
Abstract
Antibiotics remain one of our most important pharmacological tools for the control of infectious disease. However, unlike most other drugs, the use of antibiotics selects for resistant organisms and erodes their clinical utility. Resistance can emerge within populations of bacteria by mutation and be retained by subsequent selection or by the acquisition of resistance elements laterally from other organisms. The source of these resistance genes is only now being understood. The evidence supports a large bacterial resistome—the collection of all resistance genes and their precursors in both pathogenic and nonpathogenic bacteria. These genes have arisen by various means including self-protection in the case of antibiotic producers, transport of small molecules for various reasons including nutrition and detoxification of noxious chemicals, and to accomplish other goals, such as metabolism, and demonstrate serendipitous selectivity for antibiotics. Regardless of their origins, resistance genes can rapidly move through bacterial populations and emerge in pathogenic bacteria. Understanding the processes that contribute to the evolution and selection of resistance is essential to mange current stocks of antibiotics and develop new ones.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Antibiotics: Essential Medicines in Peril
The first compounds purposely discovered to combat infection were the arsenicals such as Salvarsan reported by Paul Ehrlich at the turn of the twentieth century. These were highly toxic molecules that nevertheless were the first successful drugs for the treatment of syphilis, then a worldwide scourge caused by the bacterial pathogen Treponema pallidum (and now in the twenty-first century re-emerging in antibiotic-resistant forms). The success of these compounds in infection control inspired the development of the sulpha drugs 15 years later and ushered in the “Age of Antibiotics” that we continue to enjoy. The mid-twentieth century saw the discovery of the vast majority of antibiotic chemical scaffolds now in clinical use. This era of discovery, roughly from 1940 to 1960, was followed by a period of great innovation where medicinal chemists modified these scaffolds to improve the pharmacological properties of antibiotics, making them better medicines. At the same time, chemical modification of antibiotic scaffolds was a means to overcome another problem associated with antibiotics—resistance.
Resistance to all classes of antibiotics occurs after their use in the clinic or on the farm. In the case of penicillin, resistance was actually characterized before clinical deployment (Abraham and Chain 1940)! The emergence of resistance, often quite rapidly after being made available to the clinical community, helps to stimulate the discovery of new antibiotic scaffolds and their subsequent chemical modification (when possible) to structures less susceptible to resistance. This has resulted in many cases in several “generations” of antibiotics (e.g., Fig. 1) each with improved properties including expanded bacterial spectrum, favorable pharmacology, and “resistance to resistance.”

Antibiotic improvement through chemical derivatization. The majority of antibiotics are natural products that were first introduced in the form isolated from producing organisms (First Generation). Subsequent modification of the active core scaffold by medicinal chemistry, as shown for the examples in the penicillin and tetracycline classes improve their drug properties and avoid prominent resistance mechanisms
Despite these cycles of antibiotic discovery and innovation, resistance remains a significant and growing challenge to the successful treatment of infectious disease. A principal reason for this is that new drug development over the past decade has lagged behind the unavoidable selection for resistance. Challenged with fewer leads for new antibiotic scaffolds and in some cases approaching the limits of productive chemical modification of known scaffolds, the growing costs of drug development, and a challenging regulatory environment, many players in the pharmaceutical industry have abandoned new antibiotic discovery (Spellberg et al. 2004; Williams and Bax 2009; Gwynn et al. 2010). The result is a growing unmet clinical need and an ever-widening gap in early stage antibiotic innovation and delivery of new antibiotics to the clinic (Fischbach and Walsh 2009).
This alarming trend has occurred at the same time that resistance to all antibiotics continues to evolve and spread across the globe (Boucher et al. 2009). Furthermore, pathogens are now very often multidrug resistant, harboring many resistance genes (sometimes dozens) and frequently they are associated with increased virulence. In some cases, highly successful multidrug resistant bacteria are emerging as epidemic and pandemic strains (Croucher et al. 2009; Freitas et al. 2010; Peirano and Pitout 2010). The resulting erosion of drug efficacy is now being widely viewed as a global health crisis that needs to be recognized and addressed with new drug discovery (Infectious Diseases Society of America 2010).
The root cause of this crisis is antibiotic resistance. Recent efforts to understand the evolution and origins of resistance are building on decades of research in epidemiology and fundamental biology to formulate a unifying hypothesis that ties drug resistance to bacterial genomics and physiology. This concept, the antibiotic resistome, provides a framework to understand how and why resistance emerges and offers the prospect for clinicians and drug discoverers to predict the emergence of clinical resistance and probe the long-term efficacy of new drugs. The origins and evolution of antibiotic resistance elements that comprise the resistome are the focus of this chapter.
2 The Antibiotic Resistome
The predictable and often rapid emergence of antibiotic resistance in previously susceptible pathogenic bacteria was recognized very early in the history of antibiotic drug discovery. Indeed, Fleming, the discoverer of penicillin, warned of the prospect of resistance in his Nobel Prize speech in 1945 noting: “It is not difficult to make microbes resistant to penicillin in the laboratory by exposing them to concentrations not sufficient to kill them, and the same thing has occasionally happened in the body.” (Fleming 1945). Why are all antibiotics susceptible to resistance and why does resistance appear so quickly?
Antibiotics are small organic molecules (MW ~100–2,000 Da) composed of C, H, N, O, S, and other biologically available elements. Microbes live their lives surrounded by these and countless similar molecules. Among them are nutrient sources (carbohydrates, peptides, lipids, vitamins, and cofactors), communication compounds produced by bacteria and other microbes (e.g., quorum-sensing molecules), siderophores made by fungi and bacteria to secure iron that is essential for their growth, assorted compounds produced by microbes (including antibiotics, surfactants, hormones, etc.), and countless molecules including terpenoids, steroids, and alkaloids produced by plants and other eukaryotes such as protists and algae. The world of bacteria is a world of small molecules. They are their language and their life source.
