
Abstract
The emissions of carbon dioxide and methane from aquatic ecosystems to the atmosphere are a significant component of the carbon cycle in the Amazon basin. Recent estimates of inundated areas, of gas transfer velocities, and of concentrations and fluxes of carbon dioxide in diverse aquatic habitats are used to update, and extend to the whole lowland basin, estimates of carbon dioxide emissions. Total floodable area derived from remotely sensed data with a spatial resolution of about 100 m in the Amazon basin <500 m above sea level is approximately 800,000 km2. Combination of channel geometry and lengths for streams and rivers results in an estimate of approximately 60,000 km2 for channel areas. Gas transport velocities derived from mechanistic models and measurements in a variety of standing and flowing waters indicate values are higher than those used in prior calculations of carbon dioxide emission. Combination of updated inundated areas and gas transfer velocities with numerous new measurements of carbon dioxide concentrations and fluxes from streams, rivers, lakes, reservoirs, and wetlands results in an estimate up to approximately 1800 Tg C year−1 for the whole lowland Amazon basin. The majority of the basin-wide evasion of carbon dioxide is likely derived from atmospheric carbon dioxide fixed by photosynthesis by higher plants in aquatic environments. Further research is needed on carbon inputs, sedimentation, and emission of both carbon dioxide and methane from all habitats with particular attention to those with flooded forests and floating herbaceous plants, and on areal coverage and emission from streams.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
The processing of carbon by aquatic ecosystems of inland waters is now recognised as a significant component of regional and global carbon dynamics. In particular, the high rates of sedimentation in lakes and reservoirs and considerable evasion of carbon dioxide and methane from many rivers, lakes, and wetlands lead to fluxes disproportionately large relative to the area of inland waters (Cole et al. 2007; Downing 2009; Battin et al. 2009; Aufdenkampe et al. 2011; Butman and Raymond 2011; Raymond et al. 2013; Stanley et al. 2015). Although the magnitude and variability of these fluxes remain uncertain, especially in tropical regions, recent studies are improving our understanding of carbon dynamics in the streams, rivers, lakes, reservoirs, and wetlands of the Amazon basin. The Large-scale Biosphere Atmosphere Experiment in Amazonia (LBA) and related activities have resulted in numerous relevant publications, many of which have been summarised in a recent monograph (Gash et al. 2009). In particular, Richey et al. (2009) described carbon processing from streams and rivers, and Melack et al. (2009) examined ecosystem processes in inundated areas. From these studies, it appears that evasion of carbon dioxide from Amazonian rivers, lakes, and temporally inundated aquatic habitats is of similar magnitude to net ecosystem exchanges in non-inundated upland forests (terra firme forests) derived from eddy covariance measurements. In addition, Amazonian aquatic ecosystems account for a significant proportion of the global methane flux from natural wetlands.
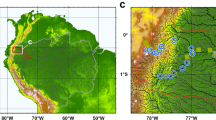
With a focus on exchanges of carbon dioxide and methane between inland waters and the atmosphere, the purpose of this chapter is to extend the regional extent of previous analyses to the whole lowland Amazon basin (regions <500 m above sea level; 5.05 million km2; Fig. 7.1; not including the Tocantins basin) by synthesis and critical evaluation of recent publications. Remote sensing analyses of inundation and wetland habitats, inundation modelling, measurements in rivers, reservoirs and other types of wetlands, and improved estimates of gas exchange coefficients for standing and flowing water are included. Regional estimates of gas fluxes are complemented with consideration of sources of the carbon being released, and remaining uncertainties and research needs are discussed. A discussion of climate trends and variability, exceptional events, and human impacts, as factors potentially altering carbon dynamics in the Amazon basin, is presented.

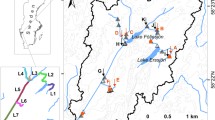
Lowland Amazon basin (area ≤500 m; grey) with floodable areas (white), as described in Melack and Hess (2010) and Hess et al. (2003). Small areas of wetlands on the northern and southern edges are not shown because remote sensing data used to develop the wetland distribution were not available there. Ba Balbina Reservoir; Ca Lake Calado; Cu Lake Curuai; Cn Lake Canaçari; Ja Jau River; JiP Ji-Paraná River; Ju Juruena watershed; Moxos Llanos de Moxos; Ro Roraima; UN upper Negro interfluvial wetlands
Several current projects are in the process of publishing their results, and will continue to contribute to our understanding of carbon dynamics in the Amazon, though only a portion is available for inclusion here. These projects include HYBAM (http://www.ore-hybam.org/index.php/eng/documents), CARBAMA (http://carbama.epoc.u-bordeaux1.fr/PUBLICATIONS.html), the ‘Rede Beija Rio’ network (http://boto.ocean.washington.edu/story/Amazon), A Biogeoquímica do Carbono e Mercúrio na Bacia Amazônica (Barbosa 2013) (http://www.biologia.ufrj.br/limnologia/projeto-carbono.php), and GEOMA (http://www.geoma.lncc.br/).
2 Inundation and the Variable Extent of Aquatic Habitats: Remote Sensing and Modelling
Carbon dynamics and carbon dioxide and methane exchanges in the Amazon basin vary among the diverse aquatic habitats. Junk et al. (2011) used information on climate, hydrology, water and sediment chemistry, and botany to delineate 14 major types of naturally occurring wetlands in the lowland Amazon. The amplitude, duration, frequency, and predictability of inundation are key criteria in this classification. While the classification has broad utility, data on all the criteria, including variations in inundation, are lacking for many parts of the Amazon. Hence, assigning spatial detail and areal extent to the various wetland types remains a challenge.
With a focus on gas exchange at the air–water interface, it is essential to have estimates of the surface area of the aquatic habitats, a daunting requirement given the large size of the Amazon basin and the wide range in dimensions of the habitats from headwater streams (<1 m across) to floodplains fringing major rivers (tens of km wide). While remote sensing can provide excellent information, currently available data on a regional scale have a spatial resolution of about 100 m, and sensors that allow seasonal and inter-annual variations in inundation to be recorded have spatial resolution of tens of km. Therefore, additional approaches are needed, especially for small rivers and streams, including geomorphology and modelling.
2.1 Remote Sensing
A variety of remote sensing approaches using passive and active microwave, laser, visible, and near-infrared and gravity anomaly detection systems are available and have been applied to tropical aquatic environments (Melack 2004). Melack and Hess (2010) applied the methodology of Hess et al. (2003), based primarily on mosaics of synthetic aperture radar (SAR) data obtained during a period of low and a period of high river stage, to the lowland Amazon basin to determine floodable area, inundated area, and areal extent of major habitats permanently or periodically inundated. Total floodable area within the lowland basin was estimated as 800,000 km2. However, portions of southern Bolivia were not covered by available SAR data, and floodplains in the south-western Brazilian Amazon were not well delineated owing to the timing of data acquisitions. Open water, floating macrophytes or grasslands, and flooded or unflooded forests were distinguished, and the spatial resolution of the products was about 100 m. Areal extent of the aquatic habitats was reported for 31 river basins and five reaches of the Solimões-Amazonas River. While not so specific as the classification proposed by Junk et al. (2011), these habitats and divisions by river basin are relevant to the biogeochemical processes considered here. High-resolution, remotely sensed products are available for specific locations in the Amazon and provide information appropriate for validation of basin-scale products (e.g. Silva et al. 2010; Renó et al. 2011; Hawes et al. 2012; Arnesen et al. 2013).
Several remote sensing approaches have been used to estimate the areas of open water in the lowland Amazon basin. Open water area was determined by Melack and Hess (2010) as 64,800 km2 from high water data. The Shuttle Radar Topography Mission (SRTM) offers digital elevations with 90 m horizontal postings (Jarvis et al. 2008) and a water body product (http://dds.cr.usgs.gov/srtm/version2_1/SWBD/). Open water area derived from the SRTM data is 72,000 km2, with river channels wider than approximately 300 m covering 52,000 km2. Hanson et al. (2013) provide a global composite of Landsat data at 30 m resolution from which an open water of 92,000 km2 can be derived (Forsberg, personal communication). Global datasets of lakes, such as those in Lehner and Döll (2004), need refinement to be applicable to the dynamic and spatially complex conditions in the Amazon.
Remote sensing of seasonal variations in inundated area depends on systems with coarse spatial resolution, and time series products are available at a 25-km scale. Hamilton et al. (2002, 2004) used data from satellite-borne passive microwave sensors to determine monthly inundation on the mainstem Amazon River floodplain (Brazil), the Llanos de Moxos (Bolivia), the Bananal (Brazil), and Roraima savannas (Brazil and Guyana). Regressions between flooded area and stage heights in nearby rivers were used to extend the records of inundation for nearly a century for the Amazon mainstem and for several decades in the other floodplains. Prigent et al. (2007), Papa et al. (2008, 2010), and Prigent et al. (2012) provide inundation estimates at 0.25° resolution, derived from several satellite-borne sensors for the period from 1993 to 2007. Comparison of these products with SAR-based estimates of inundation areas in the Amazon basin indicated generally good correspondence for moderate to large inundated units. Aires et al. (2013) used the 25-km resolution products in conjunction with SAR-based images to develop a wetland dataset for the Amazon for a 15-year period with a resolution of about 500 m. Schröder and McDonald (pers. comm.) are producing a monthly inundation product for the period from 2002 to 2009 from AMSR-E (passive microwave) and QSCAT (radar scatterometry) data at 0.25° resolution.
Gravity anomalies detected by the GRACE satellites provide estimates of changes in water volumes partially associated with seasonal variations in inundation at a scale of 100,000 s of km2, and have been used basin-wide in the Amazon (Alsdorf et al. 2010; Xavier et al. 2010). Frappart et al. (2005) combined satellite-derived SAR and altimetry data with in situ gauges to calculate water storage and inundation for the Negro River basin
2.2 Geomorphological Approaches to River Areas
Beighley and Gummadi (2011) combined relationships developed from hydraulic geometry with a high-resolution drainage network to estimate cumulative channel lengths and surface areas. Their analysis was done for drainage areas from 1 to 431,000 km2. For channels >2 m in width, they estimated that the Amazon basin contains c. 4.4 million km of channels with a combined area of 59,700 km2. Channels over 150 m in width were estimated to represent 29,500 km2 of the combined area.
To determine the area of streams and rivers in the Ji-Paraná basin (Fig. 7.1), Rasera et al. (2008) developed empirical relationships between drainage area and channel width combined with river lengths derived from a digital river network (Mayorga et al. 2005a). Areas of rivers from third to sixth order covered on average 342 km2 within the 75,400 km2 Ji-Paraná basin, representing 0.45 % of the basin. Assuming that a similar relationship applied to the whole lowland Amazon basin, an area of 22,700 km2 would result for rivers from third to sixth order. This value is similar to that estimated by Beighley and Gummadi (2011) for rivers in that size range.
Richey et al. (2002) estimated areas of moderate to small rivers and streams as a geometric series relating stream length and width to stream order. They calculated a total channel area of 21,000 km2 at low water for a 1.77 × 106 km2 area in the central Amazon, representing a fractional area of 1.2 %. If this fractional proportion is extrapolated to the whole lowland basin, a total channel area of 60,600 km2 would result.
