
Abstract
Myxozoans evoke important economic losses in aquaculture production, but there is almost a total lack of disease control methods as no vaccines or commercial treatments are currently available. Knowledge of the immune responses that lead to myxozoan elimination and subsequent disease resistance is vital for shaping the future development of disease control measures. Different fish immune factors triggered by myxozoan parasites are reviewed in this chapter. Detailed information on the phenotypic and underlying molecular aspects of innate and adaptive responses, at both cellular and humoral levels, is provided for some well-studied fish-myxozoan systems. The importance of the local immune response, mainly at mucosal sites, is also highlighted. Myxozoan tactics to disable or avoid immune responses, such as modulation of immune gene transcription and immune evasion, are also reviewed. The existence of innate and acquired resistance to some myxozoan species suggest promising possibilities for controlling myxozooses through immune-based strategies, such as genetic selection for host resistance, vaccination, immune therapies and administration of immunostimulants.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Immune evasion
- Host resistance
- Innate response
- Adaptive response
- Transcriptional modulation
- Histopathology
1 Introduction
It is generally agreed that the teleost and mammalian immune systems share a similar repertoire of molecules and cells involved in innate and adaptive immune responses. In recent years, our knowledge of the immune system of fish (Zhang et al. 2010a; Fillatreau et al. 2013; Fischer et al. 2013) and immune responses against some parasites (e.g. flagellates (Woo and Ardelli 2014) and microsporidia (Rodríguez-Tovar et al. 2011)) has greatly increased. Nevertheless, information is only available for particular fish species and many questions regarding parasite infections remain. This chapter reviews the immune responses of fish, the secondary hosts in the complex life cycle of myxozoans (see Chaps. 2, 3, and 10), by focussing on innate and adaptive immune processes evoked by myxozoan infection. We also review how myxozoans may deal with immune responses, including modulation of immune gene transcription and immune evasion.
The different immune factors of fish and their interactions with parasite stages are schematically presented in Fig. 14.1. When infectious stages of myxozoans contact fish mucosal surfaces, they may be rejected by immune molecules present in fish mucous. If not eliminated, myxozoan stages will encounter a formidable array of cell-mediated and humoral immune responses that arise in the hosts’ mucosal or epithelial tissues. These must, in some way, be subverted by myxozoan parasites in order to reach target tissues. After passing through mucosal or epithelial barriers, parasitic stages reach target tissues, including immunoprivileged sites (see Sect. 14.6.1), via the blood where they display different immune evasion strategies. Some myxozoans are eliminated in the blood owing to the expression of an array of cellular and humoral immune factors. Once in the target host tissues, myxozoan parasites evoke the activation of host immune mechanisms that are regulated by the interplay of both immunoactivating and immunosuppressive cytokine molecules. Acute or chronic disease pathology then develops, depending on the host-myxozoan model. Eventually myxozoan parasites may be encapsulated or finally cleared from the host. Some parasites, however, may survive within granulomatous tissue, leading to chronic disease pathology and even death of the host with the subsequent release of the parasite to the environment.

Diagrammatic representation of parasite strategies and host immune factors activated in myxozoan infections. Infective stages of myxozoans (from actinospores or myxosporean proliferative stages) get in contact with the host through mucosal surfaces at the gills, skin and the gastrointestinal tract. At this first contact, the parasite can be rejected by the many molecules present in the mucous, which is made of a mesh of mucins in which commensal microbiota, lysozyme, lectins, and immunoglobulins can interact. If the parasite survives this first barrier, it encounters different cell types (macrophages, granulocytes including mast cells, B cells, T cells) and immunoglobulins at the subsequent mucosal layer (composed of epithelium and dermis/lamina-propria-submucosa). After passing the mucosal barrier, parasitic stages can proliferate and travel through the blood to the final target tissue or arrive at immunoproviledged sites such as eyes, gonads or brain, or, otherwise, display different immune evasion strategies such as intracellular disguise (a), antigen masking (b) or fast proliferation (c). Some myxozoans can be eliminated during their blood passage thanks to different cellular (granulocytes/mast cells) and humoral factors (lysozyme, complement, antiproteases, immunoglobulins), whereas others survive. Once in the target tissues, the host engages again different cellular factors and Th1- Th2, Th17 and Treg cytokines play an important role in activating or suppressing them and the molecules they produce. This interplay induces acute and chronic inflammation that, depending on the host-myxozoan model, may end up with encapsulation and final clearance of the parasite. However, some parasites may survive within the granuloma or the inflammation can lead to immunopathological consequences and even death of the host, with the subsequent release of the parasite to the environment
2 Innate Immune Response and Histopathology
The innate or non-specific immune response is the first line of defence of fish against parasites and is based on cellular and humoral factors. Many myxozoan species cause little or no host cellular response, especially coelozoic forms (Lom and Dyková 1992). In some cases, the absence of a cellular reaction is due to development in immunoprivileged sites of the organism (see Sect. 14.6). On the other hand, some myxozoans trigger an excessive immune response leading to an immunopathological condition. Below we review insights about innate immune responses and the associated histopathology based on several well-studied fish-myxozoan systems.
2.1 Formation of Granulomata and Melanomacrophage Centres
The most common histopathological reaction to myxozoan infections is the formation of granulomata , which encapsulate parasitic stages by connective and epithelioid tissue layers, with the aim of isolating the parasite and preventing its dispersal to surrounding tissues (Fig. 14.2f). This occurs in Ceratomyxa shasta-resistant hosts, in which parasites captured in granulomatous lesions are lysed (Hallett and Bartholomew 2012). The efficacy of encapsulation is often limited. For instance, in Myxobolus pendula gill infections, the capsule formed around the complex cysts does not sufficiently inhibit diffusion of oxygen and nutrients to the parasite (Koehler et al. 2004). In Sphaerospora testicularis-infected European sea bass (Dicentrarchus labrax), large granulomata reduce germinal tissue of the testis in the following spawning season (Fig. 14.2g) (Sitjà-Bobadilla and Álvarez-Pellitero 1993).

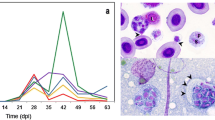
Photomicrographs showing a variety of immune responses in fish tissues infected by different myxozoan parasites. a, b Rainbow trout kidney infected by Tetracapsuloides bryosalmonae, showing the typical granulomatous reaction; arrows in a point to parasite stages and arrows in b points to a thrombus in a vessel. c Rainbow trout intestine showing a massive leucocyte proliferation in the submucosa (arrowed line with *) due to Ceratomyxa shasta. d Granulomatous reaction in the rainbow trout cartilage infected by Myxobolus cerebralis, showing infiltration with mainly macrophages (*) and partly degenerated myxozoan parasites (arrows). e Hyperplasic submucosa (arrowed line with *) of gilthead sea bream infected by Enteromyxum leei; arrow points to the parasites located in the epithelium. f Fibrotic encapsulation of Sphaerospora (formerly Polysporoplasma) sparis in the glomerulus of gilthead sea bream, spores appear necrotic (arrow). g Fibrotic encapsulation of Sphaerospora testicularis in the tesis of European sea bass; the parasite is completely destroyed (*). h Accumulation of melanomacrophage centres (MMC) around a S. sparis-infected glomerulus in the trunk kidney of gilthead sea bream. Spores are still visible in the glomerulus (arrow). i Spleen of gilthead sea bream exposed to E. leei showing a high number of MMC (dark spots). j MMC interspersed in the kidney of a Myxidum giardi-infected eel (arrows: myxospores). k Macrophages engulfing Kudoa thrysites spores (arrows) free in the muscle of Atlantic salmon. l Engulfment of a Ceratomyxa sparusaurati trophozoite (arrow) by a gilthead sea bream head kidney macrophage after 24 h of in vitro incubation. m Transmission electron micrograph of a European sea bass macrophage with an engulfed and partially destroyed spore (*) of Sphaerospora dicentrarchi. Stainings: Hematoxylin and eosin (a–e, i, j), toluidine blue (f–h, k), May-Grunwäld Giemsa (i). Scale bars: 100 µm (i), 50 µm (d) 25 µm (a, b), 20 µm (j, h), 10 µm (f, g, k, l), 0.35 µm (m). Images courtesy of: Dr. J. Bartholomew (Oregon State University, USA) (c), Dr. K. Molnár (Hungarian Academy of Sciences) (j). Images original from the authors: A S-B (e–i, k–m), H S-P (a, b, d)
The principal leucocyte types involved in granuloma formation are macrophages and lymphocytes. Other cell types can also be involved, including neutrophils, eosinophilic granule cells, multinucleated giant cells and fibroblasts. Granulomata are often accompanied by focal development of melanomacrophage centres (MMC) (Fig. 14.2h) (e.g. in lymphohaemopoietic organs of the head kidney and spleen). An increase in the number of MMCs was reported in the spleen of Enteromyxum leei-infected gilthead sea bream (Sparus aurata) (henceforth referred to as GSB) (Fleurance et al. 2008) and Enteromyxum scophthalmi-infected turbot (Sitjà-Bobadilla et al. 2006; Ronza et al. 2013). However, a significant increase in the percentage of splenic surface occupied by MMCs was observed only in GSB exposed to but not infected by E. leei (Fig. 14.2i) (Estensoro et al. 2014). These increases are associated with the inflammatory response onset and in some cases with the presence of engulfed parasites in MMCs. It is possible that rapid MMC proliferation in the spleen may help to combat the parasite. MMCs have been suggested to be the primitive analogues of the germinal centres of lymph nodes in mammals (Tort et al. 2003; Vigliano et al. 2006). Their role in the fish immune response involves antigen retention and processing and thus in the maintainience of humoral memory (Agius and Roberts 2003). Macrophages are not only involved with forming MMCs, but also engulf parasites directly (Fig. 14.2j–m).
2.2 Other Cellular Responses
In Tetracapsuloides bryosalmonae-infected rainbow trout (Oncorhynchus mykiss), the proliferation of leucocytes (mainly macrophages and lymphocytes) invokes a more diffuse granulomatous-like reaction (Fig. 14.2a, b), leading to renal tissue hyperplasia (Proliferative Kidney Disease, PKD) (Chilmonczyk et al. 2002; Schmidt-Posthaus et al. 2012). Similarly, in C. shasta-infected salmonids, a vigorous host response consisting of lymphocytes and eosinophilic granular leucocytes is mounted (Fig. 14.2c) (Bartholomew et al. 1989). The pronounced inflammatory response of channel catfish (Ictalurus punctatus) against Henneguya ictaluri in Proliferative Gill Disease contributes to the breakdown of the gill filament cartilage (Lovy et al. 2011). The chronic cartilaginous inflammation induced by Myxobolus cerebralis trophozoites in susceptible salmonids is responsible for cartilage degeneration (Fig. 14.2d) (MacConnell and Vicent 2002). A massive hyperplasia of the submucosa occurs in the intestine of E. leei-infected sparids due to the proliferation of heterogeneous leucocytes (Fig. 14.2e). Some authors have pointed out that such severe inflammatory reactions are found in unusual hosts for such myxozoans (Kent and Hedrick 1985). Furthermore, reactions against a myxozoan, even within a given host, can vary considerably depending on the infected tissue. For example, Parvicapsula minibicornis induces inflammatory cells to infiltrate the gills of sockeye salmon (Oncorhynchus nerka), whereas no cellular host response is noticed in the mesonephros around or within infected glomeruli (Bradford et al. 2010).
The types of leucocytes involved in local responses against myxozoans vary depending on the myxozoan-fish model and site of infection. For example, when T. bryosalmonae first enters the fish gills a granulomatous reaction, dominated by infiltration with macrophages, is induced in the filaments (Schmidt-Posthaus et al. 2013). Once in the kidney, macrophages surround the parasites in the vessels and proliferating lymphocytes account for the lymphoid hyperplasia and gross kidney swelling response that is characteristic of PKD. Finally, granulomatous-like reactions are established in the renal interstitium during the advanced stages of this disease (Hedrick et al. 1993; Bettge et al. 2009).
In general, phagocytes and granulocytes are the most abundant immune cells in mucosal infections (skin, gills and intestine). Neutrophils and other granulocytes (mast cells and eosinophil-like cells) dominate in the gill inflammatory infiltrate found in Proliferative Gill Disease (Lovy et al. 2011). Polymorphonuclear leucocytes, fibroblasts and macrophages are abundant in C. shasta-infected intestines (Bartholomew et al. 1989). A succession of granulocytes and lymphocytes is described in the oedemata surrounding M. pendula branchial cysts in creek chub (Semotilus atromaculatus) (Martyn et al. 2002). Some granulocyte subsets increase during enteromyxosis in sharpsnout sea bream (Diplodus puntazzo) (Fig. 14.3a) (Álvarez-Pellitero et al. 2008), whereas acidophils decrease at both local and systemic levels in GSB (Estensoro et al. 2014). Acidophilic granulocytes act as professional phagocytes in the latter. Considered the functional equivalents of mammalian neutrophils, acidophilic granulocytes can be rapidly recruited from head kidney and are the most abundant circulating granulocytes (Sepulcre et al. 2002; Chaves-Pozo et al. 2005; Estensoro et al. 2014). Mast cells are massively recruited to the intestine in E. leei infections (Fig. 14.3d, e) (Katharios et al. 2011; Estensoro et al. 2014), and are often associated with chronic inflammation. When activated, mast cells degranulate releasing hormonal mediators and histamine analogues in the extracellular environment, where they act as pro-inflammatory moderators of the inflammation reaction. Importantly, however, only fish belonging to the order Perciformes show typical mast cells containing histamine (Mulero et al. 2007). Large numbers of degranulated cells are also observed within the gill lesions produced by H. ictaluri (Lovy et al. 2011). Immune cell degranulation acts selectively against the parasite, but, when excessive, can have deleterious immunopathological consequences for the host, even causing death.

