
Abstract
Adaptive radiations of plants and animals play an important role as model systems in speciation research. Rapid emergence of biological diversity provides opportunities to study adaptive and non-adaptive factors leading to speciation, including the role of spatial factors and ecological, behavioral and genetic mechanisms potentially driving speciation processes. The radiation of sailfin silversides (Atheriniformes: Telmatherinidae) endemic to “Wallace’s Dreamponds”, i.e., the Malili Lakes in Central Sulawesi (Indonesia), allows for testing hypotheses of speciation processes under different geographic settings. Compared with other well-known freshwater fish radiations, the Telmatherinid system is of intermediate size in terms of both, geographical size and organismic diversity. Phylogenetic analyses provide evidence for multiple clades that were connected secondarily through reticulate evolution, but combined analyses support an ancient monophyletic origin of all Telmatherinidae clades in Lake Matano. The consensus view is that the lake contains two reciprocally monophyletic groups of sailfin silversides, highly diverse “sharpfins” heavily introgressed by stream populations, and less diverse “roundfins” not affected by allochthonous introgression. Genetic, morphological, habitat-utilization, trophic, and mate-choice data demonstrate that the most plausible hypothesis for the origin of roundfins is by sympatric speciation. Substantial but not absolute restrictions in gene flow coupled with morphological and behavioral adaptations to distinct ecological niches support the hypothesis that natural selection coupled with assortative mating drives speciation processes in roundfins. In contrast, discrete male color polymorphisms can be explained by sexual selection, but appear not to be associated with population divergence. In line with evidence for adaptive radiation in sharpfins, natural selection for distinct resources is certainly a major force shaping diversity in this lacustrine radiation. However, the role of divergent sexual selection on divergence of ecologically diverse sharpfins remains to be tested.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
1 Introduction
The question if speciation essentially depends on the isolating effect of strict geographical barriers or not has initiated and recurrently refuelled intense debates among evolutionary biologists (Bolnick 2004; Coyne 2007; Dieckmann and Doebeli 1999; Jiggins 2006; Mallet 2001). These discussions have severe implications for our understanding of evolutionary processes, as they are directly connected to the mechanisms driving speciation. Based on the geographical distribution of evolving populations, three major concepts have directed speciation research in the last decades (Coyne and Orr 2004). Mayr (1942) introduced the framework of allopatric speciation, which assumes absence of gene flow due to a strict geographical barrier. Reproductive isolation separating species after secondary contact is expected to emerge as a by-product of geographic isolation, which in turn is thought to result from selection or drift. Based on its intuitive plausibility, allopatric speciation has generally been accepted as the norm, and is often still used as a null hypothesis for other geographic speciation scenarios (Coyne and Orr 2004; but see Bolnick and Fitzpatrick 2007 for a critical view).
In contrast to Mayr’s concept of extrinsic separation, sympatric speciation assumes the evolution of restrictions in gene flow by intrinsic factors. Divergent selection coupled with assortative mating and intraspecific competition is thought to split a single ancestral population into two or more sister species under sympatric conditions. The third scenario, parapatric speciation, combines elements of sym- and allopatric speciation. Diverging populations are expected to have contact but no overlap areas, thereby facilitating local adaptation, which is less likely to evolve in sympatry (Gavrilets and Vose 2005; Gavrilets et al. 2000). However, unambiguous cases for the parapatric origin of species remain to be detected in nature (Gavrilets et al. 2000; Coyne and Orr 2004).
The idea that speciation can proceed without extrinsic separation preventing homogenization of diverging gene pools has raised substantial scepticism, especially regarding the likelihood of divergence in the face of ongoing gene flow (Coyne and Orr 2004; Mayr 1963). However, recent theoretical work shows that even moderately strong divergent selection may overcome very high rates of gene flow (Bolnick and Fitzpatrick 2007; Doebeli and Dieckmann 2000; Gavrilets and Vose 2005; see Gavrilets 2004 and Fitzpatrick et al. 2008a for discussion). Indeed, a growing number of field studies support the view that species can remain distinct despite of substantial gene flow, or even diverge under such conditions (Arnold 2006; Danley et al. 2000; Fitzpatrick et al. 2008b; Samonte et al. 2007; Schilthuizen et al. 2005). According to this view, individual members of diverging populations represent genomic mosaics consisting of comparatively few divergently selected loci and a remaining majority of the genome remaining functionally neutral and undifferentiated (Templeton 1981; Via and West 2008; Wu 2001). Understanding the genomic details of divergence processes leading to speciation remains a major challenge, but the rapidly evolving genomic techniques nowadays allow the tackling of population genomic approaches in non-model organisms (Luikart et al. 2003; Nosil et al. 2009).
2 Speciation Research in Adaptive Radiations
Ecologically isolated host races, evolving species pairs or emerging radiations are prime objects for testing hypotheses of the mechanisms promoting early stages of speciation (Berlocher and Feder 2002; Coyne and Orr 2004; Via and West 2008). Especially, island radiations such as Darwin’s finches (Grant and Grant 2003), Antillean lizards (Thorpe et al. 2008) or the Hawaiian silverswords (Baldwin and Sanderson 1998) have the advantage of allowing for restricting geographic scenarios involved in speciation processes (Emerson 2002). Analogously, freshwater fish endemic to “reverse islands”, i.e., isolated freshwater lakes, rank among the most exciting models for speciation research. These include well-known systems like northern sticklebacks (Schluter 2000) and lake whitefish (Rogers and Bernatchez 2007), cichlid radiations of the East African rift lakes (Kocher 2004), and radiations confined to tiny lakes or small crater lakes or lagoons (Schliewen and Klee 2004; Schliewen et al. 1994, 2001; Strecker et al. 1996). The adaptive character of most of these radiations suggests that natural selection is a major force shaping their diversity; however, sexual selection, drift, hybridization and isolation scenarios are among the possible complementary or alternative explanations (Danley and Kocher 2001; Dieckmann et al. 2004; Mallet 2007). Recent findings of sensory drive speciation uncovered in rock dwelling Lake Voctoria cichlids (Seehausen et al. 2008) suggests that, in line with evidence for reticulate evolution in species flocks (Schliewen and Klee 2004; Seehausen 2004), at least some of these alternatives may contribute significantly to divergence in species flocks.
