
Abstract
Arbuscular mycorrhizal associations imply a remarkable reprogramming of functions in both plant and fungal symbionts. The consequent alterations on plant physiology have a clear impact on the plant response to biotic stresses. In this chapter we discuss the effects of the mycorrhizal symbiosis on plant susceptibility/resistance to potential deleterious organisms, including root and shoot pathogens, root parasitic plants and phytophagous insects, highlighting the mechanisms that may be operating in each particular case. Special attention is given to the modulation of plant defence responses in mycorrhizal systems, as it may affect all interactions. Finally we focus on the priming of jasmonate regulated plant defence mechanisms that seem to mediate the induction of resistance by arbuscular mycorrhizas.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Biotic stress
- Bioprotection
- Induced resistance
- Priming
- Plant defence
- Biocontrol
- Defence signalling
- Jasmonates
- Pathogens
- Insects
1 Introduction
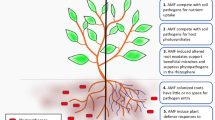
The establishment of the arbuscular mycorrhizal (AM) symbiosis implies remarkable changes in the physiology of the host plant. The changes span from alterations in the hormonal balance and transcriptional profile to altered primary and secondary metabolism (Hause et al. 2007; Liu et al. 2007; Schliemann et al. 2008; López-Ráez et al. 2010). This global reprogramming of plant functions has an impact on the plant interaction with the environment, modifying its responses to biotic and abiotic stresses. As a result, mycorrhizal plants are generally more tolerant to environmental stresses. The consequences go beyond the individual level as they may influence plant diversity and productivity in terrestrial ecosystems (van der Heijden et al. 2008).
It should be noted that the impact of the symbiosis in terms of resistance/tolerance to biotic stresses differs among AM fungal isolates for a given plant-pathogen interaction. Moreover, such impact can be modulated by environmental conditions. Despite of this variability, general trends emerge from the multiple studies dealing with mycorrhiza in diverse pathosystems. Generally, enhanced resistance to soil-borne pathogens has been reported in AM plants. Furthermore, the symbiosis can also impact plant interactions with above-ground attackers. In this case, the outcome ranges from enhanced resistance to increased susceptibility, largely depending on the attacker life-style (Pozo and Azcón-Aguilar 2007).
Early works on mycorrhizas and biotic stresses were mostly descriptive (for reviews see Schonbeck and Dehne 1989; Sharma et al. 1992; Linderman 2000). Generally, reports have focussed on beneficial effects of the symbiosis, aiming at using AM as potential biocontrol agents in integrated management programmes for disease control (Sharma and Adholeya 2000; Harrier and Watson 2004; Whipps 2004; Mukerji and Ciancio 2007).
A key factor determining the effect of the symbiosis on interactions with other organisms seems to be the extension of root colonization by the AM fungi (AMF). With some exceptions (Caron et al. 1986; García-Garrido and Ocampo 1988; St-Arnaud et al. 1997; Kapoor 2008), reports on mycorrhizal protection against pathogens show the requirement of a well established symbiosis prior to the challenge with the attacker (Rosendahl 1985; Cordier et al. 1998; Slezack et al. 2000; Khaosaad et al. 2007). The first mechanism proposed to be involved in mycorrhiza-induced protection was the improvement of plant nutrition and the consequent compensation of the damages caused by the pathogen. However, studies including nutrient supplemented controls showed that AM effects cannot be regarded as a mere consequence of improved phosphorus nutrition (Trotta et al. 1996; Fritz 2006; Liu et al. 2007). As we advance in our knowledge on the physiology and regulation of the AM symbiosis, we start to understand the diversity of mechanisms underlying the impact of the symbiosis on plant interactions with other organisms. In addition to the nutritional aspects, changes in the plant architecture, root exudation and in the microbial populations in the rhizosphere, and the activation of plant defence mechanisms may all be relevant. Their individual contribution to the final outcome will depend on the organisms involved and the timing of the interactions (Azcón-Aguilar and Barea 1996; Whipps 2004). In this chapter we will discuss the possible mechanisms affecting the different types of plant-attacker interactions, with special emphasis in those involving plant defence responses.
2 Impact of the AM Symbioses on Soil-Borne Pathogens
It is widely accepted that AM symbioses reduce the damage caused by soil-borne pathogens. Many studies revealed a reduction of the incidence and/or severity of diseases as root rot or wilting caused by diverse fungi such as Fusarium, Rhizoctonia, Macrophomina or Verticillium, bacteria as Erwinia carotovora, and oomycetes as Phytophthora, Pythium and Aphanomyces. A comprehensive review of those studies was compiled by Whipps (2004). Similarly, a reduction of the deleterious effects by parasitic nematodes such as Pratylenchus and Meloidogyne has been reported in mycorrhizal plants (Pinochet et al. 1996; de la Peña et al. 2006; Li et al. 2006). Additional reports showed protection to other soil pathogens as Armillaria melea in grapevine (Nogales et al. 2009), broadening the range of pathosystems in which AM symbioses may have a protective effect. The effectiveness against such diverse range of attackers confirms the broad spectrum character of the induced resistance associated to the AM symbiosis.
Studies comparing different fungal species or isolates highlighted that the degree of protection is highly dependent on the AMF involved (Kobra et al. 2009). Interestingly, many studies point to a higher protector effect of Glomus mosseae in comparison to other AMF (Pozo et al. 2002; Utkhede 2006; Ozgonen and Erkilic 2007).