Photomicrographs of fish tissues infected by different myxozoan parasites. a Eosinophilic granular cells (encircled) in the intestinal epithelium of sharpsnout sea bream infected by Enteromyxum leei. Arrows: parasitic stages. b Intestine of E. leei-infected gilthead sea bream showing acidophilic granulocytes immunostained (brown cells) in the submucosa with the G7 antibody. c Detail of the G7-immunostained cells. d Intestine of E. leei-infected gilthead sea bream showing mast cells immunostained (brown cells) in the submucosae with an anti-histamine antibody. e Detail of mast cells (arrows) at the base of the epithelium. f Intestine of E. scophthalmi-infected turbot with abundant iNOs+ cells in the submucosae (brown cells). g Intestine of E. leei-infected gilthead sea bream showing scarce goblets cells (in blue, arrows); parasite stages appear fuchsia-coloured. h Intestine of Ceratomyxa shasta-infected rainbow trout showing Ig+ cells in the epithelium and the submucosae (brown cells). i Intestine of E. leei-infected gilthead sea bream showing IgM+ cells (brown cells) in the submucosae and epithelium. j Detail of IgM+ cells in the epithelium, surrounding parasite stages (*). k Intestine of E. scophthlami-infected turbot showing IgM+ cells (brown cells) in the epithelium, surrounding parasite stages (*). l IgT+ intraepithelial lymphocyte (arrow) in the intestine of a naïve rainbow trout. m Sphaerospora testicularis myxospores (*) in the lumen of a seminiferous tubule of European sea bass. n Intracellular Kudoa thyrsites spores in the muscle of Atlantic salmon. Stainings (when not immunostained): Hematoxylin and eosin (a), alcian-blue PAS (g), toluidine blue (m, n). Scale bars: 100 µm (g), 50 µm (f, n), 20 µm (b, d, e, h, j), 10 µm (a, c, k, l, m), 5 µm (i). Illustrations courtesy of: Dr. J. Bartholomew (Oregon State University, USA) (h), Dr. M.I. Quiroga (University of Santiago de Compostela, Spain) (f, k), Dr. C. Tafalla (INIA, Spain) (l). Illustrations original from the authors: A S-B (a–e, g, i, j, m, n). n was photographed from material provided by Dr. W. Marshall (Centre for Aquatic Health, Canada)
Phagocytes play important roles in the host’s fight against parasites but their activity can be modulated by myxozoan infections. Some enteromyxoses induce an increase in the respiratory burst of circulating phagocytes (Sitjà-Bobadilla et al. 2006, 2008), but a decrease in the activity of head kidney phagocytes (Cuesta et al. 2006b). Furthermore, in vitro studies demonstrated that some myxozoans induce the production of reactive oxygen species (ROS) (Muñoz et al. 2000a). Reactive nitrogen species are also produced by activated phagocytes and are used as cytotoxic effector molecules against fish pathogens. The production of reactive nitrogen species is controlled by the inducible nitric oxide synthase (iNOS) enzyme. The number of putative iNOS immunoreactive cells in the intestine and lymphohaematopoietic organs of E. scophthalmi-infected turbot was significantly higher and increased as infection progressed (Fig. 14.3e). The high presence of iNOS+ cells in the gut lamina propria in severe enteromyxosis might contribute to epithelial injury and desquamation (Losada et al. 2012). In E. leei-infected sharpsnout sea bream, serum nitric oxide values gradually increased after exposure to the parasite (Golomazou et al. 2006), whereas in GSB they were reduced or unchanged in fish fed a fish oil or vegetable diet, respectively (Estensoro et al. 2011). Species more susceptible to enteromyxosis may exhibit increased levels of nitric oxide (NO) and iNOS+ cells.
Another leucocyte type involved in innate immunity in mammals is the natural killer (NK) cell. Two types of NK cell homologues have been described in fish: nonspecific cytotoxic cells (NCC) and NK-like cells. Cell-mediated cytotoxicity is known to increase in E. leei-exposed GSB (Cuesta et al. 2006b).
Rodlet cells (RC) generally increase in infected tissues or are observed in close vicinity to myxozoans, sometimes even discharging their rods (Leino 1996; Palenzuela et al. 1999; Muñoz et al. 2000b; Gorgoglione and Sommerville 2010). RCs are, however, very scarce in sharpsnout sea bream, even when infected by E. leei (Álvarez-Pellitero et al. 2008). RCs are thought to be connected with inflammatory cells and host defence against parasites (Reite 2005), but their function is still controversial and their specific role in myxozoan infections requires clarification.
2.3 Humoral Responses
Some humoral innate factors, such as peroxidases , lysozyme or complement, are variably involved in myxozoan infections. Upon E. leei exposure, serum peroxidase increases in sharpsnout sea bream (Muñoz et al. 2007), whereas serum and leucocyte peroxidases are depleted in GSB (Cuesta et al. 2006a, b; Sitjà-Bobadilla et al. 2008). Lysozyme activity is significantly depleted in GSB naturally infected with Sphaerospora (formerly Polysporoplasma) sparis (Karagouni et al. 2005) and in E. leei-exposed GSB (Sitjà-Bobadilla et al. 2008). In contrast, in other myxozoan infections or after immunisation, lysozyme levels are increased (Muñoz et al. 2000b; Sitjà-Bobadilla et al. 2006; Foott et al. 2004).
Fish serum complement is also variably involved in myxozoan infections. Plasma complement activity of infected salmon is not altered until advanced ceratomyxosis (Foott et al. 2004). In Enteromyxum spp. infections, the activity of the complement alternative pathway is initially increased (or unaltered) in response to parasite exposure, and generally decreases at later stages of infection (Cuesta et al. 2006a; Sitjà-Bobadilla et al. 2006). Some parasite-specific glycans or surface-associated carbohydrate moieties may activate the complement system through the lectin pathway (Kaltner et al. 2007; Redondo et al. 2008; Redondo and Álvarez-Pellitero 2009; Estensoro et al. 2013a). The lowered serum in vitro activity following heat inactivation and withdrawal of bivalent metal ions (Kallert et al. 2012) suggests that complement or complement induced factors are responsible for cellular disintegration of some myxozoans (M. cerebralis, Henneguya nuesslini and Myxobolus pseudodispar). The exhaustion of complement can have immunopathological consequences, due to the accumulation of immune complexes that are not lysed by complement. For instance, the immune deposits found at the basement membrane of the glomeruli of Myxobolus-infected barbels (Barbus graellsii) seem to be the cause of membranous glomerulonephritis (Peribáñez et al. 1993).
Several myxozoan parasites are known to produce proteases. For example, the early stages of H. ictaluri release proteases that degrade collagen and support trophozoite growth within the gill filament (Lovy et al. 2011). Serine proteases of M. cerebralis digest and cause lysis of rainbow trout cartilage (Kelley et al. 2004), and Kudoa thyrsites proteases are reponsible for histolysis characteristic of the invoked disease (Langdon 1991; Funk et al. 2008). Host anti-proteases counteract the action of parasitic proteases. Examples include the increase in serum α-2 Macroglobulin (α-2 M), (one of the most versatile anti-proteases) in E. scophthalmi-parasitized turbot (Sitjà-Bobadilla et al. 2006) and of total serum antiproteases in E. leei-parasitized sharpsnout sea bream (Muñoz et al. 2007).
The mucosal surface is relevant for the outcome of the disease, not only because it is a complex organisation of epithelium, immune cells and resident microbiota (Rombout et al. 2011; Gómez et al. 2013), but also because the mucous layer may favour or impede pathogen entrance. Mucins are the main structural component of the mucous and are secreted by goblet cells, forming a mesh-like structure. Turbot (Bermúdez et al. 2009) and GSB (Fleurance et al. 2008; Estensoro et al. 2012a) with advanced enteromyxoses had decreased numbers of goblet cells. In heavily parasitized intestinal areas goblet cells were even absent in GSB. The mucous of infected GSB had higher glycosylation levels and terminal glycosylation of mucous proteins, depending on the intestinal segment affected (Estensoro et al. 2013b).
3 Adaptive Immune Response
The very first attempts to detect specific antibodies against myxozoans failed (Pauley 1974; Halliday 1976; Siau 1980; Bartholomew et al. 1989). This led to the suggestion that fish were unable to mount an adaptive or specific immune response against myxozoans. The presence of specific antibodies has, however, been unambiguously reported in fish infected by M. cerebralis (Hedrick et al. 1998), Myxobolus artus (Furuta et al. 1993), T. bryosalmonae (Saulnier and de Kinkelin 1996), C. shasta (Bartholomew 2001), E. scophthalmi (Sitjà-Bobadilla et al. 2004) and E. leei (Estensoro et al. 2010a). The speed of antibody production is relatively low in comparison with available data for fish immunized or vaccinated with other parasites. Specific antibodies were not present until 12 weeks after exposure of rainbow trout to M. cerebralis (Ryce 2003), whereas anti-T. bryosalmonae antibodies were detected as early as 6 weeks post exposure (p.e.) (Hedrick et al. 1991). Specific antibodies were detected as soon as 48 days p.e. in turbot exposed to E. scophthalmi if fish belonged to previously exposed stocks. If fish belonged to naïve stocks, they developed the disease and died without producing antibodies at 40–49 days p.e. (Sitjà-Bobadilla et al. 2006, 2007b). Further information on gene expression of immunoglobulins (Ig) is provided in Sect. 14.4.
Myxozoan infections evoke changes in numbers and distributions of B cells and plasma cells . For instance, there is a significant decrease in the number of B cells positive for IgM (the main serum immunoglobulin in fish) (IgM+ B cells) in the head kidney and spleen of turbot (Bermúdez et al. 2006), but an increase in number in the head kidney of GSB in well-established infections (Estensoro et al. 2014). Both fish species demonstrate a very high recruitment of IgM+ B cells and plasma cells at the intestine (Fig. 14.3i–k) (Bermúdez et al. 2006; Estensoro et al. 2014), particularly in the lamina propia-submucosa and epithelium, where they can be observed surrounding parasite stages. These observations challenge the paradigm that teleost intraepithelial lymphocytes (IEL) are exclusively T cells, and confirm recent observations in rainbow trout (Ballesteros et al. 2013) (Fig. 14.3l).
The speed of the recruitment of IgM+ B cells depends on the route of infection in GSB, being higher in fish infected by anal intubation than by exposure to parasite-contaminated effluent (Estensoro et al. 2012b, 2014). Interestingly, in C. shasta-infected rainbow trout a large accumulation of IgT+ cells (but not IgM+ B cells) in the lamina propria was coincident with an increase in the total amount of IgT (specialised mucosal immunoglobulin) and specific IgT antibodies in the intestinal mucous, whereas IgM could not be found at this site (Zhang et al. 2010b). These findings show once more the importance of mucosal immunity and in particular the role of a specialized IgT in fish (Salinas et al. 2011; Gómez et al. 2013).
4 Changes in Immune Gene Expression in Response to Myxozoan Infection
An important prerequisite for understanding the nature of parasite-mediated modulation of immune processes is to know the immune mechanisms and molecules that result in chronic disease or resistance to infection. The majority of immune genes examined in studies on myxozoans encode innate immune molecules. Unlike bacterial infections, myxozoan-mediated changes in gene expression have often been reported to be weak, transient or down-regulated in infected fish. Data from fish genomes and transcriptomes have tremendously increased in our understanding of fish immune genes, providing new opportunities to explore, in detail, the impact of myxozoans on fish adaptive immunity (Laing and Hansen 2011, Secombes and Wang 2012). However, it is important to keep in mind that changes in gene transcription may not necessarily reflect changes in functional proteins. Moreover, the multiple isoforms of immune molecules in some fish genomes may differ functionally (Husain et al. 2012; Hong et al. 2013; Wang et al. 2014). Nevertheless, transcriptional profiling during myxozoan infection (RNA-seq) is a powerful tool providing insights into the immune mechanisms shaping chronic disease or protective immunity. Knowledge gained by such studies may identify means of rebalancing immune processes to affect parasite clearance (Álvarez-Pellitero 2008; Sitjà-Bobadilla 2008; Gorgoglione et al. 2013). Below we review evidence for transcriptional modulation of immune genes in several well-studied myxozoan infections.
