
Abstract
The kidneys are developed from the intermediate mesoderm of the embryo. They are important for osmoregulation, regulation of acid-base balance, reabsorption of nutrients, and excretion of metabolites. In fish, the kidneys also serve as a hematopoietic, lymphoid and endocrine organ for the generation of red blood cells, the development of lymphocytes, and the production of hormones (e.g., glucocorticoids, catecholamines, and thyroid hormones). In humans and all animals, kidneys play a vital role in the metabolism and reabsorption of amino acids (AAs) and glucose. Specifically, this organ contributes to glucose synthesis from AAs, lactate and pyruvate via the gluconeogenesis pathway; regulates acid-base balance via inter-organ metabolism of glutamine; and synthesizes arginine, tyrosine, and glycine, respectively, from citrulline, phenylalanine, and 4-hydroxyproline. In mammals and birds, kidneys participate in creatine synthesis. Renal dysfunction adversely alters the concentrations of AAs in blood, while promoting muscle protein breakdown, inflammation, mitochondrial abnormalities, defects in the immune response, and cardiovascular diseases. Moderation of dietary AA intake has a protective and therapeutic effect on chronic kidney disease. Understanding the functions and metabolism of AAs in kidneys is essential for maintaining whole-body homeostasis, improving health and well-being, and preventing or treating renal metabolic diseases in humans and farm animals (including swine, poultry, ruminants, fish and shrimp).
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
5.1 Introduction
The main function of the kidney is to maintain the homeostasis of water, minerals and metabolites as well as acid-base balance, osmolality, and blood pressure in the body. This is achieved through filtering the blood supplied from the renal arteries, reabsorbing nutrients into the blood circulation, and excreting metabolites (e.g., ammonia, urea, homocysteine, ketone bodies, and methylarginines). The rate of blood flow through the kidneys is particularly high. For example, in humans, the kidneys receive about 20% of the blood pumped out of the heart and filter about 1.25 L of blood per minute (Kierszenbaum and Tres 2015). Kidneys also produce hormones that affect the function of other organs. As a result, well-functioning kidneys are essential to health and well-being. However, many individuals (e.g., 11.5% of the adult population in the United States) have chronic kidney disease and are at increased risk for cardiovascular events and progression to kidney failure (Tangri et al. 2011). According to the World Health Organization (WHO), globally 864,226 deaths (or 1.5% of deaths) were attributable to chronic kidney disease in 2012. Ranked the fourteenth in the list of leading causes of death, this disease accounts for 122 deaths per 1 million people (Webster et al. 2017).
Maintaining health and preventing disease outbreaks is vital to the economy and safety in animal production. This is because farm animals (including swine, poultry, cows, sheep, goats, fish, and shrimp) usually face numerous stresses, including high-density rearing, imbalance in nutrient intakes, heavy metal and non-metal containments, heat or cold environments, air and water pollution, and challenges by infectious agents (e.g., parasites, bacteria, fungi and viruses). Behavioral and physiological alterations will occur when affected animals cannot mount a successful response to one or more of these stresses (Blokhuis et al. 1998). For example, heat stress or metal toxicity can induce oxidative damage due to enhanced production of reactive oxygen species (ROS), mitochondrial dysfunction, and metabolic disorder, thereby compromising renal function (Pandey and Madhuri 2014; Belhadj Slimen et al. 2016). In fish, which lack bone marrow and lymph nodes, the kidneys serve as the hematopoietic organ and are a major lymphoid organ (Kum and Sekkin 2011). Renal dysfunction adversely affects immunity and increases susceptibility to disease, while decreasing feed intake and growth, thereby hindering animal production (Blokhuis et al. 1998; Anderson et al. 2011).
Most of amino acids (AAs) are catabolized by the kidneys via numerous pathways (including asparaginase, glutamate dehydrogenase, glutaminase, transaminases, and D-AA oxidases), and that filtered AAs are almost completely reabsorbed by the proximal convoluted tubule into the blood in normal physiological states (Levillain et al. 1997; Wu 2013a, b). The kidneys also play a vital role in the biosynthesis of some AAs and their derivatives. As a result, renal dysfunction will induce the deficiency of these AAs and their derivatives. Moreover, understanding the functions and metabolism of AAs in the kidneys is important for preventing and treating chronic renal diseases in both humans and farm animals. This review highlights the development and functions of the kidneys in different animals, as well as AA metabolism in this organ and its relationship with renal diseases.
5.2 Kidney Development and Structure
In all vertebrate species, the kidney originates from the intermediate mesoderm that is called the nephrogenic mesoderm (Cullen-McEwen et al. 2016). The embryonic development of the kidney is complex in that two or three different kidneys (depending on species) are formed in temporal and spatial sequences. In amniotes (birds, mammals, and reptiles), nephrogenesis begins at the embryonic stage and completes at birth. This process occurs through a series of successive phases: archinephros, pronephros, mesonephros, and metanephros (Cullen-McEwen et al. 2016). Archinephros occurs in the embryos as the simplest kind of excretory organ, which is nonfunctional in humans and other mammals. Then, three types of a more advanced kidney develop from the embryonic archinephros: the pronephros from its anterior section, the mesonephros from its middle section, and the metanephros from its hind section. The pronephros and mesonephros are generally transient embryonic kidneys that subsequently degenerate and have little or no function in adult mammals. However, the mesonephros is the functional kidney in fish and amphibians (Seely 2017). Generally, the metanephros develops when an outgrowth of the primary nephric duct (the ureteric bud or metanephric diverticulum) extends into the surrounding metanephric mesenchyme (Dressler 2006; Seely 2017). The metanephros is the adult kidney or the functional kidney in humans and other amniotes (Cullen-McEwen et al. 2016). The functional kidney has an extensively branched collecting duct system and a large number of nephrons (Cullen-McEwen et al. 2016; Denic et al. 2016). During postnatal growth, mammals increase their renal performance via increases in nephron size and glomerular filtration rate (Davidson 2014).
In contrast to mammals, fish form the pronephros and mesonephros, with the latter being the permanent adult kidney (Davidson 2014). Generally, fish embryos develop externally and are therefore exposed to environmental pressures from water and salts, and the pronephric kidneys play a vital role in osmoregulation. In zebrafish, the functional larval pronephros consists of two nephrons with glomeruli fused at the embryo midline just ventral to the dorsal aorta (Drummond 2005). At the juvenile stage, the mesonephros kidney is formed, which consists of hundreds of nephrons that branch distally (Davidson 2014). Unlike the mammalian metanephric kidneys, the mesonephric kidneys of fish continue to add new nephrons as their body mass increases during their lives (Davidson 2014; Upadhyay and Silverstein 2014).
5.3 Functions of the Kidneys
The nephron is the functional unit of the kidney, and is composed of a renal corpuscle (a glomerulus and glomerular capsule known as Bowman’s capsule) and renal tubules (including the proximal convoluted tubule, the loop of Henle, and the distal convoluted tubule). The glomerulus filters the blood into the renal tubule, and the glomerular filtration rate (GFR) is an indicator of overall kidney function. The GFR is defined as the total amount of fluid filtered through all of the nephrons per unit of time (Levey et al. 2015). Filtration is the process by which cells and large proteins are retained while substances of small molecular weights are filtered from the blood to make an ultrafiltrate fluid. Most of molecules (such as water, glucose, and AAs) in the ultrafiltrate are reabsorbed from the renal tubule into the peritubular capillary and blood circulation. In a healthy adult human, reabsorption by renal tubules recovers about 70 g AAs per day (Young 1991). Moreover, the kidneys excrete a variety of metabolites (such as ammonia, urea, uric acid, methylarginines, homocysteine, and ketone bodies) into urine (McNeal et al. 2018). For example, in humans, about 180 L of ultrafiltrate fluid passes into the renal tubules per day. Most of the fluid is reabsorbed by the tubular cells into the blood circulation, and only about 1.5 L of the fluid is excreted as urine (Kierszenbaum and Tres 2015). Due to their functions in excretion, reabsorption, and filtration, the kidneys play a vital role in maintaining whole-body homeostasis, such as acid-base balance, electrolyte concentrations in plasma, extracellular fluid volume, and blood pressure. The kidneys accomplish these homeostatic functions both independently and in concert with other organs, particularly the endocrine system. Various endocrine hormones coordinate the endocrine functions, including antidiuretic hormone, thyroid hormone, adrenal cortical hormone, renin, angiotensin II, aldosterone, glucocorticoids, mineralocorticoids, prolactin, prostaglandins, and atrial natriuretic peptide, gastrin, among others (McDonald et al. 1976; Afsar et al. 2016; Ahmed and Ramesh 2016). For example, antidiuretic hormone is the hormone of paramount importance in the regulation of water excretion by the mammalian kidneys, but other hormones also influence renal excretion of water. In particular, aldosterone, which is produced by the cortex of the adrenal gland, stimulates the reabsorption of water by renal tubules along with Na+, thereby increasing blood volume and decreasing urine volume. Furthermore, glucagon-like peptide-1, which is excreted by endocrine cells of the small intestine, can increase the GFR, renal plasma flow, urine output, and the excretion of Na+, K+, and HCO3− (Afsar et al. 2016). In postnatal mammals and birds, another important function of the kidneys is the regulation of erythropoiesis (the production of erythrocytes) by the bone marrow. Although the kidney is not the site of erythropoiesis in these species, it releases a hormone called erythropoietin in response to cellular hypoxia. Through erythropoietin receptor-mediated signaling cascades that involve the Janus kinase-2 and its downstream proteins, phosphoinositide-3 kinase (PI3K)/protein kinase A, the extracellular signal-regulated kinase (ERK1/2), p38 mitogen-activated protein kinase (MAPK), and signal transducer and activator of transcription 5 (STAT5), erythropoietin stimulates the bone marrow to produce red blood cells (Watowich 2011).