Downing et al. (2012) employed stream network theory combined with data on stream width to approximate the areal extent of streams and rivers, when within their channels, on continental scales, and estimated that rivers and streams were likely to cover 0.30–0.56 % of land surfaces. Stream and river areas in the conterminous United States represent 0.52 % of the land surface (Butman and Raymond 2011). If these percentages are applied to the lowland Amazon basin with an area of 5.05 × 106 km2, areas for all rivers and streams range from 15,150 to 28,300 km2.
These different areas for river and stream channels vary depending on approach and region considered, and given the basin-wide, explicit method used by Beighley and Gummadi (2011), their values are adopted here. Headwater streams are not represented by their values. For large rivers, the area derived from the SRTM data is also used. Further refinement of stream areas is required.
2.3 Modelling of Inundation
Several models of river discharge and associated inundation dynamics have recently been developed and applied to the Amazon basin (Coe et al. 2007; Beighley et al. 2009; Victoria 2010; Yamazaki et al. 2011; Getirana et al. 2012; Paiva et al. 2013). Miguez-Macho and Fan (2012) compiled a groundwater dataset for the Amazon and combined it with a hydrological model. While generally successful at calculating river discharges, the models’ ability to represent inundation in the full range of floodplain and wetlands is only moderate, largely because of the lack of sufficiently accurate and detailed digital elevation models (DEMs). For example, Paiva et al. (2013) compared their modelled inundation in 3-month intervals with 0.25° remotely sensed estimates (Papa et al. 2010) averaged for the period from 1999 to 2004. The match was good in the central basin, but modelled areas significantly underestimated flooded area in the Bolivian Amazon and lower mainstem in Brazil. In the Peruvian Amazon, where the SAR analyses of Melack and Hess (2010) indicated a large area of wetland, the model also did so, but the coarse remotely sensed data did not. To better evaluate modelled results, it would be beneficial to examine SAR-derived products for specific periods and locations. Hydraulic models of flooding applied on a mesoscale have done well in locations with high-resolution DEMs (Wilson et al. 2007; Rudorff et al. 2014a, 2014b).
3 Gas Transfer Velocity Between Water and Atmosphere
Exchange of carbon dioxide and methane between surficial water and overlying atmosphere depends on the concentration gradient between air and water and on physical processes at the interface, usually parameterised as a gas transfer velocity (k), also called a piston velocity or gas exchange coefficient. Gas transfer velocities are a function of turbulence, kinematic viscosity of the water, and the molecular diffusion coefficient of the gas; the Schmidt number is the ratio of the latter two terms and is gas specific (MacIntyre et al. 1995). Schmidt numbers used here are normalised to carbon dioxide in freshwater at 20 °C and referred to as k(600) (Engle and Melack 2000). Gas transfer velocities are influenced by atmospheric stability and, in water, are altered by currents, wind, and convection, as well as rain (Ho et al. 2007), temperature, organic surficial films, and changes in hydrostatic pressure. Methane can also exit via bubbles (ebullition; Crill et al. 1988) and pass through tissues of rooted aquatic plants, both herbaceous and woody (Brix et al. 1992; Rice et al. 2010).
In lakes, direct measurements of exchange can be made with floating chambers, as has been done in the Amazon since the 1980s (e.g. Crill et al. 1988; Guérin et al. 2007). Alternatively, measurements of gas concentrations can be combined with estimates of k to calculate diffusive fluxes. While collecting and assaying samples for carbon dioxide and methane are fairly straightforward, the selection of appropriate k values remains a challenge. In a study at Lake Calado (Fig. 7.1), Crill et al. (1988) used a surface renewal model, which has a sound theoretical basis (Banerjee and MacIntyre 2004), as the basis for determination of k. Empirical relations between wind speed and k have been applied as well (e.g. Engle and Melack 2000; Guérin et al. 2007). Rudorff et al. (2011) used three different models of k: a simple wind-based equation, a small eddy version of the surface renewal model, and a wind-based model that includes diel heating and cooling (as described by MacIntyre et al. 2010). In a short-term experiment in the open water of a floodplain lake in the central Amazon basin, Polsenaere et al. (2013) applied an eddy covariance technique to calculate fluxes and k values for CO2.
Results from Rudorff et al. (2011) and Polsenaere et al. (2013) indicate that k values for standing waters in the Amazon are underestimated if based on simple wind-based relations commonly used. Rudorff et al. (2011) reported gas transfer coefficients that take into account wind as well as heating and cooling were on the order of 10 cm h−1. Polsenaere et al. (2013) reported k values ranging from 1.3 to 31.6 cm h−1, averaging 12.2 ± 6.7 cm h−1. Under conditions with high sensible and latent heat fluxes, but low wind speeds (<2.7 m s−1), k values near or above 20 cm h−1 were recorded. Based on floating chambers deployed in Balbina Reservoir (Fig. 7.1), Kemenes et al. (2011) calculated k values from 1.1 to 24.7 cm h−1, with an average of about 12 cm h−1. In contrast, Guérin et al. (2007) reported k values of about 2.5 cm h−1, based on chambers and eddy covariance in Petit Saut Reservoir, but noted that rain led to k values for CO2 from 0.8 to 13.4 cm h−1 at rain rates of 0.6–25 mm h−1. These results indicate that k values in lakes are generally higher than those used in prior regional extrapolations; e.g. Richey et al. (2002) used k values of 2.7 ± 1 cm h−1 for floodplains and lakes.
In warm tropical waters, such as those in the Amazon basin, latent heat fluxes are especially important and lead to convective mixing and enhanced k values. Conversely, diurnal heating under strong insolation can cause stable stratification of the water column that may lead to low or high k values. Hence, given the pronounced diel cycle of heating and cooling often observed in shallow tropical lakes, it is important to measure stratification and mixing, and gas exchanges, over these diel cycles, although such measurements are very seldom done.
The lack of studies of k values and gas concentrations in vegetated habitats adds further uncertainty, especially because of the large areas of flooded forests and floating macrophytes throughout the Amazon basin (Melack and Hess 2010; Junk et al. 2011). Though winds and direct heating are lower in vegetated habitats than in open waters, convective mixing and horizontal exchanges driven by differential heating and cooling and associated eddies, as water moves through the vegetation (Ortiz et al. 2013), will likely increase k values. Release of hydrophobic organic molecules by aquatic plants may reduce gas exchange within flooded vegetation.
Spatial variations in CO2 and CH4 concentrations can be large, as reported for the Amazon basin (Rudorff et al. 2011; Polsenaere et al. 2013; Abril et al. 2014) and elsewhere (Roland et al. 2010; Hofmann 2013). The high variability in time and space of bubbling adds further variance to methane evasion rates. Though floating chambers can capture bubbles, to increase spatial and temporal coverage submerged funnels are usually used. Recent applications of hydroacoustic measurements have allowed significant improvements in estimation of ebullition (Del Sontro et al. 2011), though this method has yet to be applied in the Amazon basin.
In flowing waters, most calculations of gas exchange are based on k in combination with concentration measurements, though direct measurements with floating chambers have also been used. Values of k for Amazon waters have been derived from 222Rn mass balances and floating chambers (Devol et al. 1987; Alin et al. 2011; Kemenes et al. 2011; Rasera et al. 2013). Alin et al. (2011) reported that values of k were significantly higher in small rivers and streams (channels <100 m wide), where current velocities and depth were found to be important, than in large rivers (channels >100 m wide), where wind was important. The range of k(600) values reported by Alin et al. (2011) for large rivers in the Amazon (1.2–31.1 cm h−1) is quite similar to that reported by Rasera et al. (2013) from a multi-year study in six non-tidal rivers in the Amazon basin (1.3–31.6 cm h−1). For the Uatumã River below Balbina Reservoir, Kemenes et al. (2011) reported average k values of 10.5 cm h−1. As in the case for lakes, these results indicate that k values are generally higher in flowing waters of the Amazon basin than those used in prior regional extrapolations; e.g. Richey et al. (2002) used k values of 9.6 ± 3.8 cm h−1 (Amazon mainstem) and 5 ± 2 cm h−1 (major tributaries).
4 Carbon Dioxide and Methane Concentrations and Fluxes
Recent measurements of carbon dioxide and methane concentrations and fluxes have been made in streams, rivers, wetlands, and a few lakes and reservoirs. These results are summarised by region with the intent of providing the basis for improved basin-wide estimates. Hence, areal fluxes are calculated for each habitat as Mg C km−2 year−1, kg C km−2 day−1, or g C m−2 year−1, as averages and/or ranges, if appropriate or possible.
4.1 Streams and Rivers
Working in remote headwater streams in the southern Amazon basin, Johnson et al. (2006, 2007, 2008) determined that most of the CO2 in the streams had been terrestrially respired within soils and that almost all was evaded to the atmosphere within headwater reaches. Baseflow delivered groundwater highly supersaturated in CO2, while during storms surface run-off and direct precipitation were relatively low in CO2. Concentrations of CO2 near the source of these headwater streams were 10,000–50,000 μatm. In a year-long study of a perennial, first-order stream in southern Mato Grosso draining a forested catchment, Neu et al. (2011) recorded pCO2 concentrations from 6490 to 14,980 μatm and evasion rates from the stream surface of c. 6490 ± 680 g C m−2 year−1; methane concentrations in the stream ranged from about 290–440 μatm and evasion averaged 990 ± 220 g C m−2 year−1. Similarly, Davidson et al. (2010) measured high pCO2 levels (average value of 19,000 μatm) in headwaters in remnant forests in northeastern Pará. Vihermaa et al. (2014) sampled two small streams and the La Torre and Tambopata rivers in the Madre de Dios region of the western Amazon; they reported CO2 evasion rates from 1866 to 82,900 mg C m−2 day−1 for the Tambopata River and a perennial stream, respectively. Working on small to moderate-sized rivers in the Ji-Paraná basin (Rondônia), Rasera et al. (2008) measured CO2 evasion rates per unit of river area from 695 to 13,095 mg C m−2 day−1 for third- and fourth-order rivers and from 622 to 4686 mg C m−2 day−1 for fifth- and sixth-order rivers in the basin.
Richey et al. (2009) reported pCO2 concentrations ranging from 500 to 20,000 μatm and illustrated a positive correlation of pCO2 with discharge over 4 years for the Solimões (at Manacapuru), the Madeira (at Porto Velho), and Ji-Paraná rivers. Borges et al. (2015) sampled along the mainstem Solimões and Amazon rivers and at the mouths of major tributaries and reported a range in pCO2 concentrations from 70 to 16,880 ppm. Ellis et al. (2012) measured concentrations from 860 μatm in the Acre River to 12,900 μatm in a stream in campina vegetation. Based on a 5-year study with seasonal sampling of six non-tidal rivers (Negro, Solimões, Teles Pires, Cristalino, Araguaia, and Javaés) and one tidal river (Caxiuanã), Rasera et al. (2013) reported a range of pCO2 concentrations from 259 to 7808 μatm and demonstrated a strong correlation between pCO2 and discharge. They reported a range in CO2 flux from uptake of 830 mg C m−2 day−1 to evasion of 15,860 mg C m−2 day−1. Uptake occurred in clear water rivers at low water in conditions conducive to algal growth. Alin et al. (2011) reported a similar range of evasion rates (41–14,720 mg C m−2 day−1), as did Ellis et al. (2012) with a range from 830 to 13,170 mg C m−2 day−1. Abril et al. (2014) conducted eight 800-km cruises along the main channel of the Solimões-Amazon River and portions of its major tributaries in the central basin and measured pCO2 every minute while underway. Values of pCO2 were similar to previous studies and varied from approximately 1000–10,000 ppmv, except for those in the Tapajós River which were lower.
