
Abstract
Forests are the dominant terrestrial ecosystem occupying approximately 30% of the Earth total land area. They play an important role in the global carbon (C) cycle, and the mitigation of CO2 emission due to its large storage of soil organic C (SOC), a large part of which is stored in soils. Due to their dominance, forests management has gained much interest in science and policy discussions as one of the important options to mitigate climate change. Global forests are increasingly affected by land use change , fragmentation, changing management objectives and degradation. The area under global forests has declined by 3% from 1990 to 2015, but the area of planted forest has increased in all regions of the world and now accounts for nearly 7% of global forest land estimated at 3999 million hectares (Mha). The area of primary forests which is typically defined as lacking direct human influence, is about 34% of the total forest land, based on country reports, but this area is declining, especially in South America and Africa because of human-caused fragmentation and degradation. About 5% of global forests are plantations generally used for commercial purposes. Globally, timber production has remained stable since 1990, but forest used for non-wood forest products indicates that harvesting is taking place on a smaller proportion of total forest area. Based on trends in the area of managed forest and regional studies, historical and current forest management has been a significant determinant of current carbon stocks in forest. The established forest currently offset 30% of global emissions of CO2 from fossil fuel combustion, and there are mitigation opportunities involving forests that could increase the gross terrestrial C uptake from about 4.0 to 6.2 Pg C annually. Diversifying use of forest land may have significant consequences for maintaining or increasing the current rate of terrestrial C sequestration. Indirect human influences such as increasing atmospheric CO2 and climate change, along with the direct effects of land management and projected increasing demand for wood biofuel, are likely to become increasingly important elements that influence land management strategies and the role of forests in the global C cycle and future climate mitigation.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others
Keywords
- Forest resources
- Deforestation
- Afforestation
- Land use change
- Sustainable forest management
- Primary forest
- Gross primary productivity
- Net primary productivity
11.1 Introduction
Forests, the dominant terrestrial ecosystem on Earth, are distributed across boreal, temperate, and tropical zones and account for 80% of Earth total biomass (Kindermann et al. 2008). They also account for 75% of the terrestrial gross primary production (GPP) (Beer et al. 2010). They are locally and globally important ecosystems that provide habitat, timber resources, carbon (C) storage, recreational opportunities, as well as multiple ecosystem services and cultural and spiritual values (Hicke et al. 2007; McKinley et al. 2011; Miura et al. 2015). Forests harbor the majority of species on Earth (Jackson et al. 2005) and contain more C in the biomass and soils than that resides in the atmosphere (Pan et al. 2011; Bellassen and Luyssaert 2014). In addition to providing valuable services to society, forests also play an important role in the global C cycle. Compared to other terrestrial ecosystems, forests store the largest quantities of C per unit land area (Poeplau et al. 2011). As a result, forests represent one of the largest, most cost-effective mitigation for climate change solutions available currently. The C storage capacity in forests could be improved through afforestation or decreased by deforestation (Wei et al. 2014). For example, afforestation of croplands results in soil C accumulation at a rate of 0.38 ± 0.04 Mg ha−1 yr−1, lasting for more than 100 years (Poeplau et al. 2011). Global forests are also increasingly affected by deforestation and land use change , fragmentation, changing management objectives, degradation, as well as responding to changes in atmospheric composition especially increasing CO2 concentration, N deposition tropospheric O3, and climate change. The response to these concurrent factors will determine the sustainability of many traditional services provided by forests such as timber production, watershed protection, as well as future C sequestration.
Forests contribute to soil formation and water regulation, and are estimated to provide direct employment to as many as 10 million people and source of livelihood to millions more (FAO 2010). It is estimated that more that 200 million people from the world’s poor communities rely directly on forests for energy, shelter, and their livelihood (Hirsch 2010). Clean water is becoming more recognized as one of the most important environmental services provided by forests. At least one third of the world’s largest cities draw a significant proportion of their drinking water from forested areas (FAO 2013). In 2015, 1002 million hectare (Mha) or 25.1% of world’s forests were primarily designated for protection of soil and water, also an additional 25.4% is managed for ecosystem services (Miura et al. 2015; FAO 2015). In the context of resilience to climate change, forests are also recognized to play an important role in minimizing the risks of natural hazards such as landslides and local floods, creating resilience as well as adaptation to climate change.
Net primary productivity (NPP), defined as the difference between accumulative photosynthesis and accumulative autotrophic respiration by green plants per unit of time and space, is the important parameter for quantifying the exchanges of energy and mass by vegetation (Running and Coughlan 1988). In general, forest ecosystem NPP accounts for 35% of global and 65% of terrestrial ecosystem NPP, respectively (Waring and Schlesinger 1985; Gower et al. 1996). Forest ecosystems contains up to 80% of all aboveground terrestrial biomass C and approximately 40% of all belowground terrestrial C (Dixon et al. 1994). Compared to other terrestrial ecosystems, forests store the largest quantities of C per unit surface area of land (Poeplau et al. 2011), and therefore, they are the major component of Earth C cycling. Thus, afforestation increases C storage capacity, while deforestation decreases terrestrial C storage capacity (Poeplau et al. 2011; Wei et al. 2014). A slight change in NPP of forests will significantly influence atmospheric CO2 concentration and, consequently, climate change. The annual CO2 exchange between forests and atmosphere through photosynthesis and respiration is estimated to be ≈50 Pg C yr−1 (Beer et al. 2010), and an increase in soil respiration would increase the CO2 emission from forest ecosystems.
The net C accumulation in forest ecosystem over decadal time frame is more influenced by disturbances than climate and atmospheric CO2 concentration (Pregitzer and Euskirchen 2004). Therefore, understanding how disturbances and stand age interact could provide fundamental knowledge of the terrestrial C cycle. The C stored in forest ecosystems over the long term will be released rapidly and in considerable amounts into the atmosphere following disturbances (Page et al. 2002). In forests, time since disturbances (i.e., forest age) and forest structure are critical factors determining forest ecosystem C storage and fluxes (Song and Woodcock 2003; Litvak et al. 2003; Kashian et al. 2006; Goulden et al. 2011). Generally, forest C cycling is influenced by forest age—including biomass of coarse woody debris (Bond-Lamberty et al. 2002; Li et al. 2012), forest stand water use (Delzon and Loustau 2005), SOC (Peltoniemi et al. 2004), live biomass increment and litter decomposition (Bradford et al. 2008), size structure, NPP, and net ecosystem productivity (NEP)/net biome productivity (Hoshino et al. 2001; Litvak et al. 2003; Song and Woodcock 2003), and other biophysical properties (McMillan and Goulden 2008). Younger forests are inherently more productive than older forests (Ryan et al. 1997) and model predictions also show that successional changes influence rates of NEP through time (Thornton et al. 2002). As a result of the high variation in stand age in large forest areas, efforts to estimate ecosystem C fluxes must take into account forest age structure (Song and Woodcock 2003), especially the quantitative analysis of forest NPP changes with stand age among forest types (Chen et al. 2003) to improve the reliability. In general, quantitative research on the temporal trends of C balance related to stand age at regional and/or global scales is rare due to lack of long-term records of disturbances, and most process-based models ignore the effects of disturbances on NPP and terrestrial C sequestration (e.g., Dean et al. 2004; Thornley and Cannell 2004), mainly because of the lack of spatial data on NPP and age distributions. The C cycling and stand age relationships developed on single species have indicated that biomass accumulation peaked within 40–60 years, while a meta-analysis of published chronosequence data to explore the dependence of forest NPP on age by age classes for tropical, temperate, and boreal forest biomes suggested that peak NPP in boreal forest occurred in the 71–120 year age class while older forests >120 years were generally less productive (Pregitzer and Euskirchen 2004).
Over the past 25 years, an average of 4.6 Mha yr-1 of forests are cleared and converted for production of commodities including soy, palm oil, beef, and paper, falling from 7.3 Mha yr−1 in the 1990s (Keenan et al. 2015). In addition, infrastructure, urban expansion, mining, and fuel wood also contribute to annual forest loss. Globally, 129 Mha of natural and planted forest were deforested between 1990 and 2015, representing an annual rate of forest loss of 0.13% (FAO 2015). The net annual rate of forest loss has slowed from 0.18% in 1990s to a net annual loss of 0.08% over the last 5 years (FAO 2015). Between 2010 and 2015, there was an annual loss of 7.6 Mha and net gain of 4.3 Mha, resulting in net annual decrease of 3.3 Mha yr−1 occurring in every climatic domain (FAO 2015). However, the largest area of forest converted to other land uses between 1990 and 2015 was the tropical forests, and the continents with the greatest forest area loss are Africa and South America, where forest land is converted to agriculture and other land uses in response to population increase (FAO 2015).
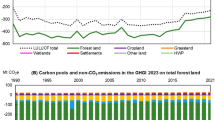
Forestry plays central role in sustainable development, global climate change, and mitigation. Decadal land use change emission of CO2, mainly from deforestation and forest degradation in tropical regions from 1990 to 2014 are estimated at 1.2 ± 0.5 Pg C yr−1, equivalent to 16% of C emissions from fossil fuels from 1990 to 2015 (Pan et al. 2011; Le Quéré et al. 2015, 2016), which is offset by tropical regrowth, boreal, temperate, and intact tropical forests C sink. Decadal residual terrestrial C sink estimated as anthropogenic emissions from fossil fuel and land use change minus ocean uptake and atmospheric growth rate for 1990–2015 is estimated at 2.8 Pg C yr−1 (Pan et al. 2011; Le Quéré et al. 2015, 2016). Decadal land use change emissions declined from 1.6 ± 0.5 Pg C yr−1 in 1990s to 1.0 ± 0.5 Pg C yr−1 in 2000s, while residual terrestrial C sinks increased from 2.6 ± 0.8 Pg C yr−1 in 1990s to 3.1 ± 0.8 Pg C yr−1 in 2000s (Le Quéré et al. 2015, 2016). Concerns about the role of tropical forests in global climate change have led to the United Nations (UN) to propose a mechanism to facilitate tropical countries participation in climate change mitigation through reducing emission from deforestation and degradation and the role of conservation (REDD+), sustainable management of forests and enhancement of forest C stocks (UNFCCC 2007). Tropical forest growth amounts for a sink of 1.6 ± 0.8 Pg C yr−1, which offsets significant portion of emissions (Nabuurs et al. 2007; Pan et al. 2011). Therefore, by participation in REDD+, tropical countries could turn tropical forests to a net C sink instead of source. Forests are also affected by climate change, and their contribution to mitigation strategies may be influenced by stresses resulting from climate change. Global forests also play significant roles socioeconomically by providing important goods, services, and financial values to the societies. Other co-benefits of forests include biodiversity and watershed conservation, provision of timber and fiber, as well as recreational services. Forest mitigation options include reducing emissions from deforestation and forest land degradation, enhancing sequestration rates in existing forests, providing fuel substitute for fossil fuels, and also providing wood products for more energy-intensive materials.
Plants take up CO2 from the atmosphere and N from soil. When they grow and redistribute it among different pools including above and below ground biomass, dead tissues, and soil organic matter (SOM). CO2, CH4 and N2O are in turn released to the atmosphere through respiration and decomposition of dead plant biomass and SOM or through combustion. Anthropogenic land use activities such as forest logging, conversion of forest lands and deforestation and afforestation cause changes superimposed on natural gaseous exchange fluxes. These activities can both lead to source and sinks of CO2 in forest. Forest management activities play key role through mitigation of climate change. However, forests are also affected by climate change, and their contribution to mitigation strategies may be influenced by stresses resulting from climate change. Forest mitigation options include (i) reducing emissions from deforestation and forest degradation, enhancing sequestration rate in existing and new forests, provisioning of wood fuel as a substitute for fossil fuel, and providing wood products for more energy intensive materials. There is an increased attention to reducing emissions from deforestation as low cost mitigation option and with significant positive side effects.
Several analyses has estimated the terrestrial C sink in the range of 1.1–3.6 Pg C yr−1 for the 1990s and 2000s on the basis of atmospheric CO2 observations and inverse modeling as well as land observation (Canadell et al. 2007b; Khatiwala et al. 2009; Le Quéré et al. 2009, 2015), while the budget residual, after accounting for atmospheric growth and ocean uptake suggest the land sink ranging from 2.6 to 4.1 (Le Quéré et al. 2015). Most forests of the world are recovering from a past disturbance, and there is a considerable uncertainty in the future trajectory and magnitude of terrestrial C sink, as many aggrading forests approach maturity following clear-cut harvesting a century or more ago (Birdsey et al. 2006). With these re-growing forests advancing beyond early aggrading phase of succession, an ecologically important transition will occur in which structurally and biologically simple forests dominated by short-lived early successional trees senesce and allow more complex stands comprising of longer lived, later successional species (Birdsey et al. 2006). Naturally regenerated mixed-deciduous forests in many regions especially in North America and Eurasia are leading to reemergence of later successional forests (Luyssaert et al. 2010; Wang et al. 2011). The objectives of this chapter is to critically assess the roles of global forest management in the global C cycle and mitigating climate change.