3 Sailfin Silversides in the Malili Lakes
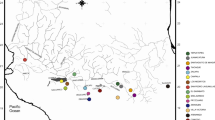
“Wallace’s dreamponds”, the Malili Lakes system in the highlands of Central Sulawesi (Indonesia), constitute, with lacustrine radiations or endemic lineages of snails, crustaceans, and fish (Kottelat 1990a, 1990b, 1991; von Rintelen and Cai 2009; T. von Rintelen et al. 2004, 2007b; K. von Rintelen et al., in press; Schubart and Ng 2008; Schubart et al. 2008), a hotspot of freshwater diversity. The system consists of three major lakes interconnected by steep rivers, and two additional satellite lakes (Fig. 1). Ancient graben-lake Matano covers an area of approx. 32 × 6 km, and is the uppermost lake of the system (Ahmad 1977; Brooks 1950). It is with 590 m depth extraordinary deep, has mostly steep walls and no major intra-lake barriers above approx. 400 m depth (Haffner et al. 2001). Recent limnological investigations demonstrated that it is anoxic below 100 m depth (Crowe et al. 2008a, b). Lake Matano is drained by the extremely steep River Petea to the comparatively small and shallow Lake Mahalona, which in turn is connected by River Tominanga to the largest lake of the system, Lake Towuti. Despite its size of approx. 560 km2, L. Towuti is less than half as deep as L. Matano. The waters of L. Towuti drain from a bay at its western shore to the sea at the Gulf of Bone.

The Malili Lakes system and its endemic sailfin silversides radiation, with focus on Lake Matano’s Telmatherina. “Roundfins”: (a) Telmatherina antoniae “large”, (b) T. antoniae “small”, (c) T. prognatha. “Sharpfins”: (d) T. sarasinorum, (e) T. sarasinorum “bigmouth”, (f) T. sarasinorum “largehead”, (g) T. sp. “thicklip”, (h) T. opudi, (i) T. abendanoni, (j) T. wahjui, (k) T. sp. “elongated”. Fish pictures inside Lake Towuti represent major groups of lacustrine groups shared with L. Mahalona; those besides streams visualize parts of the stream sailfin silverside diversity. All pictures show males, with color polymorphisms (typically yellow or blue, in some cases also blue-yellow) present in most Malili Lakes system Telmatherinidae. See Herder et al. 2006a for sailfin silverside diversity; map by T. von Rintelen, modified (with permission)
The radiation of sailfin silversides (Telmatherinidae) endemic to the Malili Lakes has received substantial interest as a new model system for studying speciation processes and the evolution of color polymorphisms. Sailfin silversides are small, atheriniform (Teleostei: Atheriniformes) freshwater fishes, which are sexually dimorphic, show conspicuous male polychromatism, and are easy to observe in the crystal clear waters of the oligotrophic lakes. Local endemism to single or some of the lakes or streams in combination with intermediate dimensions in both diversity and geographical size provide excellent preconditions for testing hypotheses regarding most of the factors actually discussed as potentially driving speciation processes.
Based on the taxonomic work by Kottelat in the early 1990s (Kottelat 1990a, 1991), the exploration of sailfin silverside diversity in lakes and streams of the area (Herder et al. 2006a) was the essential first step towards establishing the system as a model for speciation research. Surveys of the rivers and most of the permanent streams of the Malili drainage system resulted in the discovery of several new stream-dwelling sailfin silverside populations, which showed indications for local differentiation (Herder et al. 2006a). Likewise, additional lake-dwelling sailfin silversides or previously unknown color morphs were discovered. However, the major patterns of distribution in lake Telmatherinidae confirmed previous records (Kottelat 1990a, 1991; Fig. 1), with most species endemic to either L. Matano or Lakes Towuti and Mahalona (Herder et al. 2006a). Descriptions of mating behavior by Gray and McKinnon (2006) provided an important baseline for later behavioral studies focusing on evolutionary ecology and the maintenance of color polymorphisms.
4 Patterns of Hybridization
Based on individualized samples of lake- and stream-dwelling sailfin silversides, mitochondrial DNA (mtDNA) and amplified fragment length polymorphism (AFLP) markers were applied to reconstruct the phylogenetic history of the Telmatherinidae (Herder et al. 2006b). Robust phylogenies based on dense taxon sampling are a prerequisite for testing hypotheses on the evolution of radiations, and are especially important for identifying monophyletic clades which are suited as candidates for speciation studies. However, phylogenetic reconstructions of young or evolving species flocks are challenging, especially due to the possible effects of reticulate evolution (hybridization) and incomplete lineage sorting. Indeed, maternally inherited mtDNA marker showed only limited congruence to morphological concepts of Telmatherinidae (Herder et al. 2006a), and mtDNA data covering the whole flock indicated several cases of hybridization between lake- and stream-dwelling sailfin silversides (Herder et al. 2006b). In the case of Lake Matano, morphologically well-defined “roundfins” were clearly identified as a monophyletic group, whereas the highly diverse “sharpfins” carry two very distinct groups of haplotypes – one forming the sisterclade of roundfins, the other forming a separate clade comprising haplotypes shared with stream-dwelling Telmatherina.
The parallel analysis of numerous independent AFLP markers scattered randomly across the genome (Vos et al. 1995) is especially suited for the analysis of recent phylogenetic signals (Albertson et al. 1999; Schliewen and Klee 2004). Here, AFLPs served as a tool for critically evaluating phylogenetic signals derived from mtDNA, and for testing hypotheses on introgressive hybridization as deduced from inconsistencies between mtDNA and morphological data. Based on individuals of the mtDNA dataset described above, the nuclear multilocus AFLP-markers strongly supported most of the morphologically well-defined groups of sailfin silversides as monophyletic (Herder et al. 2006b; Fig. 2). In Lake Matano, AFLPs resolved “roundfins” and “sharpfins” (Figs. 1 and 2) as two monophyletic clades, sharply contrasting the pattern of three mitochondrial haplotype groups. However, monophyly of Lake Matano’s Telmatherina in toto was not significant given the whole AFLP dataset (but see below).

Distance-based phylogenetic Neighbor-Net network, reconstructed using 1,327 polymorphic AFLP fragments (redrawn from Herder et al. 2006b). In contrast to mitochondrial haplotype data, nuclear AFLPs support monophyletic “sharpfins” and “roundfins” in Lake Matano, and likewise distinguish Paratherina, Tominanga, and Telmatherina celebensis from Lakes Towuti and Mahalona from stream-dwelling Telmatherina cf. “bonti” populations. The multilocus data contain strong and significant signal for the ancient monophyly of L. Matano’s Telmatherina in toto (sharpfins + roundfins). However, this signal is confounded by alleles deriving from introgression by allochthonous stream populations into L. Matano’s sharpfins (Herder et al. 2006b)
In summary, nuclear, mitochondrial and morphological data suggested that stream-dwelling sailfin silversides most likely hybridize with lake populations. Subsequent ordination statistics and bootstrap homoplasy excess tests (Seehausen 2004) demonstrated that phylogenetic signal shared by sharpfins and stream populations is highly significant in the multilocus dataset, which provides evidence for massive hybridization. In turn, roundfins and sharpfins of Lake Matano were identified as an ancient monophylum, which is masked in the AFLP and mtDNA data due to introgression from riverine invaders (Herder et al. 2006b).
