
Abstract
This chapter discusses in detail individual honeybees fending off an enemy and the effects that can be rendered when whole colony defence is a coordinated, social effort. Enemies extend from viruses to bears. Whatever the predator, the nesting styles and nests of the bees afford a first line of defence. These are supplemented by various weapons, including stings, mandibles, legs and wings – variously used to grasp, pull, bite and sting opponents. The real defence game involves a panoply of colonial displays: hissing, shimmering, abdomen-shaking, body-twisting and rearing-up, which, if needs be, leads to socially organised defence: the awesome attacks which end in death by stinging and/or over-heating and asphyxiation by balling. The outstanding effectiveness of nest defence in honeybees is not so much based on the faculties of individual bees, but on the social coordination between thousands of individuals.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
17.1 Robbers and Thieves
Although showing considerable differences in many respects, all the Asian honeybee species share the basic feature with the western honeybee to nest in colonies containing several thousand individuals. The amassing of thousands of adult insects alone in a more or less stationary place constitutes an attractive target to predators. In the open-nesting dwarf honeybee, Apis florea, a bee mass of 6,300 individuals amounts to 0.144 kg of organic nutrition. This is by far exceeded by nests of the giant honeybee, A. dorsata, with 36,600 individuals, which are about fivefold heavier than A. florea, and adding up to a 4.3 kg biomass; while a colony of the cavity-nesting eastern honeybee, A. cerana, with 9,200 individuals still constitutes a considerable amount of 0.420 kg biomass.
This, however, is only a part of the attraction of honeybee colonies, as the cells of the wax combs, with areas of 396 cm2 in A. florea, 5,650 cm2 in A. cerana and 6,356 cm2 in A. dorsata, may contain 0.6 to 1.8 larvae and pupae per worker bee (in A. florea and A. cerana, respectively), which provide highly valuable proteins (Seeley et al. 1982; Dyer and Seeley 1991). Furthermore, the collection of considerable amounts of organic resources such as pollen and honey, with a high content of proteins or sugar, considerably enhances the nutritional value of a colony.
In view of the amount and variety of nutrition offered by a honeybee colony, it is not surprising that numerous potential predators, parasites, commensals and other pests set out to take a share of this concentrated high-value organic material. These include a wide array of taxa, notably mammals, birds and various arthropods that may target different parts of these commodities. Some only occasionally try to snatch small amounts, while others extract significant parts of their diet from this rich resource, or even show specific adaptations which facilitate overcoming the defensive measures of the honeybees.
The most notorious honey-lovers are bears (Helarctus malayanus, Melursus ursinus and Ursus thibetanus) and martens (Martes flavigula and M. foina) (Seeley et al. 1982; Thapa et al. 2000; Oldroyd and Wongsiri 2006). Bears may use their strong claws to break into nest cavities, or even attack the strongly defensive colonies of A. dorsata. They aim to feast predominantly on the combs that are filled with brood and honey, and thus their attacks are most destructive to the colonies. Martens also attack entire colonies, but have also been observed to prey on flying bees.
Occasionally, monkeys (Macaca sp.) have been observed to rob the nests of A. florea or A. cerana (Seeley et al. 1982; Thapa et al. 2000). There are few reliable data on the impact of predators on honeybee colonies, but tree shrews appear to be a notable threat to dwarf honeybee colonies in Thailand (Seeley et al. 1982). Other vertebrates of various taxa are occasionally observed to prey on honeybees, among these rats, honey badgers, foxes, lizards and toads (Caron 1978; Novogrodzki 1990). However, there is little doubt that exploitation by humans, which use all nest components, including the wax for nutrition and various other purposes, is a prominent threat to colonies of all honeybee species.
Various insect-preying birds, such as drongos and swifts, have been observed to opportunistically prey on flying honeybees (Ambrose 1978; Seeley 1994; Thapa and Wongsiri 2003). However, a range of birds has become regular predators on bees. Some display very specific behaviours. Bee-eaters, after snatching a forager bee, swish their beaks back and forth against a tree branch on which they are perching – obviously to remove the sting: a behaviour termed “bee-rubbing” (Fry 1984). In the blue-throated bee-eater (Merops virididis), it has been observed that the sting pheromone that is rubbed on the branch attracts other foragers, which then fall easy prey to the bird (Koeniger et al. 2010).
Honeybees may form a considerable part of the diet in bee-eaters, for example, the crop content of Merops orientalis and M. superciliosus could contain considerable proportions of up to 30% of A. florea (Fry 1984). Birds are mostly interested in capturing individual bees and rarely attack colonies. However, the blue-bearded bee-eater may provoke counter attacks in A. dorsata by passing close to the colonies and feed on individuals that pursue them to a perch (Kastberger and Sharma 2000). Similarly, flocks of M. orientalis were reported to launch coordinated attacks at A. dorsata nests at low ambient temperatures when the bees’ flying performance is impaired and they can be picked easily from the ground (Thapa et al. 2000). The highly specialised oriental honey buzzard (Pernis ptylorhynchos) also preys on nests of open-nesting honeybees, A. dorsata and A. laboriosa (Thapa and Wongsiri 2003; A. andreniformis – Koeniger et al. 2010), but differs as attacks are aimed at the combs to feed on brood and honey. During their specialised attacks, sometimes performed in pairs, they first arouse and mislead the defending worker force before attacking the combs. Honeyguides (Indicatoridae) are specialised wax feeders, but mainly feed on abandoned nests; the two species occurring in Asia do not constitute a major threat to honeybees (Oldroyd and Wongsiri 2006).
Besides vertebrates, other arthropods impose considerable predation pressure. A variety of different arthropods, such as mantids, crab spiders, orb-weavers and assassin bugs, have been observed to be opportunistic feeders (Seeley et al. 1982; Fuchs personal observation), causing no serious harm to colonies. In contrast, from the related Hymenoptera, two equally social groups, wasps and ants, are of major impact, either by posing a constant drain on the bees’ work force or by attacking and robbing entire colonies. In particular, the larger vespine wasps of the genus Vespa play a major role as predators of Asian honeybees (Matsuura 1988); an array of hornets which attack Asian bees are listed in Oldroyd and Wongsiri (2006). Some of the giant wasps specialise on social bees and are outstanding in their effectiveness to deplete entire colonies. Wasps commonly prey on individual flying foragers during foraging. Quite commonly, wasps catch departing or homing foragers close to the colonies (“bee-hawking”), and open-nesting dwarf and giant honeybees as well as cavity-nesting bees are targeted (Seeley et al. 1982; Ono et al. 1987; Tan et al. 2007; Kastberger et al. 2008).
Some wasps hawk honeybees at cavity-nesting colonies (Vespa multimaculata – Koeniger et al. 1996; V. velutina – Shah and Shah 1991) and employ a specialised strategy in which they hover in front of a colony entrance facing outward to catch returning foragers (“goal-keeping behaviour”, Koeniger et al. 1996). One V. multimaculata caught 14 A. nuluensis within 12 h, and V. velutina caught 17.8 in 151 (11.2%) hawking attempts (Tan et al. 2007), which could deplete a colony by 20–30% (Sakagami and Akahira 1960).
V. affinis has been observed to prey on drones in drone congregation areas, with the drones confusing them for a queen (Koeniger et al. 1994; cf. Chap. 8). Hornet attacks are not restricted to single flying individuals, but may even be fatal for a colony. A few V. tropica wasps can overpower an A. florea colony within 3 h by successively seizing and killing nest workers and gradually depleting the workforce (Seeley et al. 1982). By recruiting nestmates, V. mandarinia hornets are able to mass-attack entire A. cerana colonies and, having killed the bees, raid pupae and larvae. In order to focus their attacks, they use pheromones to mark the besieged colonies (Ono et al. 1995).
Ants are an almost omnipresent danger to honeybee colonies and may take anything for nutrition: live and dead bees, larvae, pupae and honey (Akratanakul 1987). The predominant ant species to endanger bees are the widespread tree-living weaver ants. In particular, Oecophylla smaragdina is a major threat to colonies of dwarf honeybees, who share the same habitat, but may also occasionally drive cavity-nesting A. cerana from their nests (Dyer 1991). These ants try to seize and pull down flying bees from a perch and several ants may be needed to overcome a bee. Other species, such as Monomorium indicum, Diacamma rugosum and Pheidologeton sp., were observed to molest A. cerana colonies, but without causing apparent harm. Few ant species, such as Dolichoderus bituberculatus, attack A. dorsata colonies (Seeley et al. 1982).
Only some of the most obvious and well documented examples have been mentioned, and without any doubt there are numerous other species which occasionally attack single honeybees or colonies. However, even this short overview may substantiate the threat from thieves and robbers who are trying to get their share of the spoils. However, bees are not helplessly exposed to these attacks, but have developed various means to meet such challenges. Due to strong selection pressure, these defence measures have taken various forms and led to sophisticated counter-strategies that are matched for the kind of predators. These will be the focus of the following sections.