11.2 The Role of Forests in the Global Carbon Cycle
Global C cycling is presented in Fig. 6.1, and an excerpt representing conceptual global forest C cycling is presented in Fig. 11.1. Forests hold C stocks in living and dead biomass, where living trees capture and release C through photosynthesis and respiration, dead biomass decay and release C, and a fraction of this C eventually ends up in soils. Using energy from the sun and water from the soil, trees and the understory continuously cycle C through photosynthesis, growth, respiration, death, and decay. Photosynthesis convert CO2 from the atmosphere into photosynthetic products [i.e., sugars, water, and oxygen (O2)], which are used directly for cellular respiration and root exudation, or stored as more complex molecules as an energy store or for growth of leaves, stem, and roots. Overall, photosynthesis generally exceeds respiration, making most terrestrial ecosystems net sinks of C in natural state. When the tree growth is directed towards woody tissues—stems, branches, and large roots, it is referred to as biomass accumulation. Contrary to old accepted theory that old unharvested forests are in equilibrium in terms of C sink, new observations have shown that unharvested old forests are absorbing more C than they release (Luyssaert et al. 2008), accounting for the half of the terrestrial C sinks estimated at 3.0 ± 0.8 Pg C yr−1 for 2005–2014 period (Ciais et al. 2013). The C sink of the mature forests is attributed to large-scale environmental changes such as higher atmospheric CO2 concentration and N emitted from agriculture and fossil fuel burning and deposited hundreds of kilometers away which is increasingly fertilizing forests in Europe, China and eastern USA (Solberg et al. 2009), causing violation of the steady-state conditions.

Conceptual diagram of forest carbon cycle showing forest ecosystem and forest product use
The mass of tree is composed nearly 50% C on dry mass, and natural systems dominate global C cycle. Terrestrial vegetation alone takes up an estimated 123 Pg C yr−1 and respiring 119 Pg C yr−1 (Ciais et al. 2013), which is about 13 times more than anthropogenic emissions from fossil fuels and cement production estimated at 9.3 ± 0.5 Pg C yr−1 for 2006–2015 decade (Le Quéré et al. 2015, 2016). These fluxes are dominated by forests, and the significance of forests to global C cycle is demonstrated by intra-annual variation of atmospheric CO2 concentration which is expressed in sawtooth effect showing atmospheric CO2 concentration that corresponds to the growing season in the northern hemisphere where most of land and forests exist (Fig. 6.2; Keeling 1960). For example, during northern hemisphere summers, photosynthesis exceeds respiration globally, and atmospheric CO2 declines, whereas during northern hemisphere winters, respiration exceeds photosynthesis and atmospheric CO2 concentration increases (Fig. 6.2). Forests contain 77% of all terrestrial aboveground C (Houghton 2007).
Because of forests, atmospheric CO2 concentrations are not rising as rapidly as they would be predicted by adding anthropogenic CO2 emissions to current levels in the atmosphere. Oceans are also absorbing additional 2.6 ± 0.5 Pg C yr−1 during 2006–2015 decade or 2.4 ± 0.5 Pg C since 1990 (Le Quéré et al. 2015, 2016). This phenomenon has been called a “loan from nature” and also a “buffer to climate change” (Philips and Lewis 2014). Tropical forests are the largest, most dense, and most diverse on Earth, but the high levels of deforestation within the tropics account for nearly all net forest loss and GHG emissions from forest land use across the planet. It is estimated that every year 72 Pg C cycle through tropical forests and savannahs, representing 59% of terrestrial GPP, the total influx of C from the atmosphere to plants per unit time (Beer et al. 2010). In addition to capturing C, forests also have vital interactions with climate, hydrological cycle, and nutrient cycle.
11.3 Estimation of Global Forest Area Trends and Its Carbon Pools
Climate change is predicted to create substantial shift in tree species distribution and forest structure (Gustafson et al. 2010) and these shifts require monitoring to understand how global forest resources are changing is important since global forests drive policy and resource flows at global, regional, and national levels. Assessing global forest resources and patterns of forest productivity is important for both scientific and practical purposes. For example, forest losses due to long-term land use change , especially land clearing in the tropics are important part of GHG emissions (Settele et al. 2014). Forest resources data are generally used for various purposes including estimation of global emission from land use and land use change, as well as climate change modeling (Petrescu et al. 2012; Smith et al. 2014). Such an assessment is necessary for the global C cycle studies as well as producing useful information required for planning and sustainable management of forest resources both on local and on regional scale (Corona and Marchetti 2007). The necessity of monitoring and quantifying the amount of C accumulated within forests has increased in view of mitigation of climate change efforts such as REDD+ mechanisms. However, Estimates of GHG emissions from deforestation require information on both the area of forest loss and the corresponding C stock of the land that is cleared (Houghton 2005; Gibbs et al. 2007), which has remained a challenge to accurately quantify, especially tropical deforestation and associated C losses (Ramankutty et al. 2007). Knowledge of the structure, distribution and biomass of the global forests is advancing rapidly due to improved global observation systems and analysis techniques (Saatchi et al. 2011; Asner et al. 2012). Satellite systems, airborne observation systems, improved land-based inventory systems and ecosystem models are providing high resolution maps (Running et al. 2009; Shugart et al. 2010; Masek et al. 2015). Estimates of global area under forest and global forest C pools are generally obtained from different sources of information including: (i) field inventories, (ii) country reports, (iii) book keeping models, (iv) remote sensing, and (v) modeling.
11.3.1 Field Inventories
Field inventories are based on field sampling, and this approach has been used for more than a century, mainly for assessing timber supply and monitoring forest changes. It can be a foundation of forest C monitoring as an initial assessment of C stocks under forest from which changes can be estimated either using remote sensing or modeling and also as a direct estimate of stock change from repeated inventories. One of the main aspects of forest C pool—forest biomass has traditionally been measured and monitored with field inventory method which was developed for timber supply assessment (Birdsey and Heath 2001). Inventories are particularly suitable for monitoring vital elements of forest dynamics such as growth, harvest, and mortality, and also for estimating biomass, and when combined with remote sensing it can provide a quantifiable error estimate about forest inventory.
Forest inventory involves systematic or random selection of sampling locations in large areas—as large as countries, field measurements of tree parameters such as species, diameter, and height and the development of allometric equations estimating forest variables of interest that is difficult to directly measure—such as timber volume or biomass (Pearson et al. 2007; Birdsey et al. 2013). The inventory sampling approach provides unbiased estimates with known sampling uncertainty, even though uncertainty attributed to use of allometric equations or models is infrequently estimated (Phillips et al. 2000). Monte Carlo estimation methods can be used for estimating overall national uncertainty (Heath et al. 2011). Most of global forest statistics on forest biomass and other forest attributes reported by FAO are based on forest inventories obtained at national level (FAO 2010, 2015). Global estimate of forest area and biomass for the last two decades based primarily on ground data are based on comprehensive statistics obtained from the inventory approach (Pan et al. 2011). National inventories are often targeted to assess the population of live and dead trees in forest, while other ecosystem C pools can be estimated directly using supplemental measurements added to the inventory or modeled using exogenous data (Woodall et al. 2012).
Typically, biomass equations are developed for population of trees by harvesting and weighing a small sample of trees across a range of diameter and height classes and then estimating parameters of equation relating biomass to measured variables using regression techniques (Birdsey et al. 2013). Individual tree estimates can be expanded to tree population if the probability of sampling each tree and area to which sample apply is known. Also a variation of this approach has been used (FAO 2014).
Limitation of this approach include: (1) Scarcity of representative biomass or volume equations, since only few equations are available that represent the population of trees that may be different from the population of interest (Zianis et al. 2005). This is particularly true for tropical regions where such work is generally lacking (Brown 1997). Regression equations using three independent variables—tree diameter, tree height, and wood specific gravity can be used to reduce biases in such cases where equations are lacking (Chave et al. 2005). (2) Estimating the change in biomass of live trees requires successive measurements of sample plots at an interval of several years to estimate average rate of change in tree diameter and height. Inventory estimates from USA suggests that the largest C pools is in the live biomass and soils and contributes the most to overall estimates of forest C stocks and stock changes (Heath and Smith 2000), and overall uncertainty of estimated changes in forest C stocks for USA is ~21% (EPA 2015).
11.3.2 Country Reports
Country reports, often based on national forest inventories have generally been used for compilation of regional and global forest statistics. Food and Agriculture Organization of the United Nations (FAO) in their forest resources assessment reports and also Intergovernmental Panel on Climate Change (IPCC) have relied on country reports (FAO 2010, 2015; Ciais et al. 2013). Centralizing forest statistical information through country participation makes available the national statistics which have been used in number of scientific applications (Houghton 2005; Kindermann et al. 2008). Limitation of FAO forest resource assessment include lack of internal consistence due to different definition of forest among countries and also time intervals (Grainger 2008). Country reports sometimes provide incomplete data, especially from developing countries where resources are limited. FAO compiles two types of datasets—Forest Resources Assessment at 5-year interval which provides details about forest alone, while FAOSTAT datasets are more robust, and including estimates of emissions from different land uses as well as areas under croplands, pastures forests and other lands which is also useful for estimating land use and land use changes (FAOSTAT 2016).
11.3.3 Bookkeeping Model
The book keeping model uses data on rate of land use change and per hectare change in C storage that result from changes in land use and land management (Houghton 2003, 2005). Land use change includes clearing of land for cultivation and pasture, the abandonment of agriculture land, the harvest of wood, reforestation, afforestation , shifting cultivation, and wild fires. The book keeping model tracks C in living vegetation, dead plant material, wood products removal, and soils for each unit land area harvested or reforested. Rates of land use change are generally obtained from agriculture and forest statistics, historical accounts and country reports (Houghton 2003). This approach comprises of compiling information about changes in forest area from nationally aggregated land use statistics, satellite data on land cover and satellite data on wild fires and vegetation response curves to define per hectare changes in C density as a result of land management (Houghton et al. 2012). The bookkeeping model sums the observed changes in management. The global bookkeeping estimates are good approximation of area and C changes resulting directly from human activities.
11.3.4 Remote Sensing
Aerial photographs have been used for nearly a century in forest inventories to estimate the proportion of land under forest in a given sampling area and as a first phase sample in a double sampling strategy. However, in recent decades, the Landsat satellites have provided a time series of remotely sensed digital images that are widely used for establishing historical baselines and also for monitoring current deforestation , forest degradation and natural disturbances (Tran et al. 2016; Cohen and Goward 2004; Chen et al. 2016). Landsat imagery does not directly estimate biomass, however, but spectral attributes are related to biomass and can be used together with field data and models to provide spatially explicit biomass and other vegetation attributes over large areas (Cohen et al. 1996; Masek et al. 2015). When associated with field observations or models, Landsat satellites can also be used for estimating changes in biomass and C stocks. The series of remote sensed digital images span the last 30 year period and currently widely used for monitoring biomass and C stocks. Landsat data are suitable for classifying vegetation and assessing other attributes of forest such as percent forest cover, leaf area index, and disturbances (Cohen and Goward 2004; Tran et al. 2016; Chen et al. 2016). The Moderate Resolution Imaging Spectroradiometer (MODIS) satellite also provides useful information about forest biomass, productivity and disturbances over large regions at a course spatial resolution (Running et al. 2004; Wang and D’Sa 2010; Rossi et al. 2013).
Three-dimensional remote sensing allows researchers to estimate forest canopy height, map the global forests, as well as estimating forest biomass and C stocks (Lefsky 2010; Ni et al. 2015). Many other ecological parameters such as forest succession and vegetation interactions are also assessed by three dimensional remote sensing. New tools are also providing decision support for forest management and ecological restoration. Remote sensing approaches are typically used to assess land cover and land cover change. They are not suitable for detecting land use or land use change , since it is usually difficult to separate human and natural causes of observed changes using remote sensing images without combining images with other information such as field inventory. It is also difficult to determine whether observed changes in land cover are permanent or temporary without an associated change in land use such as forest clear-cutting harvest. In general, remote sensing is combined with field sampling in national forest inventories to be more effective in identifying causes of change and improve overall monitoring efficiency.
11.3.5 Models
Varieties of ecosystem models have been used to quantify biomass and forest C dynamics through integration as well as synthesis of data covering different spatial and temporal scales ranging from detailed plot level measurement to national remote sensing products (Kurz et al. 2009; Wulder et al. 2008). Models also enable understanding of mechanisms controlling C exchanges between land and atmosphere, identify gaps in information, and guide future research to fill these gaps (Huntzinger et al. 2012). Models are also effective tools for predicting the future and comparing different scenarios to examine effects of different activities such as management or disturbance that have not been observed (Kurz et al. 2009). Available models can be grouped into process-based and empirical models. Process-based models uses information collected from intensive monitoring site such as leaf area index, soil conditions and climate variability to simulate C dynamics driven by photosynthesis processes (Running and Coughlan 1988). Bottom-up studies using dynamic global vegetation models (DGVMs) can mechanistically represent many of the key land processes and investigate how changes in the structure and function of the land ecosystems affect biogeochemical cycles and provide comprehensive analysis of surface C and mechanisms behind regional trends in C cycles (Sitch et al. 2015). Empirical models uses information derived from inventories and management plans such as wood volume yield data (Kurz et al. 2009; Masera et al. 2003).