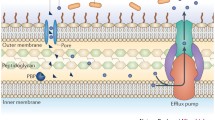
Many of these compounds, however, are toxic to bacteria. As a result, they have developed strategic countermeasures to overcome the noxious properties of chemicals that they may come across during their life cycle. These include nonspecific mechanisms to block the entry of toxic compounds into cells or decrease their intracellular concentrations. Such mechanisms are a first line of defense against toxins that a bacterium may encounter in its environment. Additionally, highly specific mechanisms to evade the activities of certain toxic molecules (among them many naturally produced antibiotics) have also evolved. Examples of such mechanisms include the alteration or mutation of the cellular targets of antibiotics and the development of degrading or modifying enzymes. The evolution of microbial defenses to guard against both general toxins and specific molecules has occurred over the millennia. The fossil record shows that stromatolites, the remnants of cyanobacteria, prodigious producers of toxic small molecules, are greater than 3.4 billion years old (Allwood et al. 2006) and land plants, equally important producers of small molecules, emerged 475 million years ago (Wellman et al. 2003). Consequently, all bacteria have developed some ability to resist toxic small molecules; indeed some microbes are prodigiously equipped to evade them.
The focus of much of the antibiotic resistance literature has (not surprisingly) been on the emergence of resistance in previously susceptible pathogens in the clinic. However, this focus has disproportionately emphasized a small subgroup of microbes. The vast majority of bacteria do not cause human or animal disease; nevertheless, they are continuously exposed to toxic small molecules, including naturally occurring antibiotics, and have consequently developed resistance. For example, a survey of ~500 soil microbes against a panel of 21 antibiotics including natural products, semisynthetic derivatives of these, and completely synthetic molecules revealed a remarkable ability of these organisms to resist antibiotics (D’Costa et al. 2006). All strains in this study were resistant to multiple drugs and on average they were impervious to 7–8 antibiotics. The source of the antibiotic did not matter; natural products or synthetic molecules were equally susceptible. This observation is consistent with evolution of both general mechanisms of detoxification, e.g., the efflux of molecules from the cell, and highly specific mechanisms such as enzymatic detoxification.
This study led us to propose the concept of an antibiotic resistome (D’Costa et al. 2006, 2007; Wright 2007, 2010). The resistome includes all the genes that directly or indirectly act to confer antibiotic resistance in bacteria. It includes the resistance elements in pathogenic bacteria and also the much more diverse and populous nonpathogenic organisms. Furthermore, the resistome includes the microbial genes that contribute to inherent antibiotic resistance in a particular organism, the intrinsic resistome (Fajardo et al. 2008). Finally, the “housekeeping” genes that can give rise to resistance elements either by mutation or overexpression are also constituents of the resistome. We have termed these “proto-resistance” elements in functional analogy to proto-oncogenes in cancer (Morar et al. 2009; Morar and Wright 2010).
The components of the resistome therefore vastly exceed the traditional narrow focus of the clinical and scientific communities on resistance in pathogens. A growing number of studies of microbes that can subsist on antibiotics (Dantas et al. 2008) and on the prevalence of resistance genes in the human microbiome (Sommer et al. 2009) and in the environmental metagenome (Riesenfeld et al. 2004; Allen et al. 2009; Donato et al. 2010; Lang et al. 2010) emphasize the point. Importantly, we now understand that unlike most complex organisms, gene flow in bacteria can readily occur vertically (from mother to daughter cell) and also horizontally (between cells) (Roy 1999; Barlow 2009). Therefore, antibiotic resistance that evolves in one bacterium will be passed on to its progeny as is common to all organisms, but it can also be mobilized (by transduction, transformation, or conjugation) into unrelated bacteria (Fig. 2). In this fashion, resistance in nonpathogenic bacteria can “escape” and be captured by pathogens. Given the vast numbers of prokaryotic cells on the planet [~5 × 1030 (Whitman et al. 1998)], their innate and evolved abilities to evade the lethal effects of antibiotics and other toxic small molecules developed over millions of years, and the relative ease of gene movement in microbes, the rapid and inevitable emergence of resistance to antibiotics in the clinic becomes completely predictable.

Genes move laterally through the resistome. Antibiotics provide a powerful selection pressure to mobilize and capture resistance elements in the resistome. The ability of genes to cross species and genus barriers by transformation, conjugation, and transduction provides a large gene pool for pathogens to draw on
2.1 Highly Efficient Resistance Elements in Nonpathogenic Bacteria
In 1973, Benveniste and Davies noted the similarity between the activity of aminoglycoside antibiotic modifying enzymes in the soil microbes that produce these drugs and the enzymes responsible for resistance in pathogens (Benveniste and Davies 1973). They identified the activities of aminoglycoside acetyltransferases, adenylyltransferases and phosphotransferases in cell free extracts of a number of producing organisms. The specificity of these enzymes matched the biochemical profiles of aminoglycoside resistance enzymes that had begun to emerge as a significant clinical problem since their first detection as horizontally acquired resistance elements in the 1950s (Davies 1995). The subsequent cloning and sequencing of aminoglycoside resistance genes from both pathogens and producers showed that the associated enzymes were closely related and demonstrated for the first time an unexpected link between mechanisms of resistance in antibiotic producers and clinically important pathogens.
In retrospect, it should not have been surprising to discover that antibiotic producers would also require intrinsic resistance elements to avoid committing suicide during drug production (Cundliffe 1989). It is now well established that secondary metabolites of bacterial origin, including antibiotics, are the products of large biosynthetic gene clusters. These clusters include the genes required for the generation of the individual components of the secondary metabolites (sugars, unusual amino acids, etc.), the assembly of these components into the proper scaffold structure (e.g., by nonribosomal peptide or polyketide synthetases), the modification of the scaffold (e.g., acylation, glycosylation, halogenation), transport of the compound from the cell, and in the case of antibiotics, resistance elements. The number of characterized biosynthetic gene clusters has increased dramatically over the past 15 years and frequently these are associated with genes that encode resistance elements that share function and structure with those found in pathogens.
