
Abstract
As obligate parasites, plant viruses, require in order to survive, to be transmitted to another plant. Experimentally, viruses such as Potato virus Y (PVY) can be transmitted by mechanical means such as wounding and grafting. In its natural environment, PVY transmission is mediated by sap-feeding aphid vector, or vegetatively through propagated organs such as potato tubers. A vast number of aphid species have been reported to transmit PVY in a non-persistent manner with variable efficiency to a large number of solanaceous and non-solanaceous plant species including weeds and ornamentals. Several sensory stimuli will influence host selection and feeding behaviour of the aphid and will strongly influence virus epidemiology. The interactions between the virus, its vector, and the environment are complex and are the focus of many studies aiming to understand the molecular basis of these interactions and their impact on disease development. This chapter will present the current knowledge of PVY transmission, epidemiology, and its management in different countries.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

1 Introduction
Plant viruses are obligate parasites, and their survival depends on their transmission (often via a vector) from an infected plant to another plant. For the vast majority of plant viruses, their infectious cycle is tightly associated with that of their vector(s) and their host(s). When a plant is infected, a virus can persist in seed or in vegetatively propagated organs, such as potato tubers, bulbs, corms, at the end of the growing season. These offer ways for a virus to survive adverse conditions and initiate new infections during the following growing season. Plants acquire virus through two pathways. The first, termed vertical or secondary transmission, occurs when a virus moves from infected planting material, e.g., potato tuber into the growing plant and, in the case of potato, into the daughter tubers. The second pathway termed horizontal transmission occurs when a plant is infected mechanically or through a vector, usually an invertebrate such as an insect. Both pathways of infection by Potato virus Y (PVY) occur with potato but foliar symptoms on infected plants are generally more severe with vertical transmission than with horizontal transmission. Nearly 765 species of aphids (Hemiptera: Aphididae) have been reported to transmit PVY with variable efficiency. PVY has a broad host range infecting solanaceous and non-solanaceous plant species including weeds and ornamentals (Kerlan 2006). Biennial and perennial arable weeds can act as reservoirs for aphid-mediated transmission (Beemster and de Bokx 1987) and can contribute to the rapid dissemination of PVY within field crops and its persistence in the environment. The interactions between the virus, its vector, and the environment are complex and are the focus of many studies aiming to understand the molecular basis of these interactions and their impact on disease development.
This chapter is intended to give an overview of the current knowledge of the transmission of PVY, and its interaction with its aphid vectors, its host (potato) and the environment.
2 Transmission of PVY
2.1 “Natural” PVY Transmission
2.1.1 Vertical Transmission: Seed-Borne Infection
The infectious cycle of a virus in a plant host starts from a single infected cell. After an initial stage of uncoating (i.e., virion CP disassembly exposing the viral RNA genome), genome replication occurs, followed by cell-to-cell (local) movement through plasmodesmata to reach phloem vessels from where the virus will be transported in the phloem sap throughout the whole plant. As is the case for all plant viruses in infected plants, PVY movement throughout the plant will follow the source-sink partitioning of photoassimilates, resulting in systemic infection of sink tissues above and below the ground. Hence, developing leaves and other tissues, including new tubers, will gradually become infected (for a review, see Hull 2002).
Infected seed potato tubers can be an important source of PVY inoculum as they can maintain the virus within a crop and enable its spread within daughter growing crops and to other potato crops in following seasons. Analyses of seed potato certification data in Scotland revealed that PVY was four times more likely to be found in a crop derived from seed potatoes from a crop which contained plants with symptoms of PVY than a crop without any symptomatic plants (Fenton et al. 2012). PVY transmission through true seed or pollen of potatoes has not been recorded (de Bokx 1972).
2.2 Horizontal Transmission
2.2.1 Transmission by Aphids
As with about 380 other plant viruses, PVY is transmitted naturally by aphids (Nault 1997). Aphids are sap (phloem)-feeding hemipteran insects, and over 190 aphid species are known to transmit plant viruses (Nault 1997), with many species capable of transmitting more than one virus species (reviewed in Katis et al, 2007). Aphid populations can rapidly reach very high numbers, primarily through asexual (parthenogenetic) reproduction rather than sexual reproduction. Aphid multiplication is very variable and depends on aphid species, geographical location, temperature, and the presence of suitable hosts. The temperature threshold for aphid flights is around 15 °C for Rhopalosiphum padi and Aphis fabae; however, it does vary among and within species (Bale et al. 2007).
The interactions between aphids and the viruses they transmit have been studied at the physiological, cellular, and molecular levels and have emerged, perhaps not surprisingly, as very complex and closely related. The host selected by an aphid will be guided primarily by its perception of plant colour, volatiles, and, once having landed on a plant, through the ingestion of plant cellular contents (Moericke 1955; Kennedy et al. 1959a, b; Kring 1972; Pettersson 1970; Pettersson et al. 2007; Giordanengo et al. 2010). Interestingly, plant viruses can promote their own transmission by manipulating host physiology to make the plant more attractive to aphids and, in some cases, increase aphid fecundity (Holmes and Bethel 1972; Ingwell et al. 2012; Casteel et al. 2014). Moreover, it has been reported that the infection of a potato plant by PVY can affect the feeding behaviour of some aphid species (Boquel et al. 2011, 2012). In the case of Myzus persicae, the most efficient aphid vector of PVY, the landing of an aphid on a PVY-infected plant increased its ingestion of phloem sap and reduced the time spent not probing (Boquel et al. 2011), which could increase the risk of PVY spreading. Consequently, a wide variety of tri-trophic interactions can be observed within the PVY/aphid/host system.
2.2.1.1 Process of Virus Transmission by Aphid
Virus transmission by aphids occurs by transferring virions (virus particles containing the viral genome, assembled coat protein monomers, and other associated virus-encoded proteins) from infected to healthy plants. After landing on plants, aphids probe (puncture) leaves by inserting their long, flexible stylet into epidermal cells to form a salivary and food canal in both host and non-host plants. The first probes last less than a minute and seem to provide sufficient information for either plant rejection (promoting aphid flight) or plant acceptance (secretion of gelling and watery saliva, followed by progression through different feeding stages until reaching phloem vessels and uptake of phloem). These stages of the aphid-host interaction have emerged as a very complex molecular interplay between host and aphid. As for other plant-pathogens interactions, aphids are likely to deliver effector molecules inside their host which will alter cellular processes, thus enabling successful establishment and fulfilment of their life cycle (Hogenhout et al. 2009; Giordanengo et al. 2010).
The process of virus transmission can be divided into four phases. The first phase is termed the “acquisition phase” in which an aphid acquires virions from an infected plant. The second phase is the “retention phase” in which an aphid will retain and carry the virions. The third phase is the “inoculation phase” in which an aphid will release the retained virions into another host, initiating a new infection. For some viruses, another period termed the “latent phase” occurs during which an aphid has acquired virions but is unable to transmit them to another host during this period of time (reviewed by Katis et al. 2007).
Based on the duration of these different phases, the types of virus transmission by aphids can be classified as “persistent”, “semi-persistent”, or “non-persistent”. In persistent transmission, virus acquisition requires a relatively long time (hours or days) before an aphid becomes infective because most persistent viruses, e.g., polerovirus such as Potato leafroll virus, are confined to phloem tissues and can either replicate (propagative viruses) or not (non-propagative viruses) in their aphid vector. Semi-persistent viruses, such as Cauliflower mosaic virus, may be acquired within minutes or hours. However, their efficiency of transmission to a new host increases with the duration of the acquisition phase (Palacios et al. 2002).
Transmission of non-persistent viruses requires only a brief period of seconds or minutes of probing of epidermal cells by an aphid’s stylet before the virus becomes infective in the aphid. The retention of virions in an aphid foregut suggests that non-persistent viruses such as PVY are acquired by ingestion of cellular contents during probing of the epidermal cells (Pirone and Perry 2002). There is no latent period for semi-persistent viruses. Aphids rapidly lose their infectivity immediately after the acquisition phase and become non-infective fairly quickly when feeding on healthy plants. However, PVY particles can persist for more than 17 h in winged forms of A. nasturtii (Kostiw 1975). The vast majority (about 75%) of aphid-transmitted viruses are transmitted in a non-persistent manner. Non-persistent viruses include viruses of the Alfamovirus, Caulimovirus, Cucumovirus, Fabavirus, Macluravirus, and Potyvirus genera, with different virus particles shape (helical and isometric), genomes composition (mono or multipartite DNA, ssRNA+ genomes).
2.2.1.2 Identification of PVY Aphid Vectors
Since the 1980s, a total of 65 aphid species or group species have been reported as having the ability to acquire and transmit PVY in a non-persistent fashion (Table 6.1).
This list includes aphid vectors that colonise potato, such as the peach-potato aphid M. persicae or the potato aphid Macrosiphum euphorbiae (Fig. 6.1), or colonise other host plants, such as black bean aphid A. fabae, bird cherry-oat aphid R. padi, cereal aphids like the rose-grain aphid Metopolophium dirhodum and the grain aphid Sitobion avenae, but nevertheless have the ability to transmit PVY in a non-persistent manner.