Several mechanisms may operate simultaneously in the enhanced resistance of mycorrhizal plants to soil pathogens. In addition to a possible competition for photosynthates between the AMF and the pathogen, competition for colonization sites has been demonstrated. For example, in tomato roots, full exclusion of Phytophthora from arbusculated cells was evidenced (Cordier et al. 1998). Mycorrhizal colonization is also known to induce changes in the root system architecture and morphology (Schellenbaum et al. 1991; Norman et al. 1996). These changes may alter the dynamics of infection by the pathogen, although direct evidences of such correlation are lacking. An altered pattern of root exudation may also impact the development of the pathogen. Mycorrhizal colonization leaded to modifications in root exudates composition that significantly reduced the sporulation of Phytophthora fragariae (Norman and Hooker 2000) and altered the chemotactic response of the zoospores of Phytophthora nicotianae (Lioussanne et al. 2008). Since root exudates are key factors in shaping soil microbial communities (Badri and Vivanco 2009), the changes in exudation into the mycorrhizosphere may result in alteration of the microbial communities including possible antagonistic organisms. This may be the reason underlying the biocontrol of pathogens in non-AM species by co-culture with mycorrhizal plants (St-Arnaud et al. 1997).
Because of the root localization of both attacker and AMF, it is difficult to discern the local or systemic character of the protection observed. However, the use of split-root experimental systems allowing physical separation between AMF and pathogens has confirmed a reduction of disease symptoms in the non-mycorrhizal parts of the mycorrhizal root systems. Systemic protection at the root level has been demonstrated against Phytophthora and Ralstonia in tomato (Cordier et al. 1998; Pozo et al. 2002; Zhu and Yao 2004), against Gaeumannomyces in wheat (Khaosaad et al. 2007), and against parasitic nematodes in banana plants (Elsen et al. 2008). The systemic character of the induced resistance pointed to the involvement of plant defences. Because of its relevance in all kinds of interactions, the implication of plant defence mechanisms will be discussed in Section 7.
3 Effects of AM Symbioses on Root Parasitic Plants
Plants of the genera Striga and Orobanche parasitize different hosts around the world, constituting one of the most damaging agricultural pests. These obligate parasites attach to the roots of many plant species and acquire nutrients and water from their host (Bouwmeester et al. 2003). Studies in African fields infested with the hemiparasite Striga hermonthica evidenced that inoculation with AMF significantly reduced the amount of parasitic plants in maize and sorghum cultures. Accordingly, the use of mycorrhizas for integrated management of parasitic weeds was proposed (Lendzemo et al. 2005; López-Ráez et al. 2009a).
Strigolactones are germinating stimulants for the seeds of root parasitic plants (Bouwmeester et al. 2007). With the discovery of strigolactones as host detection signals for AMF in the rhizosphere (Akiyama et al. 2005; see chapter by Bécard et al.) a causal connection between AM and its impact on parasitic plants could be established. Indeed, follow up experiments under controlled conditions confirmed that AMF inoculation leads to a reduction of the impact of Striga, apparently related to a reduction in strigolactones production (Lendzemo et al. 2007). Similarly, we have observed that extracts from tomatoes colonized by G. mosseae induce less germination of Orobanche ramosa seeds than those from non-mycorrhizal plants (López-Ráez et al. 2009b). Moreover, a reduced production of strigolactones in a tomato mutant correlated with less susceptibility to Orobanche (López-Ráez et al. 2008). All in all, it seems likely that a reduction in strigolactone production underlies the decrease in the incidence of root parasitic plants on mycorrhizal plants.
4 Impact of AM Symbioses on Above-Ground Interactions
Studies dealing with AM effects on above-ground diseases are less abundant, and apparently less conclusive. Early reports associated AM symbioses with enhanced susceptibility to viruses (Whipps 2004), and it was generally accepted that AM plants are more susceptible to shoot pathogens. However, recent studies dealing with pathogens of different life styles have evidenced a more complex reality. Biotrophic pathogens, such as powdery mildew and rust fungi (Blumeria, Oidium, Uromyces) seem to perform better in mycorrhizal plants, although increased tolerance was often observed in terms of plant mass and yield (Gernns et al. 2001; Whipps 2004). Concerning hemibiotrophs, the impact of the symbiosis varies from no effect to reduction of the disease, for example, against Colletotrichum orbiculare in cucumber (Lee et al. 2005; Chandanie et al. 2006). Finally, several studies evidenced a positive effect on plant resistance to other shoot pathogens, including the bacteria Xanthomonas campestris in Medicago (Liu et al. 2007), and the necrotrophic fungus Alternaria solani in tomato (Fritz et al. 2006; De La Noval et al. 2007). We have also confirmed that symbiosis with G. mosseae in tomato induces systemic resistance to the necrotrophic fungus Botrytis cinerea (Jung et al., 2009) (see Section 7). Recently, a positive effect of G. mosseae against Botrytis cinerea has also been shown in roses, although dependent on environmental factors (Moller et al. 2009).
Phytoplasma are specialized obligate parasites of phloem tissue transmitted by insect vectors. AM establishment in tomato lead to a reduction of the disease symptoms caused by a phytoplasma of the Stolbur group (Lingua et al. 2002). Because they inoculated through grafting with infected scions, potential effects on the insect vector were ruled out. Thus, the protection is related to physiological changes in the mycorrhizal plant. Tolerance to phytoplasma disease was also reported in pear (García-Chapa et al. 2004). Recently, a reduction in the titre of the Chrysanthemum yellows phytoplasma has been shown in mycorrhizal chrysanthemum (D’Amelio et al. 2007), confirming that mycorrhization can confer resistance to this type of pathogens.
In above-ground interactions of mycorrhizal plants, two main mechanisms may be operative. One would be the potential changes in nutrient levels of the host plant and alterations of the source-sink relation within it, that may affect the suitability of the plant for shoot attackers. The other would be the modulation of plant defence mechanisms, as discussed in Section 7.
5 Effects of AM Symbioses on Phytophagous Insects
The mycorrhizal status of the host plant can also influence insect herbivore performance, but the magnitude and direction of the effect depend upon the feeding mode and life style of the insect (Hartley and Gange 2009; Koricheva et al. 2009).
Many different studies cover an ample range of mycorrhizal plant-insect interactions under controlled or field conditions. Upon a comprehensive review of the published data, Hartley and Gange (2009) concluded that, generally, mycorrhizas have strong negative effects on rhizophagous insects, but effects on shoot-feeding insects are weaker and more variable. Despite of this variability, some general patterns emerge: generalist insects are usually adversely affected by mycorrhizas, whereas specialist insects may often benefit. Furthermore, aphids usually perform better on AM plants while leaf-chewing insects are usually negatively affected by the symbiosis.