In amniotes, the kidneys efficiently reabsorb water from the renal tubules into blood, and therefore a relatively small amount of water reaches the bladder (Mahasen 2016). Compared with terrestrial animals, fish have a special need to maintain their internal environment because of the constant exposure to external water and substances. Freshwater fish have a higher concentration of salts in blood than the external environment, contributing to the net osmotic gain of water and diffusional loss of salts across the gills. This conundrum is solved through several strategies, including excretion of relatively dilute urine, active uptake of salts across the gill, and possibly ingestion of more salts in the food (Evans 2002). In freshwater fish species, the kidneys generally remove, as dilute urine, a considerable amount of water that passively enters the body via the gill. This dilute urine is almost completely composed of water. Freshwater fish species have a good size of the renal corpuscle and hence a high water output. On the contrary, in marine fish, the renal corpuscle is small or absent and the renal tubule is short, thereby minimizing the glomerular infiltration of blood and conserving water as well as minimizing the reabsorption of salts by the renal tubules and producing a urine with concentrated salts (Mahasen 2016). In marine fish, excessive salts are removed largely through the kidneys. Of particular note, cartilaginous fishes (sharks, skates, rays, and chimaeras) have an ability to adapt a high-salinity marine environment through a unique urea-based osmoregulation strategy, as their kidneys reabsorb nearly all of the filtered urea in the primary urine (Hyodo et al. 2014).
In fish, each kidney contains two segments: the anterior (head) and the posterior (trunk). The filtration and urine-forming functions are carried out by the posterior kidney, as noted previously. In contrast to mammals and birds, fish lack bone marrow, lymph nodes, and adrenal gland but possess the head kidney to serve as a hematopoietic, lymphoid and endocrine organ for the production of erythrocytes (red blood cells), cytokines, antibodies, and some hormones (Shoemaker et al. 2015). Thus, hematopoiesis occurs in the head kidney in fish, instead of the bone marrow in postnatal mammals and birds. In addition, the head kidney plays an important role in both innate and adaptive immunities in fish (see Sect. 5.7). Furthermore, the head kidney of fish produces glucocorticoids from cholesterol, as well as catecholamines and thyroid hormones from tyrosine. Thus, in fish, the kidney has a high metabolic rate (including AA catabolism and ATP production) to support its integrated hematopoietic, immune, and endocrine functions.
5.4 Amino Acid Transporters in the Kidneys
Tissues in the kidney receive AAs that are supplied from the arterial blood. In addition, epithelial cells of the renal tubule reabsorb free AAs from its lumen (the AAs that are filtered through the glomerulus) into the blood. These processes require specific AA transporters and play an important role in maintaining AA homeostasis in plasma (Verrey et al. 2005, 2009). Generally, AAs are transported across the plasma membrane via: (1) simple diffusion (passive and nonsaturable), (2) Na+-independent systems (facilitated diffusion), and (3) Na+-dependent systems (active transport). Some Na+-dependent transport proteins can use Li+ instead of Na+, and a few of AA transport proteins are H+-driven. In the kidney, AA transporters are highly expressed in the luminal brush border membrane of the proximal segments of the renal tubule (Palacín et al. 1998; Verrey et al. 2009). On the basis of sequence similarity, AA transporters are grouped into solute carrier (SLC) families (Table 5.1). For example, B0AT1 (SLC6A19), XT2 (SLC6A18) and IMINO (SLC6A20) systems transport neutral AAs from the lumen of the renal proximal tubule into its epithelial cells (Bröer 2006, 2008; Verrey et al. 2009). Moreover, H+-dependent transporters, such as PAT1 (SLC36A1) and PAT2 (SLC36A2), have been identified in the kidney (Daniel et al. 2006). Besides, cationic AAs (e.g., arginine, ornithine and lysine) are taken up by cationic AA transporters in a Na+-independent manner. The cystine transporter consists of a catalytic subunit of the SLC7 family and a disulfide-linked accessory subunit referred to as a heavy chain called rBAT [related to neutral and basic (b0,+) AA transporter; SLC3A1; Verrey et al. 2009]. Mutations in the SLC3A1 gene result in cystinuria that is characterized by high concentrations of cystine (an AA with a very low solubility in water) in the urine, leading to the formation of cystine stones in the kidneys, ureter, and bladder. The transport of anionic AAs (e.g., glutamate and aspartate) requires the Na+-dependent transporters (EAAT1, EAAT2 and EAAT3). Competition of the transport of AAs by the cell occurs when they are structurally similar (e.g., among large neutral AAs, basic AAs, acidic AAs, small neutral AAs, or β-AAs).
5.5 Amino Acid Metabolism in the Kidneys
5.5.1 Gluconeogenesis
Gluconeogenesis is the metabolic pathway for the synthesis of glucose from non-glucose substrates, such as lactate, pyruvate, glycerol, and gluconeogenic AAs. A major function of gluconeogenesis is to provide the body with glucose in response to physiological needs. There are four rate-controlling reactions in gluconeogenesis that are catalyzed by pyruvate carboxylase, phosphoenolpyruvate carboxykinase (PEPCK), fructose-1,6-bisphosphatase (FBPase), and glucose-6-phosphatase (G6Pase). All of these enzymes are present in the kidneys of rats (Anderson and Stowring 1973), chickens (Shen and Mistry 1979; Watford et al. 1981), humans (Møller et al. 2000), and some fish species (Knox et al. 1980; Kirchner et al. 2008; Kumar et al. 2010). PEPCK and G6Pase are mainly expressed in the proximal tubules (Pollock 1989; Sun et al. 2002). Moreover, FBPase and PEPCK are co-localized in the kidney and liver, which contributed almost equally to glucose synthesis in fasting humans (Yánez et al. 2003). Similar to other animals, the gluconeogenic enzymes are expressed mainly in the liver and kidneys of fish species, and gluconeogenesis does not occur in skeletal muscle (Knox et al. 1980). In the kidneys of some species, whether AAs are the substrates for glucose synthesis depends on the intracellular location of PEPCK (Wu 2013a). As reported for chickens (Watford et al. 1981), the presence of PEPCK in the cytoplasm allows for the generation of NADH from glucogenic AAs and, therefore, the production of glucose from the AAs.
The mammalian kidneys synthesize glucose and release it into the blood circulation under various physiological conditions (Gerich 2010). In the post-absorptive state, renal gluconeogenesis (primarily from glutamine) contributes 20–25% of glucose production in adult humans (Stumvoll et al. 1999), and the kidneys release nearly the same amount of glucose into the circulation as the liver (Mitrakou 2011). In contrast, the contribution of alanine to gluconeogenesis occurs almost exclusively in the liver (Stumvoll et al. 1998). In the body, the production of new glucose molecules occurs mainly through gluconeogenesis in the kidneys and liver, and to a lesser extent via glycogenolysis in the liver and skeletal muscle (Stumvoll et al. 1997). In all animals, the kidney contains very little glycogen and, therefore, produces little glucose through glycogenolysis (Stumvoll et al. 1997; Gerich 2010).
Not all AAs are used for endogenous glucose synthesis (Wu 2013a). Generally, AAs are classified as “glucogenic” or “ketogenic” based on the type of intermediates that are formed from their metabolism. Glucogenic AAs are converted into either pyruvate or one of the intermediates in the Krebs cycle. Leucine and lysine are strictly ketogenic AAs because they are catabolized to acetyl CoA (Fig. 5.1). The kidneys synthesize glucose from many AAs, but glutamine, alanine, glutamate, aspartate, proline, ornithine and arginine are the main AA substrates for the renal gluconeogenesis (Krebs et al. 1963). A mixture of AAs, pyruvate, lactate, and glycerol at normal plasma concentrations can support renal gluconeogenesis at an initial rate of about 0.6 μmol/min per g of kidney (Bowman 1970). As noted previously, the kidney is the major organ for gluconeogenesis from AAs in birds (Watford et al. 1981) and contributes to about 30% of the glucose produced in the starved chickens (Tinker et al. 1986). Renal gluconeogenesis is species-dependent. For example, high expression levels of FBPase and PEPCK are present in the kidneys and liver of some fish species (e.g., cod, salmons and trout), and glucose is produced from AAs (e.g., alanine and glutamine) in their kidneys and liver (Moon and Foster 1995; NRC 2011). However, in many fish species, the kidney lacks PEPCK and is not capable of gluconeogenesis from any potential substrates (Moon and Foster 1995).

The ketogenic and glucogenic- amino acids metabolism in animals. Amino acids with red color: ketogenic only; Amino acids with green color: both glucogenic and ketogenic; Amino acids with black color: glucogenic only. G6Pase, glucose-6-phosphatase; PCL pyruvate carboxylase, PEPCK, phosphoenolpyruvate carboxykinase; PEP phosphoenolpyruvate, “----“denotes glucogenic reactions
Renal gluconeogenesis is regulated by some hormones. Insulin, growth hormone, cortisol, and catecholamines influence renal glucose release in the mammalian kidneys (Wirthensohn and Guder 1986; Schoolwerth et al. 1988). For example, expression of renal gluconeogenic genes in mice is inhibited by both insulin and glucose reabsorption via the inactivation of FoxO1 and PGC1α, respectively (Sasaki et al. 2017). The regulation of gluconeogenesis from glutamine by insulin and glucose also occurs in the kidneys of humans (Stumvoll et al. 1999). All of the metabolic pathways for renal glucose synthesis and whole-body homeostasis of glucose are altered in patients with diabetes mellitus (Gerich 2010). Patients with T2DM have an increased release of glucose into the circulation by the kidneys in the fasting and postprandial states, which impairs glucose homeostasis and leads to hyperglycemia (Triscari et al. 1979; Meyer et al. 1998). Epinephrine, norepinephrine, and prostaglandin E-2 (PGE2) also regulate gluconeogenesis in fish (Enes et al. 2009). Some fish species have poor regulation of gluconeogenesis in response to starch intake, which may result from relatively high concentrations of glucogenic AAs in the diets (NRC 2011).
5.5.2 Glutamine and Glutamate
Glutamine is the one of the most important free AAs in the blood of humans and numerous animal species, and plays key roles in diverse physiological processes, including the syntheses of DNA, RNA, protein, aminosugars, NAD, and glucose (Wu 2013a). In the kidney, glutamine catabolism is initiated via four enzymes: (1) phosphate-activated glutaminase, (2) glutamine transaminases K and L, (3) glutamine:fructose-6-phosphate transaminase, and (4) carbamoylphosphate synthase-II. Among these pathways, glutaminase contributes to most of glutamine degradation, which produces glutamate and NH4+ primarily in the proximal tubule. (Welbourne 1974). Glutamate is catabolized by glutamate dehydrogenase to NH4+ and α-ketoglutarate (α-KG). The latter enters the Krebs cycle for oxidation to CO2 and water. Overall, the complex oxidation of one mole of glutamine generates two moles of NH4+ and 2 mol of HCO3−. NH4+ is directly excreted into the lumen of the renal tubule through the apical NHE3 (sodium-hydrogen exchanger-3), whereas HCO3− is returned into the blood by crossing the basolateral membrane via the electrogenic sodium-coupled bicarbonate co-transporter (Weiner et al. 2015). In the kidneys, about 50% of NH4+ is reabsorbed into the blood primarily by the thick ascending limb of the loop of Henle via the apical membrane Na+-K+-2Cl− symporter (NKCC2) and NH4+ transporter and the basolateral membrane NHE4 (Fig. 5.2). Similar mechanisms are also present in fish species (Claiborne et al. 1982; Evans and Cameron 1986). Because of the renal reabsorption, only about 59% of the ammonia produced by the kidneys appears in the urine. Ammonia accounts for about 10% of total renal nitrogen excretion under basal conditions, but can increase substantially under a variety of clinical conditions such as acidosis, infections, manganese deficiency, urea cycle defects, and excessive intake of dietary protein or AAs (Wu 2013a).

