies on the growth and profitability of CCF have been conducted and reported during the past few years. Some studies also deal with the non-timber benefits provided by different management systems. An indication of the growing interest and activity is that two silvicultural text books on CCF have recently been published in Finland (Valkonen et~al. 2010; Pukkala et~al. 2011a).
This chapter provides a review on the recent Finnish research related to CCF. The next section describes the experiments established on CCF and the results obtained from them. The third part reviews the models developed for the stand dynamics of uneven-aged forests. Then, results on economically optimal uneven-aged management are summarized. A separate section is devoted to multifunctional forestry. Finally, the last section discusses what research is still necessary for the improved management of Finnish forests and for increasing reliability of the decision support provided by models and calculations.

Uneven-aged pine forest in South Finland (Photo: Jussi Saarinen)
2 Experiences from Silvicultural Experiments
Several experiments on CCF were established in Finland during the 1980s. The best known experiments are those of Honkamäki and Vessari in southern Finland. The forests of these experimental areas were regenerated mainly for spruce by the shelter wood method during the 1940s. The shelter trees were removed in 1957 and 1960. The experiments that compare different silvicultural methods were started in 1986 and 1987. Both experiments consist of tens of plots (92 plots altogether). Four main treatments were included: (1) RFM with different intensities of low thinning and CCF with various types of (2) single-tree selection and (3) diameter-limit cutting; and (4) no cuts at all (control). Tree species composition and post-cutting stand density were varied in low thinning and single-tree selection. Every treatment was represented by several plots, and the treatments of the plots were randomized. The plots have been measured at 3-year intervals, on average. The trees were mapped in 1994. Cuttings have been done from one to four times depending on the treatment, in 1986/1987, 1994, 2003/2004 and 2009.
At the latest measurement, in 2009, the even-aged plots had already reached the rotation age of RFM, and several of them were clear felled in autumn 2009. The other even-aged plots were treated with different variants of regenerative cuttings (seed tree and shelter wood cuttings using different intensities, and conducted as high, uniform or low thinning). Therefore, the experiments already provide growth and yield information from a period that corresponds to one full rotation (75 years) of even-aged management. In Fig. 3.1 the total yield equals the sum of ending volume, harvested volume, volume of mortality, and the volume increment of the initial shelter trees. The growth of the shelter trees during 1945–1957/1960 has been estimated at 70 m3 ha−1 in Vessari and 50 m3 ha−1 in Honkamäki.

Total volume production in different management systems in the Vessari and Honkamäki experiments during a 75-year period (typical rotation length in even-aged forestry). “Shelter trees” refers to the volume increment of shelter trees during the first years of the rotation (shelter tree cut – removal of shelter trees). RFM rotation forest management (even-aged management); STS single-tree selection; DLC diameter-limit cutting; NoC no cuttings
According to the measurements, the total wood production has been 3% (Vessari) or 11% (Honkamäki) higher in single-tree selection than in RFM. If mortality is included in the yield, control (no cuttings) is the second best in Vessari and the third in Honkamäki. In Vessari, the diameter-limit cutting removed all trees larger than 9 cm in 1986. This kind of uneven-aged management has commonly been regarded as exploitive and destructive. However, since the spruce under-storey was well developed, the total production of diameter-limit cutting was the same as in even-aged management. In the Honkamäki experiment, diameter-limit cuttings were carried out in 1987 and 2004, both removing all trees larger than 16 cm in dbh. Diameter-limit cutting has been the second best in total wood production in Honkamäki.
Assuming that the harvested timber was sold with the current stumpage prices, the net present values of different management systems, discounted to 1986/1987, are as shown in Fig. 3.2. The net present value (NPV) of the ending forest (predicted net present value of cuttings after 2009, to infinity) has been calculated with the models of Pukkala (2005), and discounted to 1986/1987. The most profitable management option in 1986/1987 had been diameter-limit cutting, both in Vessari and Honkamäki. RFM with low thinning had been the least profitable option in both experiments.

Discounted net revenue (with 3% rate) in alternative management systems in the Vessari and Honkamäki experiments. All net revenues have been discounted to 1986 or 1987 when the stands were 40 years old and the experiments were started. The net present value (NPV) due to future cuttings (NPV of ending volume) has been estimated with the models of Pukkala (2005). Some of the even-aged plots were clear felled and planted with Norway spruce in 2009 and some were left for natural regeneration without any site preparation. RFM rotation forest management; STS single-tree selection; DLC diameter-limit cutting; NoC no cuttings
Many other experiments were also established in different parts of southern and northern Finland during the 1980s, mainly for Scots pine in the north and Norway spruce in the south. The total number of plots in these scattered smaller experiments is about 150 in southern Finland. These plots have been monitored less systematically than the Vessari and Honkamäki experiments. The measurement interval has typically been 5–10 years. The drawback in these experiments is that trees have not been numbered and their coordinates have not been measured. Therefore, the growth and survival of individual trees are not known. Yet, the plots provide valuable information for growth and yield studies (e.g., Pukkala et~al. 2011b) and for validating the results based on other materials. An advantage is that the plots are reasonably large (40 m by 40 m) and each plot is surrounded by forest with similar management.
Several research articles have been published based on these experiments. For instance, regeneration in uneven-aged stands was studied in the beginning of 1990s (Lähde 1992a, b). Plots treated with commercial selective fellings 10–30 years earlier and those treated with single tree selection 4–7 years ago produced, in general, rich regeneration. In both materials, regeneration decreased with increasing stand volumes. During the first monitoring period of 11 years, the CCF-plots had almost 20% faster volume increment than the low-thinning plots of RFM (Lähde et~al. 2002a; Saksa et~al. 2003). Another study compared single-tree selection, low thinning and diameter limit cutting (Lähde et~al. 2001) and found single-tree selection to be the most productive.
In the experiment established in an old-growth Norway spruce stand in eastern Finland and treated with various modifications of CCF-management, the structural diversity index (Lähde et~al. 1999b) was clearly the highest in untreated plots (Lähde et~al. 2002b). Single-tree selection was the second best whereas even-aged management resulted in the lowest structural diversity.
Recently, new large silvicultural experiments have been established in state-owned forests. They aim at mimicking the natural disturbance dynamics occurring in forests (University of Helsinki 2010). Two large forest areas (1,200 and 650 ha) have been assigned for the studies. The idea is to analyze, not only even-aged and uneven-aged management, but also management systems that are between these extremes. An example of the in-between systems is regeneration by small canopy gaps. The purpose is to compare cuttings that mimic small disturbances (individual trees and tree groups removed), medium disturbances (many trees removed and gaps created), or heavy disturbances (large-scale clear fellings). The experiments are useful but since the study was initiated in 2009, it will take many years until major results can be expected.
Measurements of national forest inventory plots have also been used to compare even- and uneven-sized forests (e.g., Lähde et~al. 1994a, b, 2002c). These data sources have been used to analyze the structure of the forests (Pukkala et~al. 2011a) and to examine regeneration (Lähde et~al. 1999c) and tree growth in different stand types (Pukkala et~al. 2009). National forest inventory data cover all growing sites and stand types and represent the whole surface area of Finland. The drawback is that the management history of stands is not well known and only periodical growth information is available. Yet, national forest inventory data can be used for modeling the dynamics of uneven-aged forests. These models may then be used to analyze the wood production and profitability of alternative management systems.
3 Growth and Yield Modeling
The first growth and yield models developed specifically for uneven-aged forests in Finland were transition matrix models (Pukkala and Kolström 1988; Kolström 1993). A transition matrix model predicts the probability of a tree of certain size to stay in the same diameter class, or move to the next higher class (Chap. 6). Mortality is also accounted for since, in these models, trees that do not stay in the same class nor move to another class are taken as mortality. Ingrowth is also incorporated into the transition matrix models.
The above models have been used very little for analyzing the yield and profitability of uneven-aged management because the modeling data did not include stands that are known to have been treated with repeated selective high thinnings. A more recent transition matrix model was presented by Tahvonen et~al. (2010). This model is based on data collected in the Vessari and Honkamäki experiments, where more than half of the plots represent uneven-aged management. However, this new transition matrix model is only locally valid since the modeling data came from two experiments only, located close to each other.
Individual-tree models are another type of growth models suitable for simulating the dynamics of uneven-aged stands. Management planning in Finnish forestry has a long tradition of employing individual-tree models, which means that this model type is familiar to forest planners and people who develop decision support systems for forestry decision making. The first individual-tree models specifically targeted to uneven-aged stands are those of Pukkala et~al. (2009). They were based on three data sets with a total of 50,000 diameter growth measurements: (1) the experiments of Honkamäki and Vessari, (2) sample trees of the third national forest inventory (1951–1953), and (3) a set of plots measured in irregular conifer stands in Eastern Finland (158 plots). The majority of the data comes from uneven-sized stands since most forests were still uneven-sized during the third national forest inventory, and half of the plots of Vessari and Honkamäki were managed using CCF.
In the individual-tree approach the minimum set of models required for simulating stand dynamics consists of
-
individual-tree diameter increment model;
-
individual-tree survival model; and
-
ingrowth or regeneration model.
In addition, height, volume and biomass models are usually employed in calculations, but they are not necessary for simulating the stand dynamics. Pukkala et~al. (2009) fitted ingrowth models using 5 cm dbh as the ingrowth limit; the models predict the number of trees that pass the 5-cm limit during the coming 5-year period. However, since these ingrowth models were based on a rather small data set (ingrowth was unknown for the national forest inventory plots) and the model behavior was not always logical, Pukkala et~al. (2011b) fitted new models based on 140 plots measured two to four times in various silvicultural experiments in southern and central Finland. The ingrowth diameter limit was now 0.5 cm. Therefore, the best set of models currently available for simulating the dynamics of uneven-aged stands with the individual-tree approach consists of the diameter increment and survival models of Pukkala et~al. (2009) and the ingrowth models of Pukkala et~al. (2011b). In this model set, the diameter increment models are as follows:
where i d is diameter increment (cm/5 years), d is dbh (cm), G is stand basal area (m2 ha−1) and TS temperature sum (°C). TS is equal to the sum of mean daily temperatures minus 5° of those days on which the mean daily temperature is at least 5°C. MT, VT, CT and ClT are indicator variables indicating whether the site is mesic (MT), sub-xeric (VT), xeric (CT) or poorer than xeric (ClT). BAL is the basal area in trees larger than the subject tree (m2 ha−1). BAL is calculated separately from spruces (BAL Spruce) and the other tree species (BAL Others). BAL describes the competitive status of the tree within a stand. It is the most significant predictor in the models. The models indicate that small trees grow slowly and large trees grow well in an uneven-aged stand (Fig. 3.3, right). Slow early growth results in good wood quality, and fast growth at older ages maintains high relative value increment. Good diameter growth in large trees is no longer a disadvantage for wood quality.