Such patterns may arise from the differential impact of nutritional and defence aspects in the insect. While generalist insects are sensitive to plant defence mechanisms, specialist herbivores are likely to be able to circumvent the defences of their host and remain undetected. As a result, generalists may be affected by the enhanced defence capacity of AM plants, while specialists will circumvent the defences and may benefit from the improved nutritional status of the plant. The negative effect on leaf-chewers is likely related to their susceptibility to jasmonate-dependent defences (Peña-Cortés et al. 2004) potentiated in mycorrhizal plants (see Section 8). In addition, AM can also have an impact on herbivores by affecting the performance of their predators and parasitoids: in tomato, the volatile blends released by AM plants can be more attractive to aphid parasitoids than those from non-mycorrhizal ones (Guerrieri et al. 2004).
6 AM Symbiosis Modulate Host Defence Responses
As discussed earlier in this book and reviewed elsewhere (Gianinazzi-Pearson 1996; Harrison 2005; Parniske 2008) the establishment of a successful mutualistic interaction requires a high degree of coordination between both partners. Plant and fungus actively engage in the process of colonization, and a tight control of plant defence mechanisms is necessary. Interestingly, the plant is able to restrict AMF colonization once plants are already mycorrhizal, a phenomenon known as autoregulation (Vierheilig et al. 2008). The mechanisms operating in such autoregulation may also impact plant interactions with pathogens.
The levels of several phytohormones (mainly salicylic acid (SA), jasmonates (JAs), ethylene (ET) and abcisic acid (ABA)) fine-tune the defence responses in plants through an intricate regulatory network (Pieterse et al. 2009). Remarkably, the levels of these hormones seem to be altered in mycorrhizal plants (Hause et al. 2007; López-Ráez et al. 2010), probably affecting plant defence mechanisms. There is evidence for the accumulation of defensive plant compounds in mycorrhizal roots, although to a much lower extent than in plant-pathogen interactions. Activation of phenylpropanoid and oxylipin metabolism, accumulation of reactive oxygen species and of specific isoforms of defence-related enzymes has been reported in mycorrhizal roots (García-Garrido and Ocampo 2002; De Deyn et al. 2009; López-Ráez et al. 2010). These reactions, generally localized, may control the development of the fungus inside the roots (Pozo et al. 2002; Dumas-Gaudot et al. 2000; García-Garrido and Ocampo 2002). Indeed, as obligate biotrophs, AMF share similarities with biotrophic pathogens (Paszkowski 2006) and transcriptional profiling of plant responses to AMF revealed some overlap with responses to biotrophic pathogens (Güimil et al. 2005). Coherently, SA, a key regulator of plant defences against biotrophs (Glazebrook 2005), seems to have a negative effect on AM colonization (García-Garrido and Ocampo 2002; López-Ráez et al. 2010). Thus, it is plausible that AMF repress SA-dependent responses in the host in order to achieve a compatible interaction. Indeed, a delay in the accumulation of PR-1 proteins, common markers of SA-dependent responses, has been observed in mycorrhizal roots (Dumas-Gaudot et al. 2000). Even repression of defence responses triggered upon pathogen attack has been reported when G. intraradices was co-inoculated with Rhizoctonia solani (Guenoune et al. 2001). Although AMF are able to trigger plant defence responses as evidenced in myc mutants, only weak and transient defence responses are activated during compatible AM interactions (Liu et al. 2003). Thus, AM establishment seems to require inhibition of certain SA-regulated defence responses. Remarkably, inhibition of SA responses is also necessary for the Rhizobium–legume symbiosis (Stacey et al. 2006).
Modulation of plant defences during AM formation does not only occur in the roots, but also in the shoots. Accumulation of insect anti-feedant compounds (Gange 2006; Pozo et al. 2009) and transcriptional up-regulation of defence-related genes (Liu et al. 2007; Pozo et al. 2009) have been described in leaves of mycorrhizal plants. Also a repression of certain defences may take place: a delay in the systemic accumulation of PR1 upon treatment with SA or analogs has been reported in mycorrhizal tobacco shoots (Shaul et al. 1999) and suppression of certain chemical defences has also been reported (Bennett et al. 2009). This modulation may affect the interaction with shoot attackers. To this regard, the reciprocal influence of below-ground and above-ground interactions through their impact on plant defences is receiving increasing interest (Bezemer and van Dam 2005; Erb et al. 2009). An additional level of complexity is related to the altered volatile profile released by AM plants (Guerrieri et al. 2004; Rapparini et al. 2008). Volatiles may play key roles in defence, for example, by attracting natural enemies of potentially harmful insects, or by priming distal parts of the plant for a more efficient activation of defences (Heil and Ton 2008).
We have recently shown that mycorrhizal colonization in tomato leads to increases in the expression of defence related genes known to be regulated by JA (Pozo et al. 2009). JA is a key regulator of plant defences against insects and necrotrophic pathogens (Peña-Cortés et al. 2004; Pozo et al. 2005). Because of SA and JA signalling pathways are interconnected, mostly in an antagonistic way (Pieterse et al. 2009) their interplay may explain the pattern of enhanced resistance/susceptibility of AM plants. If AM inhibits SA-regulated responses, the plant would be more susceptible to pathogens resisted through these responses, i.e. biotrophic pathogens. On the contrary, an induction of the JA signalling pathway would make mycorrhizal plants more resistant to necrotrophic pathogens and JA-sensitive insects (Pozo and Azcón-Aguilar 2007). Such pattern is more obvious in shoot interactions, where modulation of plant defences seems to be the main mech-anism. In roots, the relevance of this altered balance will be lower since other mechanisms are operating simultaneously (see Section 3), and a reduction of the disease is the most general outcome.