Introgression of L. Matano’s sharpfins by stream populations induced some problems in studies relying solely on mitochondrial haplotype groups. Based on the assumption that Lake Matano’s Telmatherina radiation is physically isolated from all remaining sailfin silversides, Roy et al. (2004) did not incorporate outgroup samples from the other lakes or rivers and streams of the lakes system into phylogenetic analyses. Hence, the three mtDNA haplotype clades present in Lake Matano’s Telmatherina were discussed as three major lineages of sailfin silversides evolved in Lake Matano, rather than identifying the introgressed character of sharpfins and clearly monophyletic roundfins. Accordingly, results and predictions derived from a set of studies (Roy et al. 2004, 2007a, b) will have to be carefully linked to concepts based on nuclear monophyly Footnote 1.
Pronounced phenotypic, and apparently also ecological differences between and among roundfins and sharpfins, led to the conclusion that sailfin silversides of Lake Matano most likely represent an adaptive radiation (Herder et al. 2006b). Conspicuously, introgressed sharpfins are more diverse in terms of body shape than the clearly non-introgressed sympatric roundfins. This led to the idea that introgression might force the evolution of adaptive novelties (Nolte et al. 2005) – ideas that are the object of further, ongoing focal studies critically testing for correlations between increased phenotypic diversity and reticulate evolution (Herder and Schliewen, in preperation).
5 A Key Role of River Petea?
The Telmatherina population inhabiting River Petea – the single outlet of L. Matano – is the only known population of river-dwelling sharpfins (Herder et al. 2006a, b). Carrying the “introgressed” mitochondrial haplotype, this population at the “bottleneck” area separating the L. Matano flock from the downstream diversity of Telmatherinidae is especially suited for testing if the introgression detected in sharpfins is a past event, or possibly a still ongoing phenomenon. Evaluating potential gene flow along River Petea (Fig. 1) also appeared appropriate given the fact that Roy et al. (2004) based speciation scenarios on the assumption that this river constitutes a major barrier preventing gene flow between L. Matano and the rest of the Malili Lakes system. Hence, a focus study targeted genotypic and phenotypic variation along this remarkable river, losing 72 m altitude along its only approx. 9 km length (Schwarzer et al. 2008). The datasets consisting of population-level AFLPs, geometric morphometric shape analyses, and additional measurements suggest high levels of gene flow between both, upper and lower stretches of R. Petea, as well as between Telmatherina sp. “Petea” and lacustrine sharpfins. Phenotypic and genotypic similarity decreases with distance along this short geographic range, indicating genetic exchange along the river and between lacustrine and stream-dwelling sharpfins. In line with these results, field work along River Petea demonstrated that the major waterfall supposed to serve as strict barrier preventing dispersal (Roy et al. 2004, 2006) is absent.
In summary, the studies available support two well-separated clades of sailfin silversides within Lake Matano, which constitute together an ancient monophylum. Morphologically highly diverse “sharpfins” are introgressed by stream populations, a process which appears ongoing in the light of results showing gene flow between L. Matano and River Petea. In contrast, sympatric roundfins show no indications of introgression, neither in terms of mitochondrial haplotypes, nor in AFLP signatures. Widely lacking phylogenetic species-level resolution within both clades indicates that speciation processes are ongoing in sharpfins and roundfins, which in turn is a prime prerequisite for analyzing mechanisms shaping divergence (Dieckmann et al. 2004; Nosil et al. 2009; Via and West 2008).
6 Sympatric Speciation in Lake Matano
Indications for adaptive speciation within the confined space of ancient Lake Matano, coupled with evidence for their monophyly, renders both clades of Lake Matano’s Telmatherina possible candidates for sympatric speciation. Evidently not introgressed, with only three morphospecies of limited phenotypic diversity, and characterized by conspicuous male color polymorphisms, roundfins were the first monophyletic clade of sailfin silversides to be analyzed in detail (Herder et al. 2008). The three roundfin morphospecies are large and deep-bodied Telmatherina antoniae “large,” small and slender T. antoniae “small,” and large, slender T. prognatha (Fig. 1a–c). Males of all three morphospecies occur in either yellow, blue, or blue–yellow courtship coloration, whereas females are dusky gray (Herder et al. 2006a). Morphometric analyses confirmed that the morphospecies are distinct according to size and shape of head and body, but did not support any differentiation among the conspicuous color morphs (Herder et al. 2008; Fig. 3).

Endemic “roundfin” Telmatherina from ancient and extraordinary deep Lake Matano. (a) The three polychromatic morphospecies (i) Telmatherina antoniae “large”, (ii) Telmatherina antoniae “small” and (iii) Telmatherina prognatha are distinct according to body shape (*∼98% correct and significant assignments when leaving out 10–30% of individuals in Jackknife estimates of assignment performance; pictures show adult males). (b) Population structure of roundfins significantly reflects the three morphospecies, but shows no indications for strict reproductive isolation (data shown are reassignment results based on AFLP data; black morphospecies-specific reassignment, gray assignment to another morphospecies). Morphospecies show different patterns of trophic resource use (food importance from stomach contents; green terrestrial arthropods, brown molluscs, blue copepods, yellow fish, white remaining food items). Data from Herder et al. (2008); map by T. von Rintelen, modified (with permission)
All roundfin phenotypes occurred in full sympatry at six sampling locations distributed around the lake, but showed differences in abundance and habitat use. Telmatherina antoniae “small” is by far the most abundant morphospecies, courting and spawning in great numbers mainly in the morning hours at open soft-bottom beach habitats. However, these fish leave spawning areas later in the day, but not towards other benthic inshore areas. Records of T. antoniae “small” in non-breeding mood in the offshore area supported the hypothesis that inshore habitats serve this morphospecies predominantly as courting and spawning grounds, whereas they feed predominantly in the pelagic offshore area. In contrast to the pelagic ecology of T. antoniae “small, T. antoniae “large” inhabit inshore areas. Habitats of these sailfin silversides are typically steep sites characterized by gravel or rock bottom. The rare Telmatherina prognatha occur most frequently at similar sites, but mainly in shallower areas providing shelter by dense canopy of overhanging vegetation and structured by submerged vegetation.
Trophic ecology of roundfins fits predictions of body shape and habitat use (Herder et al. 2008). Stomachs of Telmatherina antoniae “small” mainly contained copepods, which are likely to be available offshore. In contrast, “large” T. antoniae mainly contained small molluscs and terrestrial arthropods like winged ants, corresponding to inshore feeding. Telmatherina prognatha, characterized by a predator-like appearance, indeed contained remains of small fish in addition to terrestrial arthropods. In summary, ecological data suggest that the three roundfin morphospecies are fully sympatric in Lake Matano, sharing spawning grounds, but inhabiting distinct ecological niches.