(a) Myzus persicae (peach-potato aphid); (b) Metopolophium dirhodum (rose-grain aphid); (c) Rhopalosiphum padi (bird cherry-oat aphid); (d) Macrosiphum euphorbiae (potato aphid); (e) Sitobion avenae (grain aphid); (f) Brachycaudus helichrysi (leaf curl plum aphid) (Panels a and d courtesy of SASA, Edinburgh, UK Crown copyright. Panels b, c, e and f courtesy of Rothamsted Research, Harpenden, UK)
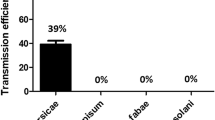
All these aphid species exhibit various degrees of efficiency in transmitting PVY. This capacity is defined by a relative efficiency factor (REF). The principle of REF determination for PVY transmission to potato relies on catching live alate aphids of different species present in potato crops, starving them for a period, allowing them to probe a PVY-infected potato and finally transferring each individual aphid to a virus-free potato plant or another bait plant. The transmission rate is calculated by dividing the total number of bait plants infected for each aphid biotype by the total number of plants infected for the M. persicae reference biotype. REF values are then calculated for each aphid species by dividing the transmission rate value of each individual species by the value of M. persicae transmission rate (which will have a value of 1). Since 1980s, several researchers have applied this methodology to define the REF factor for specific aphid species and isolates of PVYO, PVYN strain groups, and PVYNTN and PVYN-Wi variants (Kostiw 1979; van Hoof 1980; van Harten 1983; Sigvald 1984, 1992; Harrington and Gibson 1989; de Bokx and Piron 1990; Halbert et al. 2003: Verbeek et al. 2010) (Table 6.2).
However, differences in REF values for specific virus aphid combinations can be observed, e.g., Acyrthosiphon pisum and PVYO. Verbeek et al. (2010) concluded that the two most important factors influencing an assessment of an REF value were biotype of an aphid species and isolate of PVY, while acknowledging that a number of difficulties still occurred in standardising this type of experiment, particularly virus load in source plants. Differences in transmission rates as measured by REFs values among biotypes of an aphid species suggested that genetic variability among biotypes may impact on the transmission of PVY (data not shown, Verbeek et al. 2010). Moreover, as observed with A. pisum, A. fabae, Aphis nasturtii, Aphis spp., Phorodon humuli, and R. padi, recombinant isolates of PVY (PVYNTN and/or PVYN-Wi) tended to be more efficiently transmitted than non-recombinant (PVYN) isolates (Verbeek et al. 2010). These differences may occur because the transmission of PVY depends on specific interactions between receptors present in an aphid’s stylet and the PVY helper component proteinase (HC-Pro) and virion coat protein (CP). Consequently, mutations within specific sites in at least one of these two viral proteins could impair the capacity of aphids to transmit a virus (Blanc et al. 1998). Fox et al (2016) used the Verbeek method as a standardised approach to investigate PVY virus strains and aphid biotypes in the UK. This study found that two species, Cavariella aegopodii and R. padi, were able to transmit PVYNTN isolates almost as efficiently as M. persicae. This study also highlighted that the largest source of variability in transmission efficiency within a given aphid species was the virus source plant, confirming the findings from Verbeek et al. (2010). Although differences in REFs were recorded among the various laboratories, M. persicae remained the most efficient aphid vector of PVY with the greatest REF value, irrespective of the isolate tested.
In addition to these studies, PCR-based methods have been used to detect PVY in individual aphids (He et al. 2006), even in their stylets (Zhang et al. 2013), enabling species that may act as potential vectors to be identified. This approach was applied to preserved viral RNA from aphids caught in pan traps in potato crops (Nie et al. 2011; Pelletier et al. 2012). Aphid catches from the various crops were amalgamated and sorted by species. The stylets of a maximum of 12 aphids from each sample were detached from their body parts and stored for RT-PCR detection of PVY. The remaining corresponding aphid body parts were kept for taxonomic identification using a DNA barcoding approach. The sequence of the cytochrome c oxidase subunit 1 (COI) barcode region was compared with a set of ~6600 aphid sequences representing 730 species in the Barcode of Life Data Systems project (Foottit et al. 2008). Using this approach, 65 aphid taxa were identified from the samples that tested positive for PVY. Among these, 45 taxa had never been previously recorded as being able to transmit PVY and 7 had previously been labelled as non-vectors (Pelletier et al. 2012). These results indicate that many aphid species that feed on PVY-infected plants can carry PVY particles. However, detecting PVY in aphids by RT-PCR or other methods does not mean that a specific aphid is able to transmit a virus; further experiments are required to establish whether or not probing by these PVY-containing aphids can result in infection of a healthy potato plant.
Knowledge of the REF values of aphid species and the number of aphids caught in traps is of practical importance for virus management because it allows the cumulative vector pressure to be calculated and the risk of PVY transmission to potato crops to be forecast (van Harten 1983, Sigvald 1986). Indeed, while some non-potato colonising aphid species have relatively low REF values, their importance as PVY vectors is directly dependent on their phenology, because large numbers of aphids with an attributed small REF value may contribute significantly to PVY transmission in crops (DiFonzo et al. 1997; Boiteau et al. 1998; Pickup et al. 2009; Kirchner et al. 2011; Fenton et al. 2012, Fox et al. 2016). In addition to how efficient an aphid transmits virus (REF values), virus spread in potato crops will be influenced by the activity of vector species and their behaviour. These parameters need to be taken into account in virus forecasting models and control systems (Sigvald 1986). These elements will be discussed in this chapter.
2.2.1.3 Molecular Mechanisms Involved in PVY- Aphid Interactions
As with all non-persistent viruses, PVY has a basic simple virion structure with its RNA encapsidated in rod-shaped particles by a single type of CP that is required for PVY transmission. PVY requires a non-structural protein or “helper component”, the helper component proteinase (HC-Pro), which acts in its homodimer active form (Thornbury et al. 1985) as a molecular “bridge” between the virions and an aphid’s mouthparts. In relation to aphid transmission, HC-Pro N-terminal domain (KITC – Lysine/Isoleucine/Threonine/Cysteine) is involved in specific binding to an aphid’s stylet (Blanc et al. 1998), more precisely with receptors located in the extreme tip of the stylet called the acrostyle (Uzest et al. 2007), while its C-terminal domain (PTK – Proline/Threonine/Lysine) has been shown to be involved either directly or indirectly in HC-Pro binding to the DAG motif (Aspartic acid/Alanine/Glycine) at the CP N-terminus (Fig. 6.2). Mutagenesis studies performed on the N-terminal region of the CP protein of other non-persistent viruses have demonstrated that amino acids in the vicinity of the DAG motif influence the transmission efficiency of aphids (Pirone and Blanc 1996).