11.4 Forest Area, Status and Trends
Global forests are increasingly affected by land use change , land fragmentation, changing forest management objectives, and degradation. Despite these factors, since 1990 the biomass in forest has consistently increased in global forests, suggesting that global drivers of elevated CO2 may be enhancing biomass gains (Pan et al. 2013). In addition, all forests are responding to changes in atmospheric composition, especially increased NOx and N deposition, increasing CO2 concentration, and climate change. Sustainability of the future forests and the associated services provided by forests such as timber production, watershed protection, biodiversity, and the role of forests in removing anthropogenic CO2 emissions depend on response of the global forests to these concurrent factors. Currently, the global C sink in established forest is estimated at 28% of anthropogenic CO2 emissions (Pan et al. 2011; Le Quéré et al. 2015, 2016), and if this sink is reduced, the global efforts to mitigate climate change will require even further emissions reductions than currently projected (IPCC 2014). According to FAO, forest is defined as land with tree crown cover or stocking level of more than 10% and an area of more than 0.5 hectare (ha), with trees able to reach a minimum of 5 m at maturity in situ (FAO 2015). It may consist of closed forest formations with various open continuous forest vegetation cover in which tree crown exceeds 10% or various storeys and undergrowth cover with high proportion of ground. Young natural stands and plantations established for forest purposes which have not yet reached a crown density of 10% or tree height of 5 m as a result of human intervention causes such as clear-cutting are included under forest because these areas are expected to revert to forest. However, land that is predominantly agricultural or urban is not included, even if such land has some tree cover which meets the forest definition. The global land under tree cover can be classified into primary forest , protected forest, unmanaged natural forest, managed forest, plantation forest, and other wooded land (Table 11.1; FAO 2010, 2015). Although not technically considered as forest based on standard forest definition, trees outside forests are valuable source of many products and services found in forest.
Based on FAO estimate, the global forests currently covers about 3999 Mha of Earth’s land surface equivalent to 31% of global land area in year 2015, distributed among all six continents (Table 11.2; FAO 2015). Tropical, sub-tropical, temperate, and boreal forests account for 44.3, 8.0, 17.1, and 30.6% of the area, respectively, (Table 11.3; Keenan et al. 2015). The bulk of global forest is natural (comprising the sum of primary forest and other naturally regenerated forest), amounting to 93% or 3700 Mha in 2015 (Table 11.4; FAO 2015). A further 1204 Mha are covered by other wooded land which does not fully meet the criteria for forest land distributed among tropical (43%), sub-tropical (33%), temperate (14%), and boreal (10%) regions (Table 11.3; Keenan et al. 2015). Europe (including the Russian Federation) has the largest forest area than any other region (25%), followed by South America (21%) and North and Central America. Three quarters of all forest is in high-income and upper middle-income countries, with about 25% of the total forests in countries with economies classified as lower or middle income. Various natural and anthropogenic induced disturbances exert profound impacts on global forests. About 60% of the world’s forest land area is recovering from the past disturbance and nearly 3% of the forest land is disturbed annually by logging, fire, pests, or weather related catastrophes such as drought. The FAO estimates of forest land area established from forest inventories and country reports differ from similar estimates based on remote sensing. For example, spatially explicit estimates of forest area based on remote sensed land cover indicated that FAO estimates based on land use are 94% of land cover based estimates (Erb et al. 2007). FAO has also used a satellite-based land cover approach to complement the inventory approach, and these independent results show slightly smaller global forest area that is 96% of the inventory approach (D’Annunzio et al. 2014). These differences are attributed to inherent contrasts in land use versus land cover indicators. Estimates based on tree cover may include land with trees that does not meet the inventory definition of forest because the observed tree cover may be below the percentage threshold used for classifying forest from inventory perspective or sometimes another land use that has significant tree cover such as commonly observed on urban or developed land. Land use indicators used in inventory estimates usually include land that are temporarily without trees such as recently harvested but not regenerated as forest land, whereas these areas can be classified as non-forest land type based satellite-based or remote sensed land cover because they lack observable tree cover.
Overall, the global forest area decreased by 3.1% between 1990 and 2015 (Table 11.2). Agriculture expansion has been the most important cause of recent forest loss, accounting for 80% of deforestation worldwide, and primarily during 1980s and 1990s through conversion of tropical forests (Gibbs et al. 2007; Houghton 2007). Climate change induced by anthropogenic GHG emissions is also becoming an important factor shaping forests globally (Walther 2010). Climate change triggers changes in disturbance regimes such as increased frequency and intensity of wild fires, windstorms, and insect outbreaks (Dale et al. 2001). In addition, altered rainfall patters and increased global temperature causes drought and heat stress around the world, resulting into increased tree mortality and sometimes forest die-off as well as decreased forest productivity as a result of interaction of multiple factors associated with the global climate change (Allen et al. 2010; Kurz et al. 2008; Phillips et al. 2009).
Although the total global area of forest land has declined, the category of planted forest has been increasing (Table 11.4). The area of protected forest has also increased, and management intensity of forest has increased significantly with substantial gains in the area under planted forests and forests with management plan (Keenan et al. 2015; FAO 2015). Consequently, the trends in unmanaged natural forest have been sharply declining (Table 11.4). Therefore, human-induced deforestation from 1990 to 2015 was partly offset by increases in forest area that has both natural regrowth after abandonment and human causes such as plantation forests. The net change in forest area is a result of net effect of forest clearance and conversion to other land uses (deforestation) and afforestation (where forest is planted or regenerates naturally on previously cleared land) and reforestation (where trees are planted or regenerates naturally on lands classified as forest). The rate of net forest loss has halved over the 25-year period, falling from 7.3 Mha yr−1 in the 1990s to 3.3 Mha yr−1 between 2010 and 2015 (Table 11.2), although this reduction is not equivalent to the reduced rates of human-induced deforestation . Deforestation or forest conversion is more complicated than just forest area decrease, because globally, forest gain and losses occur continuously and new forests of different type and environmental benefits are generally created, which are much harder to monitor (FAO 2015). Natural forest change is a better indicator of natural habitat and biodiversity dynamics. The primary forest is defined as: “naturally regenerated forest of native species where there are no clearly visible indications of human activities and the ecological processes are not significantly disturbed” (Morales-Hidalgo et al. 2015). Their key characteristics include: showing natural forest dynamics, such as natural tree species composition, occurrence of dead wood, natural age structure, and natural regeneration processes; the area is large enough to maintain its natural characteristics; there has been no known significant human intervention or the last significant human intervention was long ago enough to have allowed the natural species composition and processes to have become re-established. Global primary forest area accounted for 35% in 2015 (FAO 2015). The global primary forest area declined by 6% between 1990 and 2015 (Table 11.4; Keenan et al. 2015), due to forest clearance followed by conversion to other land use (i.e., deforestation ) in some areas, forest fire, or drought that has led to total deforestation. Fire is the most significant cause of forest loss in boreal forests (Potapov et al. 2008), while deforestation is the dominant cause of forests loss in the tropical ecozone (Hansen et al. 2013). The tropical dry forests of South America have the highest rate of tropical forest loss due to deforestation . The largest loss (11%) occurred in the tropics, while temperate regions gained natural forest, mainly from regeneration in Europe and Asia (Table 11.4; FAO 2015; Keenan et al. 2015). Primary forests are generally reclassified as other naturally regenerated forests after the disturbance. Overall, disturbances varying in type, scale, intensity, and frequency create complex mosaic of forest distribution globally and high landscape scale diversity, while also setting the initial conditions for dynamics and structural development (Swanson et al. 2011).
Natural forest generally describes forest vegetation that evolved naturally in an area, while plantation forest includes intensively managed forest plantations purposely established, commonly composed of single tree species, established to give priority to wood production. Conversion of natural forest area into an intensively managed forest plantation of exotic tree species may increase timber production potential but will generally reduce its biodiversity and ecosystem services. Some plantation forests are also established for land conservation, coastal stabilization, biodiversity conservation or other purposes. Loss of natural forest in some parts are offset by afforestation and plantation forest elsewhere. Protective forest area remained fairly constant over the 25-year period with a mean of 35.9% of the total forest area. The main categories for the protective forest area in 2015 were clean water (3.5%), coastal stabilization (0.83%), desertification control (3.6%), avalanche control (3.1%), public recreation (4.5%), C storage (1.3%), cultural services (1.9%), and other services (2.0%) (Miura et al. 2015). The land area with trees outside forest has increased as the developed land and urban area have increased (Table 11.3; Guo et al. 2014). Also, some of formerly dry woodlands not satisfying the definition of forest have greened (Brandt et al. 2015; Piao et al. 2015).
In addition to both natural causes such as drought, natural fires, storms, and diseases and pests and also anthropogenic activities such as clearance for agriculture, changes in land use and land cover from forest to non-forest are also associated with over-exploitive timber harvesting, the expansion of settlements, and infrastructure development. Change of forest to other land uses are linked to a complex and multifaceted set of underlying driving forces including population growth, poverty, government policies, technological development, rural to urban migration, changes in cultural attitudes towards forests lack of stronger incentives for conservation.
11.5 Global Forest Biomass Accumulation and Productivity Trends
Plants fix CO2 as organic compounds through photosynthesis, a C flux also known at the ecosystem level as gross primary production (GPP). The terrestrial GPP is the largest global C flux of the global C cycle in the terrestrial ecosystem, and it drives several ecosystem functions such as respiration and growth, while impacting all of the global C cycle (Beer et al. 2010). It also forms the basis for both human and animal food, and also fiber and wood production. In addition, GPP together with respiration is one of the major processes controlling land-atmosphere CO2 exchange, and accurate quantification of GPP provides a sound scientific foundations for predicting future changes in atmospheric CO2 through understanding C sources and sinks, while also defining the management options for the global C cycling (Yu et al. 2014). The global GPP is estimated at 122 Pg C yr−1, and 49% of this occurs in forest (Beer et al. 2010), while other lands, including croplands, grasslands, and savannahs also have significant rates of GPP (Table 11.5). The global NPP is estimated to be approximately half of GPP (Table 11.5), but C use efficiency (CUE, i.e., the ratio of NPP to GPP) describes the capacity of forests to transfer C from the atmosphere to the tree biomass, and it varies among biomes (DeLucia et al. 2007).
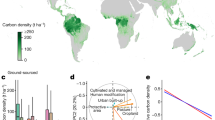
On an annual basis, the tropics have the highest biomass accumulation, followed by temperate and the boreal (Table 11.5). Boreal forests have very high NPP in their growing season but the growing season is very short in boreal regions. Boreal forests occupy large areas of northern hemisphere, mainly found in Canada, Russia, Alaska and Scandinavian region. Biodiversity is generally low in boreal forests. It is estimated that boreal forests contain about 60–100 Mg C ha−1, of which, about 80% is in the aboveground biomass (Luyssaert et al. 2007; Malhi et al. 1999). Decomposition of OM is generally slow due to low temperatures, and therefore, large accumulation of C exists in soil pool, estimated at 116–343 Mg C ha−1 (Malhi et al. 1999; Amundson 2001). Temperate forests are found in climate with distinct seasons—a well-defined winter and regular precipitation. The overall C store has been estimated to be between 150 and 320 Mg C ha−1, of which, plant biomass in the form of large woody aboveground and deep coarse root system accounts for about 60% and the soil C is about 40% (Amundson 2001). The warm and wet tropical moist forests result in rapid plant growth and most of its C is found in the vegetation with biomass estimates of 150–250 Mg C ha−1 (Amundson 2001; Chave et al. 2008; Lewis et al. 2009b). On average, the tropical forests are estimated to store 160 Mg C ha−1 in the aboveground biomass, and about 40 Mg ha−1 belowground in roots. Soil C stocks are estimated at about 90 Mg C ha−1 and somewhat lower than biomass stock (Amundson 2001). Currently tropical forests are considered to be a C sinks with recent research indicating an annual global uptake of about 1.3 Pg C yr−1, Central and South America is estimated to takes about 0.6 Pg C yr−1, African forests about 0.4 Pg C yr−1, and Asian forests about 0.25 Pg C yr−1 (Lewis et al. 2009a). Tropical forests account for two-thirds of all terrestrial biomass, while temperate and boreal forests are each equivalent to ~20% of tropical biomass. Globally, forests account for 92% of all biomass, and therefore, the distribution of forests is equivalent to the global distribution of biomass (Prentice et al. 2011).
Ice core records reveal that glacial periods were associated with lower atmospheric CO2 concentrations (Siegenthaler et al. 2005; Luethi et al. 2008; Higgins et al. 2015). Although low CO2 concentrations are not the cause of glaciations, the concentrations of 170–200 ppm were repeatedly reached during the glacial maxima. The last glacial maximum (LGM) from 26,500 to 19,000 years ago (Clark et al. 2009) is the cold glacial period with the most comprehensive paleodata. The atmospheric CO2 concentration was also ~75 ppm lower than Holocene period. Studies from paleoecology and modeling suggest that the global C storage in vegetation and soils was smaller, with GPP and NPP estimated at approximately 86.3 and 41.0 Pg C yr−1, respectively, and the global terrestrial aboveground biomass C of ~340 Pg C (Adams et al. 1990; Prentice et al. 2011). After the LGM, however, the global C storage almost doubled and peaked at ~770 Pg C during the pre-agricultural disturbance era ~10,000 years ago (Adams et al. 1990; Prentice et al. 2011). Based on global forest aggregation field observations, current aboveground forest biomass estimates are 470 Pg C (Table 11.7) and the global C of the forest living biomass is estimated at ~270 Pg C (Kohl et al. 2015), suggesting large potential for C sequestration in global biomass. Human utilization of biomass products is mostly responsible for the actual and potential biomass globally. It is estimated that currently humans are removing ~16.3 Pg C yr−1, approximately a quarter of the global NPP (Haberl et al. 2007) in the form of biomass products such as food, fiber, wood products and bioenergy. In addition, forest degradation which lowers biomass density has played significant role in reducing the capacity of forests to provide the needed goods and services globally. Human-caused reduction in forest biomass C varies across the biomes. For example, in the tropical zones forest biomass is 74%, while in boreal zones where forests occur naturally, the current biomass is only 30% of what the potential would be without human use of the land for food production, fiber, and other non-forest land uses (Table 11.5). Even as many countries in temperate zone are experiencing transitions from net deforestation to net reforestation in terms of land area, the land area under forest and also biomass C density still remain lower than that during the pre-agricultural state (Table 11.5).