Population-level AFLP genotyping supported substantial but incomplete reproductive isolation of the three morphospecies, but did not indicate strict barriers for gene flow between color morphs – neither in the “small”, nor in the “large” morphospecies of T. antoniae. Assignment tests showed strong but by far not complete consistency of genotype groups defined by morphology, which supports the hypothesis that differences between morphospecies are significant but not unequivocally distinct in each individual. Focusing on single AFLP loci, only small proportions of the nuclear multilocus dataset turned out to be significantly differentiated, which in turn suggests that footprints of selection are restricted to only small parts of the genome – a result fitting recent ideas about initial stages of ecological speciation (Fitzpatrick et al. 2008b; Via and West 2008; Wu 2001; reviewed by Nosil et al. 2009). In line with significant but incomplete genetic differentiation among roundfin morphospecies, observational transect data highlight very strong but also not absolute morphospecies-assortative mating. Interestingly, significant intrapopulation structure detected between sampling sites in T. antoniae “large” clearly does not increase with intralake distances. This indicates that these predominantly benthic populations are spatially structured, possibly as a result of low dispersal and shoaling. However, para- or allopatric differentiation, which serves as null hypothesis for sympatric modes of speciation, would predict differentiation to increase with geographical distance or the presence of barriers. As neither was supported, this null hypothesis is rejected.
Consistent with evidence for a predominant pelagic ecology derived from transect data and offshore sampling, AFLP data did not detect restrictions of gene flow among benthic sample sites in the small morph of T. antoniae (Herder et al. 2008). A recent study based on microsatellite loci focusing on dispersal in T. antoniae confirmed absence of spatial structure (Walter et al. 2009a). Population clusters detected in that study which do not coincide with sample sites may result from different roundfin morphospecies, which were not explicitly accounted for in this study (Walter et al. 2009a). Similar levels of genetic structure detected [FST = 0.03 (microsatellites; Walter et al. 2009a), FST = 0.019 (AFLPs; Herder et al. 2008)] support this interpretation. Likewise, a second microsatellite study focusing on color polymorphisms (Walter et al. 2009b) also confirmed absence of differentiation among yellow and blue male color morphs in T. antoniae “small”. The latter study also provided first indications for a lack of color morph–specific pattern in male-male competition of T. antoniae “small”.
Combined morphological, ecological, behavioral and genetic data are consistent with a sympatric mode of speciation in Lake Matano’s roundfins, according to the criteria proposed by Coyne and Orr (2004). Roundfins are clearly also sympatric on the micorgeographic level, as all phenotypes regularly encounter each other directly. Based on genetic data and mate choice observations, their reproductive isolation is substantial but only affects small parts of the genome, and allo- or parapatric scenarios are highly unlikely. The criterion of sister group relationship (Coyne and Orr 2004) appears inadequate in the present case, as speciation is obviously not di- but trichotomus in the roundfin flock. However, the intention of this criterion is claiming evidence for divergence within a monophyletic group, which is clearly provided in case of L. Matano’s roundfins.
7 On the Mechanisms Driving Speciation Processes
Answering the question about the geographic scenarios allowing or even promoting speciation processes is important, but remains only an initial step towards understanding the mechanisms driving processes of divergence (Fitzpatrick et al. 2008a). Studies like that of L. Matano’s roundfins demonstrate, in line with several others (Feder et al. 2005; Filchak et al. 2000; Noakes 2008; Rolan-Alvarez 2007; Savolainen et al. 2006; Schliewen et al. 1994, 2001; Steinfartz et al. 2007), that speciation does not depend essentially on the isolating effect of extrinsic barriers, and draw attention to the question how population specific differential adaptation helps to overcome the homogenizing effect of sexual reproduction among incipient species (Coyne 2007; Bolnick and Fitzpatrick 2007; Jiggins 2006).
There are strong indications that response to ecological selection is the causal root of speciation in L. Matano’s Telmatherina (Herder et al. 2006b). Habitat use and trophic ecology concordantly support fine-scaled niche differences in roundfins (Herder et al. 2008), which correspond to morphological traits discussed as adaptive in fish radiations, i.e., body depth and head morphology (Albertson et al. 2003; Kassam et al. 2003; Rüber and Adams 2001). Additional support for adaptation comes from geometric morphometric analyses comparing body shape between roundfins, sharpfins and stream-dwelling populations most likely involved in introgressive hybridization (Herder et al. 2006b). Multivariate axes explaining most of the shape variation demonstrated substantial segregation in body shape among all three groups, which most likely reflects differential adaptation to stream and lake habitats. Complementary to these two lines of evidence supporting ecological selection acting as a major force in speciation of Telmatherina in Lake Matano, a focus study on “trait utility” in sharpfins provides further substantial support for ecological adaptation (Pfaender et al., in press). Trait utility, the performance of traits in terms of fitness, is a central criterion for the recognition of adaptive radiation (Schluter 2000), and can serve as evidence for adaptation due to ecological selection pressure. Pfaender et al. (in press) related expression of potential key traits such as shapes of upper and lower jaw bones, pharyngeal jaws, body shape, gill raker counts and body size to stomach contents, and found surprisingly fine-scaled patterns of morphological differentiation among groups of sharpfins defined by stomach contents. Fish-, shrimp- and egg-feeders were most distinct, with trait expression being widely consistent to expectations derived from other fish radiations.
Morphological adaptations outlined above and distinct patterns of habitat- and mate-choice evident in roundfins (Herder et al. 2008) strongly suggest that divergence in Telmatherina may also affect behavior. Indeed, a series of studies conducted by the group of Gray and McKinnon (Gray and McKinnon 2006; Gray et al. 2007, 2008a) demonstrated in line with our own focus study (Cerwenka et al., in review) highly derived mating and foraging behavior in Telmatherina sarasinorum, a color polymorphic sharpfin species (Fig. 1d) feeding mainly on eggs of con- and heterospecific Telmatherina (Gray and McKinnon 2006; Kottelat 1991). Sailfin silverside eggs are by far the dominating food source of T. sarasinorum at spawning grounds of the roundfin sailfin silverside Telmatherina antoniae “small” (Fig. 1b), and are obtained using two different behavioral strategies correlated with host density (Cerwenka et al., in review). This also affects the egg-feeding species itself, with filial cannibalism correlating with the numbers of cuckolders involved (Gray et al. 2007). A spectacular example of behavioral egg-feeding adaptation in T. sarasinorum has been reported just recently: male T. sarasinorum have been observed courting and enticing female T. antoniae to spawn, and then eating the eggs (Gray et al. 2008a). Gray et al. termed this unique behavioral tactic “sneaky eating,” and suggested that it might have evolved as a possible extension of conspecific egg-feeding in the low resource environment of Lake Matano.