Dependence of diameter increment on diameter in an uneven-aged stand growing on a mesic site in southern Finland when the stand basal area is 25 m2 ha−1. In the left diagram, BAL has a fixed value of 10 m2 ha−1. On the right, BAL decreases from 25 m2 ha−1 to zero when diameter increases from zero to 40 cm
The survival models are:
where p is the probability that a tree survives during the coming 5 years. According to the models, large trees have a greater chance to survive than small ones. Trees facing much competition (having large BAL) die more often. A few percents of trees smaller than 10 cm die during a 5-year period. Trees larger than 10 cm die infrequently. Spruce trees survive better than pines and birches.
The ingrowth models are (Fig. 3.4):

Five-year ingrowth of different species in pure stands (left) and in a mixed stand (right) on mesic site according to the ingrowth models. The share of each species in the mixed stand is 33% of basal area
where Ni is the number of trees that pass the 0.5-cm diameter limit during the coming 5-year period. Indicator variable MT − equals 1 if the site is mesic or poorer, otherwise MT − = 0.
The above model set has been used in all the optimizations and calculations discussed in this chapter. By using the models, one 5-year time period of stand development is simulated as follows:
-
Calculate the predictors of the models
-
Calculate the survival probabilities of trees and multiply the frequencies of trees by their survival probabilities
-
Calculate the diameter increments of trees and add them to the current tree diameters
-
Calculate ingrowth and add new trees to the tree list
-
Calculate tree height, volume, biomass and other characteristics as required
A forest stand may be represented by a list of trees in different size classes or by a plot in which each tree is known and treated individually in the simulation. In the first case, every tree has its own frequency, i.e., it represents a certain number of trees per hectare, and mortality is simulated by reducing the tree’s frequency. In the latter case, mortality is simulated by assigning the tree either as a survivor or non-survivor. This can be done by means of random numbers (the predicted survival probability is compared to a uniformly distributed random number) or using a certain limiting survival probability (e.g., 0.5) beyond which a tree is assigned as a survivor. In the analyses of this chapter, a stand was represented by a list of sample trees, and mortality was simulated by multiplying tree frequencies by the trees’ survival probabilities. When a list of sample trees corresponding to a certain diameter distribution is generated, sample trees are taken at 0.5- or 1-cm intervals over the range of breast height diameters.
4 Optimization of CCF Management
4.1 Problem Formulations and Optimization Methods
We have used the following simple formula to calculate the net present value for the steady-state management of an uneven-aged forest (Chang 1981):
where N T is the net income obtained regularly at T-year intervals (after T, 2T, 3T,… years), T is the cutting cycle, i is discount rate (percentage divided by 100) and A T is the stumpage value of the post-thinning stand. The NPV of a certain post-thinning steady state diameter distribution is calculated as follows. (1) Calculate the value of the growing stock (A T ); (2) simulate stand development for T years using the models explained above; (3) reduce the frequencies of diameter classes back to their initial levels and calculate the net income (N T ) from the harvesting costs and roadside values of the removal; and (4) calculate NPV from Eq. 3.1 (see also Chang and Gadow 2010).
If the purpose is not to find the optimal steady-state structure but to optimize the management of an existing stand, one possibility is to simulate the growth and management for a long enough time and calculate the NPV from the simulation results. If the simulation period is short or discounting rate is low, the net present value of the ending growing stock should be taken into account. One way of doing this is to continue simulation until the forest reaches a steady state, then calculate the periodical income of the steady-state forest, and add the NPV of this periodical income to the NPV accrued before the steady state. It is also possible to constrain the optimization to reach a predefined steady-state uneven-aged structure during a conversion period. In this case, the last simulated cutting, which converts the stand into the steady-state structure, is not optimized.
The most straightforward way to formulate the optimization problem is to use the post-cutting frequencies of different diameter classes as decision variables. This choice has the advantage of being flexible, i.e., the diameter distribution is not constrained to a certain shape (such as the “inverse J” shape). A drawback is that the number of decision variables becomes large, especially if a series of successive transformation cuttings or mixed stands are optimized, in which different tree species have their own, species-specific diameter distributions.
To analyze the effect of detail in describing the stand structure, Pukkala et~al. (2010) compared four alternative sets of decision variables in the optimization of the steady-state structure of a spruce stand (Fig. 3.5):

Alternative ways to describe the post-thinning diameter distribution of an uneven-aged stand in optimization. On the left, the decision variables are the frequencies of diameter classes, which may be smoothed with a spline function to avoid stepwise distributions. On the right, the decision variables are the parameters of the Weibull or Johnson’s SB function
-
1.
Frequencies of 4-cm wide diameter classes
-
2.
Smoothed frequencies of 4-cm wide diameter classes
-
3.
Parameters of the Weibull distribution
-
4.
Parameters of the Johnson’s SB distribution
In all cases, the distribution was sampled at 0.5-cm intervals. The result was a set of sample trees that represented the stand in simulation. When the frequencies of 4-cm diameter classes were used as decision variables (option 1), the frequencies of all sample trees falling within a given diameter class were equal (Fig. 3.5). In the case of smoothed frequencies, a cubic spline function was fitted to the decision variables, and the frequencies of sample trees were obtained from the spline function (Fig. 3.5). Therefore, the frequencies of sample trees within a 4-cm diameter class were unequal.
When using the Weibull or Johnson’s SB function, the optimized decision variables were the parameters of the function, together with the total number of trees per hectare. The Weibull density function is defined as follows:
where d is diameter, f(d) is frequency of diameter d, a is minimum diameter, b is a location parameter and c is a shape parameter. The Johnson’s SB function is:
with
where ξ is the minimum and λ the range of the distribution, and γ and δ are shape parameters.
In all four methods to describe the post-thinning diameter distribution, the maximum diameter retained after a thinning was used as an additional decision variable; post-thinning frequencies of all trees larger than this diameter were taken as zero. Therefore, in the case of the Weibull function, the optimized variables were the total number of trees, parameters a, b, and c of the distribution function, and the maximum diameter (5 decision variables). There was one more decision variable when Johnson’s SB was used because this function has two shape parameters. When frequencies were used, the number of diameter classes was 10 (with class mid-points of 7, 11, 15, …, 43 cm), which means that the number of optimized decision variables was 11.
The growth of a forest stand was simulated for 20 years (T = 20 years), after which a felling was simulated, which reduced the frequencies of diameter classes to their initial levels. In this analysis, N T was calculated from the stumpage value of the removal and A T from the stumpage value of the initial stand. A penalty function was used to guarantee that the remaining frequencies of diameter classes, after simulating a cutting at the end of cutting cycle, were not less than the initial frequencies. This requirement may be violated if the ingrowth to a certain class is less than the transition of trees to larger diameter classes. Thus, the objective function, which was maximized in the optimizations, was NPV minus a penalty:
with
where \( {f^\prime_i} \) is the frequency of trees in diameter class i at the end of cutting cycle, i.e., after simulating stand development for T years and simulating a thinning treatment at the end of the cycle, f i is the initial frequency of trees in diameter class i, and n is the number of diameter classes. The multiplier i in the formula ensures that a lack of large trees was penalized more than a lack of small trees. Another optimization was made in which the saw log harvest was maximized instead of NPV.
Four different optimization techniques were used to solve the problems for a Norway spruce stand growing on medium site in South Finland. The methods were Differential Evolution, Particle Swarm Optimization, the Nelder-Mead method (also called Amoeba Search or Polytope Search), and Evolution Strategy Optimization (Pukkala 2009).
Based on the results, it was concluded that the Weibull function (together with maximum retained diameter) and Evolution Strategy Optimization was the best combination of decision variables and optimization method (Fig. 3.6). Therefore, the decision variables used in the optimizations described in this chapter use Evolution Strategy Optimization with the following decision variables: parameters a, b, and c of Weibull distribution (aW, bW and cW), number of trees per hectare (N), and maximum diameter retained in cutting (Dmax). Later on, it was found that assuming aW to be equal to zero does not usually deteriorate the result. Therefore, the optimized decision variables have mostly been bW, cW, N and Dmax in the calculations reported in this chapter. We recognize that the use of the Weibull distribution function constrains the diameter distribution to be descending or unimodal, but this does not seem to be a problem, at least when steady-state stand structures are optimized.