Integrated overview of renal ammonia metabolism. Renal ammoniagenesis occurs primarily in the proximal tubule, involving glutamine uptake, glutamine metabolism forming ammonium and bicarbonate, and apical NH4+ (H+) secretion. Note: ammonia reabsorption in the thick ascending limb, involving apical NKCC2-mediated uptake results in medullary ammonia accumulation. Ammonia is secreted in the collecting duct via parallel H+ and NH3 secretion. α-KG, α-ketoglutarate
The reabsorption of bicarbonate (HCO3−) and excretion of hydrogen ions (H+) in the kidneys play an important role in maintaining the acid–base balance (Vercoutère et al. 2004). This can be achieved partly via: (1) the regulation of glutamine uptake through the apical Na+-dependent neutral AA transporter-1 and the basolateral Na+-coupled neutral AA transporter-3 (Weiner et al. 2015); and (2) expression of glutaminase. Under acidotic conditions, glutaminase activity, along with the uptake and catabolism of glutamine by the kidney, is greatly enhanced to generate NH3 for removing excess H+ as NH4+ (Wu 2013a). In rats, acidotic kidneys extract glutamine and produce NH3 at rates that are 4–5 times greater than those in nonacidotic kidneys (Welbourne 1974). An inhibition of glutamine synthetase contributes to an increase in intracellular ammonia concentration in the kidneys of acidotic rats (Hems 1972). In contrast, in response to alkalosis, the kidneys excrete more HCO3− by decreasing H+ secretion from the tubular epithelial cells as well as the rates of glutamine catabolism and ammonium excretion (Fig. 5.2).
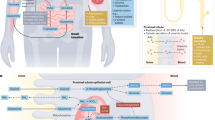
Besides the kidneys, the gut and skeletal muscle also play an important role in inter-organ metabolism of glutamine, which is a major substrate for endogenous synthesis of citrulline and arginine in most mammals (Fig. 5.3). Although the small intestine takes up glutamine from the arterial blood at all times albeit at different rates, the liver either releases or takes up glutamine depending on various physiological conditions (Welbourne 1987). For example, in acidotic animals, the release of glutamine by skeletal muscle is enhanced, the liver becomes a net producer of glutamine, and the uptake of arterial glutamine by the small intestine is reduced, resulting in an increased provision of glutamine for ammoniagenesis in the kidneys. Studies with rats have shown that skeletal muscle and liver contribute about 55% and 45%, respectively, of the glutamine extracted by the kidneys during chronic acidosis (Schrock and Goldstein 1981). In contrast to mammals, the skeletal muscle of some fish (e.g., holostean and teleost fish) does not appear to have a net synthesis of glutamine due to a greater activity of glutaminase than glutamine synthetase (Chamberlin et al. 1991). However, this does not necessarily mean a lack of net glutamine synthesis by the skeletal muscle of aquatic animals because of the complex intracellular compartmentation of these two enzymes. In support of this view, Zhou et al. (2018) reported that the skeletal muscle of hybrid striped bass synthesized glutamine from branched-chain AAs as the amino-group donors and release glutamine from the tissue.

Arginine synthesis in most mammalian animals. The small intestine of most mammals (including humans, pigs, cattle, sheep, and rats) converts glutamine and proline into citrulline, and releases citrulline into the blood. In adult mammals, the kidneys are the major site for the synthesis of arginine from citrulline. Birds lack an ability to form citrulline from glutamine or proline, but their tissues (including the kidneys are capable of converting citrulline into arginine. At present, little is known about endogenous synthesis of arginine in aquatic animals (e.g., fish and shrimp). Note: there is no net synthesis of arginine by the liver under physiological conditions. The conversion of glutamine and glutamate into citrulline occurs exclusively in the mitochondria of enterocytes. Arginine is mainly formed from citrulline in the cytoplasm of almost all cell types. ASL argininosuccinate lyase, ASS argininosuccinate synthase, NAG N-acetylglutamate, OAT ornithine aminotransferase, OCT ornithine carbamoyltransferase, PO proline oxidase, PDG phosphate-activated glutaminase, P5CS pyrroline-5-carboxylate synthase. This figure is adapted from Wu (2013a, b)
As in mammals and birds, glutamine is also crucial for the renal regulation of acid-base balance in fish (Li et al. 2020). The activities of both glutaminase and glutamine synthetase are high in the kidneys of holostean and teleost fish (Chamberlin et al. 1991). There is a metabolic channeling between glutaminase and glutamine synthetase in the kidneys of the dogfish shark to regulate their ammonia production (King and Goldstein 1983). This pathway involves the increased uptake of glutamine in the arterial blood by the kidneys and the increased degradation of glutamine into ammonia and α-KG during acidosis. However, the majority of ammonia and/or urea is excreted across the gills rather than through the kidney in most fish species (Ip et al. 2001). Both Na+/H+ exchange and vacuolar-type H+-ATPase are present in the epithelium of gills to export H+ from the fish to the environment (Cameron and Kormanik 1982). The gills generally excrete much more acids from the body than the kidneys during acidosis, but the renal reabsorption of HCO3− is required in the systemic regulation of acid-base balance as noted previously (Perry et al. 2003).
5.5.3 Arginine and Nitric Oxide (NO) Production
Arginine is a basic AA in physiological fluids and a precursor for the syntheses of proteins, NO, urea, polyamines, proline, glutamate, creatine and agmatine (Wu and Morris 1998; Wu et al. 2009). The metabolism and functions of arginine has been well summarized (Morris Jr. 2016; Wu et al. 2009, 2016). In adult mammals, the endogenous synthesis of arginine involves the intestinal-renal axis (Reyes et al. 1994; Wu and Morris 1998; Brosnan and Brosnan 2004). Specifically, citrulline is synthesized from glutamine, glutamate and proline in the mitochondria of enterocytes, released from the small intestine, and taken up primarily by kidneys for arginine production. About 85% of the gut-derived citrulline is taken up by the kidneys for quantitative conversion into arginine, which is then released into the renal vein (Brosnan and Brosnan 2004). In neonates, most of the citrulline synthesized in enterocytes is converted locally into arginine (Wu and Knabe 1995; Wu 1997). In cats, the small intestine does not produce citrulline due to the deficiency of pyrroline-5-carboxylate (P5C) synthase, and therefore there is no de novo synthesis of arginine (Rogers and Phang 1985). For fish, the endogenous synthesis of arginine is likely limited (NRC 2011), but no information is available regarding P5C synthase activity in any of their tissues. It should be borne in mind that a lack of the hepatic urea cycle is not the reason for possible absence of de novo arginine synthesis in some fish. The fact that cats and possibly many species of fish have survived without the ability to synthesize arginine may be attributed to their usual consumption of animal-source foods (containing high-arginine content) as carnivores.
In the kidneys of mammals and birds, citrulline is readily converted into arginine via argininosuccinate synthetase and argininosuccinate lyase, which are localized in their proximal tubule (Silbernagl 1988; Brosnan and Brosnan 2004). An adult human produces about 2 g arginine/day, which is 40%–50% of 4–5 g of dietary arginine intake per day (Brosnan and Brosnan 2004). The kidneys have a high capacity for converting citrulline into arginine but arginine synthesis is limited in vivo by the rate of delivery of citrulline (Dhanakoti et al. 1990). It is unknown whether the kidneys can convert citrulline into arginine in aquatic animals (e.g., fish).
Production of arginine by the kidneys is important for the health of animals. This amino acid has versatile physiological functions directly or indirectly through its metabolites (e.g., NO, agmatine, and ornithine; Wu 2013a,b). As a signaling molecule, NO regulates blood flow, angiogenesis, embryogenesis, immune response, hormone secretion, and protein synthesis (Wu 2013a). Interestingly, the concentration of arginine in plasma declines as the aging kidneys develop progressive injury, which could contribute to endothelial dysfunction and decreased NO production in chronic renal disease (Baylis and Corman 1998). An increase in renal arginine production may serve to sustain systemic NO production in response to endotoxemia (Hallemeesch et al. 2002). As a result, arginine may be regarded as a nutritionally essential amino acid for aging subjects, and dietary supplementation with arginine may be required to maintain sufficient substrate levels for NO production (Weinstein and Anderson 2010). The reduced NO-generating capacity in aging subjects may also result, in part, from deficiencies of NO synthase and its cofactors such as tetrahydrobiopterin and NADPH (Mistry et al. 2002; Delp et al. 2008).
5.5.4 Methylarginine and Sulfur AA Metabolism
Dimethylarginines, such as asymmetric dimethylarginine (ADMA), NG-monomethyl-l-arginine (NMMA) and symmetrical dimethylarginine (SDMA), result from the degradation of methylated proteins (Wu 2013a). Production of NO from arginine is reduced by ADMA and NMMA (competitive inhibitors of NOS) as well as SDMA (an inhibitor of arginine transport) (Tsikas et al. 2018). The kidneys play an important role in the metabolism and disposal of these endogenous arginine analogues (Van De Poll et al. 2004). Moreover, the kidneys have an ability to degrade ADMA and NMMA to citrulline via NG-dimethylarginine dimethylaminohydrolase. Renal excretion also plays a role in the elimination of endogenous dimethylarginines. In humans, approximately 4.5% of the ADMA generated in the body is excreted in the urine, and the remainder is metabolized by the kidneys and liver (Ogawa et al. 1987; Van De Poll et al. 2004). Concentrations of free NMMA, ADMA, and SDMA in the plasma are low in healthy subjects (<1 μM) (Wu 2013a). However, renal dysfunction elevates ADMA or NMMA levels, thereby inhibiting NO synthesis, impairing endothelial function, and promoting atherosclerosis (Sibal et al. 2010; Cooke and Ghebremariam 2011).