4.1 Transcriptional Modulation During T. bryosalmonae Infections
Expression of tumour necrosis factor (TNF)-α, interleukin (IL)-1β, and cyclooxygenase (COX)-2 isoforms in rainbow trout is refractory towards T. bryosalmonae infection, potentially reflecting the unresponsiveness of immune cells expressing these molecules due to a skewing of pro-inflammatory mechanisms towards an anti-inflammatory phenotype (Gorgoglione et al. 2013). Evidence for this is provided by the over-expression of putative anti-inflammatory molecules, including IL-6, IL-10a/b, IL-11, nIL-1F, (the IL-1β antagonist), and suppressors of cytokine signalling (SOCS) 1/3 (Costa et al. 2011; Wang et al. 2011; Gorgoglione et al. 2013). Importantly, the severe PKD pathology in rainbow trout (in Europe), provoked by extrasporogonic stages, does not occur to the same extent in brown trout (Salmo trutta) (Morris and Adams 2008). Fish IL-10 has been shown to suppress pro-inflammatory cytokines, phagocytosis and respiratory burst activity, all of which are refractory or suppressed by PKD (Chilmonczyk et al. 2002; Grayfer et al. 2011). The antimicrobial proteins, cathelicidin isoforms 1 and 2 and hepcidin are highly up-regulated during PKD. The former has been ascribed cytokine modulating activities in fish and so may contribute to the apparent anti-inflammatory phenotype in PKD (Bridle et al. 2011; Gorgoglione et al. 2013). A dominant anti-inflammatory phenotype could be attributed to the proliferation of lymphocytes characteristic of PKD. Although fish macrophage marker genes are refractory to or suppressed by PKD, trout macrophages remain abundant in granulomatous lesions (Hedrick et al. 1993; Gorgoglione et al. 2013). Furthermore, the up-regulation of molecules indicative of macrophage activity (e.g. IL-6, IL-12, and IL-34) suggests macrophages to be important in PKD pathogenesis (Gorgoglione et al. 2013; Wang et al. 2013, 2014).
PKD studies suggest a link between disease pathology and dysregulated over-expression of T helper (TH) cell-like genes (Gorgoglione et al. 2013). For instance, aberrant T cell activity has been linked to pathogen-mediated chronic pathology and over-expression of antibodies in mammals (Taylor et al. 2012). Unregulated TH17 and Treg responses are linked to granulomatous lesion formation and reduced vaccine responsiveness both of which are observed during PKD, thus, implicating TH17 and Treg-like activities in PKD pathogenesis (Chilmonczyk et al. 2002; Wen et al. 2011; Taylor et al. 2012). TH1-like activities in PKD are exemplified by up-regulation of the master TH1 transcription factor, T-bet, and TH1 cytokines (Wang et al. 2010). TH2 and Treg-like activities in PKD were evidenced by up-regulation of the master transcription factors, GATA3 and FOXP3 , and their associated cytokines, respectively (Gorgoglione et al. 2013). Rainbow trout homologues of mammalian TH17 cytokines were up-regulated during PKD despite the refractoriness of the master TH17 transcription factor, ROR-γ (Gorgoglione et al. 2013). Functional studies in rainbow trout have demonstrated pro-inflammatory activities of IL-17A/F2a and confirmed IL-21 as a key regulator of B and T cell-like activities upregulating IL-10, IL-22, interferon (IFN)-γ, CD4, T-bet and GATA3, whilst maintaining CD8 and IgM transcripts in head kidney leucocytes (Wang et al. 2011; Monte et al. 2013).
4.2 Transcriptional Modulation During E. leei Infections
Pro-inflammatory cytokines were down-regulated and refractory in infected and non-infected E. leei-challenged GSB, respectively (Sitjà-Bobadilla et al. 2008). Expression of α-2 M was up-regulated in the intestine of E. leei-infected GSB (Sitjà-Bobadilla et al. 2008). α-2 M silences pathogen proteases and inhibits pro-inflammatory cytokines in higher vertebrates (James 1990; Armstrong and Quigley 1999). Up-regulation of this molecule could account for the concurrent down-regulation of pro-inflammatory cytokines during enteromyxosis. cDNA microarray expression analysis of E. leei-challenged resistant and susceptible GSB signified a global down-regulation of innate immune and acute phase response genes in infected GSB relative to non-infected fish (Davey et al. 2011). Interplay between host and parasite proteases, protease inhibitors, apoptotic mechanisms, cell proliferation, and antioxidant defense genes was evident in both E. leei challenged groups. Down-regulation of innate immune or acute phase genes was associated with the complement system and normal homeostatic mechanisms in infected and non-infected fish, respectively. Mannose binding lectin (MBL)2, a known pathogen resistance marker in other fish species and an activator of the lectin-mediated complement pathway (Holmskov et al. 2003), was down-regulated in E. leei infected GSB (Davey et al. 2011). Conversely, MBL2 expression along with other genes, such as LPS-binding protein and MHC class II, could signify a global antiparasitic response.
Endocrine factors have important immune modulatory activities in fish (Yada 2007). Growth hormone receptor (GHR)-1 and insulin-like growth factor (IGF)-1 were down-regulated in the kidney of E. leei-infected fish and up-regulated in non-infected fish (Sitjà-Bobadilla et al. 2008). Other immune-related genes examined in the intestine of E. leei-infected GSB include mortalin (a heat-shock protein) and glutathione peroxidase (an anti-oxidant enzyme), the former being up-regulated and the latter down-regulated (Sitjà-Bobadilla et al. 2008). Modulation of endocrine and other immune-related genes was also observed in a recent transcriptomic study in order to assess changes associated with a vegetable oil replacement (66VO) diet compared to a fish oil diet in E. leei infected GSB (Calduch-Giner et al. 2012). Disease outcome was more pronounced in fish fed 66VO (see other effects in Sect. 14.5), consistent with the number and magnitude of differentially expressed genes. Four expression clusters reflected disease progression. Clusters 1 and 2 contained strongly and moderately up-regulated genes, respectively, with the first including a marker of alternative macrophage activation (arginase-1) and the second, genes encoding Ig chains, IL-6, and IFN-related genes. Strongly and moderately down-regulated genes were included in cluster 3 (with complement components and lectins among other immune genes) and 4 (grow factors, e.g. GHR1 and IGF-binding protein 4), respectively. The pro-inflammatory molecule, metallothionein was strongly down-regulated by infection, especially in 66VO fish, suggesting a potential role in parasite resistance.
Recent studies have linked mucin expression to E. leei susceptibility in GSB, with the molecule I-Muc being down-regulated in parasitized fish (Pérez-Sánchez et al. 2013). Such changes in mucins could be responsible for changes in bacterial adherence to the host intestinal epithelium (Estensoro et al. 2013b). Mucins are directly antiparasitic and involved in immune regulation during parasite infection with transcriptional up- and down-regulation being linked to parasite expulsion and chronic infection, respectively (Hasnain et al. 2013).
4.3 Transcriptional Modulation During M. cerebralis Infections
M. cerebralis infection of rainbow trout is apparently characterized by a transient up-regulation of pro-inflammatory genes, including IL-1β 1/2 and COX-2 (Severin and El-Matbouli 2007). Thus, in whirling disease, pro-inflammatory mechanisms may remain intact at the early stages of infection. In support of this, the anti-inflammatory molecule, transforming growth factor (TGF)-β, was initially down-regulated in susceptible fish and up-regulated at later stages. Intriguingly, a cDNA microarray study revealed the prominence of the interferon system in both TL and GR infected fish (Baerwald et al. 2008). The putative antiviral molecules ubiquitin-like protein 1, interferon regulatory factor (IRF)1, IRF7, and GCHV-induced gene 2 (Gig2) were up-regulated in both resistant and susceptible fish. Metallothionein isoform B was specifically up-regulated in parasite exposed resistant fish (Baerwald et al. 2008), whereas a trout metallothionein homologue was up-regulated during T. bryosalmonae infection (Burki et al. 2013). Thus, the potential roles of metallothioneins in myxozoan infection require further clarification.
Recent M. cerebralis studies have focused on known upstream immune regulators of metallothioneins, including IL-1β, the signal transducer and activator of transcription (STAT)3 and innate immune effector molecules (including IFN-γ, IRF1, and iNOS) (Baerwald 2013). Trout metallothionein isoforms A and B were up-regulated in parasite-challenged resistant fish, further implicating metallothionein isoforms in parasite resistance mechanisms. IL-1β exhibited a biphasic up-regulated expression profile in challenged susceptible and resistant fish, STAT3 exhibited a biphasic profile in resistant fish implicating STAT3 as an additional biomarker of M. cerebralis resistance (Baerwald 2013). IFN-γ and IRF1 were up-regulated in both susceptible and resistant trout 24 h post challenge, albeit to a greater extent in susceptible fish, whereas iNOS was only up-regulated in challenged susceptible fish. Trout homologues of the macrophage marker gene (e.g. natural resistance–associated macrophage protein (NRAMP α/β)) are implicated in pathogen killing by macrophages, phagocytosis, and regulation of pro-inflammatory genes (Lang et al. 1997). Both isoforms of NRAMP were down-regulated in susceptible relative to resistant fish, implicating these genes in M. cerebralis resistance. iNOS was confirmed to be up-regulated in M. cerebralis challenged susceptible fish, whilst arginase-2, was up-regulated in both susceptible and resistant challenged fish (Rucker and El-Matbouli 2007).
4.4 Comparing and Contrasting Transcriptional Modulation
The prominent involvement of macrophages suggests that innate immune responses play a key role in whirling disease susceptibility and resistance. In contrast, PKD pathology is linked to lymphoid tissue proliferation, specifically in relation to IgM-negative lymphocytes (Chilmonczyk et al. 2002). Both IgM and IgT transcripts were up-regulated during PKD, with IgT strongly correlating with both parasite burden and disease progression (Gorgoglione et al. 2013). IgT transcripts are also highly up-regulated (correlating with accumulation of IgT+ B cells) in the intestines of trout surviving infection with C. shasta (Zhang et al. 2010b). The same laboratory has also described the elevation of the anti-inflammatory cytokine, IL-10 in IgM+ and IgT+ B cells that could signify the presence of regulatory B cells in fish (Takizawa et al. 2013). In E. leei-infected GSB intestine, IgM up-regulation correlated closely with the accumulation of IgM+ B cells, particularly in fish fed the 66VO diet (Estensoro et al. 2012b). As indicated in Sect. 14.3, an important role of IgT in mucosal immunity has been proposed (Zhang et al. 2010b), although its role in non-mucosal immunity needs to be clarified. Interestingly, only IgM and IgT transcripts corresponding to the secretory forms were up-regulated during PKD (Gorgoglione et al. 2013). Whether the proliferating lymphocytes are IgT+ B cells or T cells is currently unknown. In C. shasta infected intestine, IgM+ and IgT+ B cells express CCR7, a chemokine receptor binding the mammalian homeostatic chemokines CCL19 and CCL21 (Ordás et al. 2012). Interestingly, upregulation of a trout CCL19-like chemokine (CK10) during PKD was closely correlated to disease progression, thus indicating a link between over-expressed homeostatic activity and lymphocyte proliferative responses (Gorgoglione et al. 2013).
The transcriptional profiling of myxozoan infections will continue to prove highly valuable in assessing the underlying immune mechanisms shaping susceptibility and resistance in the context of disease pathogenesis and parasite clearance, respectively. However, there is a growing need to develop new functional tools, in order to fully elucidate fish-myxozoan interactions and fish diseases in general.
5 Host and Environmental Factors Affecting Fish Immune Response
Environmental factors affecting the immune response of fish are well known (Bowden 2008), and myxozoan infections are typically influenced by environmental conditions (for more details see also Chap. 15). The effects of these environmental factors can involve changes in disease severity and disease.
5.1 Water Temperature
Water temperature can modulate the immune system efficiency due to the poikilothermic nature of fish. Temperatures below or above the thermal limit can influence both innate and adaptive immune responses (Ainsworth et al. 1991; Le Morvan et al. 1998; Martins et al. 2011). It is still unclear if these changes are directly driven by temperature or if they are due to changes in oxygen supply. Furthermore, it is still unclear whether temperature-dependent differences in myxozoan infections, such as those due to T. bryosalmonae (Bettge et al. 2009; Schmidt-Posthaus et al. 2013), M. cerebralis (Vincent 1998), C. shasta (Ray et al. 2012) and Enteromyxum spp. (Redondo et al. 2002; Yanagida et al. 2006; Estensoro et al. 2010b), are driven exclusively by changes in the immune response or also by effects on myxozoan proliferation.
5.2 Altitude
Increasing altitude involves changes in water temperature and ultraviolet B irradiation (UVB). UVB radiation is known to have various effects on the immune system of fish. Some authors have demonstrated reduced resistance to bacteria and parasites in irradiated fish (Markkula et al. 2007). To date no experimental data are available on the effects of UVB in myxozoan infections. Indirect evidence comes exclusively from epidemiological data. For instance, brown trout infected with T. bryosalmonae were almost exclusively below an altitude of 800 m above sea level (Wahli et al. 2008) (see also Chap. 15).
5.3 Stress
Stress , induced by different external or internal factors, is known to modulate the immune system of fish. This may occur through the interaction of corticosteroid receptors and pro-inflammatory cytokines (Stolte et al. 2008; Verbug-Van Kemenade et al. 2011). The direct effect of steroids on myxozoan infections is poorly known, but the little evidence available indicates that steroids increase fish susceptibility. High mortality of sharpsnout sea bream heavily infected with Ceratomyxa diplodae was sustained under a long term treatment with reproductive steroids (Katharios et al. 2007). Rainbow trout treated with cortisol implants had 20 times the density of T. bryosalmonae interstitial stages compared with non-treated fish, but showed less interstitial inflammation (Kent and Hedrick 1987). Exposure of T. bryosalmonae-infected rainbow trout to low or high levels of dietary estradiol had no influence on disease prevalence, parasite load or cumulative fish mortality (Burki et al. 2013).
