
Abstract
After reviewing the current knowledge on paleoecological tracking using carbon and nitrogen stable isotopes in fossil bones and teeth, the contribution of this new approach to key questions of Neanderthal diet and ecology is examined. In particular, the determination of ungulate habitat (open environment versus closed forest) is discussed. Thanks to the carbon and nitrogen isotopic differences observed in the main ungulates available as prey to Neanderthal of OIS 3, it is possible to evaluate quantitatively the contribution of different prey in the diet of Neanderthals. The results of this approach suggest that megaherbivores, such as mammoth and woolly rhinoceros, were the most important providers of proteins to the Neanderthal from Saint-Césaire and probably also to one Neanderthal specimen from Spy. In addition, the conclusions of zooarcheological and isotopic dietary determinations are not always in agreement, possibly due to taphonomic biases and site specialization.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
Introduction
One hundred and fifty years after the discovery of the Neanderthal fossils in Feldhofer cave, scholars are still debating about the reasons why this hominid form is not around any longer. Several hypotheses dealing with the extinction of Neanderthals involve the diet and ecology of this hominid. One may think that after one century and a half of scientific studies, these key aspects of the paleobiology of this hominid are reasonably well known. Indeed, an impressive amount of data from various scientific fields, such as paleoanthropology, paleontology, paleobotany, geology, and paleoclimatology has been gathered thanks to the investigations of many research groups around the world. These works lead to interesting hypotheses on the paleobiology of Neanderthals, but the conclusions of these investigations do not always provide a consistent picture of the Neanderthal way of life and many aspects, including diet and ecology, are still controversial. For instance, questions such as: “was the diet of Neanderthals different from the diet of anatomically modern humans?” and, “were Neanderthals more cold adapted than anatomically modern humans?” still remain with no firm and definitive answers (e.g., recent reviews in Churchill 1998; Marean and Assefa 1999; Hockett and Haws 2005; Weaver and Steudel-Numbers 2005; Adler et al. 2006).
One of the main problems is the geographical disjunction between the paleoclimatic indicators, which are mainly recovered from polar icecaps, as well as marine, lacustrine, and loessic sediments, and the sites where the Neanderthal fossils and settlements are found. Potential paleoenvironmental indicators are also found in the sites themselves, in the form of fossil mammal and plant remains and geological features. However, it is generally not straightforward to associate the last two tracers with human occupation, and using fossil mammals as paleoecological tracers is not without some difficulties. Indeed, some of these fossil mammals belong to extant species, such as reindeer, red deer, roe deer and horse, while others belong to extinct species, such as woolly rhinoceros and woolly mammoth. In the case of extant species, the basis of the paleoecological interpretations is taxonomic uniformitarianism, which implies that ancient representatives of a species have the same ecological requirements, such as range of temperature, humidity, and food resources, than the modern representatives of the same species. Such an assumption is a reasonable starting point, but presents some problems if used too rigidly. A first caveat is that the modern geographical and climatic range of the taxa does not extend to the maximum area and climatic conditions which the animals might tolerate. This can be due to recent distribution areas restricted by interfering human impact and to possible changes of physiological limits through time between populations considered to belong to the same species by paleontologists. A second difficulty in this approach is that fossil assemblages are often mixtures of species that do not cohabit in the modern world. Is this due to time averaging of the assemblages, or to the actual existence of “non-analogue communities” (communities including species that have non-overlapping distributions in the modern world), meaning that the ecological requirements of a species can change through time? This question raises interesting implications about the fact that fossil taxa may not be identical to modern ones, as they may represent different genotypic populations (Stewart et al. 2003). In addition, some community inertia has been demonstrated for some terrestrial mammal communities during the Pleistocene (McGill et al. 2006), which makes paleoenvironmental inferences based on community composition even more complex to raise. In the case of extinct species, the reconstruction of their paleoecology is even more complicated since assumptions are based on skeletal remains, sometimes fragmentary, and they are therefore less secure than the conclusions based on extant species. Tracers that would be taxa-independent but environment-dependent and measured on the fossils themselves would greatly improve our knowledge of the paleobiology of Pleistocene mammals relevant to issues related to Neanderthals, especially ancient diets and environments.
In this context a new disciplinary field, based on the isotopic biogeochemistry of vertebrate skeletal fossil tissues, yields new data since about 15 years in situations relevant to the questions of Neanderthal paleobiology (e.g., Bocherens et al. 1991, 1999, 2005b; Fizet et al. 1995). The results of this novel approach, in addition to those of other fields of research, have yielded valuable insights on the diet and ecology of Neanderthals. This approach is based on the fact that during an individual’s life, its tissues, including bones and teeth, incorporate carbon and nitrogen atoms that reflect the isotopic composition of the consumed food (e.g., DeNiro and Epstein 1978, 1981). Since different food items can be distinguished through their carbon (13C/12C) and nitrogen (15N/14N) stable isotopic signatures, information about the diet of fossil hominids can be retrieved (e.g., Drucker and Bocherens 2004; Lee-Thorp and Sponheimer 2006), as well as information about the life environment of animals found in prehistoric sites (e.g., Koch et al. 1989; Bocherens et al. 1994, 1995, 1997, 2006, 2007; Iacumin et al. 1997; Drucker 2005, 2007; Bocherens and Rousseau 2008). The application of this approach requires that diagenetic modifications suffered by the specimen since its burial in the sediment did not change significantly the isotopic signatures recorded during the life period, and that the rules of isotopic variations linked to dietary and environmental factors are reasonably well-known. This paper will review the current status of this field of research, emphasizing the basic rules of application, and describing some of the most important finds so far.
Isotopic Records in Bones and Teeth as a Tracer of Paleobiology: Potential Use and Limitations
Bones and teeth include mineral and organic components, both containing carbon, while only the organic fraction of skeletal tissues contains significant amounts of nitrogen. Both fractions are subject to modifications during the fossilization process, and great care has to be taken before interpreting the isotopic signatures measured on fossil material in paleobiological terms (e.g., DeNiro 1985; Bocherens et al. 2005a; Lee-Thorp and Sponheimer 2006). Carbon in the mineral fraction occurs as carbonate ions incorporated into the bioapatite (calcium phosphate) of bone, dentine and enamel. Although carbon isotopic compositions of carbonates can be measured in almost any fossil bone or tooth, not all values have survived unchanged since the animal’s death. The poorly crystallized minerals of the organic-rich bone and dentine are very sensitive to alteration and generally do not preserve their biogenic isotopic signature for more than a few thousand years contrarily to the highly mineralized enamel crystals which are very resistant to diagenesis and usually preserve their biogenic isotopic signature for several million years (e.g., Lee-Thorp and van der Merwe 1991; Ayliffe et al. 1994; Koch et al. 1997; Lee-Thorp and Sponheimer 2003).
Collagen represents about 25% of the dry weight of fresh bone and dentine and exhibits very invariable chemical characteristics across taxa. Collagen from mammal bones and teeth has a very narrow range of carbon and nitrogen content, a property that is routinely used to monitor the reliability of collagen extracted from fossil bones (e.g., DeNiro 1985; Ambrose 1990). It is widely accepted that any extract with a C/N atomic ratio lower than 2.9 or higher than 3.6 is to be discarded since these values are clearly outside the range exhibited by collagen extracted from fresh bone. However, the lower acceptable limit for carbon and nitrogen percentages in fossil bone extracts is evaluated case by case. Collagen-like extracts with carbon and nitrogen percentages lower than those measured in collagen extracted from fresh bones, i.e. 36% and 12%, respectively (Rodière et al. 1996), should be considered very cautiously, especially if they are outliers compared to other collagen extracts from the same site.
Different techniques can be used to extract collagen from fossil bones (e.g., Longin 1971; Bocherens et al. 1997; Higham et al. 2006), all of them aiming at the elimination of any component other than collagen while losing a minimal proportion of the remaining collagen. The contaminations include the mineral fraction containing carbonate as well as exogenous organic matter that could have impregnated the bone sample during burial. Although collagen remnants have been identified in fossils as old as dinosaurs (e.g., Wyckoff 1969; Bocherens et al. 1988; Ostrom et al. 1993; Asara et al. 2007), most applications of collagen isotopic biogeochemistry have been restricted to the last 200 kyr (Jones et al. 2001) and to material coming from regions with temperate or cold climates since the rate of collagen loss is temperature dependant (e.g., Holmes et al. 2005). Under warmer climatic conditions, as in Mediterranean areas, and in less protected environments, as in open-air sites, bone is usually less well preserved and collagen is often too altered to yield reliable isotopic signatures. These are cases where it is nevertheless possible to measure carbon isotopic signatures in the mineral fraction of tooth enamel (e.g., Bocherens et al. 1995; Lee-Thorp and Sponheimer 2006; Bocherens and Rousseau 2008).