Besides predators, honeybee colonies are also exploited by parasites, commensals, macroparasites and microparasites. Although these also include interesting and highly sophisticated interactions, they are not dealt with here. Bee mites and bee diseases are however reviewed in separate chapters of this book (cf. Chaps. 15 and 16).
17.2 Nesting and Defence Styles
It has been argued that predation has been a major selective force that is shaping the lifestyles of honeybees and, indeed, the striking differences among the Asian honeybees, dwarf, giant and cavity-nesting species, provide excellent insight into the different basic strategies to counter predation (Seeley et al. 1982; Dyer 1991; Dyer and Seeley 1991).
When facing an enemy, there are two opposing primary strategies: to retreat or to (counter-) attack. In honeybees, both are found – with differing emphasis between colonies (Kastberger et al. 2009a) and between the different honeybee species, taking very diverse forms in relation to varieties in predation patterns. Retreat and attack strategies may also occur in combination or mixed forms and may be switched due to different intensities of the predators’ attacks. The propensity for attack or retreat strategies is highly dependent on nest structures and placements; the basic relationships between the ease of detection, approach and consumption have been outlined by Dyer (1991).
In the cavity-nesting species, the primary defence measure is to live in cavities. These offer strong protection by restricting accessibility to the nest and at the same time constitute a limited and valuable resource in themselves, to be actively defended if in danger. Cavity-nesting is assumed to be a derived “primitive” trait (Lindauer 1956), but because this is shared with many social Hymenoptera, this view has been questioned (Koeniger 1976; cf. Chap. 2).
Open-nesting bees are exposed to environmental stresses, such as rain, wind and temperature changes, and need to defend the entire nest surface against predators. It needs to be emphasised, however, that cavity- and open-nesting are not entirely separate modes, as during a cavity-nesting bees’ lifecycle, reproductive or migrating swarms interrupt the cavity-sheltered mode, and will require some defence adaptations in order to reduce vulnerability during this open-nesting stage. Moreover, A. cerana occasionally creates open nests if no cavities are available. Matsuura (2003) – (cited from Sugahara and Sakamoto 2009) – reported that about 10% of A. cerana colonies in Japan are open-nesting.
In the open-nesting bees, the dwarf honeybees rely predominantly on retreat strategies, such as hiding by placing their small nests in bushes and preferring cover by leaves, with little emphasis on attack options. If challenged, they tend to abandon their nests. In contrast, the open-nesting giant honeybees prefer inaccessible, but often conspicuous, nesting sites in high trees or cliffs. In case of any attacks, they resort to exceptionally strong counter-attacks. This dichotomy in the predominant defence strategy may be related to body size (Dyer 1991), but may also be a selective force driving worker body sizes into the disparate “dwarf” and “giant” directions (Koeniger et al. 2010; cf. Chap. 2).
17.3 Bee Weapons: Stings, Mandibles, Legs and Wings
On an individual level, honeybees possess limited means and actions for defence. As for other insects, mandibles can be used for biting; however, honeybee mandibles are rather a multi-purpose tool that is also used for a range of other tasks and not specifically shaped for fighting. Nevertheless, they are useful for grabbing, and small predators such as ants may be crushed. Furthermore, legs may be used for grabbing, and thus aiding their defence actions. Wings can also be used to produce air blasts, which can be useful in defence actions to blow away intruders (Seeley et al. 1982; Dyer 1991; Yang et al. 2010).
These are, however, rather general and unimpressive faculties which are by far outclassed by the one highly sophisticated and specialised defence weapon that bees are famous for, which is the sting. As in the western honeybee, stinging is the main defence against vertebrate predators in all Asian honeybee species. All possess barbed stingers which they stab into vertebrate skin where they become anchored, while the bee is detached from the sting apparatus by autotomy. Without the bee, the sting is less likely to be removed by the victim and will continue to inject poison that is driven by movements of the detached sting apparatus, containing a nerve ganglion and a sophisticated structure of muscles and sclerotised plates.
There are no complete investigations on the sting apparatus for all Asian species, but some studies cover A. cerana, A. florea and A. dorsata as examples for cavity-nesting, dwarf and giant honeybees (Weiss 1978; Jayasvati 1989). From this limited evidence, it appears that there are only minor differences in the general morphology of the stinging apparatus. However, the size of the sting apparatus varies approximately in proportion to body size differences. Lancet length is 2.10 mm, 1.55 mm and 1.06 mm in A. dorsata, A. cerana and A. florea, respectively (Ramya and Rajagopal 2008). There is some variation in the numbers and expression of barbs on the stinger sheath and lancet, which appear to give better fixing of the stings in the open-nesting bees, A. florea and A. dorsata, in comparison to the cavity-nesting bee, A. cerana.
Although there is a surprising paucity of studies, the composition of sting poison is apparently fairly similar in all of the species investigated, with the most prominent active ingredients being the peptide mellittin and phospholipase. However, some minor components may differ (Natzir et al. 1999) from A. mellifera. The amino acid sequences of melittin show differences between A. cerana, A. dorsata and A. florea in five positions, which are of relevance in phylogenetic, but not in functional considerations (Kreil 1975).
Honeybee venom toxicity is predominantly aimed at vertebrates (Schmidt 1990) and is apparently similar among honeybee species. It does not differ if the bee venoms of A. dorsata, A. florea and A. cerana are applied to mice (~3 mg/kg mice). However, the amount of venom per worker bee is much higher in A. dorsata as compared to A. cerana or A. florea (218 μg, 43 μg, 27 μg, respectively) – even exceeding that of A. mellifera (138 μg). The percentage of poison to total body weight is 0.65%, 0.28% and 0.28%, respectively, and the higher percentage in A. dorsata is in line with the high defensiveness of this giant honeybee (Schmidt 1995). It has been estimated that the LD50 of A. mellifera bee stings for non-hypersensitive humans are between 500 and 1,500 stings (Vetter and Visscher 1998), which would be equal to about 600 stings from A. dorsata, 3,200 from A. cerana and 5,100 from A. florea.
17.4 Socially Organised Defence
The stunning and outstanding effectiveness of nest defence in honeybees is not so much based on the faculties of individual bees, as on the social coordination between thousands of individuals. Biting, grabbing and wing-buzzing of a single bee are hardly effectual, but the combined efforts of many individuals add up to astounding performances. In particular, social coordination opens up new possibilities on a higher level of colonial, behavioural and structural patterns that are not available to an individual. The great fascination of honeybee defence lies within this social dimension, and the Asian Apis species offer a unique opportunity to study the variations of defence patterns and special adaptations in response to the specific predator environments to which they are exposed.
17.4.1 Stinging
Even stinging, for which individual foraging bees might already be respected, achieves entirely new dimensions of awe if embedded within the patterns of a socially coordinated attack. In A. dorsata, such attacks are characterised by signs of unrest in the colonies if disturbed, which build up until entire squads of bees drop from the free-hanging colonies in several defence waves. The waves of bees will follow retreating intruders up to several kilometres or continue to search the vicinity of the nest for hours until they return to the nest (Lindauer 1956; Morse and Laigo 1969).
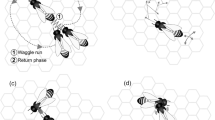
Such counter-attacks are likely to follow the general scheme for bee attacks as proposed by Breed et al. (2004), which describes the basic behavioural sequence for socially organised defence (Fig. 17.1). The typical chain of increasing defence responses would follow a sequence of primary perception and orientation of the target by a single bee, leading to either discrimination or identification. If this confirms an enemy situation, the bee is further alerted, approaches the target and, after secondary discrimination or identification based on close contact cues, may attack. Threat behaviour may provide an alternative; indeed an astounding portion of honeybee behaviour is shaped to frighten an enemy away, instead of engaging in outright attacks. At any of these steps, bees may disengage if stimulus intensity does not exceed the continuation thresholds.

Organisation of social defence: Pathways of perception and interactions leading to social attack in honeybees (from Breed et al. 2004)
This sequence alone would be apt to ensure a basic pattern of effective defence organisation. Assuming different behavioural thresholds, it could even explain some modulation of total defence efforts to a threat in a social context, based on the independent actions of the individuals. However, the main boost in the system’s responsiveness comes from its built-in feedback loops. By alerting and recruiting, further nestmates join the sequence – possibly even skipping the initial steps of the sequence – and an entire colony might catapult into a completely different behavioural state. In this, the workforce now acts as a coherent unit that is governed by mechanisms of social regulation, opening up the domain of integrated, higher-level actions way beyond the scope of the individual. These actions in turn are subject to selection, to be shaped into the organised group defence for which bees are respected and feared.