Sequencing of the complete genome of the erythromycin producer Saccharopolyspora erythraea provides an example (Oliynyk et al. 2007). The genome sequence revealed the expected ~55 kb erythromycin biosynthetic gene cluster including ermE, an associated resistance gene. Erm genes are common resistance elements in pathogenic bacteria, associated with the MLSB drug resistance phenotype, so called because they provide overlapping resistance to the structurally unrelated macrolide antibiotics such as erythromycin, the lincosamides such as clindamycin, and the type B streptogramins (Maravic 2004). Erm gene products are methyltransferases that specifically modify the exocyclic amine of a key adenosine residue (A2058 Escherichia coli numbering) of the 23S rRNA that is an essential component of the large ribosomal subunit (Mukhtar and Wright 2005). The structure of the ribosome in complex with MLSB antibiotics reveals that these compounds bind to the same region of the ribosome, the peptide exit tunnel, and that methylation of A2058 results in a steric and ionic barrier to drug binding (Tu et al. 2005). The presence of erm genes in many other macrolide producers (Cundliffe et al. 2001; Ward et al. 2004; Karray et al. 2007) demonstrates that the 23S rRNA methylation is a common link between pathogens and antibiotic-producing environmental bacteria.
Resistance to glycopeptide antibiotics such as vancomycin provides yet another example of the links between the clinic and antibiotic resistance in producing organisms. Vancomycin binds to the acyl-d-Alanine-d-Alanine terminus of the bacterial peptidoglycan, an essential component of the cell wall that protects bacteria from cell lysis. This noncovalent interaction through five hydrogen bonds prevents the growth of the peptidoglycan that is essential during cell division providing the rationale for the antibacterial activity of glycopeptides. Resistance to vancomycin emerged in the late 1980s in clinical isolates of Enterococci (Leclercq et al. 1988; Courvalin 2006). Subsequent determination of the mechanism of resistance on associated genes revealed that resistance was the result of the concerted action of five gene products (Walsh et al. 1996). Two of these, VanR and VanS, make up a two-component regulatory system that senses the presence of the antibiotic (Koteva et al. 2010). This triggers the expression of three genes: vanH, vanA, and vanX. VanH produces d-Lactate from pyruvate and VanA is an ATP-dependent ligase that forms an ester bond between d-Alanine and d-Lactate. The d-Ala-d-Lac ester (depsipeptide) is a substrate for MurF that adds the depsipeptide to the peptidoglycan monomer, which is then exported to the exterior of the cell to be grafted onto the growing peptidoglycan polymer.
Replacing d-Ala-d-Ala-terminating peptidoglycan with a d-Ala-d-Lac terminus has a dramatic effect on antibiotic action. The loss of the amide hydrogen of d-Ala-d-Ala (replaced by an ester oxygen in d-Ala-d-Lac) removes one of the key hydrogen bond interactions between the drug and the peptidoglycan (Bugg et al. 1991). The result is a 1,000-fold decrease in drug affinity and clinical resistance. Because the cell continues to make d-Ala-d-Ala even in the presence of vancomycin, the cell wall could be a mixture of d-Ala-d-Ala and d-Ala-d-Lac terminating peptidoglycan, thereby diminishing resistance. The expression of VanX ensures that the peptidoglycan is dominated by d-Ala-d-Lac by selectively hydrolyzing any d-Ala-d-Ala in the cell (Wu et al. 1995).
The origin of this highly effective and complex mechanism of resistance was the subject of significant debate when vancomycin resistance first emerged in the clinic. The resistance genes in Enterococci were associated with a transposon and mobilized on a plasmid. Subsequent sequencing of glycopeptide antibiotic biosynthetic gene clusters and the associated resistance genes revealed that the likely origins of the vanHAX operon are in drug-producing bacteria (Marshall et al. 1998). The operon has now been found in most glycopeptide producers. Furthermore, vanHAX was later identified in glycopeptide-resistant environmental strains of Paenibacillus and many other soil organisms, which do no impact human health, demonstrating that glycopeptide resistance was not restricted to human pathogens (Hong et al. 2004; Guardabassi et al. 2005; D’Costa et al. 2006; Guardabassi and Agerso 2006). The most parsimonious explanation of the available evidence suggests that the vanHAX operon originated in glycopeptide producers as a means of self-resistance and has spread laterally to neighboring nonproducing organisms and, with the selection offered in hospitals due to increased use of vancomycin to treat infections caused by drug-resistant Gram-positive pathogens, eventually to pathogenic Enterococci. However, the high GC content of the vanHAX genes in producing bacteria is significantly different to that in the operons found in Enterococci suggesting that gene escape and transfer was not a recent event. Given the distribution of vanHAX in nonproducing bacteria, it is likely that there is a very large pool of these genes in environmental bacteria.
The similarity of resistance elements in antibiotic-producing bacteria and pathogenic bacteria demonstrates a clear link between the environment and the clinic. However, as noted earlier for the vancomycin resistance genes, even nonproducing bacteria have acquired the operon. Similarly, in addition to ermE, the sequence of the S. erythraea genome revealed many predicted antibiotic resistance genes, including 17 β-lactamases, 2 aminoglycoside phosphotransferases, and an aminoglycoside acetyltransferase, even though this organism does not produce β-lactams or aminoglycosides. This observation is recapitulated in the sequences of many bacterial genomes. Antibiotic resistance elements are not rare.
The question of whether these environmental organisms present a reservoir for contemporary transfer of resistance genes to pathogens has been answered with the case of the emergence of the CTX-M β-lactamases. These enzymes emerged in the 1980s as elements that confer resistance to the oxyimino-cephalosporins such as cefotaxime and have become significant clinical problems (Canton and Coque 2006). The genes were associated with plasmids that were horizontally transferred between Enterobacteriaceae. The origins of the ctx-m genes were unknown as their products showed only ~40% similarity to known β-lactamases. This changed with the cloning of the β-lactamases genes from several species of the nonpathogenic bacterium Kluyvera, which are often found in the environment. These chromosomal genes showed up to 100% identify to the ctx-m genes found in clinical pathogens and often included flanking genes shared between Kluyvera chromosomes and circulating plasmids in Enterobacteriaceae (Decousser et al. 2001; Humeniuk et al. 2002; Poirel et al. 2002; Olson et al. 2005). These results point to a recent escape of CTX-M from Kluyvera with subsequent spread among pathogenic bacteria via lateral gene transfer. The net result has been termed the CTX-M pandemic (Canton and Coque 2006).