The isotopic analysis of animals raised on controlled diets or monitored in natural environments yielded the general rules of isotopic record in bone collagen and the carbonate fraction of bioapatite. Dietary reconstruction is based on the isotopic composition of carbon (13C/12C) and nitrogen (15N/14N). These ratios are commonly expressed using the “δ” (delta) value as follows: δ13C = [(13C/12C)sample/(13C/12C)reference – 1] × 1,000 (‰) and δ15N = [(15N/14N)sample/(15N/14N)reference – 1] × 1,000 (‰), with the international reference being V-PDB for δ13C values and atmospheric nitrogen (AIR) for δ15N values. Carbon is provided by the whole food consumed by an organism, while nitrogen is provided by the protein fraction of the food. The carbon isotopic signature of an animal is globally equivalent to that of its average food (DeNiro and Epstein 1978). However, different fractions of the body present varying carbon isotopic signatures compared to the whole body. For example, the collagen of an organism raised on a homogenous diet exhibits δ13C values enriched by 5‰ while the carbonate fraction exhibits δ13C values enriched by 9–14‰ relative to the diet and the whole body (e.g., DeNiro and Epstein 1978; Cerling and Harris 1999; Passey et al. 2005). The variations of this enrichment in the carbonate fraction seem to relate to different dietary physiology and are higher in large herbivores than in small herbivores and carnivores (e.g., Passey et al. 2005). These differences may be partly linked to the impact of 13C-enriched CO2 generated through methane production especially high in ruminants (e.g., Hedges 2003; Passey et al. 2005). As a consequence, the difference between the δ13C values of collagen and carbonate bioapatite is higher in herbivores (7.9 ± 1.1%) than in carnivores (4.7 ± 0.4%) (Bocherens 2000). Flesh is slightly enriched compared to the average diet, which is why the δ13C values of the collagen of a predator are enriched by around 1‰ (from 0.8 to 1.3‰) compared to the δ13C values of the collagen of its average prey(Bocherens and Drucker 2003).
Contrary to carbon, the nitrogen-15 content is higher in a given organism than in its food, with the δ15N values of an organism being 3–5‰ more positive than those of its average diet (DeNiro and Epstein 1981; Schoeninger and DeNiro 1984). When the δ15N values of the bone collagen of a predator are compared to those of its average prey, a similar enrichment of 3–5‰ is found (Bocherens and Drucker 2003). Some tentative links have been suggested between some variations in this enrichment factor and physiological or environmental parameters. For instance, the fact that a given herbivorous species exhibits increasing δ15N values with decreasing annual precipitation in Africa and Australia led to the suggestion that aridity would increase the 15N enrichment between plant food and body tissues in herbivores, due to changes in water and nitrogen metabolism under water and dietary stressed conditions (e.g., Ambrose and DeNiro 1986; Heaton et al. 1986; Sealy et al. 1987; Ambrose 1991; Gröcke et al. 1997). However, more recent works based on a larger dataset seem to indicate that the observed increase in δ15N values is due to an increase in the δ15N values of plants with aridity and a relatively constant fraction of nitrogen between plant food and herbivores (Murphy and Bowman 2006). This does not preclude some variability in the trophic enrichment of 15N. For instance, in herbivorous mammals, such variations were observed in controlled feeding experiments of different herbivorous species such as alpaca, cattle, goat, and horse with diets differing in their protein levels (Sponheimer et al. 2003). The extreme fractionation values are, however, obtained with diets exceeding the nitrogen requirements of the animal or leading to starvation and weight loss. Moreover, they were measured on tissues recording short-term variations, such as hair and blood. These results may thus not be relevant for the isotopic record of tissues averaging long periods of life such as bone in animals living under natural conditions. Some differences have also been observed in omnivorous mammals fed on diets with differing protein contents. For instance, a review of fractionation values measured for bears suggests an increasing difference between blood and food δ15N values for bears fed on diets presenting decreasing nitrogen content, such as fish (2.3 and 3.6‰), flesh (3.8 and 4.1‰), and fruits (4.2 and 5.8‰) (Robbins et al. 2005). Such variations may also exist in humans according to their diet quality (review in Hedges and Reynard 2007).
Collagen and carbonate reflect the isotopic composition of the food consumed during the period during which they were synthesized. In large mammals, bone collagen reflects several years of an individual’s lifetime before its death, while tooth collagen and enamel carbonate sampled along a tooth crowned reflect a much shorter period, during which this tooth part was formed, usually during the first months or years of an individual’s life. This situation has important implications in mammals, especially in the case of young individuals and of high-crown teeth of herbivorous mammals with prolonged growth periods. Young individuals are raised on their mother’s milk, which exhibits δ15N values around 3‰ higher than those of the adult diet (Fogel et al. 1989). For these reasons, the δ15N values of tooth collagen do not always reflect the adult diet in mammals and they should be treated cautiously if used in paleodietary reconstruction (Bocherens et al. 1994; Bocherens and Mariotti 1997). For the same reason, bone collagen of young individuals, from birth until a few years after weaning, is affected by the consumption of 15N-enriched milk, and several studies have demonstrated the consequences in the isotopic variations of the youngest individuals, up to 3 years old human babies, in modern and archeological human populations (e.g., Fogel et al. 1989; Katzenberg and Pfeiffer 1995; Fuller et al. 2006; Clayton et al. 2006). Similar patterns have been shown in the bones of young animals (e.g., Balasse et al. 1997). The isotopic signatures of such individuals are therefore difficult to address for paleodietary reconstruction. In high-crowned herbivorous mammals, such as horse and bovids, serial sampling of dentine and enamel bands along the crown perpendicularly to the growth direction yields varying isotopic results that correspond to seasonal or ontogenic changes in diet (e.g., Koch et al. 1989; Balasse et al. 2001; Drucker et al. 2001; Nelson 2005).
In summary, carbon isotopic signatures, measured on collagen or enamel carbonate bioapatite, yield direct information on the type of plants consumed by herbivores, and therefore on the vegetation surrounding the Neanderthals. In the case of predators, their δ13C values are informative in the case of potential preys with differing carbon isotopic signatures. Nitrogen isotopic signatures of bone collagen reflect the trophic position of an individual within its food web. Some variations in plants linked to environmental parameters are transferred to herbivores and their predators, shifting the δ15N values of the whole food web but keeping the nitrogen isotopic differences between predators and their prey within a predictable range. Examples relevant to the case of Neanderthals will be presented in the following sections.
Isotopic Variations and Ecology of the Middle and Late Pleistocene
Carbon-13 in Herbivores and the Vegetation Around Neanderthal
Carbon isotopic signatures, measured on collagen or enamel carbonate bioapatite, yield direct information on the type of plants consumed by herbivores, and therefore on the vegetation surrounding the Neanderthals. The main carbon isotopic distinction in terrestrial vegetation is due to the photosynthetic pathway used by the plants, i.e. the so-called C3 and C4 pathways. C4 plants are absent or very limited in environments with a temperate or cold growing season, as in Europe, including the Mediterranean area, northern latitudes and high altitudes (e.g., Ehleringer et al. 1997). The geographical extension of Neanderthals was deeply rooted in Europe, with representatives in the Middle East and central Asia (e.g., Bolus and Schmitz 2006; Finlayson and Carrión 2007). Within this range, C4 plants may be present as a significant but minor component in the Middle East and some dry sections of Central Asia (e.g., Bocherens et al. 2000). When present, these C4 plants are grasses or forbs. In environments where all plants use the C3 photosynthetic pathway, which correspond to the large majority of sites where Neanderthals occurred, an isotopic distinction can be seen between plants growing under a closed canopy, which have lower δ13C values than plants growing at the top of the canopy or in open environments, such as open woodland, grassland, steppe and tundra (reviews in Tieszen 1991; Heaton 1999). The tissues of herbivores consuming plants from a closed canopy also exhibit lower δ13C values than those of herbivores consuming plants from an open environment (e.g., van der Merwe and Medina 1991; Cerling and Harris 1999; Drucker et al. 2003b, 2008; Cerling et al. 2004; Drucker 2005, 2007). The differences in δ13C values can reach 5‰ in boreal and temperate contexts between open and closed areas. Shifts of δ13C values of plants and their consumers can also be due to other environmental factors such as altitude and humidity. The isotopic shifts are, however, more limited, of about 1.1‰ increase in δ13C values per 1,000 m increase in altitude and δ13C values 0.5‰ lower in humid grassland compared to drier ones (e.g., Schnyder et al. 2006; Männel et al. 2007).