Based on pre-agricultural biomass content in the global biomass, it is estimated that global forests currently contain about half of the biomass that would be present without human use of land for food, fiber, and other non-forest uses (Table 11.5; Prentice et al. 2011; Pan et al. 2013). However, how much of the potential increase in C stocks could be realized as the part of mitigation programs to offset the CO2 increase will mostly depend on societal economic and social constraints. Increasingly, C stock are not the only management goal of forests or other lands. Nabuurs et al. (2007) conducted a comprehensive analysis and estimated that the global forests activities have mitigation potential to sequester an additional 3.8 Pg C yr−1 from afforestation , reduced deforestation , and improved forest management at a cost of $50 to 100 per ton of CO2. About 1.6 Pg C yr−1 of the estimated 3.8 Pg C yr−1 could be achieved from reduced deforestation (Nabuurs et al. 2007). This will increase the gross C uptake by forests estimates from 4.0 Pg yr−1 (Pan et al. 2011) to about 6.2 Pg C yr−1, a potentially maximum C sequestration by global forest ecosystems that can be expected.
The live biomass C store of the global forest was estimated by FAO in 2015 to be 296 Pg C in both above- and below-ground biomass, about half of total C stored in forests. The global average C density is estimated at 74 Mg ha-1, with the highest densities in South America and Western and Central Africa storing 120 Mg C ha-1 in living biomass (FAO 2015). Pan et al. (2011, 2013) reported live biomass C stock densities for the tropical forests of 134 Mg C ha-1, with an estimated tropical live biomass C of 248 Pg C (Table 11.6). In contrast, Saatchi et al. (2011) reported live biomass C stock densities for tropical forests of 100.5 Mg C ha-1, similar to those repoted by FAO (2010, 2015), suggesting an overestimation for both tropical forest C stock density and biomass C by Pan et al. (2011, 2013). The FAO biomass C stock densities for tropical forests sub-regions rang around those of Saatchi et al (2011) estimate, with the highest being South America at 122.4 Mg C ha-1 and the lowest being Eastern and South Africa at 69.5 Mg C ha-1 (FAO 2010, 2015). Compared to 1990, the global biomass declined by 0.52% with largest decline occurring in subtropical forests (−8.3%). The temperate forests saw 27.1% increase in biomass relative to 1990 (Table 11.6). Although the global forest biomass has been fairly stable since 1990, the biome-specific changes reflects the status and trends of recent human use of biomass and the environmental factors in different regions of the Earth. The biomass of temperate forests is increasing, probably because of combination of factors that include continuing regrowth following abandonment from agricultural land, logging, and environmental factors such as N deposition, increasing atmospheric CO2 concentration and changing climate (Pan et al. 2011). Timber production has remained relatively stable since 1990, although areas of forest used for non-wood forest products indicate that harvesting is taking place on smaller proportion of total forest area while harvesting for biofuel has increased in recent days. The indirect effect of concentrating timber production on a smaller land base and establishment of plantation forests is to reduce harvesting on other forest areas where C stocks may increase in the absence of harvest. However, these areas are subject to higher C losses from natural disturbances including drought, as evidenced in Western USA and other areas (Allen et al. 2010; Hicke et al. 2012).
11.5.1 Emission Trends and Drivers
Estimating anthropogenic component of gross and net forest land use emissions is difficult compared to other sectors since (i) it is not always possible to separate anthropogenic and natural GHG fluxes from forest land, (ii) the input necessary to estimate GHG emission globally and regionally often based on country level statistics or on remote sensing information are highly uncertain, (iii) methods for estimating GHG emissions uses a range of approaches from simple default methodologies such as the IPCC guidelines (IPCC 2006) to more complex estimates based on the terrestrial C cycle modeling and/or remote sensing information.
Fluxes resulting directly from forest land use activities are dominated by CO2, primarily emissions from deforestation , but also an uptake due to reforestation/regrowth. Non-CO2 GHG emissions from forest land use are small and mainly arise from peat degradation through drainage and biomass fires. Estimated land use change emissions from 1750 to 2014 are 190 ± 65 Pg C, about 31% of anthropogenic CO2 emission, and 10% of anthropogenic CO2 emission in 2004–2015 (Houghton et al. 2012; Le Quéré et al. 2015, 2016). The declining fraction is largely the result of the increasing fossil fuel emissions and also declining emission from land use change. The net flux of CO2 from the land use change is also the most uncertain term in the C budget, accounting for emissions of 1.4 ± 0.5 Pg C yr−1 in 1980s, 1.6 ± 0.5 Pg C yr−1 in 1990s, and 1.1 ± 0.5 Pg C yr−1 in 2000s (Houghton et al. 2012; Le Quéré et al. 2015). The terrestrial land C sink from 1750 to 2015, mostly as a result of forest uptake, estimated as a residual C after accounting for atmospheric CO2 concentration increase and ocean sink, is 165 ± 70 Pg C (Le Quéré et al. 2015, 2016), suggesting a net emission of ~25 Pg C from land use change during the Industrial Era.
Human-induced global environmental changes exert complex effects on forest productivity and C storage (Friedlingstein et al. 2006). Some of the factors driving the environmental changes are direct and physically alter forest areas and structures—such as tropical deforestation which destroy vegetation structures and habitat, and immediately releases large amounts of C stored in biomass as CO2. However, some factors operate through tree physiology and ecological processes—such as changes in climate (temperature, precipitation, radiation) and atmospheric composition (e.g., CO2 concentration, N deposition, O3), and other pollutants (Boisvenue and Running 2006; Lewis et al. 2009a). Further, these factors may interact (i.e., drought induced forest diebacks, changes in forest structure and morphology) and interfere with C metabolism and dynamics (Walther 2010). The average global land use emissions from 2000 to 2015 are estimated at 1.0 ± 0.5 Pg C yr−1, which is lower than 1990s emissions estimated at 1.6 ± 0.7 Pg C yr−1 (Friedlingstein et al. 2010; Le Quéré et al. 2015, 2016).
Global forest land use CO2 fluxes estimates are based on wide range of data sources and includes different processes, definitions, and approaches to calculating emissions. These include CO2 fluxes from deforestation , afforestation , forest degradation and harvesting activities (logging), regrowth of forests following wood harvest or abandonment of agriculture. Some of these managements lead to emission of CO2 to the atmosphere, while others leads to CO2 sinks. Deforestation and forest degradation, primarily in tropical regions accounted for 12–20% of global anthropogenic GHG emissions in 1990s and early 2000s (van der Werf et al. 2009). It is also expected that deforestation and forest degradation in the tropics will have the major impact for the future potential of forests to sequester the additional anthropogenic CO2 emissions (Chave et al. 2008; Lewis et al. 2009b; Field et al. 1998). Over the period 1750–2014, forest was a net source of CO2 and the land as a whole was a net sink of CO2 since 1950s as confirmed by inventory measurements in managed forests in temperate and tropical regions (Phillips et al. 1998; Luyssaert et al. 2008; Pan et al. 2011). A sink is due to indirect effects of human activity on ecosystems, i.e., environmental change effects such as the fertilizing effects of increased levels of CO2 and N in the atmosphere and the effects of on C storage (Le Quéré et al. 2009; Canadell et al. 2007a). However, some of it may be due to direct forest management change activities and not accounted for in the current estimates (Erb et al. 2013). A substantial fraction of current C sink is currently incorrectly attributed to environmental change, when it is a result of changes in management practices (Erb et al. 2013). This sink capacity is relevant to forest mitigation through forest protection. There has been a decline in forest land use emissions over the most recent years largely due to a decrease in the rate of deforestation (FAO 2015; FAOSTAT 2016). Forest land use emissions peaked in the 1980s in Asia and Latin America regions and declined thereafter, consistent with reduced rate of deforestation, notably in Brazil and some areas of afforestation in China, Vietnam, and India (FAOSTAT 2016). In the Middle East and Africa, the bookkeeping model showed continuing emissions from 1970 to 2000 (Houghton et al. 2012), while models show continuing increase from 1970 to 2000 and a small sink in the 2000s (Kato et al. 2013). The general picture in temperate and boreal regions is of declining emissions and/or increasing sinks. The temperate regions include a large areas of managed forests subjected to harvest and regrowth and areas of reforestation e.g., following cropland abandonment in USA and Europe.
The bookkeeping estimate (Houghton 2003, 2012) uses regional biomass growth and decay rates from the country inventories that are generally not adjusted for changes in climate or CO2 concentration (i.e. CO2 fertilization). It takes into account the forest management associated with shifting cultivation in tropical forest regions and also global wood harvest and regrowth cycles. The primary source of data for bookkeeping model is FAO forest area data and wood harvest (FAO 2015). The quality and reliability of the country inventory data varies considerably because not all countries have well-established forest inventory and monitoring programs. The FAO estimate of CO2 emission with reference to 2001–2010 decade indicated that forest emissions were 0.9 Pg C yr−1, of which, deforestation was 1.04 Pg C yr−1, forest degradation and management was −0.5 Pg C yr−1, biomass fires emissions were 0.1 Pg C yr−1 and drained peatlands contributed 0.25 Pg C yr−1 (FAOSTAT 2016). The FAO estimated that decadal net forest emissions decreased from 1.1 Pg C yr−1 in 1991–2000 to 0.9 Pg C yr−1 in 2001–2010 (FAOSTAT 2016). Using forest inventory data and long-term C studies, Pan et al. (2011) estimated a total global forest C sink of 2.4 ± 0.4 Pg C yr−1 for 1990–2007. The emission source from tropical land use change was also estimated at 1.3 ± 0.7 Pg C yr−1 of which tropical deforestation emissions of 2.9 ± 0.5 Pg C yr−1 is partially offset by C forest regrowth of 1.6 ± 0.5 Pg C yr−1 (Pan et al. 2011). The global forest C fluxes therefore, comprised of a net global sink of 1.1 ± 0.8 Pg C yr−1, however, tropical estimates have the highest uncertainties. The FAO global forest resources assessment data show that the growing forest biomass volume has increased in East Asia, western and Central Asia, North America, Europe, Caribbean, and Oceania, while Africa and South America observed significant decrease in C pools between 1990 and 2015 (Table 11.7, FAO 2015). Overall, FAO reported decrease in global forest C pools between 1990 and 2015 (Table 11.8), mainly driven by decrease in South America and Africa.
11.6 Climate Change Mitigation Options Under Forest
Forests and climate change are intrinsically linked beyond C sequestration. Climate change and global warming could change forest landscape worldwide. Higher mean annual global temperatures, altered precipitation patterns, and more frequent extreme weather events may have adverse effects on forests, including stress, compositional and fractional changes and changes in the capacity of forests to provide products and services, and these effects are poorly understood. Since forest ecosystems capture and store CO2 and make major contribution in the mitigation of climate change, when forests are destroyed through over-harvesting or burning they become a major source of CO2 emissions. Conversion of forests to agricultural land also leads to the loss of soil organic C (SOC). The largest loss in SOC stock was reported when temperate (52%) followed by tropical forests (41%) and boreal forests (31%) were cleared (Wei et al. 2014). Due to the observed large losses of SOC resulting from conversion of forests to agricultural land, C storage capacity of the land will be increased through afforestation or decreasing deforestation (Poeplau et al. 2011; Wei et al. 2014). Afforestation of temperate croplands creates C sink of about 116% higher than cropland for the period of 100 years, and no new equilibrium was reached within 120 years (Poeplau et al. 2011).
Concerns about the role of forests in global climate have led to creation of collaborative mechanism for reducing emissions from deforestation and forest degradation under the United Nations Framework Convention on Climate Change (UNFCCC) with the goal of significantly reducing emissions from deforestation and forest degradation (REDD+) global partnership and collaboration (UN-REDD 2010). The UNFCCC has outlined two major goals with respect to forests: (i) using forests for biological C capture and storage, thus reducing the emissions of GHGs, and therefore, forests becomes part of mitigation strategy for climate change, (ii) using forests and tree biomass as part of strategy to cope with impacts of climate change, therefore forests become part of a climate strategy for adaptation (Van Bodegom et al. 2009). Forest-based climate change mitigation measures are low-cost and effective in short to medium term. Mitigation activities in forest can reduce climate forcing through: (i) conservation of existing C pools in forest (i.e., conservation of forest biomass, peatlands and soil C that could otherwise be mineralized, (ii) reduction of C losses from biota and soils through management changes within the same land use type (e.g., reducing deforestation , recreating wetlands and peat lands), (iii) enhancement of soil C sequestration, increasing C uptake by biota and long-lived products through tree planting—reforestation and/or afforestation , (iv) increasing C storage per unit area such as increasing stocking density in forests and wood use in construction, (v) changes in albedo resulting from land cover change that result in increased reflection of visible light, and (vi) use of forest products to replace high emission products such as replacing concrete and steel in construction with wood and also use of forest byproducts for bioenergy to replace fossil fuels. Summarized mitigation options associated with forest land use are presented in Table 11.9.
