
Abstract
Climate change research has progressed rapidly over the last two decades, with model projections of future climate conditions gaining enough consensus to enable downscaling these changes to regional scales. While the field of HAB research has come far since its early roots, our current understanding is not well suited to utilize advances in climate modeling to project how HABs’ prevalence and character may differ in the future oceans. This situation largely is due to the complexity of interspecies competition, but it also can be attributed to the “insular” nature of HAB research. HAB studies focus more on the HAB organism, often in isolated cultures, than on the phytoplankton community in which it may or may not flourish. Even though GEOHAB fostered comparative research, it most often is not possible to quantitatively compare among published HAB studies because nonuniform methods are used. Most HAB observational programs are triggered only when toxic species abundances become high enough to threaten human health, so the specific conditions that catalyze these bloom developments cannot be described. It is difficult then to project how changing environmental conditions may influence the development of HABs, particularly given that these events generally are comparatively rare within the context of dynamic coastal ecosystems. Participants in an international workshop considered these issues and identified a number of specific steps, presented in small part here, that will help HAB research to obtain compelling evidence that climate change is impacting HAB distribution, prevalence, or character. New HAB research strategies are needed if we are to develop informed projections for HABs under anticipated end-of-century conditions in the future oceans.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
- Harmful Algal Blooms (HAB)
- Future Ocean
- Coastal Ecosystem Dynamics
- Colored Dissolved Organic Matter (CDOM)
- CDOM Concentrations
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
There is emerging evidence that climate change already may be causing shifts in the composition of coastal and offshore marine plankton and benthic communities, and there is reasonable expectation that future climate scenario’s will lead to changes in the spatial and temporal ranges of HAB species. The concern is that these changes may increase the frequency and severity of harmful algal blooms (HABs), which can be broadly categorized as toxin-producing or high-“biomass” events. But while there are many intuitive linkages, such as greater stratification leading to increased prevalence of HABs (Moore et al. 2015), these scenarios are founded on limited and often conflicting data. As a consequence, scientific evidence cannot presently establish that climate change will increase HABs, let alone what magnitude of change might be expected. More problematic, the HAB research community has been largely unprepared to rigorously address this question. International consensus is needed to help collectively guide new research strategies, a goal that will be facilitated under the umbrella of the new Global Harmful Algal Bloom (GlobalHAB) Programme.
The central challenge facing HAB research is achieving some consensus about the most efficient research and management strategies to obtain unequivocal evidence that climate will, or has, altered the frequency, distribution, or character of HABs. These strategies should include identifying meaningful hypotheses (e.g., multistressor effects) that can be tested across species and studying HABs in specific environments, such as the Arctic, where climate change impacts are projected to be most significant (IPCC 2013). Creating synergies among these focused research strategies and observations, and assimilating these findings into ecosystem-based models, will be critical if our knowledge of HAB-climate change interactions presage actual impacts.
One of the first steps towards developing some community consensus will be to appraise what is understood about the environmental conditions that favour initiation and maintenance of different types of HAB events. This understanding is limited and fraught with structural holes, as demonstrated by our current abilities to correctly forecast HAB events, but it will serve as a springboard to focus attention on the research issues of greatest importance over the next decade. It also will help to proactively identify the fundamental parameters and research infrastructure needed to effectively hindcast current changing HAB distributions: the first step in gaining the capacity to forecast future HAB patterns in a changing climate.
The case for high-biomass HABs and climate change is more nuanced. The fundamental driver of many high-biomass HABs is well understood: elevated anthropogenic nutrient inputs [see Chaps. 4, 12, Glibert et al. (2018a, b)]. Nevertheless, climate has the potential to affect the intensity and distribution of high-biomass HAB events through, for example, changes in runoff patterns and dynamics stemming from altered precipitation regimes. The challenge will be to discriminate these effects from those associated with regional-scale shifts in human society.
A small international workshop was organized in spring 2013 at the Friday Harbor Laboratories, University of Washington, to preliminarily assess how well the current status of HAB research can serve as a foundation for projecting the effects of climate change on future HAB prevalence and character. The expertise of participants spanned the ecophysiology and nutrient acquisition of HAB species, their nutritional quality and implications for food web structure and ecosystem health, and HAB-suitable observational platforms, time series analyses, and prediction. One objective of the workshop was to produce a summary paper identifying the keystone parameters and research infrastructure needed to test the purported HAB-climate change linkage (Wells et al. 2015). The summary of those published findings that follows represents the dedicated efforts of those participants.
2 Direct Effects of Temperature on HABs
Temperature influences all metabolic processes, is a primary physiological constraint to species range extension, and is clearly shown to have increased in the surface mixed layer of the oceans (Beardall et al. 2009). But warming is not globally uniform, in either latitudinal or regional extents (Roemmich et al. 2012; Stocker et al. 2013), so this potential change in the physical environment will differ greatly among locations. The greatest temperature changes by far have been and will continue to be at high latitudes, but it is possible that small temperature shifts at lower latitudes might have equally significant impacts on HAB prevalence (e.g., benthic HABs). Perhaps of greater significance, though, is how temperature may expand the “windows of opportunity” for existing HAB regions, assuming that physiological limits are not exceeded during peak temperatures (i.e., late summer). However, the central question is whether temperature directly influences the competitive advantage of HAB species over non-HAB species or whether indirect temperature effects on other physical processes (e.g., increased stratification, altered winds, etc.) have the primary role in regulating potential HAB-temperature relationships.
