
Abstract
Approximately 6% of the world’s flowering plant species have specialised stamen morphologies that require mechanical stimulation (vibration) by bees in order to release pollen concealed within. This has given rise to the study of the phenomenon of buzz pollination. Although buzz pollination sits squarely within the discipline of biotremology, this link rarely has been made explicit. Our aim in this chapter is to bridge the gap that historically has existed between buzz pollination research and the discipline of biotremology. We will discuss what we know about bee-induced floral vibrations and compare them to other kinds of plant-borne vibrational signals. We will also highlight how certain experimental approaches developed by biotremology researchers have helped buzz pollination investigators better understand the complex behavioural and ecological interactions occurring between buzz pollinated plants and their bee visitors. We will then provide an overview of research methodologies for buzz pollination scientists and describe some of the more commonly used experimental approaches for recording and playback of bee-induced floral vibrations. By highlighting the many common themes existing between studies in buzz pollination and biotremology we hope to stimulate others to explore the many exciting new research avenues in this unique biotic interaction.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


1 Introduction
The floral vibrations produced by some bees when visiting specialised types of flowers have puzzled researchers for more than a century (Teppner 2018). The relationship between flowers in which pollen is tightly protected inside floral structures that restrict pollen access and bees that use thoracic vibrations to remove pollen from these flowers has given rise to the phenomenon of buzz pollination (Buchmann 1983). The last few years have seen renewed and rapidly rising interest in the study of buzz pollination using a variety of approaches, including mechanical studies of floral and bee vibrations, phylogenetic analyses, plant-pollinator ecological investigations in the field, and bee behaviour (reviewed in Vallejo-Marín 2019). In this chapter, we provide an overview of the intimate and complex association between bees and flowers as it relates to the production of vibrations, and how plants respond to these vibrations through pollen release. Our aim is twofold: first, to make explicit the connection buzz pollination has to the discipline of biotremology, which, despite the many common themes that link them, have rarely been considered jointly (Hill 2008; Vallejo-Marín 2019) and second, to provide an overview of recording and playback methodologies used in buzz pollination research to assist scientists new to this field who want to become familiar with some of the more commonly used experimental approaches. Accordingly, we hope to emphasise the important contributions that buzz pollination can make to the discipline of biotremology and highlight how the study of buzz pollination is enriched by the experimental methods and perspectives developed by biotremology researchers. In doing so, we aim to encourage the development of exciting new research avenues on buzz pollination.
1.1 What Is Buzz Pollination?
Buzz pollination is a type of pollination that captures the interaction between a specific bee behaviour (the production of vibrations to extract pollen, sometimes called “sonication”) and the pollination of certain types of flowers (often, but not exclusively with poricidal floral morphologies) (Buchmann 1983). The terms “buzz pollination” and “floral sonication” are derived from the fact that there is an audible (i.e., airborne) sound that occurs as vibrations radiate off the body of the bee (and perhaps also the flower) into the surrounding air (Buchmann and Hurley 1978). It is important to note, however, that the airborne sound is just an incidental by-product since the relevant component of the bee’s action is vibrational in nature. Strictly speaking then, it may be more correct to refer to the actual behaviour as “floral vibration” (Vallejo-Marín 2019). Regardless, there is considerable ambiguity in the literature in the use of terminology to describe buzz pollination and its components, and often sonication and buzz pollination are used as synonyms. Future work will benefit from clearly distinguishing the bee behaviour of producing substrate-borne plant vibrations (sonication or floral vibration) from the interaction (buzz pollination) that arises between vibrating bees and flowers with specialised morphologies (poricidal flowers) (Vallejo-Marín 2019).
During buzz pollination, a female pollen-foraging bee (Fig. 11.1a) mechanically shakes a flower (or part of it) in order to release pollen concealed within a poricidal structure (Buchmann 1983; De Luca and Vallejo-Marín 2013). Typically, these structures are anthers, but in some flower species the anthers are kept within other floral parts such as corolla tubes, and thus in these cases the bee will vibrate the corolla in order to extract the pollen (Macior 1968; Corbet and Huang 2014). A female bee typically adopts a characteristic “C” body posture (Fig. 11.1b) when vibrating a flower (King 1993). She will grasp one or more anthers (or the corolla) with her mandibles, curl the ventral part of her body around them while in direct physical contact and then rapidly contract her indirect thoracic flight muscles. Vibrations are transmitted through the head, mandibles, and ventral thoracic and abdominal sclerites into the anthers, which cause the pollen grains inside to rapidly gain energy and exit in a large cloud through apical pores or slits (Buchmann and Hurley 1978; King and Buchmann 2003). The bee will then groom herself to collect the pollen, which will be transported back to the nest to feed developing larvae. The pollen that cannot be fully groomed from the body may then be incidentally transferred to the stigma of another conspecific flower the bee visits, thus facilitating fertilisation for the plant.

Buff-tailed bumblebee (Bombus terrestris audax L.) buzz pollinating a buffalo bur flower (Solanum rostratum Dunal). (a) Bee approaching the flower. (b) Bee vibrating the central anther cone. A bee typically grasps one or more anthers in her mandibles and presses the ventral part of her body tightly against them when producing vibrations. Photos by M Vallejo-Marín
Floral vibration behaviour is exclusively associated with bees (Anthophila; but see Buchmann et al. (1978) for a possible exception from a hoverfly, Copestylum mexicanum Macquart, mimicking a carpenter bee). Among bees, floral vibration behaviour is widespread and has been reported in 74 genera distributed among six families, which encompasses 58% of the approximately 20,000 described species (Cardinal et al. 2018). There are some notable exceptions, such as honeybees (Apis mellifera L.), which have never been reported to vibrate flowers for pollen collection (Buchmann 1983). Bees use floral vibrations to extract pollen from a variety of flowers, including those with non-poricidal morphologies (e.g., Pedicularis, Rhododendron, Rosa) (Buchmann 1983; De Luca and Vallejo-Marín 2013). Bees probably use floral vibrations as a mechanism to extract pollen when other strategies, such as gathering pollen with the legs, are inefficient or ineffective (Buchmann 1985; Russell et al. 2016; Papaj et al. 2017). Among flowering plants, roughly 6% of the 352,000 described species distributed among >72 families possess poricidal stamen morphologies that probably require mechanical stimulation to release pollen and are thus buzz pollinated. Buzz pollination also occurs in species of agricultural importance such as tomatoes (Solanum lycopersicum), blueberries (Vaccinium spp.) and kiwifruit (Actinidia deliciosa) (Buchmann 1983; De Luca and Vallejo-Marín 2013).
1.2 How Does Buzz Pollination Fit into Biotremology?
The vibrations produced by bees are transmitted to a flower via direct physical contact and are thus imparted as substrate-borne vibrations. This places buzz pollination studies directly within the sphere of the discipline of biotremology. However, for the most part, even though buzz pollination researchers rarely cite biotremology studies (the discipline of ‘biotremology’ was officially named relatively recently (Endler 2014; Hill and Wessel 2016)), we use the term synonymously with ‘vibrational communication’, which is how this field was traditionally referenced (Cocroft et al. 2014a). We conducted a survey of journal articles that specifically examined vibrational aspects of buzz pollination behaviour—published after the seminal Buchmann (1983) buzz pollination review and leading up to the present--and found 24 relevant papers (The author will share the list upon request). Of these, only 5% of the references within each paper were to biotremology (vibrational communication) studies (average number of references per paper = 33.29 (range: 9–63); average number of biotremology studies cited per paper = 1.67 (range: 0–12)). Furthermore, nearly 60% (14/24) of these papers contained no biotremology references at all. Our survey highlights the communication gap between buzz pollination research and biotremology, as a discipline.
Buzz pollination, however, represents a rapidly bourgeoning field within the discipline of biotremology. In recent years, botanists, pollination ecologists and animal behaviourists, many of whom are new to the discipline of biotremology, are beginning to utilise experimental approaches developed in the context of examining vibrational communication to explore questions about plant-insect mutualisms, co-evolutionary relationships between plants and pollinators, and the role of pollinators in mediating plant community structure at various ecological levels. For the remainder of this chapter, we will discuss what we know about bee-induced floral vibrations and compare them to other kinds of plant-borne vibrational signals. We will also highlight how certain experimental approaches developed by biotremology researchers have helped buzz pollination investigators better understand the complex behavioural and ecological interactions occurring between buzz pollinated plants and their bee visitors.