Summary of option for mitigation under forests are presented in Table 11.9. Forest ecosystems contain up to 80% of all aboveground terrestrial C (Dixon et al. 1994). Forests stores vast pools of C globally and even small shifts in the balance between photosynthesis and ecosystem respiration result in a large change in the uptake or emission of CO2 from forests to the atmosphere. Processes that increase terrestrial C sequestration include land and soil management practices that enhance the storage of C, or reduce CO2 emissions. Photosynthesis process can be enhanced by the processes that deliberately augment natural C uptake in plants. The rate of net CO2 uptake by forests varies in a predictable way as forests grow from establishment or recover from the past disturbances (Pregitzer and Euskirchen 2004). Productivity in forest is generally a function of light and N supply (Hardiman et al. 2013). Investigations of forest NPP over the course of ecological succession support a general trend of declining production with forest age, but with knowledge gaps for late successional, mixed forests (Gough et al. 2008). Quantitative syntheses reports lower NPP in old-growth rather than young aggrading forests (Pregitzer and Euskirchen 2004; Luyssaert et al. 2008). Understanding factors that influence the rate of CO2 uptake by vegetation is the basis for improving the forest management (Birdsey et al. 2006).
Forest C storage can be increased by planting of new forests on previously unforested lands (i.e., afforestation ) or on previously forested lands (i.e., reforestation, for example, on lands that were previously cleared for agriculture) (Sochacki et al. 2012; Bustamante et al. 2014). This can include mixed species or monocultures. Afforestation and reforestation also provides social, economic, and other environmental benefits. Forest restoration in the degraded forests and protecting secondary forests with low C densities while allowing them to sequester C either by natural or artificial regeneration and long term fallows will also increase C sequestration in forest (Reyer et al. 2009). Other activities that promote C storage in forests include improved forest management and reducing deforestation or deforestation avoidance (van Bodegom et al. 2009). Improved management to enhance productivity, controlling diseases and fire management during the forest lifecycle, extending rotation cycles, and reducing logging waste also increases C sequestration (DeFries and Rosenzweig 2010). Protecting forests form wild fires increases C stocks. However, with climate change effects, it may also increase a risk of larger releases of stored C (Westerling et al. 2006). Afforestation of abandoned agricultural land and degraded land generally increases soil C in addition to producing wood biomass (Guo and Gifford 2002). Avoiding deforestation and forest degradation preserves existing C stock that would otherwise be lost to the atmosphere. C storage in managed forests can also be enhanced by timber harvest and tree planting practices that increase C density (i.e., OC per unit area) and reduce forest degradation. The C sequestration can also be enhanced through management of forest stands on farms (agroforestry) and in urban and suburban settings (urban forestry). However, full accounting for different management action requires knowledge of changes in ecosystem C pools (Echeverria et al. 2004), harvesting wood products (Schlamadinger and Marland 1996) and fossil fuel emission associated with growing harvesting, and manufacturing (Schlamadinger et al. 1997).
One of the climate mitigation potential of forest which has not received enough attention is the biophysical effects of land management on climate which goes beyond C storage to account for surface albedo and evapotranspiration which have direct effects on climate. Some aspects of this have been evaluated with respect to land use change . However, Luyssaert et al. (2014) noted that biophysical changes within land class may have an effect of similar magnitude to that of changes in land class.
11.7 Conclusions
Forests, climate, climate change, and climate change mitigation are intricately linked. Forests cycle large amounts of C and, with proper management they could be utilized to remove anthropogenic CO2 emissions from the atmosphere. Annual land CO2 fluxes are dominated by forests, and it could become a large source of CO2 emissions or substantial part of the mitigation depending on how they are treated internationally and managed locally. Terrestrial ecosystems, dominantly forests, have so far played an important role in controlling the increase in atmospheric CO2 concentrations by removing about one third of the anthropogenic CO2 emissions each year. However, the historical consequence of increasing intensity of global land management has been the long-term reduction of terrestrial C stocks as land has been converted to crops, pasture, and settlement for human life support. As a result, climate mitigation policies of potential to increase C stocks on lands with lower C stocks than expected compared with lands such as highly stocked old growth forests that have experienced relatively less disturbance is high. Generally, much of the world’s forest land has been permanently converted to other uses such as food production and urban settlement, and therefore, are not likely to revert back to forest. Also, other areas of forest are needed for fiber production and other social and societal use, and increasing tree stocking and C density in this category can occur without negative effects on provision of other services. Currently, tropical forests are subjected to highest level of deforestation , and therefore, account for nearly all net emissions from forests. Large quantities of C are released when forests are cleared and burnt, and more GHGs are emitted from subsequent land uses after conversion from forest to the land uses such as agriculture. Industrial agriculture and commercial logging are the main drivers of deforestation and degradation, especially in tropical South America and Southeast Asia, the hotspots for forest-related CO2 emissions. Emissions from land use change have been declining since 1990. However, this trend is misleading, because emission from other have been increasing at higher rates. Although several attempts have been made to estimate the forest potential mitigation, important C accounting considerations regarding forests is still lacking. This include C that is removed from forest in harvest operation and retained in wood products pools or discarded in landfills, which is not immediately returned to the atmosphere. Another C pool which is not fully accounted for is the substitution for using wood instead of other materials for construction which is not generally documented. These limits the ability of conducting lifecycle analysis to determine the energy consumption, emissions and sequestration throughout the full cycle of growing, harvesting, and processing wood compared to other materials. Compared to other mitigation strategies, forest-related activities, especially reducing emissions from tropical deforestation are among the most economically feasible and cost-effective options and the most viable strategy in the land sector. Reducing CO2 emissions from forestry plays a relatively large role in reducing total CO2 emissions in the short term and buy time to develop other mitigation and adaptation strategies. Forests are already removing significant quantities of CO2 from the atmosphere naturally, which, coupled with other mitigation strategies remains essential for meeting long-term climate targets.
References
Adams JM, Faure H, Faure-Denard L, McGlade JM, Woodward FI (1990) Increases in terrestrial carbon storage from the Last Glacial Maximum to the present. Nature 348(6303):711–714
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J-H, Allard G, Running SW, Semerci A, Cobb N (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259(4):660–684. doi:10.1016/j.foreco.2009.09.001
Amundson R (2001) The carbon budget in soils. Annu Rev Earth Planet Sci 29(1):535–562. doi:10.1146/annurev.earth.29.1.535
Asner GP, Mascaro J, Muller-Landau HC, Vieilledent G, Vaudry R, Rasamoelina M, Hall JS, van Breugel M (2012) A universal airborne LiDAR approach for tropical forest carbon mapping. Oecologia 168(4):1147–1160. doi:10.1007/s00442-011-2165-z
Beer C, Reichstein M, Tomelleri E, Ciais P, Jung M, Carvalhais N, Roedenbeck C, Arain MA, Baldocchi D, Bonan GB, Bondeau A, Cescatti A, Lasslop G, Lindroth A, Lomas M, Luyssaert S, Margolis H, Oleson KW, Roupsard O, Veenendaal E, Viovy N, Williams C, Woodward FI, Papale D (2010) Terrestrial gross carbon dioxide uptake: global distribution and covariation with climate. Science 329(5993):834–838. doi:10.1126/science.1184984
Bellassen V, Luyssaert S (2014) Managing forests in uncertain times. Nature 506(7487):153–155
Birdsey RA, Heath LS (2001) Forest inventory data, models, and assumptions for monitoring carbon flux. In: Lal R, Follett RF (eds) Soil carbon sequestration and greenhouse effect. Soil Science Society of America Inc., WI, USA, pp 125–135
Birdsey R, Pregitzer K, Lucier A (2006) Forest carbon management in the United States: 1600–2100. J Environ Qual 35(4):1461–1469. doi:10.2134/jeq2005.0162
Birdsey R, Angeles-Perez G, Kurz WA, Lister A, Olguin M, Pan Y, Wayson C, Wilson B, Johnson K (2013) Approaches to monitoring changes in carbon stocks for REDD. Carbon Manag 4(5):519–537. doi:10.4155/cmt.13.49
Boisvenue C, Running SW (2006) Impacts of climate change on natural forest productivity—evidence since the middle of the 20th century. Glob Change Biol 12(5):862–882. doi:10.1111/j.1365-2486.2006.01134.x
Bond-Lamberty B, Wang C, Gower ST (2002) Annual carbon flux from woody debris for a boreal black spruce fire chronosequence. J Geophys Res Atmos 108(D3). doi:10.1029/2001jd000839
Bradford JB, Birdsey RA, Joyce LA, Ryan MG (2008) Tree age, disturbance history, and carbon stocks and fluxes in subalpine Rocky Mountain forests. Glob Change Biol 14(12):2882–2897. doi:10.1111/j.1365-2486.2008.01686.x
Brandt M, Mbow C, Diouf AA, Verger A, Samimi C, Fensholt R (2015) Ground- and satellite-based evidence of the biophysical mechanisms behind the greening Sahel. Glob Change Biol 21(4):1610–1620. doi:10.1111/gcb.12807
Brown S (1997) Estimating biomass and biomass change of tropical forests: a primer, vol 134. Food & Agriculture Org
Bustamante M, Robledo-Abad C, Harper R, Mbow C, Ravindranat NH, Sperling F, Haberl H, Pinto AdS, Smith P (2014) Co-benefits, trade-offs, barriers and policies for greenhouse gas mitigation in the agriculture, forestry and other land use (AFOLU) sector. Glob Change Biol 20(10):3270–3290. doi:10.1111/gcb.12591
Canadell J, Pataki D, Gifford R, Houghton R, Luo Y, Raupach M, Smith P, Steffen W (2007a) Saturation of the terrestrial carbon sink. In: Canadell J, Pataki D, Pitelka L (eds) Terrestrial ecosystems in a changing world. Global change—the IGBP series. Springer, Berlin, pp 59–78. doi:10.1007/978-3-540-32730-1_6
Canadell JG, Le Quéré C, Raupach MR, Field CB, Buitenhuis ET, Ciais P, Conway TJ, Gillett NP, Houghton RA, Marland G (2007b) Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proc Natl Acad Sci USA 104(47):18866–18870. doi:10.1073/pnas.0702737104
Chave J, Andalo C, Brown S, Cairns MA, Chambers JQ, Eamus D, Folster H, Fromard F, Higuchi N, Kira T, Lescure JP, Nelson BW, Ogawa H, Puig H, Riera B, Yamakura T (2005) Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145(1):87–99. doi:10.1007/s00442-005-0100-x