Among others, Morse and Laigo (1969) give some impressive accounts for stinging defence actions in A. dorsata, which they describe as the “most ferocious stinging insect on earth”. Within a certain threshold, colonies may be approached and cautiously investigated, but when disturbance by animals is mounting, social chain reactions lead fairly suddenly to integrated attack patterns. Bees from the surface start to move down to the lower edge of the comb, from where they drop into the air in successive defence waves and seek the disturber in bands of flying defenders – actively seeking open areas between the plants in clouds of bees to detect intruders.
The most prominent feedback loop is activation by alarm pheromones. Although mandibular pheromones play some role, the main alarm pheromones are associated with the sting sheath of the sting apparatus (Cassier et al. 1994). An exception is A. laboriosa, which apparently relies on a mandibular pheromone for alarming (Blum et al. 2000). In almost all of the Asian bees investigated thus far, the major active constituent is isopentyl-acetate (Morse et al. 1967; Koeniger et al. 1979; cf. Chap. 9). This volatile substance is released when the sting shaft is exposed and alerts other bees to an enemy. Thus, attacking bees, if returning to the nest, will trigger off further guards to join in defence, until up to 10% of the total workforce might become airborne. Koeniger (1975) reported that between 50 and 5,000 workers of A. dorsata were activated by single alarmed workers who were returning to the colony.
The volatile isopentyl-acetate, evaporating within a few minutes, is particularly appropriate as a fast-acting colony defence activator. By autotomy, the barbed sting of successful defenders becomes fixed in a vertebrate enemy’s skin, and the exuded alarm pheromone then guides others to these vulnerable spots. Interestingly, in two open-nesting species, A. dorsata and A. florea, a second active component, 2-decen-1-yl-acetate, is present (Veith et al. 1978; Koeniger et al. 1979), which is far less volatile and seemingly acts as a more persistent enemy-marker pheromone in these bees. Following this scent, fleeing predators and humans are reported to be pursued by A. dorsata for several hundred metres or even kilometres (Lindauer 1956; Morse and Laigo 1969). At 50 m distance from a nest, a man stung by a single bee was attacked by 500 more, and the numbers of attackers increased even further at 1.6 km distance from the nest, decreasing slowly only at a distance of 3.2 km away (Koeniger 1975). The pheromonal marking is so persistent that bees recognise the smell even the next day (Lindauer 1956).
A. dorsata has 10–20 times more isopentyl-acetate in its sting sheath than other bees (Morse et al. 1967; Koeniger et al. 1979). This implies a very strong feedback loop in the dynamic of the social defence organisation of this bee. It also underlines how colony-level characteristics of defence reactions may be shaped by simple variables, such as the amount of alarm substances in single bees; in this case towards a very strong dynamic that is tuned for high responsiveness, due to a high amplification factor. The resulting integrated social attacks pose a serious threat to vertebrate intruders, including humans (Morse and Laigo 1969). Lindauer (1956) reported the deaths of three water buffalos in Sri Lanka due to an attack by A. dorsata. Kastberger and Sharma (2000) recorded about 700–800 worker bees taking part in attacks against a blue-bearded bee-eater (Nyctyornis athertoni) that was approaching a nest of A. dorsata. Even highly specialised bee predators, such as the honey buzzard (Pernis ptylorhynchos) might be stung to death by A. dorsata in spite of protective feather shielding on its head (Thapa and Wongsiri 2003). However, a main effect likely comes from scaring away enemies through the coordinated attacks of hundreds of individuals, even if only a few stings are actually administered (Koeniger 1975). In fact, threatening is an alternative because lost bees are costly to the colony, and A. cerana is known for its reluctant use of stinging (Sakagami and Akahira 1960). Threatening is fortified by the continuous mock-attacking and biting of moribund A. mellifera bees, even after stinging, until their death due to sting loss injury (Cunard and Breed 1998), as has similarly been observed in A. dorsata by Koeniger (1975).
In contrast to the highly defensive open-nesting A. dorsata (Fig. 17.2), the stinging defence in cavity-nesting species, such as A. cerana, is far less pronounced. Living in cavities offers the option of retreating as a typical first response to slight disturbances (Seeley et al. 1982), and bees cease flights due to a predator’s approach (Tan et al. 2007). Only if a colony is massively disturbed will it respond by stinging attacks. However, due to comparatively undeveloped sting barbs and characteristic “revolving” stinging behaviour, A. cerana avoids autotomy and the cost of the stinging bee’s death (Sakagami and Akahira 1960), thus simultaneously providing a comparatively low feedback loop of sting pheromones in the colony’s attack dynamic. Further, absconding is frequent if colonies are often disturbed (Sakagami 1960; Punchihewa 1994). Apparently, in Borneo, A. koschevnikovi is even less defensive as compared to A. cerana (Mathew and Mathew 1988), but there is very little known about defensiveness in the other cavity-nesting bees of Asia: A. nuluensis and A. nigrocincta.

Workers blocking the nest attachment line of the colony to the tree trunk in the open-nesting bees, Apis dorsata, so as to prevent access by ground-walking predators (photo: N. Koeniger)
Although the open-nesting dwarf honeybees predominantly rely on retreat strategies such as hiding and absconding (Seeley et al. 1982), they are nevertheless capable of issuing decisive stinging counter-attacks. A. florea is fairly docile if not provoked, but will first attack vertebrates in small groups of workers, which are later re-enforced by hundreds of workers if more persistently disturbed (Seeley et al. 1982). Stings fix to the skin with their pronounced barbs and provoke further stinging through the stimulus 2-decen-1-yl-acetate: the “enemy-marking pheromone” (Koeniger et al. 1979). Their stings are relatively painless to humans and may not repel decisive attacks by tree shrews, monkeys or humans (Seeley et al. 1982). A. andreniformis is far more prone to attacking and may possess a different alarm pheromone (Wongsiri et al. 1996).
17.4.2 Grasping, Pulling, Biting and Other Means
As with other insects, bees have mandibles, legs and wings as the main mechanical tools for dealing with the world, and which may also be used for defence. Again, the comparative ineffectiveness of these actions if employed as individuals is multiplied within the social context and new defensive means emerge in their combined use. Stinging is the predominant weapon in vertebrate defence, though A. cerana may use their mandibles also for biting (Sakagami 1960). Stinging, however, is of limited value in defence against a wide range of arthropod predators (Schmidt 1995). Here, bees rely on the more conventional weapons, gaining advantage from their socially coordinated exertion.
There is a wide range of solitary insects and other arthropods that are occasionally trying to prey on bees or their stores, and bees have little difficulty in deterring them due to their higher numbers. The more serious threats come from other social insects, namely ants and wasps, which, like bees, can make use of the advantages of sociability, join efforts and in particular may recruit nestmates for reinforcement to attack in great numbers. While vertebrate attacks are often deleterious to bee colonies, ants and wasps are a less conspicuous, but no less serious, threat to honeybees – only rarely destroying the colonies, but causing a constant drain. In dealing with these predators, Asian bees have developed highly refined social actions so as to keep at par with the dangers.
The prominent species of ants that are endangering Asian bees is the weaver ant, Oecophylla smaragdina (Seeley et al. 1982; Duangphakdee et al. 2005, 2009), which shares the tree environment, particularly with dwarf honeybees. Usually, these ants perch along the flight path of the bees, trying to grab single forager bees with their mandibles when leaving or returning. The small A. florea forager bees have little chance of breaking free if grabbed by a single O. smaragdina worker. However, the bigger A. cerana falls victim to the ants only if there are several of them grabbing a worker in a joint effort. In addition, several A. cerana workers can overcome an ant, grabbing the legs and tearing them apart; two A. cerana may suffice to kill a weaver ant. A. florea can also overcome O. smaragdina should they happen to fall on the nest, where they are mobbed by 10–15 workers and crushed (Seeley et al. 1982).
Ants and bees are effectively matched in fighting abilities, using similar means. Encounters are mainly decided by size differences and the relative numbers of opponents engaged. However, bees have one advantage over ants: that they can fly. This allows them to use the air route for trafficking from and to the nest, independent of the walking routes that ants are restricted to. Thus they are able to rely predominantly on isolating their nests from the ant environment, thereby minimising actual conflicts in spite of a situation of constant siege. In cavity-nesting bees, if any openings are sealed, the only remaining point of possible attack is at the flight entrance. This remaining conflict area can be structurally improved by carefully smoothing the surface at the nest entrance so as to weaken the grip of the ants and diminish their fighting chances. For defence, the entrance is staffed by vigilant guard bees, facing outward to ward off intruding ants with their mandibles. In addition, specific behaviours were observed making use of the wings. Guards may turn around and use air blasts to deter the ants or hover in buzzing flights above attackers to distract them (Seeley et al. 1982).
Most of the nest surface of the free-hanging nests of open-nesting bees is completely safe from ants, and bees can enter and leave without coming close to any ants. However, nests are accessible where they are attached to the substrate. In the case of the giant honeybees, this may be in the form of an extended line fixed to overhanging cliffs or the lower side of big branches. At the boundary, worker bees line up side by side in several rows, forming a permanent barrier effective against any walking insects (Seeley et al. 1982). Due to their preferential nesting in high and remote places, ant attacks are not very intense, but have been observed to occur, for example, by the arboreal ant species, Dolichoderus tuberculata and Polyrhachis armata. However, they seem to be easily kept at bay by giant honeybees (Seeley et al. 1982).