Direct effects of temperature on phytoplankton physiology and metabolism in general are well documented (Beardall and Raven 2004; Bissenger et al. 2008; Eppley 1972; Kamykowski and McCollum 1986; Raven and Geide 1988), with recognition that the optimal and inhibitory limits differ among metabolic thresholds and species. In the case of at least dinoflagellates, increased temperatures (along with light) can stimulate, but also inhibit, germination (e.g., Anderson and Rengefors 2006; Hallegraeff et al. 1998; Itakura and Yamaguchi 2005). Temperature regimes largely constrain the biogeographical boundaries of phytoplankton species (Longhurst 1998; Okolodkov 1999, 2005), likely due in large part to their temperature tolerances. However with few exceptions (e.g., Rhodes et al. 1994), there are few data on the temperature-driven competitive interactions among HAB and non-HAB species. There also are indications that temperature is correlated directly with toxin production in some HAB species (Ogata et al. 1989), but this effect is not uniform (e.g., Lewis et al. 1993). Unusually long periods of warm and calm weather may favour cyanobacterial blooms in the Baltic Sea (e.g., Kahru et al. 2016). Such conditions were found during the ecosystem disruptive bloom of Prymnesium polylepis (syn. Chrysochromulina polylepis) in Scandinavia in 1988 (Gjosaeter et al. 2000).
There are many avenues for investigating the direct effects of temperature on the prevalence of HABs in the future ocean, but there are four leading questions that, addressed experimentally, would accelerate improved understanding. First, most studies of direct temperature effects on HAB species utilize single strains, but are these representative of the genetically diverse natural populations, or do isolates from regions that experience greater seasonal temperature variations display different resilience or competitive advantages (Boyd et al. 2013)? Second, what are the synergistic or additive effects of multistressors with temperature; i.e., will temperature effects be mitigated or exacerbated by changes in other physical parameters? Third, are there observational data that can now show HAB range expansion, or contraction, associated with documented temperature increases? Finally, will any changes in future HAB prevalence result more from changes in physiology or changes in the windows of opportunity for HABs to develop?
3 Direct Effects of Stratification on HABs
Climate change pressures will lead to warmer surface waters and increased precipitation and runoff in many coastal regions (IPCC 2013), both of which will lead to increased stratification of surface waters—likely more so in mid- to high-latitude regions than in tropical/subtropical waters (Behrenfeld et al. 2006). Changes in wind speed and direction will also affect stratification; altered wind speed can increase or decrease stratification. Projected increases in stratification will alter patterns of nutrient availability, in many cases leading to decreased overall nutrient supply to surface waters. In addition to altering the seasonal patterns of blooms in general, and HABs in particular (Moore et al. 2009), dinoflagellates are well adapted to exploit conditions of strong shallow nutriclines (Figueiras et al. 2006; Smayda and Trainer 2010), and this understanding serves as the precarious foundation for much of the intuitive linkage between climate change and HABs.
The fundamental uncertainty is whether increased stratification will enhance the competitive abilities of HAB organisms over non-HAB organisms, at least to the extent where toxic impacts develop in the food web. There is ample evidence that stratification favours swimmers (Peacock and Kudela 2014) and, in cases of stable stratification, the formation of thin layers that can harbour HAB organisms (e.g., Aoki et al. 2014; Farrell et al. 2012; McManus et al. 2008; Pettersson and Pozdnyakov 2013; Raine 2014; Rines et al. 2002; Ryan et al. 2010; Strom et al. 2013). Decadal patterns of increasing intensity and decreasing depth of the pycnocline in the California Current system have coincided with increased prevalence of dinoflagellates in these waters (Jester et al. 2009; Kudela et al. 2010). Widespread Pseudo-nitzschia blooms on the western seaboard of the USA in 2015 followed unusually intense late winter stratification from the onshore movement of anomalously warm oceanic waters (the “blob”), indicating that stratification may have helped precondition the system to support this massive event (McCabe et al. 2016). Increasing stratification also has been associated with an apparent recent emergence of Dinophysis species in Canada, the USA, and South Africa (Hubbart et al. 2012; Pitcher et al. 1993; Taylor et al. 2013; Trainer et al. 2013). Blooms of fish-killing flagellates have been associated with extended periods of calm weather in Scandinavia (Eckford-Soper and Daugbjerg 2016; Gjosaeter et al. 2000; Granéli and Turner 2006 and articles therein).
These apparent correlations notwithstanding, in almost all cases it is not possible to mechanistically demonstrate causation, and it is critical to remember that stratification, in the vast majority of cases, does not lead to HAB formation. Two important questions then are: how might increased stratification work in conjunction with other environmental parameters to favour the success of HAB species? And, how important is swimming behaviour, or the formation of thin layers (biological refugia), in facilitating HAB species success within phytoplankton assemblages? It will be equally important to ask under what scenarios increased stratification will not lead to HAB development (e.g., McGillicuddy et al. 2011). Insights to these factors would help to establish whether broadening of stratification in the future ocean would lead to the expansion of HABs.
4 Altered Light Field Effects on HABs
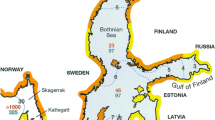
The increase in heat flux to higher latitudes over the next century is projected to cause a global increase in clouds in the mid- to high latitudes, although there remains uncertainty over these projections (IPCC 2013). Combined with trends of increased runoff that would bring higher particulate loads (Häder et al. 2010) and “brownification” (from humic substances) to coastal waters (Hansson et al. 2013), there is good reason to expect greater light attenuation in future higher latitude waters. These effects may be countered in part by shallowing of the mixed layer, but that also would lead to to increased near-surface exposure to higher-intensity photosynthetically active radiation as well as UV radiation. Despite uncertainties of the relative importance of these contrasting effects, it seems likely that light fields will be changing in much of the future ocean. Increasing levels of “brownification,” or coloured dissolved organic matter (CDOM), may lead to differences in plankton community structure. For example, mixotrophs appear to have a competitive advantage in high CDOM water. Paczkowska et al. (2017) showed that in the Gulf of Bothnia, northern Baltic Sea, where CDOM concentrations are high, the proportion of mixotrophs are higher than in the southern part of the Baltic Sea, where CDOM levels are lower.