Schematic representation of the molecular interactions between a PVY particle and an aphid stylet
HC-Pro is a multimodular protein with numerous functional domains such as a zinc-finger motif involved in viral synergism with co-infecting viruses (Vance et al. 1995; Pruss et al. 1997), nucleic acid/RNA binding domains (Urcuqui-Inchima et al. 2000), peptidyl domains involved in viral genomic RNA replication and systemic movement (Cronin et al. 1995; Kasschau et al. 1997) and cysteine-protease-like domain (Oh and Carrington 1989; Guo et al. 2011). Apart from mediating aphid transmission, HC-Pro is involved in almost all processes of the viral cycle such as movement through plasmodesmata, cell-to-cell and long-distance Potyvirus movement in a plant (Kasschau and Carrington 2001; Sáenz et al. 2002), pathogenicity (Pruss et al. 1997) and suppression of antiviral RNA silencing (Llave et al. 2000).
Studies of the acquisition of purified, labelled, non-persistent viruses by aphids demonstrated that successful transmission of PVY by individual aphids of M. persicae to potato plants required fewer than 50 virus particles (Pirone and Thornbury 1988). In a more recent study using a competition system between infectious and non-infectious PVY isolates, it was estimated that aphid transmission of fewer than four PVY particles is sufficient to generate a successful infection (Moury et al. 2007).
2.2.2 Sources of PVY Inoculum
2.2.2.1 Volunteers and Ware Potato Crops
Volunteer plants, i.e., potato plants growing from true seed or potato tubers or parts of a tuber left in the soil after the preceding crop, can be a source of PVY inoculum and, if not adequately controlled, may present a risk for PVY transmission to potato crops growing in the vicinity. In England the incidence of PVY in volunteer potatoes and in ware crops of cv. Record, susceptible to PVYN, were assessed over 2 years by Jones et al. (1996). In the second year of the survey, volunteer potatoes were found to account for 4–8% of the emerged potato plants in the crops at four sites, and 31–93% of these plants were found to be infected predominantly by PVYN. Other studies have reported that virus incidence (including PVY) in ware potatoes can be extremely variable ranging from 1 to 60% (Chatzivassiliou et al. 2008). Management of inoculum sources in ware crops and volunteer potatoes is very important in minimising the risk of virus transmission to potato seed crops, particularly PVY to pre-basic category seed potato crops and will be addressed in this chapter.
2.2.2.2 Alternative Hosts of PVY
The host range for PVY includes mainly solanaceous crops such as Nicotianae spp. (Nicotiana tabacum, Nicotiana benthamiana), pepper (Capsicum spp.), and tomato (Solanum esculentum). PVY can also infect weeds such as Solanum sarrachoides (hairy nightshade) (Srinivasan et al. 2008), Solanum nigrum (black nightshade) (Chickh-Ali et al. 2008), and ornamentals such as Physalis floridana (Beemster and de Bokx 1987; Kerlan 2006). Kaliciak and Syller (2009) identified a range of commonly occurring arable weeds such as Erodium cicutarium (redstem filaree), Geranium pusillum (small-flowered crane’s bill), Lactuca serriola (prickly lettuce), and Lamium purpureum (purple deadnettle) that could be infected mechanically by PVY. The virus has also been detected in other weeds such as Plantago major (plantain), Taraxacum spp. (dandelion), and Sonchus spp. (sow thistle), Fallopia spp. (knotweed), Senecio spp. (common groundsel), Chenopodium album (goosefoot) (R. van der Vlugt, personal communication). Their importance in the secondary transmission of PVY and as reservoir of the virus is likely to vary in different regions of the world where cultural practices differ. This aspect is discussed in Chap. 6.
2.2.2.3 Water as a Source of PVY Inoculum
Water sources such as canals, rivers, streams, ponds, lakes and oceans are known to be reservoirs of many viruses. Plant viruses found in natural water sources belong mainly to seven groups, but there is limited evidence about their survival in water and their potential for water-borne infection (reviewed by Mehle and Ravnikar 2012). There is minimal evidence of water transmission of potyviruses, specifically of PVY.
Recently, Mehle et al. (2014) investigated PVYNTN survival in water and its water-mediated transmission. Survival of PVYNTN in water was monitored under controlled conditions using macerated infected leaves in tap water. Mechanical inoculation of test plants with a PVY-infested aqueous solution confirmed that PVYNTN could remain infectious in water for up to 1 week at 20 ± 4 °C. When the solution was stored at 4 °C, PVYNTN remained infectious for up to 10 weeks. The role of water as a source of PVY inoculum was also assessed experimentally using a hydroponic system. Inoculated potato plants were placed in a glass tank filled with nutrient solution while healthy (bait) potato plants were placed in separate tanks. Bait plants were irrigated with nutrient solution from the tank containing inoculated plants. PVYNTN was detected in the nutrient solution in the first month after starting the experiment. The virus was also found in the roots of bait plants in the first month and in the foliage 54 days after initiation of the experiment. After 4 months when the experiment was concluded, PVYNTN was detected in two of the six potato bait plants. In addition, it was demonstrated that PVYNTN could be released from injured tomato roots into the nutrient solution and could infect healthy tomato plants through the roots, ultimately spreading to the foliage, where it was detected more than 3 months after initiation of irrigation with the PVY-infested nutrient solution. These experiments suggest that while irrigation is not the most efficient means of transmission of PVY between plants, it can potentially represent a source of PVY inoculum at least in hydroponic systems. Hydroponic systems and/or intensive irrigation are widely used in commercial potato production; therefore, water could be an important source of PVY infection in such systems.
2.3 “Experimental” PVY Transmission
There are several “artificial” or “experimental” methods to transmit PVY. In most of the cases, these are used for research purposes, i.e., studying the pathogenicity of PVY, characterising the resistance status of a given genotype of potato. Nevertheless, in some cases, this could represent a potential means of PVY dissemination in favourable conditions.
2.3.1 Mechanical Transmission
Healthy plants can be infected by PVY by manually rubbing infectious sap from a PVY-infected plant (obtained by grinding infected material with a buffered solution) onto the leaf surface together with a fine abrasive powder (such as aluminium oxide or carborundum). This method, while requiring a relatively high titre of PVY in the sap, is a convenient way of experimentally transmitting PVY to a range of host plants for studying the pathogenicity of PVY strains (see Chap. 2). For efficient transmission, it is preferable to use freshly infected biological material and young plantlets at 4–6-leaf stage. Using this method, disease symptoms usually develop 7 days after inoculation. If viral inoculum originates from freeze or dried material, the transmission rate is generally reduced and a delay in the symptom expression can be expected.
2.3.2 Transmission by Wounding
Under environmentally controlled conditions, Fageria et al. (2015) found that PVYO, PVYN:O, and PVYNTN strains could be transmitted by different types of wounding (i.e., squashing or squeezing leaf and stems or leaf petioles, repeated contacts of leaves through air flow between infected and bait plants). However, PVY was not transmitted by the process of cutting seed potato tubers. This study showed that wounding inflicted by the manipulation of plants in the greenhouse could result in PVY transmission and spread under particularly favourable conditions (Fageria et al. 2015; de Bokx 1972). However, de Bokx (1972) reported previously that, under field conditions, transmission of viruses through wounding is not as efficient as under greenhouse conditions, perhaps because plants are less brittle and PVY infectiveness is significantly reduced outdoors.
2.3.3 Transmission by Grafting
PVY can also be transmitted from infected to healthy plants by grafting. This method consists of joining together an infected piece of potato stem with another solanaceous plant such as potato, tobacco, tomato, or pepper. An apical scion from a virus-infected plant is placed into a slit cut the stem of the stock plant so that a vascular junction is created allowing PVY to move in the phloem sap into the healthy plant (Fig. 6.3). For PVY, this approach is essentially used to study the reaction of potato genotypes to infection by PVY. If a potato cultivar carries the hypersensitive resistance N gene, top necrosis symptoms will be observed in the grafted stock.

Representation of the grafting method of potato on to potato. Photographs were provided by J.P. Dantec (INRA, Ploudaniel, France). (1) a healthy potato plant is used as stock; (2) select an apical part of a virus-infected potato plant to prepare the scion, cut downward away from the apex to expose a large area of stem tissue; (3) split the stock’s stem at mid-height, keep the split slightly open, insert the scion to ensure the cambia are in contact, and hold the assembly together with Parafilm; (4) wait a few days to be sure the graft has taken (Adapted from de Bokx 1972)
3 Epidemiology and Dissemination of PVY
3.1 Characteristics of the Transmission and Dissemination of PVY Strains Under Field Conditions
The prevalence of PVY in cultivated potato crops depends on many biotic and abiotic factors. As detailed below, the PVY spread depends essentially on the incidence of PVY inoculum in the seed potatoes, climatic conditions, the genetic background of the potato cultivar (i.e., relative resistance to PVY strains), and, as discussed earlier, the pressure from PVY vectors between plant emergence and haulm destruction prior to harvest (Rolot 2005; Steinger et al. 2014). In the USA, Nolte et al. (2004) reported that the incidence of PVY in daughter tubers derived from virus-free seed potatoes varied significantly among cultivars ranging from 1.9 to 13.2%. While REF values for aphid species and PVY strains (Verbeek et al. 2010) have been assessed in environmentally controlled transmission experiments, transmission of PVY strains in the field will be affected by various parameters, e.g., temperature, aphid phenology, and the abundance of sources of inoculum.
Information on the dynamics of transmission of various PVY strains in the field is limited. The transmission of five isolates of four PVY strains was studied in small plots, each consisting of 50 plants with 4 replications, over a 3-year period (Dupuis and Schwaerzel 2011). Plant infection was assessed by testing a sample of tubers collected from each plant at harvest time. The incidence of plant infection varied significantly among strains and cultivars (Fig. 6.4). The PVYN-Wi strain produced the highest incidence of plant infection while infection by PVYO strain resulted in a significantly lower incidence. There was no significant difference in the incidence of plant infection for the PVYN, PVYNTN, and PVYN-Wi strains. However, the incidence of infection for the PVYNTN strain was greater for isolate 1080 than for isolate 1150, indicating that differences in transmission characteristics may exist among isolates of the same strain. No interaction was detected between the year of experiment and the incidence of plant infection produced by the PVY strains. A second study in larger plots, each of consisting 100 plants with 6 replications, was undertaken over a 2 year period in order to compare the transmission efficiency of a PVYN-Wi and a PVYNTN strain (Dupuis et al. 2014). The incidence of plant infection on the two cultivars was greater for PVYN-Wi strain than for PVYNTN strain over both years of the experiment (Fig. 6.5). These results suggest that the transmission of PVY strains to potato crops may vary significantly among strains and cultivars with an overall risk of greater transmission with PVYNTN and PVYN-Wi strains than with PVYO, potentially explaining the increased prevalence of these variants in potato crops.