2 Production and Characteristics of Bee Vibrations
Floral vibrations, as with other types of mechanical waves, can be characterised by their duration, frequency, and amplitude characteristics (Fig. 11.2). In this section, we discuss how these characteristics vary in floral vibrations, and we also compare them to other kinds of plant-borne vibrations that are used in arthropod vibrational communication. A summary comparing buzz pollination and communication vibrations is provided in Table 11.1.

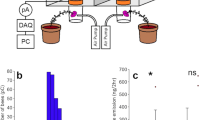
(a) Oscillogram (waveform) of four consecutive floral vibrations (buzzes) from a buff-tailed bumblebee (Bombus terrestris audax L.) worker on a watermelon nightshade flower (Solanum citrullifolium A. Braun), recorded with an accelerometer that was placed at the flower’s receptacle. (b) Frequency spectrum of the third vibration (indicated in purple inside the dashed lines in (a). The fundamental frequency in this vibration is also the dominant frequency (321 Hz). The first eight harmonics are shown with numbered dashed lines. (c) Spectrogram showing the magnitude of vibrations (relative amplitude, dB) across a range of frequencies (0–4 kHz). Lighter areas (yellow) represent frequencies of higher amplitude (data from Pritchard and Vallejo-Marín 2020)
2.1 Duration
Floral vibrations consist of one or multiple ‘buzzes’ or pulses produced in rapid succession (Fig. 11.2a). The duration of a single floral vibration ‘buzz’ usually lasts between 0.1 and 3.0 s (De Luca and Vallejo-Marín 2013). An individual bee will often produce several pulses within a single visit to a flower, and in these cases, there is often considerable variation in the durations of each pulse (and in the time interval between successive pulses). During floral visitation, a bee often quickly changes its position in consecutive buzzes, grasping and releasing an anther with its mandibles and usually moving from the base of the anther to its tip and probing different anthers (Vallejo-Marín, M. personal observation). Why this behaviour occurs is currently unclear, but it may allow the bee to maximise pollen release or assess pollen availability. Previous work has also shown that bees change the length of their floral vibrations depending on pollen reward levels (Buchmann and Cane 1989; De Luca et al. 2013; Russell et al. 2016) and this dynamic adjustment may allow them to maximise pollen release (De Luca et al. 2013). Floral vibrations thus lack the stereotyped higher order temporal patterns such as chirp trains or trills often exhibited in communication vibrations, which are used in species identification or sex-specific behavioural interactions (Čokl and Virant-Doberlet 2003; Cocroft and Rodrìguez 2005). The temporal variation in floral vibrations observed in bees is therefore likely to be due to dynamic responses to the interaction with flowers of varying characteristics and resource (pollen) levels.
2.2 Frequency
Bees generate floral vibrations through the contraction of the thoracic indirect flight muscles. Since the flight muscles are asynchronous, they can contract several times for each nervous stimulation (King et al. 1996). Floral vibrations produced by bees tend to have relatively pure-tone fundamental frequencies ranging between 100 and 400 Hz (De Luca and Vallejo-Marín 2013). The fundamental frequency is also the dominant frequency (i.e., has the greatest relative energy, Fig. 11.2b), as it is common for floral vibrations to exhibit harmonics (i.e., integer multiples of the fundamental) extending up to 8 kHz (Fig. 11.2c) (Buchmann et al. 1978; King and Lengoc 1993). However, the energy of harmonic frequencies is substantially lower and thus, they are not thought to be functionally significant (King 1993; but see Arceo-Gómez et al. 2011). It is interesting to note that floral vibration fundamental frequencies seem to have an upper limit around 400 Hz (Burkart et al. 2011; De Luca and Vallejo-Marín 2013; Switzer and Combes 2017; Rosi-Denadai et al. 2018; De Luca et al. 2019), even though it is possible for asynchronous flight muscle to contract at rates exceeding 500 Hz (Pringle 1949; Josephson et al. 2000; Tercel et al. 2018). Whether this limitation is a result of a physiological constraint for bees, or because frequencies above 400 Hz are not necessary for pollen ejection, certainly warrants further investigation.
For arthropods that communicate using plant-borne vibrations, signals are produced in a variety of ways (drumming, stridulation, tremulation, tymbals) using muscles located in the thorax and/or abdomen (Ossiannilsson 1949; Zeigler and Stewart 1977; Henry 1980; Morris 1980; Rovner and Barth 1981; Mitomi et al. 1984; Čokl et al. 2000; Miles et al. 2017; Eberhard et al. 2019). Fundamental frequencies of these signals are like floral vibrations, ranging between 50 and 500 Hz (Virant-Doberlet and Čokl 2004). Taxa that have been well studied include insect groups such as Hemiptera (true bugs), Neuroptera (lacewings), Orthoptera (crickets and katydids), Mantophasmatodea (heelwalkers), and Plecoptera (stoneflies), and arachnid groups such as Araneae (spiders). However, unlike bees, many other arthropods also possess some form of frequency-multiplier mechanism. This usually takes the form of a stridulatory device or tymbal and enables the production of vibrational signals with much higher broad band carrier frequencies that may extend up to 5 kHz (Michelsen et al. 1982; Čokl and Virant-Doberlet 2003; Elias et al. 2006). One function of these higher frequency vibrations may be related to the ability to localise a signaller on a plant. Generally, higher frequencies attenuate (lose energy and dissipate) faster than lower frequencies, and thus an individual may be able to discriminate the distance and direction of a signaller through evaluation of the specific frequencies it receives (Čokl and Virant-Doberlet 2003; Cocroft and Rodrìguez 2005; Gibson and Cocroft 2018).
2.3 Amplitude
The amplitude of a floral vibration refers to the magnitude (strength or intensity) of the vibrational wave as it propagates within a medium. It can be expressed as displacement, velocity or acceleration (Fig. 11.3). In addition, different summary statistics can be used to express magnitude. For example, peak amplitude (PA) refers to the maximum absolute value of a vibration, peak-to-peak (PK-PK) amplitude refers to the difference between the highest (peak) and lowest (trough) absolute values, and root mean square (RMS) amplitude refers to the square root of the peak amplitude (Speaks 1999). Buzz pollination studies that have measured amplitude usually report either peak velocity or peak acceleration (De Luca and Vallejo-Marín 2013). In simple sinusoidal vibrations (as floral vibrations are), knowledge of fundamental frequency and any of the three forms of amplitude (e.g., velocity) allows calculation of the other two (e.g., acceleration and displacement) (Vallejo-Marín 2019). The transducer a researcher uses to record vibrations determines which amplitude component should be reported. For example, phonograph cartridges are sensitive to displacement, laser Doppler vibrometers to velocity, and piezo-electric accelerometers to acceleration (Cocroft and Rodrìguez 2005). Amplitude is an extremely important parameter in buzz pollination research because it indicates the magnitude of a floral vibration, and therefore the forces that are transmitted to the flower that ultimately affect pollen release (Buchmann and Hurley 1978; King and Lengoc 1993; Harder and Barclay 1994; King and Buchmann 1996; De Luca et al. 2013; Rosi-Denadai et al. 2018; Switzer et al. 2019).

Amplitude waveform of a floral vibration plotted as (a) acceleration, (b) velocity, and (c) displacement. Velocity and displacement values were obtained by numerical integration of the acceleration waveform. The vibration was produced by a buff-tailed bumblebee (Bombus terrestris audax L.) worker and recorded with an accelerometer positioned at the receptacle of a watermelon nightshade (Solanum citrullifolium A. Braun) flower (data from Pritchard and Vallejo-Marín 2020)
For vibrationally communicating arthropods on plants, amplitude is an important signal property. Amplitude gradients experienced by an individual at different points on a plant may allow it to localise a signaller (Čokl and Virant-Doberlet 2003; Čokl et al. 2004; Gibson and Cocroft 2018), and peak amplitude may even provide information about individual signaller characteristics such as age or condition (De Luca and Cocroft 2009; De Luca 2015). Vibration amplitude is strongly influenced by substrate properties such as the thickness and stiffness of stems, branches and leaves, and it is also affected by the heterogeneity of the signalling environment, i.e., the complex three-dimensional structure of a plant (Michelsen et al. 1982; Casas et al. 2007; Cocroft et al. 2014b; Gibson and Cocroft 2018). These effects may be less important in floral vibrations, where bees directly contact floral structures containing pollen or when the distance travelled by the vibration is very short (e.g., between adjacent anthers). However, it is a much more complex issue in animal communication where there usually is some measurable distance between a sender and receiver. The extent to which floral structures affect the vibrations imparted by a bee is just beginning to be understood (King 1993; King and Buchmann 1995; Arroyo-Correa et al. 2019), and more work is needed to understand the effects of changes in the transmission properties of flowers for pollen release during buzz pollination.