Thus nonpathogenic bacteria, including antibiotic producers, are reservoirs of antibiotic resistance genes that can make their way into populations of pathogens by later gene transfer.
2.2 De Novo Evolution of Resistance
Resistance can also emerge spontaneously within bacterial populations. This can be the result of mutagenesis with subsequent selection of favorable resistance traits, for example in a patient undergoing antibiotic therapy. The error rate of DNA polymerase is ~1 base in 109. Therefore for E. coli with a genome size of ~5 × 106, one random mutation occurs in the genome every 200 cell divisions. In a population of bacteria in an infection, the number of mutants can be considerable. Of course many of these are silent or even fatal, but some may offer some advantage. This is especially true for slow growing organisms such as Mycobacterium tuberculosis, where mutations are the principal cause of drug resistance and chemotherapy lasts for months. In a study on the development of resistance to vancomycin and other antibiotics in a patient undergoing chemotherapy, Tomasz’s group sequenced the genomes of increasingly resistant Staphylococcus aureus isolates recovered from a patient’s bloodstream over a period of ~4 months (Mwangi et al. 2007). During this time, the strain accumulated 35 point mutations in 31 loci. The susceptibility to vancomycin decreased from 1 μg/mL at the beginning of therapy to 8 1 μg/mL after 12 weeks when the patient died. Furthermore, these mutations also conferred increased resistance to daptomycin.
The accumulation of point mutations with selection therefore has an important role in the evolution of resistance in pathogens. Another example of the significance of point mutations is in the development of resistance to the quinolone antibiotics. These are synthetic compounds, first identified in the early 1960s and developed in earnest in the 1970s, 1980s, and 1990s when 6-fluoroderivatives (the fluoroquinolones) were found to have excellent broad spectrum activity against a number of important pathogens, including multidrug resistant strains. These antibiotics target bacterial topoisomerases that catalyze double-strand breaks in DNA such as the type IIA enzymes DNA gyrase (induces supercoiling and unwinds strands at the replication fork; it is comprised of dimers of two subunits, GyrA and GyrB) and type IV topoisomerases (also a tetramer, e.g., of ParC/ParE, that decatenates daughter strands following DNA replication). These enzymes form a covalent adduct with the DNA during cleavage to enable controlled strand passage during catalysis (Laponogov et al. 2010). Quinolones have little affinity for the free enzyme, but rather bind to this cleavage complex, blocking enzyme activity. This potently arrests cell division and results in cell death (Hooper 2001).
Resistance to the quinolones arises most frequently as a result of point mutations in gyrA or parC. A region of the enzyme termed the Quinolone Resistance Determining Region (QRDR) is most frequently associated with resistance. Mutations in key residues such as Ser83 and Asp87 (E. coli GyrA numbering) are among the most common in the clinic. Additional mutations in parC and secondary mutations in both genes incrementally increase resistance (Hooper 1999). Mutations in the QRDR region are not confined to resistant pathogenic bacteria, but these have also been found in environmental organisms (Waters and Davies 1997; D’Costa et al. 2006). These presumably arise without the selective pressure of exposure to the synthetic fluoroquinolones and thus reflect either natural variation in these genes, or the selective pressure of an as yet undiscovered class of natural products that impact the same target.
The presumption has been that such mutations have associated with them fitness costs that will be selected against in the absence of antibiotics. Indeed, this is true in many cases [reviewed in (Andersson 2006; Andersson and Hughes 2010)]. This would imply that removal of antibiotics would select for loss of resistance. However in many cases, compensatory secondary mutations in the target gene or others on the chromosome improve the fitness of resistant bacteria [see Table 2 in ref (Andersson and Hughes 2010)]. Thus, resistant mutations can propagate vertically in bacteria populations even in the absence of selection.
Antibiotic exposure is stressful to bacteria, and there have been several studies showing that exposure to an antibiotic or biocide alters the expression of hundreds of genes (Shaw et al. 2003; Cirz et al. 2006; Dwyer et al. 2007; Bailey et al. 2010). This complex response can in some cases converge on important pathways. For example, E. coli produced toxic reactive oxygen species including hydroxyl radicals in response to bactericidal antibiotics (Dwyer et al. 2007). Importantly, this was further correlated with increases in the genome mutation rate leading to the hypothesis that antibiotics transiently increase the bacterial mutation rate by generating DNA damage (Kaufmann and Hung 2010; Kohanski et al. 2010). Similar results we noted upon exposure of Pseudomonas aeruginosa to ciprofloxacin where the error-prone DNA repair and synthesis proteins were up-regulated (Cirz et al. 2006). This could allow antibiotic-resistant bacteria to be selected at an elevated frequency while under pressure from antibiotics and could account for the emergence of resistance by mutation during chemotherapy.
Point mutations are not the only mechanism of de novo resistance development. Gene duplication or higher order amplification is also common and has been linked to gene evolution by providing “substrate” for mutation and innovation (Sandegren and Andersson 2009). For example, some antibiotics (for example, fluoroquinolones) are often associated with the selection in vitro and in vivo of multidrug resistant bacteria due to over-production of RND (Resistance-Nodulation-cell Division) type efflux pumps in Gram-negative bacteria (e.g., in Enterobacteriaceae AcrAB–TolC, MexAB–OprM in P. aeruginosa and MtrCDE in Neisseria spp.) (Piddock 2006). These pumps confer innate resistance to many of the agents commonly used to treat infections by Gram-positive bacteria, and when deleted not only does the Gram-negative bacterium become hyper-drug susceptible but virulence can also be attenuated (Piddock 2006). Control and regulation of expression of the components of MDR efflux pumps is still not fully understood but can occur at either a local gene level (e.g., for acrAB by AcrR and mexAB-oprM by MexR) or at a global level such as in E. coli via MarA (White et al. 1997; Pomposiello and Demple 2000), RamA in Klebsiella pneumoniae and Salmonella enterica (Ricci and Piddock 2009) and MtrR in Neisseria gonorrheae (Folster et al. 2009). Mutations in these control elements can result in the amplification of RND pump expression and associated resistance.
