
Abstract
Paralysis after stroke or neurotrauma is among the leading causes of long term disability in adults. The development of brain–computer interface (BCI) systems that allow online classification of electric or metabolic brain activity and their translation into control signals of external devices or computers have led to two major approaches in tackling the problem of paralysis. While assistive BCI systems strive for continuous high-dimensional control of robotic devices or functional electric stimulation (FES) of paralyzed muscles to substitute for lost motor functions in a daily life environment (e.g. Velliste et al. 2008 [1]; Hochberg et al. 2006 [2]; Pfurtscheller et al. 2000 [3]), restorative BCI systems aim at normalization of neurophysiologic activity that might facilitate motor recovery (e.g. Birbaumer et al. 2007, 2009 [4, 5]; Daly et al. 2008 [6]). In order to make assistive BCI systems work in daily life, high BCI communication speed is necessary, an issue that by now can only be achieved by invasive recordings of brain activity (e.g. via multi-unit arrays, MUA, or electrocorticogram, ECoG). Restorative BCI systems, in contrast, were developed as training tools based on non-invasive methods such as electro- or magnetoencephalography (EEG/MEG). More recently developed approaches use real-time functional magnetic resonance imaging (rtfMRI) or near-infrared spectroscopy (NIRS). Here, we provide an overview of the current state in the development and application of assistive and restorative BCI and introduce novel approaches to improve BCI control with brain stimulation such as transcranial direct current stimulation (tDCS). The outlook of using BCI in rehabilitation of stroke and neurotrauma is discussed.
Access provided by Autonomous University of Puebla. Download conference paper PDF
Similar content being viewed by others


Keywords
- Transcranial Magnetic Stimulation
- Motor Imagery
- Functional Electric Stimulation
- Local Field Potential
- Anodal tDCS
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Introduction
Since the development of electroencephalographic measurements (EEG) in the early twentieth century based on Hans Berger’s discovery of electric brain oscillations [7], the idea of reading out thoughts from brain activity fired the imagination of many scientists. Most recent advances in sensor technology and computational capacities led to the development of brain–computer interfaces (BCI). These systems allow direct translation of electric or metabolic brain activity into control signals of external devices or computers. While BCI systems based on classification of action potential spike trains recorded by single or multi-unit electrodes or local field potentials (LFP) recorded by electrocorticography (ECoG) require implantation (invasive BCI) [1, 8–11], well established techniques such as electroencephalography (EEG) and magnetoencephalography (MEG) allow non-invasive BCI control [12–16]. More recent developments use near-infrared-spectroscopy (NIRS) or real-time functional magnetic resonance imaging (rtfMRI) in BCI systems [17].
By creating an alternative efferent pathway of the brain, BCI were successfully used for communication [12, 18] or control of orthotic devices that would allow hemiplegic patients e.g. to grasp [19].
Stroke and neurotrauma belong to the leading causes for long-term disability worldwide and the number of affected people increases every year due to demographic change and increasing survival rates [20]. Up to 30% of all stroke victims experience very limited motor recovery and depend on assistance to manage their daily living activities [21, 22]. Enabling those patients to regain the ability to move their paralyzed limbs, respectively improving their capacity for motor behavior, could substantially improve their quality of life. While there are encouraging studies providing evidence that e.g. constrained-induced therapy (CIT) or bilateral arm training might be useful strategies for rehabilitation of stroke patients with paretic upper extremities [23, 24], there is no accepted and efficient rehabilitation strategy for severely affected stroke patients with completely paralyzed muscles, precisely those who cannot participate in common training-based rehabilitative treatments.
Assistive and Restorative BCI in Neurorehabilitation
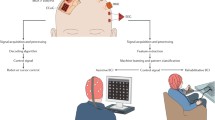
Depending on the aim of BCI use in rehabilitation of paralysis, two major approaches can be distinguished: while assistive BCI systems aim at high dimensional control of robotic limbs or functional electric stimulation (FES) that specifically activate paralyzed muscles to substitute a lost motor function in daily life [1, 2, 25], restorative BCI aim at selective induction of use-dependent neuroplasticity to facilitate motor recovery [26–29].
These two approaches derive from different research traditions and are not necessarily related to the invasiveness of the approach: In early work of Eberhard Fetz (1969) a monkey learned, based on operant conditioning, to use cortical unit activity to deflect a lever delivering reward [30]. Two decades later, decoding of different movement directions from single neurons was achieved [31, 32]. Since then the reconstruction of complex movements from neuronal activity became possible. Firing patterns of single cells of the motor cortex [33] or parietal neuronal pools [34] in animals were remarkably successful to reconstruct movement trajectories. Monkeys learned to control cursors towards moving targets on a computer screen in a predetermined sequence by successively activating neurons in motor, premotor and parietal motor areas. In a particularly successful preparation, 32 cells were sufficient to move an artificial arm and perform skilful reaching movements after extensive training [1]. This technique enabled a monkey to feed himself. The plasticity of the cortical circuits allowed learned control of movements directly from the cellular activity even outside the primary or secondary homuncular representations of the motor cortex [9], a circumstance that supports the assumption that operant conditioning is a key factor for learning BCI control irrespective the area of recording. In an encouraging experiment Hochberg (2006) implanted densely packed microelectrode arrays of up to several hundred microelectrodes in two quadriplegic human patients [2]. Within a few training sessions, the patients learned to use LFP to move a computer cursor in several directions. This kind of BCI control with two degrees of freedom (DoF) could be used e.g. to switch on lights, a TV or to make a phone call. However, in contrast to the studies in healthy animals none of the invasive procedures allowed restoration of skilful movement in paralyzed humans. It is not clear why so far the human preparations have achieved only limited results in terms of application to activities of daily living. There are a couple of major challenges that are unsolved particularly stability, encapsulation and general safety issues [35–37]. While the primary motor cortex (M1) has the most non-ambiguous influence on the motor neurons of the upper limb, it is tightly connected with supplementary (SMA) and other non-primary motor areas [38] that are involved in the integration of complex skilled movements [39, 40]. Studies using retrograde transport tracers from the arm area of M1 in macaques showed, however, that up to 60% of all cortical projections to the spinal cord originate in pre-motor areas [41]. This indicates that complex motor behavior might not be exclusively decodable from the primary motor cortex and may in fact require multiple recording sites from various brain areas that integrate complex networks. In a very encouraging recent study, though, sufficient information could be extracted from a 4 × 4 mm grid with 96 silicon-based electrodes placed over M1 of a macaque to reconstruct 25 measured joint angles representing an estimated 10 DoF [42]. This electrode system (BrainGate II®) is currently investigated in a pilot human clinical trial to address reliability and safety.Footnote 1
Studies using the less invasive approach of epidural implanted ECoG electrode grids showed that a subject could learn to control cursor movements with only a few minutes of training [43, 44]. Besides a better topographical resolution and recording bandwidth compared to non-invasive approaches [45, 46], ECoG based BCI have a better signal-to-noise ratio due to absence of electromyographic contaminations and other artifacts [47]. Recent work showed that prediction of a monkey’s 3D hand trajectories and 7 DoF arm joint angles are possible with accuracy similar to recordings based on single-cell-recordings [48]. The level of DoF that can be achieved with an ECoG-grid by decoding movement associated LFP in patients with brain lesions is unclear, though, and matter of investigation.