7 AM Symbiosis Primes JA-Dependent Responses
Upon detection of a potential attacker, a rapid and strong activation of the defence mechanisms is crucial for resistance. Accordingly, pre-conditioning of plant tissues for a quick and more effective activation of defences upon attack has important ecological fitness benefits, and seems to be a common feature of the plant’s immune system. This boost of basal defences is known as priming (Conrath et al. 2006; Goellner and Conrath 2008). Priming seems to be the strategy followed by several beneficial micro-organisms to enhance resistance in plants, avoiding a direct activation of defences which would be too expensive for the host in the absence of challenging attackers (Pozo et al. 2005; Van Wees et al. 2008).
Evidences for primed defence responses in mycorrhizas were first reported in root tissues. Mycorrhizal transformed carrot roots displayed stronger defence reactions at challenge sites by Fusarium (Benhamou et al. 1994). Similarly, mycorrhizal potatoes showed amplified accumulation of phytoalexins upon Rhizoctonia infection (Yao et al. 2003). Priming for callose deposition seems to be responsible for the protection achieved by G. intraradices against Colletotrichum in cucumber (Lee et al. 2005). Recently, primed accumulation of phenolic compounds in AM date palm trees has also been related to protection against F. oxysporum (Jaiti et al. 2008). Remarkably, priming is not restricted to AMF colonized areas of the roots, but to the whole root system. This was first illustrated in tomato plants during P. parasitica infection (Cordier et al. 1998; Pozo et al. 2002). Only AM plants, even in non-mycorrhizal parts of the root system, formed papilla-like structures around the sites of pathogen infection, preventing further spreading of the pathogen. They also accumulated more PR-proteins than non-mycorrhizal plants upon challenge (Cordier et al. 1998; Pozo et al. 1999). Mycorrhizal protection of grapevine against Meloidogyne incognita has also been associated with primed systemic expression of a chitinase gene in response to the nematode (Li et al. 2006). But the primed response is not restricted to the root system. Recently, we have shown priming of defences also in shoots of mycorrhizal plants (Pozo et al. 2009).
Evidence is accumulating that priming associated to systemic resistance induced by beneficial organisms is regulated by similar jasmonate signalling pathways (Van Wees et al. 2008). Indeed, studies on rhizobacteria induced systemic resistance (ISR) in Arabidopsis revealed the requirement of a functional JA signalling pathway for the efficient induction of resistance (Pieterse et al. 1998; Pozo et al. 2008). The JA signalling pathway is also required for rhizobacteria ISR in tomato (Yan et al. 2002) and for the induction of resistance by the beneficial fungi Trichoderma and Piriformospora (Shoresh et al. 2005; Stein et al. 2008). Interestingly, JA accumulation have been proposed to mediate plant “memory” of previous challenges (Galis et al. 2009), a possible basis for the primed state.
Jasmonates are key regulators in the AM symbiosis, and elevated endogenous levels of JA have been confirmed in mycorrhizal roots (reviewed in Hause et al. 2007 and Hause and Schaarschmidt, 2009). We have found a significant increase in JAs in mycorrhizal tomato roots (López-Ráez et al. 2010), but the levels were not altered in the shoots (López-Ráez and Pozo, unpublished). However, we found small, yet significant, increases in the expression of marker genes for JA responses, a result that may indicate an enhanced sensitivity to the hormone.
To confirm whether AM leads to priming of JA-dependent responses in the shoots, we compared the response of non-mycorrhizal and AM plants to foliar application of different defence-related stimuli. Transcript profiling of leaves 24 h after treatment with JA revealed a stronger induction of JA-regulated genes in mycorrhizal plants, particularly in G. mosseae-colonized plants, confirming a primed response in those plants (Pozo et al. 2009). To address the biological relevance of such primed response, we tested whether priming occurs during interactions with pathogens. AM and non-mycorrhizal plants were challenged with B. cinera, and the expansion of necrotic lesions was markedly lower in the leaves of AM plants (Fig. 1a). The levels of pathogen in the tissues were also lower in mycorrhizal plants at all time points tested (Fig. 1b), confirming induced systemic resistance. Remarkably, this pattern inversely correlated with the expression of the defence-related, JA-marker gene Pin II. Pin II expression was induced in response to B. cinerea in all plants, but this induction was markedly higher in AM plants (Jung et al. 2009). The results support that Mycorrhiza Induced Resistance (MIR) to Botrytis is associated to priming of JA-dependent defences.

Mycorrhizal tomato plants are more resistant to the necrotrophic foliar pathogen Botrytis cinerea. Left – Photograph of necrotic lesions developed 6 days after drop inoculation of tomato leaves with Botrytis cinerea . Right – Quantification of Botrytis cinerea in infected leaves by real time PCR analysis of expression of its tubuline gene. Nm: Non mycorrhizal plants, Gm: Glomus mosseae-colonized plants
8 Conclusions and Outlook
AM symbiosis affect the outcome of the host plant interaction with other organisms. Although the effect is influenced by plant, AMF and environmental factors, generally there is a clear protector effect of the symbiosis over soil-borne deleterious organisms. However, the impact on shoot interactions is more variable and relies largely on the attacker lifestyle. Besides nutritional effects and local changes in the plant (affecting mainly soil-borne pathogens), modulation of plant defences associated to AM seems to play a crucial role. Instead of constitutive activation of defences, MIR is mediated by priming for an efficient activation of defences. Experimental evidences and the general spectrum of protection by mycorrhiza point to a central role of jasmonates in MIR. Despite of the relevance of this process, its precise regulation awaits elucidation. Increasing our knowledge on the modifications of plant physiology in AM, as well as in the biology of the potential attackers is essential in order to define markers of induced resistance and to generate predictive models for the outcome of particular mycorrhiza-pathogen interactions. Another challenge ahead is to decipher the connections in plant responses to biotic and abiotic stresses. Experimental evidences point to common regulatory nodes in the signalling pathways governing responses to both types of stresses, and those nodes may be the target of biotechnological strategies for optimization of plant protection by arbuscular mycorrhizas. Finally, it is important to consider mycorrhiza in a multitrophic context, as the impact of the symbiosis on plant interactions can be modified by other organisms in the system.