The incidence of plant infection caused by five PVY strains (PVYN-Wi isolate 1121, PVYNTN isolate 1080, PVYNTN isolate 1150, PVYN isolate 605 and PVYO isolate 803) on six potato cultivars, as assessed by testing daughter tubers collected at harvest (mean of a 3 year trial). Letters in brackets shows the homogeneity groups of the Newman–Keuls mean comparison test

The incidence of plant infection caused by PVYNTN and PVYN-Wi strains in cultivars Nicola and Charlotte. The incidence was assessed by testing daughter tubers collected before haulm killing and 4 weeks later at harvest time. Error bars show the standard error of the mean
3.2 Modelling Approach to Determine the Relative Importance of Aphid Species in Transmitting PVY
Modelling can be used to predict the relative importance of aphid species known to be PVY vectors. In the high grade (HG) seed potato production zone in Finland, over 30,000 winged aphids from 6 or 7 potato fields were caught over three growing seasons using yellow pan traps that were emptied twice a week (Kirchner et al. 2011). Of the aphids caught, 37% belonged to nine species known to transmit PVY. The seasonal increase of PVY (incidence of PVY in harvested daughter tubers compared with that in seed tubers), aphid counts in traps, aphid REF values, and PVY resistance status of potato cultivars were used as explanatory variables. Early-season flights of aphids were the most important factor accounting for the incidence of PVY. Aphis fabae was found to be the most important vector, because only models including this aphid species showed a strong statistical model fit with the incidence of PVY in the harvested tubers (Kirchner et al. 2011).
In a different study in Scotland, a similar approach was adopted by correlating weekly incidence of virus in experimental test plants and weekly aphid catches over several years (Pickup et al. 2009; Fenton et al. 2012). A logistic regression model based on binomial response data was used to relate PVY infection to aphid counts for individual species. The cereal aphids M. dirhodum and S. avenae were identified as the most important PVY vectors (Fig. 6.6).

Weekly incidence (%) of PVYO in bait plants (thick black line), suction trap counts for M. dirhodum (red line) and S. avenae (blue line), and index of vector pressure (dashed black line) over 3 years (Fenton et al. 2012; Pickup et al. manuscript in preparation)
3.3 Aphid Monitoring, Phenology, and Current Status of PVY Transmission Worldwide
3.3.1 Aphid Monitoring and Aphid Trapping Techniques
The association of virus spread in potatoes with aphid flights is documented in numerous publications (Ashby 1976; Boiteau et al. 1988; DiFonzo et al. 1997; Harrington et al. 1986; Robert et al. 2000; Basky 2002; Radcliffe and Ragsdale 2002; Sigvald 1989; Steinger et al. 2015; Kirchner et al. 2011).
The activity of aphids can be monitored using several techniques:
-
Sticky fishing-line traps made of a wooden frame and transparent sticky polyamide threads, the frame being fixed vertically and oriented into the wind (Labonne et al. 1983)
-
Traps which collect insects flying in the air by suction at a specific flow rate (Fig. 6.7).
Fig. 6.7 
Suction trap (Rothamsted-type 12.2 m high) (right panel) and water pan trap (right panel) currently used for aphid monitoring (Courtesy of SASA, Crown copyright)
-
Coloured pan traps (Moericke 1951) which attract and trap alate aphids (water pan traps or sticky tile traps) (Fig. 6.7).

Suction traps and sticky fishing-line traps do not rely on attracting aphids and thus sample aphids in an unbiased way. The latter can easily be placed in crops at the height of foliage providing information on aphid activity within crops. Suction traps which are around 12 m high (Johnson and Taylor 1955) might not reflect exactly the abundance and composition of aphid populations in specific crops; however, it is generally accepted that suction traps can give a good representation of aphid phenology for larger areas (up to 30–60 km wide) and are easier to manage on a daily basis. Suction traps can provide information on aphid populations more rapidly and are widely used as a warning system for the risk of virus transmission. For more localised studies, i.e., the spread of a virus in a crop, the use of trap(s) positioned in a crop is recommended.
Strong correlations between aphid catches in suction traps and yellow pan traps have been found in Belgium, even when traps were several kilometres apart (Rolot 2005). Similar studies conducted in other locations showed that the range of aphid species and numbers of each species collected differed between yellow pan traps and suction traps (Robert et al. 1987; Labonne et al. 1989, Seco et al. 1990). Numbers of A. fabae and M. euphorbiae were greater in yellow water traps than in suction traps, while M. persicae and M. dirhodum made up a greater proportion of aphids caught in suction traps.
The efficacy of green (GWT) and yellow (YWT) water pan traps and suction trap has been assessed in the Ardennes region of Belgium. Water pan traps were positioned in three different fields of potato with a suction trap (ST) situated within a few hundred metres to 3 km from the fields. Aphids were counted and identified over a 10-week period. A significant difference was found between YWT and GWT in their efficacy in catching aphids (F(1,2), P = 0.0065), with more being caught in YWT than in GWT (Fig. 6.8).

Total number of aphids caught in yellow water traps (YWT) and green water traps (GWT) in three locations in Belgium between weeks 21 and 31 (left panel). Relative abundance of each of the 9 selected species expressed as a percentage of total number of aphids caught in YWT, GWT, and suction trap (ST) (right panel)
The relative abundance of each species in each type of water trap (YWT, GWT) was also analysed by testing the interaction “Species × Colour” against “Species × Colour x Sites”. These results suggest that, at least in this case, trap colour might not have had a significant impact on the relative proportion of aphid species caught in each potato field (F(8,16), P = 0.3849, data not shown). However, other reports on aphid response to colour in field experiments suggest that colour, brightness, and contrast with surroundings crops might influence aphid trapping (Baldy and Rabasse 1983; Doring and Chittka 2007). The data from the water pan traps were also compared with suction trap data (Fig. 6.8 right panel). A discrepancy in aphid trapping efficacy between ST and YWT or GWT was observed for Aphis spp., Brachycaudus helichrysi, M. persicae, R. padi, and S. avenae. This might reflect differences in flying patterns and behaviour among various aphid species. In addition, because that M. persicae is the most efficient vector of PVY, the difference between traps may be important in predicting the risk of PVY transmission in potato crops.
3.3.2 Cases Studies on the Current Status of PVY Transmission and Aphid Species Phenology Worldwide
3.3.2.1 Finland
The HG seed potato production zone in Finland (latitude 64°) is one of the northernmost intensive crop production areas of the world. PVY was not common there until 2005 when many seed potato lots were down-graded in certification due to a sudden increase in the incidence of PVY, mainly PVYNTN-like strains. In a study conducted between 2007 and 2012, aphids were monitored in potato crops in the HG zone using yellow pan traps. Over 58,000 aphids covering 83 aphid taxa were caught and 34 species were further characterised by DNA barcoding. The number of aphids caught peaked every 3 years in a recurring cycle. Most of the agriculturally relevant species occurred late in the potato growing season and did not contribute significantly to the spread of PVY (Kirchner et al. 2013). However, A. fabae was one of the most common species caught in yellow traps being found mainly in the first 2–3 weeks after emergence of the potato plants when they are most susceptible to virus infection. A. fabae is known to be a vector of PVY, and modelling analysis determined that it was the main vector of PVY in the HG zone (see Chap. 4.3; Kirchner et al. 2011). The snowball tree (Viburnum opulus) grown in gardens is the only known winter host of A. fabae there. Based on this information, four approaches were designed to reduce PVY infection pressure on seed potato crops in the HG zone: (1) Straw mulch to be spread on potato crops at plant emergence using a bale chopper in order to reduce the landing of A. fabae on plants (Kirchner et al. 2014). This new practice was readily adopted by growers because barley is grown as an intercrop in the seed potato farms and cattle farmers use the straw and have bale choppers. (2) The seed potato growers sponsored and conducted a campaign to replace snowball trees in gardens with other ornamental species, in order to reduce the local overwintering population of A. fabae. (3) Production of seed potatoes of the potato cultivar most commonly grown in Finland was terminated because it did not show symptoms when infected by PVYNTN-like strains and roguing of infected plants was, therefore, not possible. The cultivar was considered to be a major reservoir of PVY and a source of PVY inoculum for seed potato crops in the HG zone. 4) The recurring triennial cycle of aphid abundance in the HG zone was utilised in planning the production of seed potatoes of cultivars susceptible to PVY. Since these proposals were implemented, the incidence of PVY in seed potato crops has decreased in the HG zone and the downgrading of seed potato lots at certification is now as rare as it was prior to 2005.
3.3.2.2 Switzerland
In Switzerland, the flight activity of aphids has been monitored since 1987 using a standard Rothamsted suction trap located at the Agroscope Agricultural Research Station in Nyon. A second trap located in Reckenholz (Zürich) ceased operation in 2010. The traps are part of the Swiss seed potato certification scheme managed by Agroscope, who also contribute the aphid data to the European EXAMINE project. The following species are identified and counted on a daily basis during the main flight period of aphids (April–November): A. pisum, A. fabae, Aphis sp., Aulacorthum solani, Brachycaudus helichrysi, Brevicoryne brassicae, Cavariella sp., M. euphorbiae, M. dirhodum, Metopolophium festucae, Myzus ascalonicus, M. persicae, Phorodon humuli, Rhopalosiphum insertum, Rhopalosiphum maidis, R. padi, and S. avenae. In addition, the total number of aphids of the family Aphididae and non-Aphididae are counted.
Aphid data from the suction trap is used to calculate vector pressure, an indicator for the risk of PVY transmission in seed potato crops. Information is updated weekly and published on the internet platform “Agrometeo” (www.agrometeo.ch) to inform growers about virus risk. Furthermore, the aphid data is fed into a forecast model which predicts the incidence of tuber infection by PVY under a range of epidemiological conditions using factors such as cultivar, presence of virus inoculum in seed potatoes, number of mineral oil applications, altitude of the field, and date of haulm destruction. The relative contributions of these factors to PVY development in potato crops were obtained from a statistical analysis of compiled seed certification data spanning two decades (Steinger et al. 2014). The seed potato certification body at Agroscope uses model prediction along with field observations to set the optimal date of haulm destruction, which growers are obliged to follow.
The initial model employed to forecast post-harvest PVY incidence in Switzerland (named TuberPro) used vector pressure estimated from catches of 11 aphid species in suction traps as one of the input variables (Nemecek 1993). Aphid counts were weighted using REF values for each species. Myzus persicae had the highest weighting. Recently, a simple linear regression model, using the cumulative number of B. helichrysi caught as a predictor variable, was found to provide a more accurate forecast of PVY risk than M. persicae (Fig. 6.9) (Steinger et al. 2015). This new model superseded TuberPro in 2013. B. helichrysi is an aphid species that does not colonise potato plants but flies relatively early in the growing season, which may at least partly explain its importance as a vector of PVY. Interestingly, the abundance of M. persicae, often considered to be the main vector of PVY, was not associated with PVY incidence in Swiss seed potatoes (Fig. 6.9).