Although not entirely impossible [49, 50], extraction and online decoding of movement trajectories from non-invasively recorded brain activity is difficult [51]. However, in contradiction to Skinner’s proposal that operant and classical conditioning require involvement of the musculoskeletal system [52], voluntary control of brain oscillations is possible, opening the door to utilize this circumstance for non-invasive BCI control. The average communication rate achieved with non-invasive BCI technology in humans ranges between 5 and 25 bits/min [53], i.e. up to 25 binary (yes/no) choices can be correctly classified per minute.
Patients that are otherwise incapable to communicate, i.e. locked-in-patients suffering from amyotrophic lateral sclerosis (ALS), a disease characterized by a combined degradation of upper and lower motor neurons, can significantly benefit from non-invasive BCI use [12, 18, 54, 55]: patients are first trained to produce positive or negative slow cortical potentials (SCP) [75] upon the command of an auditory cue and after achieving more than 70% control, letters or words were presented on a computer screen. A particular letter was selected by creating SCP after its appearance [12, 14, 56, 57]. Over 40 patients with ALS at various stages of their disease were trained to use the SCP-BCI, eventually seven of these patients arrived at the locked-in-state (LIS) and were able to continue to use the BCI. All patients who began training after entering the complete locked-in-state (CLIS), however, were unable to achieve lasting BCI control [58, 59] – a finding that might be of relevance for the understanding of voluntary modulation of brain activity and BCI control.
Besides SCP, sensori-motor rhythms (SMR) are among the most investigated electro-physiologic signals used for non-invasive BCI control. The discovery of SMR dates back to the early 1950s: the observation of a local and frequency specific signal-amplitude reduction in the range of 8–13 Hz over the rolandic area during motor preparation or execution became introduced as μ-rhythm after a suggestion by Gastaut [60, 61]. Based on location, frequency and reactivity to sensory input or output, different components of the μ-rhythm were postulated [3]. The discovery of event-related desynchronization (ERD) and synchronization (ERS) during motor-related activities [62] was the basis for the development of SMR-based BCI. ERD/ERS offers quantification of stimulus-locked brain activity within the time-frequency and spatial domain. It is assumed that ERD reflects extensive information processing within the sensory-motor system [63], while ERS is associated with increased synchronous idling of sensory-motor neuron networks [64]. The accessibility by cognitive manipulation makes SMR an ideal candidate to drive a BCI system. Use of SMR modulation for BCI control was extensively investigated by the Pfurtscheller group in Graz [3, 13], the Wolpaw group in Albany [65, 66] and the Birbaumer group in Tübingen [67]. In 2003, Pfurtscheller’s BCI-group introduced the first SMR-based BCI that was used to enable a quadriplegic patient to control grasping through functional electrical stimulation activated by motor imagery [3].
Another well-tested BCI controller is the P300-BCI based on event-related brain potentials (ERP) by Donchin [68]. While SCP- and SMR-control is learned through visual and auditory feedback often requiring up to ten training sessions before reliable control is achieved, the P300-BCI needs no extensive training. Information rates of P300-BCI can reach 20–25 bits/min [69] but requires a very high attention level – a requirement often not met by people with neurologic or psychiatric disorders.
Most recently also a BOLD-signal based rtfMRI-BCI has been introduced [70–73]. In 2003 Weiskopf et al. [70] proposed that the development of fMRI-BCIs might be a powerful tool in the treatment of various disorders and diseases. It was shown that intracortical activity is highly correlated with local blood flow change and the BOLD signal [74] and that volitional regulation of BOLD activity in cortical and sub-cortical areas such as amygdala, anterior cingulate, insula and parahippocampal gyrus was associated with changes of connectivity between those areas [73]. DeCharms et al. [72] demonstrated that use of a real-time fMRI-BCI can affect pain perception.
In addition to the fMRI-BCI approach, near infrared spectroscopy (NIRS) is also a non-invasive technique based on measuring metabolic changes of the brain. Using multiple pairs or channels of light sources and light detectors operating at two or more discrete wavelengths at near infrared range (700–1,000 nm) cerebral oxygenation and blood flow of particular regions of the cortical surface can be determined. The degree of increase in regional cerebral blood flow (rCBF) exceeds that of increase in regional cerebral oxygen metabolic rate (rCMRO2) resulting in a decrease in deoxygenated hemoglobin in venous blood during higher oxygen demand. Therefore, an increase in total hemoglobin and oxygenated hemoglobin with a decrease in deoxygenated hemoglobin can be measured in activated active brain areas. The recent development of portable systems makes NIRS a promising tool in non-invasive BCI research [17, 28].
In contrast to this work aiming at assistive appliance of invasive and non-invasive BCI technology, the development of restorative BCI systems is tightly associated with the development and successes of neurofeedback (NF) and its use to purposefully up-regulate or down-regulate brain activity – a quality that showed to have some beneficial effect in the treatment of various neurological and psychiatric disorders associated with neurophysiologic abnormalities [5]. In NF subjects receive visual or auditory on-line feedback of their brain activity and are asked to voluntarily modify e.g. a particular type of brainwave [5]. The feedback contains the information on the degree of success in controlling the signal and delivers the reward for correct modification. NF was successfully used in the treatment of epilepsy [76, 77], ADHD [78–80], chronic pain syndrome [81] and complete paralysis after stroke [3].