A direct screen of the potential role of gene amplification in antibiotic resistance was recently reported (Soo et al. 2011). The authors screened an E. coli library of ~4,000 overexpressing clones for increased resistance to 237 antibiotics and other toxic compounds. Resistance to 86 of these was observed including many antibiotics (β-lactams, aminoglycosides, macrolides, and antifolates) upon overexpression of 61 genes. Eighteen of these genes included global mechanisms of resistance (efflux, regulators, stress proteins, capsule synthesis). The remaining 41 genes were specific to individual antibiotics including 12 that encode metabolic enzymes that could contribute to compound modification. These results in a “wild type” drug susceptible organism point to the relative ease of selecting resistance by gene amplification mechanisms and points to the importance of such events as a first step in the evolution of highly specific resistance elements that can be transferred vertically to progeny and can escape to move laterally in bacterial populations.
2.3 Proto-Resistance
The case described earlier where 12 metabolic genes were identified that can confer antibiotic resistance upon gene amplification in E. coli (Soo et al. 2011) demonstrates the capacity of microorganisms to capitalize on weak, cryptic, or promiscuous activities to evolve new metabolism. This has been termed the innovation–amplification–divergence model of gene evolution (Bergthorsson et al. 2007). The challenge in the evolution of genes with new functions after a gene duplication event is the maintenance of both the original gene and the duplicate while enabling divergence in the duplicate. Antibiotics provide the strong selective pressure to ensure that the duplicate gene is not lost in subsequent generations. Furthermore, it provides the requisite selection for innovation and evolution in the new gene. The determination of the gene (and protein) sequences of highly efficient resistance elements along with their biochemical and structural properties provides an opportunity to hypothesize on the origins of many resistance proteins. We have termed these proto-resistance genes and proteins (Morar and Wright 2010).
Among the first links between resistance biochemistry and metabolic processes was noted between peptidoglycan biosynthesis proteins and β-lactamases (Kelly et al. 1986). The β-lactamases utilize one of two general mechanisms to hydrolytically cleave the β-lactam ring of the antibiotics. Metallo-β-lactamases use an active site Zn2+ ion (more often two) to active a water molecule for attack on the β-lactam carbonyl carbon resulting in a ring-opening drug inactivation. These enzymes were previously rare in the clinic but are growing in importance as they confer resistance to virtually all β-lactams including the “last resort” carbapenems (De Pascale and Wright 2010). The more prevalent mechanism in pathogens operates through an active site Asp-His-Ser catalytic triad. The Ser hydroxyl group attacks the carbonyl of the β-lactam ring forming a covalent-enzyme intermediate and subsequent cleavage of this complex by water generates the inactive antibiotic. The first step of the reaction, formation of the acyl-enzyme intermediate, is highly reminiscent of the activity of β-lactams on their cellular targets. β-Lactam antibiotics covalently inactivate enzymes involved in peptidoglycan biosynthesis (Massova and Mobashery 1998). These so-called penicillin-binding proteins (PBPs) also operate by an active site Asp-His-Ser catalytic triad. Analogously, the Ser attacks the β-lactam ring resulting in a covalent intermediate. However unlike the β-lactamases, subsequent hydrolysis does not occur and the enzyme is permanently in the inactive form. This results in an inability to generate new peptidoglycan and subsequent inhibition of cell division.
This convergence in biochemical function was further strengthened with the determination of the structures of β-lactamases and PBPs. The first was the comparison of the 3D structure of the d-alanyl-d-alanine-peptidase from Streptomyces R61 with the β-lactamase from Bacillus licheniformis (Kelly et al. 1986). Despite a lack of significant amino acid sequence similarity, these enzymes showed remarkable conservation of structure and mechanism. This 25-year-old observation has been supported by the determination of the structure and function of many PBPs and β-lactamases (Maveyraud et al. 1998). PBPs are therefore the most likely β-lactamase proto-resistance elements.
Similarly, aminoglycoside phosphotransferases have the same protein fold as Ser/Thr/Tyr protein kinases despite a lack of sequence similarity (Hon et al. 1997; Nurizzo et al. 2003; Young et al. 2009; Fong et al. 2010; Toth et al. 2010; Stogios et al. 2011). Furthermore, these enzymes have the capacity to phosphorylate peptides and proteins (Daigle et al. 1998) and their activities blocked by protein kinase inhibitors (Daigle et al. 1997). These enzyme families likely share a common ancestor.
There are many other examples emerging of the ties between resistance and metabolic processes [reviewed in (Morar and Wright 2010)]. A contemporary example is the emergence of a variant of an aminoglycoside antibiotic inactivating acetyltransferase that has the ability to inactivate a subset of fluoroquinolone antibiotics (Robicsek et al. 2006; Vetting et al. 2008). This determinant has since spread worldwide by lateral gene transfer (Strahilevitz et al. 2009). The strong selective pressure provided by antibiotics along with the number of both pathogens and nonpathogens exposed to antibiotics across the globe predicts that the innovation–amplification–divergence model of evolution will continue to offer new examples of the co-opting of metabolic genes for drug resistance.
3 Summary, Challenges and Opportunities
Antibiotic resistance has proven to be inevitable. The reasons are now clear. The movements of genes horizontally through bacterial populations do not respect the artificial pathogen/nonpathogen boundary. Bacteria are ancient organisms that have adapted to virtually all environmental challenges on the planet. They live in environments dominated by small molecules and have evolved both specific and nonspecific mechanisms to evade or detoxify noxious compounds including antibiotics. In addition to these ancient resistance elements, contemporary resistance to any new drug can arise by gene mutation and amplification. From these processes new alleles can emerge that confer drug resistance. There are no irresistible antibiotics.
This fact explains our experience with 6 decades of antibiotic discovery and use. Nevertheless, despite the prevalence of resistance and the mechanisms that can rise to it, antibiotics remain foundational medicines for modern health care. A world without antibiotics would mean a world without many surgeries, transplants, cancer treatments, longevity of patients with cystic fibrosis, etc.; it would mean increased deaths of infants and the elderly by diseases now commonly tractable by antibiotics. Our clinical experience with antibiotics over the past decades and our growing understanding of the molecular basis of resistance must inform the discovery and use of future antibiotics.