Chave J, Olivier J, Bongers F, Chatelet P, Forget P-M, van der Meer P, Norden N, Riera B, Charles-Dominique P (2008) Above-ground biomass and productivity in a rain forest of eastern South America. J Trop Ecol 24:355–366. doi:10.1017/s0266467408005075
Chen JM, Ju WM, Cihlar J, Price D, Liu J, Chen WJ, Pan JJ, Black A, Barr A (2003) Spatial distribution of carbon sources and sinks in Canada’s forests. Tellus B 55(2):622–641. doi:10.1034/j.1600-0889.2003.00036.x
Chen W, Moriya K, Sakai T, Koyama L, Cao CX (2016) Mapping a burned forest area from landsat TM data by multiple methods. Geomatics Nat Hazards Risk 7(1):384–402. doi:10.1080/19475705.2014.925982
Ciais P, Sabine CL, Govindasamy B, Bopp L, Brovkin V, Canadell J, Chhabra A, DeFries R, Galloway J, Heimann M, Jones C, Le Quéré C, Myeneni R, Piao S, Thornton P (2013) Carbon and other biogeochemical cycles. In: Stocker TF, Qin D, Plattner G-K et al (eds) Climate change 2013: physical science basis. Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, UK and New York, USA, pp 465–570
Clark PU, Dyke AS, Shakun JD, Carlson AE, Clark J, Wohlfarth B, Mitrovica JX, Hostetler SW, McCabe AM (2009) The last glacial maximum. Science 325(5941):710–714. doi:10.1126/science.1172873
Cohen WB, Goward SN (2004) Landsat’s role in ecological applications of remote sensing. Bioscience 54(6):535–545. doi:10.1641/0006-3568(2004)054[0535:lrieao]2.0.co;2
Cohen WB, Harmon ME, Wallin DO, Fiorella M (1996) Two decades of carbon flux from forests of the Pacific northwest. Bioscience 46(11):836–844. doi:10.2307/1312969
Corona P, Marchetti M (2007) Outlining multi-purpose forest inventories to assess the ecosystem approach in forestry. Plant Biosyst 141(2):243–251. doi:10.1080/11263500701401836
D’Annunzio R, Lindquist E, MacDicken KG (2014) Global forest land—use change from 1990 to 2010: an update to a global remote sensing survey of forests. Food and Agricultural Organization (FAO), Rome, Italy, p 6
Dale VH, Joyce LA, McNulty S, Neilson RP, Ayres MP, Flannigan MD, Hanson PJ, Irland LC, Lugo AE, Peterson CJ, Simberloff D, Swanson FJ, Stocks BJ, Wotton BM (2001) Climate change and forest disturbances. Bioscience 51(9):723–734. doi:10.1641/0006-3568(2001)051[0723:ccafd]2.0.co;2
Dean C, Roxburgh S, Mackey BG (2004) Forecasting landscape-level carbon sequestration using gridded, spatially adjusted tree growth. For Ecol Manag 194(1–3):109–129. doi:10.1016/j.foreco.2004.02.013
DeFries R, Rosenzweig C (2010) Toward a whole-landscape approach for sustainable land use in the tropics. Proc Natl Acad Sci USA 107(46):19627–19632. doi:10.1073/pnas.1011163107
DeLucia EH, Drake JE, Thomas RB, Gonzalez-Meler M (2007) Forest carbon use efficiency: is respiration a constant fraction of gross primary production? Glob Change Biol 13(6):1157–1167. doi:10.1111/j.1365-2486.2007.01365.x
Delzon S, Loustau D (2005) Age-related decline in stand water use: sap flow and transpiration in a pine forest chronosequence. Agric For Meteorol 129(3–4):105–119. doi:10.1016/j.agrformet.2005.01.002
Dixon RK, Brown S, Houghton RA, Solomon AM, Trexler MC, Wisniewski J (1994) Carbon pools and flux of global forest ecosystems. Science 263(5144):185–190. doi:10.1126/science.263.5144.185
Echeverria ME, Markewitz D, Morris LA, Hendrick RL (2004) Soil organic matter fractions under managed pine plantations of the southeastern USA. Soil Sci Soc Am J 68(3):950–958
EPA (2015) Inventor of US greenhouse gas: emissions and sinks 1990–2013. US Environmental Protection Agency, EPA 430-R-15-004, Washington, DC, USA
Erb K-H, Gaube V, Krausmann F, Plutzar C, Bondeau A, Haberl H (2007) A comprehensive global 5 min resolution land-use data set for the year 2000 consistent with national census data. J Land Use Sci 2(3):191–224. doi:10.1080/17474230701622981
Erb K-H, Kastner T, Luyssaert S, Houghton RA, Kuemmerle T, Olofsson P, Haberl H (2013) Commentary: bias in the attribution of forest carbon sinks. Nat Clim Change 3(10):854–856
FAO (2010) Global forest resource assessment 2010. Food and Agriculture Organization of the United Nations, Rome, Italy
FAO (2013) Forests and water—international momentum an action. Food and Agriculture Organization of the United Nations, Rome, Italy, 76 pp
FAO (2014) State of the world’s forests: enhancing the socioeconomic benefits from forests. Food and Agriculture Organization of the United Nations, Rome, Italy, p 133
FAO (2015) Global forest resources assessment 2015. Food and Agriculture Organization of the United Nations, pp 245. Available online at http://www.fao.org/forest-resources-assessment/en/
FAOSTAT (2016) Food and agriculture organization global statistics web page. Food and Agriculture Organization of the United Nations, Statistical Division. http://faostat3.fao.org/. Accessed Aug 2016
Field CB, Behrenfeld MJ, Randerson JT, Falkowski P (1998) Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281(5374):237–240. doi:10.1126/science.281.5374.237
Friedlingstein P, Cox P, Betts R, Bopp L, Von Bloh W, Brovkin V, Cadule P, Doney S, Eby M, Fung I, Bala G, John J, Jones C, Joos F, Kato T, Kawamiya M, Knorr W, Lindsay K, Matthews HD, Raddatz T, Rayner P, Reick C, Roeckner E, Schnitzler KG, Schnur R, Strassmann K, Weaver AJ, Yoshikawa C, Zeng N (2006) Climate-carbon cycle feedback analysis: results from the (CMIP)-M-4 model intercomparison. J Climate 19(14):3337–3353. doi:10.1175/jcli3800.1
Friedlingstein P, Houghton RA, Marland G, Hackler J, Boden TA, Conway TJ, Canadell JG, Raupach MR, Ciais P, Le Quéré C (2010) Update on CO2 emissions. Nat Geosci 3(12):811–812. doi:10.1038/ngeo1022
Gibbs HK, Brown S, Niles JO, Foley JA (2007) Monitoring and estimating tropical forest carbon stocks: making REDD a reality. Environ Res Lett 2(4). doi:10.1088/1748-9326/2/4/045023
Gough CM, Vogel CS, Schmid HP, Su HB, Curtis PS (2008) Multi-year convergence of biometric and meteorological estimates of forest carbon storage. Agric For Meteorol 148(2):158–170. doi:10.1016/j.agrformet.2007.08.004
Goulden ML, McMillan AMS, Winston GC, Rocha AV, Manies KL, Harden JW, Bond-Lamberty BP (2011) Patterns of NPP, GPP, respiration, and NEP during boreal forest succession. Glob Change Biol 17(2):855–871. doi:10.1111/j.1365-2486.2010.02274.x
Gower ST, McMurtrie RE, Murty D (1996) Aboveground net primary production decline with stand age: potential causes. Trends Ecol Evol 11(9):378–382. doi:10.1016/0169-5347(96)10042-2
Grainger A (2008) Difficulties in tracking the long-term global trend in tropical forest area. Proc Natl Acad Sci USA 105(2):818–823. doi:10.1073/pnas.0703015105
Guo LB, Gifford RM (2002) Soil carbon stocks and land use change: a meta analysis. Glob Change Biol 8(4):345–360. doi:10.1046/j.1354-1013.2002.00486.x
Guo ZD, Hu HF, Pan YD, Birdsey RA, Fang JY (2014) Increasing biomass carbon stocks in trees outside forests in China over the last three decades. BioGeosciences 11(15):4115–4122. doi:10.5194/bg-11-4115-2014
Gustafson EJ, Shvidenko AZ, Sturtevant BR, Scheller RM (2010) Predicting global change effects on forest biomass and composition in south-central Siberia. Ecol Appl 20(3):700–715. doi:10.1890/08-1693.1
Haberl H, Erb KH, Krausmann F, Gaube V, Bondeau A, Plutzar C, Gingrich S, Lucht W, Fischer-Kowalski M (2007) Quantifying and mapping the human appropriation of net primary production in earth’s terrestrial ecosystems. Proc Natl Acad Sci USA 104(31):12942–12945. doi:10.1073/pnas.0704243104
Hansen MC, Potapov PV, Moore R, Hancher M, Turubanova SA, Tyukavina A, Thau D, Stehman SV, Goetz SJ, Loveland TR, Kommareddy A, Egorov A, Chini L, Justice CO, Townshend JRG (2013) High-resolution global maps of 21st-century forest cover change. Science 342(6160):850–853. doi:10.1126/science.1244693
Hardiman BS, Gough CM, Halperin A, Hofmeister KL, Nave LE, Bohrer G, Curtis PS (2013) Maintaining high rates of carbon storage in old forests: a mechanism linking canopy structure to forest function. For Ecol Manag 298:111–119. doi:10.1016/j.foreco.2013.02.031
Heath LS, Smith JE (2000) Soil carbon accounting and assumptions for forestry and forest-related land use change. In: Joyce LA, Birdsey R (eds) Impact of climate change on America’s forests: a technical document supporting the 2000 Usda forest service RPA assessment, vol 59. US Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, Colorado, USA, pp 89–101
Heath LS, Smith JE, Skog KE, Nowak DJ, Woodall CW (2011) Managed forest carbon estimates for the US greenhouse gas inventory, 1990–2008. J For 109(3):167–173
Hicke JA, Jenkins JC, Ojima DS, Ducey M (2007) Spatial patterns of forest characteristics in the western United States derived from inventories. Ecol Appl 17(8):2387–2402. doi:10.1890/06-1951.1
Hicke JA, Allen CD, Desai AR, Dietze MC, Hall RJ, Hogg EH, Kashian DM, Moore D, Raffa KF, Sturrock RN, Vogelmann J (2012) Effects of biotic disturbances on forest carbon cycling in the United States and Canada. Glob Change Biol 18(1):7–34. doi:10.1111/j.1365-2486.2011.02543.x
Higgins JA, Kurbatov AV, Spaulding NE, Brook E, Introne DS, Chimiak LM, Yan Y, Mayewski PA, Bender ML (2015) Atmospheric composition 1 million years ago from blue ice in the Allan Hills, Antarctica. Proc Natl Acad Sci USA 112(22):6887–6891. doi:10.1073/pnas.1420232112
Hirsch T (2010) Global biodiversity outlook 3. Secretariat of the Convention on Biological Diversity of the United Nations, Montreal, Quebec, Canada, p 93
Hoshino D, Nishimura N, Yamamoto S (2001) Age, size structure and spatial pattern of major tree species in an old-growth Chamaecyparis obtusa forest, Central Japan. For Ecol Manag 152(1–3):31–43. doi:10.1016/s0378-1127(00)00614-9
Houghton RA (2003) Revised estimates of the annual net flux of carbon to the atmosphere from changes in land use and land management 1850–2000. Tellus B 55(2):378–390. doi:10.1034/j.1600-0889.2003.01450.x
Houghton RA (2005) Aboveground forest biomass and the global carbon balance. Glob Change Biol 11(6):945–958. doi:10.1111/j.1365-2486.2005.00955.x
Houghton RA (2007) Balancing the global carbon budget. Annu Rev Earth Planet Sci 35:313–347. doi:10.1146/annurev.earth.35.031306.140057
Houghton RA (2012) Carbon emissions and the drivers of deforestation and forest degradation in the tropics. Curr Opin Environ Sust 4(6):597–603. doi:10.1016/j.cosust.2012.06.006
Houghton RA, House JI, Pongratz J, van der Werf GR, DeFries RS, Hansen MC, Le Quéré C, Ramankutty N (2012) Carbon emissions from land use and land-cover change. BioGeosciences 9(12):5125–5142. doi:10.5194/bg-9-5125-2012
Huntzinger DN, Post WM, Wei Y, Michalak AM, West TO, Jacobson AR, Baker IT, Chen JM, Davis KJ, Hayes DJ, Hoffman FM, Jain AK, Liu S, McGuire AD, Neilson RP, Potter C, Poulter B, Price D, Raczka BM, Tian HQ, Thornton P, Tomelleri E, Viovy N, Xiao J, Yuan W, Zeng N, Zhao M, Cook R (2012) North American carbon program (NACP) regional interim synthesis: terrestrial biospheric model intercomparison. Ecol Model 232:144–157. doi:10.1016/j.ecolmodel.2012.02.004
IPCC (2006) 2006 national greenhouse gas inventory guidelines. In: Eggleston S, Buendia L, Miwa K, Ngara T, Tanabe K (eds) 2006 IPCC guidelines for national greenhouse gas inventories. Institute of Global Environmental Strategies (IGES), Kanagaw, Japan, 20 pp
IPCC (2014) Climate change 2014: synthesis report. In: Pachauri RK, Meyer L, Team CW (eds) Contribution of working groups I, II and III to the fourth assessment report of the intergovernmental panel on climate change. Intergovernmental Panel on Climate Change, Geneva, Switzerland, 151 pp
Jackson RB, Jobbagy EG, Avissar R, Roy SB, Barrett DJ, Cook CW, Farley KA, le Maitre DC, McCarl BA, Murray BC (2005) Trading water for carbon with biological sequestration. Science 310(5756):1944–1947. doi:10.1126/science.1119282
Kashian DM, Romme WH, Tinker DB, Turner MG, Ryan MG (2006) Carbon storage on landscapes with stand-replacing fires. Bioscience 56(7):598–606. doi:10.1641/0006-3568(2006)56[598:csolws]2.0.co;2