Mean value of objective variable (net present value with 2% discount rate or mean annual saw log production) in 10 repeated optimizations with four optimization methods and four ways to describe the post-thinning diameter distribution in a spruce stand on herb-rich heath in South Finland (DE Differential evolution; PS Particle Swarm Optimization; NM Nelder-Mead method; ES Evolution Strategy Optimization)
4.2 Optimal Steady-State Management
Tables 3.1 and 3.2 show the optimal steady state post-thinning diameter distributions of spruce and pine stands on different growing sites in South (temperature sum 1,200 d.d.) and North Finland (temperature sum 900 d.d.) when net present value is maximized with a 2% discount rate. The cutting cycle varies from the 15 years of fertile sites in South Finland to 50 years of poor sites in North Finland. When economic profitability is maximized the optimal management involves removing all trees larger than 20–25 cm. This means that all sawlog-sized trees are harvested with the exception that some small log-sized trees are retained in the spruce stands of South Finland.
The remaining stand basal areas are low, 7.5–9 m2 ha−1, and slightly less than the accepted minimum in the current forestry regulations for the thinning of an even-aged stand. However, since the number of trees is high (always more than 1,300 trees taller than 1.3 m), the post-cutting stand is not under-stocked. If the selection cutting is interpreted as a removal of the over-storey trees, then there is no discrepancy between the optimization result and the current regulations. The mean annual wood production ranges from 0.9 m3 ha−1 on the poor sites of North Finland to 6.6 m3 ha−1 on fertile sites in South Finland. If wood production were maximized, instead of profitability, the sustainable yield would be 1.5 to 7.8 m3 ha−1 a−1 (Pukkala et~al. 2010).
The results of Tables 3.1 and 3.2, as well as all the remaining optimization results discussed in this chapter, have been calculated by using roadside timber prices. The harvesting costs have been calculated with the models of Rummukainen et~al. (1995), which take into account, among other things, the forwarding distance, size of harvested trees, and harvested volume per 100 meters of extraction road. Typically, the harvesting cost per cubic meter in steady-state CCF is about the same as in a normal thinning of a mature even-aged stand. Harvesting is more costly than in clear felling, but cheaper than in the first commercial thinning of even-aged management. When taking the whole rotation, there is no major difference between the harvesting costs of CCF and RFM (Pukkala et~al. 2011a).
As expected, increasing discount rates lead to decreasing density and stumpage value of the post-cutting residual growing stock (Fig. 3.7). However, due to the requirement for constant regeneration and ingrowth, the basal area of the post-cutting residual stand is never more than 9 m2 ha−1 when NPV is maximized. The removal of pulpwood-sized trees (dbh < 19 cm) is low even with high discounting rates since the relative value increment of these trees is high. Due to these reasons, the post-thinning basal area is not particularly sensitive to the discount rate.

Effect of discount rate on the steady-state structure of a spruce stand growing on fertile herb-rich site in Central Finland
If saw log production or net income is maximized, instead of NPV, the post-cutting stand will have more large trees (Fig. 3.8). However, maximal saw-log production greatly decreases economic profitability (Schulte et~al. 1999; Pukkala et~al. 2010). On the other hand, near maximal saw-log yields can be obtained with only 10% decrease in NPV (Pukkala et~al. 2011a), which means that fairly good saw-log production and fairly good profitability can be attained simultaneously.

Optimal post-cutting diameter distribution in a herb-rich spruce stand in southern Finland when the objective variable is net income, saw log production, or net present value with 1% or 4% discount rate
The optimal steady-state stands lack large trees. Therefore, the stand structures may not be the best possible for multiple-use forests. To find management options that are more suitable for recreation, we have also optimized the post-thinning residual stand structure with a penalty for lack of large trees. In these calculations the solution was penalized if the post-thinning stand contained too few large trees. Table 3.3 shows that the requirement for large trees in the post-cutting stand decreases both economic profitability and yield. However, since economic profit is not the only objective in multiple-use or recreational forests, these constrained schedules may be the preferred ones especially in regions where nature tourism is important. It is also possible that the value of large trees increases in the near future, since timber is again favored in construction. Large trees also have the smallest share of pulpwood, and especially spruce pulpwood has a declining price.
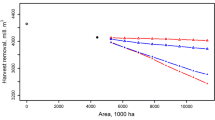
When the NPV of the optimal uneven-aged steady-state management has been compared to optimal even-aged management, uneven-aged management has been more profitable in almost all calculations (Pukkala et~al. 2010; Tahvonen et~al. 2010). The two management systems have been found equally profitable on the best growing sites of South Finland with a low discount rate (1%). In all other cases, uneven-aged management has been more profitable. The relative advantage of uneven-aged management increases with decreasing site quality and timber price, and with increasing discount rate and management costs (Fig. 3.9). The net present value of even-aged management is negative already with a 5% discounting rate (Hyytiäinen and Tahvonen 2003) whereas the optimal uneven-aged management has a positive NPV with all discount rates.

Relative net present value (optimal CCF = 100) of the current even-aged management system (clear-felling, planting, low thinning), optimal even-aged plantation forestry (optimal RFM) (clear-felling, planting, optimal high thinning) and optimal uneven-aged steady-state management (optimal CCF) in Central Finland in a spruce stand on mesic site (left) and pine stand on sub-xeric site (right) with 1%, 3% and 5% discount rates
Compared to the currently recommended even-aged management, the optimal even-aged management would have the first commercial thinning later, and the thinnings would be high thinnings instead of the prevailing practice of thinning from below (low thinning). The stand basal area would be gradually decreased towards the end of the rotation, which is opposite to the current official recommendations, which suggest increasing the stand basal area with increasing stand age.
The only way to guarantee positive net present values with high discount rates is to minimize costs, i.e., to reduce the intensity of forest management. This is possible also in even-aged management. The NPV of even-aged management is positive, irrespective of discount rate, if the clear-felled area is left to regenerate naturally, without site preparation, sowing, planting and tending operations, and timber is sold only when the stumpage price is positive, i.e., when the road side price is higher than the harvesting cost. In practice, this kind of extensive management would mean natural regeneration with the seed tree or shelter wood methods or using small canopy gaps. Because these methods may be regarded as variants of continuous cover forestry, it may be concluded that if the forest landowner wants to practice forestry that is profitable (without state subsidies) also with high discount rates, on poor growing sites and in northern parts of Finland, her only option is continuous cover management.
4.3 Species Composition in Multi-species Stands
The steady-state structure of a mixed stand with more than one tree species can be optimized by finding the optimal post-thinning diameter distributions separately for the different tree species of the stand. The cuttings return the species-specific frequencies of diameter classes to their optimal initial levels. The solution is penalized if the post-cutting frequency of any diameter class of any species is less than the initial frequency. In the problem formulation adopted in this chapter, the decision variables of the optimization problem for mixed steady-state stands are the species-specific values of Weibull parameters b and c, number of trees per hectare, and maximum diameter retained in thinning.
Realistic species mixtures on the most fertile growing sites of Finland (herb rich, OMT) are those of birch and spruce. On medium sites (mesic site, MT), all mixtures of the main tree species (spruce, pine and birch) are possible. Pine is the best-growing species on sub-xeric site (VT), but pine-spruce mixtures are also feasible since spruce frequently enters sub-xeric pine forests as an under-storey and attains reasonable dimensions. On the very poorest sites, pine is the only economically meaningful option.

Spruce enters birch forests on fertile sites (Photo: Olavi Laiho)
According to the steady-state optimizations for mixed stands, the best species composition on the best growing sites is a pure spruce stand if economic profitability is maximized, even though a small admixture of birch would increase yield. This result is mainly due to the fact that the price of spruce pulpwood is clearly better than that of birch pulpwood. However, increased use of wood as biofuel, and the decreasing demand for spruce pulpwood for mechanical mass may alter the situation making birch mixtures more profitable. An admixture of birch and other hardwoods would also improve the scenic value (Silvennoinen et~al. 2001) and structural diversity (Lähde et~al. 1999a) of the stand, and enhance regeneration. Therefore, maintaining an admixture of hardwood seems to be worthwhile on the best sites.
On medium sites the most profitable two-species combination is a mixed stand of spruce and pine, spruce being the dominant species. A mixed stand is slightly more productive and more profitable than a pure spruce stand (Fig. 3.10). Similarly as on the best growing sites, some birches should be maintained since they enhance regeneration, amenity values, and diversity.