Stroke can be associated with extensive neuroplastic changes on the synaptic, neuronal and circuit level. Besides new synapses strengthening and rewiring [82] as a consequence of long-term potentiation (LTP) or long-term depression (LTD), dendritic sprouting, extensive peri-infarct reorganization and changes of activity patterns in remote cortical regions [83] including interhemispheric inhibition [84–87] were described.
Various interventions that aim at modulation of neuroplasticity, such as reduction of somatosensory input from the intact hand [88] or increase from the affected hand [89], neuropharmacologic strategies influencing dopaminergic or adrenergic systems [90], mental training such as motor-imagery training [91–93] and non-invasive brain stimulation [93–96] showed to have beneficial effects on motor function after stroke.
It was shown that the ability to desynchronize the affected hemisphere in the SMR-range during the acute and sub-acute phase of stroke correlates with clinical motor outcome [97]. A finding that is consistent with fMRI studies performed in stroke patients that suggested an association of increased activity in the ipsilesional primary motor cortex and functional recovery while involvement of the contralesional motor cortex during movements of the affected hand was associated with poor motor recovery [98, 99].
Following these lines, a restorative BCI is based on two hypotheses: (1) By inducing CNS plasticity that produces more normal activation (e.g. in terms of lateralization), normal motor function will be restored. (2) Contingent sensory input given as reward to a specific activation pattern in the motor system induces CNS plasticity that facilitates restoration of normal motor control, potentially through rewiring and synaptic strengthening of weakened or previously inhibited motor networks.
As an important step for further development of SMR-based assistive and restorative BCI systems, a study was conducted by Buch et al. (2008) [19] to evaluate whether patients with chronic stroke would be able to learn to modulate μ-rhythm. Eight patients with chronic hand plegia resulting from stroke participated in 13–22 BCI training sessions to learn voluntary control of their μ-rhythm amplitude originating in the sensori-motor areas of the cortex. Diagnostic MRIs revealed single, unilateral subcortical, cortical or mixed lesions in the participating patients. Patients had no residual finger extension function. Before the actual training, the patients had to imagine several distinct movements of the upper and lower extremity as well as the tongue. While doing this, the ipsilesional area with the strongest oscillatory MEG response in the μ-range was identified. Based on the area’s location, three MEG sensors were selected for BCI control. During the training, μ-desynchronization was translated in cursor-movements on a screen. After approximately 4 s of either up or down regulation, the affected hand was either opened or closed by a hand-orthosis affixed to the participant’s paralyzed fingers. At the end of the training, SMR control was associated with increased range and specificity of μ-rhythm modulation as recorded from sensors overlying central ipsilesional (four patients) or contralesional (two patients) regions. However, two patients were unable to gain BCI control. One patient started with high success rates of BCI control (approximately 85%) at the beginning of the training and did not improve much further. This study demonstrated for the first time that most patients with chronic stroke, even with complete hand paralysis, could learn to control SMR-based BCI-systems.
However, the applied BCI training was not associated with notable clinical improvement. Up to 1 h of BCI training per day for 2–3 weeks might be insufficient to induce relevant motor recovery in patients with chronic paralysis after stroke. Other reasons might have been the limited translation of BCI-associated movements into daily-life activities (“transfer package”) [100] and the delay of BCI-driven somato-sensory input, which resulted in low temporal contingence of brain activation and sensory feedback. Larger clinical studies using BCI systems that couples highly specific temporo-spatial brain activation patterns online with contingent sensory feedback might help to elucidate the viability of SMR-based BCI systems for restoration of paralysis. Unpublished data by Buch et al. indicate that fronto-parietal connectivity plays a key-role for successful SMR-based BCI learning after stroke [101].
Most recently Broetz et al. (2010) [26] published a proof-of-principle study on the combination of BCI training and goal-directed physical therapy in chronic stroke. A 67-year-old hemiplegic patient who suffered from a subcortical bleeding received three blocks of BCI training coupled with goal-directed physical therapy over the course of 12 months. Before the training he had no active finger movements, depended on assistance for personal hygiene and dressing and used a wheelchair for all distances greater than half a mile. Each BCI training block consisted of daily SMR-based BCI training over 30 days. For the first BCI training block a 275-sensor MEG was used to translate motor imagery associated SMR-modulation on the affected hemisphere into opening or closing of the paralyzed hand [25]. The second and third training block was based on EEG-recordings. Goal-directed physical therapy was continued throughout the 12 months. Arm motor function as well as gait (using Fugl-Meyer Assessment, FMA, Wolf Motor Function Test, WMFT, and Ashworth Scale) and brain reorganization was assessed repeatedly during the study. After 1 year, FMA, WMFT and Ashworth scores improved by a mean of 46.6%. The patient was able to extend his fingers and to open his affected hand to grasp. He regularly walked distances over half a mile and did not use the wheelchair anymore. Analysis of spectral amplitudes in MEG data reflecting cortical activity revealed a significantly stronger SMR-desynchronization during motor imagery and motor execution on the affected hemisphere.
A multimodal neuroimaging approach based on fMRI and diffusion tensor imaging (DTI) was used to further examine neuroplastic changes in parallel with the longitudinal clinical evaluation [27]. Psycho-physiological interaction (PPI) analysis was used to assess functional connectivity and showed that activity of ipsilesional pre-motor cortex (PMC) positively co-varied with ipsilesional primary and secondary sensorimotor areas across all measurements. Cortico-spinal tract (CST) integrity was estimated based on DTI analysis and tractography showing a significant decrease of detectable ipsilesional CST fibers by 98% in the anterior part of the posterior capsula interna, while leaving evidence of most preserved fibers in the anterior part of the internal capsule. It was proposed that the anterior fibers of the CST originating from anterior parts of the primary motor cortex (M1) or PMC might constitute the anatomical pre-requisite for the observed clinical improvement. Analysis of fMRI data revealed increased activity in the ipsilesional dorsal premotor region and supplementary motor areas at the end of the last BCI training block, and a significant increase in fractional anisotropy (FA) reflecting white matter microstructure’s integrity in the affected CST. This proof-of-principle study provided encouraging data that SMR-based BCI training coupled with goal-directed physiotherapy might induce beneficial used-dependent plasticity in the perilesional areas facilitating motor recovery.