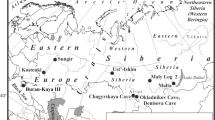
The use of δ13C values measured in the bones and teeth of herbivores found in prehistoric sites provide a tool to investigate whether animals dwelling in dense forests were part of the subsistence strategies of Neanderthals living during periods of forest development. As expected, the δ13C values of herbivores found in sites dated from OIS 3 are all within the range of animals consuming plants from open environments without dense canopy forests, as in Saint-Césaire, in Les Pradelles, in layer 1A in Scladina cave, and in layers 6a and 7a in Kůlna (e.g., Fizet et al. 1995; Bocherens et al. 1997, 2005a; Patou-Mathis et al. 2005) (location map on Fig. 8.1).


Location of the sites discussed in the text. The underlined names correspond to sites that yielded bones from Neanderthals
Only sites from more temperate periods, such as some phases of OIS 5, yielded herbivores with δ13C values negative enough to be consistent with dense canopy vegetation that could generate low δ13C values in plants. This was the case in layer 4A from Scladina cave, and in layer G from Payre (Bocherens et al. 1999; Bocherens and Rousseau 2008). The distribution of these low δ13C values in the different herbivorous species and in remains from animals hunted by humans or by carnivores tells us even more about the environment, the paleobiology of herbivorous mammals, and the subsistence strategies of Neanderthals. In layer 4A from Scladina cave, fallow deer exhibit low δ13C values, which is not surprising for this species of which extant representatives dwell in well forested landscapes. However, the occurrence of two horse specimens with δ13C values as low as those of fallow deer shows that some representatives of this species could be present even under forested conditions. Similar results were found for horses from Holocene sites in France (Bocherens et al. 2005a). These results show that ancient horses were not systematically associated to open environments, such as grasslands or steppes, but could also be present in forested environments. In both cases, horses were nevertheless a minor component of the fauna which was dominated by taxa more linked to forested environments such as cervids. In layer 4 from Scladina cave, it is striking that although a majority of analyzed herbivorous mammals yielded δ13C values indicative of a dense canopy forest, the δ13C values directly measured on a Neanderthal specimen indicate consumption of resources from open environments (Bocherens et al. 1999). This could indicate that, in a landscape with open and forested dietary resources, Neanderthals from layer 4A of Scladina cave focused on food from the open landscape. Due to limited data such a conclusion cannot be generalized to all Neanderthal populations living under temperate conditions. Indeed, the results obtained in layer G from Payre on large bovids and deer teeth indicate that these species were hunted by Neanderthals in dense forest environments (Bocherens and Rousseau 2008). The isotopic approach allows thus to determine the origin of hunted game within a mosaic landscape where open and forested areas were present, and to compare the subsistence strategies of different Neanderthal populations through space and time (Fig. 8.2). The development of this approach should be fruitful in the debate over the ecological tolerance of Neanderthals (e.g., Roebroeks et al. 1992).


Carbon isotopic signatures of the average diet reconstructed from collagen (Scladina, layer 4A; Bocherens et al. 1999) and from enamel carbonate apatite (Payre, layer G; Bocherens and Rousseau 2008) δ13C values. Neanderthal from Scladina layer 4A (open circle) exhibits δ13C values indicative of open environment, contrarily to some herbivores found in the same level, such as fallow deer and horse. In Payre layer G, some of the hunted herbivores, especially deer, exhibit δ13C values typical of dense forest environment, thus indicating that Neanderthals in Payre exploited herbivores from forested environments (Bocherens 2008)
Nitrogen-15 in Herbivores: The Effectof Environmental Factors
As described in the previous section, δ15N values of adult herbivores reflect those of the consumed vegetation. A recent review of variations of δ15N values in soils and plants demonstrated that, on a global scale, temperature and humidity are the main factors impacting on the δ15N values of plants, which decrease with decreasing temperature and increasing humidity (Amundson et al. 2003). The same factors have been shown to have similar effects on the δ15N values of plants and their consumers at more local scales, for instance with increasing altitude and on the scale of a continent such as Australia (Murphy and Bowman 2006; Männel et al. 2007).
In the context of European Late Pleistocene, large variations of δ15N values in herbivore collagen have been reported.The pattern of such variations in western Europe during the time range from 35 to 10,000 years ago has convincingly demonstrated that temperature played a driving role in the decreasing of δ15N values in herbivore collagen during and just after the Last Glacial Maximum, between 20 and 12,000 years ago, with an amplitude increasing when the distance to the polar front decreases (Drucker et al. 2003a). In the peri-Mediterranean realm, this drop in herbivore δ15N values during the Last Glacial Maximum is not observed, as in the site Paglicci in southern Italy (Iacumin et al. 1997). This clearly illustrates that different environmental changes occurred in different areas during the climatic fluctuations of the Late Pleistocene.
Other episodes of shifts of herbivore δ15N values have been reported in the Late Pleistocene, but the environmental cause is not so well understood. For instance, at a time that seems to correspond to the beginning of the Aurignacian and in localities from different areas of Europe, the δ15N values of horse, large bovids, as well as of a predator, the wolf, are 3–5‰ more positive than those of the same species before and after the Middle to Upper Paleolithic transition (Drucker and Bocherens 2004). Such an isotopic shift is very difficult to link to climatic changes, since it seems to correspond to a time of decreasing temperature, which should lead to a decrease of δ15N values in plants and herbivores as seen during the Last Glacial Maximum, and not to an increase in δ15N value as it is observed. Further research should help to better characterize this isotopic shift chronologically and spatially. One hypothesis to be tested is the possible link between this isotopic shift and the Campanian ignimbrite eruption, which occurred around this time and is likely to have disrupted ecosystems on a large scale in Europe (Fedele et al. 2002).
There are also some hints of such increases in δ15N values during earlier periods. One such episode was recognized in the study of Les Pradelles, where specimens belonging to reindeer, horse, bison, and wolf from layer 7 exhibited significantly higher δ15N values than specimens from the same species coming from layers under- and overlying (Fizet et al. 1995). Paleoclimatological proxies point to a very dry and cold climate during the deposition of this layer (Fizet et al. 1995). Reports of unusually high δ15N values have also been made for bison bones from England older than 55 ka (Jacobi et al. 2006), and for bison and proboscideans from a 200 ka old English site (Jones et al. 2001). Aridity is often presented as the most likely cause of these positive excursions of δ15N values (Fizet et al. 1995; Jones et al. 2001), but the biogeochemical behavior of nitrogen under the unique environmental conditions of the Late Pleistocene in Europe is not yet well understood and requires further investigations. In particular, it would be necessary to consider the possible influence of the herbivore community on the biogeochemistry of nitrogen, since grazing is a factor known to increase the δ15N values of plants and soils (e.g., Neilson et al. 2002), especially through manuring (e.g., Simpson et al. 1999; Choi et al. 2002; Frank et al. 2004; Dijkstra et al. 2006).
Therefore, changes in the herbivore communities through the Late Pleistocene may be also responsible for some variations observed in the δ15N values of terrestrial ecosystems. Even if the actual causes of these isotopic fluctuations are still not fully understood, it is essential to take them into account in the paleodietary interpretations, as it will be illustrated in the following sections.
