
Abstract
Lack of homogeneity in lineages of tumour cell population, in addition to constant evolution of abnormal cells, results in intratumour heterogeneity. Tumour prognosis, to a great extent, depends on the permutations and combinations in which one or more clonal lineage prevails. This directly affects therapeutic decisions as tumour sensitivity, or resistance to a particular treatment is reflected in the heterogeneity it presents. In this context, geographic predisposition presents a different dimension which may drive this heterogeneity in a particular direction and is probably the larger set of cause followed by subsets of causes like clonal evolution or cancer stem cells which result in the idiopathic nature of tumour heterogeneity. However, the geographic pervasiveness of cancers has not been studied in depth in context to tumour heterogeneity. The suffix “-omics” denotes a study in total of that particular stream of science. For example, the study of genome is referred to as genomics. Off late, multi-omics approaches encompassing a systemic understanding of cancer tissues have shed immense light in understanding the molecular basis of tumour heterogeneity with regard to the classical models traditionally proposed to explain the heterogeneity. Genomics, proteomics, metabolomics and transcriptomics have received an enormous boost with emergence and evolution of high-throughput technology like mass spectrometry and next-generation sequencing. This chapter discusses briefly about the type of tumour heterogeneity noticed especially in context of the developing world and how OMICS-based research can bring about a revolution in understanding any geographic bias that may exist and contribute to tumour heterogeneity.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Tumour heterogeneity
- Geographic variation
- Developing country
- Endemic
- Personalised therapeutics
- Multi-OMICS
2.1 Introduction
Heterogeneity in the tumours broadly depends on the origin (tissue and cell type) of the tumour (intertumour heterogeneity) (Marusyk et al. 2012). Sometimes, despite having the same origin, tumour heterogeneity (intratumour) can be observed (Burrell et al. 2013). Intratumour heterogeneity is complex , but is a widely observed phenomenon and has been best explained using Peter Nowell’s clonal evolution model and existence of cancer stem cells (Shackleton et al. 2009). Primarily, the genomic instability leading to a high genetic diversity in the tumour cell population is attributed as the primary reason or the source of heterogeneity which is driven across the clonal generations through somatic evolutions. At the same time, the non-genetic heterogeneity of deterministic and stochastic cellular phenotypes is attributed to the cancer stem cell model (Stingl and Caldas 2007; Marusyk et al. 2012). The micro environment of the tumour also has a significant influence on the epigenetic landscapes of these cells further enhancing this heterogeneity (Gupta and Massagué 2006). Furthermore, intratumour heterogeneity gets more complex in nature when a single form of clonal progeny has a pronounced dominance over the rest (Marusyk et al. 2012). There could be a situation where we have mixed dominance of two or more different clonal progenies. When such a tumour metastasises to another tissue, the heterogeneity builds up from the subtle differences depending on the differences in the tissue type between the primary tumour site and the secondary tumour site (Gupta and Massagué 2006; Bedard et al. 2013). In each of these situations, the tumour would present a different response to therapeutics and therefore poor prognosis. Heterogeneity existing in the tumour cell population exhibits some clinically important phenotypic features which renders it sensitive or resistant to certain therapies (Junttila and de Sauvage 2013), thus posing as a major hindrance in accurate diagnostic and therapeutic decision (Bedard et al. 2013). Although such phenotypic and genetic factors influencing tumour heterogeneity have been well explored, there exists a dearth of research in the field of the geographic variations which can be believed to enhance the tumour heterogeneity to another dimension.
2.2 Understanding Cancer Demographics and its Influence on Tumour Heterogeneity
A clear evidence to support the geographic pervasiveness of cancer is the fact that the cancer types that plagues the developed world is very different from the kind of cancers which burden the developing world (Rastogi et al. 2004; Gupta et al. 2014). This indicates that various pockets of populations across the globe have a predisposition towards particular cancers, and therefore conversely there are cohorts which are more “protected” from certain cancers than some other populations. If we segregate the global population into firstly two major sections—developed and developing world —there are several incidences of certain types of cancer specifically in developing nations, while there are negligible incidences for the same types of cancer in the developed world and vice versa (Gupta et al. 2014). The enormous diversities in socio-epidemiological background, ethnicity, lifestyle, diet, exposure to various environmental risk factors and infectious agents, hormonally related variables and other external epigenetic factors can be thought to contribute to cancer heterogeneity in addition to genetic factors.