Arginine methylation reactions involve the modification of guanidino N atoms and require S-adenosylmethionine (SAM), which is a metabolite of methionine (Wu 2013a). In mammals, the kidneys play an important role in sulfur AA metabolism (Fig. 5.4). In the reaction catalyzed by protein arginine N-methyltransferase, SAM is converted into S-adenosylhomocysteine (SAHC). The latter is also a metabolic precursor of homocysteine in tissues. Garibotto et al. (2010) suggested that the mammal kidneys have a good ability to remove SAHC, but not homocysteine from the bloodstream. Thus, elevated concentrations of SAHC in the whole blood are associated with renal dysfunction, and SAHC may modulate one-carbon flux (Stam et al. 2004). Considering that SAHC is a feedback inhibitor of most methyltransferases, the kidneys may play a major role in the control of the overall transmethylation rates and the circulating levels of homocysteine. Moreover, homocysteine may also inhibit NG-dimethylarginine dimethylaminohydrolase, causing ADMA to accumulate and suppressing NO synthesis (Stühlinger et al. 2001; Holven et al. 2003). This could impair endothelium-mediated NO-dependent vasodilatation. Of note, the regeneration of methionine from homocysteine is regulated by the one carbon cycle which is mediated by nutrients, such as folic acid, vitamin B12, and vitamin B6. Thus, dietary supplementation with these vitamins can reduce blood homocysteine levels and provide an effective therapy to improve outcomes in patients undergoing coronary angioplasty (Schnyder et al. 2002). In addition, intake of plant-source proteins, which contain less methionine and cysteine than animal-source proteins (Hou et al. 2019; Li and Wu 2020), may be beneficial for patients with hepatic and renal diseases who have a reduced ability to metabolize homocysteine or excrete this metabolite in urine.

The methylarginines and sulfur amino acid metabolism pathway in animals. Taurine is synthesized from cysteine in the liver of most mammals (except for cats and possibly some of the other carnivores) and birds. Taurine plays important roles as an antioxidant and as well as a regulator of cell signaling and metabolism in the kidneys. ADMA asymmetric dimethylarginine, DDAH, NG-dimethylarginine dimethylaminohydrolase; H4-folate, tetrahydro-folate; NMMA NG-monomethyl-l-arginine, NO nitric oxide, PRMTs, protein arginine methyltransferases; SAM S-adenosyl-L-methionine, SAH S-adenosyl-L-homocysteine, SDMA symmetrical dimethylarginine
5.5.5 Creatine Synthesis
In mammals and birds, the kidneys participate in the inter-organ synthesis of creatine from arginine, glycine, and methionine (Fig. 5.5). This metabolic pathway requires arginine:glycine amidinotransferase (AGAT) and guanidinoacetate N-methyltransferase (GAMT) (Silva et al. 2014). AGAT transfers the guanidino group from arginine to glycine to produce guanidinoacetate and ornithine. This enzyme is expressed primarily in the renal tubule, pancreas, and to much lesser extent, in the liver and other tissues (Wu 2013a). The kidneys are the major site for the production of guanidinoacetate in mammals and birds, as the renal activity of AGAT is the primary determinant of in vivo creatine synthesis (Wyss and Kaddurah-Daouk 2000). The liver takes up guanidinoacetate from the blood and converts this metabolite into creatine via GAMT, which requires SAM as the cofactor. In skeletal muscle, creatine is spontaneously degraded to creatinine, which is excreted in the urine. The lost creatine must be replaced from diets or de novo synthesis to maintain its total pool in the body (Barcelos et al. 2016). In rats, the kidneys produce a sufficient quantity of guanidinoacetate to replace creatinine lost in the urine (Silva et al. 2014). A 70-kg healthy adult synthesizes 1.7 g creatine per day from 2.3 g arginine, 1.0 g glycine, and 2.0 g methionine (Wu and Morris 1998) to match the daily irreversible loss of creatine (1.7 g/day) as creatinine via the urine (Stead et al. 2006; Edison et al. 2007).

The synthesis of creatine in animals: Creatine is synthesized from arginine, glycine, and methionine in animals via inter-organ cooperation. The creatine can be further converted into creatinine in skeletal muscle. AGAT arginine:glycine amidinotransferase
In contrast to mammals and birds, skeletal muscle is the organ for de novo creatine synthesis in fish. Borchel et al. (2014) reported that: (a) AGAT is nearly absent from the kidneys of the rainbow trout, (b) GAMT is weakly expressed in their liver, and (c) both AGAT and GAMT are strongly expressed in their skeletal muscle. Similar results were obtained for other four species of fish: maraena whitefsh (Coregonus maraena), pikeperch (Sander lucioperca), European perch (Perca fuviatilis), and the Atlantic herrings (Borchel et al. 2019). Thus, in the skeletal muscle of fish, the guanidinoacetate generated by AGAT is locally used for creatine production by GAMT. In these aquatic animals, besides the skeletal muscle, the kidneys also strongly express GAMT (Borchel et al. 2014, 2019) and, therefore, are capable of converting diet- and blood-derived guanidinoacetate into creatine.
In mammals, creatine synthesis is regulated by the availability of substrates and renal AGAT activity. Studies with rats have shown that the expression of this enzyme at the pre-translational level is down-regulated by dietary creatine intake but up-regulated by growth hormone (McGuire et al. 1984; Guthmiller et al. 1994; Edison et al. 2007). Thus, dietary supplementation with creatine to rats decreases renal AGAT mRNA levels and activity as well as the production of guanidinoacetate by the kidneys (Edison et al. 2007). In addition, AGAT is inhibited by ornithine (Sipilä 1980) but activated by arginine (Edison et al. 2007). Therefore, arginine supplementation enhances the production of guanidinoacetate and creatine (Edison et al. 2007), which is beneficial for endothelial function (Bodamer et al. 2005). At present, little is known about creatine biosynthesis in fish.
The main function of creatine is to store energy in tissues, primarily skeletal muscle and brain, via creatine kinase, which interconverts creatine and phosphocreatine (Brosnan and Brosnan 2016). This enzyme is expressed at high levels in most of the cells and tissues that have high energy requirements, including the brain, kidneys, retinal photoreceptor cells, spermatozoa, testis, uterus, placenta, sensory hair cells of the inner ear, as well as skeletal, cardiac, and smooth muscles (Wu 2013a). Because of the action of creatine kinase, the concentration of ATP in the brain varies little despite large and rapid changes in turnover rates (Kekelidze et al. 2001). For example, the rate of the creatine kinase-catalyzed reaction rapidly increases with increased ATP demand (e.g., in seizures) but decreases with decreased ATP synthesis (e.g., hypoxia). In a resting state, skeletal muscle transfers excessive ATP to creatine, generating phosphocreatine; in a physically active state, skeletal muscle hydrolyzes phosphocreatine, releasing energy. Approximately 95% of creatine plus phosphocreatine in the body is present in skeletal muscle and brain, indicating the importance of creatine in maintaining ATP homeostasis.
5.5.6 Tyrosine Synthesis
The sources of tyrosine can be diets, protein breakdown, and phenylalanine hydroxylation. The liver and kidneys express phenylalanine hydroxylase, which irreversibly converts phenylalanine into tyrosine (Hufton et al. 1995). In female rats, the kidney (and the liver) is devoid of phenylalanine hydroxylase on Day 20 of gestation, but at birth contains 20% of the adult activity (McGee et al. 1972). In contrast to the liver, the kidneys have a limited ability to degrade tyrosine (Møller et al. 2000; Boirie et al. 2004). In adults, the kidneys extract phenylalanine from the blood and release tyrosine (Kopple 2007). In the post-absorptive state, the human kidneys synthesize an appreciable amount of tyrosine from phenylalanine at the rate of 5.2 μmol/min, compared with the rate of 3.0 μmol/min in the splanchnic bed (the portal-drained viscera plus liver; Møller et al. 2000). Humans and other animals (e.g., dogs and rats) with renal and hepatic disease may be at risk for phenylalanine overloading and tyrosine deficiency (Møller et al. 2000; Kopple 2007), raising the possibility that tyrosine is a nutritionally essential AA under these conditions (e.g., end-stage renal disease; Kopple 2007). The rate of conversion of phenylalanine into tyrosine is approximately 50% lower in patients with end-stage renal disease in comparison with healthy subjects (Boirie et al. 2004). Tyrosine has important physiological functions, including the syntheses of thyroid hormones and neurotransmitters (Wu 2013a,b). In chronic kidney failure, the concentrations of tyrosine and the ratio of tyrosine/phenylalanine are reduced in plasma and many tissues, with either no change or an increase in phenylalanine concentrations (Kopple 2007).
5.5.7 Glycine and Serine Synthesis
Glycine plays an important role in metabolic regulation, anti-oxidative reactions, and neurological function, such as (1) preventing tissue injury; (2) enhancing anti-oxidative capacity; (3) promote protein synthesis and wound healing; (4) improving immunity; and (5) treating metabolic disorders in obesity, diabetes, cardiovascular disease, ischemia-reperfusion injuries, cancers, and various inflammatory diseases (Wang et al. 2013; Wang et al. 2014a,b). Typical intake of dietary glycine meets at most 20% and 14% of daily glycine needs in young pigs (Hou et al. 2015a) and adult humans (Wu 2020b). Thus, these mammals must synthesize a majority of glycine needed daily to ensure their optimal health, growth, and feed efficiency (Li and Wu 2018). Available evidence shows that glycine is a nutritionally essential AA for maximal protein accretion in milk-fed piglets (Wang et al. 2014a).
The pathways for glycine synthesis via different sources are summarized in Fig. 5.6. In rats, the metabolism of hydroxyproline by the kidneys may contribute to a significant production of both glycine and serine. All the enzymes (hydroxyproline oxidase, hydroxyoxoglutarate aldolase, and alanine glyoxalate transaminase) involved in the metabolism of hydroxyproline to glycine are present in the kidneys and cortical tubules (Lowry et al. 1985). Alanine-glyoxylate aminotransferase is responsible for the nearly irreversible transfer of the amino group from alanine to glyoxylate, yielding glycine and pyruvate (Wang et al. 2013). Therefore, the rate of hydroxyproline metabolism is increased by alanine as a nitrogen donor (Lowry et al. 1985). Wu et al. (2019) have estimated that dietary plus endogenously-derived hydroxyproline contributes to most (64%) of the total glycine synthesis in 7-day-old pigs, whereas glucose plus glutamate contributes to ~25% of total glycine synthesis in the neonates. Consistent with this view, milk and plasma contain large amounts of hydroxyproline-rich small peptides (Hu et al. 2017).