Net present value (left) and sustained annual production (right) of a pine-spruce stand on mesic site in Central Finland when NPV is maximized with 2% discount rate, and the share of pine in the post-thinning stand is 0%, optimal (15%), 50% or 100% of basal area
The optimal structure of a sub-xeric site is a pine-spruce mixture, in which pines are larger than spruces (Fig. 3.11). The stand therefore looks like a two-layered stand where the spruce layer consists of smaller trees than the pine layer. However, the pine and spruce layers overlap. Many current forests already show this type of structure, except that there are too few small pines compared to the optimal uneven-aged structure. In many cases, the lower spruce storey is denser than the optimal post-thinning distribution suggests. The challenge of sustainable steady-stage management of a mixed conifer stand is therefore to maintain the regeneration and ingrowth of pine. This can be done by reducing the stand basal area to a lower level than currently used, and thinning the lower storey of spruce. Creating small gaps or very sparse areas may also be necessary to promote the regeneration of pine and hardwoods. Another question is how meaningful it is to maintain a mixed stand in a steady state. It might be easier and more profitable to alternate between periods of pine and spruce dominance on medium sites, and between hardwood and spruce dominance on fertile sites.

Optimal steady-state structure and management of pine and spruce mixture on sub-xeric site in Central Finland with 20-year cutting cycle when NPV with 2% discount rate is maximized

Spruce enters pine stands on medium sites (Photo: Timo Pukkala)
4.4 Optimizing Transformation Cuttings
Another optimization problem is the conversion of an even-aged stand, or any existing stand, to the optimal steady state structure. As explained earlier in this chapter and in the other chapters of the book, this can be done in two different ways. One is to optimize the steady state diameter distribution simultaneously with the transformation cuttings. A slightly easier way is to optimize a certain number of transformation cuttings with a constraint that the next cutting (after the optimized ones) converts the stand into a predefined steady-state structure. In both cases, the optimized variables are the diameter distributions of several successive post-thinning stands; a sequence of diameter distributions is optimized. The time points of cuttings can be fixed or they can be optimized simultaneously with the distributions.
This section presents optimization results for fertile spruce stands in which the number of transformation cuttings is four and their interval is 15 years (cutting years are 0, 15, 30 and 45). The fifth cutting, after 60 years, converts the stand into a predefined steady state structure. The optimized variables are the post-thinning diameter distributions of the first four cuttings, and each distribution is described by the same variables (bW, cW, N and Dmax). The solution is not penalized if there are too few remaining trees in some diameter classes after the four transformation cuttings. However, shortage is not allowed in the fifth cutting, since the treatment from that moment onwards should represent sustainable steady-state management. Because of this, the optimization in fact consists of finding a sequence of optimal cutting limits for the diameter classes; all excessive trees, as compared to the cutting limit, are removed.
In a mature even-aged spruce stand the optimal sequence of cuttings consists of an immediate shelter tree cut, removal of the shelter trees gradually in two cuttings after 15 and 30 years, and using normal uneven-aged management thereafter (Fig. 3.12, left). It is noteworthy that it is not optimal to manage the mature stand with repeated light high thinnings. Instead, it is more profitable to regenerate the stand first. Another important observation is that shelter trees should be selected from among the smallest trees of the stand. These trees have the best relative value increment and the lowest opportunity cost when left to continue growing. Dominated and co-dominant trees also have narrow crowns, and their removal therefore causes less damage than the removal of large trees with wide crowns.

Optimal path of transformation cuttings on a herb-rich site in a mature even-aged spruce stand (left) and in two-layered spruce stand (right) when the optimal steady-state diameter distribution must be reached in the 5th cutting in year 60. Black: residual, grey: removed trees
In a two-layered stand the optimal management involves removing the over-storey in one or several cuttings (in two cuttings in the example shown in Fig. 3.12). The removal should be started with the largest trees, and pulp-wood sized trees should be left to continue growing until they are log-sized. By the time when all over-storey trees have been removed, the diameter distribution of the under-storey has already reached a near-optimal post-thinning distribution of a steady-state stand.
In young even-aged stands it is optimal to wait until a fair removal of log-sized trees can be obtained. From this moment onwards, the stand is managed with high thinnings that soon convert the stand into the optimal steady state structure.
Also Tahvonen et~al. (2010) optimized the future management of different initial stands. The optimization problem was solved in its most general dynamic form. When NPV was maximized, uneven-aged management was the choice in almost all cases. The only exception was a mature even-aged stand with a low discount rate. In this stand, which had passed the stage of financial maturity, the optimal management involved immediate clear cutting and regeneration. However, the next tree generation should be managed as an uneven-aged forest.
4.5 Temporal Variation of Harvests
Some studies (e.g., Haight et al. 1985) suggest that the profitability of uneven-aged management may improve if the stand density is varied in successive cuttings. There may be a very strong cutting every now and then, resulting in good regeneration, followed by a period of light cuttings with small removals. Then the normal cutting level is used, followed again by a strong regenerative cutting. This kind of periodical uneven-aged management may be optimized by replacing the single steady-state post-thinning diameter distribution by a series of successive distributions, which are repeated with a longer cycle. For example, if the normal cutting cycle is 20 years, four successive distributions may be optimized, representing the remaining frequencies of diameter classes in the first, second, third and fourth thinning, conducted after 0, 20, 40 and 60 years. The fifth thinning, after 80 years, returns the first diameter distribution and begins a new 80-year cycle. The initial investment (A T in Eq. 3.1) is equal to the value of the first post-thinning stand. The net incomes from the entire 80-year period need to be calculated and discounted to the first year to obtain the periodical NPV, which is converted to the NPV of an infinite series of 80-year periods.
To analyze the effect of temporal variation in stand structure, we replaced the single post-thinning diameter distribution by a sequence of 2–6 distributions. The analyses were done for South Finland for a herb-rich spruce forest (OMT), sub-xeric pine forest (VT), and xeric pine forest (CT). The cutting cycle was 15 years in OMT, 20 years in VT, and 30 years in CT. These cutting cycles result in high enough cutting removals, 70–110 m3 ha−1.
Optimizing a sequence of distributions improved the NPV of the spruce stand on the herb-rich site by 2–4% (Fig. 3.13). However, differences between successive post-thinning diameter distributions were small. In VT pine, replacing the single post-thinning diameter distribution by a sequence of 2–6 distributions never improved profitability and all distributions were almost similar. The result means that the optimal management does not require temporal variation in post-thinning stand structure.

Net present value in the optimal solution for three growing sites when a sequence of 1–6 post-thinning diameter distributions is optimized. Calculation of the NPV assumes that the cycle of 1–6 diameter distributions is repeated to infinity
The result was different for CT pine, where temporal variation clearly increased the NPV. The increase was the highest, 5–12% when a sequence of 4, 5 or 6 distributions was optimized. All these optimizations involved cutting the stand to a very low basal area of 1.5–3 m2 ha−1 once during a 120–180-year period (Fig. 3.14). All trees larger than 12 cm in dbh were removed in these cuttings and almost all trees 8–12 cm in dbh were also removed. The remaining stand after this cutting was a young uneven-aged seedling-sapling stand. Optimization resulted in a schedule which is between strictly even-aged and strictly uneven-aged management.

Optimal management of a pine stand on a xeric site in southern Finland when the cutting interval is 30 years. The optimal management involves a sequence of 5 post-thinning diameter distributions, which are repeated with a 150-year cycle. The post-thinning stand in year 150 is similar to the initial stand in year 0
In this stand, the NPV was the highest when a sequence of five diameter distributions was optimized. In the optimal schedule, the post-cutting stand of year 90 was a young stand without any large trees. This stand was uniformly and lightly thinned 30-years later, after which three consecutive high thinnings were done (years 150, 30 and 60 in Fig. 3.15).