Choice of Prey by Predators
Carbon isotopic signatures yield direct information on the type of plants consumed by herbivores. In the case of predators, their δ13C values are informative in the case of potential preys with differing carbon isotopic signatures, such asherbivores from dense forests, herbivores from open environments, freshwater resources, and marine resources. Nitrogen isotopic signatures of bone collagen reflect the trophic position of an individual within its food web. Some variations in plants linked to environmental parameters are transferred to herbivores and their predators, shifting the δ15N values of the whole food web but keeping the nitrogen isotopic differences between predators and their prey within a predictable range. Therefore, in a given paleoecosystem, using the δ13C and the δ15N values from collagen allows to determine which prey were consumed by predators, if different prey species exhibit significant differences in their isotopic signatures.
A compilation of carbon and nitrogen isotopic data obtained on fossil mammals dated to OIS 3, between 24 and 60 ka, showed that a consistent pattern of isotopic variations existed between ungulates living in the so-called “steppe-tundra” or “mammoth steppe”, from western France to Alaska (see review in Bocherens 2003). Although the environmental setting of this unique biome has no real modern equivalent, it seems that the isotopic variations observed in various herbivorous mammals such as reindeer, large bovids, horse, and mammoth, seem to relate to isotopic differences in their preferred plant food items that follow the same isotopic discrimination as modern plants in boreal and arctic environments. Lichens, which exhibit less negative δ13C values than vascular plants, lead to more positive δ13C values in reindeer collagen than in coeval herbivores due to the high proportion of these plants in reindeer diet (e.g., Fizet et al. 1995; Drucker et al. 2003a). The herbivores with the most positive δ15N values, such as mammoths and large bovids, seem to be the ones relying most on grass rather than shrub leaves. These two types of plants differ greatly in their strategy of nitrogen intake, grass having no symbiotic association with fungi, while most shrubs and trees obtain most of their nitrogen from symbiotic fungi, with significant isotopic fractionation leading to more negative δ15N values in shrubs and trees than in grass (e.g., Högberg et al. 1996; Michelsen et al. 1996; Commisso and Nelson 2006; Hobbie and Hobbie 2006).
The large range of variation of δ15N values in plants from arctic and boreal environments linked to their differences in nitrogen metabolism is sufficient to explain the pattern of isotopic variation exhibited by herbivores, especially since the nitrogen isotopic fractionation between herbivore tissues and their diet varies less than originally believed (see review in previous paragraphs). Even the large difference in δ15N values of around 4‰ observed between the most 15N-enriched and the most 15N-depleted herbivore in a given paleoecosystem, such as mammoth and cave bear in layer 1A of Scladina cave, is well within the observed range of variation of δ15N values between grass and shrubs. These observations cast serious doubts upon recent works based on models that advocate physiological factors such as hyperphagia, lactation and suckling as the cause for cave bears having lower δ15N values than those of sympatric herbivores (Balter et al. 2006), especially when considering the absence of difference between the δ15N values of male and female cave bears (Bocherens, unpublished data), a side-effect predicted by this theory, and the absence of such an effect in modern bears, which do have hyperphagia and hibernation but do not differ from other carnivores in the fractionation of their nitrogen compared to their food (Hilderbrand et al. 1996; Felicetti et al. 2003).
The consequences of these significant isotopic differences in terrestrial prey species available to Neanderthals and coeval predators is the possibility to evaluate quantitatively the proportions of the different prey that they consumed. In two case studies dealing with Neanderthals and coeval herbivores and carnivores dated to the end of OIS 3, around 40–35 ka, and located in two different regions, Western France and Belgium, the isotopic pattern exhibited by the different herbivorous and carnivorous species are strikingly similar (Fig. 8.3).


Carbon and nitrogen isotopic signatures of collagen from herbivorous and carnivorous mammals in Western France (Saint-Césaire,La Berbie, Camiac) and Belgium (Spy, layer 1A in Scladina cave) (Modified from Bocherens et al. (2005b))
In both cases, mammoths exhibit the most positive and reindeer the least positive δ15N values of herbivores, while reindeer exhibit the least negative and horse and mammoth exhibit the most negative δ13C values of herbivores. Hyenas present δ15N values slightly more positive than those of mammoths and about 3–4‰ more positive than those of the bulk of herbivores, while Neanderthals are about 2‰ more positive than hyenas.
In the case of Saint-Césaire Neanderthal, the use of a mathematical model was possible thanks to the diversity and the richness of the coeval fauna that provided robust end-members for potential prey species available to the predators and allowed quantitative estimates for the contribution of various prey in the average diet of Neanderthals and hyenas (Bocherens et al. 2005b). The results point to a much larger proportion of mammoth and rhinoceros in the diet of Neanderthals than for hyenas, while hyenas consumed more reindeer than Neanderthals. The relative contribution of the other ungulates, such as horse, large bovids and large deer, were ranging similarly for both predators. This result indirectly suggests that mammoth and rhinoceros were hunted rather than scavenged. Otherwise hyenas would also have a better access to these carcasses which was not the case. An alternative explanation for the more positive δ15N values in Neanderthals than in hyenas is the consumption of freshwater resources by the humans (Drucker and Bocherens 2004; Balter and Simon 2006). However, the complete lack of archeological evidence for exploitation of fish in sites occupied by Neanderthals, as well as the very tight clustering of the isotopic values of Neanderthal specimens from a given site, make it unlikely that fish were significantly involved in the diet of the studied Neanderthals. Moreover, the trace-element data presented to support fish consumption by Neanderthals by Balter and Simon (2006) are affected by diagenesis and lack faunal comparison data, which make them unreliable for dietary reconstructions (Lee-Thorp and Sponheimer 2006).
In the case of Belgian samples, no detailed mathematical reconstruction has been attempted yet since the association between the Neanderthal specimen from Spy and the mammalian fauna from Scladina is not as secure as in western France. But the similar isotopic pattern among species suggest a similar result for dietary reconstruction, with megaherbivores such as mammoth and rhinoceros representing a much higher percentage in the diet of Neanderthals than in the diet of hyenas. These results suggest the possibility of a niche partitioning between Neanderthals and hyenas, the hominids aiming at the largest herbivorous prey while the hyena relied more on smaller sized prey, such as large bovids, cervids and horses. The hunting technology of Neanderthals could have given them an advantage in the hunting of very large herbivores, opening them an ecological niche unavailable to other predators and allowing the cohabitation of several large predators in late Pleistocene Europe without too much dietary overlap between predatory species.
Comparison of Isotopic and Zooarcheological Dietary Reconstruction
The exploitation of megaherbivores such as proboscideans and rhinoceros by Neanderthals and other fossil European hominids has been previously suggested based on the study of animal remains found in prehistoric sites (e.g., Scott 1980; Thieme and Veil 1985; Auguste 1995; Auguste et al. 1998; Bratlund 2000; Patou-Mathis 2000, 2006; Conard and Niven 2001; Moncel 2001; Piperno and Tagliacozzo 2001). However, the consumption of such a large proportion of up to 70% mammoth and up to 56% rhinoceros meat as reconstructed with the isotopic approach is not generally deduced from zooarcheological studies of Neanderthal food leftovers (Bocherens et al. 2005b). For instance, in the case of Saint-Césaire, the zooarcheological study points to a large proportion of bovinae in the meat consumed by Châtelperronian Neanderthals, while mammoth and rhinoceros would account for 16% and 9%, respectively (Patou-Mathis 2006). One common conclusion of isotopic and zooarcheological investigations is the very low proportion of deer and reindeer in the diet of Neanderthals (Fig. 8.4).


Range of proportions of different prey species in the diet of Neanderthal from Saint-Césaire and hyenas from western France (Modified from Bocherens et al. 2005b). The vertical arrows correspond to the proportions of meat weight of different prey speciescalculated from the results of zooarcheological study (Patou-Mathis 2006)
Therefore both approaches agree that Neanderthals have consumed more meat from large mammals than from small ones. The difference in the conclusions of both approaches regarding mammoth and rhinoceros could be due to the consequence of transport decision, since bones of very large herbivores are not expected to be transported from the butchery to the occupation site, only meat filets would be transported leading to an underrepresentation of the role of large-bodied animals in Middle Paleolithic diet (e.g., Rabinovitch and Hovers 2004).