The concept of a global antibiotic resistome offers an ecological view of resistance that can be applied in new drug development. The screening of candidate molecules based not only on their activity against traditional panels of pathogens but also nonpathogenic environmental bacteria will serve to identify “weak spots” in antibiotic structures and targets. The early identification of resistance elements can propel the development of diagnostics that will ensure epidemiological monitoring of resistance and the proper use of drugs at the bedside.
The discovery of an intrinsic resistome also offers a number of important opportunities for drug development. The systematic inactivation of nonessential genes, either through precise gene deletions (Baba et al. 2006) or by more traditional transposon mutant libraries, has offered an opportunity to examine the antibiotic susceptibility of bacteria upon inactivation of selected genes—so-called chemical synthetic interactions. By screens such libraries in E. coli (Liu et al. 2010) or P. aeruginosa (Breidenstein et al. 2008; Alvarez-Ortega et al. 2010), a large number of genes whose inactivation sensitizes bacteria to antibiotics have been identified. These offer a unique set of potential targets for the development of combination drugs to extend the efficacy of our current arsenal of antibiotics well into the future. Similarly, compounds that block resistance elements, already proven as successful clinical agents with the inhibitors of β-lactamases, can also be explored.
Whether such approaches will be adopted by physicians and the drug development industry remains to be seen. What is certain is that innovation and understanding of the molecule basis of the problem is vital. As noted by Joshua Lederberg: “Pitted against microbial genes, we have mainly our wits.”
References
Abraham EP, Chain E (1940) An enzyme from bacteria able to destroy penicillin. Nature 146:837
Allen HK, Moe LA et al (2009) Functional metagenomics reveals diverse beta-lactamases in a remote Alaskan soil. ISME J 3(2):243–51
Allwood AC, Walter MR et al (2006) Stromatolite reef from the Early Archaean era of Australia. Nature 441(7094):714–8
Alvarez-Ortega C, Wiegand I et al (2010) Genetic determinants involved in the susceptibility of Pseudomonas aeruginosa to beta-lactam antibiotics. Antimicrob Agents Chemother 54(10):4159–67
Andersson DI (2006) The biological cost of mutational antibiotic resistance: any practical conclusions? Curr Opin Microbiol 9(5):461–5
Andersson DI, Hughes D (2010) Antibiotic resistance and its cost: is it possible to reverse resistance? Nat Rev Microbiol 8(4):260–71
Baba T, Ara T et al (2006) Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Mol Syst Biol 2:0008
Bailey AM, Ivens A et al (2010) RamA, a member of the AraC/XylS family, influences both virulence and efflux in Salmonella enterica serovar Typhimurium. J Bacteriol 192(6):1607–16
Barlow M (2009) What antimicrobial resistance has taught us about horizontal gene transfer. Methods Mol Biol 532:397–411
Benveniste R, Davies J (1973) Aminoglycoside antibiotic-inactivating enzymes in actinomycetes similar to those present in clinical isolates of antibiotic-resistant bacteria. Proc Natl Acad Sci USA 70:2276–2280
Bergthorsson U, Andersson DI et al (2007) Ohno’s dilemma: evolution of new genes under continuous selection. Proc Natl Acad Sci USA 104(43):17004–9
Boucher HW, Talbot GH et al (2009) Bad bugs, no drugs: no ESKAPE! An update from the Infectious Diseases Society of America. Clin Infect Dis 48(1):1–12
Breidenstein EB, Khaira BK et al (2008) Complex ciprofloxacin resistome revealed by screening a Pseudomonas aeruginosa mutant library for altered susceptibility. Antimicrob Agents Chemother 52(12):4486–91
Bugg TDH, Wright GD et al (1991) Molecular basis for vancomycin resistance in Enterococcus faecium BM4147: biosynthesis of a depsipeptide peptidoglycan precursor by vancomycin resistance proteins VanH and VanA. Biochemistry 30:10408–10415
Canton R, Coque TM (2006) The CTX-M beta-lactamase pandemic. Curr Opin Microbiol 9(5):466–75
Cirz RT, O’Neill BM et al (2006) Defining the Pseudomonas aeruginosa SOS Response and Its Role in the Global Response to the Antibiotic Ciprofloxacin. J Bacteriol 188(20):7101–10
Courvalin P (2006) Vancomycin resistance in gram-positive cocci. Clin Infect Dis 42(Suppl 1):S25–34
Croucher NJ, Walker D et al (2009) Role of conjugative elements in the evolution of the multidrug-resistant pandemic clone Streptococcus pneumoniaeSpain23F ST81. J Bacteriol 191(5):1480–9
Cundliffe E (1989) How antibiotic-producing organisms avoid suicide. Annu Rev Microbiol 43:207–33
Cundliffe E, Bate N et al (2001) The tylosin-biosynthetic genes of Streptomyces fradiae. Antonie Van Leeuwenhoek 79(3–4):229–34
Daigle DM, McKay GA et al (1997) Inhibition of aminoglycoside antibiotic resistance enzymes by protein kinase inhibitors. J Biol Chem 272:24755–24758
Daigle DM, McKay GA et al (1998) Aminoglycoside phosphotransferases required for antibiotic resistance are also Serine protein kinases. Chem Biol 6:11–18
Dantas G, Sommer MO et al (2008) Bacteria subsisting on antibiotics. Science 320(5872):100–3
Davies J (1995) Vicious circles: looking back on resistance plasmids. Genetics 139(4):1465–8
D’Costa VM, McGrann KM et al (2006) Sampling the antibiotic resistome. Science 311(5759):374–7
D’Costa VM, Griffiths E et al (2007) Expanding the soil antibiotic resistome: exploring environmental diversity. Curr Opin Microbiol 10(5):481–9
De Pascale G, Wright GD (2010) Antibiotic resistance by enzyme inactivation: from mechanisms to solutions. Chembiochem 11(10):1325–34