8 Sexual Selection and the Evolution of Colour Polymorphisms
Theory suggests that disruptive sexual selection may promote color polymorphisms (Chunco et al. 2007; Gray and McKinnon 2007) and possibly speciation processes (Kawata et al. 2007), but empirical evidence for speciation triggered by sexual selection is restricted to only a very few cases (Seehausen et al. 2008). The spectacular male color polymorphisms of several sailfin silverside species (Herder et al. 2006a) appeared highly promising for testing hypotheses linking male coloration and speciation processes. However, population-level AFLP or microsatellite data did not support restrictions in gene flow between the conspicuous male color morphs in roundfins, either in T. antoniae “small” or in T. antoniae “large” (Herder et al. 2008; Walter et al. 2009b). Likewise, morphometric data did not indicate any difference in body shape between roundfin color morphs, which might be hypothesized to accumulate under restricted gene flow. Surprisingly, however, explorative analyses of the multilocus AFLP data set revealed significant signal for yellow male coloration across all roundfins included, which suggests heritability of color traits. This heritability is, however, clearly not coupled with significant population structure. Stable color polymorphisms maintained by fluctuating or heterogeneous environments, like lighting conditions changing with daytime or season, or differing with habitat structure, may provide an appropriate explanation for this phenomenon (Chunco et al. 2007; Gray and McKinnon 2007). Recent support for the hypothesis that spatial heterogeneity of the visual environment can influence sexual selection on male coloration comes from observational studies on color polymorphic T. sarasinorum (Gray et al. 2008b; Fig. 1d). Blue and yellow males each turned out to have significantly increased reproductive fitness in one of two alternative habitats tested, which are characterized by opposing lighting conditions corresponding to likewise increased contrast of coloration with the background. Hence, sexual selection decoupled from speciation processes can explain the existence of color polymorphic sailfin silversides; however, widespread presence of different kinds of color polymorphisms in the Malili Lakes radiation, including river- and stream-dwelling populations inhabiting very different kinds of habitats, raises the question whether habitat heterogeneity alone is likely to explain the flock-wide pattern. A combination of different external factors generating visual heterogeneity, including daytime and seasonal effects, might explain these patterns.
9 Perspectives
In summary, sailfin silversides of “Wallace’s dreamponds” have been successfully established and used as a new model system for speciation research, and for the study of selection maintaining color polymorphisms. The comparatively small to medium size of each of the five Malili Lakes as well as the multifaceted geographical structure of the lakes system allows incorporating sympatrically, parapatricallyand allopatrically distributed radiations into comprehensive analyses.
This setting is hardly comparable to any other aquatic model systems, neither to the very large and complex East Africal Great Lakes, nor to the tiny crater lake species assemblages in Cameroon and Nicaragua. Hence, the sailfin silverside system offers great potential for evolutionary biology, especially for speciation research. Based on the initial results reviewed here, four major research topics have emerged as especially promising. First, the adaptive character of both sailfin silverside radiations of Lake Matano offers the chance for testing the idea of selection shaping adaptive fitness landscapes (Gavrilets 2004; Kingsolver and Pfennig 2007) in a spatially confined environment. Linking individual fitness correlates to morphological character expression and their ecological utility, this approach has the potential for providing deeper insight into processes of adaptive speciation in sympatry. Second, and complementary, massive introgression of known stream-dwelling populations into the dynamically evolving sharpfins of Lake Matano has set the optimal stage for testing hypotheses regarding the role of hybridization and introgression on the generation of increased phenotypic and genotypic diversity (Rieseberg et al. 1999; Seehausen 2004; Stelkens and Seehausen 2009). Third, and probably most important, the Malili system offers unique opportunities for testing hypotheses on different genomic consequences of sympatric versus parapatric or allopatric speciation (Nosil et al. 2009). Adaptive sympatric speciation of L. Matano’s roundfins is opposed to parapatric and allopatric settings in the Towuti-Mahalona system, where lakes are connected by a gentle river that may serve as corridor for gene flow. Genome scans applied to L. Matano’s roundfins (Herder et al. 2008) have served as a first step towards the genomic analysis of the system. Availability of the complete medaka (Oryzias latipes) genome (Kasahara et al. 2007), a species much more closely related to Telmatherina than other fully sequenced species like stickleback or zebrafish, may allow insights into the basis of speciation relevant genes. Last, but not least, the dominance of color polymorphisms in the whole flock and genetic data supporting heritability and environment contingent fitness consequences suggest that heterogeneous environments maintain color polymorphism in most telmatherinid species, but its mechanisms and potential implications for speciation remain only partially understood. Although a significant role for population divergence of roundfins based on selective female choice for alternative male color polymorphism is unlikely, this is not necessarily the case for sharpfins of Lake Matano or other sailfin silversides. Speciation through sensory drive (Seehausen et al. 2008) remains a plausible hypothesis for the origin of ecologically divergent species, some of which are polymorphic while others are not.
A major task remaining is to provide a time scale for the sailfin silverside radiation. Preliminary results derived from distance-based divergence estimates support an age of 0.95–1.9 Mya for the split separating roundfin and “non-stream” sharpfin mtDNA haplotypes (Roy et al. 2007b; Clades I and II), which roughly corresponds to the estimated geological age of L. Matano (1–2 mya; cited in von Rintelen et al. 2004). However, application of model-based approaches incorporating all relevant clades of the systems (Stoeger et al. in preparartion) appears appropriate to address this rather fundamental aspect.
The Malili Lakes system does not only harbor several independent fish radiations or endemic lineages. The data now available on the evolution of pachychilid snails (Glaubrecht and von Rintelen 2008; von Rintelen et al. 2004, 2007b, this volume), athyid shrinps (K von Rintelen and Cai 2009; K von Rintelen et al. in press) and gecarcinucid freshwater crabs (Schubart and Ng 2008; Schubart et al. 2008) remain to be assembled with the sailfin silverside data. Such an approach linking patterns of divergence in fundamentally different freshwater organisms restricted to the same area may finally provide insight into major environmental effects shaping the endemic diversity, including barriers for dispersal, and might allow incorporating potential effects of coevolution (see T. von Rintelen et al. 2004, this volume; von Rintelen et al. 2007a). Wallace´s dreamponds undoubtedly provide a rich environment for evolutionary biologists after Darwin to study the origin of species – beyond “simple” sympatric speciation.
Notes
- 1.