In contrast, dwarf honeybees nesting in shrubs formed by bushes or small trees live in the midst of “ant land”. In Thailand, 37% of 76 A. florea nests were under attack from weaver ants (Seeley et al. 1982). As nests completely enclose the thin branches to which they are attached at the upper side of the comb, the accessible nest boundary is minimised and restricted to the outlet of the branches. Here, dwarf bee workers form similar permanent defence lines, as do giant honeybees, guarding this potential way of access. In order to strengthen these possible access points, dwarf bees create a further barrier by covering the branches with rings of sticky plant resins and deliberately use the material to spread it over opponents so as to immobilise them (Seeley et al. 1982; cf. Chap. 6). This resin substance of A. florea and A. andreniformis has a strong repellent effect on O. smaragdina ants (Duangphakdee et al. 2005, 2009). A. florea eagerly maintains the integrity of this defence line and takes great care to maintain these barriers, even fortifying them if challenged by ants (Duangphakdee et al. 2005, 2009). They react immediately if the barrier is bridged by leaves, either falling on the nest or otherwise touching the nest. In this case, they persistently push with their heads and mandibles, either to remove the object or at least to keep it at some distance, and specialised workers take turns over many hours (SenSarma et al. 2000; Fig. 17.3).

Defending nest integrity in Apis florea by pushing off bridges. Head-pushing is performed by a specialised group of workers, which take turns lasting a minute to almost an hour (photo: S. Fuchs)
Dwarf honeybee nests are challenged from all sides by ants and other insects trying to snatch single bees from nearby perching places on leaves or twigs. Dwarf bees are therefore extremely watchful in their immediate environment and attack such perching predators by trying to grab them with their legs and pull them onto the nest, where they are then mobbed and overcome by the crowd. In order to bridge the distance, ants may pile up one on top of the other to form bee ladders, which eventually collapse due to their weight if piling up too high (Fig. 17.4). Thus, Asian honeybees try to minimise conflicts with ants by isolating terrestrial access to the nest as far as possible, by creating structural barriers and forming ordered defence lines at accessible points – decisively attacking any ant that is coming close, which could otherwise recruit other ants if successful.

Apis florea workers piling up, one on top of the other, to reach an assassin bug (Reduviidae), perched from the microphone that is fixed above the colony, to snatch single workers. Bee heaps become unstable if too high and collapse (photo: S. Fuchs)
The second arthropod group that seriously threaten Asian bees are large vespine wasps. In contrast to ants, wasps also attack on the wing and cannot be as easily excluded by the blocking of walk-ways. Furthermore, some of these wasps outclass the bees that they prey on, in size, and are virtually sting-proof due to the strong armouring of their body surfaces. Relying on animal protein to raise their brood, together with communication systems for recruiting nestmates, these groups are true “wolves” for bees.
Commonly, wasps prey on flying foragers and there is little possibility of a single bee defending itself once caught. However, wasps frequently try to catch leaving or returning foragers at the colony. Bees try to decrease the risk of being caught. When one or two V. velutina wasps are bee-hawking in front of a colony in the “goal-keeper” manner (Koeniger et al. 1996), flight activity decreases and almost ceases completely after 12 min of wasp exposure. In addition, returning foragers make a rapid straight-line flight path into the flight entrance, nearly halving their time of exposure to the wasp.
Overcoming an entire colony is more difficult. Bees may use grabbing and mobbing in defence, overcoming even significantly bigger wasps due to their numbers. In A. cerana, the numbers of guards increased approximately eightfold in response to V. velutina predation and guards formed tongue-like groups covering the area below the entrance in order to increase their chances to grab and ball a wasp (Tan et al. 2007). Wasps thus avoid approaching too close to a hive entrance or an open-nesting bees’ protective curtain. Nevertheless, wasps might still continuously kill individual bees until a colony is weakened and no longer able to defend itself. V. tropica was observed in bands of two or three, successively killing single workers perching above an A. florea colony. A. florea was unsuccessful in mobbing wasps, which, due to their size, could retreat in spite of 12 workers grabbing them. On the second day, the rest of the bees absconded, leaving the brood to the wasps.
Not all bees are as defenceless to wasp attacks. In Japan, A. cerana colonies are besieged by the enormous Vespa mandarinia japonica, reaching body sizes of 50 mm in length, about five to six times that of the worker bees. On top of this physical superiority of the wasps, they mark colonies by a scent from their abdominal Van der Vecht gland so as to attract further wasps to strengthen the attack force. A. cerana recognises this scent and increases the numbers of guard bees at the colony entrance. Attacked by a V. mandarinia wasp, the guard bees first retreat into the nest. If the wasp tries to follow, it meets a superior force of defenders, intensely mobbing the wasp and entirely enclosing it in a ball of bees containing about 500 workers – killing the wasp. Interestingly, this killing is not by stinging, because the stings cannot effectively penetrate the sclerotised armouring of the wasps. Instead, the bees apparently create such high temperatures within the ball that the wasps are killed by heat. Temperatures within the balls were 47°C, still below the upper thermal limit of 48–50°C in A. cerana, but higher than the thermal limit of 44–46°C in V. mandarinia (Ono et al. 1995).
Similar heat-balling in A. cerana japonica was described against V. simillima xanthoptera, where bees generated temperatures of 46°C within the balls, while the upper thermal limit of the wasps was 45–47°C (Ono et al. 1987). In China, when A. cerana is defending against V. velutina, about 30 workers ball the wasps within less than 1 s, and temperatures within the balls reach 45°C within 3 min, reaching the upper lethal thermal of 45°C for the wasps, while A. cerana survives at 50°C (Tan et al. 2005). Abrol (2006) reported that A. cerana killed about 12% of attacking V. magnifica by balling, with temperatures ranging between 45 and 46.8°C within balls.
Heat-balling is not restricted to A. cerana and was also documented in A. nuluensis (Koeniger et al. 1996). Tethered V. multimaculata brought into contact with guard bees at the hive entrance were rapidly covered by approximately 40 bees. Inner-ball temperature was not measured, but the balls felt warm on the hand (>37°C), and the bees showed no stinging behaviour. Heat-balling may also occur in open-nesting bees. Kastberger and Stachl (2003) documented that A. dorsata will grasp a tethered wasp as soon as it is brought into contact with the bees on the nest surface. The wasp was balled and killed within 10 min, during which time the recorded inner-ball temperatures were between 45 and 48°C. Heat-balling is likely to occur in most of the other Asian Apis species, although documentation is incomplete.
However, it is not entirely clear if heat is the only factor contributing to the death of the wasps within the balls. Abrol (2006) reported that during balling, A. cerana guards would seize, bite, pull, hold and sting V. magnifica and reported some degree of mutilation, particularly of the wings, in the killed wasps. Stinging may not be successful against the big V. mandarinia, but successful stinging was found in V. simillima that had been balled by A. mellifera (Ono et al. 1987).
Sugahara and Sakamoto (2009) determined a further factor contributing to the death of balled wasps. In their study, they recorded that V. mandarinia could survive temperatures of 47°C for 10 min, while inner-ball temperatures remained at temperatures below 45.9°C, insufficient to kill the wasps. However, they found that the CO2 level within balls would increase to 3.6%, at the same time lowering the wasp’s lethal thermal limit by 2°C, without affecting the upper thermal limit of the bees. They thus concluded that, in combination with heat, CO2 is a major factor in killing wasps.
A recent reinvestigation of balling behaviour in the Cyprian subspecies of A. mellifera defending against V. orientalis supported the idea that temperature within the balls would be inadequate to explain the killing success. Inner-ball temperatures in A. mellifera are 1.5–3.3°C lower than in A. cerana (Ono et al. 1987; Tan et al. 2005). In A. m. cypriaca, the temperature within balls was 44°C, clearly below the upper lethal temperature of the wasps (approximately 50°C) which is about that of the worker bees. In a detailed examination, it was found that the killing of the wasps was accomplished by still another method: the balling bees would asphyxiate the hornets by imposing pressure on the abdomen, interfering with the wasp’s breathing movements and blocking their spiracles (Papachristoforou et al. 2007).
Thus heat might be a less prominent factor in A. mellifera wasp-balling as compared to A. cerana. It appears that thermal insulation and heating up, intoxication by increased CO2 levels and possibly asphyxiation by pressure on the abdomen all contribute to the killing of wasps. Additional factors involved throughout the genus still need to be determined.