2.4 Behavioural Considerations in the Production of Floral Vibrations
Floral vibrations differ from vibrations produced in other behavioural contexts such as flight and defence (Macior 1968; De Luca et al. 2014; Pritchard and Vallejo-Marín 2020). In Bombus terrestris L., floral vibrations have higher frequency, velocity and acceleration than defence vibrations, which are also produced without wing deployment (Pritchard and Vallejo-Marín 2020), suggesting that bees can modulate the characteristics of the vibrations they produce on flowers. Floral vibrations produced by bees contain both innate and learned components, but the extent to which bees can actively modulate vibrations to match specific foraging conditions is unclear. Naïve bees quickly begin producing floral vibrations on buzz pollinated flowers (Morgan et al. 2016), and the basic motor routines of buzz pollination behaviour seem to be innate (Russell et al. 2016). Yet, properties of their floral vibrations are modified significantly as naïve bees gain foraging experience. Studies have shown that duration, frequency and amplitude can all change with increased foraging experience, albeit in species-specific ways. For example, in B. impatiens Cresson both duration and amplitude increase over the first 100 vibrations on Solanum houstonii Dunal flowers (Russell et al. 2016), while in B. terrestris individuals that gained experience foraging on S. rostratum Dunal flowers show a decrease in both frequency and amplitude of their vibrations over ten foraging bouts (Morgan et al. 2016). In an ingenious recent study, naïve B. impatiens were allowed to forage on synthetic flowers where pollen release could be controlled by the experimenter. Here, larger individuals exhibited greater flexibility in producing vibrations that varied in frequency and amplitude in response to different pollen availability conditions, suggesting that size influences how bees modulate buzz pollination behaviour (Switzer et al. 2019). Experienced bees also demonstrate flexibility in behaviour when visiting rewarding vs unrewarding buzz pollinated flowers by adjusting the duration of vibrations (Buchmann and Cane 1989), or by actively switching between buzz pollination and scrabbling (collecting pollen without vibrating anthers) (Russell et al. 2017).
For vibrationally communicating insects, within-individual changes in signal properties have been documented, but these are thought to be more the result of age or condition-dependent effects than learning or experience per se (Kumar and Saxena 1985; Zeigler and Stewart 1985; Moreira 1993; De Luca and Cocroft 2009; Eberhard et al. 2019). However, communication signals may be actively modified through information gained from a receiver, as in male wolf spiders, which increase the rate at which vibrational courtship signals are produced in response to female receptivity cues (Sullivan-Beckers and Hebets 2014). Accordingly, in both animal communication and buzz pollination, within-individual changes in the production of vibrations appear to be influenced by a combination of ontogenetic, ecological and social factors, affecting both physical and behavioural aspects of the way the vibrations are modulated for the specific conditions being encountered.
3 Recording and Playback of Bee Vibrations on Flowers
3.1 Practical Aspects of Recording Floral Vibrations
For an experimenter there are two clear (and obvious) locations from which to record floral vibrations: the bee and the anthers (or corolla tubes). However, in practice, both locations have proven to be technically challenging. A buzzing bee does not typically remain stationary as it vibrates a flower and so attempting to measure directly from a moving target (for example with a laser vibrometer) is often difficult (but see Nunes-Silva et al. 2013). Similarly, measuring vibrations directly from anthers is problematic because they are usually concealed by a bee as it curls its body around them when vibrating. To get around these constraints, researchers using an appropriate vibration transducer often measure floral vibrations from another part of the flower that is more easily accessible. Two often used locations are the base (e.g., pedicel, calyx or receptacle) (Arroyo-Correa et al. 2019; Switzer et al. 2019) or the petals (De Luca et al. 2013, 2018). In the absence of a vibration transducer, however, one commonly used alternative is to record the airborne (acoustic) component of a floral vibration with a microphone (e.g., Macior 1968; Burkart et al. 2011; Corbet and Huang 2014; De Luca et al. 2014). The hypothesis here is that the buzzing sound constitutes a faithful reproduction of the substrate-borne vibration and therefore can be used as a proxy in situations where using a vibration transducer is not practical (Burkart et al. 2011). Do floral vibrations recorded from the flower, or airborne buzzing sounds recorded with a microphone, faithfully reproduce what bees impart into anthers? We explore this (see Sect. 11.3.1.1) when we discuss practical aspects of measuring and reproducing floral vibrations, using equipment and methodologies borrowed from the field of biotremology (see Chap. 8, for a comparison of the performance among industry-standard equipment for vibration recording and playback and some inexpensive alternatives). As in Sect. 11.2, we compare recording and playback methodologies between arthropod vibrational communication and buzz pollination studies, and we provide a summary of our comparisons in Table 11.2.
3.1.1 Types of Transducers
There are two types of recording devices that are commonly used in buzz pollination studies: (1) transducers that are sensitive to the vibrational (plant-borne) component, and (2) microphones that are sensitive to the acoustic (airborne) sounds of floral vibrations. In this section, we will provide an overview of each type and highlight the methodological advantages and disadvantages of each.
Vibration Transducers
The relevant mechanical component of the action of a buzz pollinating bee is vibrational in nature. Accordingly, using an appropriate vibration transducer remains the best method for recording these vibrations. The first type we will discuss are laser Doppler vibrometers. These devices are a non-contact method that utilises a laser beam, which reflects off the vibrating surface to provide frequency and amplitude (measured as velocity) information about the vibration. Laser vibrometers are best suited for laboratory studies but portable models (e.g., Polytec PDV100; Tustin, CA, USA) are available and can be transported to the field and powered by a battery. Lasers such as the PDV100 offer both digital and analogue ports that enable recordings to be stored as either digital files (typically in S/P-DIF format) or as a voltage time series, both of which preserve the absolute amplitude information of the vibration. During recording, the laser should be situated so that the beam is perpendicular to the direction of wave propagation for maximum sensitivity (McNett et al. 2006). Signal to noise ratios are highest with good reflection of the laser from the measured surface, and thus it is usually necessary to mount a small piece of reflective tape on the vibrating surface to increase reflectance of the laser beam. For high fidelity recordings the laser beam must be in sharp focus on the vibrating surface; therefore, strong external disturbances on the vibrating surface of interest will usually result in noisy and unusable recordings. For example, wind-induced vibrations on plants in the field are a significant source of noise that can mask vibrations (McNett et al. 2010). In the laboratory where environmental conditions can be controlled, laser vibrometry is a powerful method for obtaining floral vibrations.
The second type of vibration transducer commonly used in buzz pollination research is the piezo-electric accelerometer, which measures the acceleration of a vibrating surface. Accelerometers are attached directly to the vibrating object, and in buzz-pollination studies, they are commonly attached to the pedicel, calyx or receptacle of the flower. Typically, glue or beeswax is used to affix an accelerometer to the vibrating surface, but an alternative method is to glue an insect pin to the accelerometer and then attach (insert or firmly contact) the pin on the plant structure of interest (Arroyo-Correa et al. 2019; Switzer et al. 2019; Pritchard and Vallejo-Marín 2020). In contrast to the laser vibrometer, the main drawback of accelerometers is that they impose an extra weight on the vibrating surface. The additional weight of the accelerometer (and associated cables connecting the accelerometer with the signal recorder) can affect the transmission properties of the substrate being investigated (Cocroft and Rodrìguez 2005). Using lightweight models is desirable in order to minimise the effects of the extra mass loading (Cocroft and Rodrìguez 2005). In mechanical engineering, a rule of thumb is to use accelerometers that weigh less than 5% of the mass of the vibrating object, but with floral weights commonly in the range of less than a gram (e.g., Solanum) even miniature piezoelectric accelerometers (0.2–0.8 g) represent a significant fraction of the coupled system.