5.4 Pollution
Pollution can affect myxozoan infections. For example, PKD prevalence increased at polluted sites in comparison to sites with lower levels of organic compounds (El-Matbouli and Hoffmann 2002). An increase in the prevalence of different myxozoans in relation to eutrophication (Marcogliese and Cone 2001), sewage effluents (Bucher and et al. 1992) or polluted waters (Cone et al. 1997) has also been reported. However, it is unclear whether these effects are due to increased invertebrate host populations, increased host stress, impaired immune response, respiratory impairment (e.g. gills irritation) or less oxygen.
5.5 Nutrition
Nutrition acts directly on the fish immune system (Oliva-Teles 2012), and the complex relationship between nutrition, immune status and parasitic infections is well recognized (Fekete and Kellems 2007). The only well documented case for the effect of host nutrition on myxozoan infections is for E. leei. Replacement of fish oils by vegetable oils in a plant protein based diet (66VO) increased disease progression in GSB (Estensoro et al. 2011) (see also Chap. 15). The main difference between the two diets tested was the fatty acid profile (the 66VO diet had lower levels of n-3 long-chain polyunsaturated fatty acids). Dietary fatty acids are capable of modulating the immune system in mammals (De Pablo and De Cienfuegos 2000) and n-3 long-chain polyunsaturated fatty acids have well recognized anti-inflammatory properties (Calder 2007). Nevertheless, other factors (such as an effect of nutrition on mucosal immunity or the integrity of the intestinal tract) may have contributed to disease progression.
5.6 Host Factors
Aspects of hosts clearly affect the disease outcome in different myxozoan infections, the most studied being host age and host size . There is a minimum size or age for the development of a specific immune response. Young fish are generally more susceptible to diseases and vaccination has no effect in very early stages (Ellis 1988). However, the situation may be more complex, additional interacting factors may intervene (see also Chap. 15). For example, brown trout (>1 year) infected with T. bryosalmonae showed lower infection prevalence and less severe pathology compared to young-of-the-year brown trout. The inflammatory responses of both age classes were, however, comparable (Schmidt-Posthaus et al. 2013).
6 Immune Evasion Strategies
Parasites have evolved a range of mechanisms to evade the immune response and host immune systems have, in turn, evolved means of counteracting them. Such coevolutionary dynamics are characteristic of host-parasite interactions. Myxozoans, like many other parasites (Sitjà-Bobadilla 2008), can evade host responses by masking their presence or developing strategies to combat them.
6.1 Exploiting Immunologically Privileged Sites
One form of immune evasion is to occupy immunologically privileged sites (IPS) within the host body, where the immune system has either highly specific or reduced surveillance capability. The most common IPS in fish are: the central nervous system (CNS), the eyes, and the testes. The immune privilege of the CNS is indispensable in view of damage limitation to a sensitive organ with poor regenerative capacity. A number of Myxobolus species show tropism for the brain and spinal cord (see Chap. 15) and others multiply exclusively in nervous tissue before reaching the target tissue for sporogony. The penetrating triactinomyxon-sporoplasms of M. cerebralis reach the cartilage via peripheral nerves and the CNS (El-Matbouli et al. 1995). It is likely that this route is chosen to avoid immune responses whilst allowing proliferation in large numbers before reaching other tissues with greater immune surveillance. Neurotropic myxobolids, such as M. neurophilus in Perca flavescens (Khoo et al. 2010) or M. hendricksoni in fathead minnows (Mitchell et al. 1985), induce minimal or no inflammation, even when large portions of the brain (optic lobes, cerebellum, ventricles and meninges) are replaced by the parasite.
Eyes constitute another IPS because portions of the eyes are isolated during embryonic development and thus are recognized as non-self (Koevary 2000). Several myxospores can be found in the eyes (either exclusively or in addition to the brain) and most of them belong to the genus Myxobolus (see Chap. 15). Extrasporogonic stages of Sphaerospora elegans have been found in the rete mirabile of the eye (Lom and Dyková 1992).
The immuneprivileged nature of testis is not only due to the blood-testis barrier, but also to differential immunological factors (Fijak and Meinhardt 2006). The main function of this IPS is to avoid any autoimmune reaction against gametes, which have undergone recombination during meiosis and are thus allogenic for the host. Damage to gametes would pose a threat to the survival of offspring. S. testicularis efficiently exploits the IPS condition of the seminiferous tubules of European sea bass, proliferating within their lumen without invoking any cellular reaction (Fig. 14.3m). Only when the testicular cysts are disrupted does the infection spread to the interstitial tissue and the immune cellular effectors appear (Sitjà-Bobadilla and Álvarez-Pellitero 1993). The route of entry of this myxosporean into the seminiferous tubules and the immune evasion strategy that enables it to reach the target site are unknown.
6.2 Intracellular and Antigen Disguises
Another evasion strategy used by myxozoans is intracellular disguise . This enables them to hide from the immune system within a host cell. Myxozoans that develop inside oocytes not only avoid the immune response, but also find a highly suitable habitat rich in nutrients. Plasmodia and spores of Kudoa ovivora proliferate within the internal oocyte layer and vitelline granules. The latter are gradually reduced until oocytes are fully occupied by parasitic stages and exceed the dimensions of healthy oocytes (Swearer and Robertson 1999). The cnidarian, Polypodium hydriforme, demonstrates a similar strategy, being an intracellular parasite of eggs of acipenseriform fish (Raikova 1994). Other Kudoa species are intracellular in myocytes (Fig. 14.3n), and do not elicit any cellular response as long as the host cell remains intact (Morado and Sparks 1986).
Antigen-based strategies include antigen mimicry or masking, antigen sequence diversity and polymorphism, and sharing and shedding of parasite antigens. Such strategies have been extensively documented for several mammalian parasites. Limited evidence also suggests that myxozoans exhibit such strategies. An example may be the modified antigen expression, especially involving the glycoprotein Mcgp33, that has been reported in different developmental stages of M. cerebralis (Knaus and El-Matbouli 2005a).
6.3 Dealing with Host Immune Responses
Some myxozoans induce immunosuppression to avoid host reactions. For instance, M. cerebralis-infected rainbow trout challenged with the bacterium Yersinia ruckeri exhibit lower survival, a more rapid onset of mortality, or both, compared to non-parasitized fish (Densmore et al. 2004). Immunosuppression can be a consequence of parasite-induced anorexia. Reduced food intake in infected hosts, which can end in cachexia, has been described for several myxozoans (Wang et al. 2005; Palenzuela 2006). Immunosuppression can also be caused by reducing the proliferative capacity of lymphocytes or the phagocytic ability of macrophages, by inducing apoptosis of host leucocytes, or by exhausting innate humoral factors. The phagocytic and/or the respiratory burst activities were decreased in GSB infected by S. (ex Polysporoplasma) sparis (Karagouni et al. 2005). Other myxozoan infections selectively depress phagocytic activity (Chilmonczyk et al. 2002), lymphocyte proliferative capacity (Densmore et al. 2004), or the number of some types of leucocytes in lymphohaematopoietic organs (Cuesta et al. 2006b; Álvarez-Pellitero et al. 2008) or in blood (Sitjà-Bobadilla et al. 2006).
Innate immune humoral factors can be significantly decreased in parasitized fish as indicated in Sect. 14.2. Transcriptional studies may provide us with important clues to the immune mechanisms exploited by myxozoan parasites to evade host immunity. The refractoriness of myxozoan resistance marker genes (such as STAT3 and metallothionein in M. cerebralis and E. leei susceptible fish) may suggest that myxozoan parasites exploit immune pathways involving these molecules to evade host immunity (Baerwald et al. 2008; Calduch-Giner et al. 2012; Baerwald 2013). Similarly, the modulation of genes encoding innate, pro-inflammatory and immunosuppressive molecules (SOCS-1 and -3) may also illustrate immune evasion strategies of myxozoans (Wang et al. 2011; Gorgoglione et al. 2013). Host SOCS molecules may have been targeted by pathogens ranging from viruses to eukaryotic parasites owing to the cytokine modulating activities of these molecules (Delgado-Ortega et al. 2013). Similarly, the suppression of MBL-2 during enteromyxosis may be indicative of parasite immune evasion, as has been demonstrated for fungal pathogens (Koneti et al. 2008; Davey et al. 2011).
7 Immune-Based Strategies for the Control of Myxozoans
As inferred from the above sections, we are far from a comprehensive understanding of fish immune responses elicited against myxozoans. However, the information gathered to date enables us to identify promising opportunities for controlling these organisms.
7.1 Genetic Selection for Host Resistance
One possibility for disease control is to exploit the innate or natural resistance against Myxozoa observed in some fish species and strains, by means of genetic selection. The presence of natural resistance is implied by both interspecific and intraspecific differences in susceptibility in several systems (Table 14.1)
. In only a few cases have particular immune factors involved in such differences been suggested. For example, brown trout, Atlantic (Salmo salar) and coho (Oncorhynchus kisutch) salmon showed a considerably higher number of eosinophilic granule cells in M. cerebralis-affected areas compared to rainbow trout (Hedrick et al. 1999). In sharpsnout sea bream no lysozyme was detected in either E. leei-infected or healthy animals (Golomazou et al. 2006; Álvarez-Pellitero et al. 2008). The absence of lysozyme could therefore contribute to the high pathogenicity of E. leei in sharpsnout sea bream (Álvarez-Pellitero et al. 2008).
It is important to appreciate the potential complexity of resistance and its heritable basis. For example, rainbow trout strains resistant to C. shasta are susceptible to M. cerebralis, suggesting that different mechanisms might be specifically involved in the resistance to each myxozoan (Hedrick et al. 2001). Quantitative genetic studies and genome-wide mapping (Baerwald et al. 2011; Fetherman et al. 2012) provide evidence that resistance to M. cerebralis is heritable; and is based on a single quantitative trait locus (QTL) which explains a large percentage of the phenotypic variance contributing to whirling disease resistance in rainbow trout (Dionne et al. 2009) found that different MHC alleles were associated with different levels of susceptibility and resistance to M. cerebralis. In contrast, resistance to C. shasta is linked to multiple loci in rainbow trout (Nichols et al. 2003), however, protection can be overwhelmed by high and continuous exposure to C. shasta (Bjork and Bartholomew 2010).
7.2 Vaccination
A second strategy for controlling myxozoan disease is the development of vaccines to promote acquired resistance in fish upon re-exposure. For M. cerebralis, acquired immunity was found only among previously exposed fish that developed active infections (Hedrick et al. 1998). Similarly, trout that had recovered from clinical PKD were found to be resistant to reinfection if an active infection was previously induced (Foott and Hedrick 1987). Sharpsnout sea bream that recovered from E. leei infection were refractive to the disease in a second exposure (Golomazou et al. 2006). E. leei-infected red sea bream (Pagrus major) surviving mortalities in Japanese farms do not seem to have recurrent infections (Yanagida et al. 2008). The association between this acquired immunity and the production of specific antibodies has been demonstrated for turbot surviving E. scophthalmi epizootic outbreaks (Sitjà-Bobadilla et al. 2007b). It is speculated that these protective responses reflect a repertoire of non-specific immune factors that limit ceratomyxosis (Bartholomew 1998) and contribute to M. cerebralis resistance (Ryce 2003). In turbot enteromyxosis, antibody responses are slow in onset (50–100 days p.e.) compared to cellular and humoral effectors (20 days p.e.) (Sitjà-Bobadilla et al. 2006, 2007b). If innate immune responses to myxozoans can be activated more rapidly and to a greater extent this may accelerate the production of specific antibodies.
There is an empirical basis to search for suitable parasite antigens capable of triggering a protective immune response for several myxozooses. This is a complex task since myxozoans have several parasitic stages possibly with different antigenic profiles owing to adaptation to different hosts and environments in their life cycle. As an example, monoclonal antibodies raised against mature K. thyrsites spores do not react with stages found in exposed fish, suggesting they are antigenically distinct stages (Young and Jones 2005). Antigenic profiles have been obtained using classical approaches, such as enzyme-linked immunosorbent assay (ELISA), dot-blot, western-blot, lectin-blot, indirect fluorescent antibody technique (IFAT) and immunohistochemistry for a number of myxozoans, including S. dicentrarchi (Muñoz et al. 2000c), E. leei (Estensoro et al. 2013a), M. cerebralis (Kaltner et al. 2007; Knaus and El-Matbouli 2005b), K. thyrsites (Chase et al. 2001), H. salminicola (Clouthier et al. 1997), M. rotundus (Lu et al. 2002; Zhang et al. 2006) and T. bryosalmonae (Saulnier et al. 1996; Saulnier and de Kinkelin 1996).
More recently, other molecular approaches have become available such as Expression Library Immunization (ELI). One such study, integrating cDNA-based ELI (cDELI) with next generation sequencing approaches, is currently underway in the search for protective T. bryosalmonae antigens (Secombes and Holland, personal communication). ELI or cDELI consists of the systemic screening of any given genome or transcriptome to identify potential vaccine candidates. The essential concept is to employ the host immune system to select the best vaccine candidates against a particular disease. The entire genome or transcriptome of a pathogen is cloned into genetic vectors optimized for eukaryotic expression under the control of a powerful cytomegalovirus (CMV) promoter to create a library that would express all the open reading frames (ORFs) of a pathogen. Purified plasmid DNA from this library is inoculated into animals, usually in subgenomic pools of clones, to induce immune responses against the cloned antigens. The immunized animals are then challenged with the pathogenic organism to see which clones induced protective immunity and therefore which pool of antigens should be further deconvoluted to individual protective antigens (Talaat and Stemke-Hale 2005).