There are no known PAR requirements or tolerances that competitively separate HAB species from non-HAB species, although differing optical properties among algal taxa are well known. However, high surface irradiance can influence the production of UV absorbing toxic mycosporine-like amino acids (MAA). Although MAA are not unique to HAB species, as they are found in many non-toxic cyanobacteria and raphidophytes (Carreto and Carignan 2011; Jeffrey et al. 1999; Mohlin et al. 2012), they are presumed to contribute to the high-light tolerances of these and other HAB species.
There is equivocal evidence that high light intensities influence the “traditional” toxicity of HAB organisms. For example, saxitoxin content increases in Alexandrium fundyense with increasing light (Etheridge and Roesler 2005), but there is no general trend among Alexandrium spp. (Fu et al. 2012). In other cases, the more toxic strains of Microcystis aeruginosa and Pseudo-nitzschia spp. are observed to have greater tolerances to high light conditions. None of these or other light relationships are characterized well enough to generalize, but there are reasons to ask whether changing light fields, both increasing and decreasing maximum irradiances, will affect HAB species or alter their toxin production. In the case of tropical/subtropical benthic HABs currently prolific in high light environments, will increases in PAR/UV help to expand their range towards higher latitudes, or will light tolerance thresholds be surpassed causing contraction of their contemporary tropical distributions? Although there are a number of unresolved questions regarding light field effects, understanding how future changes in light fields may affect HAB prevalence depends upon climate models gaining better insight on the distribution and magnitude of changes in global cloud cover.
5 Effects of Ocean Acidification on HABs
Ocean acidification (OA) in surface waters is the consequence of atmospheric CO2 dissolution rates into surface waters being greater than the rate of surface water exchange with the deep ocean. The end-of-century surface water pH is projected to be 0.3–0.4 units lower than present day, with most of this change occurring in high-latitude regions (IPCC 2013). These changes will affect carbon acquisition in phytoplankton, with increasing pCO2(aq) de-emphasizing the value of carbon-concentrating mechanisms (CCMs) utilized by some phytoplankton (Beardall et al. 2009; Hansen 2002; Hinga 2002). Species that contain the low-affinity Form II Rubisco (ribulose-1,5-bisphosphate carboxylase-oxygenase) and those having limited CCM capabilities, including some toxic dinoflagellates and perhaps all raphidophytes, in particular could benefit from increased pCO2(aq) (Dason and Colman 2004; Nimer et al. 1997), although findings so far have been mixed (Cho et al. 2001; Lundholm et al. 2004; Sun et al. 2011; Tatters et al. 2012).
Changes in pH also will affect other aspects of cell metabolism, including membrane potential, enzyme activity, and energy partitioning (Beardall and Raven 2004; Giordano et al. 2005). Increases in hydrogen ion activity can influence nutrient uptake, through alterations to chemical speciation or transmembrane transport, and flagellar motion, which could affect movement to favourable nutrient regimes (Hallegraeff et al. 2012). There also is evidence, though limited and conflicting, that toxin production can increase in at least some strains of Alexandrium at lower pH (Kremp et al. 2012), and this increased toxicity can be enhanced in Pseudo-nitzschia spp. under nutrient limitation (Sun et al. 2011; Tatters et al. 2012).
There are numerous unresolved questions about OA effects on HABs. For example, does increasing CO2 availability benefit HAB species versus non-HAB species, and, if so, under what conditions might this competitive advantage affect the prevalence of HAB events? More fundamentally, are our current experimental methods adequate to enable the replication needed to gain consensus about OA effects on the toxicity and competitive interactions of HAB species, given the inherent difficulties in conducting pH manipulation experiments?
6 Effects of Nutrients on HABs
The majority of high-biomass HABs stem from impacts of cultural eutrophication, which will continue to be the major driving mechanism in the future ocean [see also Chaps. 4, 12, Glibert et al. (2018a, b)]. Changes in precipitation regimes are likely to diminish these HABs in some regions while exacerbating them in others. Gaining a better understanding of these changes largely waits for the adequate downscaling of global models to regional-scale predictions of environmental change. The comparative simplicity of mechanistic control for high-biomass HABs means that advances in projecting their response to climate change will be coupled to advances in downscaled climate forecasts.
Decades of HAB research has provided no clear evidence that changes in natural macronutrient supply drives a shift towards HAB species, although great progress has been made [see, however, Chaps. 4, 12, Glibert et al. (2018a, b)]. Of all nutrients, nitrogen (N) supply and forms have attracted the greatest interest, and in some cases there appear to be linkages between N supply or forms and cell toxicity (e.g., Auro and Cochlan 2013; Dyhrman and Anderson 2003; Leong et al. 2004; Levasseur et al. 1995). However, there is no overall consistent pattern among laboratory studies to suggest that specific N sources generate greater success or toxicity of HAB species—a perspective that is consistent with observations in at least eastern boundary current upwelling regimes (Kudela et al. 2010).
The cell toxicity of Pseudo-nitzschia spp. appears to consistently increase under N, P, or Si limited senescence (cf. reviews by Bates and Trainer 2006; Lelong et al. 2012; Trainer et al. 2012). Earlier and more rapid onset of warming in temperate waters can lead to more intense spring blooms and greater depletion of nutrients (McGillicuddy et al. 2011), which potentially could lead to more toxic Pseudo-nitzschia scenarios in the future.