Inferences of evolutionary history of radiations from phylogenetic data require an adequate taxon sampling, including the relevant outgroups (see Herder et al. (2006a, 2006c), Schliewen et al. (2006) and Schwarzer et al. (2008) for discussion with respect to publications on fish radiations in Sulawesi (Roy et al. 2004, 2007a, b) and in Nicaraguan crater lakes (Barluenga et al. 2006). In addition, quality control of a posteriori inferences critically depends on the availability of “individualized” voucher material, in order to enable cross-checking of results. This is especially important for studies of incipient speciation, where phenotypic variation is not necessarily discrete and unambigous assignment of individuals to (emerging) species is not always possible.
References
Ahmad W (1977) Geology along the Matano Fault Zone East Sulawesi, Indonesia. In: Proceedings of the regional conference on the geology and mineral resources of South East Asia, Jakarta:1–15
Albertson RC, Markert JA, Danley PD, Kocher TD (1999) Phylogeny of a rapidly evolving clade: the cichlid fishes of Lake Malawi, East Africa. Proc Natl Acad Sci USA 96:5107–5110
Albertson RC, Streelman JT, Kocher TD (2003) Directional selection has shaped the oral jaws of Lake Malawi cichlid fishes. Proc Natl Acad Sci USA 100:5252–5257
Arnold GM (2006) Evolution through genetic exchange. Oxford University Press, New York
Barluenga M, Stölting KN, Salzburger W, Muschick M, Meyer A (2006) Sympatric speciation in crater lake cichlid fish. Nature 439:719–723
Baldwin BG, Sanderson MJ (1998) Age and rate of diversification of the Hawaiian silversword alliance (Compositae). Proc Natl Acad Sci USA 95:9402–9406
Berlocher SH, Feder JL (2002) Sympatric speciation in phytophagous insects: moving beyond controversy? Annu Rev Entomol 47:773–815
Bolnick DL (2004) Waiting for sympatric speciation. Evolution 58:895–899
Bolnick DL, Fitzpatrick BM (2007) Sympatric speciation: models and empirical evidence. Annu Rev Ecol Evol Syst 38:459–487
Brooks JL (1950) Speciation in ancient lakes (concluded). Q Rev Biol 25:131–176
Cerwenka AF, Schliewen UK, Herder F Egg-feeding in Lake Matano’s adaptive sailfin silversides radiation. (in review)
Chunco AJ, McKinnon JS, Servedio MR (2007) Microhabitat variation and sexual selection can maintain male color polymorphisms. Evolution 61:2504–2515
Coyne JA (2007) Sympatric speciation. Curr Biol 17:R787–R788
Coyne JA, Orr HA (2004) Speciation. Sinauer, Sunderland
Crowe SA, O'Neill AH, Katsev S, Hehanussa P, Haffner GD, Sundby B, Mucci A, Fowle DA (2008a) The biogeochemistry of tropical lakes: a case study from Lake Matano, Indonesia. Limnol Oceanogr 53:319–331
Crowe SA, Jones CA, Katsev S, Magen C, O’Neil AH, Sturm A, Canfield DE, Haffner GD, Mucci A, Sundby B, Fowle DA (2008b) Photoferrotrophs thrive in an Archean Ocean analogue. Proc Natl Acad Sci USA 105:15938–15943
Danley PD, Kocher T (2001) Speciation in rapidly diverging systems: lessons from Lake Malawi. Mol Ecol 10:1075–1086
Danley PD, Markert JA, Arnegard ME, Kocher TD (2000) Divergence with gene flow in the rock-dwelling cichlids of Lake Malawi. Evolution 54:1725–1737
Dieckmann U, Doebeli M (1999) On the origin of species by sympatric speciation. Nature 400:354–357
Dieckmann U, Doebeli M, Metz JAJ, Tautz D (2004) Adaptive speciation. Cambridge University Press, Cambridge
Doebeli M, Dieckmann U (2000) Evolutionary branching and sympatric speciation caused by different types of ecological interactions. Am Nat 156:S77–S101
Emerson BC (2002) Evolution on oceanic islands: molecular phylogenetic approaches to understanding pattern and process. Mol Ecol 11:951–966
Feder JL, Xie X, Rull J, Velez S, Forbes A, Leung B, Dambroski H, Filchak KE, Aluja M (2005) Mayr, Dobzhanski, and Bush and the complexities of sympatric speciation in Rhagoletis. Proc Natl Acad Sci USA 102:6573–6580
Filchak KE, Roethele JB, Feder JL (2000) Natural selection and sympatric divergence in the apple maggot Rhagoletis pomonella. Nature 407:739–742
Fitzpatrick BM, Fordyce JA, Gavrilets S (2008a) What, if anything, is sympatric speciation? J Evol Biol 21:1452–1459
Fitzpatrick BM, Placyk JS, Niemiller ML, Casper GS, Burghardts GM (2008b) Distinctiveness in the face of gene flow: hybridization between specialist and generalist gartersnakes. Mol Ecol 17:4107–4117
Gavrilets S (2004) Fitness landscapes and the origin of species. Princeton University Press, Princeton and Oxford
Gavrilets S, Li H, Vose MD (2000) Patterns of parapatric speciation. Evolution 54:1126–1134
Gavrilets S, Vose A (2005) Dynamic patterns of adaptive radiation. Proc Natl Acad Sci USA 102:18040–18045
Glaubrecht M, von Rintelen T (2008) The species flocks of lacustrine gastropods: Tylomelania on Sulawesi as models in speciation and adaptive radiation. Hydrobiologia 615:181–199
Grant BR, Grant PR (2003) What Darwin's finches can teach us about the evolutionary origin and regulation of biodiversity. Bioscience 53:965–975
Gray SM, McKinnon JS (2006) A comparative description of mating behaviour in the endemic telmatherinid fishes of Sulawesi’s Malili Lakes. Environ Biol Fish 75:471–482
Gray SM, McKinnon JS (2007) Linking color polymorphism maintenance and speciation. Trends Ecol Evol 22:71–79
Gray SM, Dill LM, McKinnon JS (2007) Cuckoldry incites cannibalism: male fish turn to cannibalism when perceived certainty of paternity decreases. Am Nat 169:258–263
Gray SM, McKinnon JS, Tantu FY, Dill LM (2008a) Sneaky egg-eating in Telmatherina sarasinorum, an endemic fish from Sulawesi. J Fish Biol 73:728–731
Gray SM, Dill LM, Tantu FY, Loew ER, Herder F, McKinnon JS (2008b) Environment-contingent sexual selection in a colour polymorphic fish. Proc R Soc Lond B 275:1785–1791
Haffner GD, Hehanussa PE, Hartoto D (2001) The biology and physical processes of large lakes of Indonesia: Lakes Matano and Towuti. In: Munawar M, Hecky RE (eds) The great lakes of the world (GLOW) food web, health and integrity. Backhuis, Leiden, pp 182–192