Relationship between the incidence of PVY in tubers of susceptible potato cultivars and suction trap catches of Brachycaudus helichrysi and Myzus persicae in Switzerland. Dots represent the annual percentage of PVY-infected tubers averaged over all tested seed lots (N = 150–611 seed lots per year) (Adapted from Steinger et al. 2015)
3.3.2.3 Slovenia
While data on the aphid species responsible for PVY transmission in Slovenia is not currently available, their occurrence in potato crops was assessed between 2003 and 2006 in the four main areas of seed potato production: Komenda, Jablje, Øentvid pri Stioeni, and Libelice. Yellow water (Moericke) traps were used to determine the number and species of aphids present in potato crops during the growing season (Modic and Urek 2008). The majority of aphids of the family Aphididae that were caught belonged to the genus Aphis. However, some species previously unreported in Slovenia were recorded: Aphis spiraecola (Patch), Amphorophora gei (Börner), Chaitophorus leucomelas (Koch), Chaitophorus populeti (Panzer), Drepanosiphum aceris (Koch), Cinara sp., Macrosiphum cholodkovskyi (Mordvilko), Macrosiphum gei (Koch), Myzocallis castanicola (Baker), Myzocallis coryli (Goeze), Protrama flavescens (Koch), Protrama ranunculi del Guercio, R. maidis (Fitch), Thelaxes dryophila (Schrank), Therioaphis luteola (Börner), Trama rara (Mordvilko), and Tuberolachnus salignus (Gmelin). On the basis of previous publications and this survey, 180 species of true aphids of the family Aphididae have now been identified in Slovenia. In this study, 197 aphid species were found in seed potato crops.
3.3.2.4 France
France, as with 18 other European countries, has collaborated since 2000 in the EXAMINE programme (Exploitation of Aphid Monitoring in Europe – an EU-funded project within the Concerted Action Programme, with the remit of collecting national data on aphid distribution, phenology and abundance throughout Europe) to monitor aphid flights with a network of suction traps. In addition, the National Federation of Seed Potato Growers (FNPPPT, Fédération Nationale des Producteurs de Plants de Pomme de Terre) together with its regional producer organisations, established in 1970s an aphid trapping network of yellow water (Moericke) traps in the three main seed potato production areas. Currently, there are 18 traps: 5 in the west, 10 in the north, and 3 in the centre and the south of France. Trapping is done during the potato growing season, from April to the end of July, and the content of traps is analysed twice a week to determine the number and species of aphids. However, not all specimen are identified to the species level. Routinely, the assessment focuses on 11 species and/or group species known to transmit PVY: four that colonize potato (M. persicae, Aulacorthum solani, M. euphorbiae, Aphis spp.) and seven that do not colonise potato (R. padi, S. avenae, M. dirhodum, A. pisum, Brevicoryne brassicae, Phorodon humuli, Cavariella sp.). The data on the flights of these aphids is transmitted to potato inspectors and seed potato producers by fax with a recommendation on protection measures appropriate for the aphid pressure. In addition, this information is also integrated into plant health news bulletins which alert growers on pests present in crops. In 2006, the monitoring of nine yellow traps from west, north, and centre of France showed differences in the aphid species recorded at the various sampling sites (Fig. 6.10) (Le Hingrat 2007). M. persicae was the most abundant aphid species caught in the west of France, whereas in the north and the centre, the most abundant aphids were mainly the non-colonising M. dirhodum and Cavariella sp. In 2014, in west area, the situation was reversed compared with 2006, namely, a preponderance of non-colonising aphid species (M. dirhodum, Cavariella sp., Aphis spp., S. avenae). It was concluded that aphid species diversity depends on environment factors, notably the climate and neighbouring crops.

Relative abundance of some aphid species caught in 2006 and 2014 in yellow water traps in the three main seed potato areas of France (West, North, Center-South)
Although aphid flights can be monitored and the efficiency of an aphid species to transmit PVY can be evaluated, it is still a challenge to determine accurately those aphid species responsible for causing PVY infection of potato crops. Indeed, the latent period between aphid transmission of PVY to a plant and the expression of symptoms, (or when the PVY infection could be detected by ELISA), is about 10–15 days in the environmental conditions in France. To improve knowledge of the interactions between aphids, PVY, and the environment and to identify the parameters involved in PVY epidemiology, monitoring of PVY infection in seed and ware potato crops was undertaken as part of a 3-year research project (INRA-Rennes-Le Rheu and FNPPPT) (Boisgontier et al. 2013). Factors associated to landscape, meteorological data (temperature, rainfall, wind speed, and direction), aphid flights (number, species), and potato PVY infection were monitored, e.g., meteorological data every 15 min each day, PVY infection survey once per week, and aphid catches three times per week. Depending on the season, 18–42 aphid species or group species were identified in the trap samples from potato crops, with M. persicae, M. dirhodum, A. pisum, and Brevicorynae brassicae being the most prevalent aphid species probably due to the proximity of cereal, maize, and rape crops in the vicinity of the potato crops. The analysis of quantitative and qualitative data linked to aphid flights and PVY infection in crops identified the presence of aphids throughout the growing crop season as a key factor in the incidence of PVY (Fig. 6.11). It was concluded that modelling PVY incidence at post-harvest in relation to the initial PVY incidence of the parent crop, the spread of PVY infection, and aphid phenology is very complex and additional parameters such as environmental conditions affect the model significantly. However, cultural practices (using or not insecticide and mineral oil) and initial quality of planted seed are two of the most important parameters affecting PVY incidence post-harvest.

Relationship between prevalence of aphids (black line) and PVY infection (red circle) in a ware potato crop from the planting to haulm killing. The green circle corresponds to the incidence of PVY in seed potatoes. Aphid species listed in the graph correspond to those most commonly identified in peaks of aphid numbers
3.3.2.5 South-Africa
The South African aphid monitoring network consists of 13 Rothamsted-type suction traps distributed in the Western Cape, Northern Cape, Free State, North West, and Limpopo. The national network was initiated in 2005 by Potatoes South Africa and is coordinated by the University of Pretoria. Aphid assessments are conducted on a daily or weekly basis. Bulletins are issued to growers to alert them to the risk of virus spread. These provide information on the activity of aphids and cumulative vector pressure based on the abundance of aphid species and their REF values following van Harten (1983) (mean values derived from published literature and South Africa unpublished results), together with weekly SMS notifications. The suction trap network is funded by Potatoes South Africa, the Western Cape Department of Agriculture, the KwaZulu-Natal Department of Agriculture, the Winter Cereal Trust, the Agricultural Research Council, and the Technology and Human Resources for Industry Programme (THRIP). In addition, yellow water traps are used locally to monitor the prevalence of aphids in potato crops.
The most important of the 79 species or species groups identified are A. pisum, Aphis gossypii, and other Aphis spp., M. euphorbiae, M. persicae, and the cereal aphids M. dirhodum, R. padi, and S. avenae. Their contribution to vector pressure varies with region. For example, in some areas in the summer rainfall region, such as the western Free State (summer crop), which is dominated by grassland, R. padi is the most abundant vector of PVY, followed by Aphis spp. In KwaZulu-Natal (summer crop), R. padi, A. gossypii, and other Aphis spp. are the most abundant vectors. In the Sandveld in the Western Cape, a part of the winter rainfall region, seed potatoes are planted in March (winter crop), although potatoes are grown throughout the year. In this region, M. euphorbiae, M. persicae, A. pisum and the cereal aphids are important vectors of PVY in seed potatoes, but PLRV is considered to be a greater threat than PVY. Although R. padi is the most abundant aphid species in most of the regions monitored, it is a weak vector of PVYNTN. In tests with individual adult aphids, none transmitted the PVYNTN isolate to healthy potato seedlings, whereas the transmission efficiency for M. persicae for the same isolate was 0.33 (K. Krüger, unpublished data).
A simple model was developed to determine the influence of changes in climate on the abundance of M. persicae, the most efficient vector of PVY and thus indirectly on the risk of virus spread to potatoes (van der Waals et al. 2013). The results suggest that the South African seed potato industry may incrementally experience greater problems with PVY in the future due to an increase in temperature during the potato growing season and consequently in aphid abundance over the 90-year period in the areas modelled (Sandveld, Eastern Free State and Limpopo; Fig. 6.12). Although the results suggest an increase in aphid abundance, population growth may be limited by high temperature in areas such as Limpopo and the Sandveld, where aphids may reach their thermal limits during the hottest months. Shifts in main planting times for potatoes in some regions may lessen vector pressure and consequently the risk of PVY transmission.