Raymond et al. (2013) included regional information in their global estimates of carbon dioxide emissions from streams and rivers. Carbon dioxide concentrations in streams and rivers of whole Amazon basin (including the Andes) averaged about 6890 μatm and their calculated efflux, using a k(600) of 29 cm h−1, averaged 19,100 mg C m−2 day−1, expressed in relation to surface area of rivers and streams, not in relation to land area, as given in Raymond et al. (2013). The k(600) value used is at the high end of those reported by others for the rivers of the lowland Amazon as it includes streams and rivers in highland portions of the basin. Hence, the areal efflux is probably too high as a basin-wide average.
Sawakuchi et al. (2014) reported measurements of methane concentrations and fluxes from the mainstem Solimões-Amazon River and five tributaries (Negro, Madeira, Tapajós, Xingu, and Para) based on floating chambers. Sixteen of the 34 sites were sampled during low and high water. Dissolved, near-surface methane concentrations ranged from 0.02 to 0.5 μM. Overall, average riverine flux was 16.8 kg C km−2 day−1.
4.2 Lakes
In Lago Grande de Curuai (Fig. 7.1), a floodplain composed of interconnected lakes with a flooded area ranging seasonally from 850 to 2274 km2, Rudorff et al. (2011) noted gradients in CO2 concentration with higher concentrations near littoral regions with floating macrophytes than farther off shore as well as seasonal differences in concentrations. Polsenaere et al. (2013) used an equilibrator connected to an infrared gas analyser to measure pCO2 on transects in Lake Canaçari (Fig. 7.1), 450 km2 in area, during a 4-day measurement period. Incorporation of extensive spatial sampling in these two studies permitted recognition of spatial patterns not possible from other studies having far fewer samples. Based on transects of pCO2 and eddy covariance-based k values, Polsenaere et al. (2013) calculated mean evasion of 612 kg C km−2 day−1 during a 4-day low water period. Rudorff et al. (2011) estimated mean fluxes of CO2 from open water in L. Curuai of 2930, 4180, 4450, and 4370 kg C km−2 day−1 during receding, low, rising, and high water levels, respectively. Abril et al. (2014) include five large lakes as part of their continuous transects of pCO2 in the central Amazon and reported variations from approximately 20–20,000 ppmv. They found that the carbon dioxide efflux increased as the percentage of floating, emergent aquatic vegetation in the lakes increased.
Raymond et al. (2013) calculated carbon dioxide concentrations in lakes of whole Amazon basin (including the Andes) as averaging about 1906 μatm and an efflux, using a k(600) of 5.8 cm h−1, of 1230 kg C km−2 day−1, expressed in relation to surface area of rivers and streams, not in relation to land area, as given in Raymond et al. (2013).
4.3 Wetlands
The Negro River basin includes extensive flooded forests (locally called igapó) and large areas of interfluvial wetlands (Hess et al. 2003). Based on multi-temporal synthetic aperture radar data and field measurements with floating chambers of methane flux in the Jau River basin (Fig. 7.1) made by Rosenqvist et al. (2002), mean annual emission of methane from igapó was 23 Mg C km−2 year−1. Upper Negro interfluvial wetlands are a mosaic of emergent grasses, sedges, shrubs, and palms with shallow permanent water or seasonal flooding. Belger et al. (2011) measured methane uptake on unflooded lands, evasion from flooded areas as diffusive and ebullitive fluxes with chambers and funnels, and as transport through rooted plants. Carbon dioxide fluxes were calculated from measurements of CO2 concentrations in air and water and a k value of 2.7 cm h−1. Based on annual emission from two interfluvial wetlands representative of the region (Fig. 7.1), Belger et al. (2011) estimated average areal emission from wetland areas as 770 Mg C km−2 year−1 for CO2 and 21 Mg C km−2 year−1 for CH4.
Large savanna floodplains occur in the Llanos de Moxos (Bolivia) and in Roraima (Brazil) (Fig. 7.1) (Hamilton et al. 2002; Ferreira et al. 2007). Based on measurements in similar systems elsewhere, Melack et al. (2004) approximated mean annual methane emission from these two areas as 70 Mg C km−2 year−1. Jati (2013) made monthly measurements of carbon dioxide and methane flux with floating chambers in 80 wetlands near Boa Vista (Roraima); mean values from his results were about 9670 kg C km−2 day−1 and 9.6 kg C km−2 day−1, for CO2 and CH4, respectively. These are quite high CO2 fluxes and rather low CH4 fluxes compared to other Amazonian habitats. Emissions from cultivated rice in Roraima are not available.
4.4 Reservoirs
Five hydroelectric reservoirs (Tucuruí, Balbina, Samuel, Curuá-Una, Serra da Mesa), covering about 6300 km2, currently exist in the lowland Amazon in Brazil. All were constructed decades ago and continue to release both carbon dioxide and methane from their surfaces and through their turbines and to enhance releases in downstream rivers. Only Balbina has data collected from multiple upstream and downstream stations over a full year as well as measurements of fluxes associated with turbines (Kemenes et al. 2007, 2011). Though not in the Amazon basin, multiyear studies at Petit Saut (French Guiana), located in tropical forest, include measurements in the reservoir and downstream (Abril et al. 2005). Data scattered through the years at other Amazonian reservoirs are also available (see citations in Melack et al. 2004; Guérin et al. 2006). Kemenes et al. (2016) report carbon dioxide and methane evasion via degassing through turbines and downstream for Tucuruí, Samuel, and Curuá-Una reservoirs. Barros et al. (2011) summarised much of the data from Amazonian reservoirs, though methane emission from Balbina is listed as 10 mg C m−2 day−1 rather than 47 mg C m−2 day−1, as reported in Kemenes et al. (2007). Moreover, degassing through turbines and downstream is not included for Balbina or other Amazonian reservoirs.
Carbon dioxide emissions from the surface of Balbina and Petit Saut reservoirs averaged 1296 Mg C km−2 year−1 and 473 Mg C km−2 year−1, respectively. In the case of CO2 total annual emission from the Balbina hydroelectric system, including the reservoir, turbine outflow, and river channel extending 30 km downstream, was 1340 Mg C km−2 year−1, when expressed relative to the average reservoir area. Average CH4 emissions from the reservoir surface were 18 Mg C km−2 year−1, and total emissions were 39 Mg C km−2 year−1, indicating the importance of degassing of methane through the turbines and downstream. Though Barros et al. (2011) present mean fluxes from Amazonian reservoirs for CO2 of 400 Mg C km−2 year−1 and for CH4 of 50 Mg C km−2 year−1, these values should be used with caution since the data from which they were obtained are based on only a few samples from most of the reservoirs without adequate seasonal sampling and without data on downstream fluxes.
4.5 Airborne Surveys
Airborne campaigns have provided integrated coverage of subregions of the Amazon basin and permitted calculation of carbon balance (Lloyd et al. 2007) and methane emission (Beck et al. 2012; Miller et al. 2007). Miller et al. (2007) collected vertical profiles of methane over 4 years at sites near Santarém and Manaus and calculated average emissions of 20 kg C km−2 day−1. Wetlands are likely the major source of methane, at least during seasons with extensive inundated areas. Other sources include fires, urban areas, termites, and, perhaps emissions associated with terra firme forests. Beck et al. (2012) described results of airborne campaigns in November and May, periods representing generally low and large inundation, during which continuous, in-flight measurements of CH4 and sampling for isotopic analyses were conducted. The flights extended over much of the lowland Amazon in Brazil and were concentrated in the central basin. Isotopic measurements indicated that biogenic methane predominates, and wetlands are likely the major source though near Manaus anthropogenic sources, such as waste decomposition, contribute. A signature of biomass burning was detected in samples collected during the dry season, but this source appeared to be minor for CH4. Beck et al. (2012) estimated a CH4 flux for the lowland Amazon during November as 27 ± 9 kg C km−2 day−1 and during May as 32 ± 14 kg C km−2 day−1.
5 Regionalisation of Fluxes
5.1 Prior Estimates
Though various estimates of regional carbon dioxide and methane fluxes have been made through the years, only recent estimates that used data available through the beginning of the twenty-first century are summarised here. Richey et al. (2002) and Melack et al. (2004) were the first to use regional analyses of microwave remote sensing data to establish inundated areas and habitats. Both applied Monte Carlo error propagation to establish uncertainties.
Richey et al. (2002) used carbon dioxide measurements primarily from the Solimões-Amazon River, its fringing floodplain and mouths of major tributaries, and conservatively low piston velocities to calculate an outgassing rate of 830 ± 240 Mg C km−2 year−1 for the annual mean flooded area. The areal flux was combined with remote sensing-derived inundated areas at high and low stages for rivers and floodplains over 100 m across (Hess et al. 2003) to determine total outgassing in a 1.77 million km2 quadrant in the central basin. The seasonal variation in inundated area was assumed to track river stage, as measured by the sparse network of gauges. A comparison of inundated area-derived passive microwave data versus stage for large floodplains supports this assumption (Hamilton et al. 2002). Areas of moderate to small rivers and streams in this quadrant were approximated as a geometric series relating stream length and width to stream order. To extrapolate to the whole Amazon basin (6.07 million km2), an areal flux of half that used for the central quadrant was applied to the 4.3 million km2 outside the central quadrant to yield a value of 470 Tg C year−1 and that applied to the area of the lowland basin yields an evasion of 390 Tg C year−1.
Rasera et al. (2013) extrapolated their results to a central Amazon quadrant (1.47 million km2). To do so, they combined (1) areas of streams (<100 m wide) and the areas of rivers and floodplains (>100 m wide) for high and low stage as reported in Richey et al. (2002); (2) k(600) values based on recent work for rivers and streams; and (3) pCO2 values from measurements and from a relationship between measured values and average soil cation exchange capacity. If the annual total evasion of CO2 calculated in this manner were increased in proportion to the slightly larger central basin area used by Richey et al. (2002), 432 ± 78 Tg C year−1 would result. This annual rate is about twice that reported by Richey et al. (2002) and reflects higher k(600) values and improved data for streams. It is important to recognise that the fluxes calculated for the mainstem Amazon and tributaries in Rasera et al. (2013) include floodplain areas, not just river channels; if extrapolated to the area of the lowland basin a flux of 1240 ± 206 Tg C year−1 results.