Another study by Ang and colleagues (2010) [102] compared two groups of sub-acute and chronic stroke patients (1–35 months after stroke) with predominantly sub-cortical brain lesions (80%) who received either a standardized (n = 10) or BCI-driven (n = 8) robotic rehabilitation, which was applied over 12 sessions within 4 weeks. During the standardized robotic rehabilitation training the participant’s affected arm was strapped to a robotic device (MIT-Manus). Participants were instructed to move their paretic hand according to a visually presented goal on a screen in front of them. If the subject could not perform the movement on their own, the robot would assist and actively guide the subject’s arm towards the goal. In the BCI group, assistive movements were only performed if SMR-ERD was detectable over the affected hemisphere during the trial. Both groups were clinically evaluated using FMA before and after the training. FMA scores ranged between 4 and 61 points (mean 29.7 +/− 17.7) before training onset. Correcting for age and gender among the subjects with positive gain, the BCI group improved more and yielded a higher gain 2-month post-rehabilitation compared to the group that received standardized robotic rehabilitation. Besides limitations of this study due to the small sample size as well as heterogeneity of the groups regarding lesion site, age and time after stroke, it provided supportive data on the potential benefit of BCI training in the context of stroke rehabilitation.
A study performed by Daly et al. (2009) [29] combined an EEG-BCI training with FES of paralyzed finger muscles. A 43-year-old 10-months post-stroke patient with a lesion affecting the left cortical and sub-cortical regions of the frontal and parietal lobe underwent nine sessions of BCI-FES training within 3 weeks. During the training the patient had to either imagine or attempt finger movements on the affected side in alternation with attempted relaxation. Before the training, the patient could not actively extent the affected index finger. Sustained motor-related ERD was translated in activation of the FES device. During the BCI sessions the patient achieved good BCI control (over 88% in eight of nine sessions for attempted movement) and regained 26 degrees of volitional isolated index finger extension after session nine.
While all these reports are encouraging, larger controlled clinical studies are necessary to further evaluate the potential role of non-invasive assistive and restorative BCI technology in the rehabilitation of stroke and neurotrauma. Anatomical and functional pre-requisites for successful SMR-based BCI learning and mechanisms underlying clinical improvements need to be identified and well characterized.
As some of the stroke patients did not gain SMR-based BCI control, strategies to improve BCI learning would be of particular importance. In this context, techniques that can be used to modulate cortical plasticity such as transcranial direct current stimulation (tDCS) or transcranial magnetic stimulation (TMS) could be helpful tools to develop better BCI training protocols in patient groups.
Improving BCI Performance with Brain Stimulation
It was shown that transcranial direct current stimulation (tDCS), a non-invasive and well-tolerated method based on application of weak direct currents (e.g. 1 mA delivered for 20 min) through saline-soaked sponges attached to the head, can induce polarity specific changes of excitability in M1 [103] and, thus, enhance activity within M1. Further studies suggested that tDCS over M1 could be used to improve motor learning and consolidation [104–106]. After stroke, modulation of M1 excitability of the affected hemisphere by anodal tDCS was associated with motor function improvements of the paretic hand [94, 107]. A pilot study on combined tDCS and robot-assisted arm training by Hesse et al. (2007) [94] indicated beneficial effects on motor function (assessed by FMA) and aphasia in several participants.
Another non-invasive and well-established technique to modulate brain excitability is transcranial magnetic stimulation (TMS). In TMS a magnetic field is used to induce a small electric current that can lead to depolarization of cortical neurons. Depending on the intensity and frequency of stimulation, TMS can have lasting effects on the excitability of the brain when delivered repetitively (rTMS). Also, based on the finding that rTMS can elicit long lasting excitatory or inhibitory effects, use of rTMS as a therapeutic tool in neurological and psychiatric disorders, such as depression [108], chronic pain [109], epilepsy [110] or movement disorders [111] became investigated. In stroke, low-frequency (inhibitory) rTMS was used to reduce cortical excitability in the unaffected primary motor cortex and resulted in transient improvements of motor function [112–114]. Targeting the affected hemisphere of stroke patients with high-frequency (excitatory) rTMS, motor function improvements were reported [115, 116].
As SMR-related ERD can be interpreted as an electrophysiological correlate of cortical activation [117], anodal tDCS or high-frequency rTMS applied to the affected motor cortex might be a useful tool to improve ERD-dependent BCI performance in stroke patients and hence enhance practicability of assistive and restorative BCI systems.
Prospects of BCI Applications in Stroke and Neurotrauma
The development of assistive and restorative BCI technology for rehabilitation of stroke or neurotrauma is an exciting emerging field that yields notable potential to improve quality of life for many affected people. So far, only few studies on application of invasive or non-invasive BCI technology in patients with stroke or neurotrauma are available [19, 26, 29, 102]. Mechanisms underlying voluntary SMR-modulation for BCI control are not well understood. Optimal settings and algorithms for BCI training protocols in patients with brain lesions are unknown. Therefore, studies based on MEG recordings for BCI control are of particular importance as MEG allows precise and relatively artifact-free post-hoc analysis of cortical activity patterns.
EEG-based BCI systems have the best potential for widespread clinical use. However, preparation time and sensitivity to muscle artifacts limit their practicability. The development of dry-electrode-systems with portable amplifiers offers a promising perspective. In this context the combination with FES systems and simultaneous electric brain stimulation represents a propitious vista for both, assistive and restorative BCI systems. While costly at present, BCI systems based on NIRS might become an attractive alternative to EEG.
New concepts for innovative BCI approaches based on measures that are currently too complex for online applications, such as dual-core beamforming [118] that allow identification of simultaneously active correlated networks, might become feasible once required computational capacities are available.
More and larger clinical studies are needed to develop optimal protocols for both, assistive and restorative BCI applications. Due to the heterogeneity of patient populations, multimodal approaches to evaluate subject specific characteristics in terms of anatomy and function including e.g. fMRI, DTI, MEG and diagnostic TMS are an important pre-requisite for a better understanding of BCI related neuroplasticity and might help to develop new strategies for BCI use in neurorehabilitation.
Conclusion
Assistive and restorative BCI technology might be a powerful tool to improve rehabilitation strategies in patients with brain lesions and severe motor paralysis, such as stroke or traumatic brain injury.