Optimal sequence of diameter distributions in a pine forest growing on a xeric site in South Finland
The result may be termed as any-aged management according to Haight and Monserud (1990a, b). However, it is still continuous cover management since the site is continuously covered by trees. The optimal solution for CT pine suggests that it is not always enough to optimize a single post-cutting diameter distribution but options allowing temporal variation in stand structure should also be analysed. This is evident when the cuttings of a certain, non-optimal initial stand are optimized (Tahvonen 2009; Tahvonen et~al. 2010). However, also the steady state management may involve a long cycle consisting of shorter and unequal sub-cycles.
This kind of CCF management was optimal in only one out of three cases and maximally 12% better than the optimal one-cycle steady-state management. The conclusion may change if species dynamics and mixed stands were analysed. For example, the optimal management of a fertile site may involve a heavy regeneration cut in a spruce stand so as to increase the regeneration of hardwoods. Then, the largest spruces are removed, the residual stand being a young rather even-aged hardwood stand where spruce begins to enter again as under-storey. The next cuttings gradually decrease the share of hardwoods and convert the stand into a spruce stand. Because regeneration often gets more difficult with increasing spruce dominance, another very strong cutting may be required, which initiates a new hardwood-dominated period.
4.6 Variable-Density Thinning as a Means to Increase Non-timber Benefits
The economically optimal management of an uneven-aged Norway spruce stand growing on a fertile site in South Finland involves cutting all trees larger than 20 cm at 15-year intervals. The newly harvested stand resembles a young sparse forest. The recreationist walking in the forest does not feel like being in a mature forest. The scenic value of the post-cutting forest is not particularly high due to the absence of large trees. The amount of epiphytic lichens, growing on the branches of trees is most probably low due to absence of old trees since it takes time from the lichens to develop (e.g., Johansson 2008). Together with wild berries and mushrooms, lichen constitutes an important food and nest source for several animal species.
One possibility to avoid too sparse post-cutting stands is to divide the uneven-aged forests into segments and harvest only one segment at a time. The densest places with the highest stand volume or greatest number of large trees are harvested first, the second densest places in the next cutting, and so on. A stand having an optimal cutting interval of 20 years may be divided into four segments, each of which is treated at 20-year intervals. Thus, the stand will be visited at 5-year intervals. With two segments, the stand needs to be visited at 10-year intervals and with tree segments at 6–7-year intervals.
This kind of management results in spatially heterogeneous stands. The temporal continuity of large trees is better since a newly cut stand has large trees in the uncut segments. This is an advantage for both scenic amenity and biodiversity maintenance. Many real stands already are spatially heterogeneous with dense and sparse places, and it is logical to restrict the cutting to the densest places. If the post-thinning density of the treated places is lower than the stand density of the non-treated places, it is equally logical to skip the newly thinned places in the next cutting. In this way, the spatial heterogeneity will be maintained in the future.
When using cutting segments the removals are concentrated on smaller areas with a consequence that the visual impact of cutting and the rate of logging injuries may be smaller than when treating the whole stand simultaneously. If the optimal interval between the cuttings of the same place is longer than it would be without segmentation, the removal per hectare and the mean size of harvested trees will be larger, too. This will result in cheaper harvesting. The optimal post-thinning density of the zone may be lower than the optimal density without segmentation, which may have a favourable effect on regeneration. On the other hand, the stand needs to be visited more often, which increases the harvesting cost.
This section explains a simple way of optimizing the management of a segmented stand, in which the steady-state management is optimized using only one post-tinning diameter distribution (per species). The distribution is defined by the same variables as in previous examples (N, bW, cW and Dmax). Therefore, the number of optimized variables is the same as in a non-segmented forest, but simulation is slightly more complicated. With three cutting segments and 15-year cutting cycle, the simulation is now done as follows:
-
1.
Using N, bW, cW, and Dmax, generate the post-thinning stand for cutting segment 1
-
2.
Simulate the development of segment 1 for 5 years
-
3.
Generate the post-thinning stand for segment 2
-
4.
Simulate the development of segment 1 for 5 years (10 years from beginning)
-
5.
Simulate the development of segment 2 for 5 years (5 years from beginning)
-
6.
Generate the post-thinning stand for segment 3
-
7.
Simulate the development of segment 1 for 5 years (15 years from beginning)
-
8.
Simulate the development of segment 2 for 5 years (10 years from beginning)
-
9.
Simulate the development of segment 3 for 5 years (5 years from beginning)
-
10.
Reduce the frequencies of diameter classes of segment 1 to their initial values; calculate the per hectare removal, roadside value of removed trees, harvesting costs, and net income of cutting: divide the net income by 3 to obtain the per hectare net income for the entire stand
-
11.
Calculate the value of the post-thinning stand using all three segments
-
12.
Calculate the NPV of the schedule
The post-thinning structure of the entire forest consisting of three segments is found in Step 6 after simulating the development of an initial diameter distribution for 0, 5 and 10 years. Another 5-year time step produces the pre-thinning structure of the entire forest (Step 9). Reducing the frequencies of diameter classes to their initial levels in the densest segment gives the harvest removal and periodical net income, and returns the forest into a post-thinning state (Step 10).
This type of simulation assumes that the stand dynamics of a segment do not depend on adjacent segments; the simulation is “distance-independent” although the management of spatially heterogeneous forest is optimized.
We have optimized the management of segmented spruce stands growing on a fertile herb-rich site in Central Finland with several combinations of segments and cutting cycles (Pukkala et~al. 2011d). All optimizations have been done also by using a modified objective function, which includes a penalty if the post-thinning stand includes too few large trees; if the number of trees with dbh between 20 and 30 cm was less than 10 ha−1 a penalty of 10,000 € ha−1 was paid in the cutting year. Another 10,000 € ha−1 was paid if the number of trees with dbh > 30 cm was less than 5 ha−1. The post-thinning number of large trees was counted within the whole stand (not only in the thinned segment).
According to the optimizations, segmentation decreased net present values when the cutting cycle was short. The cost of visiting the stand more often was higher than the benefits obtained from segmentation. However, when lack of large trees was penalized spatially heterogeneous stand was always more profitable (Fig. 3.16). The optimal cutting cycle of a segment became longer when the penalty for lack of large trees was added to the objective function, 20–30-year cutting cycles being optimal instead of the 15-year cycle which was optimal without the penalty.

Net present value (NPV) of the optimal management schedule with the requirement of having large trees continuously present in the stand
The requirement for a continuous presence of large trees always reduced profitability. Without segmentation the decrease was 20–30% depending on the cutting cycle. However, segmentation greatly reduced the cost of maintaining large trees; with cutting cycles of 20 years or more, the reduction was less than 5%.
These optimizations suggest that, when economic profitability should be combined with non-timber services, segmentation with spatially heterogeneous management and stand structure should be pursued. Even without the requirement for large trees, segmentation leads to wider post-thinning diameter distributions, continuous presence of larger trees, and smaller temporal variation in the forest. In addition, a spatially heterogeneous stand may be experienced more interesting by people who visit the forest.
Segmentation is not much different from having smaller stands. However, if the segments are small enough or narrow enough, they may be impractical as stands. In the calculations described above, it was assumed that the reason behind segmentation is reduced temporal variation in stand characteristics and better continuity of large standing trees, so as to improve diversity and amenity. When designing the width and size of the zones, the essential thing is how people and animal species experience the cut and un-cut segments. If the segments are so narrow that large trees in the uncut segments are perceived as belonging to the same stand as the cut places, the use of neighbouring uncut segments as providers of large trees is justified. If people or animal species experience the segmented forest as a single mature uneven-aged forest, segmentation is beneficial. For practical reasons, the minimum width of the cutting segment should be equal to the distance between adjacent extraction roads, i.e., 20–30 m. If the narrow cutting segments are not straight, it is unlikely that they are perceived as distinct stands, especially if the thinning intensity within a cutting zone is not uniform. To minimize harvesting injuries, the best option is to harvest most near the extraction road and reduce the thinning intensity gradually with increasing distance from the strip road. Trees that are difficult to fell without causing logging injuries could be left as retention trees.
Another advantage of stand heterogeneity is high flexibility in marketing; the owner can harvest when the price is good but she is not forced to do that. In a uniform stand, especially in even-aged forestry, management is more tightly related to the stage of stand development; timber sales are not possible during some stages whereas cutting is almost obligatory at another stages. This reduces the possibilities to adjust timber sales according to the market situation.
5 Multifunctional Management
This section analyzes the economic performance of alternative management systems when non-timber benefits are added to the landowner’s objective function. The non-timber benefits are wild berries and carbon sequestration. Biodiversity maintenance and scenic beauty are also briefly discussed.
5.1 Methods
In the optimization of multifunctional management the objective function may be a formula that calculates the net present value obtained from several products and services (Faustmann or Hartman formula). Another possibility is to use a utility function. A third option is to employ Hartman formula or utility function augmented with a penalty function. It is also possible to have NPV as a component in a utility function.
If the non-timber products and services can be expressed in monetary units it is logical to convert all benefits into net incomes and discount them to the present. Examples of products and services that can be measured in monetary units are berry yields, mushroom yields and carbon sequestration. We have analysed the optimal management and profitability of even- and uneven-aged management of pine and spruce stands when timber, bilberry (Vaccinium myrtillus L.) and carbon benefits are all expressed in monetary units and included in the analysis (Pukkala et~al. 2011c). Both of the stands were growing in Central Finland, spruce on mesic site and pine on sub-xeric site.
The discounted bilberry and carbon benefits were first calculated for a cutting cycle as follows:
where B t is the value of bilberry harvest in year t, and C t is the value of carbon sequestration in year t. If the carbon balance is negative, C t is also negative (carbon release tax is paid). The NPV of a cycle was then converted into the NPV of an infinite series of cutting cycles:
The total NPV of uneven-aged management was computed from
with
where N T is the net income obtained regularly at T-year intervals (after T, 2T, 3T,… years), T is the cutting cycle, i is discount rate and A T is the stumpage value of the post-thinning stand.
The empirical bilberry yield models of Miina et~al. (2009) modified for uneven-aged stands (Pukkala et~al. 2011c) were used to predict the bilberry production along with stand development. The models first predict the coverage of bilberry and then the annual berry yield as a function of stand characteristics and bilberry coverage. The bilberry yield predictions were multiplied by 0.75 assuming that 75% of the total yield of the season is actually harvested (Raatikainen and Niemelä 1983). Bilberry price was taken as 3 € kg−1. In 2008 the average market price paid to pickers in eastern Finland was 1.9 € kg−1 for non-cleaned and 4.6 € kg−1 for cleaned bilberries (Miina et~al. 2010).
The annual carbon balance was calculated based on Romero et~al. (1998) and Díaz-Balteiro and Romero (2003):
where C is the annual carbon balance, a is carbon content of biomass (proportion of carbon of dry mass), ΔB is the annual change in biomass, H is the biomass of annual harvest, R is the biomass of new cutting residues, D is the biomass of annual mortality, HR are harvesting releases, PR are processing releases, S are substitution effects, ΔP is the decomposition (decrease of dry mass) of earlier products, ΔR is the decomposition of earlier residues, and ΔD is the decomposition of earlier deadwood. All biomasses are in tons of dry mass per hectare and C, HR, PR and S are in tons of carbon per hectare. Stumps and roots were included in the biomass of living trees, cutting residues and dead trees.
Decomposition of different components of dead trees, cutting residues, and harvested trees was simulated using
where B t is the remaining dry mass after t years, B 0 is the initial dry mass and k is the annual decomposition rate. Parameter k values for branches and needles were based on literature (e.g., Hyvönen et~al. 2000). For stems and stumps (including roots), the annual decomposition rate depended on tree species and breast height diameter of the tree (Pukkala 2006).
The sawlog part of each cut tree was divided into four end product classes: long-term product (sawn wood and plywood), mechanical mass, chemical mass and biofuel (e.g., Karjalainen et~al. 1994; Liski et~al. 2001). The pulpwood part was divided into three classes: mechanical mass, chemical mass and biofuel. The product classes, their proportions and decomposition rates were adapted from Karjalainen et~al. (1994) and Liski et~al. (2001). The decomposition rates of products were based on the estimated time-spans during which half of the mass is decomposed (Liski et~al. 2001). This time was taken as 50 years for sawn wood and plywood, 3 years for mechanical and chemical mass, and 1 year for biofuel.
Harvesting emissions were calculated from
where HR% is harvesting release (released carbon in percent of carbon contained in harvested wood) and Dq is the quadratic mean diameter of removed trees (cm). Biofuel collection rate was taken as 0.67 (2/3 of the biomass of branches is collected and 1/3 remains in the forest) which corresponds to the current efficiency. Stumps and roots were not collected for biofuel. The price of carbon dioxide (tax or subsidy) was assumed to be 15 € ton−1 (http://www.carbonpoint.com), which corresponds to a carbon price of 55 € ton−1.
The decomposition simulator was initialized with near optimal steady-state amounts of decomposing materials in the beginning of a rotation or cutting cycle. The near optimal values were obtained from preliminary optimizations. To guarantee that the carbon dynamics represented a steady state, three rotations were always simulated in even-aged management and four cutting cycles were simulated in uneven-aged management. The annual carbon balances were calculated from the last rotation or cycle.
5.2 Results
The optimal uneven-aged management of the spruce stand yielded the highest NPV of timber benefits, the highest NPV of carbon benefits, and the highest NPV of bilberry benefits (Fig. 3.17). In the pine stand, the optimal even-aged management was better in terms on carbon benefits but uneven-aged management was clearly better in terms of timber and bilberry benefits. The currently recommended even-aged management was much worse than the optimized systems. The main differences between the current and optimal even-aged management systems are that the optimal management postpones the first commercial thinning, uses longer rotation lengths, and applies high thinning instead of the currently favoured low thinning. Summing the net present values of timber production, bilberry collection, and carbon sequestration reveals that the optimal uneven-aged management was by far the best in both species, and the current even-aged management system was by far the worst.