Another case of discrepancy between the conclusions of zooarcheological and isotopic dietary studies is provided by the Les Pradelles site, in Charentes (France). Three Neanderthal specimens from this site have yielded reliable collagen that was analyzed isotopically, as well as specimens of coeval horse, large bovids, reindeer, wolf and hyena (Fizet et al. 1995; Bocherens et al. 2005b). Based on zooarcheological analysis of the fauna from layers 9 and 10 where the Neanderthals come from suggest a site where essentially reindeer were hunted, with 55 individuals, while other species yielded much less individuals, i.e. 3 red deer, 11 horse and 8 large bovids (Costamagno et al. 2006). The isotopic results indicate that the contribution of prey with high δ15N values, i.e. large bovids, has to be important in order to fit the observed isotopic values for all three Neanderthals (Fig. 8.5).


Carbon and nitrogen isotopic signatures of collagen fromherbivorous and carnivorous mammals from Les Pradelles, compared to those of coeval Neanderthal specimens (Modified from Bocherens et al. 2005b). Rectangles represented the reconstructed range of collagen for predators consuming exclusively one prey species, i.e. Bovinae, Horse or Reindeer. This reconstruction is based on the carbon and nitrogen isotopic fractionations for predators compared to their prey presented by Bocherens and Drucker (2003)
The apparent discrepancy in terms of hunted individuals is partially corrected if we calculate the maximum percent meat obtainable from the different species, taking into account that an adult reindeer yields around 50 kg meat, and adult horse 200 kg and an adult large bovid 750 kg (based on Patou-Mathis 2006). Due to these differences in size, eight large bovid individuals would provide around half the meat of all herbivores found in Les Pradelles. The isotopic results suggest that bovids should be even more important in the Neanderthal diet, unless other large herbivores with high δ15N values, such as rhinoceros and mammoths, were hunted but not represented in the site. This is not to say that reindeer were unimportant to Les Pradelles Neanderthals or that zooarcheology yielded false results. Les Pradelles could well correspond to a site where reindeer hunting and processing was a major activity, but that, on average, the main protein source of the three analyzed Neanderthal specimens was meat from large bovids.
Conclusions and Perspectives
Isotopic investigations of fossil bones relevant to questions of Neanderthal paleoecology are only at their beginning. So far, only about 25 Neanderthal individuals have been attempted using this new isotopic approach, and only 12 specimens yielded well-preserved collagen, ten of them having been published (Bocherens et al. 2005b). The present paper discussed isotopic results obtained on only 5 Neanderthal individuals as the other additional specimens for which isotopic data were published lack either suitable comparison fauna, are juvenile for which interference from suckling milk could not be excluded or their collagen presents preservation problems (see Bocherens et al. 2005b for detailed discussion). There are still a huge number of prehistoric sites dated to the Late Pleistocene that should yield fossil bones and teeth suitable for collagen and/or enamel apatite isotopic studies, allowing paleoecological reconstruction with high spatial and chronological resolution in the coming years. One question that will certainly benefit from the further isotopic investigations is the Middle to Paleolithic transition, especially by testing the possibility of dietary competition between late Neanderthals and early Anatomically Modern Humans in Europe. The results obtained so far are still ambiguous (Richards et al. 2001 contra Drucker and Bocherens 2004). The addition of new biogeochemical tracers, such as sulfur isotopic signatures, could help to solve these controversies (e.g., Richards et al. 2003). In parallel, a better integration of paleoecological results from different indicators, including isotopic tracers, should allow more robust reconstructions of local environmental conditions, which will be fruitfully compared with paleoclimatic reconstructions based on glacial and marine global tracers. Another promising field of research is the combination of isotopic and paleogenetic data that will help to understand the mechanism of population and species evolution in the changing environment of the Late Pleistocene (e.g., Barnes et al. 2002; Bocherens et al. 2006, 2007).
References
Adler, D. S., Bar-Oz, G., Belfer-Cohen, A., & Bar-Yosef, O. (2006). Ahead of the Game. Middle and Upper Palaeolithic hunting behaviors in the Southern Caucasus. Current Anthropology, 47, 89–118.
Ambrose, S. H. (1990). Preparation and characterization of bone and tooth collagen for isotopic analysis. Journal of Archaeological Science, 17, 431–451.
Ambrose, S. H. (1991). Effects of diet, climate and physiology on nitrogen isotope abundances in terrestrial foodwebs. Journal of Archaeological Science, 18, 293–317.
Ambrose, S. H., & DeNiro, M. J. (1986). The isotopic ecology of East African mammals. Oecologia, 69, 395–406.
Amundson, R., Austin, A. T., Schuur, E. A. G., Yoo, K., Matzek, V., Kendall, C., Uebersax, A., Brenner, D., & Baisden, W. T. (2003). Global patterns of the isotopic composition of soil and plant nitrogen. Global Biogeochemical Cycles, 17(1), 1031.
Asara, J. M., Schweitzer, M. H., Freimark, L. M., Phillips, M., & Cantley, L. C. (2007). Protein sequences from Mastodon and Tyrannosaurus rex revealed by mass spectrometry. Science, 316, 280–285.
Auguste, P. (1995). Chasse et charognage au paléolithique moyen: l’apport du gisement de Biache-Saint-Vaast (Pas-de-Calais). Bulletin de la Société Préhistorique Française, 92, 155–167.
Auguste, P., Moncel, M.-H., & Patou-Mathis, M. (1998). Chasse ou “charognage”: acquisition et traitement des Rhinocéros au Paléolithique moyen en Europe Occidentale. In J.-P. Brugal, L. Meignen, & M. Patou-Mathis (Eds.), Actes du XIIIème Colloque d’Antibes, Economie préhistorique: les comportements de subsistance au Paléolithique, 23–25 octobre 1997 (pp. 133–151). CNRS Edition, APDCA, Sophia Antipolis.
Ayliffe, L. K., Chivas, A. R., & Leakey, M. G. (1994). The retention of primary oxygen isotope compositions of fossil elephant skeletal phosphate. Geochimica et Cosmochimica Acta, 85, 5291–5298.
Balasse, M., Bocherens, H., Tresset, A., Mariotti, A., & Vigne, J.-D. (1997). Émergence de la production laitière au néolithique ? Contribution de l’analyse isotopique d’ossements de bovins archéologiques. Comptes rendus de l’Academie des Sciences Paris. Série IIA. Science de la terre et des planètes, 325, 1005–1010.
Balasse, M., Bocherens, H., Ambrose, S. H., & Mariotti, A. (2001). Detection of dietary changes by intra-tooth carbon and nitrogen isotopic analysis: An experimental study of dentine collagen of cattle (Bos taurus). Journal of Archaeological Science, 28, 235–245.
Balter, V., & Simon, L. (2006). Diet and behavior of the Saint-Césaire Neanderthal inferred from biogeochemical data inversion. Journal of Human Evolution, 51, 329–338.
Balter, V., Simon, L., Fouillet, H., & Lécuyer, C. (2006). Box-modelling of 15N/14N in mammals. Oecologia, 147, 212–222.
Barnes, I., Matheus, P., Shapiro, B., Jensen, D., & Cooper, A. (2002). Dynamics of Pleistocene population extinctions in Beringian brown bears. Science, 295, 2267–2270.
Bocherens, H. (2000). Preservation of isotopic signals (13C, 15N) in Pleistocene mammals. In M. A. Katzenberg & S. H. Ambrose (Eds.), Biogeochemical approaches to Paleodietary analyses (pp. 65–88). New York: Kluwer Academic/Plenum.
Bocherens, H. (2003). Isotopic biogeochemistry and the paleoecology of the mammoth steppe fauna. In W. F. Reumer, F. Braber, D. Mol, & J. de Vos (Eds.), Advances in Mammoth research (pp. 57–76). Rotterdam: Deinsea.
Bocherens, H., & Drucker, D. (2003). Trophic level isotopic enrichments for carbon and nitrogen in collagen: Case studies from recent and ancient terrestrial ecosystems. International Journal of Osteoarchaeology, 13, 46–53.
Bocherens, H., & Mariotti, A. (1997). Comments on: Diet, physiology and ecology of fossil mammals as inferred from stable carbon and nitrogen isotope biochemistry: Implications for Pleistocene bears by Bocherens et al. - Reply. Palaeogeography, Palaeoclimatology, Palaeoecology, 128, 362–364.