Increased intensities or duration of stratification will decrease new nutrient inputs to surface waters, and these conditions favour flagellates (Smayda 1998). While it is clear that these changes in nutrient flux will regulate phytoplankton community development, the complexity of the response among individual species frustrates attempts to link nutrient supply to toxic HAB events. However, more detailed field observations may help refine this understanding. For example, the massive Pseudo-nitzschia bloom event along the western coast of N. America in 2015 may have stemmed from “preincubation” of coastal waters under low-nutrient conditions. Pseudo-nitzschia spp. have superior nutrient uptake capabilities relative to many centric diatoms under low-nutrient conditions (e.g., Auro and Cochlan 2013; Kudela et al. 2010; Wells 2003), so extended low-nutrient conditions may have selected for Pseudo-nitzschia spp. success once upwelling conditions returned (McCabe et al. 2016).
Fundamental questions remain about how climate-induced changes in nutrient fields may impact HABs. First, are the nutrient uptake kinetics and preferences of HAB species sufficiently different from non-HAB species to inform on likely competitive outcomes [see also Chap. 6, Glibert et al. (2018c)]? If so, then can improved linkages between observed nutrient fields and physical, ecological, and climate change models improve our competence in forecasting contemporary HAB patterns? Any advances in this area would add confidence to projecting how changes in nutrient fluxes may impact future HAB trends. Finally, while the vast majority of studies on nutrient/HAB interactions are restricted to the macronutrients N, P, and Si, how do changes in micronutrient and vitamin supply affect the competitive success of HAB species in coastal waters?
7 Grazer Effects on HABs
Most studies on HAB development focus on how environmental conditions affect the growth of HAB species. Far fewer studies have considered “top-down” controls that grazers exert on the success, or failure, of HAB species competition, and the complexity of these potential controls is daunting. The tiers of grazers include microbial pathogens, microzooplankton, copepods and other mesozooplankton, benthic invertebrates, and fish, each with unique modes of predation, prey preference, susceptibility to toxins, and ecologies (Smayda 1992). Indeed, some HAB organisms can be mixotrophic, potentially enhancing their relative abundance in the plankton community (e.g., Flynn et al. 2013; Jeong et al. 2010; see also Chap. 7, Flynn et al. 2018). Some HABs are apparently immune to grazing pressure, such as ecosystem disruptive blooms (Buskey et al. 2001), while grazing pressures are too sluggish to significantly affect other HABs (Schultz and Kjørboe 2009).
The dominant grazers on HAB species are microzooplankton (Turner 2006), and while toxic HAB prey do not appear to kill microzooplankton, their chemical defences can serve to impair grazer activity (e.g., Strom 2002). The presence of toxic prey species in some cases reduces copepod grazing rates (Engström-Ost et al. 2002; Koski et al. 1999; Schultz and Kjørboe 2009), while in others there are no effects (Engström-Ost et al. 2002). Even when prey toxins do influence copepod grazing rates, these effects are partially modulated by repeated exposures to the toxins (Colin and Dam 2002; Zheng et al. 2011).
Perhaps the greatest difficulty in assessing grazer effects on HABs in general, let alone extrapolation of this knowledge to future climate scenarios, is constructing experimental conditions that are representative of natural systems. That is, selection of appropriate prey densities, predator-prey pairings (including mixotrophs), mixed prey opportunities, and strain differences among prey and predators creates considerable logistical difficulties for designing experiments. It is possible that the current trophic-ecological paradigms may shift to altered systems that are not well represented under current environmental regimes.
Given the diversity of prey ecology and predator feeding strategies, it is unlikely that a general predator-prey interaction model will be sufficient to characterize climate change effects on grazer/HAB interactions. While modelling efforts are improving (e.g., Mitra and Flynn 2006), there remain many questions. For example, what are the best approaches to study grazing effects on HAB success? Will climate-driven changes in the coastal environment alter some or many of the known predator-prey relationships by temporally decoupling the grazer and prey life histories? To what extent will top-down control of the phytoplankton community be influenced by OA (e.g., see pH effects on sensory capacity—Lonnstedt et al. 2013), and what might this change mean for HABs in particular? Progress on understanding climate impacts on grazer/HAB species coupling will depend upon intensive study of a narrow subset of specific grazer-prey interactions under standardized future climate conditions.
8 General Strategies to Accelerate Understanding of Climate Change Impacts on HABs
Climate change already is causing shifts in the seasonal dynamics of production and phytoplankton community composition, but the projections of how these changes will affect the distribution, character, and intensity of HABs remains largely speculative. Some consensus is needed within the HAB research community on steps that will accelerate our understanding in both scientific and management perspectives, if our knowledge is to advance faster than any climate-induced changes in HABs. This is particularly true for high-latitude northern regions where climate change impacts are most rapid. Proactive approaches are needed that develop rigorous, testable hypotheses to guide scientists, managers, and the public on what changes are projected, and then put into place the necessary observing infrastructures (research studies, detection schemes, observing systems) to capture these critical data. Modelling or prediction of HABs under changing environmental conditions will have little hope of being accurate without the foresight to obtain the relevant data.
As a start, the fundamental limitations of single stressor or factor experiments must be recognized. Although there are many known “unknowns” on how HAB and non-HAB organisms respond to individual climate change factors, it is more relevant to ask how these species might alter their responses under multiple stressors that better represent future ocean conditions. Multistressor experiments should become the standard experimental approach. New experimental systems such as community-scale mesocosms need to be developed as tools for testing multiple stressor effects on ecosystem responses that enhance HAB species competitive success or toxicity. Effective utilization of quantitative projection models as experimental tools depends heavily on having data on these interactive outcomes.