Cumulative relative development rates (cRDR) of Myzus persicae as vector of PVY and PLRV on potatoes for the periods 1961–1970 (••••••), 2001–2010 (—–) and 2041–2050 (----) in (a) the Sandveld (winter rainfall), (b) the Eastern Free State (summer rainfall) and (c) Limpopo (summer rainfall) regions in South Africa (Adapted from van der Waals et al. 2013)
3.3.2.6 Scotland/UK
In Scotland, Science and Advice for Scottish Agriculture (SASA-Scottish Government) provides a diagnostic service for viruses in seed potato crops (growing crops and post-harvest) and is part of a network of suction traps operated by Rothamsted Research (England, UK) in England and Scotland. Information on the weekly catches of aphids is published in a bulletin as part of the Rothamsted Insect Survey and advice is also provided on the risk of virus transmission and the need for aphid control. The data is contributed to the EU EXAMINE program. An Aphid Monitoring programme was introduced into the Seed Potato Classification Scheme in 1992 to identify seed potato crops in which potato aphids had been poorly controlled. Subsequent classification of seed potatoes from these crops was dependent upon a satisfactory post-harvest tuber test for the presence of viruses. This programme was discontinued because it was relatively unsuccessful in identifying crops with a high risk of virus infection.
The Agriculture and Horticulture Development Board-Potatoes (AHDB-Potatoes) funds 100 water traps located in potato crops in the major seed potato growing areas of Great Britain. The contents of each trap are analysed weekly at the Food and Environment Research Agency in York (UK). Registered users receive e-mail and SMS alerts when the peach-potato aphid (M. persicae) is found in their region or when aphid catches in any trap in their region exceed a specified weekly threshold. Comparative information with previous seasons is also available.
The pattern of changing incidence of PVYN in Scottish seed potato crops closely follows the incidence of three species of cereal aphid: the rose-grain aphid (M. dirhodum), the grain aphid (S. avenae), and the bird cherry-oat aphid (R. padi). Using a model based on these three abundant cereal aphids, the incidence of PVYN in seed potatoes was found to be strongly correlated with the abundance of these aphids up until day 210 (29 July) (Fig. 6.13). The incidence of PVYN in seed potato crops in Scotland can be predicted using the incidence of PVY in the previous year and geometrical mean of the catches of these three aphids prior to day 210 in three standards Rothamsted suction traps (Elgin, Dundee and Edinburgh, UK).

Relationship between observed PVYN incidence in Scottish seed potato crops and predicted PVYN incidence based on suction trap catches of M. dirhodum, S. avenae, and R. padi
3.3.2.7 Belgium
In Belgium, aphid monitoring is operated using two suction traps located in Libramont (since 1983) and in Gembloux (since 1981) under the supervision of the Walloon Agriculture Research Center. Aphids are counted and identified on a daily basis each year from mid-April (week 16) until mid-October (week 41). For seed potatoes, a set of 12 species or species groups are followed during the cropping season based on their overall relative abundance in the catches and their efficiency for PVY transmission. The vegetative period for seed potatoes is generally between week 21 (first emergence of plants end of May) and week 31 (last date of destruction of foliage, end of July). A strong correlation was established between the abundance of flying aphids and the quality of the seed lots, especially regarding PVY which is the most prevalent virus in seed potato crops in Belgium. The downgrading rate of seed lots following post-harvest testing depends on: (1) the abundance of the selected species as measured by suction traps during the vegetative period, (2) the relative efficiency of each of the prescribed species in transmitting PVY (REF value), and (3) the mature plant resistance status of the plants (MPR-Table 6.3). These three factors enable a weekly index for the PVY infection pressure (IP) to be calculated using the equation:
-
IPj: infection pressure during week j; i….p: aphid species, Ni: aphids number of the considered species during week j, r.e.fi : REF for species; MPRj: MPR status during week j.
The weekly values can be accumulated to get a value of IP for the whole season which is strongly related to the downgrading rate of seed lots after laboratory testing. Moreover, the weekly cumulative IP can be plotted and compared in a graph with values for reference seasons (low-pressure and high-pressure seasons) so that the current IP can be compared with previous ones (Fig. 6.14) to provide advice to growers.