Decousser JW, Poirel L et al (2001) Characterization of a chromosomally encoded extended-spectrum class A beta-lactamase from Kluyvera cryocrescens. Antimicrob Agents Chemother 45(12):3595–8
Donato JJ, Moe LA et al (2010) Metagenomics reveals antibiotic resistance genes encoding predicted bifunctional proteins in apple orchard soil. Appl Environ Microbiol 76:4396–4401
Dwyer DJ, Kohanski MA et al (2007) Gyrase inhibitors induce an oxidative damage cellular death pathway in Escherichia coli. Mol Syst Biol 3:91
Fajardo A, Martinez-Martin N et al (2008) The neglected intrinsic resistome of bacterial pathogens. PLoS One 3(2):e1619
Fischbach MA, Walsh CT (2009) Antibiotics for emerging pathogens. Science 325(5944):1089–93
Fleming A (1945) Penicillin. Nobel Prize Lecture http://nobelprize.org/nobel_prizes/medicine/laureates/1945/fleming-lecture.pdf
Folster JP, Johnson PJ et al (2009) MtrR modulates rpoH expression and levels of antimicrobial resistance in Neisseria gonorrhoeae. J Bacteriol 191(1):287–97
Fong DH, Lemke CT et al (2010) Structure of the antibiotic resistance factor spectinomycin phosphotransferase from Legionella pheumophila. J Biol Chem 285(13):9545–55
Freitas AR, Tedim AP et al (2010) Global spread of the hyl(Efm) colonization-virulence gene in megaplasmids of the Enterococcus faecium CC17 polyclonal subcluster. Antimicrob Agents Chemother 54(6):2660–5
Guardabassi L, Agerso Y (2006) Genes homologous to glycopeptide resistance vanA are widespread in soil microbial communities. FEMS Microbiol Lett 259(2):221–5
Guardabassi L, Perichon B et al (2005) Glycopeptide resistance vanA operons in Paenibacillus strains isolated from soil. Antimicrob Agents Chemother 49(10):4227–33
Gwynn MN, Portnoy A et al (2010) Challenges of antibacterial discovery revisited. Ann N Y Acad Sci 1213:5–19
Hon WC, McKay GA et al (1997) Structure of an enzyme required for aminoglycoside antibiotic resistance reveals homology to eukaryotic protein kinases. Cell 89(6):887–95
Hong HJ, Hutchings MI et al (2004) Characterization of an inducible vancomycin resistance system in Streptomyces coelicolor reveals a novel gene (vanK) required for drug resistance. Mol Microbiol 52(4):1107–21
Hooper DC (1999) Mechanisms of fluoroquinolone resistance. Drug Resist Updat 2(1):38–55
Hooper DC (2001) Mechanisms of action of antimicrobials: focus on fluoroquinolones. Clin Infect Dis 32(Suppl 1):S9–S15
Humeniuk C, Arlet G et al (2002) Beta-lactamases of Kluyvera ascorbata, probable progenitors of some plasmid-encoded CTX-M types. Antimicrob Agents Chemother 46(9):3045–9
Infectious Diseases Society of America (2010) The 10 × ‘20 Initiative: pursuing a global commitment to develop 10 new antibacterial drugs by 2020. Clin Infect Dis 50(8):1081–3
Karray F, Darbon E et al (2007) Organization of the biosynthetic gene cluster for the macrolide antibiotic spiramycin in Streptomyces ambofaciens. Microbiology 153(Pt 12):4111–22
Kaufmann BB, Hung DT (2010) The fast track to multidrug resistance. Mol Cell 37(3):297–8
Kelly JA, Dideberg O et al (1986) On the origin of bacterial resistance to penicillin: comparison of a beta-lactamase and a penicillin target. Science 231(4744):1429–31
Kohanski MA, DePristo MA et al (2010) Sublethal antibiotic treatment leads to multidrug resistance via radical-induced mutagenesis. Mol Cell 37(3):311–20
Koteva K, Hong HJ et al (2010) A vancomycin photoprobe identifies the histidine kinase VanSsc as a vancomycin receptor. Nat Chem Biol 6(5):327–9
Lang KS, Anderson JM et al (2010) Novel florfenicol and chloramphenicol resistance gene discovered in Alaskan soil using functional metagenomics. Appl Environ Microbiol 76:5321–5326
Laponogov I, Pan XS et al (2010) Structural basis of gate-DNA breakage and resealing by type II topoisomerases. PLoS One 5(6):e11338
Leclercq R, Derlot E et al (1988) Plasmid-mediated resistance to vancomycin and teicoplanin in Enterococcus faecium. N Engl J Med 319(3):157–61
Liu A, Tran L et al (2010) Antibiotic sensitivity profiles determined with an Escherichia coli gene knockout collection: generating an antibiotic bar code. Antimicrob Agents Chemother 54(4):1393–403
Maravic G (2004) Macrolide resistance based on the Erm-mediated rRNA methylation. Curr Drug Targets Infect Disord 4(3):193–202
Marshall CG, Lessard IA et al (1998) Glycopeptide antibiotic resistance genes in glycopeptide-producing organisms. Antimicrob Agents Chemother 42(9):2215–20
Massova I, Mobashery S (1998) Kinship and diversification of bacterial penicillin-binding proteins and beta-lactamases. Antimicrob Agents Chemother 42(1):1–17
Maveyraud L, Mourey L et al (1998) Structural basis for the clinical longevity of carbapenem antibiotics in the face of challenge by the common A beta-lactamasees from the antibiotic-resistnat bacteria. J Am Chem Soc 120:9748–9752
Morar M, Wright GD (2010) The genomic enzymology of antibiotic resistance. Annu Rev Genet 44:25–51
Morar M, Bhullar K et al (2009) Structure and mechanism of the lincosamide antibiotic adenylyltransferase LinB. Structure 17(12):1649–59
Mukhtar TA, Wright GD (2005) Streptogramins, oxazolidinones, and other inhibitors of bacterial protein synthesis. Chem Rev 105(2):529–42
Mwangi MM, Wu SW et al (2007) Tracking the in vivo evolution of multidrug resistance in Staphylococcus aureus by whole-genome sequencing. Proc Natl Acad Sci USA 104(22):9451–6
Nurizzo D, Shewry SC et al (2003) The crystal structure of aminoglycoside-3'-phosphotransferase-IIa, an enzyme responsible for antibiotic resistance. J Mol Biol 327(2):491–506