Abbreviations
- AM:
-
Arbuscular mycorrhiza
- AMF:
-
Arbuscular mycorrhizal fungi
- MIR:
-
mycorrhiza-induced resistance
- JA:
-
jasmonate
- SA:
-
Salicylic acid
- Nm:
-
Non-mycorrhizal plants
- Gm:
-
Glomus mosseae colonized plants
- Hpi:
-
hours post inoculation
References
Akiyama K, Matsuzaki K, Hayashi H (2005) Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 435:824–827
Azcón-Aguilar C, Barea JM (1996) Arbuscular mycorrhizas and biological control of soil-borne plant pathogens – an overview of the mechanisms involved. Mycorrhiza 6:457–464
Badri DV, Vivanco JM (2009) Regulation and function of root exudates. Plant Cell Environ 32:666–681
Benhamou N, Fortin JA, Hamel C, St-Arnaud M, Shatilla A (1994) Resistance responses of mycorrhizal Ri T-DNA-transformed carrot roots to infection by Fusarium oxysporum f sp chrysanthemi. Phytopathol 84:958–968
Bennett A, Bever J, Deane Bowers M (2009) Arbuscular mycorrhizal fungal species suppress inducible plant responses and alter defensive strategies following herbivory. Oecologia 160:771–779
Bezemer TM, van Dam NM (2005) Linking aboveground and belowground interactions via induced plant defenses. Trends Ecol Evol 20:617–624
Bouwmeester HJ, Matusova R, Zhongkui S, Beale MH (2003) Secondary metabolite signalling in host-parasitic plant interactions. Curr Opin Plant Biol 6:358–364
Bouwmeester HJ, Roux C, López-Raez JA, Becard G (2007) Rhizosphere communication of plants, parasitic plants and AM fungi. Trends Plant Sci 12:224–230
Caron M, Fortin JA, Richard C (1986) Effect of inoculation sequence on the interaction between Glomus intraradices and Fusarium oxysporum f. sp. radicis-lycopersici in tomatoes. Can J Plant Pathol 8:12–16
Conrath U, Beckers GJM, Flors V, García-Agustín P, Jakab G, Mauch F, Newman MA, Pieterse CMJ, Poinssot B, Pozo MJ, Pugin A, Schaffrath U, Ton J, Wendehenne D, Zimmerli L, Mauch-Mani B (2006) Priming: getting ready for battle. Mol Plant-Microbe Interact 19:1062–1071
Cordier C, Pozo MJ, Barea JM, Gianinazzi S, Gianinazzi-Pearson V (1998) Cell defense responses associated with localized and systemic resistance to Phytophthora induced in tomato by an arbuscular mycorrhizal fungus. Mol Plant-Microbe Interact 11:1017–1028
Chandanie WA, Kubota M, Hyakumachi M (2006) Interactions between plant growth promoting fungi and arbuscular mycorrhizal fungus Glomus mosseae and induction of systemic resistance to anthracnose disease in cucumber. Plant Soil 286:209–217
D’Amelio R, Massa M, Gamalero E, D’Agostino G, Sampò S, Berta G, Faoro F, Iriti M, Bosco D, Marzachì C (2007) Preliminary results on the evaluation of the effects of elicitors of plant resistance on chrysanthemum yellows phytoplasma infection. Bull Insectol 60:317–318
De Deyn GB, Biere A, Van Der Putten WH, Wagenaar R, Klironomos JN (2009) Chemical defense, mycorrhizal colonization and growth responses in Plantago lanceolata L. Oecologia 160:433–442
De La Noval B, Perez E, Martinez B, Leon O, Martinez-Gallardo N, Delano-Frier J (2007) Exogenous systemin has a contrasting effect on disease resistance in mycorrhizal tomato (Solanum lycopersicum) plants infected with necrotrophic or hemibiotrophic pathogens. Mycorrhiza 17:449–460
de la Peña E, Rodriguez-Echeverria S, van der Putten WH, Freitas H, Moens M (2006) Mechanism of control of root-feeding nematodes by mycorrhizal fungi in the dune grass Ammophila arenaria. New Phytol 169:829–840
Dumas-Gaudot E, Gollotte A, Cordier C, Gianinazzi S, Gianinazzi-Pearson V (2000) Modulation of host defence systems. In: Kapulnik Y, Douds DDJ (eds) Arbuscular mycorrhizas: physiology and function. Kluwer, The Netherlands, pp 173–200
Elsen A, Gervacio D, Swennen R, De Waele D (2008) AMF-induced biocontrol against plant parasitic nematodes in Musa sp.: a systemic effect. Mycorrhiza 18:251–256
Erb M, Flors V, Karlen D, Lange E, Planchamp C, D’Alessandro M, Turlings TCJ, Ton J (2009) Signal signature of aboveground-induced resistance upon belowground herbivory in maize. Plant J 59:292–302
Fritz M, Jakobsen I, Lyngkjaer MF, Thordal-Christensen H, Pons-Kuehnemann J (2006) Arbuscular mycorrhiza reduces susceptibility of tomato to Alternaria solani. Mycorrhiza 16:413–419
Galis I, Gaquerel E, Pandey SP, Baldwin IT (2009) Molecular mechanisms underlying plant memory in JA-mediated defense responses. Plant Cell Environ 32:617–627
Gange AC (2006) Insect-mycorrhizal interactions: patterns, processes, and consequences. In: Ohgushi T, Craig TP, Price PW (eds) Indirect interaction webs: nontrophic linkages through induced plant traits. Cambridge University Press, Cambridge, pp 124–144
García-Chapa M, Batlle A, Laviña A, Camprubí A, Estaún V, Calvet C (2004) Tolerance increase to pear decline phytoplasma in mycorrhizal OHF-333 pear rootstock. Acta Hort 657:437–441
Garcia Garrido JM, Ocampo JA (1988) Interaction between Glomus mosseae and Erwinia carotovora and its effects on the growth of tomato plants. New Phytol 110:551–555
García-Garrido JM, Ocampo JA (2002) Regulation of the plant defence response in arbuscular mycorrhizal symbiosis. J Exp Bot 53:1377–1386