Heat generation during fights is an obvious by-product of agitation, accompanying any event in which an intruder is mobbed by a group of nest defenders. Under most circumstances, this would add to the stress of the opponent during attack. From simple mobbing of small predators to dense balling, turning on heat and increasing CO2 levels provide outstanding examples of how the social dimension opens up completely new means for defending, which are then perfected in their own right.
17.5 Colonial Displays
Coordinated stinging defence and balling are socially coordinated behaviours in which hundreds or thousands of individuals may participate. However, Asian bees have particularly impressed observers by conspicuous defence-related behavioural coordination in a much grander style, involving high proportions of the workers in activities, simultaneously giving the appearance that the entire colony is a coherent acting unit.
Since Roepke (1930) first mentioned coordinated waves of excitation moving over the combs of A. dorsata, there have been various accounts of conspicuous coordinated displays over the years. On closer examination, these proved to consist of several fairly distinct behavioural patterns, all sharing coordination between neighboured workers, but separated by different motor patterns, conditions of release and some degree of species-specific variation. The main dividing lines between these behaviours are the releasing stimuli, either by substrate vibration and air movement or by the perception of visual object movements, the vertical or transverse direction of their body axis movement, and the different relation to attack–retreat strategies. At the same time, some of these behaviours are apparently interrelated or show transitions according to the functional context, which still need to be clarified in more detail (cf. Chap. 18).
In early observations, variations in intensity of disturbance and unaccounted mixtures of different stimuli created variable outcomes and overlaps between these behaviour patterns. Fairly commonly, they were jointly addressed as “shimmering” (Butler 1954), and in past and recent literature, the different behaviour patterns were frequently misnamed or confused. By now, with more and more precise investigations available, the relations and functions of these behaviours can be outlined more clearly. Although knowledge is still incomplete, we will try here to summarise their characteristics, tentative functions and possible relationships. To date, at least four different behaviours can be distinguished: “hissing”, “rearing-up”, “abdomen-shaking” and “body-twisting”. “Hissing” and “rearing-up” are both elicited by mechanical stimulation and constitute elements of defence against vertebrates. In contrast, “abdomen-shaking” and “body-twisting” are elicited exclusively by moving visual stimuli and are closely related subforms of “shimmering” behaviour, originally described by Butler (1954), serving to deter various flying predators, predominantly wasps.
17.5.1 Hissing Behaviour
Hissing behaviour was first described in A. cerana by Sakagami (1960) but erroneously likened to “shimmering” earlier described by Butler (1954) in A. dorsata. Later, the behaviour was investigated in A. cerana in more detail by Koeniger and Fuchs (1973) and was termed “hissing behaviour”. In this behaviour, individual A. cerana workers, in a slight and inconspicuous movement, draw their entire body closer to the substrate, either to the comb or the bees they are sitting on. During this movement they brace their wings slightly, by about 45°, and produce a short 0.5–1 s burst of wing vibration sound, consisting of a 0.5–6 kHz energy spectrum lacking harmonic structure. This behaviour is contagious, such that neighbouring workers will be induced to perform the same behaviour within a short time delay. In effect, a hissing wave is produced which spreads at approximately 25 cm/s among the bees on a comb and may wander over the edge to the bees on the opposite side of the comb or even spread to other combs if these are connected by bee bridges. The excitation can be propagated by short-range air movements, without direct bee-to-bee contact or without a connecting comb. The joint activity of the bees produces the characteristic hiss, lasting until the hissing wave is exhausted. Series of more than 10 successive hisses may follow each other in 3–5 s intervals (Sakagami 1960), which points to an individual latency of hissing as a prerequisite for wave-like propagation.
Hissing behaviour is most readily released by substrate vibrations, such as by knocking or poking a colony or tapping the supporting branch of a open-nesting colony, and also by locally applied gusts of air. Substrate vibrations which are effective are in a low-frequency range, and sine waves exceeding 600 Hz, soon become ineffective (Fuchs and Koeniger 1974). In contrast, moving visual stimuli are not effective at all. Thus, the array of stimuli points to reactions that are tuned to approaching vertebrates, rather than flying predators. Hissing was mentioned to occur in A. florea and A. cerana by Seeley et al. (1982), was investigated in more detail in A. florea by SenSarma et al. (2002) and is widespread in the Apis.
Colony displays may serve to create the impression of the entire nest being a coherent organism rather than an aggregation of thousands individuals, and integrated actions such as “shimmering” waves can underline this by mimicking total movements of this entire structure. Hissing behaviour has thus been interpreted in terms of a threat, with the hissing sound serving as a warning signal (Ghatge 1949; Fuchs and Koeniger 1974; Akratanakul 1977; Seeley et al. 1982). Hissing sounds are common aposematic signals that are understood as a warning across many species (Kirchner and Röschard 1999). There are no conclusive experiments proving an effect of hissing behaviour on potential predators, but Koeniger and Fuchs (1973) demonstrated that sun bears, kept in an zoo (Heliarctos malayanus), could be temporarily repelled if hissing sounds were played back inside a box in which they had first found a honeycomb, thus providing at least anecdotal evidence that predators could be deterred.
Hissing alters the colonial state. After hissing, bees are rather less defensive than aroused (Fuchs and Koeniger 1974). In particular, nest activities of the bees cease, and foraging flights commence (SenSarma et al. 2002). In the open-nesting A. florea, during hissing, the nest bees on the surface draw closer to the comb surface and nests appear smaller, and, due to the absence of surface movements, are clearly less conspicuous. Thus, rather than a counter-attack strategy, hissing is related to a retreat strategy, by which bee nests become less noticeable and foragers are protected from individual predation.
17.5.2 “Rearing-up”
A second less widely recognised behaviour, which is superficially similar to “hissing”, has rarely been mentioned and was observed only in colonies of the open-nesting species. On gross disturbances, workers in the colony turn their heads towards the colony and stretch their abdomens outwards. This behaviour is accompanied by wing vibrations and sound production, although of a lower frequency. However, it is set apart from hissing by the body movement where, after stimulation, the individual bees draw their thorax closer to the comb and simultaneously stretch their abdomen outward. It still needs to be clarified whether this is accompanied by exposure of the sting apparatus and/or the release of alarm pheromones. The movement pattern is contagious between neighbouring bees as it spreads over the colony and results in an increase of the colony’s outline and a rough surface structure.
A detailed description of the behaviour in A. florea was given by Pirk et al. (2002), who, however, did not propose a particular name for the pattern, but which might be termed “testudo behaviour”. Kastberger and Sharma (2000) described spreading waves of abdomen stretching in A. dorsata, which differs from “shimmering” behaviour by the slow speed of the propagation, and introduced the term “rearing-up”, which we will maintain here.
“Rearing-up” as a colony display increases rather than decreases the colony’s outline, and a small A. florea colony reacting to a disturbance resembles an angry vertebrate ruffling its fur, which, together with the sound production, is likely to have some impact on potential predators (Fig. 17.5). In relation to “hissing”, “rearing-up” and subsequent attacking appears to be more easily elicited in A. andreniformis than in A. florea, whose colonies are in general more bent on attacking intruders (Oldroyd and Wongsiri 2006). In A. dorsata, “rearing-up” apparently corresponds to the “defensive roar” described by Morse and Laigo (1969), which precedes attacks from the bees and certainly does not fail to warn off humans to retreat before getting stung.

Display of “rearing-up” behaviour in a colony of Apis florea after disturbance by air gusts. Left: before stimulation, right: after stimulation
“Hissing” and “rearing-up” are both elicited by substrate concussions or air gusts; the movement patterns are related, and both are accompanied by sound production. However, a main difference between the behaviours is their different relationships to defensive or counter-attack strategies. While hissing has a distinctly defensive connotation, quieting the nest and rendering it inconspicuous, “rearing-up”, with its outward extension of the sting apparatus, is closely connected to the organisation of counter-attack behaviour. “Rearing-up” typically occurs at higher stimulus intensities or after more persistent stimulation. Thus, in spite of their positioning at opposite ends of the defence/attack axis, the two behaviours connect in a reaction cascade, where more intense stimulation after the initial defensive hissing responses leads to attack-related “rearing-up” behaviour. An apparent relationship between the response types of “releaser” or “retreater” colonies to mechanical stimulation in A. mellifera (Kastberger et al. 2009a, b) might be a further reflection of the two different strategies as possible reactions to variable levels of the same threat.
If disturbance is further continued, “rearing-up” is followed by a general commotion among the bees, a loosening of the bee curtain and the organising into attack flights if an enemy is recognised. This results in bees moving down and hanging at the lower rim of the comb, where they then start, in attacking waves, to drive away any intruder (Morse and Laigo 1969). As a result of the extension of the bee curtain along the bottom of the comb, the optical outline of the colony is augmented. This may take quite impressive dimensions, as demonstrated in the apparent area of an A. dorsata colony more than doubling (Koeniger et al. 2010). This enhancement of the appearance of colony size presumably adds to the threatening effects of “rearing-up”, and the defensive roar might contribute to repel intruders. A particularly interesting effect of these bee beards was observed in A. dorsata by Koeniger and Koeniger et al. (2010), who reported that the oriental honey buzzard, ptylorhynchos, could be misled to erroneously attack the extended bee curtain, and thus fail to successfully prey on the comb and its contents. This observation provides remarkable evidence as to the functionality of colony displays, where direct proof is generally sparse.