Regional extrapolation of fluxes of carbon dioxide from streams and small rivers is especially difficult because of the few measurements among the millions of kilometres of these systems and the large spatial and temporal variations observed. Johnson et al. (2008) approximated potential evasion of CO2 from headwater streams basin-wide (an area of 6.07 million km2, which included the Andean highlands) as 114 Tg C year−1 or 19.5 Mg C km−2 year−1, excluding inundated areas and accounting for human modified land uses, and where the areal rate is expressed per km2 of total land area, not stream areas. Variations in annual water balances for the period from 1976 to 1996 would introduce about a 10 % increase or decrease between wet or dry years. Their regionalisation approach is based on groundwater fluxes, determined as the difference between average annual precipitation and evapotranspiration, and estimates of soil pCO2 from carbonate equilibrium reactions at a spatial scale of 0.1°. Though an interesting approach, it requires validation based on actual measurements for a variety of headwaters, such as Andean, blackwater, or savanna, streams that are different from those examined by Johnson et al. (2006). Rasera et al. (2008) extrapolated from the Ji-Paraná River basin to the Amazon basin and arrived at a value substantially higher than can be calculated from the data presented. Based on the total CO2 evasion from the Ji-Paraná River basin divided by the area of the basin and multiplied by the area of third- to sixth-order rivers (derived from the fractional area of these rivers in the Ji-Paraná River basin; see above), the annual evasion is about 21 Tg C for these rivers in the lowland Amazon basin.
Melack et al. (2004) used available measurements of habitat-specific methane fluxes; all the data were from the central Amazon basin and based mainly on methane captured in floating chambers. By combining these values with remote sensing-based estimates of areal extent of aquatic habitats (open water in lakes, rivers, flooded forests, and aquatic macrophytes), they calculated regional estimates for the mainstem Solimões-Amazon in Brazil and for the same 1.77 million km2 quadrant used by Richey et al. (2002). Seasonal changes in the areal extent of the aquatic habitats were approximated by interpolating between remote sensing-derived areas obtained for low and high water periods. A time series of inundation extent along the mainstem Solimões-Amazon floodplain, based on passive microwave data, was used to calculate inter-annual variations in methane flux for 3 years. To extrapolate to the whole lowland basin (5.05 million km2), a single, habitat-averaged value of 30 Mg C km−2 year−1, calculated from the mean annual emission estimated for the mainstem Solimões-Amazon in Brazil and the mean annual flooded area of this reach, was used, resulting in a flux of c. 22 Tg C year−1. If expressed as the greenhouse gas warming potential equivalence of CO2, this mean flux amounts to about 0.2 Pg C year−1 (not 0.5 Pg C year−1, as noted in the original paper).
Sawakuchi et al. (2014) subtracted estimates of Landsat-based lake areas from the water body category in an AVHRR 1-km regional land cover product (Brown et al. 2003) to obtain a large river channel area of approximately 91,000 km2, a value considerably larger than others discussed earlier. When combined with their methane fluxes, they calculated an annual average flux of 0.37 Tg C as methane from the large rivers of the Amazon basin. If the areal estimate of Beighley and Gummadi (2011) is used, the flux is 0.12 Tg C year−1, and if the SRTM areal estimate is used, the flux is 0.21 Tg C year−1. These methane fluxes represent 0.5–1.6 % of the basin-wide emission calculated by Melack et al. (2004) and indicate a minor role for the large river channels. Methane emission from the mainstem Solimões-Amazon River in Brazil was estimated by Melack et al. (2004) to represent only 0.06 % of the total flux for that reach including the fringing floodplains.
Barros et al. (2011) estimated emission from extant tropical Amazonian reservoirs, based on an area of 20,000 km2, of 8 Tg C year−1 as CO2 and 1.0 Tg C year−1 as CH4. We question these values based on the area of Amazonian reservoirs being 6300 km2 and based on areal emission estimates available from Balbina and Petit Saut, as described above.
Global or continental scale calculations derived from satellite retrievals of atmospheric concentrations and inverse modelling provide coarse spatial resolution methane emission values for the Amazon basin (e.g. Frankenberg et al. 2008). Beck et al. (2012) examined the performance of CH4 inversion models, constrained by observations from surface stations and SCIAMACHY retrievals, in comparison to their airborne campaigns for the Amazon basin. In this comparison, the global models used the same transport model but different prior CH4 inputs from wetlands, none of which were well suited to the Amazon basin. One result derived from the transport modelling suggested that the Amazon basin is influenced by an atmospheric region larger than the basin. Furthermore, Beck et al. (2012) concluded that a reliable annual methane budget will require regional airborne campaigns over a full year.
5.2 New Estimates
In principle, total basin-wide fluxes of CO2 and CH4 (F, in units of Tg C year−1) could be calculated using a general expression similar to that in Melack et al. (2004):
where F is the flux of CO2 or CH4 for each habitat (expressed as kg C km−2 day−1); j is each habitat: (1) headwaters, (2) streams and moderate-sized rivers, (3) large rivers, (4) floodplains, (5) wetlands, and (6) hydroelectric reservoirs; A is an estimate of average flooded area of each habitat per month; t is the number of days per month; and i is each month incremented from 1 to 12. Depending on available data, several habitat categories could be subdivided: e.g. large rivers (e.g. Solimões-Amazon and white water tributaries, Negro and black water tributaries, Tapajós, and other clear waters), floodplains (e.g. open water lakes, flooded forests, aquatic macrophytes), and wetlands (e.g. upper Negro interfluvial, Roraima savanna, Llanos de Moxos). If data from multiple years were available, such as the inundation time series of Papa et al. (2010) or Paiva et al. (2013), or calculated from an empirical or mechanistic model of carbon dioxide and methane dynamics, Eq. (7.1) could be evaluated repeatedly to determine inter-annual variability.
Several challenges make it difficult to apply Eq. (7.1). In particular, sufficient information about the spatial and temporal variations of inundated areas, habitat characteristics, and associated fluxes on a basin-wide scale for multiple years is lacking. A modelling system that combines climatic and hydrological processes with biogeochemical and ecological processes is required.
As an alternative, a combination of the published values, summarised in prior sections, for specific regions and calculations based on averaged measurements of carbon dioxide from (1) moderate-sized rivers, (2) large rivers, (3) floodplains and wetlands, and (4) hydroelectric reservoirs as noted in Table 7.1 are used to estimate carbon dioxide emissions. These fluxes include those measured with floating chambers and those calculated with new estimates of gas exchange velocities and in situ gas concentrations. Remote sensing-based estimates of inundated areas at low and high water levels and modelled variations in inundation are used to approximate annual values for floodplain and wetland environments. River channel areal estimates from Beighley and Gummadi (2011) and SRTM are used. The estimates are standardised to the lowland basin below 500 m as delineated from JERS-1 synthetic aperture radar data (5.05 million km2). Insufficient new measurements for methane are available to improve upon Melack et al. (2004).
Carbon dioxide evasion from rivers and streams is estimated to be c. 200 Tg C year−1. Though fluxes per unit area for rivers and streams are based on updated k(600) and recent data from a range of stream and river sizes, a smaller surface area for these habitats than previous basin-wide estimates is being used. The annual flux from the category called floodplain and wetland habitats, which includes lakes, plus those from other wetlands and reservoirs is estimated to be approximately 1600 Tg C year−1. These fluxes incorporate updated k(600) values; however, the lack of information about k values in vegetated areas is a concern because Melack and Hess (2010) estimate that about 79 % of the lowland basin is characterised by woody vegetation with another 13 % predominately herbaceous vegetation. Since k values are likely to be less in these areas in comparison to those in lakes, the annual fluxes would be lower. Furthermore, basin-wide modelled inundation fractions and remote sensing-based areas do not represent well the seasonality in areas or fluxes. With these and other issues noted above acknowledged, total evasion of carbon dioxide for the lowland Amazon basin is estimated to be c. 1.8 Pg C year−1. Though a formal analysis of uncertainty cannot be done based on the heterogeneous information used for these estimates, based on spatial and temporal variability and uncertainty in measurements, an overall uncertainty of at least ±50 % is reasonable.
Raymond et al. (2013) calculated global annual evasion of carbon dioxide from streams and rivers ranging from 1.5 to 2.1 Pg C and evasion from lakes and reservoirs ranging from 0.06 to 0.84 Pg C. The ranges represent 5th and 95th confidence intervals derived from a Monte Carlo analysis. Wetlands were not included. That the value reported here for the lowland Amazon basin is similar to the sum of these global estimates indicates the importance of including tropical floodplains and other wetlands in calculations of carbon dioxide evasion from inland waters.
6 Sources and Decomposition of Organic Carbon
Floodplains and other wetlands are productive aquatic environments in which most of the production and evasion of CO2 and CH4 is likely derived from metabolic processing of the carbon fixed by aquatic plants. These environments also export considerable amounts of carbon to rivers and accumulate sediments. Estimates of carbon balances for floodplains at several spatial scales provide supporting evidence: Calado (Melack and Engle 2009), Curuai (Rudorff 2013), the mainstem Solimões-Amazon floodplain in Brazil (Melack and Forsberg 2001), and a 1.77 million-km2 quadrant in the central Amazon (Melack et al. 2009; Abril et al. 2014). In particular, Abril et al. (2014) suggest that Amazonian wetlands export about half of their primary productivity to neighbouring waters where it is metabolised and much is released to the atmosphere. Further evidence is provided by estimates of root respiration by herbaceous and woody aquatic plants (Hamilton et al. 1995; Worbes 1997), isotopic studies of microbial respiration (Waichman 1996), calculation of aquatic macrophyte growth and decay (Engle et al. 2008; Silva et al. 2009, 2013), rates of methane oxidation in exposed wetland sediments (Koschorreck 2000), and the enrichment of δ13C of CO2 in lowland rivers, expected if C4 grasses are significant sources of respired carbon (Mayorga et al. 2005b).
Understanding the relevance of evasion of carbon dioxide from aquatic environments to the carbon balance of terra firme forests requires (1) determination of the proportion of the carbon fixed within aquatic ecosystems versus that imported from uplands as inorganic or organic carbon and (2) measurements of aquatic respiration and of decomposition of these sources of organic carbon. Though further work is needed, recent results indicate that the carbon inputs to aquatic systems that are eventually emitted as CO2 or CH4 vary among habitats and are related to hydrological conditions and proximity of carbon sources.
Headwater streams in the Juruena watershed (Fig. 7.1), and presumably elsewhere, receive most of the CO2 degassed in their uppermost reaches directly from terrestrially derived respiration in soils with small contributions of organic C from riparian and upland litter that is gradually processed downstream (Johnson et al. 2006, 2007). In slightly larger streams and small rivers, relatively labile dissolved and particulate organic carbon and lateral inputs of dissolved CO2 support evasion (earlier work summarised in Richey et al. 2009; Davidson et al. 2010). There is regional heterogeneity in the carbon sources, with 13C-depleted CO2 in streams draining sandy soils in the Negro basin indicative of C3 plants, while the 13C-enriched CO2 in streams passing through pastures in Rondônia indicating C4 grasses. Based on direct assays of 14CO2 outgassed from small streams and rivers in the western Amazon, Vihermaa et al. (2014) report that a portion of the carbon dioxide is derived from sedimentary rock and carbonate weathering.