There are multiple pathways to degrade glycine in mammals and birds (Coon et al. 1974; Wu 2013a). In the liver and kidneys of these animals, glycine can be converted into serine by serine hydroxymethyl transferase at a low rate or degraded to NH3 and CO2 by glycine cleavage system (GCS) at a higher rate, with the regeneration of tetrahydrofolate. The low activities of renal GCS and serine hydroxymethyl transferase are necessary for the net production of glycine by the kidneys (Lowry et al. 1985; Wu et al. 2019). Thus, although the kidneys of mammals and birds may release serine into the blood circulation, the concentration of serine in plasma is enhanced only moderately by dietary glycine supplementation (Wang et al. 2013). Interestingly, the kidneys of fish have a higher activity of serine dehydratase (converting serine into pyruvate and NH4+) as compared with mammals, and are a major site for serine catabolism in the body (Jürss and Bastrop 1995).

The synthesis of glycine and serine from hydroxyproline, choline or D-3-phosphoglycerate. These synthetic pathways in mammals, fish and birds are cell- and tissue- specific. SHMT serine hydroxymethyl transferase
5.5.8 Branched-Chain Amino Acid (BCAA) Metabolism
BCAAs play an important role in whole-body nitrogen metabolism under both physiological and pathological conditions (Cano et al. 2006). In dogs, the kidneys take up BCAAs (valine, leucine, and isoleucine) after consuming AAs in a meal (Kuhlmann and Kopple 1990). In the post-absorptive state, whole-blood-renal AA exchanges are characterized by the release of leucine from the kidneys that accounts for one-third of whole-body leucine production, with no net renal exchange for valine or isoleucine (Tizianello et al. 1983). This indicates a different metabolic fate of leucine than the other two BCAAs in the kidneys. The plasma pool of BCAAs in the post-absorptive state is regulated by their release from tissues (e.g., skeletal muscle and kidneys) due to proteolysis, their uptakes by the tissues, and their oxidation in a tissue-specific manner (Felig 1975; Abumrad and Miller 1983; Price et al. 1998). Catabolic factors, such as acidosis and inflammation, are responsible for increases in intramuscular protein breakdown and BCAA degradation (Kopple et al. 2005), in association with an enhanced activity of branched-chain α-ketoacid (BCKA) dehydrogenase (May et al. 1987; Lim et al. 1998). In mammals and birds, BCKAs are extensively oxidized to CO2 and water in the kidneys due to a high activity of BCKA dehydrogenase. Likewise, in fish species, the kidneys seem to be important for the catabolism of BCAAs because a high activity of BCAA transaminase is present in the posterior kidney (Hughes et al. 1984). Dietary supplementation with leucine or valine increases the activity of BCAA transaminase in the posterior kidney of Lake trout, Salvelinus namaycush (Hughes et al. 1984). Thus, in all animal species studied, the kidneys play an important role in regulating whole-body BCAA homeostasis.
5.5.9 Homoarginine Synthesis
L-Homoarginine (hArg) has an additional −CH2 group on its main carbon chain than L-arginine. Thus, hArg is a structural homologue of arginine. Synthesis of hArg by rats and humans was discovered by Ryan and Wells (1964), with the major sites of synthesis including the kidneys and liver (Ryan et al. 1968, 1969; Hou et al. 2015b). Less than 0.025% and < 0.045% of ingested arginine is metabolized to hArg in pigs and rats, respectively (Hou et al. 2016). Concentrations of hArg in plasma are relatively low (approximately 2 μM) in healthy humans (Marescau et al. 1985) and rats (Hou et al. 2015b), increase up to 20 μM in hyperargininemic patients (Marescau et al. 1985), and decrease in diabetic mice (Wetzel et al. 2019). The enzyme responsible for hArg synthesis in animals is unknown, but there are suggestions that mitochondrial arginine:glycine amidinotransferase (AGAT) catalyzes the transfer of the amidino group from L-arginine to L-lysine to form hArg, with L-ornithine being a product (Tsikas and Wu 2015). The concentrations of hArg in the brain, kidney and liver of rats are about 1.5, 100 and 115 μM, respectively (Hou et al. 2015b). Although AGAT activity in the liver is 20 times greater than that in the kidneys, concentrations of hArg in the liver are only 15% greater than those in the kidneys, possibly due a relatively low concentration of arginine in the liver (~0.05 mM) as compared with a much higher concentration (~1.5 mM) in the kidneys, and (b) the possibility of a higher rate of hArg catabolism in the liver than in the kidney.
hArg can regulate the metabolism of arginine and other nutrients by inhibiting arginine transport across the cell membranes, arginase, as well as liver and bone alkaline phosphohydrolases, while serving as a substrate for NO synthase (Tsikas et al. 2018). Whether hArg has a beneficial or an adverse effect on NO production likely depends on cell type, extracellular and intracellular concentrations of arginine, and activities of competing pathways for hArg and arginine metabolism. There is evidence that low concentrations of hArg in plasma are associated with a high risk of cardiovascular (Atzler et al. 2015) and renal (Wieczorek-Surdacka et al. 2019) diseases in humans and animals. Accordingly, dietary supplementation with hArg [via either drinking water (50 mg/L) or a mini-osmotic pump (0.72 mg/kg body weight per day)] for 12 weeks prevents kidney damage in diabetic mice (Wetzel et al. 2019).
5.6 Benefits of AAs on Renal Function
The current recommended protein intake for healthy adult humans with minimal physical activity is 0.8 to 1.0 g/kg body weight/day (Wu 2016). Excessive protein intake may promote renal damage by chronically increasing glomerular pressure and hyperfiltration in mammals (Brenner et al. 1982; Martin et al. 2005; Beasley et al. 2014). In dogs, transition from a carbohydrate meal to a meat meal resulted in a 50–100% increase in glomerular filtration rate (GFR; King and Levey 1993). High protein intake may accelerate renal disease in adults with mild renal insufficiency or in peritoneal dialysis patients, leading to a progressive loss of renal capacity and function (Johnson et al. 2003; Knight et al. 2003). In another study, increased GFR and renal hypertrophy as well as hormonal changes occur at moderate rates after the consumption of a high-protein diet (e.g., 2.6 g protein/kg body weight/day vs 0.1 to 0.4 and 1.0 to 1.4 g protein/kg body weight/day for 2 weeks; King and Levey 1993; Martin et al. 2005). However, an early study indicated that glucagon, insulin and growth hormone were not involved in an increase of GFR induced by protein intake (Bergstrom et al. 1985), indicating a complex interplay among nutritional and physiological factors. Reddy et al. (2002) reported that consumption of low-carbohydrate and high-protein diets (19 g carbohydrate and 164 g protein for the first 2 weeks and 33 g carbohydrate and 170 g protein for the subsequent 4 weeks) increased risks for stone formation and bone loss, compared with a regular (control) diet (285 g carbohydrate and 91 g protein). However, interpretation of this result is confounded because the dietary intakes of energy and minerals (including calcium) were lower in the high-protein group, compared with the control group. Nonetheless, dietary protein restriction is a common treatment for patients with renal disease by alleviating uremic symptoms due to the better control of hyperparathyroidism, hyperphosphatemia and hyperkalemia as well as improvements in the epithelial integrity of the renal tubule (Pedrini et al. 1996; Chauveau et al. 2007). Changes from a low to a high intake of dietary protein can allow for adaptive alterations in renal size and function without adverse effects (Skov et al. 1999). Results of clinical studies indicate that consumption of a weight-loss diet containing 90–120 g protein per day does not affect renal function in overweight subjects or in obese adults with type-II diabetes, compared with the counterparts consuming 55–70 g protein per day (Wu 2016). Aquatic animals naturally require 100–200% greater dietary protein than land mammals (Wu 2018), it is unclear whether such a high intake of protein over a prolonged period of time may adversely affect the health of fish and shrimp. Thus, it is important to study the roles of AAs not only as major metabolic fuels for the kidneys of aquatic animals but also as protectors of their renal health.
Restricted intakes of dietary protein below physiological requirements for AAs may be harmful to health by accelerating the development of protein-energy wasting, leading to adverse consequences such as malnutrition and increased risk for death (Garibotto 1999). Protein-energy wasting is a strong predictor of the adverse outcomes that are characterized by a decline in body protein mass and energy reserves. Unfortunately, this metabolic condition is often under-appreciated in early to moderate stages of chronic kidney disease (Kovesdy et al. 2013). A protein-restricted diet supplemented with some AAs or the α-ketoacids or hydroxyacid analogues of AAs (e.g., phenylalanine and methionine) may have potential beneficial effects on improving renal function and survival (Abel, et al. 1973; King and Levey 1993). For example, oral administration of tyrosine, tyrosine-containing dipeptides, or N-acetyl-tyrosine can replete the plasma and intracellular pools of tyrosine and improve nitrogen balance in chronic renal failure patients on a low protein diet (Druml et al. 1989). The AA supplements for those patients include valine, leucine, isoleucine, phenylalanine, threonine, tryptophan, lysine, methionine, tyrosine, and histidine (Alvestrand et al. 1983; Cano et al. 2006). Patients with chronic kidney disease on dialysis have been recommended to reduce protein intake from 1.2 to < 0.8 or even < 0.6 g/kg body weight per day (Kalantar-Zadeh et al. 2011). BCAA and BCKA supplements may be integrated into a therapeutic strategy that includes protein restriction for these subjects (Cano et al. 2006). However, high levels of dietary BCAAs can exert a deleterious effect on renal disease and should be avoided at all times (Pillai and Verrey 2019).
Functional AAs hold great promise in prevention and treatment of metabolic diseases, such as renal dysfunction (Wu 2013b). A deficiency of AAs or glutathione depletion may contribute to disturbances in renal structure and function (Epstein et al. 1982). An increase in renal blood flow in response to a short-term AA infusion can protect the kidneys from acute ischemic insults in animal models (King and Levey 1993). Glycine has a positive effect on reducing medullary injury in perfused kidneys (Silva et al. 1991) and mild ischemia-reperfusion injury in the kidneys in vivo, in part by decreasing initial damage and preventing chronic hypoxia (Yin et al. 2002). Thus, glycine can be used to prevent or treat ischemic and/or toxin-induced injury to the kidneys. Similarity, Baines et al. (1990) reported that small neutral AAs, such as glycine and alanine, prevented tubular disruption through their physicochemical effects on stabilizing the tertiary structure of membrane proteins. Some AAs play a role in the renal cell cycle and apoptosis, function as osmolytes during the stress response, scavenge reactive oxygen species (ROS), and modulate blood flow (Chesney et al. 2010). For example, taurine reduces oxidant levels in diabetic nephropathy (Trachtman et al. 1995), and protects against Cd-induced renal oxidative damages (Manna et al. 2009) or cyclosporine A- induced hypertension and nephrotoxicity (Hagar et al. 2006). In addition, arginine corrects renal failure-associated endothelial dysfunction (Hand et al. 1998) and hypertension (Wu et al. 2000). L-Citrulline may also be beneficial for ameliorating renal disorders, while facilitating the removal of ammonia through arginine synthesis by proximal renal tubules and other cell types.