An advantage over laser vibrometers is that accelerometers are much easier to set up and reposition; therefore, they tend to work better under field conditions. As with lasers, environmental sources of noise such as wind can induce unwanted vibrations in a flower, which can potentially mask floral vibrations. Output from an accelerometer is analogue (a change in voltage over time), which can be converted to acceleration using the calibration reference information specific to the type and model being used (see Arroyo-Correa et al. 2019; Switzer et al. 2019; Pritchard and Vallejo-Marín 2020). Regardless of the type of vibration transducer, it is important to consider how the transmission path between the source of the vibration and the sensor may affect vibrational properties. The material and mechanical properties of the substrate can affect both the frequency and magnitude of the vibrations. Factors such as plant characteristics, and the distance between a signaller and recording device, can have unpredictable effects on vibration amplitude and thus need to be accounted for (see Sect. 11.2.3; Cocroft et al. 2014b). For example, a researcher might be interested in assessing the vibrations produced by different species of bees buzzing a flower by placing a sensor somewhere in the flower (e.g., the pedicel). Because the vibrations measured on the flower will depend not only on the bee species but on how the vibrations are changed as they pass through the flower, it is necessary to account for this potentially confounding floral effect. A solution is to empirically estimate the plant’s ‘coupling factor’, which can be done by stimulating anthers with a vibration of known amplitude and then measuring its value with the transducer positioned at the desired recording location. The difference in the amplitude ratio between the two locations is then used to calculate the coupling factor and estimate the vibrations produced at the source (bee) (King 1993; Arroyo-Correa et al. 2019). However, it is important to remember that this is just a rough approximation, as replicating exactly the way in which a bee manipulates a flower during buzzing will be difficult. Yet, this may be the best experimental approach at this time. Very few studies have estimated the coupling factor of different types of flowers (e.g., King 1993), but it seems that even closely related plant taxa have statistically different coupling factors (Arroyo-Correa et al. 2019).
Microphones
An alternative method of recording buzz pollination vibrations is to focus on the acoustic component of floral vibrations using a microphone. Here, the signal of interest is the airborne buzzing sound that accompanies the production of floral vibrations (Buchmann 1983). This method has been used for over 50 years (Macior 1968), and it continues to be a popular choice for researchers, particularly in the field (Burkart et al. 2011; Corbet and Huang 2014; Switzer and Combes 2017; De Luca et al. 2019). The advantage of this approach is ease of use: modern handheld digital recorders are easy to carry and move around, and so rather than waiting (often patiently!) by a flower that was chosen in advance for a bee to visit with a laser or accelerometer, a researcher can instead actively follow a foraging bee as it moves from flower to flower. This permits the collection of a large sample of recordings in a relatively short amount of time compared to what can be obtained with vibration transducers. For the optimal recording of airborne buzzing sounds, the microphone should be positioned perpendicular to the dorsal surface of the bee’s thorax (where the sound radiates most intensely) and be as close as possible without disturbing the natural behaviour of the bee. The distance between the bee and the microphone should be recorded in the data in order to later ensure that the sample was taken outside the near field, but also to be able to calculate attenuation with distance and control for that with multiple samples. Recordings of airborne buzzing sounds have been shown to faithfully reproduce spectral (frequency) and temporal (duration) patterns of floral vibrations, thus their use as proxies for their corresponding vibrational components can be argued (De Luca et al. 2018). However, measures of acoustic power (e.g., dB SPL) are not strongly correlated with vibration amplitude (measured as peak velocity) and should not be used (De Luca et al. 2018). Relative measures of acoustic power have been used in comparative studies, and are appropriate, if the signals being compared are recorded in exactly the same way (De Luca et al. 2014; Morgan et al. 2016). This means ensuring that the distance and orientation of bees to the microphone is kept the same, and that the microphone gain setting is kept identical between recordings.
One consideration of using microphones is background acoustic noise, which can often mask the sounds of buzz pollination vibrations. Sources can include wind, passing vehicles, machinery and animals (especially nearby singing birds); therefore, researchers should be aware of these potential confounding influences and compensate accordingly either during recording (e.g., using wind screens to reduce wind noise) or during analysis (e.g., using high pass filters to reduce low-frequency noise below 100 Hz). Filters used post-data collection may be of limited use if a recording contains noise at frequencies that overlap those of floral vibrations (i.e., within the 100–400 Hz range). We refer readers to Sueur (2018) for an excellent and detailed introduction to sound analysis, including the use of digital filtering.
Another consideration of using microphones is that on some occasions identifying the correct fundamental frequency of a floral vibration can be difficult if the researcher only focuses on the dominant frequency value in a recording. Recordings made with a vibration transducer (e.g., laser or accelerometer) show the fundamental frequency as the dominant frequency. However, acoustic recordings of floral vibrations may sometimes result in a harmonic frequency being dominant (Fig. 11.4). This difference is probably associated with several factors, including the difference in transmission properties of mechanical vibrations in air vs solid mediums, the sensitivity of a microphone brand to a specific frequency range, and because the effect the size of a vibrating object has on the sound (airborne) frequencies that radiate most efficiently from it (Michelsen and Nocke 1974; Michelsen et al. 1982; Bennet-Clark 1989, 1998). Therefore, although the overall spectral pattern (fundamental and associated harmonics) of a floral vibration recorded with an acoustic microphone could be used as a proxy for that recorded with a vibration transducer in some situations, the specific vibration frequency that transmits best in the flower often does not match the sound frequency that transmits best in the air (Michelsen et al. 1982). Accordingly, researchers using acoustic recordings should examine frequency spectra carefully to ensure that a higher frequency harmonic (>500 Hz) is not used to approximate the fundamental within plant tissue simply because it is the dominant frequency recorded in air. However, the main point to make (in our view) is that acoustic recordings can be a proxy (with the described limitations) only for the bee’s behaviour, the buzzing, and not the mechanical response of the flower itself.

Example of a floral vibration simultaneously recorded with a Polytec PDV100 laser Doppler vibrometer (a, c) and a Zoom H4 acoustic microphone (b, d) from an Eastern bumblebee (Bombus impatiens Cresson) worker on a ‘diente de burro’ nightshade (Solanum houstonii Dunal) flower. The microphone was positioned 14 cm from the flower and the laser beam was directed on one of the flower’s petals, 5 mm from the base of the anthers. Top panels (a, b) show the entire floral vibration in the time domain, and the bottom panels (c, d) show the frequency domain of the section marked with black vertical dashed lines in the top panels. Vertical dashed lines in c and d indicate the position for the fundamental (1) and harmonic (2–8) frequencies calculated from the laser signal. In the spectrum from the laser recording (bottom left panel) the fundamental (339 Hz) is also the dominant frequency. In contrast, in the acoustic recording (bottom right panel) the dominant frequency is at the second harmonic (738 Hz) (data from De Luca PA, unpublished)
Videography
Some of the earliest experimental investigations into buzz pollination were conducted using high-speed cameras (Macior 1964). This approach has enabled researchers to construct detailed ethograms describing the sequence of behaviours involved in floral sonication (Macior 1968; Russell et al. 2016). Furthermore, when video is combined with acoustic recording it enables researchers to connect physical movements of the bee to changes in buzzing properties (e.g., fluctuations in duration or frequency) as the vibration is imparted into the flower (Switzer et al. 2016).
3.2 Playback of Vibrations on Flowers
Characterising properties of bee-induced floral vibrations represents just one side of the buzz pollination story. The other equally important side concerns the effect bee vibrations have on pollen release (Buchmann and Hurley 1978; Harder and Barclay 1994; King and Buchmann 1995, 1996). Although we know a great deal about causes of variability in bee vibrations, we are still in the beginning stages of understanding the factors affecting vibratile ejection of pollen from anthers. The reason is perhaps practical—recording vibrations from bees is comparatively easier to do than performing the kinds of intricate playback experiments needed to adequately evaluate how bee vibrations influence pollen release. Accordingly, this is where knowledge of the challenges involved in the playback of vibrational communication signals can help buzz pollination researchers. The good news is that recent biotremology studies have specifically dealt with evaluating the most common playback issues on plant substrates. We have reviewed the few studies that have examined the effect of floral vibrations on pollen release (see Sect. 11.3.2.1) and then evaluated the equipment and methodologies that are available for investigating this phenomenon and provide some recommendations for performing well-designed playback experiments (see Sect. 11.3.2.2).