Gernns H, von Alten H, Poehling HM (2001) Arbuscular mycorrhiza increased the activity of a biotrophic leaf pathogen – is a compensation possible? Mycorrhiza 11:237–243
Gianinazzi-Pearson V (1996) Plant cell responses to arbuscular mycorrhizal fungi: Getting to the roots of the symbiosis. Plant Cell 8:1871–1883
Glazebrook J (2005) Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Ann Rev Phytopathol 43:205–227
Goellner K, Conrath U (2008) Priming: it’s all the world to induced disease resistance. Eur J Plant Pathol 121:233–242
Guenoune D, Galili S, Phillips DA, Volpin H, Chet I, Okon Y, Kapulnik Y (2001) The defense response elicited by the pathogen Rhizoctonia solani is suppressed by colonization of the AM-fungus Glomus intraradices. Plant Sci 160:925
Guerrieri E, Lingua G, Digilio MC, Massa N, Berta G (2004) Do interactions between plant roots and the rhizosphere affect parasitoid behaviour? Ecol Entomol 29:753–756
Güimil S, Chang HS, Zhu T, Sesma A, Osbourn A, Roux C, Ioannidis V, Oakeley EJ, Docquier M, Descombes P, Briggs SP, Paszkowski U (2005) Comparative transcriptomics of rice reveals an ancient pattern of response to microbial colonization. Proc Nat Acad Sci 102:8066–8070
Harrier LA, Watson CA (2004) The potential role of arbuscular mycorrhizal (AM) fungi in the bioprotection of plants against soil-borne pathogens in organic and/or other sustainable farming systems. Pest Manage Sci 60:149–157
Harrison MJ (2005) Signaling in the arbuscular mycorrhizal symbiosis. Ann Rev Microbiol 59:19–42
Hartley SE, Gange AC (2009) Impacts of plant symbiotic fungi on insect herbivores: mutualism in a multitrophic context. Ann Rev Entomol 54:323–342
Hause B, Mrosk C, Isayenkov S, Strack D (2007) Jasmonates in arbuscular mycorrhizal interactions. Phytochem 68:101–110
Hause B, Schaarschmidt S (2009) The role of jasmonates in mutualistic symbioses between plants and soil-born microorganisms. Phytochem 70:1589–1599
Heil M, Ton J (2008) Long-distance signalling in plant defence. Trends Plant Sci 13:264–272
Jaiti F, Kassami M, Meddich A, El Hadrami I (2008) Effect of arbuscular mycorrhization on the accumulation of hydroxycinnamic acid derivatives in date palm seedlings challenged with Fusarium oxysporum f. sp. albedinis. J Phytopathol 156:641–646
Jung SC, García-Andrade J, Verhage A, Fernández I, García JM, Azcón-Aguilar C, Pozo MJ (2009) Arbuscular mycorrhiza confers systemic resistance against Botrytis cinerea in tomato through priming of JA-dependent defense responses. In: Mauch-Mani B, Schmidt A (eds) Induced resistance: chances and limits, IOBC/wprs Bulletin, Working Group “Induced resistance in plants against insects and diseases.” Proceedings of the meeting at Granada (Spain) 8–16 May 2009 (in press)
Kapoor R (2008) Induced resistance in mycorrhizal tomato is correlated to concentration of jasmonic acid. OnLine J Biol Sci 8:49–56
Khaosaad T, García-Garrido JM, Steinkellner S, Vierheilig H (2007) Take-all disease is systemically reduced in roots of mycorrhizal barley plants. Soil Biol Biochem 39:727–734
Kobra N, Jalil K, Youbert G (2009) Effects of three Glomus species as biocontrol agents against verticillium-induced wilt in cotton. J Plant Protect Res 49:185–189
Koricheva J, Gange AC, Jones T (2009) Effects of mycorrhizal fungi on insect herbivores: a meta-analysis. Ecology 90:2088–2097
Lee CS, Lee YJ, Jeun YC (2005) Observations of infection structures on the leaves of cucumber plants pre-treated with arbuscular mycorrhiza Glomus intraradices after challenge inoculation with Colletotrichum orbiculare. Plant Pathol J 21:237–243
Lendzemo VW, Kuyper TW, Kropff MJ, van Ast A (2005) Field inoculation with arbuscular mycorrhizal fungi reduces Striga hermonthica performance on cereal crops and has the potential to contribute to integrated Striga management. Field Crops Res 91:51–61
Lendzemo VW, Kuyper TW, Matusova R, Bouwmeester HJ, van Ast A (2007) Colonization by arbuscular mycorrhizal fungi of sorghum leads to reduced germination and subsequent attachment and emergence of Striga hermonthica. Plant Signal Behav 2:58–62
Li HY, Yang GD, Shu HR, Yang YT, Ye BX, Nishida I, Zheng CC (2006) Colonization by the arbuscular mycorrhizal fungus Glomus versiforme induces a defense response against the root-knot nematode Meloidogyne incognita in the grapevine (Vitis amurensis Rupr.), which includes transcriptional activation of the class III chitinase gene VCH3. Plant Cell Physiol 47:154–163
Linderman RG (2000) Effects of mycorrhizas on plant tolerance to diseases. In: Kapulnik Y, Douds DDJ (eds) Arbuscular mycorrhizas: physiology and function. Kluwer, Dordrecht, The Netherlands, pp 345–365
Lingua G, D’Agostino G, Massa N, Antosiano M, Berta G (2002) Mycorrhiza-induced differential response to a yellows disease in tomato. Mycorrhiza 12:191–198
Lioussanne L, Jolicoeur M, St-Arnaud M (2008) Mycorrhizal colonization with Glomus intraradices and development stage of transformed tomato roots significantly modify the chemotactic response of zoospores of the pathogen Phytophthora nicotianae. Soil Biol Biochem 40:2217–2224
Liu J, Blaylock LA, Endre G, Cho J, Town CD, Van den Bosch KA, Harrison MJ (2003) Transcript profiling coupled with spatial expression analyses reveals genes involved in distinct developmental stages of an arbuscular mycorrhizalsymbiosis. Plant Cell 15:2106–2123