Another antigen screening option is the Serial Analysis of Gene Expression (SAGE ) of the pathogen (Ye et al. 2002). This is also promising for myxozoans, since the transcriptome and genome of several species is already engaged. This will offer a comprehensive catalogue of genes encoding all the potential proteins of the pathogen, with the potential to rationally select vaccine candidates rather than empirically test them one at a time. Furthermore, the strategy of antigen prediction is independent of the need to culture the pathogen in vitro, a bottleneck for myxozoan research. A complete genome sequence can be screened using bioinformatic algorithms to select ORFs encoding putative surface-exposed or secreted proteins, to select proteins with homology to known virulence factors or protective antigens from other pathogens, and to filter for proteins with no similarity to proteins of the host. After candidate antigens are identified in silico, they are produced as recombinant proteins or DNA vaccines and their immunogenicity is assayed to measure their potential as vaccine candidates.
7.3 Immune Therapies
An alternative strategy to vaccination for disease control is the development of specific immune therapies , designed to rebalance immune responses. This approach aims to alleviate the chronicity and intensity of myxozoan-mediated immunopathology in fish hosts. This has been demonstrated recently in mammals through the development of small peptide antagonists of SOCS proteins, by SOCS gene knockdown (Delgado-Ortega et al. 2013). Antibody-based neutralization of over-expressed IL-17 (one of the causative factors of chronic immunopathology and granuloma formation in helminth infections) has been shown to enhance parasite-specific antibodies and to reduce disease pathology and parasite prevalence (Wen et al. 2011). Mammalian MBL has some benefits towards relieving the intensity of chronic infections through the use of recombinant protein (Casanova and Abel 2004). Trout SOCS and IL-17 homologues up-regulated during PKD and E. leei-mediated suppression of GSB MBL-2 could be similarly controlled to facilitate immune protection and parasite clearance. Looking further ahead, tailoring fish vaccines by incorporating molecular adjuvants may enable the construction of highly efficacious vaccines priming the immune response and providing long-lasting immunological memory. This approach has been highly successful in mammalian vaccines utilizing a variety of cytokines, including IL-12, IL-18, and granulocyte monocyte colony stimulating factor (GMCSF) (Toka et al. 2004; Kathuria et al. 2012). Such advances, however, will need to go hand-in-hand with the continued functional characterization of fish immune molecules and pathways and, ultimately, the immune cell populations shaping disease pathogenesis and pathogen resistance.
7.4 Immunostimulants
Different commercial aquafeeds are nowadays formulated to include immunostimulant compounds, such as vitamins, nucleotides, minerals, glucans, prebiotics, probiotics, and herbal extracts. They are thought to enhance the fish basal immune system, the mucosal barriers, and the overall potential to fight against pathogens (Dalmo and Bøgwald 2008; Merrifield et al. 2010; Kiron 2012). However, their usefulness in myxozoan infections has not been fully determined, and it depends on the type of immune factor enhanced and its involvement in the immune response against each specific myxozoan. A diet including a glucan from yeast and the antiparasitic drug fumagilin decreased the number of O. mykiss infected with C. shasta, but did not improve the overall survival rate (Whipple et al. 2002). Similarly, bioactive, natural extract-supplemented feeds, although not preventing infection, decreased the intensity of infection and associated intestinal lesions of E. scophthalmi infected turbot and significantly increased survival rate (Palenzuela et al. 2009).
8 Conclusions
In the current chapter we have shown the complexity of the different immune factors evoked by the challenge of aquacultured fish species with myxozoan parasites, and the parasite strategies that have evolved to evade host immune responses. The importance of the local immune response and the interplay of host and environmental factors have been highlighted. Differences in the knowledge of the many fish-myxozoan models are outstanding, with most of the information coming from few species, and nothing is known about how invertebrates fight myxozoan infections. The existence of innate and acquired resistance to some myxozoan species opens promising possibilities to prevent myxozooses and therefore to alleviate the important economic losses provoked by them worldwide. We have come a long way in characterising the fish immune response, but still many aspects await to be deciphered. Only integrated and concerted research efforts will enable the identification of means to control diseases caused by myxozoans and to avoid their global dispersal in farmed fish stocks.
9 Key Questions for Future Studies
-
What are the key immunoregulatory molecules associated with disease susceptibility and intensity?
-
What methods can be explored to culture myxozoans in vitro?
-
Are myxozoan strategies to deal with host immune response similar in fish and invertebrates?
-
What are the protective mechanisms involved in disease resistance, and hence the pathways that need to be triggered by vaccination?
-
What are the main myxozoan virulence-associated targets for the development of drugs and vaccines?
References
Agius C, Roberts RJ (2003) Melano-macrophage centres and their role in fish pathology. J Fish Dis 26:499–509
Ainsworth AJ, Deixang C, Waterstraat PR, Greenway T (1991) Effect of temperature on the immune system of channel catfish (Ictalurus punctatus) I. Leucocyte distribution and phagocyte function in the anterior kidney at 10 °C. Comp Biochem Physiol—A Mol Integ Physiol 100:907–912
Álvarez-Pellitero P (2008) Fish immunity and parasite infections: from innate immunity to immunoprophylactic prospects. Vet Immunol Immunopathol 126:171–198
Álvarez-Pellitero P, Palenzuela O, Sitjà-Bobadilla A (2008) Histopathology and cellular response in Enteromyxum leei (Myxozoa) infections of Diplodus puntazzo (Teleostei). Parasitol Int 57:110–120
Arkush KD, Hedrick RP (1990) Experimental transmission of PKX, the causative agent of proliferative kidney disease, to three species of Pacific salmon. J Appl Ichthyol 6:237–343
Armstrong PB, Quigley JP (1999) Alpha (2) macroglobulin: an evolutionary conserved arm of the innate immune system. Dev Comp Immunol 23:375–390
Baerwald MR (2013) Temporal expression patterns of rainbow trout immune-related genes in response to Myxobolus cerebralis exposure. Fish Shellfish Immunol 35:965–971
Baerwald MR, Welsh AB, Hedrick RP, May B (2008) Discovery of genes implicated in whirling disease infection and resistance in rainbow trout using genome-wide expression profiling. BMC Genom 9:37
Baerwald MR, Petersen JL, Hedrick RP, Schisler GJ, May B (2011) A major effect quantitative trait locus for whirling disease resistance identified in rainbow trout (Oncorhynchus mykiss). Heredity 106:920–926
Ballesteros NA, Castro R, Abos B, Rodríguez Saint-Jean SS, Pérez-Prieto SI, Tafalla C (2013) The pyloric caeca area is a major site for IgM+ and IgT+ B cell recruitment in response to oral vaccination in rainbow trout. PLoS ONE 8:e66118
Bartholomew JL (1998) Host resistance to infection by the myxosporean parasite Ceratomyxa shasta: a review. J Aquat Anim Health 10:112–120
Bartholomew JL (2001) Parasites, their hosts and why disease occurs (or doesn’t?). In: 131st Annual Meeting American Fisheries Society, Phoenix, pp 20–24
Bartholomew JL, Smith CE, Rohovec JS, Fryer JL (1989) Characterization of a host response to the myxosporean parasite, Ceratomyxa shasta (Noble), by histology, scanning electron microscopy and immunological techniques. J Fish Dis 12:509–522
Bermúdez R, Vigliano F, Marcaccini A, Sitjà-Bobadilla A, Quiroga MI, Nieto JM (2006) Response of Ig-positive cells to Enteromyxum scophthalmi (Myxozoa) experimental infection in turbot, Scophthalmus maximus (L.): a histopathological and immunohistochemical study. Fish Shellfish Immunol 21:501–512
Bermúdez R, Faílde LD, Losada AP, Redondo MJ, Álvarez- Pellitero P, Quiroga MI (2009) Morphopathological and ultrastructural studies on enteromyxosis of turbot (Psetta maxima L.). In: Abstracts of the 14th international conference of the EAFP on diseases of fish and shellfish, Prague, 14–19 Sept 2009
Bettge K, Segner H, Burki R, Schmidt-Posthaus H, Wahli T (2009) Proliferative kidney disease (PKD) of rainbow trout: temperature-and time-related changes of Tetracapsuloides bryosalmonae DNA in the kidney. Parasitology 136:615–625
Bjork SJ, Bartholomew JL (2010) Invasion of Ceratomyxa shasta (Myxozoa) and comparison of migration to the intestine between susceptible and resistant fish hosts. Int J Parasitol 40:1087–1095
Bjork SJ, Zhang Y-A, Hurst CN, Alonso-Naveiro ME, Alexander JD, Sunyer JO, Bartholomew JL (2014) Defenses of susceptible and resistant Chinook salmon (Oncorhynchus tshawytscha) against the myxozoan parasite Ceratomyxa shasta. Fish Shellfish Immunol 3:87–95
Blazer VS, Densmore CL, Schill WB, Cartwright DD, Page SJ (2004) Comparative susceptibility of Atlantic salmon, lake trout and rainbow trout to Myxobolus cerebralis in controlled laboratory exposures. Dis Aquat Org 58:27–34
Bowden TJ (2008) Modulation of the immune system of fish by their environment. Fish Shellfish Immunol 25:373–383
Bradford MJ, Lovy J, Patterson DA, Speare DJ, Bennett WR, Stobbart AR, Tovey CP (2010) Parvicapsula minibicornis infections in gill and kidney and the premature mortality of adult sockeye salmon (Oncorhynchus nerka) from Cultus Lake, British Columbia. Can J Fish Aquat Sci 67:673–683
Bridle A, Nosworthy E, Polinski M, Nowak B (2011) Evidence of an antimicrobial-immunomodulatory role of Atlantic salmon cathelicidins during infection with Yersinia ruckeri. PLoS ONE 6:e23417
Bucher F, Hofer R, El-Matbouli M (1992) Prevalence and pathology of Zschokella nova (Myxosporea) in the liver of bullhead Cottus gobio from a polluted river. Dis Aquat Org 14:137–143
Burki R, Krasnov A, Bettge K, Rexroad CE III, Afanasyev S, Antikainen M, Burkhardt-Holm P, Wahli T, Segner H (2013) Molecular crosstalk between a chemical and a biological stressor and consequences on disease manifestation in rainbow trout. Aquat Toxicol 127:2–8
Calder PC (2007) Immunomodulation by omega-3 fatty acids. Prost Leuk Essent Fatty Acids 77:327–335
Casanova J-L, Abel L (2004) Human mannose-binding lectin in immunity: friend, foe, or both? J Exp Med 199:1295–1299
Calduch-Giner JA, Sitjà-Bobadilla A, Davey GC, Cairns MT, Kaushik S, Pérez-Sánchez J (2012) Dietary vegetable oils do not alter the intestine transcriptome of gilthead sea bream (Sparus aurata), but modulate the transcriptomic response to infection with Enteromyxum leei. BMC Genom 13:470
Chase JC, Dawson-Coates JA, Haddow JD, Steward MH, Haines LR, Whitaker DJ, Kent ML, Olafson RW, Pearson TW (2001) Analysis of Kudoa thyrsites (Myxozoa: Myxosporea) spore antigens using monoclonal antibodies. Dis Aquat Org 45:121–129
Chaves-Pozo E, Mulero V, Meseguer J, Ayala AG (2005) Professional phagocytic granulocytes of the bony fish gilthead seabream display functional adaptation to testicular microenvironment. J Leukoc Biol 78:345–351
Chilmonczyk S, Monge D, de Kinkelin P (2002) Proliferative kidney disease: cellular aspects of the rainbow trout, Oncorhynchus mykiss (Walbaum), response to parasitic infection. J Fish Dis 25:217–226
Clouthier SC, Gunning DJ, Olafson RW, Kay WW (1997) Antigenic characterization of Henneguya salminicola. Mol Biochem Parasitol 90:543–548