3.2.1 Review of Experimental Studies to Date
Only a handful of experimental studies have directly assessed how variability in the properties of floral vibrations influences pollen ejection. Buchmann and Hurley (1978) developed a biophysical model that examined how pollen grains inside an anther might behave when the anther was vibrated. A key result was that both vibration frequency and amplitude (expressed as velocity) were important predictors of how quickly pollen grains gain energy and are expelled through apical pores in the anther tips. Accordingly, subsequent experiments have attempted to reproduce bee vibrations on anthers to quantify pollen ejection. Buchmann et al. (1978) used a tuning fork to vibrate the anthers of two Solanum species. Although significant quantities of pollen were ejected, no attempt was made to modulate the duration or amplitude of the vibrations emanating from the tuning fork to approximate what a bee might naturally produce. Also, the tuning fork generated a 512 Hz pure tone, which is more than 100 Hz higher than the maximum floral vibration frequencies reported for bees (De Luca and Vallejo-Marín 2013; De Luca et al. 2019). Five subsequent studies were more systematic in their methodology and provide more compelling results. Here, researchers generated artificial stimuli that varied in key properties such as frequency, amplitude and duration, and applied them to anthers of various flowers (i.e., Actinidia, Dodecatheon, Rhododendron and several Solanum species), using either a loudspeaker to which a fine wire was glued (Corbet et al. 1988), or a vibration exciter (King and Lengoc 1993; Harder and Barclay 1994; King and Buchmann 1995, 1996). Three of these studies varied amplitude and found more pollen was ejected as amplitude increased (measured as either displacement or acceleration). However, results were equivocal with respect to frequency. Two studies showed more pollen was released with stimuli that contained fundamental frequencies well above what bees naturally produce (i.e., 400–1000 Hz) (Corbet et al. 1988; Harder and Barclay 1994), while two reported greater pollen ejection within the range used by bees (i.e., 100–400 Hz) (King and Buchmann 1995, 1996). These studies verified that amplitude and frequency were indeed important functional properties of floral vibrations, as varying them affected the quantity of pollen that was ejected from anthers. However, the methodologies in these papers were sometimes vague regarding the number of stimuli used, and it was not made abundantly clear how values for some properties (e.g., duration) were adjusted between different playback exemplars. A study by De Luca et al. (2013) adopted the same experimental approach but the researchers here greatly expanded the number of stimuli that were used, while describing how stimuli were generated. They created 294 different stimuli that simultaneously varied in frequency, duration and amplitude (in seven discrete steps, i.e., mean ± 1, 2, 3 SD) based on measurements taken from 54 workers of Bombus terrestris L. bumblebees foraging on Solanum rostratum Dunal flowers. Results of the playback experiment revealed that amplitude and duration were positively correlated with pollen release (with amplitude having four times the influence as duration). However, varying frequency had a weak significant quadratic effect on the amount of pollen ejected. They also found significant correlational effects (e.g., a positive interaction between amplitude and duration), suggesting that pollen ejection is a complex process that likely involves different vibrational properties interacting with one another. Most recently, Rosi-Denadai et al. (2018) evaluated pollen ejection in tomatoes (S. lycopersicum L.) using 40 artificially generated stimuli that varied in amplitude and frequency (duration was kept constant at 2 s across stimuli). Here, frequency was varied across a wider range (100–1600 Hz) than what bees naturally produce, but their results indicated no single frequency was optimal to maximise pollen release. Rather, variation in amplitude had the greatest effect on the quantity of pollen extracted from anthers, with higher amplitudes releasing more pollen. The results of these eight studies established the important functional role that floral vibrations play in affecting pollen release and highlighted the relative importance of properties such as amplitude, duration and frequency. However, we still have much to learn about how flowers respond to bee-induced vibrations. For example, little comparative data currently exists for flowers with different stamen morphologies (e.g., Solanum vs. Pedicularis) regarding their responses to floral vibrations. Accordingly, more experiments investigating the biophysical responses of anthers (and of pollen grains) are needed to help us better understand how vibrations transmitted from the bee to the anther translate into useful work to affect pollen release. This will require an integrative approach combining analyses of bee behaviour and floral biomechanics, and how these factors are connected through buzz pollination.
3.2.2 Descriptions of Playback Systems and Experimental Approaches
When designing a playback experiment, selecting the right equipment to correctly address the question is always a prime concern. There are several vibration playback systems that are available, and the only limitation for researchers is likely to be cost. The most important methodological consideration for buzz pollination researchers is constructing an apparatus that transmits bee vibrations to anthers in a biologically realistic manner. All the studies discussed in the previous section utilised different methods to excite anthers that we further explain in this section.
Vibration Exciters
The use of an electrodynamic vibrator (also called a mini-shaker) is by far the most common device that has been used in vibrational playback experiments. Electrodynamic shakers such as the popular Brüel & Kjær (B & K) 4810 shaker (Brüel & Kjær, Naerum, Denmark) are relatively small and can be positioned in any spatial orientation with appropriate clamps. These types of vibrators can handle a wide range of frequencies and amplitudes and thus are quite versatile. The main technical challenge is constructing an appropriate accessory piece to connect to the mounting base to adequately transmit vibrations to flowers in a manner that realistically approximates the action of a buzzing bee. This has ranged from simply attaching anthers directly to the shaker base with tape (King and Lengoc 1993; Harder and Barclay 1994; King and Buchmann 1996), fashioning metal hooks that wrap around the base of anthers (Rosi-Denadai et al. 2018), or using featherweight forceps that grip anthers analogous to the way a bee grips them in her mandibles (De Luca et al. 2013).
Loudspeakers
Another method of transmitting vibrations to flowers is with an audio loudspeaker. They are easily modified into a substrate-borne transducer by removing the membrane and affixing a metal wire or pin to the centre moving coil, which is then pressed against the anthers (Corbet et al. 1988). Those interested in utilising this approach are referred to Rodrigues et al. (2018) for further information. The authors provide detailed step-by-step instructions for constructing an affordable playback apparatus using a loudspeaker that is powered by a laptop and uses freely available software (e.g., Audacity: http://audacity.sourceforge.net/) to generate stimuli.
3.3 Calibrating Stimuli
Regardless of the playback system used, it is imperative to ensure that playback stimuli are correctly calibrated in the properties of interest prior to conducting the experiment. This will usually entail performing a preliminary test in which stimuli are applied to the substrate (e.g., anthers) and the resulting vibrations are recorded and inspected for accuracy. Our opinion is that amplitude constitutes the most important parameter for attention in playback studies of floral vibrations. Accordingly, experimenters should ensure that the appropriate amplitude component (acceleration, velocity or displacement) is faithfully reproduced. Recall that alternative recording transducers are sensitive to different amplitude components, and so if vibrations are recorded with an accelerometer it is recommended that playback stimuli are calibrated to reproduce correct acceleration values, and if a laser vibrometer is used then stimuli should be calibrated to velocity. To our knowledge, phonograph cartridges (which are sensitive to displacement) have never been used to record floral vibrations, and so we have no experience in their use as a method of calibrating playback stimuli. However, Cocroft et al. (2014b) discuss their use in vibrational communication studies and so we refer interested readers to that chapter for more information.
One difference in vibrational playback studies between buzz pollination and animal communication concerns the calibration of frequency. Plants, just as other substrates, act as filters that may drastically alter the frequency components of a broad band signal (Michelsen et al. 1982). This is important in animal communication studies because many vibrational signals often contain a range of frequencies, and so it is crucial to ensure that initial playback stimuli are conditioned, or matched to the specific plant filtering properties. By the time these playback stimuli reach the intended receiver, any alteration in the initial playback frequencies due to plant-substrate filtering (which cannot be controlled) will result in the correct signal properties required for a realistic proxy for a naturally occurring signal. Thus, playback signal in will serve as a reasonable proxy for natural signal out. A variety of compensation methods are available (reviewed in Cocroft et al. 2014b), including recently developed software tools that make their implementation straightforward (Michael et al. 2019), which ensures that playback signals containing a range of frequencies are correctly transmitted to a plant. In contrast, floral vibrations may be less affected by these filtering effects as they consist of pure tones (fundamental and a few harmonics) that decay in magnitude with distance, and thus are less likely to be adversely affected by substrate filtering effects when compared to broad band communication vibrations (Cocroft et al. 2014b). Moreover, since the experimental protocol for anther stimulation involves direct contact of the playback transducer to the anthers, there is little opportunity for floral vibrations to be altered before they reach their intended location.