Net present value of different benefits in the optimal uneven-aged, optimal even-aged, and current even-aged management. Discount rate is 3%, bilberry price is 3 € kg−1, and carbon dioxide price is 15 € ton−1. In the pine stand, the NPV of carbon benefits is close to zero in the optimal uneven-aged management
The low net present value of even-aged management, as compared to uneven-aged management, is partly explained by the unfavourable temporal distribution of costs and incomes. Even-aged management has early stand establishment and tending costs but the first incomes are obtained only after 45–50 years since planting. The first incomes of uneven-aged management are obtained after one cutting cycle, i.e., after 20 years. The optimal rotation lengths of the multifunctional forestry were 15–25 years longer than currently recommended or obtained when only timber production is optimized. Therefore, considering carbon sequestration and bilberry production in forestry decision-making leads to the use of longer rotations.
The timber yields were of the same magnitude in all three management systems (Fig. 3.18). The current even-aged management system yielded most timber in the spruce stand, and the optimal uneven-aged management was the most productive in pine. In the spruce stand, the saw log yields were nearly the same in all three management systems but pulpwood and biofuel harvests were higher in the currently recommended even-aged management schedule. Optimal uneven-aged management was the best in terms of carbon sequestration and bilberry yield, in both spruce and pine (Fig. 3.18). With the used substitution rates (0.2 for sawn wood and 0.4 for biofuel), spruce forestry was always a carbon source whereas pine forestry was a carbon sink.

Mean annual wood harvest, carbon balance and bilberry harvest in the optimal uneven-aged, optimal even-aged, and current even-aged management. Discount rate is 3%, bilberry price is 3 € kg−1, and carbon dioxide price is 15 € ton−1. The dry mass of branches collected for biofuel has been converted into cubic metre equivalents and included in the “Fuel” assortment. “Fuel” includes the tops and 2/3 of branches of cut trees
In spruce, the bilberry harvest was very sensitive to the management system so that the optimal uneven-aged management yielded five times more bilberries than the current even-aged management system. The density of the uneven-aged spruce stand was almost constantly near the optimal value for bilberry. Differences in bilberry harvest between the three management systems were small in pine stand.
The carbon balance of the first years of an even-aged rotation is very negative because there are plenty of decomposing cutting residues, stumps and roots, as well as products with short life spans (biofuel and pulp) from the previous clear-felling. The annual balance turns positive after about 30 years (the lines representing accumulated carbon in Fig. 3.19 start to ascend) since most of the fast-decomposing materials have already decomposed and carbon sequestration into new biomass is fast. However, due to discounting, the negative carbon balances of the first years have a much stronger influence on the NPV than the positive balances of later years.

Temporal development of accumulated carbon balance in the optimal uneven-aged, optimal even-aged, and current even-aged management. Discounting rate is 3%, bilberry price is 3 € kg−1, and carbon dioxide price is 15 € ton−1
The NPV of the carbon benefits is much better for the optimal even-aged management schedule than for the currently recommended schedule. This is because the optimal even-aged management employs heavy high thinnings during the latter half of the rotation, which decreases the clear-felling removal. As a consequence, much decomposition and carbon releases occur at the end of the rotation when the cutting residues and short-term products of the thinnings decompose, leaving less decomposing material to the first years of the next rotation.
Other important services of Finnish forests are biodiversity maintenance and scenic values. The most recent models for the landscape preferences of Finns are those of Silvennoinen et~al. (2001), which predict the scenic quality of a forest stand as a function of mean tree height, skewness of the diameter distribution, number of trees per hectare, volume of pine, and volume of birch. Figure 3.20 shows the scenic beauty index of Silvennoinen et~al. (2001) for the stands and management systems analyzed in this section. It can be seen that uneven-aged and mature even-aged stands are close to each other in terms of scenic beauty, but a young even-aged forest is clearly inferior. Therefore, the long-term mean scenic value is better for uneven-aged management.