References
Velliste M, Perel S, Spalding MC, Whitford AS, Schwartz AB (2008) Cortical control of a prosthetic arm for self-feeding. Nature 453:1098–1101
Hochberg LR, Serruya MD, Friehs GM, Mukand JA, Saleh M, Caplan AH, Branner A, Chen D, Penn RD, Donoghue JP (2006) Neural ensemble control of prosthetic devices by a human with tetraplegia. Nature 442:164–171
Pfurtscheller G, Guger C, Muller G, Krausz G, Neuper C (2000) Brain oscillations control hand orthosis in a tetraplegic. Neurosci Lett 292:211–214
Birbaumer N, Cohen LG (2007) Brain–computer interfaces: communication and restoration of movement in paralysis. J Physiol 579:621–636
Birbaumer N, Ramos Murguialday A, Weber C, Montoya P (2009) Neurofeedback and brain–computer interface clinical applications. Int Rev Neurobiol 86:107–117
Daly JJ, Wolpaw JR (2008) Brain–computer interfaces in neurological rehabilitation. Lancet Neurol 7:1032–1043
Berger H (1929) Ueber das Elektrenkephalogramm des Menschen. Archiv für Psychiatrie und Nervenkrankheiten 87:527–570
Serruya MD, Hatsopoulos NG, Paninski L, Fellows MR, Donoghue JP (2002) Instant neural control of a movement signal. Nature 416:141–142
Taylor DM, Tillery SI, Schwartz AB (2002) Direct cortical control of 3D neuroprosthetic devices. Science 296:1829–1832
Carmena JM, Lebedev MA, Crist RA, O’Doherty JA, Santucci DM, Dimitrov DF, Patil PG, Henriquez CS, Nicolelis MAL (2003) Learning to control a brain–machine interface for reaching and grasping by primates. PLoS Biol 1:1–16
Donoghue JP, Nurmikko A, Black M, Hochberg LR (2007) Assistive technology and robotic control using motor cortex ensemble-based neural interface systems in humans with tetraplegia. J Physiol 579:603–611
Birbaumer N, Ghanayim N, Hinterberger T, Iversen I, Kotchoubey B, Kubler A, Perelmouter J, Taub E, Flor H (1999) A spelling device for the paralyzed. Nature 398:297–298
Pfurtscheller G, Graimann B, Huggins JE, Levine SP (2004) Brain–computer communication based on the dynamics of brain oscillations. Suppl Clin Neurophysiol 57:583–591
Wolpaw JR, Birbaumer N, McFarland DJ, Pfurtscheller G, Vaughan TM (2002) Brain–computer interfaces for communication and control. Clin Neurophysiol 113:767–791
McFarland DJ, Krusienski DJ, Sarnacki WA, Wolpaw JR (2008) Emulation of computer mouse control with a noninvasive brain–computer interface. J Neural Eng 5:101–110
Blankertz B, Dornhege G, Krauledat M, Muller KR, Curio G (2007) The non-invasive Berlin brain–computer interface: fast acquisition of effective performance in untrained subjects. Neuroimage 37:539–550
Sitaram R, Caria A, Birbaumer N (2009) Hemodynamic brain–computer interfaces for communication and rehabilitation. Neural Netw 22:1320–1328
Kübler A, Nijboer F, Mellinger J, Vaughan TM, Pawelzik H, Schalk G, McFarland DJ, Birbaumer N, Wolpaw JR (2005) Patients with ALS can use sensorimotor rhythms to operate a brain–computer interface. Neurology 64:1775–1777
Buch E, Weber C, Cohen LG, Braun C, Dimyan MA, Ard T, Mellinger J, Caria A, Soekadar S, Fourkas A, Birbaumer N (2008) Think to move: a neuromagnetic brain–computer interface (BCI) system for chronic stroke. Stroke 39:910–917
Organization WH (2003) The World Health report: shaping the future. World Health Organization, Geneva, p 2003
Kwakkel G, Kollen BJ, van der Grond J, Prevo AJ (2003) Probability of regaining dexterity in the flaccid upper limb: impact of severity of paresis and time since onset in acute stroke. Stroke 34:2181–2186
Rosamond W, Flegal K, Furie K, Go A, Greenlund K, Haase N, Hailpern SM, Ho M, Howard V, Kissela B, Kittner S, Lloyd-Jones D, McDermott M, Meigs J, Moy C, Nichol G, O’Donnell C, Roger V, Sorlie P, Steinberger J, Thom T, Wilson M, Hong Y (2008) Heart disease and stroke statistics – 2008 update: a report from the American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Circulation 117:e25–e146
Wolf SL, Winstein CJ, Miller JP, Taub E, Uswatte G, Morris D, Giuliani C, Light KE, Nichols-Larsen D, EXCITE Investigators (2006) Effect of constraint-induced movement therapy on upper extremity function 3 to 9 months after stroke – the EXCITE randomized clinical trial. JAMA 296:2095–2104
Luft AR, McCombe-Waller S, Whitall J, Forrester LW, Macko R, Sorkin JD, Schulz JB, Goldberg AP, Hanley DF (2004) Repetitive bilateral arm training and motor cortex activation in chronic stroke: a randomized controlled trial. JAMA 292:1853–1861
Pfurtscheller G, Müller GR, Pfurtscheller J, Gerner HJ, Rupp R (2003) ‘Thought’ – control of functional electrical stimulation to restore hand grasp in a patient with tetraplegia. Neurosci Lett 351:33–36
Broetz D, Braun C, Weber C, Soekadar SR, Caria A, Birbaumer N (2010) Combination of brain–computer interface training and goal-directed physical therapy in chronic stroke: a case report. Neurorehabil Neural Repair 24:674–679
Caria A, Weber C, Brötz D, Ramos A, Ticini LF, Gharabaghi A, Braun C, Birbaumer N (2010) Chronic stroke recovery after combined BCI training and physiotherapy: A case report. Psychophysiology 48:578–582
Nagaoka T, Sakatani K, Awano T, Yokose N, Hoshino T, Murata Y, Katayama Y, Ishikawa A, Eda H (2010) Development of a new rehabilitation system based on a brain–computer interface using near-infrared spectroscopy. Adv Exp Med Biol 662:497–503
Daly JJ, Cheng R, Rogers J, Litinas K, Hrovat K, Dohring M (2009) Feasibility of a new application of noninvasive brain computer interface (BCI): a case study of training for recovery of volitional motor control after stroke. J Neurol Phys Ther 33:203–211