Bocherens, H., & Rousseau, L. (2008). Chapitre 8: Le cadre environnemental des occupations humains. Isotopes sur grands mammifères et plancher stalagmitique. In M.-H. Moncel (Ed.), Le site de Payre: occupations humaines dans la vallée du Rhône à la fin du Pleistocène moyen et au début du Pleistocène supérieur. Mémoire de la Société préhistorique française (46, pp. 113–122). Société préhistorique française, Paris.
Bocherens, H., Fizet, M., Cuif, J.-P., Jaeger, J.-J., Michard, J.-G., & Mariotti, A. (1988). Premières mesures d'abondances isotopiques naturelles en 13C et en 15N de la matière organique fossile de Dinosaure. Application à l'étude du régime alimentaire du genre Anatosaurus (Ornithischia, Hadrosauridae). Comptes rendus de l’Academie des Sciences Paris. Série IIA. Science de la terre et des planètes, 306, 1521–1526.
Bocherens, H., Fizet, M., Mariotti, A., Lange-Badré, B., Vandermeersch, B., Borel, J.-P., & Bellon, G. (1991). Isotopic biogeochemistry (13C, 15N) of fossil vertebrate collagen: Implications for the study of fossil food web including Neandertal Man. Journal of Human Evolution, 20, 481–492.
Bocherens, H., Fizet, M., & Mariotti, A. (1994). Diet, physiology and ecology of fossil mammals as inferred by stable carbon and nitrogen isotopes biogeochemistry: Implications for Pleistocene bears. Palaeogeography, Palaeoclimatology, Palaeoecology, 107, 213–225.
Bocherens, H., Fogel, M. L., Tuross, N., & Zeder, M. (1995). Trophic structure and climatic information from isotopic signatures in a Pleistocene cave fauna of Southern England. Journal of Archaeological Science, 22, 327–340.
Bocherens, H., Billiou, D., Patou-Mathis, M., Bonjean, D., Otte, M., & Mariotti, A. (1997). Isotopic biogeochemistry (13C, 15N) of fossil mammal collagen from Scladina cave (Sclayn, Belgium). Quaternary Research, 48, 370–380.
Bocherens, H., Billiou, D., Patou-Mathis, M., Otte, M., Bonjean, D., Toussaint, M., & Mariotti, A. (1999). Paleoenvironmental and paleodietary implications of isotopic biogeochemistry of late interglacial Neandertal and mammal bones in Scladina Cave (Belgium). Journal of Archaeological Science, 26, 599–607.
Bocherens, H., Billiou, D., Charpentier, V., & Mashkour, M. (2000). Aleoenvironmental and archeological implications of bone and tooth isotopic biogeochemistry (13C, 15N) in Southwestern Asia. In H. Buitenhuis, M. Mashkour, & F. Poplin (Eds.), Archeozoology of the Near East IV (pp. 104–115). Groningen: ARC 32 (B).
Bocherens, H., Drucker, D., & Billiou, D. (2005a). Paléoenvironnements et Hominidés: Approches biogéochimiques. In A. Tuffreau (Ed.), Peuplements humains et variations environnementales au Quaternaire (BAR International Series 1352, pp. 55–68). Oxford: Archaeopress.
Bocherens, H., Drucker, D. G., Billiou, D., Patou-Mathis, M., & Vandermeersch, B. (2005b). Isotopic evidence for diet and subsistence pattern of the Saint-Césaire I Neanderthal: Review and use of a multi-source mixing model. Journal of Human Evolution, 49, 71–87.
Bocherens, H., Hofreiter, M., Münzel, S. C., & Conard, N. J. (2006). Human and bear population dynamics and evolutionary trends in Southwestern Germany. 48. Tagung der Hugo Obermaier-Gesellschaft für Erforschung des Eiszeitalters und der Steinzeit (Köln, 18–22 April 2006), Erlangen, 12.
Bocherens, H., Conard, N. J., Germonpré, M., Hofreiter, M., Münzel, S., Stephan, E., & Tütken, T. (2007). Isotopic biogeochemistry and the evolution of cave bear ecology during Marine Oxygen Isotopic Stage 3 in Western and Central Europe. In R. Musil & V. Vávra (Eds.), 13th international Cave Bear symposium, September 20–24. Czech Republic Proceedings. Masaryk University, Brno.
Bolus, M., & Schmitz, R. W. (2006). Der Neandertaler. Ostfildern: Thorbecke Verlag.
Bratlund, B. (2000). Taubach revisited. Jahrbuch des Römisch-Germanischen Zentralmuseums Mainz, 46, 61–174.
Cerling, T. E., & Harris, J. M. (1999). Carbon isotope fractionation between diet and bioapatite in ungulate mammals and implications for ecological and paleoecological studies. Oecologia, 120, 347–363.
Cerling, T. E., Hart, J. A., & Hart, J. B. (2004). Stable isotope ecology in the Ituri-Forest. Oecologia, 138, 5–12.
Choi, W. J., Lee, S. M., Ro, H. M., Kim, K. C., & Yoo, S. H. (2002). Natural 15N abundances of maize and soil amended with urea and composted pig manure. Plant and Soil, 245, 223–232.
Churchill, S. E. (1998). Cold adaptation, heterochrony, and Neandertals. Evolutionary Anthropology, 7, 46–61.
Clayton, F., Sealy, J., & Pfeiffer, S. (2006). Weaning age among foragers at Matjes River Rock Shelter, South Africa, from stable nitrogen and carbon isotope analyses. American Journal of Physical Anthropology, 129, 311–317.
Commisso, R. G., & Nelson, D. E. (2006). Modern plant δ15N values reflect ancient human activity. Journal of Archaeological Science, 33, 1167–1176.
Conard, N. J., & Niven, L. (2001). The paleolithic finds from Bollschweil and the question of Neandertal mammoth hunting in the Black Forest. In G. Cavarretta, P. Giola, M. Mussi, & M. R. Palombo (Eds.), The world of Elephants (pp. 194–200). Roma: Consiglio nazionale delle ricerche.
Costamagno, S., Meignen, L., Beauval, C., Vandermeersch, B., & Maureille, B. (2006). Les Pradelles (Marillac-le-Franc, France): A Mousterian reindeer hunting camp? Journal of Anthropological Archaeology, 25, 466–484.
DeNiro, M. J. (1985). Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to paleodietary reconstruction. Nature, 317, 806–809.
DeNiro, M. J., & Epstein, S. (1978). Influence of diet on the distribution of carbon isotopes in animals. Geochimica et Cosmochimica Acta, 42, 495–506.
DeNiro, M. J., & Epstein, S. (1981). Influence of diet on the distribution of nitrogen isotopes in animals. Geochimica et Cosmochimica Acta, 45, 341–351.
Dijkstra, P., Menyailo, O. V., Doucett, R. R., Hart, S. C., Schwartz, E., & Hungate, B. A. (2006). C and N availability affects the 15N natural abundance of the soil microbial biomass across a cattle manure gradient. European Journal of Soil Science, 57, 468–475.
Drucker, D. (2005). Ecologie des rennes chassés par les hommes préhistoriques: que peuvent dire les isotopes du carbone? Annales de la Fondation Fyssen, 20, 90–97.
Drucker, D. (2007). Les cervidés durant le tardiglaciaire et l’holocène ancien en Europe occidentale: Approche isotopique. In S. Beyries & V. Vaté (Eds.), Les civilisations du renne d’hier et d’aujourd’hui. Approches ethnohistoriques, archéologiques et anthropologiques. XXVII e rencontres internationales d’archéologie et d’histoire d’Antibes (pp. 243–253). Antibes: APDCA.
Drucker, D., & Bocherens, H. (2004). Carbon and nitrogen stable isotopes as tracers of change in diet breadth during Middle and Upper Paleolithic in Europe. International Journal of Osteoarchaeology, 14, 162–177.
Drucker, D., Bocherens, H., Pike-Tay, A., & Mariotti, A. (2001). Isotopic tracking of seasonal dietary change in dentine collagen: Preliminary data from modern caribou. Comptes rendus de l’Academie des Sciences Paris. Série IIA. Science de la terre et des planètes, 333, 303–309.