Cone D, Eurell T, Axler R, Rau D, Beasley V (1997) Intense infections with a variant of Myxobolus procerus (Myxosporea) in muscle of trout-perch (Percopsis omiscomaycus) in Duluth Harbor, Lake Superior. Folia Parasitol 44:7–11
Costa MM, Maehr T, Diaz-Rosales, P, Secombes CJ, Wang T (2011) Bioactivity studies of rainbow trout (Oncorhynchus mykiss) interleukin-6: effects on macrophage growth and antimicrobial peptide gene expression. Mol Immunol 48:1903–1916
Cuesta A, Muñoz P, Rodríguez A, Salinas I, Sitjà-Bobadilla A, Álvarez-Pellitero P, Esteban MA, Meseguer J (2006a) Gilthead seabream (Sparus aurata L.) innate defence against the parasite Enteromyxum leei (Myxozoa). Parasitology 132:95–104
Cuesta A, Salinas I, Rodriguez A, Muñoz P, Sitjà-Bobadilla A, Álvarez-Pellitero P, Meseguer J, Esteban MA (2006b) Cell-mediated cytotoxicity is the main innate immune mechanism involved in the cellular defence of gilthead seabream (Teleostei: Sparidae) against Enteromyxum leei (Myxozoa). Parasite Immunol 28:657–665
Dalmo RA, Bøgwald J (2008) β-glucans as conductors of immune symphonies. Fish Shellfish Immunol 25:384–396
Davey GC, Calduch-Giner JA, Houeix B, Talbot A, Sitjà-Bobadilla A, Prunet P, Pérez-Sánchez J, Cairns MT (2011) Molecular profiling of the gilthead sea bream (Sparus aurata L.) response to chronic exposure to the myxosporean parasite Enteromyxum leei. Mol Immunol 48:2102–2112
De Pablo MA, De Cienfuegos GA (2000) Modulatory effects of dietary lipids on immune system functions. Immunol Cell Biol 78:31–39
Delgado-Ortega M, Marc D, Dupont J, Trapp S, Berri M, Meurens F (2013) SOCS proteins in infectious diseases of mammals. Vet Immunol Immunopathol 151:1–19
Densmore CL, Blazer VS, Cartwright BDD, Schill WB (2001) A comparison of susceptibility to Myxobolus cerebralis among strains of rainbow trout and steelhead in field and laboratory trials. J Aquat Anim Health 13:220–227
Densmore CL, Ottinger CA, Blazer VS, Iwanowicz LR (2004) Immunomodulation and disease resistance in postyearling rainbow trout infected with Myxobolus cerebralis, the causative agent of whirling disease. J Aquat Anim Health 16:73–82
Dionne M, Miller KM, Dodson JJ, Bernatchez L (2009) MHC standing genetic variation and pathogen resistance in wild Atlantic salmon. Phil Trans Roy Soc B: Biol Sci 364:1555–1565
El-Matbouli M, Hoffmann RW (2002) Influence of water quality on the outbreak of proliferative kidney disease—field studies and exposure experiments. J Fish Dis 25(8):459–467
El-Matbouli M, Hoffmann RW, Mandok C (1995) Light and electron microscopic observations on the route of the triactinomyxon-sporoplasm of Myxobolus cerebralis from epidermis into rainbow trout cartilage. J Fish Biol 46:919–935
Ellis AE (1988) Ontogeny of the immune system in teleost fish. In: Ellis AE (ed) Fish vaccination. Academic Press, San Diego, pp 20–31
Estensoro I, Redondo MJ, Álvarez-Pellitero P, Sitjà-Bobadilla A (2010a) Detection of specific antibodies against Enteromyxum leei (Myxozoa: Myxosporea) in gilthead sea bream (Sparus aurata) by ELISA and immunohistochemistry. In: Abstracts of the 1st international symposium of the european organisation of fish immunology, Viterbo, May 2010
Estensoro I, Redondo MJ, Álvarez-Pellitero P, Bobadilla A (2010b) Novel horizontal transmission route for Enteromyxum leei (Myxozoa) by anal intubation of gilthead sea bream Sparus aurata. Dis Aquat Org 92:51–58
Estensoro I, Benedito-Palos L, Palenzuela O, Kaushik S, Sitjà-Bobadilla A, Pérez-Sánchez J (2011) The nutritional background of the host alters the disease course in a fish-myxosporean system. Vet Parasitol 175:141–150
Estensoro I, Redondo MJ, Salesa B, Kaushik S, Pérez-Sánchez J, Sitjà-Bobadilla A (2012a) Effect of nutrition and Enteromyxum leei infection on gilthead sea bream Sparus aurata intestinal carbohydrate distribution. Dis Aquat Org 100:29–42
Estensoro I, Calduch-Giner JA, Kaushik S, Pérez-Sánchez J, Sitjà-Bobadilla A (2012b) Modulation of the IgM gene expression and IgM immunoreactive cell distribution by the nutritional background in gilthead sea bream (Sparus aurata) challenged with Enteromyxum leei (Myxozoa). Fish Shellfish Immunol 33:401–410
Estensoro I, Álvarez-Pellitero P, Sitjà-Bobadilla A (2013a) Antigenic characterization of Enteromyxum leei (Myxozoa: Myxosporea). Dis Aquat Org 106:149–162
Estensoro I, Jung-Schroers V, Álvarez-Pellitero P, Steinhagen D, Sitjà-Bobadilla A (2013b) Effects of Enteromyxum leei (Myxozoa) infection on gilthead sea bream (Sparus aurata) (Teleostei) intestinal mucus: glycoprotein profile and bacterial adhesion. Parasitol Res 112:567–576
Estensoro I, Mulero I, Redondo MJ, Álvarez-Pellitero P, Mulero V, Sitjà-Bobadilla A (2014) Modulation of leukocytic populations of gilthead sea bream (Sparus aurata) by the intestinal parasite Enteromyxum leei (Myxozoa: Myxosporea). Parasitology 141:425–440
Fekete SG, Kellems RO (2007) Interrelationship of feeding with immunity and parasitic infection: a review. Vet Med 52:131–143
Fetherman ER, Winkelman DL, Schisler GJ, Antolin MF (2012) Genetic basis of differences in myxospore count between whirling disease-resistant and-susceptible strains of rainbow trout. Dis Aquat Org 102:97–106
Fijak M, Meinhardt A (2006) The testis in immune privilege. Immunol Rev 213:66–81
Fillatreau S, Six A, Magadan S, Castro R, Sunyer JO, Boudinot P (2013) The astonishing diversity of Ig classes and B cell repertoires in teleost fish. Front Immunol 4:28
Fischer U, Koppang EO, Nakanishi T (2013) Teleost T and NK cell immunity. Fish Shellfish Immunol 35:197–206
Fleurance R, Sauvegrain C, Marques A, Le Breton A, Guereaud C, Cherel Y, Wyers M (2008) Histopathological changes caused by Enteromyxum leei infection in farmed sea bream Sparus aurata. Dis Aquat Org 79:219–228
Foott JS, Harmon R, Stone R (2004) Effect of water temperature on non-specific immune function and ceratomyxosis in juvenile chinook salmon and steelhead from the Klamath River. Calif Fish Game 90:71–84
Foott JS, Hedrick RP (1987) Seasonal occurrence of the infectious stage of proliferative kidney-disease (PKD) and resistance of rainbow-trout, Salmo gairdneri Richardson, to reinfection. J Fish Biol 30:477–483
Funk VA, Olafson RW, Raap M, Smith D, Aitken L, Haddow JD, Wang D, Dawson-Coates JA, Burke RD, Miller KM (2008) Identification, characterization and deduced amino acid sequence of the dominant protease from Kudoa paniformis and K. thyrsites: a unique cytoplasmic cysteine protease. Comp Biochem Physiol—B Biochem Mol Biol 149:477–489
Furuta T, Ogawa K, Wakabayashi H (1993) Humoral immune response of carp Cyprinus carpio to Myxobolus artus (Myxozoa: Myxobolidae) infection. J Fish Biol 43:441–450
Golomazou E, Athanassopoulou F, Karagouni E, Tsagozis P, Tsantilas H, Vagianou S (2006) Experimental transmission of Enteromyxum leei Diamant, Lom and Dyková, 1994 in sharpsnout sea bream, Diplodus puntazzo C. and the effect on some innate immune parameters. Aquaculture 260:44–53
Gómez D, Bartholomew J, Sunyer JO (2013) Biology and mucosal immunity to myxozoans. Dev Comp Immunol 43:243–256
Gorgoglione B, Sommerville C (2010) Histopathological features of the infection by the myxosporean Zschokkella hildae Auerbach, 1910 in the kidney of Atlantic cod (Gadus morhua Linnaeus, 1758). In: 6th ISAAH—international symposium on aquatic animal health, Tampa
Gorgoglione B, Wang T, Secombes CJ, Holland JW (2013) Immune gene expression profiling of proliferative kidney disease in rainbow trout Oncorhynchus mykiss reveals a dominance of anti-inflammatory, antibody and T helper cell-like activities. Vet Res 44:55
Grabner DS, El-Matbouli M (2009) Comparison of the susceptibility of brown trout (Salmo trutta) and four rainbow trout (Oncorhynchus mykiss) strains to the myxozoan Tetracapsuloides bryosalmonae, the causative agent of proliferative kidney disease (PKD). Vet Parasitol 165:200–206
Grayfer L, Hodgkinson JW, Hitchen SJ, Belosevic M (2011) Characterization and functional analysis of goldfish (Carassius auratus) interleukin-10. Mol Immunol 48:563–571
Griffin MJ, Camus AC, Wise DJ, Greenway TE, Mauel MJ, Pote LM (2010) Variation in susceptibility to Henneguya ictaluri infection by two species of catfish and their hybrid cross. J Aquat Anim Health 22:21–35
Halliday MM (1976) The biology of Myxosoma cerebralis: the causative organism of whirling disease of salmonids. J Fish Biol 9:339–357
Hallett SL, Bartholomew JL (2012) Myxobolus cerebralis and Ceratomyxa shasta. In: Woo PTK, Buchmann K (eds) Fish parasites: pathobiology and protection. CABI, Oxfordshire, pp 131–162
Hasnain SZ, Gallagher AL, Grencis RK, Thornton DJ (2013) A new role for mucins in immunity: insights from gastrointestinal nematode infection. Int J Biochem Cell Biol 45:364–374
Hedrick RP, Wishkovsky A, Modin JC, Toth RJ (1991) Three myxosporeans found in the cranial tissues of rainbow trout in California. J. Aquat Anim Health 3:55–62
Hedrick RP, McConnell E, de Kinkelin P (1993) Proliferative kidney disease of salmonid fish. Ann Rev Fish Dis 3:277–290
Hedrick RP, El-Matbouli M, Adkison MA, MacConnell E (1998) Whirling disease: re-emergence among wild trout. Immunol Rev 166:365–376
Hedrick RP, McDowell TS, Gay M, Marty GD, Georgiadis MP, MacConnell E (1999) Comparative susceptibility of rainbow trout Oncorhynchus mykiss, and brown trout Salmo trutta to Myxobolus cerebralis, the cause of salmonid whirling disease. Dis Aquat Org 37:173–183
Hedrick RP, McDowell TS, Mukkatira K, Georgiadis MP, MacConnell E (2001) Salmonids resistant to Ceratomyxa shasta are susceptible to experimentally induced infections with Myxobolus cerebralis. J Aquat Anim Health 13:35–42
Hedrick RP, Mcdowell TS, Marty GD, Fosgate GT, Mukkatirak K, Myklebust K, El-Matbouli M (2003) Susceptibility of two strains of rainbow trout (one with suspected resistance to whirling disease) to Myxobolus cerebralis infection. Dis Aquat Org 55:37–44
Holmskov U, Thiel S, Jensenius JC (2003) Collectins and ficolins: humoral lectins of the innate immune defense. Annu Rev Immunol 21:547–578
Hong S, Li R, Xu Q, Secombes CJ, Wang T (2013) Two types of TNF-α exist in teleost fish: phylogeny, expression, and bioactivity analysis of type-II TNF-α3 in rainbow trout trout Oncorhynchus mykiss. J Immunol 191:5959–5972
Husain M, Bird S, van Zwieten R, Secombes CJ, Wang TH (2012) Cloning of the IL-1 beta 3 gene and IL-1 beta 4 pseudogene in salmonids uncovers a second type of IL-1 beta gene in teleost fish. Dev Comp Immunol 38:431–446
James K (1990) Interactions between cytokines and α-2 macroglobulin. Immunol Today 11:163–166
Jublanc E, Toubiana M, Sri Widada J, Le Breton A, Lefebvre G, Sauvegrain C, Marques A (2006) Observation of a survival case following infestation by Enteromyxum leei (Myxozoa: Myxosporea), a pathogenic myxosporidian of the digestive duct of the gilthead sea bream (Sparus aurata) in pisciculture. J Eukaryot Microbiol 53:S20
Kallert DM, Borrelli J, Haas W (2012) Biostatic activity of piscine serum and mucus on myxozoan fish infective stages. Fish Shellfish Immunol 33:969–976
Kaltner H, Stippl M, Knaus M, El Matbouli M (2007) Characterization of glycans in the developmental stages of Myxobolus cerebralis (Myxozoa), the causative agent of whirling disease. J Fish Dis 30:637–647
Karagouni E, Athanassopoulou F, Tsagozis P, Ralli E, Moustakareas T, Lytra K, Dotsika E (2005) The impact of a successful anti-myxosporean treatment on the phagocyte functions of juvenile and adult Sparus aurata L. Int J Immunopathol Pharmacol 18:121–132
Katharios P, Garaffo M, Sarter K, Athanassopoulou F, Mylonas CC (2007) A case of high mortality due to heavy infestation of Ceratomyxa diplodae in sharpsnout sea bream (Diplodus puntazzo) treated with reproductive steroids. Bull Eur Assoc Fish Pathol 27:43–47