Relation between the cumulative index of PVY infection pressure (IP) (weeks 21–31) and the downgrading rate of Elite seed potato lots after laboratory testing (left panel). IP index during the growing season: comparison of 2014 with two reference seasons (2013, low pressure – 2011, high pressure) (Libramont, Belgium) (right panel)
4 Conclusion and Future Prospects
As for many other viruses transmitted in a non-persistent fashion, the interactions between PVY and their aphid vectors and environment are very complex. As we are just beginning to uncover the molecular basis of virus-vector relationships, the ongoing and future challenge faced are to integrate the wealth of data on the molecular diversity of PVY, their interaction with their hosts-vector-environment, and how these interactions will affect transmission and spread. Ultimately, gathering knowledge from a wide variety of ecological niche should help us understand what affects PVY epidemiology at the macroscopic level and, in practice, refine PVY risk modelling.
References
Al Mrabeh A, Anderson E, Torrance L, Evans A, Fenton B (2010) A literature review of insecticide and mineral oil use in preventing the spread of non-persistent viruses in potato crops. Agriculture and Horticulture Development Board (AHDB, UK). http://www.potato.org.uk/sites/default/files/publication_upload/Review%20of%20Spray%20Oils%20Insecticides%20and%20Potyvirus%20Transmission.pdf
Ashby JW (1976) An aphid trap used in virus-vector studies. N Z Entomol 6:187
Baldy C, Rabasse J-M. (1983) Caracteristiques spectrales de pieges jaunes utilises pour la capture des aphides. Agronomie 3(2):161–166
Bale JS, Ponder KL, Pritchard J (2007) Coping with stress. In: van Emden HF, Harrington R (eds) Aphids as crop pests. CABI, Wallingford, pp 287–310
Basky Z (2002) The relationship between aphid dynamics and two prominent potato viruses (PVY and PLRV) in seed potatoes in Hungary. Crop Prot 21:823–827
Basky Z, Almasi A (2005) Differences in aphid transmissibility and translocation between PVYN and PVYO isolates. J Pest Sci 78:67–75
Basky Z, Raccah B (1990) Aphids colonizing peppers in Hungary and their importance as virus vectors. Acta Phytopathologica Entomol Hung 25:383–391
Beemster ABR, de Bokx JA (1987) Survey of properties and symptoms. In: de Bokx JA, van der Want JPH (eds) Viruses of potatoes and seed-potato production. Pudoc, Wageningen, pp 84–113
Blanc S, Ammar ED, Garcia-Lampasona S, Dolja VV, Llave C, Baker J, Pirone TP (1998) Mutations in the potyvirus helper component protein: effects on interactions with virions and aphid stylets. J Gen Virol 79:3119–33122
Boisgontier D, Orensanz J, Glais L, Boulard F, Laty P, Deudon O, Gaucher D, Golaz F, Molot B, Giovinazzo R, Jacquot E, Le Hingrat Y (2013) Spatialisation de l’Information Météorologique Pour l’Amélioration des Techniques Culturales (projet SIMPATIC). Innov Agronomiques 30:329–347
Boiteau G, Singh RP, Parry RH, Pelletier Y (1988) The spread of PVY0 in New Brunswick potato fields: timing and vectors. Am Potato J 65:639–649
Boiteau G, Singh M, Singh RP, Tai GCC, Turner TR (1998) Rate of spread of PVYN by alate Myzus persicae (Sulzer) from infected to healthy plants under laboratory conditions. Potato Res 41:335–344
Boquel S, Giordanengo P, Ameline A (2011) Divergent effects of PVY-infected potato plant on aphids. Eur J Plant Pathol 129(4):507–510
Boquel S, Delayen C, Couty A, Giordanengo P, Ameline A (2012) Modulation of aphid vector activity by Potato virus Y on in vitro potato plants. Plant Dis 96:82–86
Casteel CL, Yang C, Nanduri AC, De Jong HN, Whitham SA, Jander G (2014) The NIa-Pro protein of Turnip mosaic virus improves growth and reproduction of the aphid vector, Myzus persicae (green peach aphid). Plant J 77:653–663
Chatzivassiliou EK, Moschos E, Gazi S, Koutretsis P, Tsoukaki M (2008) Infection of potato crops and seeds with PVY and PLRV in Greece. J Plant Pathol 90:253–261
Chickh-Ali M, Katayama K, Maoka T, Natsuaki KT (2008) Significance of weed hosts for Potato virus Y protection in Syria. EPPO Bull 38:226–232
Cronin S, Verchot J, Haldeman-Cahill R, Schaad MC, Carrington JC (1995) Long-distance movement factor: a transport function of the potyvirus helper component proteinase. Plant Cell 7(5):549–559
Davis JA, Radcliffe EB, Ragsdale DW (2005) Soybean aphid, Aphis glycines Matsumura, a new vector of Potato virus Y in potato. Am J Potato Res 82(3):197–201
de Bokx JA (1972) Graft and mechanical transmission. In: C.A.P.D (ed) Viruses of potatoes and seed-potato production, pp 26–35
de Bokx JA, Piron PGM (1990) Relative efficiency of a number of aphid species in the transmission of potato virus Y N in the Netherlands. Neth J Plant Pathol 96:237–246
DiFonzo CD, Ragsdale DW, Radcliffe EB, Gudmestad NC, Secor GA (1997) Seasonal abundance of aphid vectors of Potato virus Y in the Red River Valley of Minnesota and North-Dakota. J Econ Entomol 90:824–831
Doring TF, Chittka L (2007) Visual ecology of aphids—a critical review on the role of colours in host finding. Arthropod Plant Interact 1:3. doi:10.1007/s11829-006-9000-1
Dupuis B, Schwaerzel R (2011) Potato virus Y (PVY) strains spread and symptoms on plants and progeny tubers of potato cultivars. Proceedings of the The 18th Triennial Conference of the European Association for Potato Research. Oulu, Finland 115
Dupuis B, Balmelli C, Schumpp O, Steinger T (2014) Within-field spread of two strains of potato virus Y (PVY). In: Research EaFP (ed) Proceedings of the 19th Triennial Conference of the European Association for Potato Research. European Association for Potato Research, Brussels, p 65
Edwards AR (1963) A non-colonizing aphid vector of potato virus disease. Nature 200:1233–1234
Fageria MS, Nie X, Gallagher A, Singh M (2015) Mechanical transmission of Potato virus Y (PVY) through seed cutting and plant wounding. Am J Potato Res 92(1):143–147
Fenton B, Lacomme C, Collins L, Fox A, Pickup J, Evans A, Northing P, Anderson E (2012) Aphids and virus transmission in seed crops. Potato Council and Agriculture and Horticulture Development Board (AHDB, UK) project R428 final report. http://www.potato.org.uk/sites/default/files/publication_upload/R428_Final%20Report.pdf
Fereres A, Perez P, Gemeno C, Ponz F (1993) Transmission of spanish pepper- and potato-PVY isolates by aphid (Homoptera: Aphididae) vectors: epidemiological implications. Environ Entomol 22(6):1260–1265
Foottit RG, Maw HEL, Von Dohlen CD, Hebert PDN (2008) Species identification of aphids (Insecta: Hemiptera: Aphididae) through DNA barcodes. Mol Ecol Resour 8(6):1189–1201
Fox A, Collins L, Macarthur R, Blackburn LF, Northing P (2016) New aphid vectors and efficiency of transmission of Potato virus A and strains of Potato virus Y in the UK. Plant Pathol 66:325. doi:10.1111/ppa.12561
Giordanengo P, Brunissen L, Rusterucci C, Vincent C, van Bel A, Dinant S, Girousse C, Faucher M, Bonnemain JL (2010) Compatible plant-aphid interactions: how aphids manipulate plant responses. C R Biol 333(6–7):516–523
Guo B, Lin J, Ye K (2011) Structure of the autocatalytic cysteine protease domain of potyvirus helper-component proteinase. J Biol Chem 286(24):21937–21943
Halbert SE, Corsini DL, Wiebe MA (2003) Potato virus Y transmission efficiency for some common aphids in Idaho. Am J Potato Res 80(2):87–91
Harrington R, Gibson RW (1989) Transmission of Potato virus Y by aphids trapped in potato crops in southern England. Potato Res 32(2):167–174
Harrington R, Katis N, Gibson RW (1986) Field assessment of the relative importance of different aphid species in the transmission of Potato virus Y. Potato Res 29(1):67–76
He C, Molen TA, Xiong X, Boiteau G, Nie X (2006) Cytochrome c oxidase mRNA as an internal control for detection of Potato virus Y and Potato leafroll virus from single aphids by a co-amplification RT-PCR assay. J Virol Methods 138(1):152–159
Hogenhout SA, van der Hoorn RA, Terauchi R, Kamoun S (2009) Emerging concepts in effector biology of plant-associated organisms. Mol Plant-Microbe Interact 22:115–122
Holmes JC, Bethel WM (1972) Modification of intermediate host behaviour by parasites. In: Canning EU, Wright CA (eds) Behavioural aspects of parasite transmission. Academic Press, London, pp 123–149
Hull R (2002) Matthews’ plant virology, 4th edn. Acedemic Press, London
Ingwell LL, Eigenbrode SD, Bosque-Perez NA (2012) Plant viruses alter insect behavior to enhance their spread. Sci Rep 2:578
Johnson CG, Taylor LR (1955) The development of large suction traps for airborne insects. Ann Appl Biol 43:51–61
Jones DAC, Woodford JAT, Main SC, Pallett D, Barker H (1996) The role of volunteer potatoes in the spread of PVYN in ware crops of cv record. Ann Appl Biol 129:471–478
Kaliciak A, Syller J (2009) New hosts of Potato virus Y (PVY) among common wild plants in Europe. Eur J Plant Pathol 124:707–713
Kanavaki OM, Margaritopoulos JT, Katis NI, Skouras P, Tsitsipis JA (2006) Transmission of Potato virus Y in tobacco plants by Myzus persicae nicotianae and M-persicae s.str. Plant Dis 90(6):777–782
Kasschau KD, Carrington JC (2001) Long-distance movement and replication maintenance functions correlate with silencing suppression activity of potyviral HC-pro. Virology 285(1):71–81
Kasschau KD, Cronin S, Carrington JC (1997) Genome amplification and long-distance movement functions associated with the central domain of tobacco etch potyvirus helper component-proteinase. Virology 228(2):251–262
Katis N, Tsitsipis JA, Stevens M, Powell G (2007) Transmission of plant viruses. In: van Emden HF, Harrington R (eds) Aphids as crop pests. CABI, Wallingford, pp 353–390
Kennedy JS, Booth CO, Kershaw WJS (1959a) Host finding by aphids in the field. I. Gynoparae of Myzus persicae (Sulzer). Ann Appl Biol 47:410–423
Kennedy JS, Booth CO, Kershaw WJS (1959b) Host finding by aphids in the field. II. Aphis fabae Scop. (gynoparae) and Brevicoryne brassicae L.; with a re-appraisal of the role of host-finding behaviour in virus spread. Ann Appl Biol 47:424–444
Kerlan C (2006) Potato virus Y. Descriptions of plant viruses, no. 414. http://www.dpvweb.net/dpv/showdpv.php?dpvno=414
Kirchner SM, Döring TF, Hiltunen LH, Virtanen E, Valkonen JPT (2011) Information theory-based model selection for determining the main vector and period of transmission of Potato virus Y in potato crops. Ann Appl Biol 159:414–427