Several indices have been proposed for describing the diversity of a forest stand in Finland (e.g., Lähde et~al. 1999b; Pukkala 2006; Koskela et~al. 2007). Their idea is to measure the number of different structural elements present in the stand such as tree species, tree sizes, and types of deadwood. The indices are based on the assumption that the number of species that can live in the stand increases with an increasing number of structural elements. The LLNS index of Lähde et~al. (1999b) was calculated for the three management systems compared in this section (Fig. 3.20). The results suggest that an uneven-aged stand is equal to a mature even-aged stand in terms of structural diversity, but clearly better than young even-aged stands. Taking into account that small temporal variation in the amounts of structural elements is an advantage, the results in Fig. 3.20 suggest that uneven-aged management is better than even-aged forestry also in terms of structural diversity.
The analysis suggests that, in Finnish forests, CCF is better than even-aged management for a simultaneous production of multiple benefits. It is evident that the higher is the number of forest functions included in the analysis the clearer is the superiority of CCF management. However, at the landscape level, RFM and clear-felling may be used to improve the visibility of distant vistas, and to increase the number of different habitat types. In addition, there are some forest products such as cowberry (Vaccinium vitis-ideae L.) and some mushrooms, which benefit from clear-felling. Therefore, forest management which is optimal at the landscape level would most probably use both even- and uneven-aged management. This is in accordance with the natural disturbance dynamics of Fennoscandian forests, which includes mainly small disturbances (trees and tree groups destroyed) with large disturbances (clear felling) occurring every now and then (Keto-Tokoi and Kuuluvainen 2010).
6 Discussion
Most analyses conducted so far in Finland and described in this chapter suggest that CCF is more profitable to the forest landowner than the currently recommended RFM (Tahvonen et~al. 2010; Pukkala et~al. 2010, 2011c). The relative superiority of CCF management improves with increasing discount rate and management costs, and with decreasing site productivity and timber price (Tahvonen 2009). The low profitability of RFM is mainly because of too high stand establishment cost; the low timber yields of most Finnish forests do not warrant high management costs. More extensive management with less human intervention remains the only viable option on poor growing sites, peatland sites and North Finland.
The Finnish results may appear different from some Swedish and Norwegian studies. In Sweden, Wikström (2000) obtained higher NPVs for RFM than for CCF. However, Wikström himself (2001) reminds that his results cannot be used to compare the profitability of RFM and CCF because the growth of uneven-aged stands was underestimated and ingrowth was an arbitrarily set constant. In addition, calculations were done for financially mature stands for which immediate regeneration would be optimal. However, this treatment was prevented by the constraints of the optimization problems. As a result, the uneven-aged stands were constantly too old and dense.
Andreassen and Øyen (2001) found clear felling and planting to be more profitable than selective cutting in financially mature spruce stands in Norway. Similarly to Wikström (2000), the optimal CCF-alternative, i.e. immediate natural regeneration followed by normal uneven-aged management, was not included in the analysis. In addition, the growth predictions were multiplied by 0.85 in CCF. Therefore, these Swedish and Norwegian studies only show that financially mature stands should not be treated with repeated light thinnings; they should be rejuvenated by using heavy regenerative cuttings. The studies do not show that optimal RFM is more profitable than optimal CCF.
RFM can be made profitable to forest landowner by state subsidies, and this is what is currently done in Finland. Non-profitable silviculture is recommended, and it is made profitable again with the help of subsidies. However, the society would gain more by withdrawing these subsidies and promoting economically more viable management. To promote socially optimal forest management the beneficial externalities of timber production could be subsidized instead. As explained in this chapter, the most profitable management would be continuous cover forestry.
When evaluated from the multiple-use and multi-functional forestry perspectives the superiority of CCF increases. This is mainly because of the poor performance of RFM during the first decades after clear-felling. Taking into account the changing uses of Finnish forests, with decreasing importance of timber production and increasing importance of non-timber benefits, it is evident that alternatives to the current even-aged silviculture should be studied and promoted.

Clear-felled fertile sites regenerate naturally for hardwood and spruce regenerates easily under the hardwood cover. One management option is to produce hardwood biomass for biofuel at first, and then convert the stand to a mixed uneven-sized stand, which is managed with repeated high thinnings (Photo: Olavi Laiho).
From the wood production point of view, the demand for pulpwood is slowly decreasing and the demand for biofuel is increasing. Therefore, the two principal future timber assortments may be saw log and biofuel, instead of saw log and pulp wood. This change improves the relative value of hardwoods as compared especially to spruce pulpwood. At the moment, small-sized spruce timber is clearly more valuable than small-sized hardwood but the order may soon change since the energy content of hardwood is higher than that of conifer wood. As a consequence, the prevailing practice of planting spruce on fertile sites becomes questionable since most cutting areas would regenerate naturally with different hardwood species. This regeneration could be used as biofuel when the trees are large enough for a cheap harvesting operation.
Since spruce will enter the hardwood stand as an under-storey, spruce timber will be obtained later on without any cost. The two-layered mixed stand could be managed with repeated high thinnings, as long as regeneration and ingrowth continue. On some sites, such as most peat lands and fertile moist sites, regeneration is always sufficient for CCF. In the other cases, heavier cuttings are required every now and then to enhance regeneration and re-initiate the natural species cycle. This type of CCF management follows the natural species dynamics, is cheaper, and produces assortments that are required in the future. The hardwood stage also prevents spruce butt rot from spreading to the new spruce generation, since the rot will disappear from the root systems of rotten spruce stumps during the hardwood phase.
On poorer sites, where only pine grows well, CCF requires constant regeneration and ingrowth of pine. Group selection may be used and small canopy gaps created if regeneration is otherwise insufficient. It is also possible to use variable-density thinning as well as partial shelter wood and seed tree cuttings to promote regeneration.
Improved management of Finnish forests requires experimentation with different variants of CCF. The new experiments mentioned in Sect. 2 that mimic the natural disturbance regimes occurring on different scales will provide information for a continuum of forest management options. However, additional experiments, in which strip roads are an integral part of the disturbance dynamics, are required. Extraction roads may be enlarged into small gaps, or combined with spatially uneven thinning intensity where places adjacent to strip roads are thinned more heavily. Alternatives involving partial seed tree and shelter wood cuttings should also be studied in field experiments. More growing sites with different soil properties and moisture conditions should be included in the experiments to learn about the effect of site characteristics on the regeneration dynamics.
In the management problems analyzed in this chapter, a simple steady-state management problem of a pure one-species stand was solved first. This involved finding the optimal combination of only four decision variables (bW, cW, N, Dmax) and simulating the stand development for one cutting cycle. The problem was gradually extended to involve the penalty for lack of large trees, steady-state management of mixed stands, transformation cuttings, and steady-state management in which a long management cycle consisted of a sequence of shorter cutting cycles. The joint production of several benefits was analyzed and optimized in the last example.

Extraction roads are good places for regeneration (Photo: Timo Pukkala)
Although many types of optimization have already been done in Finland, and can be made with the current models, most analyses conducted so far should be taken as the first tries and preliminary calculations. For example, the post-thinning diameter distribution was described with a Weibull function in almost all optimizations described in this chapter. This may be sufficient for the steady state management of a pure one-species stand, but it may not be a justified simplification in mixed stands or when a sequence of distributions is optimized. The intervals between cuttings may be optimized simultaneously with the diameter distributions. When conversion cuttings are optimized, a better option would be to not fix the length of the transformation period or the structure of the ending forest (Tahvonen et~al. 2010). For the near future, optimizations with higher biofuel and saw log price, and lower pulpwood price should also be conducted. Involving the species dynamics in the optimization of CCF management also needs much attention in future studies.