Both “hissing” and “rearing-up” have a common response to vibration stimuli, nest concussions and air gusts, which are likely connected to the approach of vertebrate predators. Both involve the emission of perceptible auditory signals. They can be understood as integrated elements of a graded defence cascade, aimed against vertebrates; reaching from retreat-related “hissing” behaviour to counter-attack-related “rearing-up” behaviour; ultimately leading to the organisation of a bee curtain extension and the release of attack waves.
17.5.3 “Shimmering” and “Abdomen-Shaking”
“Abdomen-shaking” was described by Sakagami (1960) in A. cerana as the violent, pendulum-like, lateral swaying of the abdomen, with the behaviour frequently performed by several groups of worker bees. Later, Schneider and Kloft (1971) gave a more detailed description of the movement pattern and recognised its function in group defensive behaviour (“Gruppenverteidigungsverhalten”) against hornets. The behaviour corresponds to that described by Butler (1954) in A. cerana, which he termed “shimmering”.
Koeniger and Fuchs (1973) analysed film sequences and confirmed the basic movement pattern consisting of approximately two to five successive lateral movements in parallel to the ground surface, which first increase and later decrease in amplitude, each lasting approximately 0.09 s. The behaviour is silent and not accompanied by any conspicuous emission of sound, other than a slight rustling of the wings. In several experiments, they confirmed that the behaviour is released exclusively by the movement of dark objects against a light background. In contrast, presentation of motionless objects (including insects), light flashes and chemical stimuli did not release any reaction. In particular, “abdomen-shaking” could not be released by ground vibrations or concussions, which constitute a fundamental difference of “body-shaking” from both “hissing” and “rearing-up” behaviour. The visual stimuli which release “abdomen-shaking” in A. cerana were tested by an optical dummy and showed that black circles of a vision at an angle of approximately 8°, moving at 140° per second over the vision field, were most effective. In particular, the reaction was surprisingly persistent and after a continuous stimulation of over 1 h, 20% of the initially participating bees were still found reacting.
In nature, the behaviour can be observed whenever flying predators of appropriate size and speed approach the bees. The synchronised movements of neighbouring bees suggest that body-shaking is contagious and, as a result of slight delays, waves of body-shaking can be observed travelling over groups of animals. The origin of the body-shaking waves is usually where the moving stimulus is at its closest; but, as the waves do not travel far, it is likely that the behaviour is mainly restricted to workers which perceive some of the stimulus by themselves.
Although body-shaking was observed to effectively discourage various insects, such as butterflies or flies, from landing, the behaviour is mainly aimed to deter predatory wasps (Butler 1954; Schneider and Kloft 1971; Koeniger and Fuchs 1973; Seeley et al. 1982; Abrol 2006; Tan et al. 2007). In the open-nesting species, efficient defence strategies to protect the freely exposed colony are a vital necessity, and this visual display may serve as an efficient initial barrier. “Abdomen-shaking” was observed in A. florea and A. andreniformis (Butler 1954; Koeniger and Fuchs 1973; SenSarma et al. 2002; Oldroyd and Wongsiri 2006), while the giant honeybees, A. dorsata and A. laboriosa, showed the related “shimmering” display of “body-twisting” described below.
Body-shaking also appears widespread in cavity-nesting species. It was long known and first described in the cavity-nesting A. cerana (Butler 1954; Sakagami 1960) and is expressed even more in A. koschevnikovi (Mathew and Mathew 1990) and A. nuluensis (Koeniger et al. 1996). There are no reports related to A. nigrocincta, but it would be an unlikely exception. In the cavity-nesting bees, body-shaking apparently defends the nest entrance against wasp intruders, as was reported in A. nuluensis defending against V. multimaculata (Koeniger et al. 1996). Occasionally, body-shaking at the nest entrance can be observed in A. mellifera when besieged by hornets (personal observation). However, body-shaking in cavity-nesting bees might also be of high relevance during unsheltered periods in reproductive swarming or migration, although no reports are available referring to these situations. Thus an effective colony surface defence is not restricted to permanently open-nesting species, but may be a prerequisite for this status.
17.5.4 “Shimmering” and “Body-Twisting”
“Shimmering” waves in the giant honeybees, A. dorsata and A. laboriosa, are an extraordinarily conspicuous and eye-catching colonial display and were described early by Roepke (1930). Even from a long distance, waves of excitation can be observed travelling across the extended comb surface of these open-nesting giant honeybees. Accordingly, these waves have attracted considerable attention, and impressive video examples are available in Kastberger et al. (1998), Kastberger and Sharma (2000) and Woyke et al. (2008). The characteristic spiralling patterns of the spreading waves closely correspond to other self-organising excitation patterns (Camazine et al. 2003) found in both animate and inanimate systems that are governed by contagious actions followed by latency periods. “Shimmering” patterns thus represent a stunning visualisation of self-organised action in social insects (cf. Chap. 18).
As has been consistently reported, the typical stimulus eliciting “shimmering” waves in giant honeybees is the visual disturbance by flying insects, such as wasps, butterflies or birds, and the behaviour can easily be elicited by moving a dummy object close to the bees. Particularly, responsive individuals play a crucial role in triggering the social waves (Schmelzer and Kastberger 2010). In a detailed study, Kastberger et al. (2008) provided convincing analytical evidence that these waves are indeed capable of driving away preying hornets, and thus for the first time giving direct proof for the efficiency of a honeybee colony-level display.
Although the stimulus releasing the behaviour clearly indicates its close relationship to body-shaking, “shimmering” waves in the giant honeybees differ from “shimmering” by “abdomen-shaking” as found in the other honeybee species, by the distinctly different body movement during which, instead of a lateral swaying of the abdomen, the abdomen is flipped upward. Closer video analysis by Woyke et al. (2008) revealed that the movement is not limited to the abdomen, but includes a rotation of the entire body around a horizontal cross-axis through the thorax, by which the head is simultaneously brought closer to the ground. They thus proposed the term, “defence body-twisting”.
Although “abdomen-shaking” and “body-twisting” are fairly diverse motor patterns, the difference might not be very fundamental. In “abdomen-shaking”, the abdomen is also somewhat raised upward, and this component of the movement may have become the predominant movement in the giant honeybees. Restricting movement to the dorso-ventral component could have been for economical reasons or advantageous for coordination over the fairly large comb areas. Whatever the reasons, a common origin of both motor patterns would be in accordance with a close relationship of the “shimmering” patterns across the species.
On the other hand, the dorso-ventral rotation, with a flipping of the abdomen, closely resembles the motor pattern of “rearing-up”. As discussed above, “rearing-up” is unrelated to “shimmering”, as it is elicited by vibrations and the response mainly addresses ground-moving vertebrate predators. One may, however, consider the possibility that bird predation might have contributed to connecting the action patterns. Birds constitute a major threat to A. dorsata nests. As moving objects, they constitute visual stimuli, and, being visually oriented predators, are unlikely to be confused or scared off by the communal “shimmering” display as are wasp predators. Instead, as highly sophisticated predators of a comparatively large body size, birds would require more substantial defence actions, such as anti-vertebrate counter-attack activities of the rearing-up response and the successive initiation of stinging raids; this could have worked to merge the honeybee defence responses.
It has been noted that in the giant honeybee’s “body-twisting”, the bees expose their Nasanov glands (Kastberger et al. 1998). This interesting observation underlines the role of the displays in social coordination between colony workers. Exposure of the Nasanov gland secretion ensures cohesion between the workers, rather than alerting them for counter-attack (Kastberger et al. 2008). This has also been observed in the “abdomen-shaking” of A. nuluensis (Koeniger et al. 1996) elicited by V. multimaculata attacks that functions to attract additional guards.
17.6 Communication About Predators
Colonial displays serve to deter or confuse attacking predators, with the main effect of the entire colony appearing as a contiguously acting unit. At the same time, these displays may function in an internal context to coordinate defence actions of the individuals. Spreading waves of contagious behaviour confer information to partaking individuals, even if they may not have perceived the releasing stimulus themselves. The well-described build-up of defensive actions in A. dorsata, from first “shimmering” responses until the coordination of coherent attack waves (Lindauer 1956; Morse and Laigo 1969; Koeniger 1975), clearly depicts a sequence in which colonial displays retroact on the individuals’ behavioural states, modifying the colony-level arousal state until decisive action is taken. This information, spread between individuals, may involve not only tactile and vibrational stimuli, but also chemical communication, as in A. dorsata’s “body-twisting” or in A. nuluensis’ “abdomen-shaking”, in which the exposure of the Nasanov gland apparently releases pheromones (Koeniger et al. 1996; Kastberger et al. 1998).