Amidst this, it is also important to open up to another perspective and acknowledge the magnitude of disease burden in developing nations. While it was thought initially that low- and middle-income nations house the bulk of infectious diseases, recent observations suggest that the trends are similar even with non-communicable diseases like cardiovascular disorders, diabetes, cancer, etc. (Wagner and Brath 2012). When specifically talking about cancer , there has definitely been a rise in the cancer-incidence rate in developed world as compared to the developing world (Are et al. 2013). According to World Cancer Report 2008 (Boyle and Levin), the incidence of newly reported cancer jumped to 56 % in 2008 compared to 15 % in 1970. Out of 7.6 million cancer deaths worldwide every year, developing world accounts for two-thirds of it (Jemal et al. 2011). Hence, while genetic stability and clonal evolution-based models of studying cancer heterogeneity, post the manifestation of the disease, are the ultimate effectors of tumour heterogeneity , we propose an angle to understand cancer heterogeneity where it is more likely to first influence the kind of heterogeneity an individual is likely to present. Perhaps genetic predisposition of a population to be prone to a type of cancer is an interesting arena for -omics researchers wherein the overshadowed facet of geographic heterogeneity and its function in determining the course of detection and treatment are studied in depth.
2.3 Predisposition of a Population to a Particular Cancer
Considering that tumour heterogeneity may arise due to a large number of variables, the very root of this heterogeneity can at its onset be detected as aberrations at the genetic level. At this point, what is of interest is the likely possibility of genetic vulnerabilities lying in the population of an area endemic to a certain type of cancer and the path of heterogeneity it takes as the disease progresses as compared to any other non-endemic region. Taking India as an example of an emerging developing country, one can observe ubiquity of a particular cancer in a particular territory. It has been proven that certain gene polymorphism prevalent within a given population renders them more vulnerable to a particular cancer (Shankarkumar and Sridharan 2011). It has been observed that Indian women residing in the northern province of the country have a polymorphic copy of CYP1A1, a biomarker known to rendering them more prone to breast cancer (Singh et al. 2007), while the association of polymorphic copy of Fas-670 gene has shown to play a role in North Indian women being prone to cervical cancer (Kordi Tamandani et al. 2008). XRCC4 is known to play a key role in nonhomologous end-joining repair pathway. Thus, polymorphisms in such DNA repair genes have been shown to reduce their DNA repair capability. XRCC polymorphisms have been implicated for the prevalence of prostate cancer in North Indian Population (Mandal et al. 2011).
Several other examples within the Indian populations can be cited wherein the age-adjusted rates (AAR) of certain cancers are among the highest reported in the globe. However, the picture that has unveiled recently is that the highest AAR of a particular malignancy is focused at its epicentre and not dispersed in the entire nation uniformly. In other developing countries like China, it has been established that IL-1B2511T/T genotypes predisposes the population residing in Shanxi which has reported high prevalence of gastric cancer as compared to Guangdong which has low prevalence of gastric cancer (Zeng et al. 2003). Similarly, Korea is known to be endemic for gastric cancer (Park et al. 2000). Gall bladder carcinoma is known to be endemic in Chile (Wistuba et al. 1995), while nasopharyngeal carcinoma and oesophageal carcinoma are known to be major tumours in southern China (Seng et al. 2007). The broader question in this scenario is to find out the extent to which these cancer endemic populations are predisposed as compared to the diseased population from sporadic occurrences. The next question is to find the nature and uniqueness of these predispositions as several factors like age, sex, socioeconomic background, education, tobacco and alcohol abuse, lifestyle changes, infections (Helicobacter pylori, human papillomavirus, hepatitis B virus, Epstein Barr virus, etc.) and environmental factors contributing to cancer risks. However, in spite of these factors, one cannot ignore these significant demographic trends within the slow-growing pandemic of cancer which need to be studied considering these factors more holistically. Thus, these observations establish the need for specifically studying the population residing in developing nations on the basis of these trends (Nandakumar et al. 2005).