Fetz EE (1969) Operant conditioning of cortical unit activity. Science 163:955–958
Georgopoulos AP, Schwartz AB, Kettner RE (1986) Neuronal population coding of movement direction. Science 233:1416–1419
Georgopoulos AP, Lurito JT, Petrides M, Schwartz AB, Massey JT (1989) Mental rotation of the neuronal population vector. Science 243:234–236
Nicolelis MA (2003) Brain–machine interfaces to restore motor function and probe neural circuits. Nat Rev Neurosci 4:417–422
Scherberger H, Jarvis MR, Andersen RA (2005) Cortical local field potentials encodes movement intentions in the posterior parietal cortex. Neuron 46:347–354
Dickey AS, Suminski A, Amit Y, Hatsopoulos NG (2009) Single-unit stability using chronically implanted multielectrode arrays. J Neurophysiol 102:1331–1339
Rousche PJ, Normann RA (1998) Chronic recording capability of the Utah intracortical electrode array in cat sensory cortex. J Neurosci Methods 82:1–15
Fountas KN, Smith JR (2007) Subdural electrode-associated complications: a 20-year experience. Stereotact Funct Neurosurg 85:264–272
Penfield W, Welch K (1951) The supplementary motor area of the cerebral cortex; a clinical and experimental study. AMA Arch Neurol Psychiatry 66:289–317
Fulton JF (1934) A note on the definition of the motor and premotor areas. Brain 57:311–316
Fulton JF (1935) Definition of the ‘motor’ and ‘premotor’ areas. Brain 58:311–316
Dum RP, Strick PL (1991) The origin of corticospinal projections from the premotor areas in the frontal lobe. J Neurosci 11:667–689
Vargas-Irwin CE, Shakhnarovich G, Yadollahpour P, Mislow JM, Black MJ, Donoghue JP (2010) Decoding complete reach and grasp actions from local primary motor cortex populations. J Neurosci 30:9659–9669
Schalk G, Miller KJ, Anderson NR, Wilson JA, Smyth MD, Ojemann JG, Moran DW, Wolpaw JR, Leuthardt EC (2008) Two-dimensional movement control using electrocorticographic signals in humans. J Neural Eng 5:75–84
Leuthardt EC, Schalk G, Wolpaw JR, Ojemann JG, Moran DW (2004) A brain–computer interface using electrocorticographic signals in humans. J Neural Eng 1:63–71
Freeman WJ, Rogers LJ, Holmes MD, Silbergeld DL (2000) Spatial spectral analysis of human electrocorticograms including the alpha and gamma bands. J Neurosci Methods 95:111–121
Staba RJ, Wilson CL, Bragin A, Fried I, Engel J Jr (2002) Quantitative analysis of high-frequency oscillations (80–500 Hz) recorded in human epileptic hippocampus and entorhinal cortex. J Neurophysiol 88:1743–1752
Ball T, Kern M, Mutschler I, Aertsen A, Schulze-Bonhage A (2009) Signal quality of simultaneously recorded invasive and non-invasive EEG. Neuroimage 46:708–716
Chao ZC, Nagasaka Y, Fujii N (2010) Long-term asynchronous decoding of arm motion using electrocorticographic signals in monkeys. Front Neuroeng 3:3
Bradberry TJ, Gentili RJ, Contreras-Vidal JL (2010) Reconstructing three-dimensional hand movements from noninvasive electroencephalographic signals. J Neurosci 30:3432–3437
Waldert S, Preissl H, Demandt E, Braun C, Birbaumer N, Aertsen A, Mehring C (2008) Hand movement direction decoded from MEG and EEG. J Neurosci 28:1000–1008
Lebedev MA, Nicolelis MA (2006) Brain–machine interfaces: past, present and future. Trends Neurosci 29:536–546
Skinner F (1953) Science and human behavior. Macmillan, New York
Wolpaw JR, Birbaumer N, Heetderks WJ, McFarland DJ, Peckham PH, Schalk G, Donchin E, Quatrano LA, Robinson CJ, Vaughan TM (2000) Brain–computer interface technology: a review of the first international meeting. IEEE Trans Rehabil Eng 8:164–173
Kübler A, Kotchoubey B, Kaiser J, Wolpaw J, Birbaumer N (2001) Brain–computer communication: unlocking the locked-in. Psychol Bull 127:358–375
Birbaumer N (2006) Breaking the silence: brain–computer interfaces (BCI) for communication and motor control. Psychophysiology 43:517–532
Birbaumer N, Hinterberger T, Kübler A, Neumann N (2003) The thought-translation device (TTD): neurobehavioral mechanisms and clinical outcome. IEEE Trans Neural Syst Rehabil Eng 11:120–123
Perelmouter J, Birbaumer N (2000) A binary spelling interface with random errors. IEEE Trans Rehabil Eng 8:227–232
Hinterberger T, Veit R, Wilhelm B, Weiskopf N, Vatine JJ, Birbaumer N (2005) Neuronal mechanisms underlying control of a brain–computer-interface. Eur J Neurosci 21:3169–3181
Wilhelm B, Jordan M, Birbaumer N (2006) Communication in locked-in syndrome: effects of imagery on salivary pH. Neurology 67:534–535
Gastaut H, Terzian H, Gastaut Y (1952) Study of a little electroencephalographic activity: rolandic arched rhythm. Mars Med 89:296–310
Howe RC, Sterman MB (1972) Cortical–subcortical EEG correlates of suppressed motor behavior during sleep and waking in the cat. J Electroencephalogr Clin Neurophysiol 32:681–695
Pfurtscheller G, Aranibar A (1979) Evaluation of event-related desynchronization (ERD) preceding and following self-paced movement. Electroencephgr Clin Neurophysiol 46:138–146
Leocani L, Toro C, Zhuang P, Gerloff C, Hallet M (2001) Event-related desynchronization in reaction time paradigms: a comparison with event-related potentials and corticospinal excitability. Clin Neurophysiol 112:923–930
Pfurtscheller G, Stancák A Jr, Neuper C (1996) Event-related synchronization (ERS) in the alpha band – an electrophysiological correlate of cortical idling: a review. Int J Psychophysiol 24:39–46
Wolpaw JR, McFarland DJ (2004) Control of a two-dimensional movement signal by a noninvasive brain–computer interface in humans. Proc Natl Acad Sci USA 101:17849–17854