A specifically sophisticated case of inner-colony transfer of information in relation to potential predators has recently been described in the piping–hissing sequence in A. florea (SenSarma et al. 2002), which also demonstrates how pre-existing colony-level behavioural components can be combined for building higher-level information-handling mechanisms. In this behaviour, single foragers who encounter a predator-like stimulus during their return from collection flights engage in impetuous vibrational signalling. They press their thoraces to the comb surface and, over several seconds, emit a series of faintly audible piping sounds, which prove to be a fairly energetic signal in substrate vibration recordings (Fig. 17.6). These vibrations are perceived by adjacent workers on the nest surface, which then react by hissing behaviour. From these initial workers, a hissing wave spreads over the comb and may encompass substantial parts of, or even the entire, colony. The colonial reaction is pattern-sensitive, as hissing is released more readily by subsequent piping within the piping series (Fig. 17.7).

An alerted A. dorsata colony, in which the guard bees have built a curtain, formed by chaining worker bees that are hanging from the lower rim of the comb, and thereby more than doubling the colony’s original outline (solid part of the nest, ruler size 60 cm, photo: N. Koeniger, from Koeniger et al. 2010)

A piping–hissing sequence in Apis florea. A pollen forager has just landed (upper left, arrow) and crawls forward, emitting a series of 10 piping sounds. In order to transmit the vibration, it presses the thorax to the comb (upper right, arrow). The workers on the comb react by stopping activities and draw closer to the comb, emitting a hissing sound while pulling their wings together. This can be clearly seen in the two bees that are marked by a red or green spot, respectively. The sonogram (below) shows a series of piping–hissing sequences. The harmonic structure of the foragers’ piping overlaps in time with the noisy hissing response of the hive workers (photo: S. Fuchs)
In this two-step mechanism, single or few foragers are able to transfer information they have picked up in the field about a potential threat to the nest, and in a second step, this information is rapidly communicated to high proportions of the nestmates. The two steps of this reaction chain are completely separated by the stimulus modalities involved. Foragers detect enemies either by vision or possibly odour. However, the hissing reaction of the workers on the nest is released by sensitivity to nest vibration and not by vision. They do not react by hissing, even if a predator is very close and moving, if the movement or proximity does not cause nest vibrations. However, the nest vibrations applied by the returning workers through piping behaviour cause an identical hissing reaction, as if the nest were directly disturbed by a predator, followed by a cessation of nest activity. There are some indications that this sequence of behaviour is not restricted to A. florea, as piping–hissing sequences have been recorded in A. cerana (S. Biswas personal communication).
The hissing reaction to the piping sound is the same reaction that the bees show to nest vibrations from outside sources, being the percussions from approaching enemies. It is thus likely that, in the evolution of the system, the bees tapped into the pre-existing connection between danger and nest concussions by using vibration signals to communicate danger. Besides the hissing sound, the most obvious reaction of the worker bees is the cessation of all activity. Stopping activity is an unspecific reaction that bees generally show to artificial vibrations applied to combs (Frings and Little 1957). Carpenter ants apply drumming series to the ground, which similarly mimic nest percussions due to their disturbance and immobilise recipient workers, while alerting them to react more strongly to any additional stimuli that may indicate an intruder (Fuchs 1976). Stop signals were emitted by bee foragers that had encountered dangers at the feeding sites and decreased foraging (Nieh 2010). Thus it seems plausible that the connection to ground vibrations indicating danger may be the common root for the various worker piping signals, which tend to induce a cessation of current activities in the recipients (“stop signal”: Nieh 1993) and may also function as modulatory signals (Thom et al. 2003).
In this particular case, the energetic worker piping of returning foragers obviously carries a strong message of imminent danger. A. florea workers detect and report enemies over distances of more than 10 m from the nest, thus truly functioning as far-reaching sensory organs for the colony (Seeley 1994). The fact that honeybees collect and communicate information about food sources inside the nests has been known since von Frisch’s (1967) studies and is regarded as an outstanding example of invertebrate communication skills. Collecting and communicating information on the equally essential environmental conditions and threats to the colony has, however, been widely overlooked and may reveal fascinating details in future research.
References
Abrol D (2006) Defensive behaviour of Apis cerana F. against predatory wasps. J Apic Sci 50:39–46
Akratanakul P (1977). The natural history of the dwarf honey bee, Apis florea F., in Thailand. Thesis, Cornell University, Ithaca
Akratanakul P (1987) Honey bee diseases and enemies in Asia: a practical guide. FAO, Rome
Ambrose JT (1978) Birds. In: Morse RA (ed) Honey bee pests, predators, and diseases. Cornell University Press, Ithaca, pp 215–226
Blum MS, Fales HM, Morse RA, Underwood BA (2000) Chemical characters of two related species of giant honeybees (Apis dorsata and A. laboriosa): possible ecological significance. J Chem Ecol 26:801–807
Breed M, Guzman-Novoa E, Hunt G (2004) Defensive behavior of honey bees: organization, genetics and comparison with other bees. Annu Rev Entomol 49:271–98
Butler CG (1954) The world of the honeybee. Collins, London
Camazine S, Deneubourg JL, Franks NR, Sneyd J, Theralauz G, Bonabeau E (2003) Self-organization in biological systems. Princeton University Press, Princeton
Caron DW (1978) Marsupials and mammals. In: Morse RA (ed) Honey bee pests, predators, and diseases. Cornell University Press, Ithaca, pp 227–256
Cassier P, Tel-Zur D, Lensky Y (1994) The sting sheath of honey bee workers (Apis mellifera L): structure and alarm pheromone secretion. J Insect Physiol 40:23–23
Cunard SJ, Breed MD (1998) Post-stinging behavior of worker honey bees (Hymenoptera: Apidae). Ann Entomol Soc Am 91:754–757
Duangphakdee O, Koeniger N, Koeniger G, Wongsiri S, Deowanish S (2005) Reinforcing a barrier – a specific social defense of the dwarf honeybee (Apis florea) released by the weaver ant (Oecophylla smaragdina). Apidologie 36:505–511
Duangphakdee O, Koeniger N, Deowanish S, Hepburn HR, Wongsiri S (2009) Ant repellent resins of honeybees and stingless bees. Insectes Soc 56:333–339
Dyer FC (1991) Coadaptation of colony design and worker performance in honey bees. In: Smith DR (ed) Diversity in the genus Apis. Westview, Boulder, pp 213–245
Dyer FC, Seeley TD (1991) Nesting behavior and the evolution of worker tempo in four honey bee species. Ecology 72:156–170
Frings H, Little F (1957) Reactions of honeybees in the hive to simple sounds. Science 125:122
Fry CH (1984) The bee-eaters. T&AD Poyser, Town Head House, Calton
Fuchs S (1976) The response to vibrations of the substrate and reactions to the specific drumming in colonies of carpenter ants (Camponotus, Formicidae, Hymenoptera). Behav Ecol Sociobiol 1:155–184
Fuchs S, Koeniger N (1974) Schallerzeugung im Dienst der Kolonieverteidigung bei Apis cerana. Apidologie 5:271–287 [in German]
Ghatge A (1949) Some research work on Apis florea. Indian Bee J 11:5–6
Jayasvati S (1989) Scanning electron microscopy analysis of honey bee Apis florea F., A. dorsata F., A. cerana F. A. mellifera L. and A. andreniformis stings in Thailand. Rec Adv Toxicol Res 2:193–204
Kastberger G, Sharma DK (2000) The predator-prey interaction between blue-bearded bee eaters (Nyctyornis athertoni Jardine and Selby 1839) and giant honey bees (Apis dorsata Fabricius 1798). Apidologie 31:727–736
Kastberger G, Stachl R (2003) Infrared imaging technology and biological applications. Behav Res Methods Instrum Comput 35:429–439