Drucker, D., Bocherens, H., & Billiou, D. (2003a). Evidence for shifting environmental conditions in Southwestern France from 33,000 to 15,000 years ago derived from carbon-13 and nitrogen-15 natural abundances in collagen of large herbivores. Earth and Planetary Science Letters, 216, 163–173.
Drucker, D., Bocherens, H., Bridault, A., & Billiou, D. (2003b). Carbon and nitrogen isotopic composition of red deer (Cervus elaphus) collagen as a tool for tracking paleoenvironmental change during the Late-Glacial and Early Holocene in the northern Jura (France). Palaeogeography, Palaeoclimatology, Palaeoecology, 195, 375–388.
Drucker, D., Bridault, A., Hobson, K. A., Szuma, E., & Bocherens, H. (2008). Can carbon-13 abundances in large herbivores track canopy effect in temperate and boreal ecosystems? Evidence from modern and ancient ungulates. Palaeogeography, Palaeoclimatology, Palaeoecology, 266, 69–82.
Ehleringer, J. R., Cerling, T. E., & Helliker, B. R. (1997). C4 photosynthesis, atmospheric CO2, and climate. Oecologia, 112, 285–299.
Fedele, F. G., Biaccio, B., Isaia, R., & Orsi, G. (2002). Ecosystem impact of the Campanian ignimbrite eruption in Late Pleistocene Europe. Quaternary Research, 57, 420–424.
Felicetti, L. A., Schwartz, C. C., Rye, R. O., Haroldson, M. A., Gunther, K. A., Phillips, D. L., & Robbins, C. T. (2003). Use of sulfur and nitrogen stable isotopes to determine the importance of whitebark pine nuts to Yellowstone grizzly bears. Canadian Journal of Zoology, 81, 763–770.
Finlayson, C., & Carrión, J. S. (2007). Rapid ecological turnover and its impact on Neanderthal and other human populations. Trends in Ecology & Evolution, 22, 213–222.
Fizet, M., Mariotti, A., Bocherens, H., Lange-Badré, B., Vandermeersch, B., Borel, J. P., & Bellon, G. (1995). Effect of diet, physiology andclimate on carbon and nitrogen isotopes of collagen in a late Pleistocene anthropic paleoecosystem (France, Charente, Marillac). Journal of Archaeological Science, 22, 67–79.
Fogel, M. L., Tuross, N., & Owsley, D. W. (1989). Nitrogen isotope traces of human lactation in modern and archeological populations. Annual Report of the Director of the Geophysical Laboratory, Carnegie Institution 1988–1989, pp. 111–117.
Frank, D. A., Evans, R. D., & Tracy, B. F. (2004). The role of ammonia volatilization in controlling the natural 15N abundance of a grazed grassland. Biogeochemistry, 68, 169–178.
Fuller, B. T., Fuller, J. L., Harris, D. A., & Hedges, R. E. M. (2006). Detection of breastfeeding and weaning in modern human infants with carbon and nitrogen stable isotope ratios. American Journal of Physical Anthropology, 129(2), 279–293.
Gröcke, D. R., Bocherens, H., & Mariotti, A. (1997). Annual rainfall and nitrogen-isotope correlation in Macropod collagen: Application as a paleoprecipitation indicator. Earth and Planetary Science Letters, 153, 279–285.
Heaton, T. H. E. (1999). Spatial, species, and temporal variations in the 13C/12C ratios of C3 plants: Implications for paleodiet studies. Journal of Archaeological Science, 26, 637–649.
Heaton, T. H. E., Vogel, J. C., von la Chevallerie, G., & Collett, G. (1986). Climatic influence on the isotopic composition of bone nitrogen. Nature, 322, 822–823.
Hedges, R. E. M. (2003). On bone collagen-apatite-carbonate isotopic relationships. International Journal of Osteoarchaeology, 13, 66–79.
Hedges, R. E. M., & Reynard, L. M. (2007). Nitrogen isotopes and the trophic level of humans in archeology. Journal of Archaeological Science, 34, 1240–1251.
Higham, T., Bronk Ramsey, C., Karavanic, I., Smith, F. H., & Trinkaus, E. (2006). Revised direct radiocarbon dating of the Vindija G1 Upper Paleolithic Neandertals. Proceedings of the National Academy of Sciences of the United States of America, 103, 553–557.
Hilderbrand, G. V., Farley, S. D., Robbins, C. T., Hanley, T. A., Titus, K., & Servheen, C. (1996). Use of stable isotopes to determine diets of living and extinct bears. Canadian Journal of Zoology, 74, 2080–2088.
Hobbie, J. E., & Hobbie, E. A. (2006). 15N in symbiotic fungi and plants extimates nitrogen and carbon flux rates in arctic tundra. Ecology, 87, 816–822.
Hockett, B., & Haws, J. A. (2005). Nutritional ecology and the human demography of Neandertal extinction. Quaternary International, 137, 21–34.
Högberg, P., Högbom, L., Schinkel, H., Högberg, M., Johannisson, C., & Wallmark, H. (1996). 15N abundance of surface soils, roots and mycorrhizas in profiles of European forest soils. Oecologia, 108, 207–214.
Holmes, K. M., Robson Brown, K. A., Oates, W. P., & Collins, M. J. (2005). Assessing the distribution of African Paleolithic sites: A predicitive model of collagen degradation. Journal of Archaeological Science, 32, 157–166.
Iacumin, P., Bocherens, H., Delgado Huertas, A., Mariotti, A., & Longinelli, A. (1997). A stable isotope study of fossil mammal remains from the Paglicci cave. S. Italy. N and C as paleoenvironmental indicators. Earth and Planetary Science Letters, 148, 349–357.
Jacobi, R. M., Higham, T. F. G., & Bronk Ramsey, C. (2006). AMS radiocarbon dating of Middle and Upper Paleolithic bone in the British Isles: Improved reliability using ultrafiltration. Journal of Quaternary Science, 21, 557–573.
Jones, A. M., O’Connell, T. C., Young, E. D., Scott, K., Buckingham, C. M., Iacumin, P., & Brasier, M. D. (2001). Biogeochemical data from well preserved 200 ka collagen and skeletal remains. Earth and Planetary Science Letters, 193, 143–149.
Katzenberg, M. A., & Pfeiffer, S. (1995). Nitrogen isotope evidence for weaning age in a nineteenth century canadian skeletal sample. In A. L. Grauer (Ed.), Bodies of evidence. Reconstructing history through skeletal analysis (pp. 221–235). New York: Wiley-Liss.
Koch, P. L., Fisher, D. C., & Dettman, D. (1989). Oxygen isotope variation in the tusks of extinct proboscideans: A measure of season of death and seasonality. Geology, 17, 515–519.
Koch, P. L., Tuross, N., & Fogel, M. L. (1997). The effects of sample treatment and diagenesis on the isotopic integrity of carbonate in biogenic hydroxylapatite. Journal of Archaeological Science, 24, 417–429.
Lee-Thorp, J. A., & Sponheimer, M. (2003). Three case studies used to reassess the reliability of fossil bone and enamel isotope signals for paleodietary studies. Journal of Anthropological Archaeology, 22, 208–216.
Lee-Thorp, J. A., & Sponheimer, M. (2006). Contributions of biogeochemistry to understanding Hominin dietary ecology. Yearbook of Physical Anthropology, 49, 131–148.
Lee-Thorp, J. A., & van der Merwe, N. J. (1991). Aspects of the chemistry of modern and fossil biological apatites. Journal of Archaeological Science, 18, 343–354.
Longin, R. (1971). New method of collagen extraction for radiocarbon dating. Nature, 230, 241–242.
Männel, T. T., Auerswald, K., & Schnyder, H. (2007). Altitudinal gradients of grassland carbon and nitrogen isotope composition are recorded in the hair of grazers. Global Ecology and Biogeography, 16, 583–592.
Marean, C. W., & Assefa, Z. (1999). Zooarcheological evidence for the faunal exploitation behavior of Neandertals and Early Modern Humans. Evolutionary Anthropology, 8, 22–37.
McGill, B. J., Hadly, E. A., & Maurer, B. A. (2006). Community inertia of Quaternary small mammal assemblages in North America. Proceedings of the National Academy of Sciences of the United States of America, 102, 16701–16706.