Liu J, Maldonado-Mendoza I, Lopez-Meyer M, Cheung F, Town CD, Harrison MJ (2007) Arbuscular mycorrhizal symbiosis is accompanied by local and systemic alterations in gene expression and an increase in disease resistance in the shoots. Plant J 50:529–544
López-Ráez JA, Charnikhova T, Mulder P, Kohlen W, Bino R, Levin I, Bouwmeester H (2008) Susceptibility of the tomato mutant high pigment-2 dg (hp-2 dg) to Orobanche spp infection. J Agric Food Chem 56:6326–6332
López-Ráez JA, Matusova R, Cardoso C, Jamil M, Charnikhova T, Kohlen W, Verstappen F, Ruyter-Spira C, Bouwmeester HJ (2009a) Strigolactones: ecological significance and use as a target for parasitic plant control. Pest Manag Sci 64:471–477
López-Ráez JA, Jung SC, Fernandez I, García JM, Bouwmeester H, Pozo MJ (2009b) Mycorrhizal symbiosis as a strategy for root parasitic weed control. In: Mauch-Mani B, Schmidt A (eds) Induced resistance: chances and limits, IOBC/wprs Bulletin, Working Group “Induced resistance in plants against insects and diseases.” Proceedings of the meeting at Granada (Spain) 8–16 May 2009 (in press)
López-Ráez JA, Verhage A, Fernández I, García JM, Azcón-Aguilar C, Flors V, Pozo MJ (2010) Hormonal and transcriptional profiles highlight common and differential host responses to arbuscular mycorrhizal fungi and the regulation of the oxylipin pathway. J Exp Bot 61: 2589–2601
Moller K, Kristensen K, Yohalem D, Larsen J (2009) Biological management of gray mold in pot roses by co-inoculation of the biocontrol agent Ulocladium atrum and the mycorrhizal fungus Glomus mosseae. Biol Control 49:120–125
Mukerji KG, Ciancio A (2007) Mycorrhizae in the integrated pest and disease management. In: Ciancio A, Mukerji KG (eds) General concepts in integrated pest and disease management. Springer, The Netherlands, pp 245–266
Nogales A, Aguirreolea J, Santa María E, Camprubí A, Calvet C (2009) Response of mycorrhizal grapevine to Armillaria mellea inoculation: Disease development and polyamines. Plant Soil 31:177–187
Norman JR, Atkinson D, Hooker JE (1996) Arbuscular mycorrhizal fungal-induced alteration to root architecture in strawberry and induced resistance to the root pathogen Phytophthora fragariae. Plant Soil 185:191–198
Norman JR, Hooker JE (2000) Sporulation of Phytophthora fragariae shows greater stimulation by exudates of non-mycorrhizal than by mycorrhizal strawberry roots. Mycol Res 104:1069–1073
Ozgonen H, Erkilic A (2007) Growth enhancement and Phytophthora blight (Phytophthora capsici Leonian) control by arbuscular mycorrhizal fungal inoculation in pepper. Crop Protection 26:1682–1688
Parniske M (2008) Arbuscular mycorrhiza: the mother of plant root endosymbioses. Nat Rev Micro 6:763–775
Paszkowski U (2006) Mutualism and parasitism: the yin and yang of plant symbioses. Curr Opin Plant Biol 9:364–370
Peña-Cortés H, Barrios P, Dorta F, Polanco V, Sánchez C, Sánchez E, Ramírez I (2004) Involvement of jasmonic acid and derivatives in plant response to pathogen and insects and in fruit ripening. J Plant Growth Regul 23:246–260
Pieterse CMJ, León-Reyes A, Van der Ent S, Van Wees SCM (2009) Networking by small-molecule hormones in plant immunity. Nat Chem Biol 5:308–316
Pieterse CMJ, Van Wees SCM, Van Pelt JA, Knoester M, Laan R, Gerrits H, Weisbeek PJ, Van Loon LC (1998) A novel signaling pathway controlling induced systemic resistance in Arabidopsis. Plant Cell 10:1571–1580
Pinochet J, Calvet C, Camprubi A, Fernandez C (1996) Interactions between migratory endoparasitic nematodes and arbuscular mycorrhizal fungi in perennial crops – A review. Plant Soil 185:183–190
Pozo MJ, Azcón-Aguilar C, Dumas-Gaudot E, Barea JM (1999) ß-1, 3-glucanase activities in tomato roots inoculated with arbuscular mycorrhizal fungi and/or Phytophthora parasitica and their possible involvement in bioprotection. Plant Sci 141:149–157
Pozo MJ, Cordier C, Dumas-Gaudot E, Gianinazzi S, Barea JM, Azcón-Aguilar C (2002) Localized vs systemic effect of arbuscular mycorrhizal fungi on defence responses to Phytophthora infection in tomato plants. J Exp Bot 53:525–534
Pozo MJ, Van Loon LC, Pieterse CMJ (2005) Jasmonates – Signals in plant-microbe interactions. J Plant Growth Regul 23:211–222
Pozo MJ, Azcón-Aguilar C (2007) Unravelling mycorrhiza induced resistance. Curr Opin Plant Biol 10:1–6
Pozo MJ, Van Der Ent S, Van Loon LC, Pieterse CMJ (2008) Transcription factor MYC2 is involved in priming for enhanced defense during rhizobacteria-induced systemic resistance in Arabidopsis thaliana. New Phytol 180:511–523
Pozo MJ, Verhage A, García-Andrade J, García JM, Azcón-Aguilar C (2009) Priming plant defences against pathogens by arbuscular mycorrhizal fungi. In: Azcón-Aguilar C, Barea JM, Gianinazzi S, Gianinazzi-Pearson V (eds) Mycorrhizas: functional processes and ecological impact. Springer, Heidelberg, pp 123–136
Rapparini F, Llusia J, Peñuelas J (2008) Effect of arbuscular mycorrhizal colonization on terpene emission and content of Artemisia annua L. Plant Biol 10:108–122