5.7 Functions of AAs in the Immune System of Fish Kidneys
As noted previously, the anterior (head) kidney is an important immune organ in fish, in addition to their thymus and spleen. The head kidney contains cytokine-producing T-lymphocytes (T-cells), antibody-producing B-lymphocytes (B-cells), and ROS-producing macrophages (Shoemaker et al. 2015). Protein synthesis in immunocytes require AAs. Our recent studies have shown that AAs, such as glutamate and glutamine, are more important energy substrates than glucose and fatty acids in the kidneys of fish (Jia et al. 2017). These findings indicate an essential role of the head kidney and AAs in innate and adaptive immunities in fish. There are many lines of evidence that functional AAs have beneficial effects on the immune system of fish via improving the kidney functions. For example, arginine stimulates the proliferation of T-lymphocytes in response to mitogens (Ochoa et al. 2001) and increases the number of cell surface receptors in fish (Cheng et al. 2011). In Jian carp, the transcript levels of inflammatory response genes in the head kidney are up-regulated by increasing dietary arginine intake (Chen et al. 2015). In addition, glutamine supplementation improves the development of B-cells in the head kidney of fish in the head kidney (Hu et al. 2015). Similarly, glutamine supplementation promotes the development of the head kidney in channel catfish (Pohlenz et al. 2012). Finally, leucine supplementation augments antioxidant activities, immune-gene expression, and ant-inflammatory responses in the head kidney of Labeo rohita fingerlings (Giri et al. 2015). Thus, AAs and their metabolism in the kidneys play an important role in mounting immune responses to challenges by bacterial, viral, parasitic, and fungal pathogens.
5.8 Conclusion and Perspectives
Results from studies with humans and different animal species indicate that the kidneys play a vital role in AA metabolism and hemostasis through the following mechanisms: the reabsorption of free AAs and other nutrients by the proximal tubules into the blood; the production of glucose from AAs via gluconeogenesis; regulation of the acid-base balance via the inter-organ metabolism of glutamine; the synthesis of arginine from citrulline and the conversion of arginine into NO; the generation of polyamines from arginine and ornithine; the generation of tyrosine from phenylalanine; glycine and serine syntheses; the catabolism of BCAAs; the metabolism of methylarginines and sulfur AAs; ATP production; and immune response. Renal dysfunction alters the pathways of AA metabolism and, therefore, whole-body AA homeostasis, leading to health problems, including excessive muscle protein breakdown, inflammation, mitochondrial dysfunction, impaired immune system, and cardiovascular diseases (Fig. 5.7). Although a reduced intake of protein is usually recommended to subjects with chronic renal failure in humans and other mammals, moderate supplementation with AAs (including valine, leucine, isoleucine, phenylalanine, threonine, tryptophan, lysine, methionine, tyrosine, histidine, glycine, glutamine, and taurine) may provide a beneficial effect to meet physiological requirements.

The diseases development from the impaired amino acids metabolism in kidney dysfunction (like chronic kidney failure). ADMA asymmetric dimethylarginine, BCAAs, branched-chain amino acids
Most of the knowledge on renal AA metabolism was gained from studies with land mammals and birds. At present, little is known about AA synthesis or catabolism in the kidneys of fish. In aquatic animals, the nutritional values of AAs are often evaluated on the basis of growth, protein accretion, survival, immunity, anti-oxidative ability, or meat quality. We propose that protective effects of AAs on renal structure and function should also be an important criterion with which to assess dietary requirements of fish for these nutrients. It is likely that renal AA metabolism differs between fish and terrestrial animals. Although much research has been done with humans, mice and rats regarding the roles of the gut, intestinal microbiome, liver, skeletal muscle and adipose tissue in metabolic disorders (including diseases), there is only a limited number of published studies regarding the improvement of renal functions in livestock and poultry production or aquaculture (Wu 2020a). Further studies are warranted to address this critical issue in order to further enhance the quantity and sustainability of animal production.
References
Abel RM, Beck CH Jr, Abbott WM, Ryan JA Jr, Barnett GO, Fischer JE (1973) Improved survival from acute renal failure after treatment with intravenous essential L-amino acids and glucose: results of a prospective, double-blind study. N Engl J Med 288:695–699
Abumrad NN, Miller B (1983) The physiologic and nutritional significance of plasma-free amino acid levels. J Parent Ent Nutr 7:163–170
Afsar B, Vaziri ND, Aslan G, Tarim K, Kanbay M (2016) Gut hormones and gut microbiota: implications for kidney function and hypertension. J Am Soc Hypertens 10:954–961
Ahmed SB, Ramesh S (2016) Sex hormones in women with kidney disease. Nephrol Dial Transplant 31:1787–1795
Alvestrand A, Fürst P, Bergström J (1983) Intracellular amino acids in uremia. Kidney Int 16(Suppl):S9–S16
Anderson JW, Stowring LI (1973) Glycolytic and gluconeogenic enzyme activities in renal cortex of diabetic rats. Am J Phys 224:930–936
Anderson PA, Berzins IK, Fogarty F, Hamlin HJ, Guillette LJ Jr (2011) Sound, stress, and seahorses: the consequences of a noisy environment to animal health. Aquaculture 311:129–138
Atzler D, Schwedhelm E, Choe CU (2015) L-Homoarginine and cardiovascular disease. Curr Opin Clin Nutr Metab Care 18:83–88
Baines AD, Shaikh N, Ho P (1990) Mechanisms of perfused kidney cytoprotection by alanine and glycine. Am J Phys 259:F80–F87
Barcelos RP, Stefanello ST, Mauriz JL, Gonzalez-Gallego J, Soares FA (2016) Creatine and the liver: metabolism and possible interactions. Minirev Med Chem 16:12–18
Baylis C, Corman B (1998) The aging kidney: insights from experimental studies. J Am Soc Nephrol 9:699–709
Beasley JM, Katz R, Shlipak M, Rifkin DE, Siscovick D, Kaplan R (2014) Dietary protein intake and change in estimated GFR in the cardiovascular health study. Nutrition 30:794–799
Belhadj Slimen I, Najar T, Ghram A, Abdrrabba M (2016) Heat stress effects on livestock: molecular, cellular and metabolic aspects, a review. J Anim Physiol Anim Nutr 100:401–412
Bergstrom J, Ahlberg M, Alvestrand A (1985) Influence of protein intake on renal hemodynamics and plasma hormone concentrations in normal subjects. Acta Med Scand 217:189–196
Blokhuis HJ, Hopster H, Geverink NA, Korte SM, Van Reenen CG (1998) Studies of stress in farm animals. Comp Haematol Int 8:94–101
Bodamer OA, Sahoo T, Beaudet AL, O’Brien WE, Bottiglieri T, Stöckler-Ipsiroglu S, Wagner C, Scaglia F (2005) Creatine metabolism in combined methylmalonic aciduria and homocystinuria. Ann Neurol 57:557–560
Boirie Y, Albright R, Bigelow M, Nair KS (2004) Impairment of phenylalanine conversion to tyrosine inend-stage renal disease causing tyrosine deficiency. Kidney Int 66:591–596
Boll M, Daniel H, Gasnier B (2004) The SLC36 family: proton-coupled transporters for the absorption of selected amino acids from extracellular and intracellular proteolysis. Pflugers Arch 447:776–779
Borchel A, Verleih M, Rebl A, Kühn C, Goldammer T (2014) Creatine metabolism differs between mammals and rainbow trout (Oncorhynchus mykiss). Springerplus 3:510
Borchel A, Verleih M, Kühn C, Rebl A, Goldammer T (2019) Evolutionary expression differences of creatine synthesis-related genes: implications for skeletal muscle metabolism in fish. Sci Rep 9:5429
Bowman RH (1970) Gluconeogenesis in the isolated perfused rat kidney. J Biol Chem 245:1604–1612
Brenner BM, Meyer TW, Hostetter TH (1982) Dietary protein intake and the progressive nature of kidney disease: the role of hemodynamically mediated glomerular injury in the pathogenesis of progressive glomerular sclerosis in aging, renal ablation, and intrinsic renal disease. N Engl J Med 307:652–659
Bröer S (2006) The SLC6 orphans are forming a family of amino acid transporters. Neurochem Int 48:559–567
Bröer S (2008) Amino acid transport across mammalian intestinal and renal epithelia. Physiol Rev 88:249–286
Brosnan ME, Brosnan JT (2004) Renal arginine metabolism. J Nutr 134:2791S–2795S
Brosnan ME, Brosnan JT (2016) The role of dietary creatine. Amino Acids 48:1785–1791
Cameron JN, Kormanik GA (1982) The acid-base responses of gills and kidneys to infused acid and base loads in the channel catfish, Ictalurus punctatus. J Exp Biol 99:143–160
Cano NJ, Fouque D, Leverve XM (2006) Application of branched-chain amino acids in human pathological states: renal failure. J Nutr 136:299S–307S
Chamberlin ME, Glemet HC, Ballantyne JS (1991) Glutamine metabolism in a holostean (Amia calva) and teleost fish (Salvelinus namaycush). Am J Phys 260:R159–R166
Chauveau P, Combe C, Rigalleau V, Vendrely B, Aparicio M (2007) Restricted protein diet is associated with decrease in proteinuria: consequences on the progression of renal failure. J Renal Nutr 17:250–257