Broad spatial and temporal observational data will be critical for informing and testing projections of climate impacts on HABs. Our current data gathering methods are largely inadequate. These programmes rely heavily on seafood safety monitoring rather than studying HAB species in their oceanographic context. The necessity for new insights to the driving mechanisms underlying HAB development helped foster the GEOHAB Programme’s focus in predictive ecology and oceanography. There is a critical need to establish, or augment, long-term HAB observer sites, both in current HAB regions and more importantly in areas (e.g., high latitudes) where HABs are not currently recognized. These sites should span citizen science-based programmes involving stakeholders (Van Dolah et al. 2016), to cutting-edge automated sensor systems (Campbell et al. 2010; Preston et al. 2011), taking advantage of the local strengths in different regions (e.g., utilizing traditional “low-tech” methods such as microscopy in regions where “higher-tech” methods are less tractable). In each case, collection of a minimum range of oceanographic and meteorological data should be encouraged to help characterize environmental conditions.
Stronger linkages with existing global observing systems are needed. For example, the Global Ocean Observing System (GOOS) include HABs as part of the essential ocean variable phytoplankton biomass and diversity (see www.goosocean.org) and the GEO Task SB-01 Oceans and Society: Blue Planet initiative (Djavidnia et al. 2014) comprises ocean information systems, forecasting networks, ecosystem monitoring, and fisheries/aquaculture management, all of which are relevant to contextualizing HAB occurrences and impacts. Indeed, HABs are specifically recognized as a task component (Bernard et al. 2014). The International Ocean Colour Coordinating Group (IOCCG) is another Programme of relevance, working towards developing remote sensing capabilities for detecting phytoplankton functional types, HABs, and water quality.
Finally, “HAB” is a scientifically arbitrary, societal-dependent definition, so understanding the full scope of climate change effects on HABs will entail ascertaining not only direct and indirect human health impacts but also the broader socioeconomic and cultural implications (Smayda 1997; Trainer and Yoshida 2014). New synergies are needed through expanding HAB research beyond HAB ecology, physiology, or direct health and economic impacts to embrace social science and economics. These broader human wellness aspects of HABs are an important part of the new IOC/SCOR-supported GlobalHAB Programme.
References
Anderson DM, Rengefors K (2006) Community assembly and seasonal succession of marine dinoflagellates in a temperate estuary: the importance of life cycle events. Limnol Oceanogr 51(2):860–873
Aoki K, Onitsuka G, Shimizu M et al (2014) Variability of factors driving spatial and temporal dispersion in river plume and Chattonella antiqua bloom in the Yatsushiro Sea, Japan. Mar Pollut Bull 81(1):131–139
Auro ME, Cochlan WP (2013) Nitrogen utilization and toxin production by two diatoms of the Pseudo-nitzschia pseudodelicatissima complex: P. cuspidata and P. fryxelliana. J Phycol 49(1):156–169
Bates SS, Trainer VL (2006) The ecology of harmful diatoms. In: Granéli E, Turner JT (eds) Ecology of harmful algae. Springer, Berlin, pp 81–93
Beardall J, Raven JA (2004) The potential effects of global climate change on microbial photosynthesis, growth and ecology. Phycologia 43:26–40
Beardall J, Stojkovic S, Larsen S (2009) Living in a high CO2 world: impacts of global climate change on marine phytoplankton. Plant Ecol Divers 2(2):191–205
Behrenfeld MJ, O’Malley RT, Siegel DA et al (2006) Climate-driven trends in contemporary ocean productivity. Nature 444(7120):752–755
Bernard S, Kudela R, Velo-Suárez L (2014) Developing global capabilities for the observation and prediction of harmful algal blooms. In: Djavidnia S, Cheung V, Ott M et al (eds) Oceans and society, blue planet. Cambridge Scholars Publishing, Newcastle upon Tyne, pp 46–52
Bissenger JE, Montagnes SJ, Atkinson D (2008) Predicting marine phytoplankton maximum growth rates from temperature: improving on the Eppley curve using quantile regression. Limnol Oceanogr 53:487–493
Boyd PW, Rynearson T, Armstrong EA et al (2013) Marine phytoplankton temperature versus growth responses from polar to tropical waters – outcome of a scientific community-wide study. PLoS One 8(5):e63091. https://doi.org/10.1371/journal.pone.0063091
Buskey EJ, Liu H, Collumb C, Bersano JGF (2001) The decline and recovery of a persistent Texas brown tide algal bloom in the Laguna Madre (Texas, USA). Estuaries 24:337–346
Campbell L, Olson RJ, Sosik HM et al (2010) First harmful Dinophysis (Dinophyceae, Dinophysiales) bloom in the US is revealed by automated imaging flow cytometry. J Phycol 46(1):66–75
Carreto JI, Carignan MO (2011) Mycosporine-like amino acids: relevant secondary metabolites. Chemical and ecological aspects. Mar Drugs 9(3):387–446
Cho ES, Kotaki Y, Park JG (2001) The comparison between toxic Pseudo-nitzschia multiseries (Hasle) Hasle and non-toxic P. pungens (Grunow) Hasle isolated from Jinhae Bay, Korea. Algae 16:275–285