Kato E, Kinoshita T, Ito A, Kawamiya M, Yamagata Y (2013) Evaluation of spatially explicit emission scenario of land-use change and biomass burning using a process-based biogeochemical model. J Land Use Sci 8(1):104–122. doi:10.1080/1747423X.2011.628705
Keeling CD (1960) The concentration and isotopic abundances of carbon dioxide in the atmosphere. Tellus 12(2):200–203
Keenan RJ, Reams GA, Achard F, de Freitas JV, Grainger A, Lindquist E (2015) Dynamics of global forest area: results from the FAO global forest resources assessment 2015. For Ecol Manag 352:9–20. doi:10.1016/j.foreco.2015.06.014
Khatiwala S, Primeau F, Hall T (2009) Reconstruction of the history of anthropogenic CO2 concentrations in the ocean. Nature 462(7271):346–349. doi:10.1038/nature08526
Kindermann GE, McAllum I, Fritz S, Obersteiner M (2008) A global forest growing stock, biomass and carbon map based on FAO statistics. Silva Fenn 42(3):387–396. doi:10.14214/sf.244
Kohl M, Lasco R, Cifuentes M, Jonsson O, Korhonen KT, Mundhenk P, de Jesus Navar J, Stinson G (2015) Changes in forest production, biomass and carbon: results from the 2015 UN FAO global forest resource assessment. For Ecol Manag 352:21–34. doi:10.1016/j.foreco.2015.05.036
Kurz WA, Dymond CC, Stinson G, Rampley GJ, Neilson ET, Carroll AL, Ebata T, Safranyik L (2008) Mountain pine beetle and forest carbon feedback to climate change. Nature 452(7190):987–990. doi:10.1038/nature06777
Kurz WA, Dymond CC, White TM, Stinson G, Shaw CH, Rampley GJ, Smyth C, Simpson BN, Neilson ET, Tyofymow JA, Metsaranta J, Apps MJ (2009) CBM-CFS3: a model of carbon-dynamics in forestry and land-use change implementing IPCC standards. Ecol Model 220(4):480–504. doi:10.1016/j.ecolmodel.2008.10.018
Le Quéré C, Raupach MR, Canadell JG, Marland G, Bopp L, Ciais P, Conway TJ, Doney SC, Feely RA, Foster P, Friedlingstein P, Gurney K, Houghton RA, House JI, Huntingford C, Levy PE, Lomas MR, Majkut J, Metzl N, Ometto JP, Peters GP, Prentice IC, Randerson JT, Running SW, Sarmiento JL, Schuster U, Sitch S, Takahashi T, Viovy N, van der Werf GR, Woodward FI (2009) Trends in the sources and sinks of carbon dioxide. Nat Geosci 2(12):831–836. doi:10.1038/ngeo689
Le Quéré C, Moriarty R, Andrew RM, Canadell JG, Sitch S, Korsbakken JI, Friedlingstein P, Peters GP, Andres RJ, Boden TA, Houghton RA, House JI, Keeling RF, Tans P, Arneth A, Bakker DCE, Barbero L, Bopp L, Chang J, Chevallier F, Chini LP, Ciais P, Fader M, Feely RA, Gkritzalis T, Harris I, Hauck J, Ilyina T, Jain AK, Kato E, Kitidis V, Goldewijk KK, Koven C, Landschuetzer P, Lauvset SK, Lefevre N, Lenton A, Lima ID, Metzl N, Millero F, Munro DR, Murata A, Nabel JEMS, Nakaoka S, Nojiri Y, O’Brien K, Olsen A, Ono T, Perez FF, Pfeil B, Pierrot D, Poulter B, Rehder G, Roedenbeck C, Saito S, Schuster U, Schwinger J, Seferian R, Steinhoff T, Stocker BD, Sutton AJ, Takahashi T, Tilbrook B, van der Laan-Luijkx IT, van der Werf GR, van Heuven S, Vandemark D, Viovy N, Wiltshire A, Zaehle S, Zeng N (2015) Global carbon budget 2015. Earth Syst Data 7(2):349–396. doi:10.5194/essd-7-349-2015
Le Quéré C, Andrew RM, Canadell JG, Sitch S, Korsbakken JI, Peters GP, Manning AC, Boden TA, Tans PP, Houghton RA, Keeling RF, Alin S, Andrews OD, Anthoni P, Barbero L, Bopp L, Chevallier F, Chini LP, Ciais P, Currie K, Delire C, Doney SC, Friedlingstein P, Gkritzalis T, Harris I, Hauck J, Haverd V, Hoppema M, Klein Goldewijk K, Jain AK, Kato E, Körtzinger A, Landschützer P, Lefèvre N, Lenton A, Lienert S, Lombardozzi D, Melton JR, Metzl N, Millero F, Monteiro PMS, Munro DR, Nabel JEMS, Nakaoka SI, O’Brien K, Olsen A, Omar AM, Ono T, Pierrot D, Poulter B, Rödenbeck C, Salisbury J, Schuster U, Schwinger J, Séférian R, Skjelvan I, Stocker BD, Sutton AJ, Takahashi T, Tian H, Tilbrook B, van der Laan-Luijkx IT, van der Werf GR, Viovy N, Walker AP, Wiltshire AJ, Zaehle S (2016) Global carbon budget 2016. Earth Syst Data 8(2):605–649. doi:10.5194/essd-8-605-2016
Lefsky MA (2010) A global forest canopy height map from the moderate resolution imaging spectroradiometer and the geoscience laser altimeter system. Geophys Res Lett 37. doi:10.1029/2010gl043622
Lewis SL, Lloyd J, Sitch S, Mitchard ETA, Laurance WF (2009a) Changing ecology of tropical forests: evidence and drivers. Annu Rev Ecol Evol Syst 40:529–549. doi:10.1146/annurev.ecolsys.39.110707.173345
Lewis SL, Lopez-Gonzalez G, Sonke B, Affum-Baffoe K, Baker TR, Ojo LO, Phillips OL, Reitsma JM, White L, Comiskey JA, Djuikouo KM-N, Ewango CEN, Feldpausch TR, Hamilton AC, Gloor M, Hart T, Hladik A, Lloyd J, Lovett JC, Makana J-R, Malhi Y, Mbago FM, Ndangalasi HJ, Peacock J, Peh KSH, Sheil D, Sunderland T, Swaine MD, Taplin J, Taylor D, Thomas SC, Votere R, Woell H (2009b) Increasing carbon storage in intact African tropical forests. Nature 457(7232):1003–1006. doi:10.1038/nature07771
Li Q, Chen J, Moorhead DL (2012) Respiratory carbon losses in a managed oak forest ecosystem. For Ecol Manag 279:1–10. doi:10.1016/j.foreco.2012.05.011
Litvak M, Miller S, Wofsy SC, Goulden M (2003) Effect of stand age on whole ecosystem CO2 exchange in the Canadian boreal forest. J Geophys Res Atmos 108(D3). doi:10.1029/2001jd000854
Luedeling E, Neufeldt H (2012) Carbon sequestration potential of parkland agroforestry in the Sahel. Clim Change 115(3–4):443–461. doi:10.1007/s10584-012-0438-0
Luethi D, Le Floch M, Bereiter B, Blunier T, Barnola J-M, Siegenthaler U, Raynaud D, Jouzel J, Fischer H, Kawamura K, Stocker TF (2008) High-resolution carbon dioxide concentration record 650,000–800,000 years before present. Nature 453(7193):379–382. doi:10.1038/nature06949
Lundgren BO, Rainee JB (1982) Sustained agroforestry. In: Nestel B (ed) Agricultural research for development: potentials and challenges in Asia. ISNER, Tha Hahue, Netherlands, pp 37–49
Luyssaert S, Inglima I, Jung M, Richardson AD, Reichstein M, Papale D, Piao SL, Schulzes ED, Wingate L, Matteucci G, Aragao L, Aubinet M, Beers C, Bernhofer C, Black KG, Bonal D, Bonnefond JM, Chambers J, Ciais P, Cook B, Davis KJ, Dolman AJ, Gielen B, Goulden M, Grace J, Granier A, Grelle A, Griffis T, Gruenwald T, Guidolotti G, Hanson PJ, Harding R, Hollinger DY, Hutyra LR, Kolar P, Kruijt B, Kutsch W, Lagergren F, Laurila T, Law BE, Le Maire G, Lindroth A, Loustau D, Malhi Y, Mateus J, Migliavacca M, Misson L, Montagnani L, Moncrieff J, Moors E, Munger JW, Nikinmaa E, Ollinger SV, Pita G, Rebmann C, Roupsard O, Saigusa N, Sanz MJ, Seufert G, Sierra C, Smith ML, Tang J, Valentini R, Vesala T, Janssens IA (2007) CO2 balance of boreal, temperate, and tropical forests derived from a global database. Glob Change Biol 13(12):2509–2537. doi:10.1111/j.1365-2486.2007.01439.x
Luyssaert S, Schulze ED, Boerner A, Knohl A, Hessenmoeller D, Law BE, Ciais P, Grace J (2008) Old-growth forests as global carbon sinks. Nature 455(7210):213–215. doi:10.1038/nature07276
Luyssaert S, Ciais P, Piao SL, Schulze ED, Jung M, Zaehle S, Schelhaas MJ, Reichstein M, Churkina G, Papale D, Abril G, Beer C, Grace J, Loustau D, Matteucci G, Magnani F, Nabuurs GJ, Verbeeck H, Sulkava M, van der Werf GR, Janssens IA, Team C-IS (2010) The European carbon balance. Part 3: forests. Glob Change Biol 16(5):1429–1450. doi:10.1111/j.1365-2486.2009.02056.x
Luyssaert S, Jammet M, Stoy PC, Estel S, Pongratz J, Ceschia E, Churkina G, Don A, Erb K, Ferlicoq M, Gielen B, Gruenwald T, Houghton RA, Klumpp K, Knohl A, Kolb T, Kuemmerle T, Laurila T, Lohila A, Loustau D, McGrath MJ, Meyfroidt P, Moors EJ, Naudts K, Novick K, Otto J, Pilegaard K, Pio CA, Rambal S, Rebmann C, Ryder J, Suyker AE, Varlagin A, Wattenbach M, Dolman AJ (2014) Land management and land-cover change have impacts of similar magnitude on surface temperature. Nat Clim Change 4(5):389–393. doi:10.1038/nclimate2196
Malhi Y, Baldocchi DD, Jarvis PG (1999) The carbon balance of tropical, temperate and boreal forests. Plant Cell Environ 22(6):715–740. doi:10.1046/j.1365-3040.1999.00453.x
Masek JG, Hayes DJ, Hughes MJ, Healey SP, Turner DP (2015) The role of remote sensing in process-scaling studies of managed forest ecosystems. For Ecol Manag 355:109–123. doi:10.1016/j.foreco.2015.05.032
Masera OR, Garza-Caligaris JF, Kanninen M, Karjalainen T, Liski J, Nabuurs GJ, Pussinen A, de Jong BHJ, Mohren GMJ (2003) Modeling carbon sequestration in afforestation, agroforestry and forest management projects: the CO2FIX V.2 approach. Ecol Model 164(2–3):177–199. doi:10.1016/s0304-3800(02)00419-2
McKinley DC, Ryan MG, Birdsey RA, Giardina CP, Harmon ME, Heath LS, Houghton RA, Jackson RB, Morrison JF, Murray BC, Pataki DE, Skog KE (2011) A synthesis of current knowledge on forests and carbon storage in the United States. Ecol Appl 21(6):1902–1924. doi:10.1890/10-0697.1
McMillan AMS, Goulden ML (2008) Age-dependent variation in the biophysical properties of boreal forests. Glob Biogeochem Cycles 22(2). doi:10.1029/2007gb003038
Miura S, Amacher M, Hofer T, San-Miguel-Ayanz J, Ernawati-Thackway R (2015) Protective functions and ecosystem services of global forests in the past quarter-century. For Ecol Manag 352:35–46. doi:10.1016/j.foreco.2015.03.039
Morales-Hidalgo D, Oswalt SN, Somanathan E (2015) Status and trends in global primary forest, protected areas, and areas designated for conservation of biodiversity from the global forest resources assessment 2015. For Ecol Manag 352:68–77. doi:10.1016/j.foreco.2015.06.011
Nabuurs GJ, Masera O, Andrasko K, Benitez-Ponce P, Boer R, Dutschke M, Elsiddig E, Ford-Robertson J, Frumhoff P, Karjalainen T, Krankina O, Kurz WA, Matsumoto M, Oyhantcabal W, Ravindranath NH, Sanchez MJS, Zhang X (2007) Forestry. In: Metz B, Davidson OR, Bosch PR, Dave R, Meye LA (eds) Climate change 2007: mitigation. Contribution of working group III to the fourth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, UK, and New York, USA, pp 541–584
Ni X, Zhou Y, Cao C, Wang X, Shi Y, Park T, Choi S, Myneni RB (2015) Mapping forest canopy height over continental china using multi-source remote sensing data. Remote Sens 7(7):8436–8452. doi:10.3390/rs70708436
Page SE, Siegert F, Rieley JO, Boehm HDV, Jaya A, Limin S (2002) The amount of carbon released from peat and forest fires in Indonesia during 1997. Nature 420(6911):61–65. doi:10.1038/nature01131
Pan Y, Birdsey RA, Fang J, Houghton R, Kauppi PE, Kurz WA, Phillips OL, Shvidenko A, Lewis SL, Canadell JG, Ciais P, Jackson RB, Pacala SW, McGuire AD, Piao S, Rautiainen A, Sitch S, Hayes D (2011) A large and persistent carbon sink in the world’s forests. Science 333(6045):988–993. doi:10.1126/science.1201609
Pan Y, Birdsey RA, Phillips OL, Jackson RB (2013) The structure, distribution, and biomass of the world’s forests. Annu Rev Ecol Evol Syst 44:593–622. doi:10.1146/annurev-ecolsys-110512-135914
Pearson TR, Brown SL, Birdsey RA (2007) Measurement guidelines for the sequestration of forest carbon. United States Department of Agriculture (USDA) Forest Service, Northern Research Station, PA, USA, p 47
Peltoniemi M, Makipaa R, Liski J, Tamminen P (2004) Changes in soil carbon with stand age—an evaluation of a modelling method with empirical data. Glob Change Biol 10(12):2078–2091. doi:10.1111/j.1365-2486.2004.00881.x
Petrescu AMR, Abad-Vinas R, Janssens-Maenhout G, Blujdea VNB, Grassi G (2012) Global estimates of carbon stock changes in living forest biomass: EDGARv4.3-time series from 1990 to 2010. BioGeosciences 9(8):3437–3447. doi:10.5194/bg-9-3437-2012
Philips O, Lewis SL (2014) Recent changes in tropical forest biomass and dynamics. In: Coomes DA, Burslem DFR, Simonson WD (eds) Forests and global change. Cambridge University Press, Cambridge and New York, pp 77–108