2.4 Perceiving Geographic Heterogeneity by Multi-OMICS Research
2.4.1 Genomic and Transcriptomics Studies
Due to the nature of the problem that tumour heterogeneity presents, genomics-based techniques have been traditionally used to detect signature abberations (Chung et al. 2002). RNA-/cDNA-based microarrays have been used to understand heterogeneous gene expression to understand cancer as a disease to greater depths (Alizadeh et al. 2000; Pennings et al. 2012). With the advent of next-generation sequencing, the power unleashed by genomic-based techniques has undeniably become the most important tool available to researchers to identify the players involved in tumour heterogeneity (Meyerson et al. 2010; Roychowdhury et al. 2011). In a study, a 54-year-old individual was subjected to integrated personal omics profile (iPOP). This involved a high-resolution examination of this whole genome sequence (WGS) and whole exome sequence (WES). This study provided his genomic and transcriptomic profile in addition to his proteomic , metabolomic and autoantibody profiles. These were analysed over a 14-month period through his healthy and diseased states which helped reveal certain medical risks (Chen et al. 2012). Such approaches can be extended to cancer-based cohorts as it would shed some light on tumour heterogeneity , particularly if one targets cancers in certain endemic populations.
2.4.2 Proteomic Studies
Although genomic studies are an effective method to study the heterogeneity presented in cancer, proteomic analysis of vulnerable populations can help elucidate the extent of their predisposition to a particular cancer. A comparative study of vulnerability with healthy populations can be sought out with an aim to comparatively analyse the proteome profile of relatively resistant groups to vulnerable groups (Alaiya et al. 2000). Alterations in the genome are often reflected in its proteome by several magnifications. Proteomics-based studies specially allow identification of novel biomarkers and protein expression signatures (Wulfkuhle et al. 2003). They also help in prediction of responses of a patient to a given drug, thus helping personalised therapy (Cancer Target Discovery and Development Network et al. 2010). In an attempt to reduce the invasiveness to diagnose cancer and to develop novel biomarkers, the plasma proteome has been proposed to be an attractive target to detect variation in cancer patients as compared to healthy individuals. High-throughput techniques like mass spectrometry have proved to be instrumental in understanding the differential expression of proteins in tumour tissues. Thus, such approaches provide a base to form a proteomic reference in an endemic population , and it also helps in comparing protein profiles of patients across different states within the country and in the world to explore relative risks and targeting therapy (Petricoin et al. 2002; Chumbalkar et al. 2005). The importance of such studies is to find if there are proteomic differences which may be location and race based. From the clinical perspective as well, proteomic findings can be immediately applied into translational validations, and hence the initiative to study those is in the interest of national health.
2.4.3 Metabolomic Studies
Cellular metabolism at the functional level is one of the key players, which define the physiological nature of the cell. Thereby, understanding tumour metabolism is not only crucial to understand the disease biology, but it also is important in terms of source of potential biomarkers. Metabolomics, the systemic study of metabolites, is an emerging field and is increasingly being adopted to unravel various facets of clinical oncology. Nuclear magnetic resonance (NMR), mass spectrometry (MS) , Fourier-transform infrared spectroscopy (FTIR), high-performance liquid chromatography coupled with electrochemical coulometric array detection (LCECA), Raman spectroscopy and metabolite arrays are few of the technologies available to study metabolomics from various biospecimens (Kaddurah-Daouk et al. 2008; Oakman et al. 2011). Metabolomics has been used extensively and easily in biofluids where sample preparation is relatively simple; however, for tissue-based metabolomics, the sample preparation has been relatively tricky (Heinemann and Zenobi 2011). With due precaution, it is possible to study large cohorts to understand systemic alterations in biofluids like serum, urine, CSF, etc., and understand population-based metabolomics signatures. To dissect the disease completely, tissue-based metabolomics followed by single cell analysis of these tissues would be of prime importance.