Colin SP, Dam H (2002) Latitudinal differentiation in the effects of the toxic dinoflagellate Alexandrium spp. on the feeding and reproduction of populations of the copepod Acartia hudsonica. Harmful Algae 1:113–125
Dason JS, Colman B (2004) Inhibition of growth in two dinoflagellates by rapid changes in external pH. Can J Bot 82(4):515–520
Djavidnia S, Cheung V, Ott M et al (eds) (2014) Oceans and society, blue planet. Cambridge Scholars Publishing, Newcastle upon Tyne, p 285
Dyhrman ST, Anderson DM (2003) Urease activity in cultures and field populations of the toxic dinoflagellate Alexandrium. Limnol Oceanogr 48(2):647–655
Eckford-Soper L, Daugbjerg N (2016) The ichthyotoxic genus Pseudochattonella (Dictyochophyceae): distribution, toxicity, enumeration, ecological impact, succession and life history – a review. Harmful Algae 58:51–58
Engström-Ost J, Viitasalo M, Jodonasdottir S et al (2002) Calanoid copepods feed and produce eggs in the presence of toxic cyanobacteria Nodularia spumigena. Limnol Oceanogr 47:878–885
Eppley RW (1972) Temperature and phytoplankton growth in the sea. Fish Bull 70:1063–1085
Etheridge SM, Roesler CS (2005) Effects of temperature, irradiance, and salinity on photosynthesis, growth rates, total toxicity, and toxin composition for Alexandrium fundyense isolates from the Gulf of Maine. Deep Sea Res II 52:2491–2500
Farrell H, Gentien P, Fernand L et al (2012) Scales characterising a high density thin layer of Dinophysis acuta Ehrenberg and its transport within a coastal jet. Harmful Algae 15:36–46
Figueiras FG, Pitcher GC, Estrada M (2006) Harmful algal bloom dynamics in relation to physical processes. In: Granéli E, Turner JT (eds) Ecology of harmful algae. Springer, Berlin, pp 127–138
Flynn KJ, Stoecker DK, Mitra A et al (2013) Misuse of the phytoplankton-zooplankton dichotomy: the need to assign organisms as mixotrophs within plankton functional types. J Plankton Res 35(1):3–11
Flynn KJ, Mitra A, Glibert PM et al (2018) Mixotrophy in HABs: by whom, on whom, why, and what next. In: Glibert PM, Berdalet E, Burford M et al (eds) Global ecology and oceanography of harmful algal blooms. Springer, Cham, pp 113–132
Fu FX, Tatters AO, Hutchins DA (2012) Global change and the future of harmful algal blooms in the ocean. Mar Ecol Prog Ser 470:207–233
Giordano M, Beardall J, Raven JA (2005) CO2 concentrating mechanisms in algae: mechanisms, environmental modulation, and evolution. Annu Rev Plant Biol 56:99–131
Gjosaeter J, Lekve K, Stenseth NC, Leinaas HP, Christie H, Dahl E, Danielssen DS, Edvardsen B, Olsgard F, Oug E, Paasche E (2000) A long-term perspective on the Chrysochromulina bloom on the Norwegian Skagerrak coast 1988: a catastrophe or an innocent incident? Mar Ecol Prog Ser 207:201–218
Glibert PM, Al-Azri A, Allen JI et al (2018a) Key questions and recent research advances on harmful algal blooms in relation to nutrients and eutrophication. In: Glibert PM, Berdalet E, Burford M et al (eds) Global ecology and oceanography of harmful algal blooms. Springer, Cham, pp 229–259
Glibert PM, Beusen AHW, Harrison JA et al (2018b) Changing land-, sea- and airscapes: sources of nutrient pollution affecting habitat suitability for harmful algae. In: Glibert PM, Berdalet E, Burford M et al (eds) Global ecology and oceanography of harmful algal blooms. Springer, Cham, pp 53–76
Glibert PM, Heil CA, Wilkerson F et al (2018c) Nutrients and HABs: dynamic kinetics and flexible nutrition. In: Glibert PM, Berdalet E, Burford M et al (eds) Global ecology and oceanography of harmful algal blooms. Springer, Cham, pp 93–112
Granéli E, Turner JT (eds) (2006) Ecology of harmful algae. Springer, Berlin, p 413
Häder DP, Helbling W, Williamson CE et al (2010) Effects of UV radiation on aquatic ecosystems and interactions with climate change. Photochem Photobiol Sci 10:242–260. https://doi.org/10.1039/C0PP90036B
Hallegraeff GM, Blackburn SI, Doblin MA et al (2012) Global toxicology, ecophysiology and population relationships of the chainforming PST dinoflagellate Gymnodinium catenatum. Harmful Algae 14:130–143
Hallegraeff GM, Marshall JA, Valentine J (1998) Short cyst-dormancy period of an Australian isolate of the toxic dinoflagellate Alexandrium catenella. Mar Freshw Res 49:415–420
Hansen PJ (2002) Effect of high pH on the growth and survival of marine phytoplankton: implications for species succession. Aquat Microb Ecol 28(3):279–288
Hansson L-A, Nicolle A, Granéli W et al (2013) Food-chain length alters community responses to global change in aquatic systems. Nat Clim Chang 3(3):228–233
Hinga KR (2002) Effects of pH on coastal marine phytoplankton. Mar Ecol Prog Ser 238:281–300
Hubbart B, Pitcher GC, Krock B et al (2012) Toxigenic phytoplankton and concomitant toxicity in the mussel Choromytilus meridionalis off the west coast of South Africa. Harmful Algae 20:30–41
Intergovermental Panel on Climate Change (IPCC) (2013) Climate change 2013: the physical science basis. In: Contribution of working group I to the fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge and New York, NY
Itakura S, Yamaguchi M (2005) Morphological and physiological differences between the cysts of Alexandrium catenella and A. tamarense (Dinophyceae) in the Seto Inland Sea, Japan. Plankton Biol Ecol 57:85–91