Herder F, Schwarzer J, Pfaender J, Hadiaty RK, Schliewen UK (2006a) Preliminary checklist of sailfin silversides (Pisces: Telmatherinidae) in the Malili Lakes of Sulawesi (Indonesia), with a synopsis of systematics and threats. Verh Ges Ichthyol 5:139–163
Herder F, Nolte A, Pfaender J, Schwarzer J, Hadiaty RK, Schliewen UK (2006b) Adaptive radiation and hybridization in Wallace’s Dreamponds: evidence from sailfin silversides in the Malili Lakes of Sulawesi. Proc R Soc Lond B 273:2209–2217
Herder F, Hadiaty RK, Schliewen UK (2006c) Diversity and evolution of Telmatherinidae in the Malili Lakes system in Sulawesi. In: Proceedings of the international symposium “the ecology and limnology of the Malili Lakes” March 20–22 Bogor, Indonesia, pp 67–72
Herder F, Pfaender J, Schliewen UK (2008) Adaptive sympatric speciation of polychromatic “roundfin” sailfin silverside fish in Lake Matano (Sulawesi). Evolution 62:2178–2195
Jiggins CD (2006) Sympatric speciation: why the controversy? Curr Biol 16:R333–R334
Kasahara M, Naruse K, Sasaki S, Nakatani Y, Qu W, Ahsan B, Yamada T, Nagayasu Y, Doi K, Kasai Y, Jindo T, Kobayashi D, Shimada A, Toyoda A, Kuroki Y, Fujiyama A, Sasaki T, Shimizu A, Asakawa S, Shimizu N, Hashimoto S, Yang J, Lee Y, Matsushima K, Sugano S, Sakaizumi M, Narita T, Ohishi K, Haga S, Ohta F, Nomoto H, Nogata K, Morishita T, Endo T, Shin-I T, Takeda H, Morishita S, Kohara Y (2007) The medaka draft genome and insights into vertebrate genome evolution. Nature 447:714–719
Kassam DD, Adams DC, Ambali AJD, Yamaoka K (2003) Body shape variation in relation to resource partitioning within cichlid trophic guilds coexisting along the rocky shore of Lake Malawi. Anim Biol 53:59–70
Kawata M, Shoji A, Kawamura S, Seehausen O (2007) A genetically explicit model of speciation by sensory drive within a continuous population in aquatic environments. BMC Evol Biol 7:99
Kingsolver JG, Pfennig DW (2007) Patterns and power of phenotypic selection in nature. Bioscience 57:561–572
Kocher TD (2004) Adaptive evolution and explosive speciation: the cichlid fish model. Nat Genet 5:288–298
Kottelat M (1990a) Sailfin silversides (Pisces: Telmatherinidae) of Lakes Towuti, Mahalona and Wawontoa (Sulawesi, Indonesia) with descriptions of two new genera and two new species. Ichthyol Explor Freshwaters 1:227–246
Kottelat M (1990b) The ricefishes (Oryziidae) of the Malili Lakes, Sulawesi, Indonesia, with description of a new species. Ichthyol Explor Freshwaters 1:151–166
Kottelat M (1991) Sailfin silversides (Pisces: Telmatherinidae) of Lake Matano, Sulawesi, Indonesia, with descriptions of six new species. Ichthyol Explor Freshwaters 1:321–344
Luikart G, England PR, Tallmon D, Jordan S, Taberlet P (2003) The power and promise of population genomics: from genotyping to genome typing. Nat Genet 4:981–994
Mallet J (2007) Hybrid speciation. Nature 446:279–283
Mallet J (2001) The speciation revolution - commentary. J Evol Biol 14:887–888
Mayr E (1942) Systematics and the origin of species. Columbia University Press, New York
Mayr E (1963) Animal species and evolution. Belknap, Cambridge, MA
Noakes DLG (2008) Charr truth: sympatric differentiation in Salvelinus species. Environ Biol Fish 83:7–15
Nolte A, Freyhof J, Stemshorn K, Tautz D (2005) An invasive lineage of sculpins, Cottus sp. (Pisces, Teleostei) in the Rhine with new habitat adaptations has originated from hybridization between old phylogenetic groups. Proc R Soc Lond B 272:2379–2387
Nosil P, Funk DJ, Ortiz-Barrientos D (2009) Divergent selection and heterogeneous genomic divergence. Mol Ecol 18:375–402
Pfaender J, Schliewen UK, Herder F (in press) Trait utility in “sharpfin” sailfin silversides of Lake Matano (Sulawesi) suggests adaptive radiation. Evol Ecol (Online First)
Rieseberg LH, Archer MA, Wayne R (1999) Transgressive segregation, adaptation and speciation. Heredity 83:363–372
Rogers SM, Bernatchez L (2007) The genetic architecture of ecological speciation and the association with signatures of selection in natural lake whitefish (Coregonus sp., Salmonidae) species pairs. Mol Biol Evol 24:1423–1438
Rolan-Alvarez E (2007) Sympatric speciation as a by-product of ecological adaptation in the galician Littorina saxatilis hybrid zone. J Molluscan Stud 73:1–10
Roy D, Docker MF, Hehanussa PE, Heath DD, Haffner GD (2004) Genetic and morphological data supporting the hypothesis of adaptive radiation in the endemic fish of Lake Matano. J Evol Biol 17:1268–1276
Roy D, Kelly DW, Fransen CHJM, Heath DD, Haffner GD (2006) Evidence of small-scale vicariance in Caridina lanceolata (Decapoda: Atyidae) from the Malili Lakes, Sulawesi. Evol Ecol Res 8:1087–1099
Roy D, Docker MF, Haffner GD, Heath DD (2007a) Body shape vs. colour associated initial divergence in the Telmatherina radiation in Lake Matano, Sulawesi, Indonesia. J Evol Biol 20:1126–1137
Roy D, Docker MF, Paterson G, Hamilton PB, Heath DD, Haffner GD (2007b) Resource-based adaptive divergence in the freshwater fish Telmatherina from Lake Matano, Indonesia. Mol Ecol 16:35–48
Rüber L, Adams DC (2001) Evolutionary convergence of body shape and trophic morphology in cichlids from Lake Tanganyika. J Evol Biol 14:325–332
Samonte IE, Satta Y, Sato AS, Tichy H, Takahata N, Klein J (2007) Gene flow between species of Lake Victoria Haplochromine Fishes. Mol Biol Evol 24:2069–2080
Savolainen V, Anstett M-C, Lexer C, Hutton I, Clarkson JJ, Norup MV, Powell MP, Springate D, Salamin N, Baker WJ (2006) Sympatric speciation in palms on an oceanic island. Nature 441:210–213
Schilthuizen M, Cabanban AS, Haase M (2005) Possible speciation with gene flow in tropical cave snails. J Zool Sys Evol Res 43:133–138
Schliewen UK, Klee B (2004) Reticulate sympatric speciation in Cameroonian crater lake cichlids. Front Zool 1:1–12