Katharios P, Rigos G, Divanach P (2011) Enteromyxum leei (Myxozoa), a lethal intruder of tropical pet fish: first case in humphead wrasse, Cheilinus undulatus (Ruppell, 1835). J Exotic Pet Med 20:138–143
Kathuria N, Kraynyak KA, Carnathan D, Betts M, Weiner DB, Kutzler MA (2012) Generation of antigen-specific immunity following systemic immunization with DNA vaccine encoding CCL25 chemokine immunoadjuvant. Hum Vaccin Immunother 8:1607–1619
Kelley GO, Zagmutt-Vergara FJ, Leutenegger CM, Adkison MA, Baxa DV, Hedrick RP (2004) Identification of a serine protease gene expressed by Myxobolus cerebralis during development in rainbow trout Oncorhynchus mykiss. Dis Aquat Org 59:235–248
Kent ML, Hedrick RP (1985) Development of the PKX myxosporean in rainbow trout Salmo gairdneri. Dis Aquat Org 1:169–182
Kent ML, Hedrick RP (1987) Effects of cortisol implants on the PKX myxosporean causing proliferative kidney disease in rainbow trout, Salmo gairdneri. J Parasitol 73:455–461
Khoo L, Rommel FA, Smith SA, Griffin MJ, Pote LM (2010) Myxobolus neurophilus: morphologic, histopathologic and molecular characterization. Dis Aquat Org 89:51–61
Kiron V (2012) Fish immune system and its nutritional modulation for preventive health care. Anim Feed Sci Technol 173:111–133
Knaus M, El-Matbouli M (2005a) Characterisation of carbohydrate binding sites in developmental stages of Myxobolus cerebralis. Parasitol Res 97:505–514
Knaus M, El-Matbouli M (2005b) Lectin blot studies on proteins of Myxobolus cerebralis, the causative agent of whirling disease. Dis Aquat Org 65:227–235
Koehler A, Romans P, Desser S, Ringuette M (2004) Encapsulation of Myxobolus pendula (Myxosporidia) by epithelioid cells of its cyprinid host Semotilus atromaculatus. J Parasitol 90:1401–1405
Koevary SB (2000) Ocular immune privilege: a review. Clin Eye Vis Care 12:97–106
Koneti A, Linke MJ, Brummer E, Stevens DA (2008) Evasion of innate immune response: evidence for mannose binding lectin inhibition of tumopur necrosis factor alpha production by macrophages in response to Blastomyces dermatitidis. Infect Immun 76:994–1002
Laing KJ, Hansen JD (2011) Fish T cells: recent advances through genomics. Dev Comp Immunol 35:1282–1295
Lang T, Prina E, Sibthorpe D, Blackwell J (1997) Nramp1 transfection transfers Ity/Lsh/Bcg-related pleiotropic effects on macrophage activation: influence on antigen processing and presentation. Infect Immunol 65:380–386
Langdon JS (1991) Myoliquefaction post-morten (“milky flesh”) die to Kudoa thyrsites (Gilchrist) (Myxosporea. Multivalvulida) in mahi mahi. Coryphaena hippurus L. J Fish Dis 14:45–54
Leino RL (1996) Reaction of rodlet cells to a myxosporean infection in kidney of the bluegill, Lepomis macrochirus. Can J Zool 74:217–225
Le Morvan C, Troutaud D, Deschaux P (1998) Differential effects of temperature on specific and nonspecific immune defences in fish. J Exp Biol 201:165–168
Losada AP, Bermúdez R, Faílde LD, Quiroga MI (2012) Quantitative and qualitative evaluation of iNOS expression in turbot (Psetta maxima) infected with Enteromyxum scophthalmi. Fish Shellfish Immunol 32:243–248
Lovy J, Goodwin AE, Speare DJ, Wadowska DW, Wright GM (2011) Histochemical and ultrastructural analysis of pathology and cell responses in gills of channel catfish affected with proliferative gill disease. Dis Aquat Org 94:125–134
Lom J, Dyková I (1992) Protozoan parasites of fishes. In: Developments in aquaculture and fisheries science, vol 26. Elsevier, Amsterdam
Lu YS, Li M, Wu YS, Wang JG (2002) Antigenic study of Myxobolus rotundus (Myxozoa: Myxosporea) using monoclonal antibodies. J Fish Dis 25:307–310
MacConnell E, Vincent ER (2002) The effects of Myxobolus cerebralis on the salmonid host. Am Fish Soc Symp 2002:95–107
Marcogliese DJ, Cone DK (2001) Myxozoan communities parasitizing Notropis hudsonius (Cyprinidae) at selected localities of the St. Lawrence River, Quebec: possible effects of urban effluents. J Parasitol 87:951–956
Markkula SE, Karvonen A, Salo H, Valtonen ET, Jokinen EI (2007) Ultraviolet B irradiation affects resistance of rainbow trout (Oncorhynchus mykiss) against bacterium Yersinia ruckeri and trematode Diplostomum spathaceum. Photochem Photobiol 83:1263–1269
Martins ML, Xu DH, Shoemaker CA, Klesius PH (2011) Temperature effects on immune response and hematological parameters of channel catfish Ictalurus punctatus vaccinated with live theronts of Ichthyophthirius multifiliis. Fish Shellfish Immunol 31:774–780
Martyn AA, Hong H, Ringuette MJ, Desser SS (2002) Changes in host-derived cellular and extracellular matrix components in developing cysts of Myxobolus pendula (Myxozoa). J Eukaryot Microbiol 49:175–182
Merrifield DL, Dimitroglou A, Foey A, Davies SJ, Baker RTM, Bøgwald J, Castex M, Ringø E (2010) The current status and future focus of probiotic and prebiotic applications for salmonids. Aquaculture 302:1–18
Mitchell LG, Seymour CL, Gamble JM (1985) Light and electron microscopy of Myxobolus hendricksoni sp. nov. (Myxozoa: Myxobolidae) infecting the brain of the fathead minnow, Pimephales promelas Rafinesque. J Fish Dis 8:75–89
Molnár K (2002) Differences between the European carp (Cyprinus carpio carpio) and the coloured carp (Cyprinus carpio haematopterus) in susceptibility to Thelohanellus nikolskii (Myxosporea) infection. Acta Vet Hung 50:51–57
Monte MM, Wang T, Holland JW, Zou J, Secombes CJ (2013) Cloning and characterization of rainbow trout interleukin (IL)-17A/F2 and IL-17RA: expression during infection and bioactivity of the recombinant IL-17A/F2. Infect Immun 81:340–353
Morado JF, Sparks A (1986) Observations on the host-parasite relations of the pacific whiting, Merluccius productus (Ayres), and two myxosporean parasites, Kudoa thyrsites (Gilchrist, 1924) and K. paniformis (Kabata and Whitaker, 1981). J Fish Dis 9:445–455
Morris DJ, Adams A (2008) Sporogony of Tetracapsuloides bryosalmonae in the brown torut Salmo trutta and the role of the tertiary cell during the vertebrate phase of myxozoan life cycles. Parasitology 135:1075–1092
Mulero I, Sepulcre MP, Meseguer J, García-Ayala MA, Mulero V (2007) Histamine is stored in mast cells of most evolutionarily advanced fish and regulates the fish inflammatory response. Proc Nat Acad Sci USA 104:19434–19439
Muñoz P, Álvarez-Pellitero P, Sitjà-Bobadilla A (2000a) Modulation of the in vitro activity of European sea bass (Dicentrarchus labrax L.) phagocytes by the myxosporean parasite Sphaerospora dicentrarchi (Myxosporea: Bivalvulida). Fish Shellfish Immunol 10:567–581
Muñoz P, Sitjà-Bobadilla A, Álvarez-Pellitero P (2000b) Cellular and humoral immune response of European sea bass (Dicentrarchus labrax L.) (Teleostei: Serranidae) immunized with Sphaerospora dicentrarchi (Myxosporea: Bivalvulida). Parasitology 120:465–477
Muñoz P, Sitjà-Bobadilla A, Álvarez-Pellitero P (2000c) Antigenic characterization of Sphaerospora dicentrarchi (Myxosporea: Bivalvulida), a parasite from European sea bass Dicentrarchus labrax (Teleostei: Serranidae). Dis Aquat Org 40:117–124
Muñoz P, Cuesta A, Athanassopoulou F, Golomazou E, Crespo S, Padrós F, Sitjà-Bobadilla A, Albiñana G, Esteban MA, Álvarez-Pellitero P, Meseguer J (2007) Sharpsnout sea bream (Diplodus puntazzo) humoral immune response against the parasite Enteromyxum leei (Myxozoa). Fish Shellfish Immunol 23:636–645
Nichols KM, Bartholomew JL, Thorgaard GH (2003) Mapping multiple genetic loci associated with Ceratomyxa shasta resistance in Oncorhynchuus mykiss. Dis Aquat Org 56:145–154
Oliva-Teles A (2012) Nutrition and health of aquaculture fish. J Fish Dis 35:83–108
Ordás MC, Castro R, Dixon B, Sunyer JO, Bjork S, Bartholomew J, Korytar T, Köllner B, Cuesta A, Tafalla C (2012) Identification of a novel CCR7 gene in rainbow trout with differential expression in the context of mucosal or systemic infection. Dev Comp Immunol 38:302–311
Palenzuela O (2006) Myxozoan infections in Mediterranean mariculture. Parassitologia 48:27–29
Palenzuela O, Álvarez-Pellitero P, Sitjà-Bobadilla A (1999) Glomerular disease associated with Polysporoplasma sparis (Myxozoa) infections in cultured gilthead sea bream, Sparus aurata L. (Pisces: Teleostei). Parasitology 118:245–256
Palenzuela O, López-Grandal E, Zarza C, Álvarez-Pellitero P (2009) Treatment of turbot enteromyxosis with antiparasitic drugs and bioactive natural extracts-suplemented feeds. In: Abstracts of the 14th international conference of the EAFP on diseases of fish and shellfish, Prague, 14–19 Sept 2009
Pauley GB (1974) Fish sporozoa: extraction of antigens from Myxosoma cerebralis spores which mimic tissue antigens of rainbow trout (Salmo gairdneri). J Fish Res Bd Canada 31:1481–1484
Pérez-Sánchez J, Estensoro I, Redondo MJ, Calduch-Giner JA, Kaushik S, Sitjà-Bobadilla A (2013) Mucins as diagnostic and prognostic biomarkers in a fish-parasite model: transcriptional and functional analysis. PLoS ONE 8:e65457
Peribáñez MA, del Cacho E, Castillo JA, Arnal MC (1993) Immune complex-mediated glomerulopathy in Barbus graellsi infected with Myxobolus spp. Histol Histopathol 8:709–714
Quigley DTG, McArdie JF (1998) Management and control of proliferative kidney disease (PKD) in a freshwater Atlantic salmon (Salmo salar L.) farm in Ireland: a case history. Fish Vet J 2:1–12
Quiroga MI, Redondo MJ, Sitjà-Bobadilla A, Palenzuela O, Riaza A, Macías A, Vázquez S, Pérez A, Nieto JM, Álvarez-Pellitero P (2006) Risk factors associated with Enteromyxum scophthalmi (Myxozoa) infection in cultured turbot (Scophthalmus maximus L.). Parasitology 133:433–442
Raikova IG (1994) Life cycle, cytology and morphology of Polypodium hydriforme, a coelenterate parasite of acipenseriform fishes. J Parasitol 80:1–22
Ray RA, Holt RA, Bartholomew JL (2012) Relationship between temperature and Ceratomyxa shasta–induced mortality in Klamath River salmonids. J Parasitol 98:520–526
Redondo MJ, Álvarez-Pellitero P (2009) Lectinhistochemical detection of terminal carbohydrate residues in the enteric myxozoan Enteromyxum leei parasitizing gilthead seabream Sparus aurata (Pisces: Teleostei): a study using light and transmission electron microscopy. Folia Parasitol 56:259–267
Redondo MJ, Cortadellas N, Palenzuela O, Álvarez-Pellitero P (2008) Detection of carbohydrate terminals in the enteric parasite Enteromyxum scophthalmi (Myxozoa) and possible interactions with its fish host Psetta maxima. Parasitol Res 102:1257–1267
Redondo MJ, Palenzuela O, Riaza A, Macías MA, Álvarez-Pellitero P (2002) Experimental transmission of Enteromyxum scophthalmi (Myxozoa), an enteric parasite of turbot Scophthalmus maximus. J Parasitol 88:482–488
Reite OB (2005) The rodlet cels of teleostean fish: their potential role in host defence in relation to the role of mast cells/eosinophilic granule cells. Fish Shellfish Immunol 19:253–267
Rodríguez-Tovar LE, Speare DJ, Markham RJF (2011) Fish microsporidia: immune response, immunomodulation and vaccination. Fish Shellfish Immunol 30:999–1006
Rombout JHWM, Abelli L, Picchietti S, Scapigliati G, Kiron V (2011) Teleost intestinal immunology. Fish Shellfish Immunol 31:616–626
Ronza P, Coscelli G, Losada AP, Bermúdez R, Quiroga MI (2013) Changes in splenic melanomacrophage centres of turbot, Psetta maxima (L.), infected experimentally with Enteromyxum scophthalmi (Myxozoa). J Comp Pathol 148:92
Rucker U, El-Matbouli M (2007) Sequence analysis of OmNramp α and quantitative expression of Nramp homologues in different trout strains after infection with Myxobolus cerebralis. Dis Aquat Org 76:223–230
Ryce EKN (2003) Factors affecting the resistance of juvenile rainbow trout to whirling disease. PhD thesis, Montana State University, Bozeman
Salinas I, Zhang YA, Sunyer JO (2011) Mucosal immunoglobulins and B cells of teleost fish. Dev Comp Immunol 35:1346–1365