Carbon dioxide evasion from large rivers appears to be supported by a wide variety of carbon compounds from a combination of nearby and distant organic carbon sources. Especially relevant results are reported by Ellis et al. (2012), who determined the δ13C of the CO2 evaded from the Amazonian rivers and found that organic carbon from C3 and C4 plants and phytoplankton was evident and spatially and temporally variable. Another valuable result from Ward et al. (2013) demonstrated that the degradation of lignin and associated macromolecules in water from the lower Amazon River was sufficient to support considerable respiration and presumably CO2 evasion. Since these compounds have been thought to be refractory and are often of terrestrial origin, this finding supports the notion that metabolism in the large rivers is supported by diverse carbon sources. Fatty acid and stable isotope analyses by Mortillaro et al. (2011, 2012) as well as studies by Kim et al. (2012), using the branched and isoprenoid tetraether index, offer further evidence of multiple carbon inputs to the lower Amazon River. Oxidation of petrogenic organic carbon has also been documented (Bouchez et al. 2010). Results by Mayorga et al. (2005b) indicated that the main source of respired carbon was <5 years old and that the dissolved CO2 was isotopically different from organic carbon in the rivers sampled at the same time. These results imply that inputs of labile carbon, which is rapidly oxidised, support the generation of the high pCO2 values observed. However, Ellis et al. (2012) found that the respired carbon was isotopically similar to that in the water. Further, as noted by Rasera et al. (2013), the high concentrations of CO2 during high river stages may reflect export of labile organic carbon from fringing floodplains. In the Amazon, Madeira and Solimões rivers Ellis et al. (2012) found that riverine respiration could account for most or all of the CO2 evaded, while in the Negro River it could account for only 15–34 %. Photo-oxidation of organic carbon appears to make small contributions to CO2 in large rivers (Amaral et al. 2013; Remington et al. 2011).
In summary, the relative importance of carbon sources originating in terra firme forests versus aquatic habitats varies as a function of position in the continuum from headwaters to large rivers with fringing floodplains or associated wetlands. Hence, a general conclusion regarding the proportion of terra firme net productivity that is emitted to the atmosphere after lateral transport to aquatic environments is difficult to state. However, it is clear that CO2 evasion, in particular, is supported by carbon fixed by terra firme plants in many streams and that this carbon contributes to metabolic processes in the largest rivers. In contrast, floodplains and wetlands likely represent environments where the CO2 and CH4 emitted to the atmosphere are derived largely from carbon fixed by aquatic plants and, to a lesser extent, algae, and that a portion of organic carbon metabolised in rivers is supplied by their floodplains. Based on this logic, the values in Table 7.1 lead to the conclusion that the majority of the basin-wide evasion of carbon dioxide is derived from plants in aquatic environments.
7 Uncertainties and Research Needs
7.1 Field Measurements
The largest uncertainties stem from the sparseness of measurements in time and space. For example, based on a Monte Carlo error analysis, Melack et al. (2004) noted that the uncertainty in actual methane fluxes, largely because of the high spatial and temporal variability, compounded by the episodic nature of ebullition, was larger than the uncertainty associated with remote sensing-based habitat extent. This result applied to the best sampled floodplains of the central basin. In many wetlands of the basin, few or no data are available. Particularly large gaps with no data exist in the Llanos de Moxos, Bananal, peatlands in the western Amazon (Lahteenoja et al. 2011), Peruvian lowlands, and habitats above 500 m. The extensive network of streams and medium-sized rivers is significantly under-sampled, and Richey et al. (2009) made a plea for collecting many spatially distributed measurements given the large variability observed among the data gathered. While reservoirs are receiving increased attention, time series data and measurements above and below the dams are required to better guide the management of greenhouse gas evasion from hydroelectric projects.
Within lakes, reservoirs, and wetlands, diel and seasonal variations in vertical stratification and horizontal advection influence the concentration gradients of gases and the transfer velocities. With the increasing availability of in situ sensors and automatic recording systems, it is possible to incorporate temporal and spatial variations in physical and chemical conditions in calculations of gas exchanges. For example, deep convective mixing at night, common in tropical waters, is likely to increase both transfer velocities and concentrations of carbon dioxide and methane in surficial waters, thus increasing fluxes.
Other components of the carbon balance in aquatic habitats can be challenging to measure, and data are lacking for many areas or are without sufficient spatial and temporal coverage even at better studied sites. Key processes that need attention are primary productivity by algae and higher plants, sedimentation in floodplains and other wetlands, transport of dissolved and particulate inorganic and organic carbon from uplands to inland waters, and rates of respiration and decomposition.
7.2 Modelling
While models of river discharge and inundation are improving, the current basin-scale models do not properly flood some important and large habitats, such as the largely rain-fed interfluvial wetlands in the upper Negro basin, the Roraima wetlands, or the Llanos de Moxos. One key to improvement is higher spatial resolution DEMs for floodplains. Biogeochemical models of carbon dioxide and methane production and evasion appropriate for conditions in the Amazon are not available (Riley et al. 2011), though relevant models are under development (Potter et al. 2016). While several models have potentially useful components or formulations (Ringeval et al. 2010; Bloom et al. 2010; Cao et al. 1996; Potter 1997; Walter and Heimann 2000), no spatially explicit model exists that incorporates the inundation dynamics and biogeochemical processes of aquatic environments in the Amazon.
8 Climate Change, Exceptional Events, and Human Impacts
Climate warming, climate variability, including exceptional droughts and floods and severe wind storms, and fires are influencing the Amazon basin (Davidson et al. 2012). Additionally, human alterations include agricultural expansion with associated deforestation, construction of dams, and fossil fuel exploration (Melack 2005; Costa et al. 2009; Pires and Costa 2013; Renó et al. 2011; Castello et al. 2013; Finer and Orta-Martinez 2010). A review of current climatic conditions in the context of aquatic environments in the Amazon basin by Melack and Coe (2013) noted the paucity of meteorological records in floodplains and other wetlands. Basin-wide, long-term warming trends in air temperature are generally 0.1–0.3 °C per decade (1960–2009; Burrows et al. 2011). Rainfall has negative trends in the northern Amazon and positive trends in the southern Amazon (1949–1999; Marengo 2004). Severe droughts occurred in 2005 and 2010 and were associated with tropical Atlantic warming and ENSO events (Marengo et al. 2008, 2009, 2011; Zeng et al. 2008; Villar et al. 2011). In contrast, an exceptional flood occurred in 2009 (Chen et al. 2010). Furthermore, increased variability in climatic conditions with increased frequency and severity of droughts and storms is projected by global models (Malhi et al. 2008; Gloor et al. 2013; Huntingford et al. 2013; Lau et al. 2013; Liu et al. 2013; Reichstein et al. 2013).
Another consequence of severe weather can be substantial disturbance of forests as trees are blown down (Negrón-Juárez et al. 2010; Chambers et al. 2007). Droughts can lead to increased incidence of fires (Aragão et al. 2008; Longo et al. 2009), even in igapó forests (Flores et al. 2014).
Melack and Coe (2013) ran simulations of inundation under altered climate and land uses specifically for the Amazon basin using a basin-wide hydrological model forced with observed climate data from 1950 to 2000. Simulations of inundation with 10 and 25 % decreases in rainfall (without seasonal and spatial variations included) resulted in reductions in inundation similar to reductions in rainfall: −5 to −20 % (10 %) and −12 to −30 % (25 %). Based on 35 % deforestation coupled to a global climate model, rainfall decreased and evapotranspiration decreased more; hence average maximum flooded area along mainstem Amazon increased slightly compared to current land cover. Others have considered actual or potential climate change or land use impacts on hydrological conditions in the Amazon basin (Coe et al. 2009, 2011; Casimiro et al. 2011, 2012; Langerwisch et al. 2012; Cox et al. 2013). In addition to the existing hydroelectric reservoirs, others are under construction (Belo Monte on the Xingu River; Santo Antonio and Jirau, run-of-river dams on the Madeira River), and many more are planned throughout the Amazon basin including the Tapajós hydroelectric complex which could inundate about 2000 km2 (Finer and Jenkins 2012). To estimate the emissions of CO2 and CH4 from these new and planned reservoirs is difficult because of differences in construction and operation and because economic and environmental issues will likely play roles. Furthermore, road construction and agriculture create numerous small impoundments (Macedo et al. 2013), of unknown total area with unmeasured fluxes.
9 Conclusion
The updated value for annual carbon dioxide emission from aquatic habitats in the lowland Amazon basin of 1800 Tg C is larger than previous estimates. Almost 90 % of this flux is likely associated with lakes, floodplains, and other wetlands. Carbon fixed by terra firme plants contributes most of the carbon dioxide emitted from streams and adds organic carbon to rivers. Lakes, floodplains, and other wetlands represent environments where the CO2 emitted to the atmosphere is derived largely from carbon fixed by aquatic plants with lesser contributions from algae, and a portion of organic carbon metabolised in rivers is supplied by their floodplains.
To further improve the updated estimates of carbon dioxide and methane evasion from aquatic habitats in the Amazon basin requires several activities. The largest uncertainties stem from the sparseness of measurements in time and space. Deployment of eddy covariance systems, continuous measurements along transects, and regional airborne campaigns would help considerably. Inclusion of habitats not well characterised, such as flooded forests, savannas, intermittently flooded regions along streams, and depressions within terra firme forests is needed. Hydroacoustic techniques and sampling releases via turbines in hydroelectric dams will improve estimation of ebullition. Seasonal and inter-annual variability of inundated areas derived from remote sensing and modelling should be incorporated. The lack of high-resolution digital elevation data are a serious limitation throughout the basin. Given the pronounced diel cycle of heating and cooling often observed in shallow tropical waters, it is important to measure stratification and mixing, and gas concentrations and exchanges, over these diel cycles. Biogeochemical models of carbon dioxide and methane production and evasion appropriate for conditions in the Amazon require further development. Climate warming, climate variability, including exceptional droughts and floods and severe wind storms, fires, agricultural expansion, and construction of dams are all influencing the Amazon basin with consequences for the role of aquatic environments in the carbon cycle and release of carbon dioxide and methane.