Wolpaw JR (2007) Brain–computer interfaces as new brain output pathways. J Physiol 579:613–619
Mellinger J, Schalk G, Braun C, Preissl H, Rosenstiel W, Birbaumer N, Kübler A (2007) An MEG-based brain–computer interface (BCI). Neuroimage 36:581–593
Farwell LA, Donchin E (1988) Talking off the top of your head: toward a mental prosthesis utilizing event-related brain potentials. Electroencephalogr Clin Neurophysiol 70:510–523
Lenhardt A, Kaper M, Ritter HJ (2008) An adaptive P300-based online brain–computer interface. IEEE Trans Neural Syst Rehabil Eng 16:121–130
Weiskopf N, Veit R, Erb M, Mathiak K, Grodd W, Goebel R, Birbaumer N (2003) Physiological self-regulation of regional brain activity using real-time functional magnetic resonance imaging (fMRI): methodology and exemplary data. NeuroImage 19:577–586
Yoo SS, Fairneny T, Chen NK, Choo SE, Panych LP, Park H, Lee SY, Jolesz FA (2004) Brain–computer interface using fMRI: spatial navigation by thoughts. Neuroreport 15:1591–1595
DeCharms RC, Maeda F, Glover GH, Ludlow D, Pauly JM, Soneji D, Gabrieli JD, Mackey SC (2005) Control over brain activation and pain learned by using real-time functional MRI. Proc Natl Acad Sci USA 102:18626–18631
Caria A, Veit R, Sitaram R, Lotze M, Weiskopf N, Grodd W, Birbaumer N (2007) Regulation of anterior insular cortex activity using real-time fMRI. NeuroImage 35:1238–1246
Logothetis N, Pauls J, Augath M, Trinath T, Oeltermann A (2001) Neurophysiological investigation of the basis of the fMRI signal. Nature 412:150–157
Elbert T, Rockstroh B, Lutzenberger W, Birbaumer N (1984) Self-regulation of the brain and behavior. Springer, New York
Seifert AR, Lubar JF (1975) Reduction of epileptic seizures through EEG biofeedback training. Biol Psychol 3:157–184
Kotchoubey B, Strehl U, Uhlmann C, Holzapfel S, König M, Fröscher W, Blankenhorn V, Birbaumer N (2001) Modification of slow cortical potentials in patients with refractory epilepsy: a controlled outcome study. Epilepsia 42:406–416
Birbaumer N, Elbert T, Rockstroh B, Lutzenberger W (1986) Biofeedback of slow cortical potentials in attentional disorders. In: McCallum WC, Zappoli R, Denoth F (eds) Cerebral psychophysiology: studies in event-related potentials. Elsevier, Amsterdam
Strehl U, Leins U, Goth G, Klinger C, Hinterberger T, Birbaumer N (2006) Self-regulation of slow cortical potentials: a new treatment for children with attention-deficit/hyperactivity disorder. Pediatrics 118:1530–1540
Fuchs T, Birbaumer N, Lutzenberger W, Gruzelier JH, Kaiser J (2003) Neurofeedback training for attention-deficit/hyperactivity disorder in children: a comparison with methylphenidate. Appl Psychophysiol Biofeedback 28:1–12
Lotze M, Grodd W, Birbaumer N, Erb M, Huse E, Flor H (1999) Does use of a myoelectric prosthesis prevent cortical reorganization and phantom limb pain? Nat Neurosci 2:501–502
Chklovskii DB, Mel BW, Svoboda K (2004) Cortical rewiring and information storage. Nature 431:782–788
Frost SB, Barbay S, Friel KM, Plautz EJ, Nudo RJ (2003) Reorganization of remote cortical regions after ischemic brain injury: a potential substrate for stroke recovery. J Neurophysiol 89:3205–3214
Murase N, Duque J, Mazzocchio R, Cohen LG (2004) Influence of interhemispheric interactions on motor function in chronic stroke. Ann Neurol 55:400–409
Duque J, Hummel F, Celnik P, Murase N, Mazzocchio R, Cohen LG (2005) Transcallosal inhibition in chronic subcortical stroke. Neuroimage 28:940–946
Grefkes C, Nowak DA, Eickhoff SB, Dafotakis M, Küst J, Karbe H, Fink GR (2008) Cortical connectivity after subcortical stroke assessed with functional magnetic resonance imaging. Ann Neurol 63:236–246
Harris-Love ML, Perez MA, Chen R, Cohen LG (2007) Interhemispheric inhibition in distal and proximal arm representations in the primary motor cortex. J Neurophysiol 97:2511–2515
Floel A, Nagorsen U, Werhahn KJ, Ravindran S, Birbaumer N, Knecht S, Cohen LG (2004) Influence of somatosensory input on motor function in patients with chronic stroke. Ann Neurol 56:206–212
Conforto AB, Kaelin-Lang A, Cohen LG (2002) Increase in hand muscle strength of stroke patients after somatosensory stimulation. Ann Neurol 51:122–125
Scheidtmann K (2004) Advances in adjuvant pharmacotherapy for motor rehabilitation: effects of levodopa. Restor Neurol Neurosci 22:393–398
Liu KP, Chan CC, Wong RS, Kwan IW, Yau CS, Li LS, Lee TM (2009) A randomized controlled trial of mental imagery augment generalization of learning in acute poststroke patients. Stroke 40:2222–2225
Malouin F, Richards CL, Doyon J, Desrosiers J, Belleville S (2004) Training mobility tasks after stroke with combined mental and physical practice: a feasibility study. Neurorehabil Neural Repair 18:66–75
Page SJ, Levine P, Leonard A (2007) Mental practice in chronic stroke: results of a randomized, placebo-controlled trial. Stroke 38:1293–1297
Hummel F, Celnik P, Giraux P, Floel A, Wu WH, Gerloff C, Cohen LG (2005) Effects of non-invasive cortical stimulation on skilled motor function in chronic stroke. Brain 128:490–499
Hesse S, Werner C, Schonhardt EM, Bardeleben A, Jenrich W, Kirker SG (2007) Combined transcranial direct current stimulation and robot-assisted arm training in subacute stroke patients: a pilot study. Restor Neurol Neurosci 25:9–15
Takeuchi N, Chuma T, Matsuo Y, Watanabe I, Ikoma K (2005) Repetitive transcranial magnetic stimulation of contralesional primary motor cortex improves hand function after stroke. Stroke 36:2681–2686