Kastberger G, Raspotnik G, Biswas S, Winder O (1998) Evidence of Nasanov scenting in colony defence of the giant honeybee Apis dorsata. Ethology 104:23–37
Kastberger G, Schmelzer E, Kranner I (2008) Social waves in giant honeybees repel hornets. PLoS ONE 3(9):e3141. doi:10.1371/journal.pone.0003141
Kastberger G, Thenius R, Stabentheiner A, Hepburn R (2009a) Aggressive and docile colony defence patterns in Apis mellifera. A retreater–releaser concept. J Insect Behav 22:65–85
Kastberger G, Weihmann F, Hoetzl T (2009b) Complex social waves of giant honeybees provoked by a dummy wasp support the special-agent hypothesis. J Integr Commun Biol 2:1–2
Kirchner WH, Röschard J (1999) Hissing in bumblebees: an interspecific defence signal. Insectes Soc 46:239–243
Koeniger N (1975) Observations on alarm behaviour and colony defence of Apis dorsata. In: Noirot C, Howse PE, LeMasne G (eds) Pheromones and defensive secretions in social insects. IUSSI, Dijon, pp 153–154
Koeniger N (1976) Neue Aspekte der Phylogenie innerhalb der Gattung Apis. Apidologie 7:357–366 [in German]
Koeniger N, Fuchs S (1973) Sound production as colony defence in Apis cerana. In: Proceedings of 7th international congress, IUSSI, London, pp 199–204
Koeniger N, Weiss J, Maschwitz U (1979) Alarm pheromones of the sting in the genus Apis. J Insect Physiol 25:467–476
Koeniger N, Koeniger G, Mardan M (1994) Mimicking a honey bee queen? Vespa affinis indosiniensis Perez 1910 hunts drones of Apis cerana F. Apidologie 27:353–359
Koeniger N, Koeniger G, Gries M, Tingek S, Kelitu A (1996) Observations on colony defense of Apis nuluensis Tingek, Koeniger and Koeniger 1996, and predatory behaviour of the hornet Vespa multimaculata Perez, 1910. Apidologie 27:314–352
Koeniger N, Koeniger G, Tingek S (2010) Honey bees of Borneo. Natural History, Borneo
Kreil G (1975) The structure of Apis dorsata melittin: phylogenetic relationships between honey bees as deduced from sequence data. FEBS Lett 54:100–102
Lindauer M (1956) Über die Verständigung bei indischen Bienen. Zeit Vergl Physiol 38:521–557 [in German]
Mathew S, Mathew K (1988) The “red” bees of Sabah. Int Bee Res Assoc Newsl Beekeep Trop Subtrop Countries 12:1
Mathew K, Mathew S (1990) Further observations on characters and behaviour of Apis koschevnikovi, the “red bee” of Sabah. Bee World 71:61–66
Matsuura M (1988) Ecological studies on vespine wasps (Hymenoptera: Vespidae) attacking honeybee colonies. Appl Entomol Zool 23:428–440
Matsuura M (2003) Biology and control of social wasps and bees in urban environments 3. Honeybee Sci 24:193–205
Morse RA, Laigo FM (1969) Apis dorsata in the Philippines. Philipp Assoc Entomol 1:1–96
Morse RA, Shearer DA, Bosh SR, Benton AW (1967) Observations on alarm substances in the genus Apis. J Apic Res 6:113–118
Natzir R, Teranishi H, Kitagawa M, Kasuya M (1999) A novel venom protein of the Asian bee (Apis cerana indica) with affinity to human α1-microglobulin. Allergol Int 48:121–129
Nieh JC (1993) The stop signal of honey bees: reconsidering its message. Behav Ecol Sociobiol 33:51–56
Nieh JC (2010) A negative feedback signal that is triggered by peril curbs honey bee recruitment. Curr Biol 20:310–315
Novogrodzki R (1990) Amphibians and reptiles. In: Morse RA (ed) Honey bee pests, predators, and diseases. Cornell University Press, Ithaca, pp 236–242
Oldroyd BP, Wongsiri S (2006) Asian honeybees. Harvard University Press, Cambridge
Ono M, Okada I, Sasaki M (1987) Heat production by balling in the Japanese honeybee Apis cerana japonica as a defensive behaviour against the hornet, Vespa simillima xanthoptera (Hymenoptera: Vespidae). Experientia 43:1031–1032
Ono M, Igarashi T, Ohno E, Sasaki M (1995) Unusual thermal defence by a honeybee against mass attack by hornets. Nature 377:334–336
Papachristoforou A, Rortais A, Zafeiridou G, Theophilidis G, Garnery L, Thrasyvoulou A, Arnold G (2007) Smothered to death: hornets asphyxiated by honeybees. Curr Biol 17:R795–R796
Pirk CWW, Hepburn HR, Radloff SE, Erlandson J (2002) Defense posture in the dwarf honeybee, Apis florea. Apidologie 33(3):289–294
Punchihewa RWK (1994) Beekeeping for honey production in Sri Lanka. Sri Lanka Department of Agriculture, Peradeniya
Ramya J, Rajagopal D (2008) Morphology of the sting and its associated glands in four different honey bee species. J Apic Res 47:46–52
Roepke DW (1930) Beobachtungen an indischen Bienen. Med Landbow Wageningen 34:1–22 [in German]
Sakagami SF (1960) Preliminary report on the specific difference of behaviour and other ecological characters between European and Japanese honeybees. Acta Hymenopt 1:171–198
Sakagami SF, Akahira Y (1960) Studies on the Japanese honeybee, Apis cerana cerana Fabricius. VIII. Two opposing adaptations in the post-stinging behaviour of honeybees. Evolution 14:29–40
Schmelzer E, Kastberger G (2010) ‘Special agents’ trigger social waves in giant honeybees (Apis dorsata). Naturwissenschaften 96:1431–1441
Schmidt JO (1990) Hymenopteran venoms: striving toward the ultimate defense against vertebrates. In: Evans DL, Schmidt JO (eds) Insect defenses: adaptive mechanisms and strategies of prey and predators. State University New York Press, New York, pp 387–463
Schmidt JO (1995) Toxicology of venoms from the honeybee genus Apis. Toxicon 33:917–927
Schneider P, Kloft W (1971) Beobachtungen zum Gruppenverteidigungsverhalten der Östlichen Honigbiene Apis cerana Fabr. Z Tierpsychol 29:337–342 [in German]
Seeley TD (1994) Honey bee foragers as sensory units of their colonies. Behav Ecol Sociobiol 34:51–62
Seeley TD, Seeley RH, Akratanakul P (1982) Colony defense strategies of the honeybees in Thailand. Ecol Monogr 52:43–63
SenSarma M, Fuchs S, Tautz J (2000) Debris removal by head pushing in A. florea Fabr. Honeybees. Naturwissenschaften 87:241–243
SenSarma M, Fuchs S, Tautz J (2002) Worker piping triggers hissing for coordinated colony defence in the dwarf honeybee Apis florea. Zoology 105:215–223
Shah FA, Shah TA (1991) Vespa velutina, a serious pest of honey bees in Kashmir. Bee World 72:161–164
Sugahara M, Sakamoto F (2009) Heat and carbon dioxide generated by honeybees jointly act to kill hornets. Naturwissenschaften 96:1133–1136
Tan K, Hepburn HR, Radloff SE, Yu Y, Liu Y, Zhou D, Neumann P (2005) Heat-balling wasps by honeybees. Naturwissenschaften 92:492–495
Tan K, Radloff SE, Li JJ, Hepburn HR, Yang MX, Zhang LJ, Neumann P (2007) Bee-hawking by the wasp, Vespa velutina, on the honeybees Apis cerana and A. mellifera. Naturwissenschaften 94:469–472
Thapa R, Wongsiri S (2003) Flying predators of the giant honey bees, Apis dorsata and Apis laboriosa in Nepal. Am Bee J 143:540–542
Thapa R, Wongsiri S, Manandhar DN (2000) Current status of predators and diseases of honey bees in Nepal. In: Proceedings of 7th international conference on tropical bees, Chiang Mai, pp 221–226
Thom C, Gilley DC, Tautz J (2003) Worker piping in honey bees (Apis mellifera): the behaviour of piping nectar foragers. Behav Ecol Sociobiol 53:199–205
Veith HJ, Weiss J, Koeniger N (1978) A new alarm pheromone (2-decen 1-yl-acetate) isolated from the stings of Apis dorsata and Apis florea (Hymenoptera: Apidae). Experientia 34: 423–424
Vetter RS, Visscher PK (1998) Bites and stings of venomous arthropods. Int J Dermatol 37: 481–496
von Frisch K (1967) The dance language and orientation of bees. Harvard University Press, Cambridge
Weiss J (1978) Vergleichende Morphologie des Stachelapparates bei den vier Apis-Arten (Hymenoptera: Apidae). Apidologie 9:19–32 [in German]
Wongsiri S, Lekprayoon C, Thapa R, Rinderer TE, Sylvester HA, Oldroyd BP, Booncham U (1996) Comparative biology of Apis andreniformis and Apis florea in Thailand. Bee World 77:23–35
Woyke J, Wilde J, Wilde M, Sivaram V, Cervancia C, Nagaraja N, Reddy M (2008) Comparison of defense body movements of Apis laboriosa, Apis dorsata dorsata and Apis dorsata breviligula honey bees. J Insect Behav 21:481–494
Yang M, Radloff S, Tan K, Hepburn R (2010) Anti-predator fan-blowing in guard bees, Apis mellifera capensis Esch. J Insect Behav 23:12–18
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Fuchs, S., Tautz, J. (2011). Colony Defence and Natural Enemies. In: Hepburn, H., Radloff, S. (eds) Honeybees of Asia. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-16422-4_17
Download citation
DOI: https://doi.org/10.1007/978-3-642-16422-4_17
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-16421-7
Online ISBN: 978-3-642-16422-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)