Saulnier D, Brémont M, de Kinkelin P (1996) Cloning, sequencing and expression of a cDNA encoding and antigen from the myxosporean parasite causing the proliferative kidney disease of salmonid fish. Mol Biochem Parasitol 83:153–161
Saulnier D, de Kinkelin P (1996) Antigenic and biochemical study of PKX, the myxosporean causative agent of proliferative kidney disease of salmonid fish. Dis Aquat Org 27:103–114
Schmidt-Posthaus H, Bettge K, Forster U, Segner H, Wahli T (2012) Kidney pathology and parasite intensity in rainbow trout Oncorhynchus mykiss surviving proliferative kidney disease: time course and influence of temperature. Dis Aquat Org 97:207–218
Schmidt-Posthaus H, Steiner P, Müller B, Casanova-Nakayama A (2013) Complex interaction between proliferative kidney disease, water temperature and concurrent nematode infection in brown trout. Dis Aquat Org 104:23–34
Secombes CJ, Wang T (2012) The innate and adaptive immune system of fish. In: Austin B (ed) Infectious disease in aquaculture: prevention and control, vol 231, Woodhead Publications Ltd., Cambridge. pp 3–68
Sepulcre MP, Pelegrín P, Mulero V, Meseguer J (2002) Characterisation of gilthead seabream acidophilic granulocytes by a monoclonal antibody unequivocally points to their involvement in fish phagocytic response. Cell Tiss Res 307:97–102
Severin VIC, El-Matbouli M (2007) Realtive quantifcation of immune-regulatory genes in two rainbow trout strains, Oncorhynchus mykiss, after exposure to Myxobolus cerebralis, the causative agent of whirling disease. Parasitol Res 101:1019–1027
Siau Y (1980) Observation immunologique sur des poissons du genre Mugil parasites par la Myxosporidie Myxobolus exiguus Thélohan, 1895. Parasitol Res 62:1–6
Sitjà-Bobadilla A (2008) Living off a fish: a trade-off between parasites and the immune system. Fish Shellfish Immunol 25:358–372
Sitjà-Bobadilla A, Álvarez-Pellitero P (1993) Pathologic effects of Sphaerospora dicentrarchi Sitjà-Bobadilla and Álvarez-Pellitero, 1992 and S. testicularis Sitjà-Bobadilla and Álvarez-Pellitero, 1990 (Myxosporea: Bivalvulida) parasitic in the Mediterranean sea bass Dicentrarchus labrax L. (Teleostei: Serranidae) and the cell-mediated immune reaction: a light and electron microscopy study. Parasitol Res 79:119–129
Sitjà-Bobadilla A, Palenzuela O (2012) Enteromyxum species. In: Woo PTK, Buchmann K (eds) Fish parasites: pathobiology and protection. CABI, Oxfordshire, pp 163–176
Sitjà-Bobadilla A, Redondo MJ, Macias MA, Ferreiro I, Riaza A, Álvarez-Pellitero P (2004) Development of immunohistochemistry and enzyme-linked immunosorbent assays for the detection of circulating antibodies against Enteromyxum scophthalmi (Myxozoa) in turbot (Scophthalmus maximus L.). Fish Shellfish Immunol 17:335–345
Sitjà-Bobadilla A, Redondo MJ, Bermudez R, Palenzuela O, Ferreiro I, Riaza A, Quiroga I, Nieto JM, Álvarez-Pellitero P (2006) Innate and adaptive immune responses of turbot, Scophthalmus maximus (L.), following experimental infection with Enteromyxum scophthalmi (Myxosporea: Myxozoa). Fish Shellfish Immunol 21:485–500
Sitjà-Bobadilla A, Diamant A, Palenzuela O, Álvarez-Pellitero P (2007a) Effect of host factors and experimental conditions on the horizontal transmission of Enteromyxum leei (Myxozoa) to gilthead sea bream, Sparus aurata L., and European sea bass, Dicentrarchus labrax (L.). J Fish Dis 30:243–250
Sitjà-Bobadilla A, Palenzuela O, Riaza A, Macias MA, Álvarez-Pellitero P (2007b) Protective acquired immunity to Enteromyxum scophthalmi (myxozoa) is related to specific antibodies in Psetta maxima (L.) (Teleostei). Scand J Immunol 66:26–34
Sitjà-Bobadilla A, Calduch-Giner J, Saera-Vila A, Palenzuela O, Álvarez-Pellitero P, Pérez-Sánchez J (2008) Chronic exposure to the parasite Enteromyxum leei (Myxozoa: Myxosporea) modulates the immune response and the expression of growth, redox and immune relevant genes in gilthead sea bream, Sparus aurata L. Fish Shellfish Immunol 24:610–619
Stolte EH, Nabuurs SB, Bury NR, Sturm A, Flik G, Savelkoul HFJ, Verburg-Van Kemenade BML (2008) Stress and innate immunity in carp: corticosteroid receptors and pro-inflammatory cytokines. Mol Immunol 46:70–79
Sugiyama A, Yokoyama H, Ogawa K (1999) Epizootiological investigation on kudoosis amani caused by Kudoa amamiensis (Multivalvulida: Myxozoa) in Okinawa prefecture, Japan. Fish Pathol 34:39–43
Swearer SE, Robertson DR (1999) Life history, pathology, and description of Kudoa ovivora n. sp. (Myxozoa, Myxosporea): an ovarian parasite of Caribbean labroid fishes. J Parasitol 85:337–353
Talaat AM, Stemke-Hale K (2005) Expression library immunization: a road map for discovery of vaccines against infectious diseases. Infect Immun 73:7089–7098
Takizawa F, Gómez D, Parra D, Zhang YA, Bjork S, Bartholomew J, Wiens GD, Sunyer JO (2013) Identification of regulatory B and T cell subsets in teleost fish. In: Abstracts of the 1st international conference of fish and shellfish immunology, Vigo, 25–28 June 2013
Taylor MD, van der Werf N, Maizels RM (2012) T cells in helminth infection: the regulators and the regulated. Trends Immunol 33:181–189
Toka FN, Pack CD, Rouse BT (2004) Molecular adjuvants for mucosal immunity. Immunol Rev 199:100–112
Tort L, Balasch JC, Mackenzie S (2003) Fish immune system. A crossroads between innate and adaptive responses. Immunologia 22:277–286
Verburg-Van Kemenade BML, Ribeiro CMS, Chadzinska M (2011) Neuroendocrine-immune interaction in fish: differential regulation of phagocyte activity by neuroendocrine factors. Gen Comp Endocrinol 172:31–38
Vigliano FA, Bermúdez R, Quiroga MI, Nieto JM (2006) Evidence for melano-macrophage centres of teleost as evolutionary precursors of germinal centres of higher vertebrates: an immunohistochemical study. Fish Shellfish Immunol 21:467–471
Vincent ER (1998) The relationship of time, temperature and fish life histories to whirling disease infections. In: Whirling disease symposium proceedings, research in progress, Fort Collins, pp 34–35, 19–21 Feb 1998
Vincent ER (2002) Relative susceptibility of various salmonids to whirling disease with emphasis on rainbow and cutthroat trout. Ameri Fish Soc Symp 29:109–115
Wagner E, Arndt R, Brough M (2002) Comparison of susceptibility of five cutthroat trout strains to Myxobolus cerebralis infection. J Aquat Anim Health 14:84–91
Wagner EJ, Wilson C, Arndt R, Goddard P, Miller M, Hodgson A, Vincent R, Mock K (2006) Evaluation of disease resistance of the Fish Lake-DeSmet, wounded man, and Harrison Lake strains of rainbow trout exposed to Myxobolus cerebralis. J Aquat Anim Health 18:128–135
Wahli T, Bernet D, Segner H, Schmidt-Posthaus H (2008) Role of altitude and water temperature as regulating factors for the geographical distribution of Tetracapsuloides bryosalmonae infected fishes in Switzerland. J Fish Biol 73:2184–2197
Wang P-C, Huang J-P, Tsai M-A, Cheng S-Y, Tsai S-S, Chen S-D, Chen SP, Chiu S-H, Liaw L-L, Chang L-T, Chen SC (2005) Systemic infection of Kudoa lutjanus n. sp. (Myxozoa: Myxosporea) in red snapper Lutjanus erythropterus from Taiwan. Dis Aquat Org 67:115–124
Wang T, Holland JW, Martin SAM, Secombes CJ (2010) Sequence and expression analysis of two T helper master transcription factors, T-beta and GATA3, in rainbow trout Oncorhynchus mykiss and analysis of their expression during bacterial and parasitic infection. Fish Shellfish Immunol 29:705–715
Wang T, Diaz-Rosales P, Costa MM, Campbell S, Snow M, Collet B, Martin SAM, Secombes CJ (2011) Functional characterization of a non-mammalian IL-21: rainbow trout Oncorhynchus mykiss IL-21 upregulates the expression of the Th cell signature cytokines, IFN-γ, IL-10, and IL-22. J Immunol 186:708–721
Wang T, Kono T, Monte MM, Kuse H, Costa MM, Korenaga H, Maehr T, Husain M, Sakai M, Secombes CJ (2013) Identification of IL-34 in teleost fish: differential expression of rainbow trout IL-34, MCSF1 and MCSF2, ligands of the MCSF receptor. Mol Immunol 53:398–409
Wang T, Husain M, Hong S, Holland JW (2014) Differential expression, modulation and bioactivity of distinct fish IL-12 isoforms: implication towards the evolution of Th1-like immune responses. Eur J Immunol 44:1541–1551
Wen X, He L, Chi Y, Zhou S, Hoellwarth J, Zhang C, Zhu J, Wu C, Dhesi S, Wang X, Liu F, Su C (2011) Dynamics of Th17 cells and their role in Schistosoma japonicum infection in C57BL/6 mice. PLoS Negl Trop Dis 5:e1399
Whipple MJ, Gannam AL, Bartholomew JL (2002) Inability to control ceratomyxosis in rainbow trout and steelhead with oral treatments of glucan immunostimulant or the fumagillin analog TNP-470. North Am J Aquacult 64:1–9
Woo PTK, Ardelli BF (2014) Immunity against selected piscine flagellates. Dev Comp Immunol 43:268–279
Yada T (2007) Growth hormone and fish immune system. Gen Comp Endocrinol 152:353–358
Yanagida T, Palenzuela O, Hirae T, Tanaka S, Yokoyama H, Ogawa K (2008) Myxosporean emaciation disease of cultured red sea bream Pagrus major and spotted knifejaw Oplegnathus punctatus. Fish Pathol 43:45–48
Yanagida T, Sameshima M, Nasu H, Yokoyama H, Ogawa K (2006) Temperature effects on the development of Enteromyxum spp. (Myxozoa) in experimentally infected tiger puffer, Takifugu rubripes (Temminck & Schlegel). J Fish Dis 29:561–567
Ye SQ, Usher DC, Zhang LQ (2002) Gene expression profiling of human diseases by serial analysis of gene expression. J Biomed Sci 9:384–394
Young CA, Jones SRM (2005) Epitopes associated with mature spores not recognized on Kudoa thyrsites from recently infected Atlantic salmon smolts. Dis Aquat Org 63:267–271
Zhang JY, Wang JG, Wu YS, Li M, Li AH, Gong XL (2006) A combined phage display ScFv library against Myxobolus rotundus infecting crucian carp, Carassius auratus auratus (L.), in China. J Fish Dis 29:1–7
Zhang JY, Yokoyama H, Wang JG, Li AH, Gong XN, Ryu-Hasegawa A, Iwashita M, Ogawa K (2010a) Utilization of tissue habitats by Myxobolus wulii Landsberg & Lom, 1991 in different carp hosts and disease resistance in allogynogenetic gibel carp: redescription of M. wulii from China and Japan. J Fish Dis 33:57–68
Zhang YA, Salinas I, Li J, Parra D, Bjork S, Xu Z, LaPatra SE, Bartholomew J, Sunyer JO (2010b) IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat Immunol 11:827–835
Acknowledgments
Gene transcription, transcriptomic, and expression library immunization studies using T. bryosalmonae as the model organism were supported financially by a Ph.D. studentship from DEFRA (Department for Environment, Food, and Rural Affairs; contract C3490) to Bartolomeo Gorgoglione, the Marine Alliance for Science and Technology for Scotland (MASTs), and the Biotechnology and Biological Sciences Research Council (BBSRC; contracts BB/F003242/1, BB/K009125/1). Part of the information on enteromyxoses gathered in this chapter has been obtained with funding from Spanish research projects (AGL2006-13158-C03-01, AGL2009-13282-C02, PROMETEO-II 2014/085 and ISIC 2012/003) and EU projects (AQUAMAX, FOOD-CT-2006-16249; AQUAEXCEL, FP7/2007-2012, no. 262336 and ARRAINA, FP7/2007/2013, no. 288925). The authors thank Dr. K. Molnár, Dr. J. Bartholomew, Dr. M.I. Quiroga and Dr. C. Tafalla for providing photomicrographs.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Sitjà-Bobadilla, A., Schmidt-Posthaus, H., Wahli, T., Holland, J.W., Secombes, C.J. (2015). Fish Immune Responses to Myxozoa. In: Okamura, B., Gruhl, A., Bartholomew, J. (eds) Myxozoan Evolution, Ecology and Development. Springer, Cham. https://doi.org/10.1007/978-3-319-14753-6_14
Download citation
DOI: https://doi.org/10.1007/978-3-319-14753-6_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-14752-9
Online ISBN: 978-3-319-14753-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)