4 Budding Buzz Pollination: Conclusions and Future Directions
The field of buzz pollination has made great strides in recent years, benefitting from the diverse viewpoints and approaches of researchers actively investigating the many facets of this specialised pollination syndrome. Although the majority of studies have focused on proximate causes, describing the behaviour and mechanisms governing the production of floral vibrations, there is rapidly growing interest in utilising more integrative approaches to examine ultimate (evolutionary) causes of bee-flower interactions, most notably the intersection between bee and floral characteristics and pollen release (Rosi-Denadai et al. 2018; Arroyo-Correa et al. 2019; Switzer et al. 2019). At the forefront of this is the inclusion of a biotremology perspective with its solid theoretical grounding and proven experimental methodology. In this context, studies of buzz pollination have excellent potential to contribute more broadly to other nascent fields, including mechanical ecology, the interface between mechanics and ecology at the organismal level (Bauer et al. 2020). Accordingly, as we conclude this chapter, we propose some future avenues of inquiry that are critical for expanding knowledge of buzz pollination, and that we hope will be enthusiastically embraced by researchers who study this unique biotic interaction.
First, there need to be more studies that address evolutionary hypotheses on the adaptive basis of buzz pollination for both bees and flowers. Specifically, we would like to see experimental tests estimating the fitness consequences of bee and floral traits under natural conditions. This may include examining the co-evolution of plant-pollinator characteristics (Solis-Montero and Vallejo-Marin 2017) or performing comparative analyses within a phylogenetic context to explore the adaptive significance of bee behaviour and poricidal floral morphologies (Cardinal et al. 2018). Second, new knowledge that builds on the groundwork previously established on the biomechanics of buzz pollination will be critical in broadening our understanding of the physical and behavioural factors governing the release of pollen from anthers, arguing that the vibrations induced in the flower are a complex of different wave forms (bending, Rayleigh, longitudinal, pressure) that could only be studied by a combination of 3-D (three dimensional)-measurements and computer simulations based on these measurements. Only then will we have a chance to understand how the bee-induced vibrations in the flower in turn affect the bee’s behaviour (see Sect. 11.2.4)—a really new research avenue in buzz pollination research! Experimental techniques already developed by biotremology researchers that integrate different recording methods (high-speed videography and laser vibrometry) and use computer simulation modelling (see Mhatre et al. 2018) offer an unparalleled approach for linking bee behaviour and vibration production with anther mechanical responses. Third, we call for more large-scale ecological studies linking the presence of sonicating bees to plant community structure. A key gap in current knowledge of buzz pollination concerns the role bees have played in driving the structure and assemblage of buzz pollinated plant communities. Although some studies have examined how bees affect the distribution of a single buzz pollinated species (Larson and Scheme 1999a, b), scaling up to the community level has rarely been addressed (Mesquita-Neto et al. 2017). Pollinators are known to filter plant community composition as a result of preferences for certain floral traits (Pellissier et al. 2012), and this may also occur in the buzz-pollination syndrome since many floral traits are hypothesised to be the product of selection resulting from the action of sonicating bees (Dulberger et al. 1994; Marazzi et al. 2007; Vallejo-Marín et al. 2010). Such community-level investigations may be particularly timely, as recent declines in bee abundances observed around the globe (Colla and Packer 2008; Cameron et al. 2010; Goulson et al. 2015) may have drastic consequences for buzz-pollinated plant communities.
References
Arceo-Gómez G, Martinez ML, Parra-Tabla V, Garcıa-Franco JG (2011) Anther and stigma morphology in mirror-image flowers of Chamaecrista chamaecristoides (Fabaceae): Implications for buzz pollination. Plant Biol 13(Suppl 1):19–24
Arroyo-Correa B, Beattie C, Vallejo-Marín M (2019) Bee and floral traits affect the characteristics of the vibrations experienced by flowers during buzz-pollination. J Exp Biol. 222:jeb198176. https://doi.org/10.1242/jeb.198176
Bauer U, Poppinga S, Müller UK (2020) Mechanical ecology--Taking biomechanics to the field. Integr Comp Biol icaa018. https://doi.org/10.1093/icb/icaa018
Bell PD (1980) Transmission of vibrations along plant stems: Implications for insect communication. Journal of the New York Entomological Society 88:210–216
Bennet-Clark HC (1989) Songs and the physics of sound production. In: Huber F, Moore TE, Loher W (eds) Cricket Behavior and Neurobiology. Cornell University Press, Ithaca, London, pp 227–261
Bennet-Clark HC (1998) Size and scale effects as constraints in insect sound communication. Philos Trans R Soc Lond B 353:407–419
Buchmann SL (1983) Buzz pollination in angiosperms. In: Jones CE, Little RJ (eds) Handbook of experimental pollination biology. Van Nostrand Reinhold Company, New York, pp 73–113
Buchmann SL (1985) Bees use vibration to aid pollen collection from non-poricidal flowers. J Kansas Entomol Soc 58:517–525
Buchmann SL, Cane JH (1989) Bees assess pollen returns while sonicating Solanum flowers. Oecologia 81:289–294
Buchmann SL, Hurley JP (1978) Biophysical model for buzz pollination in Angiosperms. J Theor Biol 72:639–657
Buchmann SL, Jones CE, Colin LJ (1978) Vibratile pollination of Solanum douglasii and S. xanti (Solanaceae) in Southern California. Wasmann Journal of Biology 35:1–25
Burkart A, Lunau K, Schlindwein C (2011) Comparative bioacoustical studies on flight and buzzing of neotropical bees. Journal of Pollination Ecology 6:118–124
Cameron SA, Lozier JD, Strange JP, Koch JB, Cordes N, Solter LF, Griswold TL (2010) Patterns of widespread decline in North American bumble bees. P Natl Acad Sci USA 108:662–667
Cardinal S, Buchmann SL, Russell AL (2018) The evolution of floral sonication, a pollen foraging behavior used by bees (Anthophila). Evolution 72:590–600. https://doi.org/10.1111/evo.13446
Casas J, Magal C, Sueur J (2007) Dispersive and non-dispersive waves through plants: Implications for arthropod vibratory communication. P Roy Soc Lond B 274:1087–1092
Cocroft RB (1996) Insect vibrational defence signals. Nature 382:679–680
Cocroft RB, De Luca PA (2006) Size-frequency relationships in insect vibrational signals. In: Drosopolous S, Claridge MF (eds) Insect Sounds and Communication: Physiology, behaviour, ecology and evolution. CRC Press, Boca Raton, pp 109–120
Cocroft RB, Rodrìguez RL (2005) The behavioral ecology of insect vibrational communication. BioScience 55:323–334
Cocroft RB, Gogala M, Hill PSM, Wessel A (2014a) Fostering research progress in a rapidly growing field. In: Cocroft RB, Gogala M, Hill PSM, Wessel A (eds) Studying Vibrational Communication, vol 3. Springer-Verlag, Berlin, pp 3–12
Cocroft RB, Hamel J, Su Q, Gibson J (2014b) Vibrational playback experiments: Challenges and solutions. In: Cocroft RB, Gogala M, Hill PSM, Wessel A (eds) Studying Vibrational Communication, vol 3. Springer-Verlag, Berlin, pp 249–274
Čokl A, Virant-Doberlet M (2003) Communication with substrate-borne signals in small plant-dwelling insects. Annu Rev. Entomol 48:29–50
Čokl A, Virant-Doberlet M, Stritih N (2000) The structure and function of songs emitted by southern green stink bugs from Brazil, Florida, Italy and Slovenia. Physiol Entomol 25:196–205
Čokl A, Presern J, Virant-Doberlet M, Bagwell GJ, Millar JG (2004) Vibratory signals of the harlequin bug and their transmission through plants. Physiol Entomol 29:372–380