Jeffrey SW, MacTavish HS, Dunlap WC et al (1999) Occurrence of UVA- and UVB-absorbing compounds in 152 species (206 strains) of marine microalgae. Mar Ecol Prog Ser 189:35–51
Jeong HJ, Yoo YD, Kim JS et al (2010) Growth, feeding and ecological roles of the mixotrophic and heterotrophic dinoflagellates in marine planktonic food webs. Ocean Sci J 45(2):65–91
Jester R, Lefebvre KA, Langlois G et al (2009) A shift in the dominant toxin-producing algal species in central California alters phycotoxins in food webs. Harmful Algae 8(2):291–298
Kahru M, Elmgren R, Savchuk OP (2016) Changing seasonality of the Baltic Sea. Biogeosciences 13(4):1009–1018
Kamykowski D, McCollum SA (1986) The temperature acclimatized swimming speed of selected marine dinoflagellates. J Plankton Res 8:275–287
Koski M, Rosenberg M, Viitasalo M et al (1999) Is Prymnesium patelliferum toxic for copepods? – Grazing, egg production, and egestion of the calanoid copepod Eurytemora affinis in mixtures of “good” and “bad” food. ICES J Mar Sci 56:131–139
Kremp A, Godhe A, Egardt J et al (2012) Intraspecific variability in the response of bloom-forming marine microalgae to changed climate conditions. Ecol Evol 2(6):1195–1207
Kudela RM, Seeyave S, Cochlan WP (2010) The role of nutrients in regulation and promotion of harmful algal blooms in upwelling systems. Prog Oceanogr 85(1–2):122–135
Lelong A, Hégaret H, Soudant P et al (2012) Pseudo-nitzschia (Bacillariophyceae) species, domoic acid and amnesic shellfish poisoning: revisiting previous paradigms. Phycologia 51(2):168–216
Leong SCY, Murata A, Nagashima Y et al (2004) Variability in toxicity of the dinoflagellate Alexandrium tamarense in response to different nitrogen sources and concentrations. Toxicon 43(4):407–415
Levasseur M, Gamache T, St.-Pierre I et al. (1995) Does the cost of NO3 reduction affect the production of harmful compounds by Alexandrium excavatum? In: Lassus, P, Arzul G, Erard E et al (eds), Harmful marine algal blooms. Technique et Documentation-Lavoisier, Intercept Ltd, pp 463–468
Lewis NI, Bates SS, McLachlan JL et al (1993) Temperature effects on growth, domoic acid production, and morphology of the diatom Nitzschia-pungens f. multiseries. In: Smayda TJ, Shimizu Y (eds) Toxic phytoplankton blooms in the sea, 3. Elsevier Science Publ B V, Amsterdam, pp 601–606
Longhurst A (1998) Ecological geography of the sea. Academic Press, San Diego, p 398
Lonnstedt OM, Munday PL, McCormick MI, Ferrari MCO et al (2013) Ocean acidification and responses to predators: can sensory redundancy reduce the apparent impacts of elevated CO2 on fish? Ecol Evol 3(10):3565–3575
Lundholm N, Hansen PJ, Kotaki Y (2004) Effect of pH on growth and domoic acid production by potentially toxic diatoms of the genera Pseudo-nitzschia and Nitzschia. Mar Ecol Prog Ser 273:1–15
McCabe RM, Hickey BM, Kudela RM et al (2016) An unprecedented coastwide toxic algal bloom linked to anomalous ocean conditions. Geophys Res Lett 43(19):10366–10376
McGillicuddy DJ, Townsend DW, He R et al (2011) Suppression of the 2010 Alexandrium fundyense bloom by changes in physical, biological, and chemical properties of the Gulf of Maine. Limnol Oceanogr 56(6):2411–2426
McManus MA, Kudela RM, Silver MW et al (2008) Cryptic blooms: are thin layers the missing connection? Estuar Coasts 31(2):396–401
Mitra A, Flynn KJ (2006) Accounting for variation in prey selectivity by zooplankton. Ecol Model 199(1):82–92
Mohlin M, Roleda MY, Pattanaik B et al (2012) Interspecific resource competition-combined effects of radiation and nutrient limitation on two diazotrophic filamentous cyanobacteria. Microb Ecol 63(4):736–750
Moore SK, Johnstone JA, Banas NS et al (2015) Present-day and future climate pathways affecting Alexandrium blooms in Puget Sound, WA, USA. Harmful Algae 48:1–11
Moore SK, Mantua NJ, Hickey BM et al (2009) Recent trends in paralytic shellfish toxins in Puget Sound, relationships to climate, and capacity for prediction of toxic events. Harmful Algae 8(3):463–477
Nimer NA, Iglesias-Rodríguez MD, Merrett MJ (1997) Bicarbonate utilization by marine phytoplankton species. J Phycol 33(4):625–631
Ogata T, Kodama M, Ishimaru T (1989) Effect of water temperature and light intensity on growth rate and toxin production of toxic dinoflagellates. In: Okaichi T, Anderson DM, Nemoto T (eds) Red tides, biology, environmental science and toxicology. Elsevier, New York, NY, pp 423–426
Okolodkov YB (1999) Species range types of recent marine dinoflagellates recorded from the Arctic. Grana 38(2–3):162–169
Okolodkov YB (2005) The global distributional patterns of toxic, bloom dinoflagellates recorded from the Eurasian Arctic. Harmful Algae 4(2):351–369
Paczkowska J, Rowe OF, Schlüter L et al (2017) Allochthonous matter: an important factor shaping the phytoplankton community in the Baltic Sea. J Plankton Res 39(1):23–34
Peacock MB, Kudela RM (2014) Evidence for active vertical migration by two dinoflagellates experiencing iron, nitrogen, and phosphorus limitation. Limnol Oceanogr 59(3):660–673
Pettersson LH, Pozdnyakov D (2013) Biology and ecology of harmful algal species. In: Pettersson LH, Pozdnyakov D (eds) Monitoring of harmful algal blooms. Springer, Berlin, pp 25–47
Pitcher GC, Horstman DA, Calder D et al (1993) The first record of diarrhetic shellfish poisoning on the South-African coast. S Afr J Sci 89(10):512–514