2.4.4 Single Cell Analysis (SCA)
The resolution of results obtained in terms of dissecting the heterogeneity in both genomics and proteomics is a huge limiting factor. A minimum of 105 cells are required for extracting DNA/RNA to perform any further genomic or sequencing studies. This large number of cell input implies that they may not be single cell-derived lineages. Thus, the results from such studies may give us an averaged or holistic impression of the genomic profile (Rosenblum and Peer). Same is the case in proteomic studies wherein tissue extracts cannot be thought to be derived from cells in isolation. Single cell sequencing is an outcome of next-generation sequencing (NGS) which has helped us to now perform high-resolution screening of the single cell genome to understand the genetic, epigenetic and transcriptomic heterogeneity, especially in case of diseased samples (Macaulay and Voet 2014). Single cell proteomics in the form of flow cytometry, single cell proteomic chips, etc., have been used to decipher signalling pathways (Irish et al. 2006; Shi et al. 2012). However, there is more scope for innovation to cater to the needs for detecting proteomic aberrations at the single cell level.
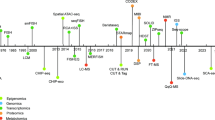
Many new commercial technologies apart from the traditional high-throughput microscopy have recently emerged to take SCA to a multi-omics platform. These technologies are essentially “Lab-on-Chip” devices which exploit microfluidics, cell trapping techniques using biophysical parameters like dielectrophoretic charges, etc. (Mannello et al. 2012). Devices from ScreenCell, Fluidigm’s C1TM Single-Cell AutoPrep System, Silicon Biosystems DEPArray, etc., are few such examples (Mannello et al. 2012). On culturing cell populations derived from each single cell in a given tumour tissue, it is possible to run a host of multi-omics-based experimentation (including genomics, transcriptomics, metabolomics, degradomics, etc.) on them which would truly represent the systemic alterations down to the single cell level and capture the entire biological heterogeneity. These studies can then be extrapolated at a cross-sectional level where cohorts from different geographical locations could be studied to find common and unique etiological alterations in cancers (Fig. 2.1).

Represents the rationale of undertaking single cell analysis to dissect intratumour heterogeneity by separating single cells in a tumour, each of which could have unique alterations of their own. These could be subjected to various multi-omics approaches like genomics, transcriptomics, proteomics , metabolomics , etc., to holistically determine the trends in a population
2.5 Tumour Heterogeneity Necessitates the Personalisation of Therapeutics and Implementation of Standard Global Models
Findings of research pursued globally, primarily in the developed world, using samples from the developed countries would be futile when extrapolated to populations residing in developing nations rendering it inconclusive. Certain strategies have been explored by various research groups globally, viz., the University of Michigan in collaboration with the International Genomics Consortium and institutes contributing to The Cancer Genome Atlas (TCGA), with an aim to personalise therapy. The success of such an approach from the Broad Institute of Harvard and MIT has been recently published where pharmacological profiles of 24 anticancer drugs were studied across 479 cancer cell lines (Barretina et al. 2012). They found that the development of personalised therapeutic regimens can be enhanced by genetic predictions of drug response in the preclinical settings and their incorporation into cancer clinical trials (Barretina et al. 2012). This study is widely being used as the central rationale for most research groups towards developing personalised medication. In fact, the same institute also reported the sensitivity of Bcl-2 family antagonist, Navitoclax, resulting due to mutations in oncogene b-catenin and its candidate dependency (Basu et al. 2013). They have used this model to hypothesise novel therapeutics to accelerate the discovery of drugs to cater to patients depending on the cancer genotype and lineage. Studies affiliated to TCGA using such approaches have been responsible for major findings like classification of glioblastoma into four distinct subtypes depending on gene expression patterns and clinical characteristics, viz., proneural, neural, classical, and mesenchymal subtypes (Verhaak et al. 2010). Aberrations in expression of EGFR, NF1 and PDGFRA/IDH1 genes result in the classical, mesenchymal, and proneural subtypes, respectively (Verhaak et al. 2010). The Pan-Cancer initiative affiliated to TCGA has profiled 12 tumour types by analysing large number of human tumours to shed light on aberrations in DNA, RNA, protein or epigenetic at the molecular level (Cancer Target Discovery and Development Network et al. 2010; Kandoth et al. 2013). Their success in utilising such strategies has helped the scientific community to understand how one can extend therapies effective in one cancer type to others with a similar genomic profile (Barretina et al. 2012). On extending these strategies in collaboration with research groups in the developing world or by using samples from the developing world, the genetic basis for the disparity in global heterogeneity in cancer can be better understood.