Phillips OL, Malhi Y, Higuchi N, Laurance WF, Nunez PV, Vasquez RM, Laurance SG, Ferreira LV, Stern M, Brown S, Grace J (1998) Changes in the carbon balance of tropical forests: evidence from long-term plots. Science 282(5388):439–442. doi:10.1126/science.282.5388.439
Phillips DL, Brown SL, Schroeder PE, Birdsey RA (2000) Toward error analysis of large-scale forest carbon budgets. Glob Ecol Biogeogr 9(4):305–313. doi:10.1046/j.1365-2699.2000.00197.x
Phillips OL, Aragao LEOC, Lewis SL, Fisher JB, Lloyd J, Lopez-Gonzalez G, Malhi Y, Monteagudo A, Peacock J, Quesada CA, van der Heijden G, Almeida S, Amaral I, Arroyo L, Aymard G, Baker TR, Banki O, Blanc L, Bonal D, Brando P, Chave J, Alves de Oliveira AC, Cardozo ND, Czimczik CI, Feldpausch TR, Freitas MA, Gloor E, Higuchi N, Jimenez E, Lloyd G, Meir P, Mendoza C, Morel A, Neill DA, Nepstad D, Patino S, Cristina Penuela M, Prieto A, Ramirez F, Schwarz M, Silva J, Silveira M, Thomas AS, ter Steege H, Stropp J, Vasquez R, Zelazowski P, Alvarez Davila E, Andelman S, Andrade A, Chao K-J, Erwin T, Di Fiore A, Honorio CE, Keeling H, Killeen TJ, Laurance WF, Pena Cruz A, Pitman NCA, Nunez Vargas P, Ramirez-Angulo H, Rudas A, Salamao R, Silva N, Terborgh J, Torres-Lezama A (2009) Drought sensitivity of the Amazon rainforest. Science 323(5919):1344–1347. doi:10.1126/science.1164033
Piao S, Yin G, Tan J, Cheng L, Huang M, Li Y, Liu R, Mao J, Myneni RB, Peng S, Poulter B, Shi X, Xiao Z, Zeng N, Zeng Z, Wang Y (2015) Detection and attribution of vegetation greening trend in China over the last 30 years. Glob Change Biol 21(4):1601–1609. doi:10.1111/gcb.12795
Poeplau C, Don A, Vesterdal L, Leifeld J, Van Wesemael B, Schumacher J, Gensior A (2011) Temporal dynamics of soil organic carbon after land-use change in the temperate zone—carbon response functions as a model approach. Glob Change Biol 17(7):2415–2427. doi:10.1111/j.1365-2486.2011.02408.x
Potapov PV, Zhuravleva IV, Manisha AE, Turubanova SA, Yaroshenko AY (2008) Identification and monitoring of world intact landscapes using remote sensing methods. Lesovedenie 2:58–67
Pregitzer KS, Euskirchen ES (2004) Carbon cycling and storage in world forests: biome patterns related to forest age. Glob Change Biol 10(12):2052–2077. doi:10.1111/j.1365-2486.2004.00866.x
Prentice IC, Harrison SP, Bartlein PJ (2011) Global vegetation and terrestrial carbon cycle changes after the last ice age. New Phytol 189(4):988–998. doi:10.1111/j.1469-8137.2010.03620.x
Ramankutty N, Gibbs HK, Achard F, Defriess R, Foley JA, Houghton RA (2007) Challenges to estimating carbon emissions from tropical deforestation. Glob Change Biol 13(1):51–66. doi:10.1111/j.1365-2486.2006.01272.x
Reyer C, Guericke M, Ibisch PL (2009) Climate change mitigation via afforestation, reforestation and deforestation avoidance: and what about adaptation to environmental change? New Forest 38(1):15–34. doi:10.1007/s11056-008-9129-0
Rossi E, Rogan J, Schneider L (2013) Mapping forest damage in northern Nicaragua after Hurricane Felix (2007) using MODIS enhanced vegetation index data. GISSci Remote Sens 50(4):385–399. doi:10.1080/15481603.2013.820066
Running SW, Coughlan JC (1988) A general model of forest ecosystem processes for regional applications I. Hydrologic balance, canopy gas exchange and primary production processes. Ecol Model 42(2):125–154. doi:10.1016/0304-3800(88)90112-3
Running SW, Nemani RR, Heinsch FA, Zhao MS, Reeves M, Hashimoto H (2004) A continuous satellite-derived measure of global terrestrial primary production. Bioscience 54(6):547–560. doi:10.1641/0006-3568(2004)054[0547:acsmog]2.0.co;2
Running SW, Nemani RR, Townshend JRG, Baldocchi DD (2009) Next-generation terrestrial carbon monitoring. In: McPherson BJ, Sundquist ET (eds) Carbon sequestration and its role in the global carbon cycle. Geophysical monograph series, vol 183. American Geophysical Union, Washington, DC, pp 49–69. doi:10.1029/2006GM000526
Ryan MG, Binkley D, Fownes JH (1997) Age-related decline in forest productivity: pattern and process. Adv Ecol Res 27:213–262. doi:10.1016/s0065-2504(08)60009-4
Saatchi SS, Harris NL, Brown S, Lefsky M, Mitchard ETA, Salas W, Zutta BR, Buermann W, Lewis SL, Hagen S, Petrova S, White L, Silman M, Morel A (2011) Benchmark map of forest carbon stocks in tropical regions across three continents. Proc Natl Acad Sci USA 108(24):9899–9904. doi:10.1073/pnas.1019576108
Schlamadinger B, Marland G (1996) The role of forest and bioenergy strategies in the global carbon cycle. Biomass Bioenerg 10(5–6):275–300. doi:10.1016/0961-9534(95)00113-1
Schlamadinger B, Apps M, Bohlin F, Gustavsson L, Jungmeier G, Marland G, Pingoud K, Savolainen I (1997) Towards a standard methodology for greenhouse gas balances of bioenergy systems in comparison with fossil energy systems. Biomass Bioenerg 13(6):359–375. doi:10.1016/s0961-9534(97)10032-0
Settele J, Scholes R, Betts R, Bunn S, Leadley P, Stonestrom DA, Nepstad, Overpeck JT, Taboada MA (2014) Terrestrial and inland water systems. In: Field CB, Barros VR, Dokken DJ et al (eds) Climate change 2014: impacts, adaptation, and vulnerability. Part A: global and sectoral aspects. Contribution of working group II to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, pp 271–359
Shugart HH, Saatchi S, Hall FG (2010) Importance of structure and its measurement in quantifying function of forest ecosystems. J Geophys Res Biogeosci 115. doi:10.1029/2009jg000993
Siegenthaler U, Stocker TF, Monnin E, Luthi D, Schwander J, Stauffer B, Raynaud D, Barnola JM, Fischer H, Masson-Delmotte V, Jouzel J (2005) Stable carbon cycle-climate relationship during the late Pleistocene. Science 310(5752):1313–1317. doi:10.1126/science.1120130
Sinare H, Gordon LJ (2015) Ecosystem services from woody vegetation on agricultural lands in Sudano-Sahelian West Africa. Agric Environ 200:186–199. doi:10.1016/j.agee.2014.11.009
Sitch S, Friedlingstein P, Gruber N, Jones SD, Murray-Tortarolo G, Ahlstrom A, Doney SC, Graven H, Heinze C, Huntingford C, Levis S, Levy PE, Lomas M, Poulter B, Viovy N, Zaehle S, Zeng N, Arneth A, Bonan G, Bopp L, Canadell JG, Chevallier F, Ciais P, Ellis R, Gloor M, Peylin P, Piao SL, Le Quéré C, Smith B, Zhu Z, Myneni R (2015) Recent trends and drivers of regional sources and sinks of carbon dioxide. BioGeosciences 12(3):653–679. doi:10.5194/bg-12-653-2015
Smith P, Bustamante M, Ahammad H, Clark H, Dong H, Elsiddig EA, Haberl H, Harper R, House J, Jafari M, Masera O, Mbow C, Ravindranath NH, Rice CW, Abad CR, Romanovskaya A, Sperling F, Tubiello F (2014) Agriculture, forestry and other land use (AFOLU). In: Edenhofer O, Pichs-Madruga R, Sokona Y et al (eds) Climate change 2014: mitigation of climate change. Contribution of working group III to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, pp 811–922
Sochacki SJ, Harper RJ, Smettem KRJ (2012) Bio-mitigation of carbon following afforestation of abandoned salinized farmland. GCB Bioenergy 4(2):193–201. doi:10.1111/j.1757-1707.2011.01139.x
Solberg S, Dobbertin M, Reinds GJ, Lange H, Andreassen K, Fernandez PG, Hildingsson A, de Vries W (2009) Analyses of the impact of changes in atmospheric deposition and climate on forest growth in European monitoring plots: a stand growth approach. For Ecol Manag 258(8):1735–1750. doi:10.1016/j.foreco.2008.09.057
Song CH, Woodcock CE (2003) A regional forest ecosystem carbon budget model: impacts of forest age structure and landuse history. Ecol Model 164(1):33–47. doi:10.1016/s0304-3800(03)00013-9
Swanson ME, Franklin JF, Beschta RL, Crisafulli CM, DellaSala DA, Hutto RL, Lindenmayer DB, Swanson FJ (2011) The forgotten stage of forest succession: early-successional ecosystems on forest sites. Front Ecol Environ 9(2):117–125. doi:10.1890/090157
Thorburn C (2013) Seeing the forest for the carbon: interrogating reduced emissions from deforestation and degradation (REDD). In: Thorburn C (ed) Critical reflections on development. Springer, pp 139–161
Thornley JHM, Cannell MGR (2004) Long-term effects of fire frequency on carbon storage and productivity of boreal forests: a modeling study. Tree Physiol 24(7):765–773
Thornton PE, Law BE, Gholz HL, Clark KL, Falge E, Ellsworth DS, Golstein AH, Monson RK, Hollinger D, Falk M, Chen J, Sparks JP (2002) Modeling and measuring the effects of disturbance history and climate on carbon and water budgets in evergreen needleleaf forests. Agric For Meteorol 113(1–4):185–222. doi:10.1016/s0168-1923(02)00108-9
Tran TV, de Beurs KM, Julian JP (2016) Monitoring forest disturbances in Southeast Oklahoma using landsat and MODIS images. Int J Appl Earth Obs Geoinf 44:42–52. doi:10.1016/j.jag.2015.07.001
UNFCCC (2007) Report of the conference of the parties on its thirteenth session, held in Bali from 3 to 15 December 2007. United Nations Framework Convention on Climate Change (UNFCCC), p 60
UN-REDD (2010) The UN-REDD programme strategy 2011–2015. United Nations, New York, p 22
van Bodegom JA, Savinije H, Wit M (eds) (2009) Forests and climate change: adaptation and mitigation. In: European tropical forest research network, Issue 50. Tropenbos International, Wagenigen, The Netherlads
van der Werf GR, Morton DC, DeFries RS, Olivier JGJ, Kasibhatla PS, Jackson RB, Collatz GJ, Randerson JT (2009) CO2 emissions from forest loss. Nat Geosci 2(11):737–738. doi:10.1038/ngeo671
Walther G-R (2010) Community and ecosystem responses to recent climate change. Philos T R Soc B 365(1549):2019–2024. doi:10.1098/rstb.2010.0021
Wang F, D’Sa EJ (2010) Potential of MODIS EVI in identifying hurricane disturbance to coastal vegetation in the Northern Gulf of Mexico. Remote Sens 2(1):1–18. doi:10.3390/rs2010001
Wang S, Zhou L, Chen J, Ju W, Feng X, Wu W (2011) Relationships between net primary productivity and stand age for several forest types and their influence on China’s carbon balance. J Environ Manag 92(6):1651–1662. doi:10.1016/j.jenvman.2011.01.024
Waring RH, Schlesinger WH (1985) Forest ecosystems: concepts and management. Academic Press, New York
Wei X, Shao M, Gale W, Li L (2014) Global pattern of soil carbon losses due to the conversion of forests to agricultural land. Sci Rep 4:4062–4066. doi:10.1038/srep04062 http://www.nature.com/articles/srep04062#supplementary-information
Westerling AL, Hidalgo HG, Cayan DR, Swetnam TW (2006) Warming and earlier spring increase western US forest wildfire activity. Science 313(5789):940–943. doi:10.1126/science.1128834
Woodall CW, Domke GM, MacFarlane DW, Oswalt CM (2012) Comparing field- and model-based standing dead tree carbon stock estimates across forests of the US. Forestry 85(1):125–133. doi:10.1093/forestry/cpr065
Wulder MA, White JC, Fournier RA, Luther JE, Magnussen S (2008) Spatially explicit large area biomass estimation: three approaches using forest inventory and remotely sensed imagery in a GIS. Sensors 8(1):529–560. doi:10.3390/s8010529
Yu G, Chen Z, Piao S, Peng C, Ciais P, Wang Q, Li X, Zhu X (2014) High carbon dioxide uptake by subtropical forest ecosystems in the East Asian monsoon region. Proc Natl Acad Sci USA 111(13):4910–4915. doi:10.1073/pnas.1317065111
Zianis D, Muukkonen P, Makipaa R, Mencuccini M (2005) Biomass and stem volume equations for tree species in Europe. Silva Fenn Monogr (4):1–2, 5–63
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Ussiri, D.A., Lal, R. (2017). Global Forests Management for Climate Change Mitigation. In: Carbon Sequestration for Climate Change Mitigation and Adaptation. Springer, Cham. https://doi.org/10.1007/978-3-319-53845-7_11
Download citation
DOI: https://doi.org/10.1007/978-3-319-53845-7_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-53843-3
Online ISBN: 978-3-319-53845-7
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)