References
Abril G, Guérin F, Richard S, Delmas R, Galy-Lacaux C, Gosse P, Tremblay A, Varfalvy L, Santos MA, Matvienko B (2005) Carbon dioxide and methane emissions and the carbon budget of a 10-year old tropical reservoir (Petit Saut, French Guiana). Global Biogeochem Cycles 19:1–16
Abril G, Martinez J-M, Artigas LF, Moreira-Turcq P, Benedetti MF, Vidal L, Meziane T, Kim J-H, Bernardes MC, Savoye N, Deborde J, Souza EL, Albéric P, de Souza MFL, Roland F (2014) Amazon River carbon outgassing fuelled by wetlands. Nature 505:395–398
Aires F, Papa F, Prigent C (2013) A long-term high-resolution wetland dataset over the Amazon basin, downscaled from multiwavelength retrieval using SAR data. J Hydrometeorol 14:594–607
Alin SR, Rasera MFFL, Salimon CI, Richey JE, Holtgrieve GW, Krusche AV, Snidvongs A (2011) Physical controls on carbon dioxide transfer velocity and flux in low-gradient river systems and implications for regional carbon budgets. J Geophys Res Biogeosci 116:G01009. doi:10.1029/2010jg001398
Alsdorf D, Han S-C, Bates P, Melack J (2010) Seasonal water storage on the Amazon floodplain measured from satellites. Remote Sens Environ 114:2448–2456
Amaral JHF, Suhett AL, Melo S, Farjalla VF (2013) Seasonal variation and interaction of photodegradation and microbial metabolism of DOC in black water Amazonian ecosystems. Aquat Microb Ecol 70:157–168
Aragão LEOC, Malhi Y, Barbier N, Lima A, Shimabukuro Y, Anderson L, Saatchi S (2008) Interactions between rainfall, deforestation and fires during recent years in the Brazilian Amazonia. Philos Trans R Soc B 363:1779–1785
Arnesen AS, Silva TSF, Hess LL, Novo EMLM, Rudorff CM, Chapman BD, McDonald KC (2013) Monitoring flood extent in the lower Amazon River floodplain using ALOS/PALSAR ScanSAR images. Remote Sens Environ 130:51–61. doi:10.1016/j.rse.2012.10.035
Aufdenkampe AK, Mayorga E, Raymond PA, Melack JM, Doney SC, Alin SR, Aalto RE, Yoo K (2011) Rivers key to coupling biogeochemical cycles between land, oceans and atmosphere. Front Ecol Environ 9:53–60
Banerjee S, MacIntyre S (2004) The air-water interface: turbulence and scalar exchange. In: Grue J (ed) Advances in coastal and ocean engineering, vol 9. World Scientific, Hackensack, NJ, pp 181–237
Barbosa PM (2013) Avaliação do fluxo difusivo de metano (CH4) em ambientes do médio-baixo Solimões. M.Sc. Thesis, Universidade Federal do Rio de Janeiro
Barros N, Cole JJ, Tranvik LJ, Prairie YT, Bastiviken D, Huszar VLM, del Giorgi P, Roland F (2011) Carbon emission from hydroelectric reservoirs linked to reservoir age and latitude. Nat Geosci 4:593–596
Battin TJ, Luyssaert S, Kaplan LA, Aufdenkampe AK, Richter A, Tranvik LJ (2009) The boundless carbon cycle. Nat Geosci 2:598–600
Beck V, Chen H, Gerbig C, Bergamaschi P, Bruhwiler L, Houweling S et al (2012) Methane airborne measurements and comparison to global models during BARCA. J Geophys Res 117:D15310. doi:10.1029/2011JD017345
Beighley RE, Gummadi V (2011) Developing channel and floodplain dimensions with limited data: a case study in the Amazon basin. Earth Surf Process Landf 36:1059–1071
Beighley RE, Eggert KG, Dunne T, He Y, Gummadi V, Verdin KL (2009) Simulating hydrologic and hydraulic processes throughout the Amazon River Basin. Hydrol Process 23:1221–1235. doi:10.1002/hyp.7252
Belger L, Forsberg B, Melack JM (2011) Factors influencing carbon dioxide and methane emissions from interfluvial wetlands of the upper Negro River basin, Brazil. Biogeochemistry 105:171–183. doi:10.1007/s10533-010-9536-0
Bloom AA, Palmer PI, Fraser A, Reay DS, Frankenberg C (2010) Large-scale controls on methanogenesis inferred from methane and gravity spaceborne data. Science 327:322–325
Borges AV, Abril G, Darchambeau F, Teodoru CR, others (2015) Divergent biophysical controls of aquatic CO2 and CH4 in the world’s two largest rivers. Sci Rep. doi:10.1038/srep15614
Bouchez J, Beyssac O, Galy V, Gaillardet J, France-Lanord C, Laurence M, Moreira-Turcq PF (2010) Oxidation of petrogenic organic carbon in the Amazon floodplain as a source of atmospheric CO2. Geology 38:255–258
Brix H, Sorrell BK, Orr PT (1992) Internal pressurization and convective gas flow in some emergent freshwater macrophytes. Limnol Oceanogr 37:1420–1433
Brown J, Loveland T, Ohlen D, Zhu Z (2003) LBA regional land cover from AVHRR 1-km version 1.2 (IGBP). Oak Ridge National Laboratory Distributed Active Archive Center, Oak Ridge, TN
Burrows MT, Schoeman DS, Buckley LB, Moore P, Poloczanska ES, Brander KM et al (2011) The pace of shifting climate in marine and terrestrial ecosystems. Science 334:652–655
Butman D, Raymond PA (2011) Significant efflux of carbon dioxide from streams and rivers in the United States. Nat Geosci 4:839–842
Cao M, Marshall S, Gregson K (1996) Global carbon exchange and methane emission from natural wetlands: application of a process-based model. J Geophys Res 101:14399–14414
Casimiro L, Waldo S, Labat D, Guyot JL, Ardoin-Bardin S (2011) Assessment of climate change impacts on the hydrology of the Peruvian Amazon-Andes basin. Hydrol Process 25:3721–3734
Casimiro L, Waldo S, Labat D, Guyot JL, Ronchail J, Espinoza Villar JC (2012) Trends in rainfall rates and temperature fluctuations in the Peruvian Amazonas-Andes basin over the last 40 years. Hydrol Process. doi:10.1002/hyp.9418
Castello L, McGrath DG, Hess LL, Coe MT, Lefebvre PA, Petry P, Macedo MN, Renó VF, Arantes CC (2013) The vulnerability of Amazon freshwater ecosystems. Conserv Lett 6:217–229
Chambers JQ, Fisher JI, Zeng H, Chapman EL, Baker DB, Hurtt GC (2007) Hurricane Katrina’s carbon footprint on U.S. Gulf Coast forests. Science 318:1107
Chen JL, Wilson CR, Tapley BD (2010) The 2009 exceptional Amazon flood and interannual terrestrial water storage change observed by GRACE. Water Resour Res 46. doi:10.1029/2010WR009383
Coe MT, Costa MH, Howard E (2007) Simulating the surface waters of the Amazon River basin: impacts of new river geomorphic and dynamic flow parameterizations. Hydrol Process 22:2542–2553
Coe MT, Costa MH, Soares-Filho BS (2009) The influence of historical and potential future deforestation on the stream flow of the Amazon River—land surface processes and atmospheric feedback. J Hydrol. doi:10.1016/j.jhydrol.2009.02.043
Coe MT, Latrubesse EM, Ferreira ME, Amsler ML (2011) The effects of deforestation and climate variability on the streamflow of the Araguaia River, Brazil. Biogeochemistry 105:119–131. doi:10.1007/s10533-011-9582-2
Cole JJ, Praire YT, Caraco NF, McDowell WH, Tranvik LJ, Striegl RR, Duarte CM, Kortelainen P, Downing JA, Middleburg J, Melack JM (2007) Plumbing the global carbon cycle: integrating inland waters into the terrestrial carbon budget. Ecosystems. doi:10.1007/s10021-006-9013-8
Costa MH, Coe MT, Guyot JL (2009) Effects of climatic variability and deforestation on surface water regimes. In: Gash J, Keller M, Silva-Dias P (eds) Amazonia and global change, Geophysical monograph series 186. American Geophysical Union, Washington, DC, pp 543–553
Cox PM, Pearson D, Booth BB, Friedlingstein P, Huntingford C, Jones CD, Luke CM (2013) Sensitivity of tropical carbon to climate change constrained by carbon dioxide variability. Nature 494:341–344
Crill PM, Bartlett KB, Wilson J, Sebacher DI, Harriss RC, Melack JM, MacIntyre S, Lesack L, Smith Morrill L (1988) Tropospheric methane from an Amazon floodplain lake. J Geophys Res 93:1564–1570
Davidson EA, Figueiredo RDO, Markewitz D, Aufdenkampe AK (2010) Dissolved CO2 in small catchment streams of eastern Amazonia: a minor pathway of terrestrial carbon loss. J Geophys Res 115. doi:10.1029/2009JG001202
Davidson EA, de Araújo AC, Artaxo P, Balch JK, Brown IF, Bustamante MMC, Coe MT, DeFries RS, Keller M, Longo M, Munger JW, Schroeder W, Soares-Filho BS, Souza CM Jr, Wofsy SC (2012) The Amazon basin in transition. Nature 481:321–328
Del Sontro T, Kunz MJ, Kempter T, Wuest A, Wehrli B, Senn DB (2011) Spatial heterogeneity of methane ebullition in a large tropical reservoir. Environ Sci Technol 45:9866–9873
Devol AH, Quay PD, Richey JE, Martinelli LA (1987) The role of gas exchange in the inorganic carbon, oxygen and 222 radon budgets of the Amazon River. Limnol Oceanogr 32:235–248
Downing J (2009) Global limnology: up-scaling aquatic services and processes to planet Earth. Verh Int Verein Limnol 30:1149–1166
Downing JA, Cole JJ, Duarte CM, Middelburg J, Melack JM, Prairie YT, Kortelainen P, Striegl RG, McDowell WH, Tranvik LJ (2012) Global abundance and size distribution of streams and rivers. Inland Waters 2:229–236
Ellis EE, Richey JE, Aufdenkampe AK, Krusche AV, Quay PD, Salimon C, da Cunha HB (2012) Factors controlling water-column respiration in rivers of the central and southwestern Amazon Basin. Limnol Oceanogr 57:527–540
Engle D, Melack JM (2000) Methane emissions from the Amazon floodplain: enhanced release during episodic mixing of lakes. Biogeochemistry 51:71–90
Engle DL, Melack JM, Doyle RD, Fisher TR (2008) High rates of net primary productivity and turnover for floating grasses on the Amazon floodplain: implications for aquatic respiration and regional CO2 flux. Glob Chang Biol 14:369–381
Ferreira E, Zuanon J, Forsberg B, Goulding M, Briglia-Ferreira SR (2007) Rio Branco: ecologia e conservação de Roraima. INPA/ACCA, Lima, p 201
Finer M, Jenkins CN (2012) Proliferation of hydroelectric dams in the Andean Amazon and implications for Andes-Amazon connectivity. PLoS One 7:e35126. doi:10.1371/journal.pone.0035126
Finer M, Orta-Martinez M (2010) A second hydrocarbon boom threatens the Peruvian Amazon: trends, projections, and policy implications. Environ Res Lett 5. doi:10.1088/1748-9326/5/1/014012
Flores BM, Piedade MTF, Nelson BW (2014) Fire disturbance in Amazonian blackwater floodplain forests. Plant Ecol Divers 7:319–327
Frankenberg C, Bergamaschi P, Butz A, Houweling S, Meirink JF, Notholt J, Petersen AK, Schrijver H, Warneke T, Aben I (2008) Tropical methane emissions: a revised view from SCIAMACHY onboard ENVISAT. Geophys Res Lett 35. doi:10.1029/2008GL034300
Frappart F, Seyler F, Martinez J-M, León JG, Cazenave A (2005) Floodplain water storage in the Negro River basin estimated from microwave remote sensing of inundation and water levels. Remote Sens Environ 99:387–399