Oliynyk M, Samborskyy M et al (2007) Complete genome sequence of the erythromycin-producing bacterium Saccharopolyspora erythraea NRRL23338. Nat Biotechnol 25(4):447–453
Olson AB, Silverman M et al (2005) Identification of a progenitor of the CTX-M-9 group of extended-spectrum beta-lactamases from Kluyvera georgiana isolated in Guyana. Antimicrob Agents Chemother 49(5):2112–5
Peirano G, Pitout JD (2010) Molecular epidemiology of Escherichia coli producing CTX-M beta-lactamases: the worldwide emergence of clone ST131 O25:H4. Int J Antimicrob Agents 35(4):316–21
Piddock LJ (2006) Multidrug-resistance efflux pumps – not just for resistance. Nat Rev Microbiol 4(8):629–36
Poirel L, Kampfer P et al (2002) Chromosome-encoded Ambler class A beta-lactamase of Kluyvera georgiana, a probable progenitor of a subgroup of CTX-M extended-spectrum beta-lactamases. Antimicrob Agents Chemother 46(12):4038–40
Pomposiello PJ, Demple B (2000) Identification of SoxS-regulated genes in Salmonella enterica serovar typhimurium. J Bacteriol 182(1):23–9
Ricci V, Piddock LJ (2009) Ciprofloxacin selects for multidrug resistance in Salmonella enterica serovar Typhimurium mediated by at least two different pathways. J Antimicrob Chemother 63(5):909–16
Riesenfeld CS, Goodman RM et al (2004) Uncultured soil bacteria are a reservoir of new antibiotic resistance genes. Environ Microbiol 6(9):981–9
Robicsek A, Strahilevitz J et al (2006) Fluoroquinolone-modifying enzyme: a new adaptation of a common aminoglycoside acetyltransferase. Nat Med 12(1):83–8
Roy PH (1999) Horizontal transfer of genes in bacteria. Microbiol Today 26:168–170
Sandegren L, Andersson DI (2009) Bacterial gene amplification: implications for the evolution of antibiotic resistance. Nat Rev Microbiol 7(8):578–88
Shaw KJ, Miller N et al (2003) Comparison of the changes in global gene expression of Escherichia coli induced by four bactericidal agents. J Mol Microbiol Biotechnol 5(2):105–22
Sommer MO, Dantas G et al (2009) Functional characterization of the antibiotic resistance reservoir in the human microflora. Science 325(5944):1128–31
Soo VW, Hanson-Manful P et al (2011) From the cover: artificial gene amplification reveals an abundance of promiscuous resistance determinants in Escherichia coli. Proc Natl Acad Sci USA 108(4):1484–9
Spellberg B, Powers JH et al (2004) Trends in antimicrobial drug development: implications for the future. Clin Infect Dis 38(9):1279–86
Stogios PJ, Shakya T et al (2011) Structure and function of APH(4)-Ia, a hygromycin B resistance enzyme. J Biol Chem 286(3):1966–75
Strahilevitz J, Jacoby GA et al (2009) Plasmid-mediated quinolone resistance: a multifaceted threat. Clin Microbiol Rev 22(4):664–89
Toth M, Frase H et al (2010) Crystal structure and kinetic mechanism of aminoglycoside phosphotransferase-2″-IVa. Protein Sci 19(8):1565–76
Tu D, Blaha G et al (2005) Structures of MLSBK antibiotics bound to mutated large ribosomal subunits provide a structural explanation for resistance. Cell 121(2):257–70
Vetting MW, Park CH et al (2008) Mechanistic and structural analysis of aminoglycoside N-acetyltransferase AAC(6')-Ib and its bifunctional, fluoroquinolone-active AAC(6′)-Ib-cr variant. Biochemistry 47(37):9825–35
Walsh CT, Fisher SL et al (1996) Bacterial resistance to vancomycin: five genes and one missing hydrogen bond tell the story. Chem Biol 3:21–28
Ward SL, Hu Z et al (2004) Chalcomycin biosynthesis gene cluster from Streptomyces bikiniensis: novel features of an unusual ketolide produced through expression of the chm polyketide synthase in Streptomyces fradiae. Antimicrob Agents Chemother 48(12):4703–12
Waters B, Davies J (1997) Amino acid variation in the GyrA subunit of bacteria potentially associated with natural resistance to fluoroquinolone antibiotics. Antimicrob Agents Chemother 41(12):2766–9
Wellman CH, Osterloff PL et al (2003) Fragments of the earliest land plants. Nature 425(6955):282–5
White DG, Goldman JD et al (1997) Role of the acrAB locus in organic solvent tolerance mediated by expression of marA, soxS, or robA in Escherichia coli. J Bacteriol 179(19):6122–6
Whitman WB, Coleman DC et al (1998) Prokaryotes: the unseen majority. Proc Natl Acad Sci USA 95(12):6578–83
Williams KJ, Bax RP (2009) Challenges in developing new antibacterial drugs. Curr Opin Investig Drugs 10(2):157–63
Wright GD (2007) The antibiotic resistome: the nexus of chemical and genetic diversity. Nat Rev Microbiol 5(3):175–86
Wright GD (2010) The antibiotic resistome. Expert Opin Drug Disc 5:779–788
Wu Z, Wright GD et al (1995) Overexpression, purification, and characterization of VanX, a D-D-dipeptidase which is essential for vancomycin resistance in Enterococcus faecium BM4147. Biochemistry 34(8):2455–63
Young PG, Walanj R et al (2009) The crystal structures of substrate and nucleotide complexes of Enterococcus faecium aminoglycoside-2″-phosphotransferase-IIa [APH(2″)-IIa] provide insights into substrate selectivity in the APH(2″) subfamily. J Bacteriol 191(13):4133–43
Acknowledgements
Research in the author’s lab on antibiotic resistance is supported by a Canada Research Chair, the Canadian Institutes of Health Research and the Natural Sciences and Engineering Research Council of Canada.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Wright, G.D. (2012). The Origins of Antibiotic Resistance. In: Coates, A. (eds) Antibiotic Resistance. Handbook of Experimental Pharmacology, vol 211. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-28951-4_2
Download citation
DOI: https://doi.org/10.1007/978-3-642-28951-4_2
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-28950-7
Online ISBN: 978-3-642-28951-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)