2.6 Concluding Remarks
Tumour heterogeneity is a challenging subject to study and is essential to understand how cancer progresses and the therapeutic implications it extends. To this end, multi-omics research has provided numerous clues which establish these facts clearly. However, there is a huge scope to study geographic bias of a disease and the heterogeneity it presents. By comparing populations and the cancers endemic to these regions using multi-omics approaches, it would not only provide us biological evidences of genetic vulnerability, if it exists, but will also help extensively in certain basic generalisations of inclusions or exclusions of therapeutic regimes according to its response in a population. Further from population centric regimes, it also provides a gateway to understand the response on an individual to a particular therapy. It is probable that in the near future, technologies like NGS and Mass Spectrometry based Targeted Proteomics would be routinely used to detect residual disease after therapy during follow-ups. With the drastic evolution of expertise, personalised medicine and targeted therapeutics are very much a reality and should be pursued extensively in order to combat the challenges presented by cancer heterogeneity.
References
Alaiya AA, Franzén B, Auer G, Linder S (2000) Cancer proteomics: from identification of novel markers to creation of artificial learning models for tumor classification. Electrophoresis 21:1210–1217. doi:10.1002/(SICI)1522-2683(20000401)21:6<1210::AID-ELPS1210>3.0.CO;2-S
Alizadeh AA, Eisen MB, Davis RE et al (2000) Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature 403:503–511. doi:10.1038/35000501
Are C, Rajaram S, Are M et al (2013) A review of global cancer burden: trends, challenges, strategies, and a role for surgeons. J Surg Oncol 107:221–226. doi:10.1002/jso.23248
Barretina J, Caponigro G, Stransky N et al (2012) The cancer cell line encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature 483:603–607. doi:10.1038/nature11003
Basu A, Bodycombe NE, Cheah JH et al (2013) An interactive resource to identify cancer genetic and lineage dependencies targeted by small molecules. Cell 154:1151–1161. doi:10.1016/j.cell.2013.08.003
Bedard PL, Hansen AR, Ratain MJ, Siu LL (2013) Tumour heterogeneity in the clinic. Nature 501:355–364. doi:10.1038/nature12627
Burrell RA, McGranahan N, Bartek J, Swanton C (2013) The causes and consequences of genetic heterogeneity in cancer evolution. Nature 501:338–345. doi:10.1038/nature12625
Cancer Target Discovery, Development Network, Schreiber SL, Shamji AF et al (2010) Towards patient-based cancer therapeutics. Nat Biotechnol 28:904–906. doi:10.1038/nbt0910-904
Chen R, Mias GI, Li-Pook-Than J et al (2012) Personal omics profiling reveals dynamic molecular and medical phenotypes. Cell 148:1293–1307. doi:10.1016/j.cell.2012.02.009
Chumbalkar VC, Subhashini C, Dhople VM et al (2005) Differential protein expression in human gliomas and molecular insights. Proteomics 5:1167–1177. doi:10.1002/pmic.200401202
Chung CH, Bernard PS, Perou CM (2002) Molecular portraits and the family tree of cancer. Nat Genet 32(Suppl):533–540. doi:10.1038/ng1038