Platz T, Kim IH, Engel U, Kieselbach A, Mauritz KH (2002) Brain activation pattern as assessed with multi-modal EEG analysis predict motor recovery among stroke patients with mild arm paresis who receive the arm ability training. Restor Neurol Neurosci 20:21–35
Calautti C, Naccarato M, Jones PS, Sharma N, Day DD, Carpenter AT, Bullmore ET, Warburton EA, Baron JC (2007) The relationship between motor deficit and hemisphere activation balance after stroke: a 3 T fMRI study. Neuroimage 34:322–331
Ward NS, Cohen LG (2004) Mechanisms underlying recovery of motor function after stroke. Arch Neurol 61:1844–1848
Taub E, Uswatte G, Mark VW, Morris DM (2006) The learned nonuse phenomenon: implications for rehabilitation. Eura Medicophys 42:241–256
Buch ER, Fourkas AD, Weber C, Birbaumer N, Cohen LG (2010) Anatomical parieto-frontal connectivity predicts performance gains in μ rhythm-based brain–computer interface (BCI) training in chronic stroke. SFN 2010, San Diego, 493.6/FFF19
Ang KK, Guan C, Chua KS, Ang BT, Kuah C, Wang C, Phua KS, Chin ZY, Zhang H (2010) Clinical study of neurorehabilitation in stroke using EEG-based motor imagery brain–computer interface with robotic feedback. Conf Proc IEEE Eng Med Biol Soc 1:5549–5552
Nitsche MA, Paulus W (2000) Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J Physiol 527:633–639
Nitsche MA, Schauenburg A, Lang N, Liebetanz D, Exner C, Paulus W, Tergau F (2003) Facilitation of implicit motor learning by weak transcranial direct current stimulation of the primary motor cortex in the human. J Cognit Neurosci 15:619–626
Reis J, Schambra HM, Cohen LG, Buch ER, Fritsch B, Zarahn E, Celnik PA, Krakauer JW (2009) Noninvasive cortical stimulation enhances motor skill acquisition over multiple days through an effect on consolidation. Proc Natl Acad Sci USA 106:1590–1595
Antal A, Nitsche MA, Kincses TZ, Kruse W, Hoffmann KP, Paulus W (2004) Facilitation of visuo-motor learning by transcranial direct current stimulation of the motor and extrastriate visual areas in humans. Eur J Neurosci 19:2888–2892
Boggio PS, Nunes A, Rigonatti SP, Nitsche MA, Pascual-Leone A, Fregni F (2007) Repeated sessions of noninvasive brain DC stimulation is associated with motor function improvement in stroke patients. Restor Neurol Neurosci 25:123–129
Miniussi C, Bonato C, Bignotti S, Gazzoli A, Gennarelli M, Pasqualetti P, Tura GB, Ventriglia M, Rossini PM (2005) Repetitive transcranial magnetic simulation (rTMS) at high and low frequency: an efficacious therapy for major drug-resistant depression? Clin Neurophysiol 116:1062–1071
Lefaucheur JP (2004) Transcranial magnetic stimulation in the management of pain. Clin Neurophysiol (Suppl) 57:737–748
Theodore WH (2003) Transcranial magnetic stimulation in epilepsy. Epilepsy Curr 3:191–197
Fregni F, Simon DK, Wu A, Pascual-Leone A (2005) Non-invasive brain stimulation for Parkinson’s disease: a systematic review and meta-analysis of the literature. J Neurol Neurosurg Psychiatry 76:1614–1623
Mansur CG, Fregni F, Boggio PS, Riberto M, Gallucci-Neto J, Santos CM, Wagner T, Rigonatti SP, Marcolin MA, Pascual-Leone A (2005) A sham stimulation-controlled trial of rTMS of the unaffected hemisphere in stroke patients. Neurology 64:1802–1804
Takeuchi N, Tada T, Toshima M, Chuma T, Matsuo Y, Ikoma K (2008) Inhibition of the unaffected motor cortex by 1 Hz repetitive transcranical magnetic stimulation enhances motor performance and training effect of the paretic hand in patients with chronic stroke. J Rehabil Med 40:298–303
Fregni F, Boggio PS, Valle AC, Rocha RR, Duarte J, Ferreira MJ, Wagner T, Fecteau S, Rigonatti SP, Riberto M, Freedman SD, Pascual-Leone A (2006) A sham-controlled trial of a 5-day course of repetitive transcranial magnetic stimulation of the unaffected hemisphere in stroke patients. Stroke 37:2115–2122
Khedr EM, Ahmed MA, Fathy N, Rothwell JC (2005) Therapeutic trial of repetitive transcranial magnetic stimulation after acute ischemic stroke. Neurology 65:466–468
Kim YH, You SH, Ko MH, Park JW, Lee KH, Jang SH, Yoo WK, Hallett M (2006) Repetitive transcranial magnetic stimulation-induced corticomotor excitability and associated motor skill acquisition in chronic stroke. Stroke 37:1471–1476
Pfurtscheller G, Klimesch W (1992) Functional topography during a visuoverbal judgment task studied with event-related desynchronization mapping. J Clin Neurophysiol 9:120–131
Diwakar M, Huang MX, Srinivasan R, Harrington DL, Robb A, Angeles A, Muzzatti L, Pakdaman R, Song T, Theilmann RJ, Lee RR (2011) Dual-core beamformer for obtaining highly correlated neuronal networks in MEG. Neuroimage 54:253–263
Acknowledgements
This contribution was supported by the NINDS intramural research program of the National Institutes of Health (NIH), the Deutsche Forschungsgemeinschaft (DFG) and the German Ministry of Education and Research (BMBF, 01GQ0831).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer
About this paper
Cite this paper
Soekadar, S.R., Birbaumer, N., Cohen, L.G. (2011). Brain–Computer Interfaces in the Rehabilitation of Stroke and Neurotrauma. In: Kansaku, K., Cohen, L.G. (eds) Systems Neuroscience and Rehabilitation. Springer, Tokyo. https://doi.org/10.1007/978-4-431-54008-3_1
Download citation
DOI: https://doi.org/10.1007/978-4-431-54008-3_1
Publisher Name: Springer, Tokyo
Print ISBN: 978-4-431-53998-8
Online ISBN: 978-4-431-54008-3
eBook Packages: MedicineMedicine (R0)