Čokl A, Zorovic M, Zunic A, Virant-Doberlet M (2005) Tuning of host plants with vibratory songs of Nezara viridula L (Heteroptera: Pentatomidae). J Exp Biol 208:1481–1488
Colla SR, Packer L (2008) Evidence for decline in eastern North American bumblebees (Hymenoptera: Apidae), with special focus on Bombus affinis Cresson. Biodiversity and Conservation 17:1379–1391
Corbet SA, Huang SQ (2014) Buzz pollination in eight bumblebee-pollinated Pedicularis species: Does it involve vibration-induced triboelectric charging of pollen grains? Ann Bot 114:1665–1674. https://doi.org/10.1093/aob/mcu195
Corbet SA, Chapman H, Saville N (1988) Vibratory pollen collection and flower form: Bumble-bees on Actinidia, Symphytum, Borago and Polygonatum. Funct Ecol 2:147–155
De Luca PA (2015) Mass correlates with increased mating success for older but not younger males in thornbug treehoppers. Ann Entomol Soc Am 108:222–228
De Luca PA, Cocroft RB (2009) Age-related changes in an insect mating signal have no effect on female choice. Behav Ecol Sociobiol 63:1787–1798
De Luca PA, Morris GK (1998) Courtship communication in meadow katydids: Female preference for large male vibrations. Behaviour 135:777–794
De Luca PA, Vallejo-Marín M (2013) What’s the buzz about? The ecology and evolutionary significance of buzz-pollination. Curr Opin Plant Biol 16:429–435
De Luca PA, Bussière LF, Souto-Vilaros D, Goulson D, Mason AC, Vallejo-Marín M (2013) Variability in bumblebee pollination buzzes affects the quantity of pollen released from flowers. Oecologia 172:805–816
De Luca PA, Cox DA, Vallejo-Marín M (2014) Comparison of pollination and defensive buzzes in bumblebees indicates species-specific and context-dependent vibrations. Naturwissenschaften 101:331–338
De Luca PA, Giebink N, Mason AC, Papaj DR, Buchmann SL (2018) How well do acoustic recordings characterize properties of bee (Anthophila) floral sonication vibrations? Bioacoustics. https://doi.org/10.1080/09524622.2018.1511474
De Luca PA, Buchmann SL, Galen C, Mason AC, Vallejo-Marín M (2019) Does body size predict the buzz-pollination frequencies used by bees? Ecol Evol 9:4875–4887
Dierkes S, Barth FG (1995) Mechanism of signal production in the vibratory communication of the wandering spider Cupiennius getazi (Arachnida, Araneae). J Comp Physiol A 176:31–44
Dulberger R, Smith MB, Bawa KS (1994) The stigmatic orifice in Cassia, Senna, and Chamaecrista (Caesalpinaceae): Morphological variation, function during pollination, and possible adaptive significance. Am J Bot 81:1390–1396
Eberhard MJB, Metze D, Küpper SC (2019) Causes of variability in male vibratory signals and the role of female choice in Mantophasmatodea. Behav Process 166:103907
Elias DO, Lee N, Hebets EA, Mason AC (2006) Seismic signal production in a wolf spider: Parallel versus serial multi-component signals. J Exp Biol 209:1074–1084
Endler JA (2014) Foreward: The emerging field of tremology. In: Cocroft RB, Gogala M, Hill PSM, Wessel A (eds) Studying Vibrational Communication. Springer, Berlin Heidelberg, pp vii–x
Gibson JS, Cocroft RB (2018) Vibration-guided mate searching in treehoppers: directional accuracy and sampling strategies in a complex sensory environment. J Exp Biol 221(6):jeb175083
Goulson D, Nicholls E, Botias C, Rotheray EL (2015) Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347:1–16
Harder LD, Barclay MR (1994) The functional significance of poricial anthers and buzz pollination: controlled pollen removal from Dodecatheon. Funct Ecol 8:509–517
Hebets EA, Elias DO, Mason AC, Miller GL, Stratton GE (2008) Substrate-dependent signalling success in the wolf spider, Schizocosa retrorsa. Anim Behav 75:605–615
Henry CS (1980) The importance of low-frequency, substrate-borne sounds in lacewing communication (Neuroptera: Chrysopidae). Ann Entomol Soc Am 73:617–621
Henry CS, Martinez-Wells ML (2006) Testing the ability of males and females to respond to altered songs in the duetting green lacewing, Chrysoperla plorabunda (Neuroptera: Chrysopidae). Behav Ecol Sociobiol 61:39–51
Henry CS, Wells MLM, Simon CM (1999) Convergent evolution of courtship songs among cryptic species of the Carnea group of green lacewings (Neuroptera: Chrysopidae: Chrysoperla). Evolution 53:1165–1179
Hill PSM (2008) Vibrational Communication in Animals. Harvard University Press, Cambridge
Hill PSM, Wessel A (2016) Biotremology. Curr Biol 26:R187–R191
Hrncir M, Schmidt VM, Schorkopf DLP, Jarau S, Zucchi R, Barth FG (2006) Vibrating the food receivers: A direct way of signal transmission in stingless bees (Melipona seminigra). J Comp Physiol A 192:879–887
Hunt RE, Morton TL (2001) Regulation of chorusing in the vibrational communication system of the leafhopper Graminella nigrifrons. Am Zool 41:1222–1228
Hunt RE, Nault LR (1991) Roles of interplant movement, acoustic communication, and phonotaxis in mate-location behavior of the leafhopper Graminella nigrifrons. Behav Ecol Sociobiol 28:315–320
Josephson RK, Malamud JG, Stokes DR (2000) Asynchronous muscle: A primer. J Exp Biol 203:2713–2722
Kilpinen O, Storm J (1997) Biophysics of the subgenual organ of the honeybee, Apis mellifera. J Comp Physiol A 181:309–318
King MJ (1993) Buzz foraging mechanism in bumble bees. J Apic Res 32:41–49
King MJ, Buchmann SL (1995) Bumble bee-initiated vibration release mechanism of Rhododendron pollen. Am J Bot 82:1407–1411
King MJ, Buchmann SL (1996) Sonication dispensing of pollen from Solanum laciniatum flowers. Funct Ecol 10:449–456
King MJ, Buchmann SL (2003) Floral sonication by bees: Mesosomal vibration by Bombus and Xylocopa, but not Apis (Hymenoptera: Apidae), ejects pollen from poricidal anthers. J Kansas Entomol Soc 76:295–305
King MJ, Lengoc L (1993) Vibratory pollen collection dynamics. Transactions of the American Society of Agricultural Engineers 36:135–140
King MJ, Buchmann SL, Spangler HG (1996) Activity of asynchronous flight muscle from two bee families during sonication (buzzing). J Exp Biol 199:2317–2321
Kumar H, Saxena KN (1985) Certain environmental factors influencing the acoustic communication in the sexual behaviour of the leafhopper Amrasca devastans (Distant) (Homoptera: Cicadellidae). Appl Entomol Zool 20:199–209
Larson BM, Scheme B (1999a) The ecology of pollen limitation in buzz-pollinated Rhexia virginica (Melastomataceae). J Ecol 87:371–381
Larson BM, Scheme B (1999b) The pollination ecology of buzz-pollinated Rhexia virginica (Melastomataceae). Am J Bot 86:502–511
Macior LW (1964) Experimental study of floral ecology of Dodecatheon meadia. Am J Bot 51:96–108
Macior LW (1968) Pollination adaptation in Pedicularis groenlandica. Am J Bot 55:927–932
Marazzi B, Conti E, Endress PK (2007) Diversity in anthers and stigmas in the buzz-pollinated genus Senna (Leguminosae, Cassiinae). Int J Plant Sci 168:371–391
McNett GD, Cocroft RB (2008) Host shifts favor vibrational signal divergence in Enchenopa binotata treehoppers. Behav Ecol 19:650–656
McNett GD, Miles RN, Homentcovschi D, Cocroft RB (2006) A method for two-dimensional characterization of animal vibrational signals transmitted along plant stems. J Comp Physiol A 192:1245–1251
McNett GD, Luan LH, Cocroft RB (2010) Wind-induced noise alters signaler and receiver behavior in vibrational communication. Behav Ecol Sociobiol 64:2043–2051
McVean A, Field LH (1996) Communication by substratum vibration in the New Zealand tree weta, Hemideina femorata (Stenopelmatidae: Orthoptera). J Zool 239:101–122
Mesquita-Neto JN, Costa BKP, Schlindwein C (2017) Heteranthery as a solution to the demand for pollen as food and for pollination—legitimate flower visitors reject flowers without feeding anthers. Plant Biol 19:942–950
Mhatre N, Sivalinghem S, Mason AC (2018) Posture controls mechanical tuning in the black widow spider mechanosensory system. bioRxiv. https://doi.org/10.1101/123456
Michael SCJ, Appel HA, Cocroft RB (2019) Methods for replicating leaf vibrations induced by insect herbivores. In: Gassmann W (ed) Plant innate immunity: methods and protocols. Humana Press, New York, pp 141–158
Michelsen A, Nocke H (1974) Biophysical aspects of sound communication in insects. Adv Insect Physiol 10:247–296