Gash J, Keller M, Silva-Dias P (eds) (2009) Amazonia and global change, Geophysical monograph series 186. American Geophysical Union, Washington, DC
Getirana ACV, Boone A, Yamazaki D, Decharme B, Papa F, Mognard N (2012) The hydrological modeling and analysis platform (HyMAP): evaluation in the Amazon Basin. J Hydrometeorol 13:1641–1665
Gloor M, Brienen RJW, Galbraith D, Feldpausch TR, Schöngart J, Guyot JL, Espinoza JC, Lloyd J, Phillips OL (2013) Intensification of the Amazon hydrological cycle over the last two decades. Geophys Res Lett 40. doi:10.1002/grl.50377
Guérin F, Abril G, Richard S, Burban B, Reynouard C, Seyler P, Delmas R (2006) Methane and carbon dioxide emissions from tropical reservoirs: significance of downstream rivers. Geophys Res Lett 33. doi:10.1029/2006GL027929
Guérin F, Abril G, Serca D, Delon C, Richard S, Delmas R, Tremblay A, Varfalvy L (2007) Gas transfer velocities of CO2 and CH4 in a tropical reservoir and its river downstream. J Mar Syst 66:161–172
Hamilton S, Sippel S, Melack JM (1995) Oxygen depletion and carbon dioxide production in waters of the Pantanal wetland of Brazil. Biogeochemistry 30:115–141
Hamilton SK, Sippel SJ, Melack JM (2002) Comparison of inundation patterns among major South American floodplains. J Geophys Res 107. doi:10.1029/2000JD000306
Hamilton SK, Sippel SJ, Melack JM (2004) Seasonal inundation patterns in two large savanna floodplains of South America: the Llanos de Moxos (Bolivia) and the Llanos del Orinoco (Venezuela and Colombia). Hydrol Process 18:2103–2116
Hanson MC, Potapov PV, Moore R, Hancher M, Turubanov SA, Tyukavina A et al (2013) High-resolution global maps of 21st century forest cover change. Science 342:850–853
Hawes JE, Peres CA, Riley LB, Hess LL (2012) Landscape-scale variation in structure and biomass of Amazonian seasonally flooded and unflooded forests. For Ecol Manage 281:163–176
Hess LL, Melack JM, Novo EMLM, Barbosa CCF, Gastil M (2003) Dual-season mapping of wetland inundation and vegetation for the central Amazon basin. Remote Sens Environ 87:404–428
Ho DT, Veron F, Harrison E, Bliven LF, Scott N, McGillis WR (2007) The combined effect of rain and wind on air-water gas exchange: a feasibility study. J Mar Syst 66:150–160
Hofmann H (2013) Spatiotemporal distribution patterns of dissolved methane in lakes: how accurate are the current estimations of the diffusive flux path? Geophys Res Lett 40:2779–2784
Huntingford C, Jones PD, Livina VN, Lenton TM, Cox PM (2013) No increase in global temperature variability despite changing regional patterns. Nature 500:327–330
Jarvis A, Reuter HI, Nelson A, Guevara E (2008) Hoe-filled seamless SRTM data v4. International Center for Tropical Agriculture, Cali. Available at http://srtm.csi.cgiar.org
Jati SR (2013) Emissão de CO2 and CH4 nas savanas úmidas de Roraima. M.Sc. Thesis, Instituto Nacional de Pesquisas da Amazonia
Johnson MS, Lehmann J, Couto EG, Novaes JP, Riha SJ (2006) DOC and DIC in flowpaths of Amazonian headwater catchments with hydrologically contrasting soils. Biogeochemistry 81:45–57
Johnson MS, Weiler M, Couto EG, Riha SJ, Lehmann J (2007) Storm pulses of dissolved CO2 in a forested headwater Amazonian stream explored using hydrograph separation. Water Resour Res 43:W11201. doi:10.1029/2007WR006359
Johnson MS, Lehmann J, Riha SJ, Krusche AV, Richey JE, Ometto J, Couto EG (2008) CO2 efflux from Amazonian headwater streams represents a significant fate for deep soil respiration. Geophys Res Lett 35:L17401. doi:10.1029/2008GL034619
Junk WJ, Piedade MTF, Schöngart J, Cohn-Haft M, Adeney JM, Wittmann F (2011) A classification of major naturally-occurring Amazonian lowland wetlands. Wetlands 31:623–640
Kemenes A, Forsberg BR, Melack JM (2007) Methane release below a hydroelectric dam. Geophys Res Lett 34:L12809. doi:10.1029/2007GL029479
Kemenes A, Forsberg BR, Melack JM (2011) CO2 emissions from a tropical hydroelectric reservoir (Balbina, Brazil). J Geophys Res Biogeosci 116:G03004. doi:10.1029/2010JG001465
Kemenes A, Forsberg BR, Melack JM (2016) Downstream emissions of CH4 and CO2 from Amazon hydroelectric reservoirs (Tucuruí, Samuel and Curuá-Una). Inland Waters (in press)
Kim JH, Zell C, Moreira-Turcq P, Pérez MAP, Abril G, Mortillaro JM, Weijers JWH, Meziane T, Sinninghe Damsté JS (2012) Tracing soil organic carbon in the lower Amazon River and its tributaries using GDGT distributions and bulk organic matter properties. Geochim Cosmochim Acta 90:163–180
Koschorreck M (2000) Methane turnover in exposed sediments of an Amazon floodplain lake. Biogeochemistry 50:195–206
Lahteenoja O, Reátegui YR, Räsänen M, Torres DC, Oinonen M, Page S (2011) The large Amazonian peatland carbon sink in the subsiding Pastaza-Maranon foreland basin, Peru. Glob Chang Biol 18:164–178
Langerwisch F, Rost S, Gerten D, Poulter B, Rammig A, Cramer W (2012) Potential effects of climate change on inundation patterns in the Amazon basin. Hydrol Earth Syst Sci Discuss 9:261–300
Lau KM, Wu HT, Kim KM (2013) A canonical response of precipitation characteristics to global warming from CMIP5 models. Geophys Res Lett 40:3163–3169
Lehner B, Döll P (2004) Development and validation of a global database of lakes, reservoirs and wetlands. J Hydrol 296:1–22
Liu J, Wang B, Cane MA, Yim S-Y, Leem J-Y (2013) Divergent global precipitation changes induced by natural versus anthropogenic forcing. Nature 493:656–659
Lloyd J, Kolle O, Fritsch H, Freitas SR, Dias MAFS, Artaxo P, Nobre AD, Araújo AC, Kruijt B, Sogocheva L, Fisch G, Thielmann A, Kuhn U, Andreae MO (2007) An airborne regional carbon balance for Central Amazonia. Biogeosciences 4:759–768
Longo KM, Freitas SR, Andreae MO, Yokelson R, Artaxo P (2009) Biomass burning in Amazonia: emissions, long-range transport of smoke and its regional and remote impacts. In: Gash J, Keller M, Silva-Dias P (eds) Amazonia and global change, Geophysical monograph series 186. American Geophysical Union, Washington, DC, pp 207–232
Macedo MN, Coe MT, DeFries R, Uriarte M, Brando PM, Neill C, Walker WS (2013) Land-use-driven stream warming in southeastern Amazonia. Philos Trans R Soc B 368:20120153, 10.1098/rstb.2012.0153
MacIntyre S, Wanninkhof R, Chanton J (1995) Trace gas exchange across the air-water interface in freshwater and coastal marine environments. In: Matson P, Harriss R (eds) Biogenic trace gases: measuring emissions from soil and water. Blackwell, Oxford, pp 52–97
MacIntyre S, Jonsson A, Jansson M, Åberg J, Turney DE, Miller S (2010) Buoyancy flux, turbulence, and the gas transfer coefficient in a stratified lake. Geophys Res Lett 37:L24604. doi:10.1029/2010GL044164
Malhi Y, Roberts JT, Betts RA, Killen TJ, Li W, Nobre CA (2008) Climate change, deforestation, and the fate of the Amazon. Science 319:169–172
Marengo JA (2004) Interdecadal variability and trends of rainfall across the Amazon basin. Theor Appl Climatol 78:79–96
Marengo JA, Nobre CA, Tomasella J, Oyama MD, de Oliveira GS, de Oliveira R, Camargo H, Alves LM, Brown IF (2008) The drought of Amazonia in 2005. J Clim 21:495–516
Marengo J, Nobre CA, Betts RA, Cox PM, Sampaio G, Salazar L (2009) Global warming and climate change in Amazonia: climate-vegetation feedback and impacts on water resources. In: Gash J, Keller M, Silva-Dias P (eds) Amazonia and global change, Geophysical monograph series 186. American Geophysical Union, Washington, DC, pp 273–292
Marengo JA, Tomasella J, Alves LM, Soares WR, Rodriguez DA (2011) The drought of 2010 in the context of historical droughts in the Amazon region. Geophys Res Lett 38:L12703. doi:10.1029/2011GL047436
Mayorga E, Logsdon MG, Ballester MVR, Richey JE (2005a) Estimating cell-to-cell land surface drainage paths from digital channel networks, with an application to the Amazon basin. J Hydrol 315:167–182
Mayorga E, Aufdenkampe AK, Masiello CA, Krusche AV, Hedges JI, Quay PD, Richey JE, Brown TA (2005b) Young organic matter as a source of carbon dioxide outgassing from Amazonian rivers. Nature 436:538–541. doi:10.1038/nature03880
Melack JM (2004) Remote sensing of tropical wetlands. In: Ustin S (ed) Manual of remote sensing, vol 4, Remote sensing for natural resources management and environmental monitoring. Wiley, New York, NY, pp 319–343
Melack JM (2005) Floodplain lakes and reservoirs in tropical and subtropical South America: limnology and human impacts. In: O’Sullivan P, Reynolds C (eds) Lakes handbook, vol 2. pp 241–257
Melack JM, Coe MT (2013) Climate change and the floodplain lakes of the Amazon basin. In: Goldman CR, Kumagai M, Robarts R (eds) Global impact of climate change on inland water systems. Wiley, New York, NY, pp 295–310
Melack JM, Engle D (2009) An organic carbon budget for an Amazon floodplain lake. Verh Int Verein Limnol 30:1179–1182
Melack JM, Forsberg B (2001) Biogeochemistry of Amazon floodplain lakes and associated wetlands. In: McClain ME, Victoria RL, Richey RE (eds) Biogeochemistry of the Amazon Basin and its role in a changing world. Oxford University Press, Oxford, pp 235–276
Melack JM, Hess LL (2010) Remote sensing of the distribution and extent of wetlands in the Amazon basin. In: Junk WJ, Piedade M, Wittmann F, Schöngart J, Parolin P (eds) Amazonian floodplain forests: ecophysiology, ecology, biodiversity and sustainable management, Ecological studies. Springer, Berlin, pp 43–59
Melack JM, Hess LL, Gastil M, Forsberg BR, Hamilton SK, Lima IBT, Novo EMLM (2004) Regionalization of methane emissions in the Amazon basin with microwave remote sensing. Glob Chang Biol 10:530–544
Melack JM, Novo EMLM, Forsberg BR, Piedade MTF, Maurice L (2009) Floodplain ecosystem processes. In: Gash J, Keller M, Silva-Dias P (eds) Amazonia and global change, Geophysical monograph series 186. American Geophysical Union, Washington, DC, pp 525–541
Miguez-Macho G, Fan Y (2012) The role of groundwater in the Amazon water cycle: 1. Influence on seasonal streamflow, flooding and wetlands. J Geophys Res 117. doi:10.1029/2012JD017539
Miller JB, Gatti LV, d’Amelio MTS, Crotwell AM, Dlugokencky EJ, Bakwin P, Artaxo P, Tans PP (2007) Airborne measurements indicate large methane emissions from the eastern Amazon basin. Geophys Res Lett 34:L10809. doi:10.1029/2006GL029213