Chen G, Liu Y, Jiang J, Jiang W, Kuang S, Tang L, Tang W, Zhang YA, Zhou X, Feng L (2015) Effect of dietary arginine on the immune response and gene expression in head kidney and spleen following infection of Jian carp with Aeromonas hydrophila. Fish Shellfish Immunol 44:195–202
Cheng Z, Buentello A, Gatlin DM III (2011) Effects of dietary arginine and glutamine on growth performance, immune responses and intestinal structure of red drum, Sciaenops ocellatus. Aquaculture 319:247–252
Chesney RW, Han X, Patters AB (2010) Taurine and the renal system. J Biomed Sci 17:S4
Claiborne JB, Evans DH, Goldstein L (1982) Fish branchial Na+/NH4+ exchange is via basolateral Na+-K+-activated ATPase. J Exp Biol 96:431–434
Cooke JP, Ghebremariam YT (2011) DDAH says NO to ADMA. Arterioscler Thromb Vasc Biol 31:1462–1464
Coon CN, Luther LW, Couch JR (1974) Effect of glycine and serine in synthetic amino acid diets upon glycine and serine metabolism in chicks. J Nutr 104:1018–1023
Cullen-McEwen L, Sutherland MR, Black MJ (2016) The human kidney: parallels in structure, spatial development, and timing of nephrogenesis. In: Little MH (ed) Kidney development, disease, repair and regeneration. Academic, New York, pp 27–40
Daniel H, Spanier B, Kottra G, Weitz D (2006) From bacteria to man: archaic proton-dependent peptide transporters at work. Physiology 21:93–102
Davidson A (2014) J: kidney regeneration in fish. Nephron Exp Nephrol 126:45–49
Delp MD, Behnke BJ, Spier SA, Wu G, Muller-Delp JM (2008) Aging diminishes endothelium-dependent vasodilation and tetrahydrobiopterin content in rat skeletal muscle arterioles. J Physiol 586:1161–1168
Denic A, Glassock RJ, Rule AD (2016) Structural and functional changes with the aging kidney. Adv Chronic Kidney Dis 23:19–28
Dhanakoti SN, Brosnan JT, Herzberg GR, Brosnan ME (1990) Renal arginine synthesis: studies in vitro and in vivo. Am J Phys 259:E437–E442
Dressler GR (2006) The cellular basis of kidney development. Annu Rev Cell Dev Biol 22:509–529
Druml W, Roth E, Lenz K, Lochs H, Kopsa H (1989) Phenylalanine and tyrosine metabolism in renal failure: dipeptides as tyrosine source. Kidney Int 27(Suppl):S282–S286
Drummond IA (2005) Kidney development and disease in the zebrafish. J Am Soc Nephrol 16:299–304
Edison EE, Brosnan ME, Meyer C, Brosnan JT (2007) Creatine synthesis: production of guanidinoacetate by the rat and human kidney in vivo. Am J Phys 293:F1799–F1804
Enes P, Panserat S, Kaushik S, Oliva-Teles AA (2009) Nutritional regulation of hepatic glucose metabolism in fish. Fish Physiol Biochem 35:519–539
Epstein FH, Brosnan JT, Tange JD, Ross BD (1982) Improved function with amino acids in the isolated perfused kidney. Am J Phys 243:F284–F292
Evans DH (2002) Osmoregulation by vertebrates in aquatic environments. Encycl Life Sci:1–4
Evans DH, Cameron JN (1986) Gill ammonia transport. J Exp Zool 239:17–23
Felig P (1975) Amino acid metabolism in man. Annu Rev Biochem 44:933–955
Garibotto G (1999) Muscle amino acid metabolism and the control of muscle protein turnover in patients with chronic renal failure. Nutrition 15:145–155
Garibotto G, Sofia A, Saffioti S, Bonanni A, Mannucci I, Verzola D (2010) Amino acid and protein metabolism in the human kidney and in patients with chronic kidney disease. Clin Nutr 29:424–433
Gerich JE (2010) Role of the kidney in normal glucose homeostasis and in the hyperglycaemia of diabetes mellitus: therapeutic implications. Diabetic Med 27:136–142
Giri SS, Sen SS, Chi C, Kim HJ, Yun S, Park SC, Sukumaran V (2015) Effect of dietary leucine on the growth parameters and expression of antioxidant, immune, and inflammatory genes in the head kidney of Labeo rohita fingerlings. Vet Immunol Immunopathol 167:36–43
Guthmiller P, Van Pilsum JF, Boen JR, McGuire DM (1994) Cloning and sequencing of rat kidney L-arginine:glycine amidinotransferase. Studies on the mechanism of regulation by growth hormone and creatine. J Biol Chem 269:17556–17560
Hagar HH, El Etter E, Arafa M (2006) Taurine attenuates hypertension and renal dysfunction induced by cyclosporine a in rats. Clin Exp Pharmacol Physiol 33:189–196
Hallemeesch MM, Soeters PB, Deutz NE (2002) Renal arginine and protein synthesis are increased during early endotoxemia in mice. Am J Phys 282:F316–F323
Hand MF, Haynes WG, Webb DJ (1998) Hemodialysis and L-arginine, but not D-arginine, correct renal failure-associated endothelial dysfunction. Kidney Int 53:1068–1077
Hems DA (1972) Metabolism of glutamine and glutamic acid by isolated perfused kidneys of normal and acidotic rats. Biochem J 130:671–680
Holven KB, Haugstad TS, Holm T, Aukrust P, Ose L, Nenseter MS (2003) Folic acid treatment reduces elevated plasma levels of asymmetric dimethylarginine in hyperhomocysteinaemic subjects. Br J Nutr 89:359–363
Hou YQ, Yin YL, Wu G (2015a) Dietary essentiality of “nutritionally nonessential amino acids” for animals and humans. Exp Biol Med 240:997–1007
Hou YQ, Jia SC, Nawaratna G, Hu SD, Dahanayaka S, Bazer FW, Wu G (2015b) Analysis of L-homoarginine in biological samples by HPLC involving pre-column derivatization with o-phthalaldehyde and N-acetyl-L-cysteine. Amino Acids 47:2005–2014
Hou YQ, Hu SD, Jia SC, Nawaratna G, Che DS, Wang FL, Bazer FW, Wu G (2016) Whole-body synthesis of L-homoarginine in pigs and rats supplemented with L-arginine. Amino Acids 48:993–1001
Hou YQ, He WL, Hu SD, Wu G (2019) Composition of polyamines and amino acids in plant-source foods for human consumption. Amino Acids 51:1153-1165
Hu K, Zhang JX, Feng L, Jiang WD, Wu P, Liu Y, Jiang J, Zhou XQ (2015) Effect of dietary glutamine on growth performance, non-specific immunity, expression of cytokine genes, phosphorylation of target of rapamycin (TOR), and anti-oxidative system in spleen and head kidney of Jian carp (Cyprinus carpio var. Jian). Fish Physiol Biochem 41:635–649
Hu S, Nawaratna G, Long BD, Bazer FW, Johnson GA, Brosnan JT, Wu G (2017) The hydroxyproline–glycine pathway for glycine synthesis in neonatal pigs. J Anim Sci 95:45
Hufton SE, Jennings IG, Cotton RG (1995) Structure and function of the aromatic amino acid hydroxylases. Biochem J 311:353–366
Hughes SG, Rumsey GL, Nesheim MC (1984) Effects of dietary excesses of branched-chain amino acids on the metabolism and tissue composition of lake trout (Salvelinus namaycush). Comp Biochem Physiol A 78:413–418
Hyodo S, Kakumura K, Takagi W, Hasegawa K, Yamaguchi Y (2014) Morphological and functional characteristics of the kidney of cartilaginous fishes: with special reference to urea reabsorption. Am J Phys 307:R1381–R1395
Ip YK, Chew SF, Randall DJ (2001) Ammonia toxicity, tolerance, and excretion. Fish Physiol 20:109–148
Jia SC, Li XY, Zheng SX, Wu G (2017) Amino acids are major energy substrates for tissues of hybrid striped bass and zebrafish. Amino Acids 49:2053–2063
Johnson DW, Mudge DW, Sturtevant JM, Hawley CM, Campbell SB, Isbel NM, Hollett P (2003) Predictors of decline of residual renal function in new peritoneal dialysis patients. Perit Dial Int 23:276–283
Jürss K, Bastrop R (1995) Amino acid metabolism in fish. In: Hochachka PW, Mommsen TP (eds) Biochemistry and molecular biology of fishes, vol 4, pp 159–189
Kalantar-Zadeh K, Cano NJ, Budde K, Chazot C, Kovesdy CP, Mak RH, Mehrotra R, Raj DS, Sehgal AR, Stenvinkel P, Ikizler TA (2011) Diets and enteral supplements for improving outcomes in chronic kidney disease. Nat Rev Nephrol 7:369–384
Kekelidze T, Khait I, Togliatti A, Benzecry JM, Wieringa B, Holtzman D (2001) Altered brain phosphocreatine and ATP regulation when mitochondrial creatine kinase is absent. J Neurosci Res 66:866–872
Kierszenbaum AL, Tres L (2015) Histology and cell biology: an introduction to pathology E-book. Elsevier Health Sciences
King PA, Goldstein LE (1983) Renal ammoniagenesis and acid excretion in the dogfish, Squalus acanthias. Am J Physiol 245:R581–R589
King AJ, Levey AS (1993) Dietary protein and renal function. J Am Soc Nephrol 3:1723–1737
Kirchner S, Panserat S, Lim PL, Kaushik S, Ferraris RP (2008) The role of hepatic, renal and intestinal gluconeogenic enzymes in glucose homeostasis of juvenile rainbow trout. J Comp Physiol B 178:429–438
Knight EL, Stampfer MJ, Hankinson SE, Spiegelman D, Curhan GC (2003) The impact of protein intake on renal function decline in women with normal renal function or mild renal insufficiency. Ann Intern Med 138:460–467
Knox D, Walton MJ, Cowey CB (1980) Distribution of enzymes of glycolysis and gluconeogenesis in fish tissues. Mar Biol 56:7–10
Kopple JD (2007) Phenylalanine and tyrosine metabolism in chronic kidney failure. J Nutr 137:1586S–1590S
Kopple JD, Kalantar-Zadeh K, Mehrotra R (2005) Risks of chronic metabolic acidosis in patients with chronic kidney disease. Kidney Int 67:S21–S27
Kovesdy CP, Kopple JD, Kalantar-Zadeh K (2013) Management of protein-energy wasting in non-dialysis-dependent chronic kidney disease: reconciling low protein intake with nutritional therapy. Am J Clin Nutr 97:1163–1177