Preston CM, Harris A, Ryan JP et al (2011) Underwater application of quantitative PCR on an ocean mooring. PLoS One 6(8):e22522. https://doi.org/10.1371/journal.pone.0022522
Raine R (2014) A review of the biophysical interactions relevant to the promotion of HABs in stratified systems: the case study of Ireland. Deep Sea Res Part II 101:21–31
Raven JA, Geide RJ (1988) Temperature and algal growth. New Phytol 110:411–416
Rhodes LL, O’Kelly CJO, Hall JA (1994) Comparison of growth characteristics of New Zealand isolates of the prymnesiophytes Chrysochromulina quadrikonta and C. camella with those of the ichthyotoxic species C. polylepis. J Plankton Res 16:69–82
Rines JEB, Donaghay PL, Dekshenieks MM et al (2002) Thin layers and camouflage: hidden Pseudo-nitzschia spp. (Bacillariophyceae) populations in a fjord in the San Juan Islands, Washington, USA. Mar Ecol Prog Ser 225:123–137
Roemmich D, Gould WJ, Gilson J (2012) 135 years of global ocean warming between the Challenger expedition and the Argo Programme. Nat Clim Chang 2(6):425–428
Ryan JP, McManus MA, Sullivan JM (2010) Interacting physical, chemical and biological forcing of phytoplankton thin-layer variability in Monterey Bay, California. Cont Shelf Res 30(1):7–16
Schultz M, Kjørboe T (2009) Active prey selection in two pelagic copepods feeding on potentially toxic and non-toxic dinoflagellates. J Plankton Res 31:553–561
Smayda TJ (1992) Global epidemic of noxious phytoplankton blooms and food chain consequences in large ecosystems. In: Sherman K, Alexander L, Gold BD (eds) Food chains, yields, models, and management of large marine ecosystems. Westview Press, San Francisco, pp 275–307
Smayda TJ (1997) What is a bloom? A commentary. Limnol Oceanogr 42(5):1132–1136
Smayda TJ (1998) Patterns of variability characterizing marine phytoplankton, with examples from Narragansett Bay. ICES J Mar Sci 55(4):562–573
Smayda TJ, Trainer VL (2010) Dinoflagellate blooms in upwelling systems: seeding, variability, and contrasts with diatom bloom behaviour. Prog Oceanogr 85(1–2):92–107
Stocker TF, Qin D, Plattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM (2013) Climate change 2013: the physical science basis. In: IPCC (ed) Contribution of working group I to the fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge and New York, NY, p 1535
Strom SL (2002) Novel interactions between phytoplankton and microzooplankton: their influence on the coupling between growth and grazing rates in the sea. Hydrobiologia 180:41–54
Strom SL, Harvey EL, Fredrickson KA et al (2013) Broad salinity tolerance as a refuge from predation in the harmful raphidophyte alga Heterosigma akashiwo (Raphidophyceae). J Phycol 49(1):20–31
Sun J, Hutchins DA, Feng Y et al (2011) Effects of changing pCO2 and phosphate availability on domoic acid production and physiology of the marine harmful bloom diatom Pseudo-nitzschia multiseries. Limnol Oceanogr 56(3):829–840
Tatters AO, Fu FX, Hutchins DA (2012) High CO2 and silicate limitation synergistically increase the toxicity of Pseudo-nitzschia fraudulenta. PLoS One 7(2):e32116. https://doi.org/10.1371/journal.pone.0032116
Taylor M, McIntyre L, Ritson M et al (2013) Outbreak of diarrhetic shellfish poisoning associated with mussels, British Columbia, Canada. Mar Drugs 11(5):1669–1676
Trainer V, Yoshida M (eds) (2014) Proceedings of the workshop on economic impacts of harmful algal blooms on fisheries and aquaculture. PICES Sci Rep No 47, p 85
Trainer VL, Bates SS, Lundholm N et al (2012) Pseudo-nitzschia physiological ecology, phylogeny, toxicity, monitoring and impacts on ecosystem health. Harmful Algae 14:271–300
Trainer VL, Moore L, Bill BD et al (2013) Diarrhetic shellfish toxins and other lipophilic toxins of human health concern in Washington State. Mar Drugs 11(6):1815–1835
Turner JT (2006) Harmful algae interactions with marine planktonic grazers. In: Granéli E, Turner JT (eds) Ecology of harmful algae. Springer, Berlin, pp 259–270
Van Dolah ER, Paolisso M, Sellner K et al (2016) Employing a socio-ecological systems approach to engage harmful algal bloom stakeholders. Aquat Ecol 50(3):577–594
Wells ML (2003) The level of iron enrichment required to initiate diatoms blooms in HNLC waters. Mar Chem 82:101–114
Wells ML, Trainer VL, Smayda et al (2015) Harmful algal blooms and climate change: learning from the past and present to forecast the future. Harmful Algae 49:68–93
Zheng Y, Dam HG, Avery DE (2011) Differential responses of populations of the copepod Acartia hudsonica to toxic and nutritionally insufficient food algae. Harmful Algae 10(6):723–731
Acknowledgments
This is NOAA ECOHAB contribution number 920.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG, part of Springer Nature
About this chapter

Cite this chapter
Wells, M.L., Karlson, B. (2018). Harmful Algal Blooms in a Changing Ocean. In: Glibert, P., Berdalet, E., Burford, M., Pitcher, G., Zhou, M. (eds) Global Ecology and Oceanography of Harmful Algal Blooms . Ecological Studies, vol 232. Springer, Cham. https://doi.org/10.1007/978-3-319-70069-4_5
Download citation
DOI: https://doi.org/10.1007/978-3-319-70069-4_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-70068-7
Online ISBN: 978-3-319-70069-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)