Gupta GP, Massagué J (2006) Cancer metastasis: building a framework. Cell 127:679–695. doi:10.1016/j.cell.2006.11.001
Gupta S, Venkatesh A, Ray S, Srivastava S (2014) Challenges and prospects for biomarker research: a current perspective from the developing world. Biochim Biophys Acta. doi:10.1016/j.bbapap.2013.12.020
Heinemann M, Zenobi R (2011) Single cell metabolomics. Curr Opin Biotechnol 22:26–31. doi:10.1016/j.copbio.2010.09.008
Irish JM, Kotecha N, Nolan GP (2006) Mapping normal and cancer cell signalling networks: towards single-cell proteomics. Nat Rev Cancer 6:146–155. doi:10.1038/nrc1804
Jemal A, Bray F, Center MM et al (2011) Global cancer statistics. CA Cancer J Clin 61:69–90. doi:10.3322/caac.20107
Junttila MR, de Sauvage FJ (2013) Influence of tumour micro-environment heterogeneity on therapeutic response. Nature 501:346–354. doi:10.1038/nature12626
Kaddurah-Daouk R, Kristal BS, Weinshilboum RM (2008) Metabolomics: a global biochemical approach to drug response and disease. Annu Rev Pharmacol Toxicol 48:653–683. doi:10.1146/annurev.pharmtox.48.113006.094715
Kandoth C, McLellan MD, Vandin F et al (2013) Mutational landscape and significance across 12 major cancer types. Nature 502:333–339. doi:10.1038/nature12634
Kordi Tamandani DM, Sobti RC, Shekari M (2008) Association of Fas-670 gene polymorphism with risk of cervical cancer in north Indian population. Clin Exp Obstet Gynecol 35:183–186
Macaulay IC, Voet T (2014) Single cell genomics: advances and future perspectives. PLoS Genet 10:e1004126. doi:10.1371/journal.pgen.1004126
Mandal RK, Singh V, Kapoor R, Mittal RD (2011) Do polymorphisms in XRCC4 influence prostate cancer susceptibility in north Indian population? Biomark Biochem Indic Expo Response Susceptibility Chem 16:236–242. doi:10.3109/1354750X.2010.547599
Mannello F, Ligi D, Magnani M (2012) Deciphering the single-cell omic: innovative application for translational medicine. Expert Rev Proteomics 9:635–648. doi:10.1586/epr.12.61
Marusyk A, Almendro V, Polyak K (2012) Intra-tumour heterogeneity: a looking glass for cancer? Nat Rev Cancer 12:323–334. doi:10.1038/nrc3261
Meyerson M, Gabriel S, Getz G (2010) Advances in understanding cancer genomes through second-generation sequencing. Nat Rev Genet 11:685–696. doi:10.1038/nrg2841
Nandakumar A, Gupta PC, Gangadharan P et al (2005) Geographic pathology revisited: development of an atlas of cancer in India. Int J Cancer J Int Cancer 116:740–754. doi:10.1002/ijc.21109
Oakman C, Tenori L, Biganzoli L et al (2011) Uncovering the metabolomic fingerprint of breast cancer. Int J Biochem Cell Biol 43:1010–1020. doi:10.1016/j.biocel.2010.05.001
Park YJ, Shin KH, Park JG (2000) Risk of gastric cancer in hereditary nonpolyposis colorectal cancer in Korea. Clin Cancer Res Off J Am Assoc Cancer Res 6:2994–2998
Pennings JLA, Van Dycke KCG, van Oostrom CTM et al (2012) Biomarker discovery using a comparative omics approach in a mouse model developing heterogeneous mammary cancer subtypes. Proteomics 12:2149–2157. doi:10.1002/pmic.201100497
Petricoin EF, Zoon KC, Kohn EC et al (2002) Clinical proteomics: translating benchside promise into bedside reality. Nat Rev Drug Discov 1:683–695. doi:10.1038/nrd891