Michelsen A, Fink F, Gogala M, Traue D (1982) Plants as transmission channels for insect vibrational songs. Behav Ecol Sociobiol 11:269–281
Miklas N, Stritih N, Čokl A, Virant-Doberlet M, Renou M (2001) The influence of substrate on male responsiveness to the female calling song in Nezara viridula. J Insect Behav 14:313–332
Miles CI, Allison BE, Losinger MJ, Su QT, Miles RN (2017) Motor and mechanical bases of the courtship call of the male treehopper Umbonia crassicornis. J Exp Biol 220:1915–1924
Miranda X (2006) Substrate-borne signal repertoire and courtship jamming by adults of Ennya chrysura (Hemiptera: Membracidae). Ann Entomol Soc Am 99:374–386
Mitomi M, Ichikawaa T, Okamoto H (1984) Morphology of the vibration-producing organ in adult rice brown planthopper, Nilaparvata lugens (STAL): Homoptera: Delphacidae. Appl Entomol Zool 19:407–417
Moreira GRP (1993) Reproductive biology of the stream-dwelling stonefly, Agnetina capitata (Pictet) (Plecoptera: Perlidae). Dissertation, Cornell University, Ithaca, NY
Morgan T, Whitehorn PR, Lye GC, Vallejo-Marín M (2016) Floral sonication is an innate behaviour in bumblebees that can be fine-tuned with experience in manipulating flowers. J Insect Behav 29:233–241
Morris GK (1980) Calling display and mating behaviour of Copiphora rhinoceros Pictet (Orthoptera: Tettigoniidae). Anim Behav 28:42–51
Nunes-Silva P, Hrncir M, Shipp L, Kevan P, Imperatriz-Fonseca VL (2013) The behaviour of Bombus impatiens (Apidae, Bombini) on tomato (Lycopersicon esculentum Mill., Solanaceae) flowers: Pollination and reward perception. J Pollinat Ecol 11:33–40
Ossiannilsson F (1949) Insect drummers. A study on the morphology and function of the sound-producing organ of Swedish Homoptera Auchenorrhyncha. Opuscula Entomol Suppl X:1–146
Papaj DR, Buchmann SL, Russell AL (2017) Division of labor of anthers in heterantherous plants: flexibility of bee pollen collection behavior may serve to keep plants honest. Arthropod Plant Interactions 11:307–315
Pellissier L, Alvarez N, Guisan A (2012) Pollinators as drivers of plant distributions and assemblage into communities. In: Patiny S (ed) Evolution of plant-pollinator relationships. Cambridge University Press, New York, pp 392–413
Pringle JWS (1949) The excitation and contraction of the flight muscles of insects. J Physiol (Lond) 108:226–232
Pritchard DJ, Vallejo-Marín M (2020) Floral vibrations by buzz-pollinating bees achieve higher frequency, velocity and acceleration than flight and defence vibrations. J Exp Biol. https://doi.org/10.1242/jeb.220541
Rebar D, Hoebel G, Rodriguez RL (2012) Vibrational playback by means of airborne stimuli. J Exp Biol 215:3513–3518
Rodrigues EV, Riguette JR, Pereira HRC, Tesch JA, Silva AG (2018) An affordable apparatus for fine-controlled emulation of buzzing frequencies of bees for the testing hypothesis in buzz interactions. Ecol Evol 8:7667–7672
Rodríguez RL, Cocroft RB (2006) Divergence in female duetting signals in the Enchenopa binotata species complex of treehoppers (Hemiptera: Membracidae). Ethology 112:1231–1238
Rodríguez RL, Sullivan LE, Cocroft RB (2004) Vibrational communication and reproductive isolation in the Enchenopa binotata species complex of treehoppers (Hemiptera: Membracidae). Evolution 58:571–578
Rodríguez RL, Ramaswamy K, Cocroft RB (2006) Evidence that female preferences have shaped male signal evolution in a clade of specialized plant-feeding insects. Proc R Soc Lond B 273:2585–2593
Rosi-Denadai CA, Araújo PCS, de Oliveira Campos LA, Cosme L Jr, Guedes RNC (2018) Buzz-pollination in neotropical bees: genus-dependent frequencies and lack of optimal frequency for pollen release. Insect Sci. https://doi.org/10.1111/1744-7917.12602
Rovner JS, Barth FG (1981) Vibratory communication through living plants by a tropical wandering spider. Science 214:464–466
Russell AL, Golden RE, Leonard AE, Papaj DR (2015) Bees learn preferences for plant species that offer only pollen as a reward. Behav Ecol 27:731–740
Russell AL, Leonard AS, Gillette HD, Papaj DR (2016) Concealed floral rewards and the role of experience in floral sonication by bees. Anim Behav 120:83–91
Russell AL, Buchmann SL, Papaj DR (2017) How a generalist bee achieves high efficiency of pollen collection on diverse floral resources. Behav Ecol 28:991–1003. https://doi.org/10.1093/beheco/arx058
Solis-Montero L, Vallejo-Marin M (2017) Does the morphological fit between flowers and pollinators affect pollen deposition? An experimental test in a buzz-pollinated species with anther dimorphism. Ecol Evol 7:2706–2715. https://doi.org/10.1002/ece3.2897
Speaks CE (1999) Introduction to sound: acoustics for the hearing and speech sciences. Singular Publishing Group, San Diego CA
Sueur J (2018) Sound analysis and synthesis with R. Springer, Cham, Switzerland
Sullivan-Beckers LE, Hebets EA (2014) Tactical adjustment of signalling leads to increased mating success and survival. Anim Behav 93:11–17
Switzer C, Combes S (2017) Bumblebee sonication behavior changes with plant species and environmental conditions. Apidologie 48:223–233. https://doi.org/10.1007/s13592-016-0467-1
Switzer C, Hogendoorn K, Ravi S, Combes S (2016) Shakers and head bangers: differences in sonication behavior between Australian Amegilla murrayensis (blue-banded bees) and North American Bombus impatiens (bumblebees). Arthropod Plant Interactions 10:1–8
Switzer C, Russell AL, Papaj DR, Combes SA, Hopkins R (2019) Sonicating bees demonstrate flexible pollen extraction without instrumental learning. Curr Zool. https://doi.org/10.1093/cz/zoz013
Teppner H (2018) The first records of vibratory pollen-collection by bees. Phyton Annales Rei Botanicae 57:135–141
Tercel MP, Veronesi F, Pope TW (2018) Phylogenetic clustering of wingbeat frequency and flight-associated morphometrics across insect orders. Physiol Entomol 43:149–157
Vallejo-Marín M (2019) Buzz pollination: studying bee vibrations on flowers. New Phytol. https://doi.org/10.1111/nph.15666
Vallejo-Marín M, Da Silva EM, Sargent RD, Scheme B (2010) Trait correlates and functional signficance of heteranthery in flowering plants. New Phytol 188:418–425
Virant-Doberlet M, Čokl A (2004) Vibrational communication in insects. Neotrop Entomol 33:121–134
Wells MM, Henry CS (1992) The role of courtship songs in reproductive isolation among populations of green lacewings of the genus Chrysoperla (Neuroptera: Chrysopidae). Evolution 46:31–42
Zeigler DD, Stewart KW (1977) Drumming behavior of eleven nearctic stonefly (Plecoptera) species. Ann Entomol Soc Am 70:495–505
Zeigler DD, Stewart KW (1985) Age effects on drumming behavior of Pteronarcella badia (Plecoptera) males. Entomol News 96:157–160
Acknowledgments
We thank Peggy Hill for inviting us to contribute to this chapter about buzz pollination and its important links to the discipline of biotremology. PAD appreciates the generosity of the University of The Bahamas for supporting this work. MVM was supported by a Leverhulme Trust Research Grant (RPG-2018-235). Kristen Brochu read an earlier version of this work and provided helpful comments that improved its clarity. This chapter was also greatly improved from the insights and suggestions offered by Peggy Hill. We dedicate this chapter to Dr. Reginald B. Cocroft, whose pioneering work examining vibrational communication in treehoppers inspired a new generation of biotremology researchers, including us. Thank you, Rex.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 Springer Nature Switzerland AG
About this chapter

Cite this chapter
De Luca, P.A., Vallejo-Marín, M. (2022). Blooms and Buzzing Bees: Bridging Buzz Pollination and Biotremology. In: Hill, P.S.M., Mazzoni, V., Stritih-Peljhan, N., Virant-Doberlet, M., Wessel, A. (eds) Biotremology: Physiology, Ecology, and Evolution. Animal Signals and Communication, vol 8. Springer, Cham. https://doi.org/10.1007/978-3-030-97419-0_11
Download citation
DOI: https://doi.org/10.1007/978-3-030-97419-0_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-97418-3
Online ISBN: 978-3-030-97419-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)