Group selection in a pine stand. Pine regenerates in canopy gaps and in heavily thinned places (Photo: Olavi Laiho)
Some of the optimization problems described in this chapter are spatial. In fact, the problem is always spatial when extraction roads are an integral part of the management system. This is the case especially in CCF where extraction roads and their vicinity are the best places for regeneration. The most convenient way to analyze spatial problems is to use distance-dependent growth models and spatial optimization. At the moment, there are already distance-dependent regeneration models for uneven-aged spruce stands, which predict the amount, survival and growth of regeneration in 4-m2 cells as a function of over-storey density within and around the cell (Eerikäinen et~al. 2007). This model, combined with distance-dependent simulation of the dynamics of larger trees, offers interesting possibilities to analyze spatially non-homogenous management systems.
Improved optimization techniques and economic analyses need improved modeling. The weakest parts in the current models for uneven-aged forests are the regeneration and ingrowth models. It is known that spruce regenerates more easily under pines and birches than under spruces, but this type of species interaction is not included in the models. Therefore, a necessary modeling task before doing more ambitious optimizations for mixed stands, are regeneration or ingrowth modeling for describing the influence that different tree species may have on the regeneration of other species. Also the inter-specific competition effects of larger trees should be modeled better than done so far. The most interesting, most challenging and most useful future research topic would be to include the species dynamics in the analysis of CCF management. This would require new field experimentation, new modeling efforts and new optimization tools.
References
Andreassen K, Øyen B-H (2001) Economic consequences of three silvicultural methods in uneven-aged mature coastal spruce forests of central Norway. Forestry 75:483–488
Chang SJ (1981) Determination of the optimal growing stock and cutting cycle for an uneven-aged stand. Forest Sci 27(4):739–744
Chang SJ, Gadow Kv (2010) Application of the generalized Faustmann model to uneven-aged forest management. J Forest Econ 16(2010):313–325
Díaz-Balteiro L, Romero C (2003) Forest management optimisation models when carbon captured is considered: a goal programming approach. Forest Ecol Manage 174:447–457
Eerikäinen K, Miina J, Valkonen S (2007) Models for the regeneration establishment and the development of established seedlings in uneven-aged, Norway spruce dominated stands of southern Finland. Forest Ecol Manage 242:444–461
Haight RG (1985) A comparison of dynamic and static economic models of uneven-aged stand management. Forest Sci 31(4):957–974
Haight RG, Monserud RA (1990a) Optimizing any-aged management of mixed-species stands: I. Performance of a coordinate search process. Can J Forest Res 20:15–25
Haight RG, Monserud RA (1990b) Optimizing any-aged management of mixed-species stands: II. Effect of decision criteria. Forest Sci 36(1):125–144
Haight RG, Brodie JD, Adams DM (1985) Optimizing the sequence of diameter distributions and selection harvests for uneven-aged stand management. Forest Sci 31(2):451–462
Hyvönen R, Olsson BA, Lundqvist H, Staaf H (2000) Decomposition and nutrient release from Picea abies (L.) Karst. and Pinus sylvestris L. logging residues. Forest Ecol Manage 126: 97–112
Hyytiäinen K, Tahvonen O (2003) Maximum sustained yield, forest rent of Faustmann: does it really matter? Scand J Forest Res 18:457–469
Johansson P (2008) Consequences of disturbance on epiphytic lichens in boreal and near boreal forests. Biol Conserv 141:1933–1944
Karjalainen T, Kellomäki S, Pussinen A (1994) Role of wood-based products in absorbing atmospheric carbon. Silva Fennica 28(2):67–80
Keto-Tokoi P, Kuuluvainen T (2010) Suomalainen aarniometsä. Maahenki, Kariston kirjapaino, Hämeenlinna, 302 pp
Kolström T (1993) Modelling the development of an uneven-aged stand of Picea abies. Scand J Forest Res 8:373–383
Koskela E, Ollikainen M, Pukkala T (2007) Biodiversity conservation in commercial boreal forestry: optimal rotation age and volume of retention. Forest Sci 53(3):443–452
Lähde E (1992a) Natural regeneration of all-sized spruce-dominated stands. In: Hagner M (ed) Silvicultural alternatives, Proceedings from an internordic workshop, June 22–25 1992. The Swedish University of Agricultural Sciences, Department of Silviculture, Reports 35, pp 111–116
Lähde E (1992b) Natural regeneration of all-sized spruce-dominated stands treated by single tree selection. In: Hagner M (ed) Silvicultural alternatives, Proceedings from an internordic workshop, June 22–25 1992. The Swedish University of Agricultural Sciences, Department of Silviculture, Reports 35, pp 117–123
Lähde E, Laiho O, Norokorpi Y, Saksa T (1991) The structure of advanced virgin forests in Finland. Scand J Forest Res 6:527–537
Lähde E, Laiho O, Norokorpi Y, Saksa T (1992a) Alternative silvicultural treatments as applied to advanced stands – research plan. In: Hagner M (ed) Silvicultural alternatives, Proceedings from an internordic workshop, June 22–25 1992. The Swedish University of Agricultural Sciences, Department of Silviculture, Reports 35, pp 66–73
Lähde E, Laiho O, Norokorpi Y, Saksa T (1992b) Stand structure of thinning and mature conifer-dominated forests in boreal zone. In: Hagner M (ed) Silvicultural alternatives, Proceedings from an internordic workshop, June 22–25 1992. The Swedish University of Agricultural Sciences, Department of Silviculture, Reports 35, pp 58–65
Lähde E, Laiho O, Norokorpi Y, Saksa T (1994a) Structure and yield of all-sized and even-sized conifer-dominated stands on fertile sites. Ann Sci Forestiéres 51(2):97–109
Lähde E, Laiho O, Norokorpi Y, Saksa T (1994b) Structure and yield of all-sized and even-sized Scots pine-dominated stands. Ann Sci Forestiéres 51(2):111–120
Lähde E, Laiho O, Norokorpi Y (1999a) Diversity-oriented silviculture in the Boreal Zone of Europe. Forest Ecol Manage 118:223–243
Lähde E, Laiho O, Norokorpi Y (1999b) Stand structure as the basis of diversity index. Forest Ecol Manage 115:213–220
Lähde E, Laiho, Norokorpi, Y (1999c) Ekometsänhoidon perusteet ja mallit. The Finnish Forest Research Institute. Research Notes 736: 1–61
Lähde E, Laiho O, Norokorpi Y, Saksa T (2001) Structure transformation and volume increment in Norway spruce-dominated forests following contrasting silvicultural treatments. Forest Ecol Manage 151:133–138
Lähde E, Laiho O, Norokorpi Y, Saksa T (2002a) Development of Norway spruce dominated stands after single-tree selection and low thinning. Can J Forest Res 32:1577–1584
Lähde E, Eskelinen T, Väänänen A (2002b) Growth and diversity effects of silvicultural alternatives on an old-growth forest in Finland. Forestry 75:395–400
Lähde E, Laiho O, Norokorpi Y, Saksa T (2002c) Zuwachs von Fichte und Birke in gleich und ungleich strukturierten Beständen. Forst und Holz 57(15/16):481–485
Liski J, Pussinen A, Pingoud K, Mäkipää R, Karjalainen T (2001) Which rotation length is favourable to carbon sequestration? Can J Forest Res 31:2004–2013
Miina J, Hotanen J-P, Salo K (2009) Modelling the abundance and temporal variation in the production of bilberry (Vaccinium myrtillus L.) in Finnish mineral soil forests. Silva Fennica 43:577–593
Miina J, Pukkala T, Hotanen J-P, Salo K (2010) Optimizing the joint production of timber and bilberries. Forest Ecol Manage 259:2065–2071
Pukkala T (2005) Metsikön tuottoarvon ennustemallit kivennäismaan männiköille, kuusikoille ja rauduskoivikoillle. Metsätieteen aikakauskirja 3(2005):311–322
Pukkala T (2006) Optimising the semi-continuous cover forestry of Finland. Allgemaine Forst und Jagd Zeitung 1677:141–149
Pukkala T (2009) Population-based methods in the optimization of stand management. Silva Fennica 43(2):261–274
Pukkala T, Kolström T (1988) Simulating the development of Norway spruce stands using transition matrix. Forest Ecol Manage 25:255–267
Pukkala T, Lähde E, Laiho O (2009) Growth and yield models for uneven-sized forest stands in Finland. Forest Ecol Manage 258:207–216
Pukkala T, Lähde E, Laiho O (2010) Optimizing the structure and management of uneven-sized stands of Finland. Forestry 83:129–142
Pukkala T, Lähde E, Laiho O (2011a) Metsän jatkuva kasvatus. Joen Forest Program Consulting, Joensuu, 229 pp
Pukkala T, Lähde E, Laiho O (2011b) Using optimization for fitting individual-tree growth models for uneven-aged stands. Eur J Forest Res 130: 829–839
Pukkala T, Lähde E, Laiho O, Salo K, Hotanen J-P (2011c) A multifunctional comparison of even-aged and uneven-aged forest management in a boreal region. Can J Forest Res 41:851–862
Pukkala T, Lähde E, Laiho O (2011d) Variable-density thinning in uneven-aged forest management – a case for Norway spruce in Finland. Forestry DOI:10.1093/forestry/cpr020
Raatikainen M, Niemelä M (1983) Mustikan poimintatarkkuuden määrittäminen. University of Jyväskylä, Department of Biology. Res Notes 32:18–23
Romero C, Rios V, Díaz-Balteiro L (1998) Optimal forest rotation age when carbon captured is considered: Theory and applications. J Oper Res Soc 49:121–131
Rummukainen A, Alanne H, Mikkonen E (1995) Wood procurement in the pressure of change – resource evaluation model till year 2010. Acta Forestalia Fennica 248:98
Saksa T, Lähde E, Laiho O, Norokorpi Y (2003) Entwicklung und Leistung von Beständen mit vorherrschender Fichte nach Niederdurchforstung und Einzelstamm-Auslese. Forst und Holz 58(8):220–222
Schulte BJ, Buongiorno J, Skog K (1999) Optimizing uneven-aged management of loblolly pine stands. Proceedings of the Society of American Foresters. 1998 National Convention, Traverse City, Michigan, September 19–23, 1998, pp 306–318
Siiskonen H (2007) The conflict between traditional and scientific forest management in 20th century Finland. Forest Ecol Manage 249:125–133
Silvennoinen H, Alho J, Kolehmainen O, Pukkala T (2001) Prediction models of landscape preferences at the forest stand level. Landsc Urban Plann 56(1–2):11–20
Tahvonen O (2009) Optimal choice between even- and uneven-aged forestry. Nat Resources Model 22:289–321
Tahvonen O, Pukkala T, Laiho O, Lähde E, Niinimäki S (2010) Optimal management of uneven-aged Norway spruce stands. Forest Ecol Manage 260:106–115
University of Helsinki (2010) Forest Research News 1/2010 (http://www.helsinki.fi/metsatieteet/laitos/ uutiskirjeet_pdf/2010_1_For_Res_News_Univ_Helsinki.pdf). 3 pp
Valkeapää A, Paloniemi R, Vainio A, Vehkalahti K, Helkama K, Karppinen H, Kuuluvainen J, Ojala A, Rantala T, Rekola M (2009) Suomen metsät ja metsäpolitiikka – kansalaisten näkemyksiä. University of Helsinki, Department of Forest Economics. Reports 55, 36 p
Valkonen S, Sirén M, Piri T (2010) Poiminta- ja pienaukkohakkuut – vaihtoehtoja avohakkuulle. Metsäkustannus Oy, Tampere. 125 pp
Wikström P (2000) A solution method for uneven-aged management applied for Norway spruce. Forest Sci 46(3):452–463
Wikström P (2001) Effect of decision variable definition and data aggregation on a search process applied to a single-tree simulator. Can J Forest Res 31:1057–1066
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Pukkala, T., Lähde, E., Laiho, O. (2012). Continuous Cover Forestry in Finland – Recent Research Results. In: Pukkala, T., von Gadow, K. (eds) Continuous Cover Forestry. Managing Forest Ecosystems, vol 23. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-2202-6_3
Download citation
DOI: https://doi.org/10.1007/978-94-007-2202-6_3
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-2201-9
Online ISBN: 978-94-007-2202-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)