Rosendahl S (1985) Interactions between the vesicular-arbuscular my-corrhizal fungus Glomus fasciculatum and Aphanomyces euteiches root rot of peas. Phytopathol Z 114:31–40
Schellenbaum L, Berta G, Ravolanirina F, Tisserant B, Gianinazzi S, Fitter AH (1991) Influence of endomycorrhizal infection on root morphology in a micropropagated woody plant species (Vitis vinifera L.). Biol J Linne Soc 68:135–141
Schliemann W, Ammer C, Strack D (2008) Metabolite profiling of mycorrhizal roots of Medicago truncatula. Phytochem 69:112–146
Schonbeck F, Dehne HW (1989) VA-mycorrhiza and plant health Interrelationships between microorganisms and plants in soil. Proc Symp Liblice 1987:83–91
Shaul O, Galili S, Volpin H, Ginzberg I, Elad Y, Chet I, Kapulnik Y (1999) Mycorrhiza-induced changes in disease severity and PR protein expression in tobacco leaves. Mol Plant-Microbe Interact 12:1000–1007
Sharma MP, Adholeya A (2000) Sustainable management of arbuscular mycorrhizal fungi in the biocontrol of soil-borne plant diseases. In: Upadhyay RK, Mukerji KG, Chamola BP (eds) Biocontrol potential and its exploitation in sustainable agriculture. Vol. I. Crop diseases. Kluwer/Plenum, New York, pp 117–138
Sharma AK, Johri BN, Gianinazzi S (1992) Vesicular-arbuscular mycorrhizae in relation to plant disease. World J Microbiol Biotechnol 8:559–563
Shoresh M, Yedidia I, Chet I (2005) Involvement of jasmonic acid/ethylene signaling pathway in the systemic resistance induced in cucumber by Trichoderma asperellum T203. Phytopathol 95:76–84
Slezack S, Dumas-Gaudot E, Paynot M, Gianinazzi S (2000) Is a fully established arbuscular mycorrhizal symbiosis required for bioprotection of Pisum sativum roots against Aphanomyces euteiches? Mol Plant-Microbe Interact 13:238–241
Stacey G, McAlvin CB, Kim SY, Olivares J, Soto MJ (2006) Effects of endogenous salicylic acid on nodulation in the model legumes Lotus japonicus and Medicago truncatula. Plant Physiol 141:1473–1481
St-Arnaud M, Hamel C, Vimard B, Caron M, Fortin JA (1997) Inhibition of Fusarium oxysporum f.sp. dianthi in the non-Vam species Dianthus caryophyllus by co-culture with Tagetes patula companion plants colonized by Glomus intraradices. Can J Bot 75:998–1005
Stein E, Molitor A, Kogel KH, Waller F (2008) Systemic resistance in Arabidopsis conferred by the mycorrhizal fungus Piriformospora indica requires jasmonic acid signaling and the cytoplasmic function of NPR1. Plant Cell Physiol 49:1747–1751
Trotta A, Varese GC, Gnavi E, Fusconi A, Sampò S, Berta G (1996) Interactions between the soilborne root pathogen Phytophthora nicotianae var. parasitica and the arbuscular mycorrhizal fungus Glomus mosseae in tomato plants. Plant Soil 1(85):199–209
Utkhede R (2006) Increased growth and yield of hydroponically grown greenhouse tomato plants inoculated with arbuscular mycorrhizal fungi and Fusarium oxysporum f. sp radicis-lycopersici. Biocontrol 51:393–400
van der Heijden MGA, Bardgett RD, van Straalen NM (2008) The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol Lett 11:296–310
Van Wees SCM, Van der Ent S, Pieterse CMJ (2008) Plant immune responses triggered by beneficial microbes. Curr Opin Plant Biol 11:443–448
Vierheilig H, Steinkellner S, Khaosaad T, Garcia-Garrido JM (2008) The biocontrol effect of mycorrhization on soil-borne fungal pathogens and the autoregulation of the AM symbiosis: one mechanism, two effects? In: Varma A (ed) Mycorrhiza: genetics and molecular biology, eco-function, biotechnology, eco-physiology, structure and systematics. Springer, Heidelberg/Germany, pp 307–320
Whipps JM (2004) Prospects and limitations for mycorrhizas in biocontrol of root pathogens. Can J Bot 82:1198–1227
Yan Z, Reddy MS, Ryu C-M, McInroy JA, Wilson WM, Kloepper JW (2002) Induced systemic protection against tomato late blight elicited by plant growth-promoting rhizobacteria. Phytopathol 92:1329–1333
Yao MK, Desilets H, Charles MT, Boulanger R, Tweddell RJ (2003) Effect of mycorrhization on the accumulation of rishitin and solavetivone in potato plantlets challenged with Rhizoctonia solani. Mycorrhiza 13:333–336
Zhu HH, Yao Q (2004) Localized and systemic increase of phenols in tomato roots induced by Glomus versiforme inhibits Ralstonia solanacearum. J Phytopathol 152:537–542
Acknowledgments
We acknowledge funding by the Marie Curie program from the European Commission (ERG; PERG02-GA-2007-224751) and by the Spanish Ministry of Science and Innovation (AGL2006-08029). JALR and SJ are supported by the JAE program of the Spanish National Scientific Research Council (CSIC).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2010 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Pozo, M.J., Jung, S.C., López-Ráez, J.A., Azcón-Aguilar, C. (2010). Impact of Arbuscular Mycorrhizal Symbiosis on Plant Response to Biotic Stress: The Role of Plant Defence Mechanisms. In: Koltai, H., Kapulnik, Y. (eds) Arbuscular Mycorrhizas: Physiology and Function. Springer, Dordrecht. https://doi.org/10.1007/978-90-481-9489-6_9
Download citation
DOI: https://doi.org/10.1007/978-90-481-9489-6_9
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-90-481-9488-9
Online ISBN: 978-90-481-9489-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)