Schliewen UK, Tautz D, Pääbo S (1994) Sympatric speciation suggested by monophylie of crater lake cichlids. Nature 368:629–632
Schliewen UK, Rassmann K, Markmann M, Markert J, Kocher T, Tautz D (2001) Genetic and ecological divergence of a monophyletic cichlid species pair under fully sympatric conditions in Lake Ejagham, Cameroon. Mol Ecol 10:1471–1488
Schliewen UK, Kocher TD, McKaye KR, Seehausen O, Tautz D (2006) Evidence for symparic speciation? Nature 444:E12–E13
Schluter D (2000) The ecology of adaptive radiation. Oxford University Press, Oxford
Schubart CD, Ng PKL (2008) A new molluscivore crab from Lake Poso confirms multiple colonization of ancient lakes in Sulawesi by freshwater crabs (Decapoda: Brachyura). Zool J Linn Soc 154:211–221
Schubart CD, Santl T, Koller P (2008) Mitochondrial patterns of intra- and interspecific differentiation among endemic freshwater crabs of ancient lakes in Sulawesi. Contrib Zool 77:83–90
Schwarzer J, Herder F, Misof B, Hadiaty RK, Schliewen UK (2008) Gene flow at the margin of Lake Matano’s adaptive sailfin silverside radiation: Telmatherinidae of River Petea in Sulawesi. Hydrobiologia 615:201–213
Seehausen O (2004) Hybridization and adaptive radiation. Trends Ecol Evol 19:198–207
Seehausen O, Terai Y, Magalhaes IS, Carleton KL, Mrosso HDJ, Miyagi R, van der Sluijs I, Schneider MV, Maan ME, Tachida H, Imai H, Okada N (2008) Speciation through sensory drive in cichlid fish. Nature 455:620–626
Strecker U, Meyer CG, Sturmbauer C, Wilkens H (1996) Genetic divergence and speciation in an extremely young species flock in Mexico formed by the genus Cyprinodon (Cyprinodontidae, Teleostei). Mol Phyl Evol 6:143–149
Steinfartz S, Weitere M, Tautz D (2007) Tracing the first step to speciation: ecological and genetic differentiation of a salamander population in a small forest. Mol Ecol 16:4550–4561
Stelkens R, Seehausen O (2009) Genetic distance between species predicts novel trait expression in their hybrids. Evolution 63:884–897, DOI: 10.111/j.1558-5646.2008.00595.x
Templeton AR (1981) Mechanisms of speciation - a population genetic approach. Annu Rev Ecol Syst 12:23–48
Thorpe RS, Jones AG, Malhotra A, Surget-Groba Y (2008) Adaptive radiation in Lesser Antillean lizards: molecular phylogenies and species recognition in the Lesser Antillean dwarf gecko complex, Sphaerodactylus fantasticus. Mol Ecol 17:1489–1504
Via S, West J (2008) The genetic mosaic suggests a new role for hitchhiking in ecological speciation. Mol Ecol 17:4334–4345
von Rintelen K, Cai Y (2009) Radiation of endemic species flocks in ancient lakes: Systematic revision of the freshwater shrimp Caridina H. Milne Edwards, 1837 (Crustacea: Decapoda: Atyidae) from the ancient lakes of Sulawesi, Indonesia, with the description of eight new species. Raff Bull Zool 57:343–452
von Rintelen K, von Rintelen T, Meixner M, Lüter C, Cai Y, Glaubrecht M (2007a) Freshwater shrimp-sponge association from an ancient lake. Biol Lett 3:262–264
von Rintelen K, Glaubrecht, M, Schubart CD, Wessel A, von Rintelen T. (in press) Adaptive radiation and ecological diversification of Sulawesi's ancient lake shrimps. Evolution
von Rintelen T, Bouchet P, Glaubrecht M (2007b) Ancient lakes as hotspots of diversity: a morphological review of an endemic species flock of Tylomelania (Gastropoda: Cerithioidea: Pachychilidae) in the Malili lake system on Sulawesi, Indonesia. Hydrobiologia 592:11–94
von Rintelen T, Wilson AB, Meyer A, Glaubrecht M (2004) Escalation and trophic specialization drive adaptive radiation of freshwater gastropods in acient lakes on Sulawesi, Indonesia. Proc R Soc Lond B 271:2541–2549
Vos P, Hogers R, Bleeker M, Reijans M, van de Lee T, Hornes M, Frijters A, Pot J, Peleman J, Kuiper M, Zabeau M (1995) AFLP: a new technique for DNA fingerprinting. Nucl Acids Res 23:4407–4414
Walter RP, Haffner GD, Heath DD (2009a) Dispersal and population genetic structure of Telmatherina antoniae, an endemic freshwater Sailfin silverside from Sulawesi. Indonesia J Evol Biol 22:314–323
Walter RP, Haffner GD, Heath DD (2009b) No barriers to gene flow among sympatric polychromatic ‘small’ Telmatherina antoniae from Lake Matano, Indonesia. J Fish Biol 74:1804–1815
Wu CI (2001) The genic view of the process of speciation. J Evol Biol 14:851–865
Acknowledgments
We thank the Indonesian Institute of Sciences (LIPI) for the permit to conduct research in Indonesia. We are especially grateful to R. K. Hadiaty for strongly supporting our project in Indonesia. PT. INCO provided outstanding logistic support in Sulawesi. A. Cerwenka, S. Chapuis, J. Frommen, R. K. Hadiaty, J. Herder, A. Nolte, J. Pfaender and J. Schwarzer contributed in the field and/or laboratory to the success of the project. W. Wickler (MPG) enabled the study by financing a pilot study. Fieldwork greatly benefited from logistic support in Indonesia by T. von Rintelen. We acknowledge T. von Rintelen for providing access to digitized maps. Comments and suggestions by Jobst Pfaender and two anonymous referees helped improving the manuscript. This study was funded by research grants from the Deutsche Forschungsgemeinschaft to UKS (DFG SCHL 567/2-1, 2, 3) and by a graduate fellowship donated by the Rheinische Friedrich Wilhelms- Universität Bonn to F.H.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2010 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Herder, F., Schliewen, U.K. (2010). Beyond Sympatric Speciation: Radiation of Sailfin Silverside Fishes in the Malili Lakes (Sulawesi). In: Glaubrecht, M. (eds) Evolution in Action. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-12425-9_22
Download citation
DOI: https://doi.org/10.1007/978-3-642-12425-9_22
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-12424-2
Online ISBN: 978-3-642-12425-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)