Kirchner SM, Hiltunen L, Döring TF, Virtanen E, Palohuhta JP, Valkonen JPT (2013) Seasonal phenology and species composition of the aphid fauna in a northern crop production area. PLoS One 8:e71030
Kirchner SM, Hiltunen LH, Santala J, Döring TF, Ketola J, Kankaala A, Virtanen E, Valkonen JPT (2014) Comparison of straw mulch, insecticides, mineral oil, and birch extract for control of transmission of Potato virus Y in seed potato crops. Potato Res 57:59–75
Kostiw M (1975) Investigation on the retention of potato viruses M and Y in two species of aphids (Myzus persicae Sulz. and Aphis nasturtii Kalt.) Potato Res 18:637–640
Kostiw M (1979) Transmission of Potato virus Y by Rhopalosiphum padi L. Potato Res 22:237–238
Kring JB (1972) Flight behaviour of aphids. Annu Rev Entomol 17:461–492
Labonne G, Fauvel G, Quiot JB (1983) Intérêt des pièges à fils dans l’étude des populations de pucerons ailés. Agronomie 3:315–326
Labonne G, Lauriaut F, Quiot J (1989) Comparison of 3 types of traps to sample alate aphid populations. Agronomie 9:547–557
Le Hingrat Y (2007) Identifier les pucerons pour adapter la lutte. La Pomme de Terre Française:36–37
Llave C, Kasschau KD, Carrington JC (2000) Virus-encoded suppressor of posttranscriptional gene silencing targets a maintenance step in the silencing pathway. Proc Natl Acad Sci 97:13401–13406
Mehle N, Ravnikar M (2012) Plant viruses in aqueous environment: survival, water mediated transmission and detection. Water Res 46:4902–4917
Mehle N, Gutierrez-Aguirre I, Prezelj N, Delić D, Vidic U, Ravnikar M (2014) Survival and transmission of Potato virus Y, Pepino mosaic virus, and Potato spindle tuber viroid in water. Appl Environ Microbiol 80(4):1455–1462
Modic S, Urek G (2008) Contribution to the knowledge of the aphid fauna (Sternorrhyncha: Aphidoidea) of Slovenia. Acta Entomol Slov 16(1):87–97
Moericke V (1951) Eine Farballe zur Kontrolle des Fluges von Blattlausen insbesondere der pfirschblattlaus, Myzodes persicae (Sulz). Nachrichtenbl Dtsch Pflanzenschutzdienst 3:23–24
Moericke V (1955) Über die Lebensgewohnheiten der geflügelten Blattläuse (Aphidina) unter besonderer Berücksichtigung des Verhaltens beim Landen. Z Angew Entomol 37:29–91
Moury B, Fabre F, Senoussi R (2007) Estimation of the number of virus particles transmitted by an insect vector. Proc Natl Acad Sci U S A 104(45):17891–17896
Nault LR (1997) Arthropod transmission of plant viruses: a new synthesis. Ann Entomol Soc Am 90:521–541
Nemecek T (1993) The role of aphid behaviour in the epidemiology of Potato virus Y: a simulation study. PhD thesis ETH Zürich
Nie X, Pelletier Y, Mason N, Dilworth A, Giguère MA (2011) Aphids preserved in propylene glycol can be used for reverse transcription-polymerase chain reaction detection of Potato virus Y. J Virol Methods 175(2):224–227
Nolte P, Whitworth JL, Thornton MK, Mcintosh CS (2004) Effect of seedborne potato virus Y on performance of russet Burbank, russet Norkotah, and Shepody potato. Plant Dis 88:248–252
Oh CS, Carrington JC (1989) Identification of essential residues in potyvirus proteinase HC-pro by site-directed mutagenesis. Virology 173(2):692–699
Palacios I, Drucker M, Blanc S, Leite S, Moreno A (2002) Cauliflower mosaic virus is preferentially acquired from the phloem by its aphid vectors. J Gen Virol 83:3163–3171
Pelletier Y, Nie X, Giguère MA, Nanayakkara U, Maw E, Foottit R (2012) A new approach for the identification of aphid vectors (Hemiptera: Aphididae) of Potato virus Y. J Econ Entomol 105(6):1909–1914
Perez P, Collar JL, Avilla C, Duque M, Fereres A (1995) Estimation of vector propensity of Potato virus Y in open field pepper crops of Central Spain. J Econ Entomol 88(4):986–991
Pettersson J (1970) Studies on Rhopalosiphum padi (L.). I. Laboratory studies on olfactometric responses to the winter host Prunus padus L. Lantbrukshögskolans Ann 36:381–389
Pettersson J, Tjallingii WF, Hardie J (2007) Host-plant selection and feeding. In: van Emden HF, Harrington R (eds) Aphids as crop pests. CABI, Wallingford, pp 87–114
Pickup J, Davie K, Fox A, Highet F, Holmes R (2009) Epidemiology of viruses in Scottish seed potatoes. Asp Appl Biol Potatoes 94:5–10
Pirone TP, Blanc S (1996) Helper-dependent vector transmission of plant viruses. Annu Rev Phytopathol 34:227–247
Pirone TP, Perry KL (2002) Aphids: non-persistent transmission. Adv Bot Res 36:1–20
Pirone TP, Thornbury DW (1988) Quantity of virus required for aphid transmission of a Potyvirus. Phytopathology 78(1):104–107
Powell G, Harrington R, Spiller NJ (1992) Stylet activities and Potato virus Y vector efficiencies by the aphids Brachycaudus helichrysi and Drepanosiphum platanoidis. Entomol Exp Appl 62(3):293–300
Pruss G, Ge X, Shi XM, Carrington JC, Bowman Vance V (1997) Plant viral synergism: the potyviral genome encodes a broad-range pathogenicity enhancer that transactivates replication of heterologous viruses. Plant Cell 9(6):859–868
Raccah B, Galon A, Eastop VF (1985) The role of flying aphid vectors in the transmission of Cucumber mosaic virus and Potato virus Y to peppers in Israel. Ann Appl Biol 106:451–460
Radcliffe EB, Ragsdale DW (2002) Aphid-transmitted potato viruses: the importance of understanding vector biology. Am J Potato Res 79:353–386
Ragsdale DW, Radcliffe EB, DiFonzo CD (2001) Epidemiology and field control of PVY and PLRV. In: Loebenstein G, Berger PH, Brunt AA, Lawson RH (eds) Virus and virus-like diseases of potatoes and production of seed-potatoes. Klumer Academic Publisher, Dordrecht, pp 237–270
Robert Y, Turpeau E, Tanguy S (1987) Complementation of suction and yellow traps. In: Cavalloro R (ed) Aphid migration and forecasting ‘Euraphid’ systems in European Community countries. Published by the Commission of the European communities, Luxembourg, pp 103–111
Robert Y, Woodford JAT, Ducray-Bourdin DG (2000) Some epidemiological approaches to the control of aphid-borne virus diseases in seed potato crops in northern Europe. Virus Res 71:33–47
Rolot JL (2005) Analyse des facteurs régulant la dissémination du virus Y de la pomme de terre (PVY) en vue de stratégies de lutte raisonnées. Dissertation. Faculté Universitaire des Sciences Agronomiques, Gembloux, Belgium
Sáenz P, Salvador B, Simón-Mateo C, Kasschau KD, Carrington JC, García JA (2002) Host-specific involvement of the HC protein in the long-distance movement of potyviruses. J Virol 76(4):1922–1931
Salazar LF (1996) Aphid transmission of potato viruses. In: C.I.P (ed) Potato viruses and their control. International Potato Center, Lima. 214 p
Seco V, Hulle M, Nafria JMN (1990) Numbers, richness and diversity of aphids trapped in Moericke and suction traps in Leon, Spain. Acta Phytopathol Entomol Hung 25:159–165
Sigvald R (1984) The relative efficiency of some aphid species as vectors of potato virus YO (PVYO). Potato Res 27(3):285–290
Sigvald R (1986) Forecasting the incidence of potato virus YO. In: McLean GD, Garrett RG, Ruesink WG (eds) Plant virus epidemics, monitoring, modelling and predicting outbreaks. Academic Press, Sydney, pp 419–441
Sigvald R (1987) Forecasting Potato virus Y. Waxtskyddsnotiser 51:131–137
Sigvald R (1989) Relationship between aphid occurrence and spread of potato virus YO (PVYO) in field in southern Sweden. J Appl Ent 108:35–43
Sigvald R (1992) Progress in aphid forecasting systems. Neth J Plant Pathol 98(suppl. 2):55–62
Srinivasan R, Alvarez JM, Bosque-Pérez NA, Eigenbrode SD, Novy RG (2008) Effect of an alternate weed host, hairy nightshade, Solanum sarrachoides, on the biology of the two most important potato leafroll virus (Luteoviridae: Polerovirus) vectors, Myzus persicae and Macrosiphum euphorbiae (Aphididae: Homoptera). Environ Entomol 37(2):592–600
Steinger T, Gilliand H, Hebeisen T (2014) Epidemiological analysis of risk factors for the spread of potato viruses in Switzerland. Ann Appl Biol 164:200–207
Steinger T, Goy G, Gilliand H, Hebeisen T, Derron J (2015) Forecasting virus disease in seed potatoes using flight activity data of aphid vectors. Ann Appl Biol 166:410–419
Thornbury DW, Hellmann GM, Rhoads RE, Pirone TP (1985) Purification and characterization of potyvirus helper component. Virology 144(1):260–267
Urcuqui-Inchima S, Maia IG, Arruda P, Haenni AL, Bernardi F (2000) Deletion mapping of the potyviral helper component-proteinase reveals two regions involved in RNA binding. Virology 268(1):104–111
Uzest M, Gargani D, Drucker M, Hebrard E, Garzo E, Candresse T, Fereres A, Blanc S (2007) A protein key to plant virus transmission at the tip of the insect vector stylet. Proc Natl Acad Sci U S A 104(46):17959–17964
Vance VB, Berger PH, Carrington JC, Hunt AG, Shi XM (1995) 5’ proximal potyviral sequences mediate potato virus X/potyviral synergistic disease in transgenic tobacco. Virology 10:583–590
van der Waals JE, Krüger K, Franke AC, Haverkort AJ, Steyn JM (2013) Climate change and potato production in contrasting south African agro-ecosystems 3. Effects on relative development rates of selected pathogens and pests. Potato Res 56:67–84
van Harten A (1983) The relation between aphid flights and the spread of potato virus YN (PVYN) in the Netherlands. Potato Res 26(1):1–15
van Hoof HA (1980) Aphid vectors of potato virus YN. Neth J Plant Pathol 86:159–162
Verbeek M, Piron PGM, Dullemans AM, Cuperus C, van der Vlugt RAA (2010) Determination of aphid transmission efficiencies for N, NTN and Wilga strains of Potato virus Y. Ann Appl Biol 156(1):39–49
Zhang J, Nie X, Nanayakkara U, Boquel S, Giguère MA, Pelletier Y (2013) Detection of Potato virus Y from the stylets of a single aphid by one-step reverse transcription polymerase chain reaction. Entomol Exp Appl 147(1):93–97
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Lacomme, C. et al. (2017). Transmission and Epidemiology of Potato virus Y . In: Lacomme, C., Glais, L., Bellstedt, D., Dupuis, B., Karasev, A., Jacquot, E. (eds) Potato virus Y: biodiversity, pathogenicity, epidemiology and management. Springer, Cham. https://doi.org/10.1007/978-3-319-58860-5_6
Download citation
DOI: https://doi.org/10.1007/978-3-319-58860-5_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-58858-2
Online ISBN: 978-3-319-58860-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)