Mortillaro JM, Abril G, Moreira-Turcq P, Sobrinh R, Perez M, Meziane T (2011) Fatty acid and stables isotopes (δ13C, δ15N) signatures of particulate organic matter in the Lower Amazon River: seasonal contrasts and connectivity between floodplain lakes and the mainstem. Org Geochem 42:1159–1168
Mortillaro JM, Rigal F, Rybarczyk H, Bernardes M, Abril G, Meziane T (2012) Particulate organic matter distribution along the lower Amazon River: addressing aquatic ecology concepts using fatty acids. PLoS One 7:e46141. doi:10.1371/journal.pone.0046141
Negrón-Juárez RI, Chambers JQ, Guimaraes G, Zeng H, Raupp CFM, Marra DM, Ribeiro GH, Higuchi N, Nelson B (2010) Widespread Amazon forest tree mortality from a single cross-basin squall line event. Geophys Res Lett 37:L16701. doi:10.1029/2010GL043733
Neu V, Neill C, Krusche AV (2011) Gaseous and fluvial carbon export from an Amazon forest watershed. Biogeochemistry 105:133–147
Ortiz AC, Ashton A, Nepf H (2013) Mean and turbulent velocity fields near rigid and flexible plants and the implications for deposition. J Geophys Res Earth Surf 118. doi:10.1002/2013JF002858
Paiva RCD, Buarque DC, Collischonn W, Bonnet MP, Frappart F, Calmant S, Mendes CAB (2013) Large-scale hydrologic and hydrodynamic modeling of the Amazon River basin. Water Resour Res 49:1226–1243
Papa F, Guntner A, Frappart F, Prigent C, Rossow WB (2008) Variations of surface water extent and water storage in large river basins: a comparison of different global data sources. Geophys Res Lett 35:L11401. doi:10.1029/2008GL033857
Papa F, Prigent C, Jimenez C, Aires F, Rossow WB (2010) Interannual variability of surface water extent at global scale, 1993–2004. J Geophys Res 115:D12111. doi:10.1029/2009JD012674
Pires GF, Costa MH (2013) Deforestation causes different subregional effects on the Amazon bioclimatic equilibrium. Geophys Res Lett 40:3618–3623
Polsenaere P, Deborde J, Detandt G, Vidal LO, Pérez MAP, Marieu V, Abril G (2013) Thermal enhancement of gas transfer velocity of CO2 in an Amazon floodplain lake revealed by eddy covariance measurements. Geophys Res Lett 40:1734–1740
Potter CS (1997) An ecosystem simulation model for methane production and emission from wetlands. Global Biogeochem Cycles 11:495–506
Potter CS, Melack JM, Engle D (2016) Modeling carbon dynamics and methane emissions from Amazon floodplain ecosystems. Wetlands 11:501–511
Prigent C, Papa F, Aires F, Rossow WB, Matthews E (2007) Global inundation dynamics inferred from multiple satellite observations, 1993–2000. J Geophys Res 112:D12107. doi:10.1029/2006JD007847
Prigent C, Papa F, Jimenez C, Rossow WB, Matthews E (2012) Changes in land surface water dynamics since the 1990s and relation to population pressures. Geophys Res Lett 39:L08403. doi:10.1029/2012GL051276
Rasera MFFL, Ballester MVR, Krusche AV, Salimon C, Montebelo LA, Alin SR, Victoria RL, Richey JE (2008) Estimating the surface area of small rivers in the southwestern Amazon and their role in CO2 outgassing. Earth Interact 12:1–16. doi:10.1175/2008EI257.1
Rasera MFFL, Krusche AV, Richey JE, Ballester MVR, Victória RL (2013) Spatial and temporal variability of pCO2 and CO2 efflux in seven Amazonian rivers. Biogeochemistry 116:241–259
Raymond PA, Hartmann J, Lauerwald R, Sobek S, McDonald C, Hoover M, Butman D, Striegl R, Mayorga E, Humborg C, Kortelainen P, Dürr H, Meybeck M, Ciais P, Guth P (2013) Global carbon dioxide emissions from inland waters. Nature 503:355–359
Reichstein M, Bahn M, Ciais P, Frank D, Mahecha MD, Seneviratne SI et al (2013) Climate extremes and the carbon cycle. Nature 500:287–295
Remington S, Krusche AV, Richey JM (2011) Effects of DOM photochemistry on bacterial metabolism and CO2 evasion during falling water in a humic and a whitewater river in the Brazilian Amazon. Biogeochemistry 105:185–200
Renó VF, Novo EMLM, Suemitsu C, Rennó CD, Silva TSF (2011) Assessment of deforestation in the lower Amazon floodplain using historical landsat MSS/TM imagery. Remote Sens Environ 115:3446–3456
Rice AL, Butenhoff CL, Shearer MJ, Teama D, Rosenstiel TN, Khalil AK (2010) Emissions of anaerobically produced methane by trees. Geophys Res Lett 37:L03807. doi:10.1029/2009GL041565
Richey JE, Melack JM, Aufdenkampe AK, Ballester VM, Hess L (2002) Outgassing from Amazonian rivers and wetlands as a large tropical source of atmospheric carbon dioxide. Nature 416:617–620
Richey JE, Krusche AV, Johnson MS, da Cunha HB, Ballester MV (2009) The role of rivers in the regional carbon balance. In: Gash J, Keller M, Silva-Dias P (eds) Amazonia and global change, Geophysical monograph series 186. American Geophysical Union, Washington, DC, pp 489–504
Riley WJ, Subin AM, Lawrence DM, Swenson SC, Torn MS, Meng L et al (2011) Barriers to predicting changes in global terrestrial methane fluxes: analyses using CLM4Me, a methane biogeochemistry model integrated in CESM. Biogeosciences 8:1925–1953. doi:10.5194/bg-8-1925-2011
Ringeval B, Noblet-Ducoudré N, Ciais P, Bousquet P, Prigent C, Papa F, Rossow WB (2010) An attempt to quantify the impact of changes in wetland extent on methane emissions on the seasonal and interannual time scales. Global Biogeochem Cycles 24:GB2003. doi:10.1029/2008GB003354
Roland F, Vidal LO, Pacheco FS, Barros NO, Assireu A, Ometto HPHB, Cimbleris ACP, Cole JJ (2010) Variability of carbon dioxide flux from tropical (Cerrado) hydroelectric reservoirs. Aquat Sci 72:283–293
Rosenqvist A, Forsberg BR, Pimentel T, Rauste YA, Richey JE (2002) The use of spaceborne radar data to model inundation patterns and trace gas emissions in the central Amazon floodplain. Int J Remote Sens 23:1303–1328
Rudorff CM (2013) Flooding and carbon dynamics on the lower Amazon floodplain. Ph.D. Thesis, University of California, Santa Barbara
Rudorff CM, Melack JM, MacIntyre S, Barbosa CCF, Novo EMLM (2011) Seasonal and spatial variability in CO2 emissions from a large floodplain lake in the lower Amazon. J Geophys Res Biogeosci 116:G04007. doi:10.1029/2011JG001699
Rudorff CM, Melack JM, Bates PD (2014a) Flooding dynamics on the lower Amazon floodplain: 1. Hydraulic controls on water elevation, inundation extent and river-floodplain discharge. Water Resour Res 50. doi:10.1002/2013WR014091
Rudorff CM, Melack JM, Bates PD (2014b) Flooding dynamics on the lower Amazon floodplain: 2. Seasonal and interanual hydrological variability. Water Resour Res 50. doi:10.1002/2013WR014714
Sawakuchi HO, Bastiviken D, Sawakuchi AEO, Krusche AV, Ballester MVR, Richey JE (2014) Methane emissions from Amazonian rivers and their contribution to the global methane budget. Glob Chang Biol. doi:10.1111/gcb.12646
Silva TSF, Costa M, Melack JM (2009) Annual net primary production of macrophytes in the eastern Amazon floodplain. Wetlands 29:747–758
Silva TSF, Costa MPF, Melack JM (2010) Spatio-temporal variability of macrophyte cover and productivity in the eastern Amazon floodplain: a remote sensing approach. Remote Sens Environ 114:1998–2010
Silva TSF, Melack JM, Novo EML (2013) Responses of aquatic macrophyte cover and productivity to flooding variability on the Amazon floodplain. Glob Chang Biol 19:3379–3389
Stanley EH, Casson NJ, Christel ST, Others (2015) The ecology of methane in streams and rivers: patterns, controls, and global significance. Ecol Monogr. doi:10.1890/15-1027.1
Victoria DC (2010) Simulacao hidrologica de bacias amazonicas utilizando o modelo de Capacidade de Infiltracao Variavel (VIC). PhD. Thesis, University of Sao Paulo, Piracicaba
Vihermaa LE, Waldron S, Garnett MH, Newton J (2014) Old carbon contributes to aquatic emissions of carbon dioxide in the Amazon. Biogeosciences 11:3635–3645
Villar E, Jhan C, Ronchail J, Guyot JL, Junquas C, Vauchel P, Casimiro L, Waldo S, Drapeau G, Pombosa Loza LR (2011) Climate variability and extreme drought in the upper Solimões River (Western Amazon Basin): understanding the exceptional 2010 drought. Geophys Res Lett 38:L13406. doi:10.1029/2011GL047862
Waichman AV (1996) Autotrophic carbon sources for heterotrophic bacterioplankton in a floodplain lake of Central Amazon. Hydrobiologia 341:27–36
Walter BP, Heimann M (2000) A process-based, climate-sensitive model to derive methane emissions from natural wetlands: application to five wetland sites, sensitivity to model parameters, and climate. Global Biogeochem Cycles 14:745–765
Ward ND, Keil RG, Medeiros PM, Brito DC, Cunha AC, Dittmar T, Yager PL, Krusche AV, Richey JE (2013) Degradation of terrestrially derived macromolecules in the Amazon River. Nat Geosci. doi:10.1038/ngeo1817
Wilson MD, Bates PD, Alsdorf A, Forsberg B, Horritt M, Melack J, Frappart F, Famglietti JS (2007) Modeling large-scale inundation of Amazonian seasonally flooded wetlands. Geophys Res Lett 34:L15404. doi:10.1029/2007GL030156
Worbes M (1997) The forest ecosystem of the floodplains. In: Junk WJ (ed) The Central Amazon floodplain, Ecological studies. Springer, Berlin, pp 223–260
Xavier L, Becker M, Cazenave A, Longuevergne L, Llovel W, Rotunno Filho OC (2010) Interannual variability in water storage over 2003–2008 in the Amazon Basin from GRACE space gravimetry, in situ river level and precipitation data. Remote Sens Environ 114:1626–1637
Yamazaki D, Kanae S, Kim H, Oki T (2011) A physically based description of floodplain inundation dynamics in a global river routing model. Water Resour Res 47:W04501. doi:10.1029/2010WR009726
Zeng N, Yoon JH, Marengo JA, Subramaniam A, Nobre CA, Mariotti A, Neelin JD (2008) Causes and impacts of the 2005 Amazon drought. Environ Res Lett 3. doi:10.1088/1748-9326/3/1/014002
Acknowledgements
S. MacIntyre, L. Hess, G. Abril, J. Richey, B.R. Forsberg, and L. Nagy provided helpful comments on the manuscript and its development. The writing and related research was supported by NASA’s Terrestrial Ecology Program.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Melack, J.M. (2016). Aquatic Ecosystems. In: Nagy, L., Forsberg, B., Artaxo, P. (eds) Interactions Between Biosphere, Atmosphere and Human Land Use in the Amazon Basin. Ecological Studies, vol 227. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-662-49902-3_7
Download citation
DOI: https://doi.org/10.1007/978-3-662-49902-3_7
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-662-49900-9
Online ISBN: 978-3-662-49902-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)