Michelsen, A., Schmidt, I. K., Jonasson, S., Quarmby, C., & Sleep, D. (1996). Leaf 15N abundance of subarctic plants provides field evidence that ericoid, ectomycorrhizal and non- and arbuscular mycorrhizal species access different sources of soil nitrogen. Oecologia, 105, 53–63.
Moncel, M.-H. (2001). Microlithic Middle Paleolithic assemblages in Central Europe and elephant remains. In G. Cavarretta, P. Giola, M. Mussi, & M. R. Palombo (Eds.), The world of Elephants, Proceedings of the 1st international Congress (pp. 314–317). Roma: Consiglio nazionale delle ricerche.
Murphy, B. P., & Bowman, D. M. J. S. (2006). Kangaroo metabolism does not cause the relationship between bone collagen δ15N and water availability. Functional Ecology, 20, 1062–1069.
Neilson, R., Robinson, D., Marriott, C. A., Scrimgeour, C. M., Hamilton, D., Wishart, J., Boag, B., & Handley, L. L. (2002). Above-ground grazing affects floristic composition and modifies soil trophic interactions. Soil Biology and Biochemistry, 34, 1507–1512.
Nelson, S. V. (2005). Paleoseasonality inferred from equid teeth and intra-tooth isotopic variability. Palaeogeography, Palaeoclimatology, Palaeoecology, 222, 122–144.
Ostrom, P. H., Macko, S. A., Engel, M. H., & Russell, D. A. (1993). Assessment of trophic structure of Cretaceous communities based on stable nitrogen isotope analyses. Geology, 21, 491–494.
Passey, B. H., Robinson, T. F., Ayliffe, L. K., Cerling, T. E., Sponheimer, M., Dearing, M. D., Roeder, B. L., & Ehleringer, J. R. (2005). Carbon isotope fractionation between diet, breath CO2, and bioapatite in different mammal. Journal of Archaeological Science, 32, 1459–1470.
Patou-Mathis, M. (2000). Neandertal subsistence behaviours in Europe. International Journal of Osteoarchaeology, 10, 379–395.
Patou-Mathis, M. (2006). Comportement de subsistance des Néandertaliens du niveau châtelperronien de Saint-Césaire (Charente-Maritime). Munib, 57, 197–204.
Patou-Mathis, M., Auguste, P., Bocherens, H., Condemi, S., Michel, V., Neruda, P., & Valoch, K. (2005). Les occupations du Paléolithique moyen de la grotte de Kůlna (Moravie, République Tchèque): Nouvelles approches, nouveaux résultats. In A. Tuffreau (Ed.), Peuplements humains et variations environnementales au Quaternaire (BAR International Series1352, pp. 69–94). Oxford: Archaeopress.
Piperno, M., & Tagliacozzo, A. (2001). The elephant butchery area at the Middle Pleistocene site of Notarchirico (Venosa, Basilicata, Italy). In G. Cavarretta, P. Giola, M. Mussi, & M. R. Palombo (Eds.), The world of Elephants, Proceedings of the 1st international Congress (pp. 230–236). Roma: Consiglio nazionale delle ricerche.
Rabinovitch, R., & Hovers, E. (2004). Faunal analysis from Amud Cave: Preliminary results and interpretations. International Journal of Osteoarchaeology, 14, 287–306.
Richards, M., Pettitt, P., Stiner, M., & Trinkhaus, E. (2001). Stable isotope evidence for increasing dietary breadth in the European mid-Upper Paleolithic. Proceedings of the National Academy of Sciences of the United States of America, 98, 6528–6532.
Richards, M. P., Fuller, B. T., Sponheimer, M., Robinson, T., & Ayliffe, L. (2003). Sulphur isotopes in paleodietary studies: A review and results from a controlled feeding experiment. International Journal of Osteoarchaeology, 13, 37–45.
Robbins, C. T., Felicetti, L. A., & Sponheimer, M. (2005). The effect of dietary protein quality on nitrogen isotope discrimination in mammals and birds. Oecologia, 144, 534–540.
Rodière, É., Bocherens, H., Angibault, J.-M., & Mariotti, A. (1996). Particularités isotopiques chez le chevreuil (Capreolus capreolus L.): Implications pour les reconstitutions paléoenvironnementales. Science de la terre et des planètes, 323, 179–185.
Roebroeks, W., Conard, N. J., & van Kolfschoten, T. (1992). Dense forests, cold steppes, and the Paleolithic settlement of Northern Europe. Current Anthropology, 33, 551–586.
Schnyder, H., Schwertl, M., Auerswald, K., & Schäufele, R. (2006). Hair of grazing cattle provides an integrated measure of the effects of site conditions and interannual weather variability on δ13C of temperate humid grassland. Global Change Biology, 12, 1315–1329.
Schoeninger, M. J., & DeNiro, M. J. (1984). Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochimica et Cosmochimica Acta, 48, 625–639.
Scott, K. (1980). Two hunting episodes of Middle Paleolithic Age at La Cotte de Saint-Brelade, Jersey (Channel Islands). World Archaeology, 12, 137–152.
Sealy, J. C., van der Merwe, N. J., Lee-Thorp, J. A., & Lanham, J. L. (1987). Nitrogen isotope ecology in southern Africa: Implications for environmental and dietary tracing. Geochimica et Cosmochimica Acta, 51, 2707–2717.
Simpson, I. A., Bol, R., Bull, I. D., Evershed, R. P., Petzke, K. J., & Dockrill, S. J. (1999). Interpreting early land management through compound specific stable isotope analyses of archeological soils. Rapid Communications in Mass Spectrometry, 13, 1315–1319.
Sponheimer, M., Robinson, T., Ayliffe, L., Roeder, B., Hammer, J., Passey, B., West, A., Cerling, T., Dearing, D., & Ehleringer, J. (2003). Nitrogen isotopes in mammalian herbivores: Hair δ15N values from a controlled feeding study. International Journal of Osteoarchaeology, 13, 80–87.
Stewart, J. R., van Kolfschoten, T., Markova, A., & Musil, R. (2003). The mammalian faunas of Europe during Oxygen Isotope Stage Three. In T. H. van Andel & W. Davis (Eds.), Neanderthals and modern humans in the European landscape during the last glaciation (pp. 103–130). Cambridge: McDonald Institute for Archeological Research.
Thieme, H., & Veil, S. (1985). Neue Untersuchungen zum eemzeitlichen Elefanten-Jagdplatz Lehringen, Lkr. Verden. Die Kunde, 36, 11–58.
Tieszen, L. L. (1991). Natural variations in the carbon isotope values of plants: Implications for archeology, ecology, and paleoecology. Journal of Archaeological Science, 18, 227–248.
van der Merwe, N. J., & Medina, E. (1991). The canopy effect, carbon isotope ratios and foodwebs in Amazonia. Journal of Archaeological Science, 18, 249–259.
Weaver, T. D., & Steudel-Numbers, K. (2005). Does climate or mobility explain the differences in body proportions between Neandertals and their Upper Paleolithic successors? Evolutionary Anthropology, 14, 218–223.
Wyckoff, R. W. G. (1969). Sur la composition de quelques protéines dinosauriennes. Comptes rendus de l’Academie des Sciences Paris. Série IIA. Science de la terre et des planètes, 269, 1489–1491.
Acknowledgements
I wish to warmly thank N.J. Conard andJ. Richter for their invitation to this symposium and to publish in the proceedings volume. I am indebted to the Alexander von Humboldt Foundation for the financial support of my research stay in Tübingen, during which this work was prepared. I also thank many colleagues for allowing the sampling of archeological material and for in-depth discussion about Neanderthal ecology, among which more particularly D. Bonjean, N.J. Conard, D. Drucker, M. Germonpré, S. Münzel, M. Otte, M. Patou-Mathis, and M. Toussaint. I want to thank an anonymous reviewer for helpful comments on an earlier version of this paper.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Bocherens, H. (2011). Diet and Ecology of Neanderthals: Implications from C and N Isotopes. In: Conard, N.J., Richter, J. (eds) Neanderthal Lifeways, Subsistence and Technology. Vertebrate Paleobiology and Paleoanthropology Series. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-0415-2_8
Download citation
DOI: https://doi.org/10.1007/978-94-007-0415-2_8
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-0414-5
Online ISBN: 978-94-007-0415-2
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)