Rastogi T, Hildesheim A, Sinha R (2004) Opportunities for cancer epidemiology in developing countries. Nat Rev Cancer 4:909–917. doi:10.1038/nrc1475
Rosenblum D, Peer D (n.d.) Omics-based nanomedicine: the future of personalized oncology. Cancer Lett. doi: 10.1016/j.canlet.2013.07.029
Roychowdhury S, Iyer MK, Robinson DR et al (2011) Personalized oncology through integrative high-throughput sequencing: a pilot study. Sci Transl Med 3:111ra121. doi:10.1126/scitranslmed.3003161
Seng TJ, Low JSW, Li H et al (2007) The major 8p22 tumor suppressor DLC1 is frequently silenced by methylation in both endemic and sporadic nasopharyngeal, esophageal, and cervical carcinomas, and inhibits tumor cell colony formation. Oncogene 26:934–944. doi:10.1038/sj.onc.1209839
Shackleton M, Quintana E, Fearon ER, Morrison SJ (2009) Heterogeneity in cancer: cancer stem cells versus clonal evolution. Cell 138:822–829. doi:10.1016/j.cell.2009.08.017
Shankarkumar U, Sridharan B (2011) Glioma Indian scenario: is there a human leucocyte antigen association? J Nat Sci Biol Med 2:205–208. doi:10.4103/0976-9668.92323
Shi Q, Qin L, Wei W et al (2012) Single-cell proteomic chip for profiling intracellular signaling pathways in single tumor cells. Proc Natl Acad Sci U S A 109:419–424. doi:10.1073/pnas.1110865109
Singh N, Mitra AK, Garg VK et al (2007) Association of CYP1A1 polymorphisms with breast cancer in North Indian women. Oncol Res 16:587–597
Stingl J, Caldas C (2007) Molecular heterogeneity of breast carcinomas and the cancer stem cell hypothesis. Nat Rev Cancer 7:791–799. doi:10.1038/nrc2212
Verhaak RGW, Hoadley KA, Purdom E et al (2010) Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 17:98–110. doi:10.1016/j.ccr.2009.12.020
Wagner K-H, Brath H (2012) A global view on the development of non communicable diseases. Prev Med 54(Suppl):S38–S41. doi:10.1016/j.ypmed.2011.11.012
Wistuba II, Sugio K, Hung J et al (1995) Allele-specific mutations involved in the pathogenesis of endemic gallbladder carcinoma in Chile. Cancer Res 55:2511–2515
Wulfkuhle JD, Liotta LA, Petricoin EF (2003) Proteomic applications for the early detection of cancer. Nat Rev Cancer 3:267–275. doi:10.1038/nrc1043
Zeng Z-R, Hu P-J, Hu S et al (2003) Association of interleukin 1B gene polymorphism and gastric cancers in high and low prevalence regions in China. Gut 52:1684–1689
Acknowledgement
This work has been supported by the Department of Biotechnology, Government of India, Project No. BT/PR4599/BRB/10/1042/2012.Competing Interests The authors declare no competing interests.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer India
About this chapter
Cite this chapter
Gupta, S., Ray, S., Talukdar, A., Sehgal, K., Moiyadi, A., Srivastava, S. (2016). Geographic Pervasiveness of Cancer: Prospects of Novel Biomarker and Therapeutic Research in Developing Countries using OMICS approaches. In: Srivastava, S. (eds) Biomarker Discovery in the Developing World: Dissecting the Pipeline for Meeting the Challenges. Springer, New Delhi. https://doi.org/10.1007/978-81-322-2837-0_2
Download citation
DOI: https://doi.org/10.1007/978-81-322-2837-0_2
Published:
Publisher Name: Springer, New Delhi
Print ISBN: 978-81-322-2835-6
Online ISBN: 978-81-322-2837-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)