Krebs HA, Bennett DA, De Gasquet P, Gascoyne T, Yoshida T (1963) Renal gluconeogenesis. The effect of diet on the gluconeogenic capacity of rat-kidney-cortex slices. Biochem J 86:22–27
Kuhlmann MK, Kopple JD (1990) Amino acid metabolism in the kidney. Semin Nephrol 10:445–457
Kum C, Sekkin S (2011) The immune system drugs in fish: immune function, immunoassay, drugs. In: Aral F, Dogu Z (eds) Recent advances in fish farms, pp 169–216
Kumar V, Sahu NP, Pal AK, Kumar S, Sinha AK, Ranjan J, Baruah K (2010) Modulation of key enzymes of glycolysis, gluconeogenesis, amino acid catabolism, and TCA cycle of the tropical freshwater fish Labeo rohita fed gelatinized and non-gelatinized starch diet. Fish Physiol Biochem 36:491–499
Levey AS, Becker C, Inker LA (2015) Glomerular filtration rate and albuminuria for detection and staging of acute and chronic kidney disease in adults: a systematic review. JAMA 313:837–846
Levillain O, Parvy P, Hassler C (1997) Amino acid handling in uremic rats: citrulline, a reliable marker of renal insufficiency and proximal tubular dysfunction. Metabolism 46:611–618
Li P, Wu G (2018) Roles of dietary glycine, proline, and hydroxyproline in collagen synthesis and animal growth. Amino Acids 50:29–38
Li P, Wu G (2020) Composition of amino acids and related nitrogenous nutrients in feedstuffs for animal diets. Amino Acids 52:523–542
Li XL, Zheng SX, Wu G (2020) Nutrition and metabolism of glutamate and glutamine in fish. Amino Acids 52:671–691
Lim VS, Yarasheski KE, Flanigan MJ (1998) The effect of uraemia, acidosis, and dialysis treatment on protein metabolism: a longitudinal leucine kinetic study. Nephrol Dial Transplant 13:1723–1730
Lowry M, Hall DE, Brosnan JT (1985) Hydroxyproline metabolism by the rat kidney: distribution of renal enzymes of hydroxyproline catabolism and renal conversion of hydroxyproline to glycine and serine. Metabolism 34:955–961
Mahasen LM (2016) Evolution of the kidney. Anat Physiol Biochem Int J 1:555554
Manna P, Sinha M, Sil PC (2009) Taurine plays a beneficial role against cadmium-induced oxidative renal dysfunction. Amino Acids 36:417–428
Marescau B, Qureshi IA, De Deyn P, Letarte J, Ryba R, Lowenthal A (1985) Guanidino compounds in plasma, urine and cerebrospinal fluid of hyperargininemic patients during therapy. Clin Chim Acta 146:21–27
Martin WF, Armstrong LE, Rodriguez NR (2005) Dietary protein intake and renal function. Nutr Metab 2:25
May RC, Hara Y, Kelly RA, Block KP, Buse MG, Mitch WE (1987) Branched-chain amino acid metabolism in rat muscle: abnormal regulation in acidosis. Am J Phys 252:E712–E718
McDonald KM, Miller PD, Anderson RJ, Berl T, Schrier RW (1976) Hormonal control of renal water excretion. Kidney Int 10:38–45
McGee MM, Greengard O, Knox WE (1972) The quantitative determination of phenylalanine hydroxylase in rat tissues. Its developmental formation in liver. Biochem J 127:669–674
McGuire DM, Gross MD, Van Pilsum JF, Towle HC (1984) Repression of rat kidney L-arginine: glycine amidinotransferase synthesis by creatine at a pretranslational level. J Biol Chem 259:12034–12038
McNeal CJ, Meininger CJ, Wilborn CD, Tekwe CD, Wu G (2018) Safety of dietary supplementation with arginine in adult humans. Amino Acids 50:1215-1229
Meyer C, Stumvoll M, Nadkarni V, Dostou J, Mitrakou A, Gerich J (1998) Abnormal renal and hepatic glucose metabolism in type 2 diabetes mellitus. J Clin Invest 102:619–624
Mistry SK, Greenfeld Z, Morris SM Jr, Baylis C (2002) The ‘intestinal–renal’arginine biosynthetic axis in the aging rat. Mech Ageing Dev 123:1159–1165
Mitrakou A (2011) Kidney: its impact on glucose homeostasis and hormonal regulation. Diabetes Res Clin Pract 93:S66–S72
Møller N, Meek S, Bigelow M, Andrews J, Nair KS (2000) The kidney is an important site for in vivo phenylalanine-to-tyrosine conversion in adult humans: a metabolic role of the kidney. Proc Natl Acad Sci U S A 97:1242–1246
Moon TW, Foster GD. Tissue carbohydrate metabolism, gluconeogenesis and hormonal and environmental influences. In: Biochemistry and molecular biology of fishes, PW Hochachka and TP Mommsen. 1995; 4: 65-100. Elsevier, New York
Morris SM Jr (2016) Arginine metabolism revisited. J Nutr 146:2579S–2586S
National Research Council (2011) Nutrient requirements of fish and shrimp. National Academies Press, Washington, DC
Ochoa JB, Strange J, Kearney P, Gellin G, Endean E, Fitzpatrick E (2001) Effects of L-arginine on the proliferation of T lymphocyte subpopulations. J Parent Enteral Nutr 25:23–29
Ogawa T, Kimoto M, Watanabe H, Sasaoka K (1987) Metabolism of NG, NG-and NG, NG-dimethylarginine in rats. Arch Biochem Biophys 252:526–537
Palacín M, Estévez R, Bertran J, Zorzano A (1998) Molecular biology of mammalian plasma membrane amino acid transporters. Physiol Rev 78:969–1054
Pandey G, Madhuri S (2014) Heavy metals causing toxicity in animals and fishes. Res J Anim Vet Fish Sci 2:17–23
Pedrini MT, Levey AS, Lau J, Chalmers TC, Wang PH (1996) The effect of dietary protein restriction on the progression of diabetic and nondiabetic renal diseases: a meta-analysis. Ann Intern Med 124:627–632
Perry SF, Shahsavarani A, Georgalis T, Bayaa M, Furimsky M, Thomas SL (2003) Channels, pumps, and exchangers in the gill and kidney of freshwater fishes: their role in ionic and acid-base regulation. J Exp Zool A 300:53–62
Pillai S, Verrey F (2019) Dietary amino acids affect the rate of chronic kidney disease progression in rats. FASEB J 33(Suppl 1):570.1
Pohlenz C, Buentello A, Criscitiello MF, Mwangi W, Smith R, Gatlin DM III (2012) Synergies between vaccination and dietary arginine and glutamine supplementation improve the immune response of channel catfish against Edwardsiella ictaluri. Fish Shellfish Immunol 33:543–551
Pollock AS (1989) Induction of renal phosphoenolpyruvate carboxykinase mRNA: suppressive effect of glucose. Am J Phys 257:F145–F151
Price SR, Wang XI, Bailey JL (1998) Tissue-specific responses of branched-chain alpha-ketoacid dehydrogenase activity in metabolic acidosis. J Am Soc Nephrol 9:1892–1898
Reddy ST, Wang CY, Sakhaee K, Brinkley L, Pak CY (2002) Effect of low-carbohydrate high-protein diets on acid-base balance, stone-forming propensity, and calcium metabolism. Am J Kidney Dis 40:265–274
Reyes AA, Karl IE, Klahr SA (1994) Role of arginine in health and in renal disease. Am J Phys 267:F331–F346
Rogers QR, Phang JM (1985) Deficiency of pyrroline-5-carboxylate synthase in the intestinal mucosa of the cat. J Nutr 115:146–150
Ryan WL, Wells IC (1964) Homocitrulline and homoarginine synthesis from lysine. Science 144:1122–1123
Ryan WL, Barak AJ, Johnson RJ (1968) Lysine, homocitrulline, and homoarginine metabolism by the isolated perfused rat liver. Arch Biochem Biophys 123:294–297
Ryan WL, Johnson RJ, Dimari S (1969) Homoarginine synthesis by rat kidney. Arch Biochem Biophys 131:521–526
Sasaki M, Sasako T, Kubota N, Sakurai Y, Takamoto I, Kubota T, Inagi R, Seki G, Goto M, Ueki K, Nangaku M (2017) Dual regulation of gluconeogenesis by insulin and glucose in the proximal tubules of the kidney. Diabetes 66:2339–2350
Schnyder G, Roffi M, Flammer Y, Pin R, Hess OM (2002) Effect of homocysteine-lowering therapy with folic acid, vitamin B12, and vitamin B6 on clinical outcome after percutaneous coronary intervention: the Swiss Heart study: a randomized controlled trial. JAMA 288:973–979
Schoolwerth AC, Smith BC, Culpepper RM (1988) Renal gluconeogenesis. Miner Electrolyte Metab 14:347–361
Schrock H, Goldstein LE (1981) Interorgan relationships for glutamine metabolism in normal and acidotic rats. Am J Phys 240:E519–E525
Seely JC (2017) A brief review of kidney development, maturation, developmental abnormalities, and drug toxicity: juvenile animal relevancy. J Toxicol Pathol 30:125–133
Shen CS, Mistry SP (1979) Development of gluconeogenic, glycolytic, and pentose-shunt enzymes in the chicken kidney. Poult Sci 58:663–667
Shoemaker C, Xu DH, LaFrentz B, LaPatra S (2015) Overview of fish immune system and infectious diseases. In: Lee CS, CLim DM, III G, Webster CD (eds) Dietary nutrients, additives and fish health. Wiley, Hoboken, pp 1–24
Sibal L, Agarwal SC, Home PD, Boger RH (2010) The role of asymmetric dimethylarginine (ADMA) in endothelial dysfunction and cardiovascular disease. Curr Cardiol Rev 6:82–90
Silbernagl ST (1988) The renal handling of amino acids and oligopeptides. Physiol Rev 68:911–1007
Silva P, Rosen S, Spokes K, Epstein FH (1991) Effect of glycine on medullary thick ascending limb injury in perfused kidneys. Kidney Int 39:653–658
Silva RP, Clow K, Brosnan JT, Brosnan ME (2014) Synthesis of guanidinoacetate and creatine from amino acids by rat pancreas. Br J Nutr 111:571–577
Sipilä I (1980) Inhibition of arginine-glycine amidinotransferase by ornithine. A possible mechanism for the muscular and chorioretinal atrophies in gyrate atrophy of the choroid and retina with hyperornithinemia. Biochim Biophys Acta 613:79–84
Skov AR, Toubro S, Bülow J, Krabbe K, Parving HH, Astrup A (1999) Changes in renal function during weight loss induced by high vs low-protein low-fat diets in overweight subjects. Int J Obesity 23:1170–1177
Stam F, van Guldener C, Wee PM, Kulik W, Smith DE, Jakobs C, Stehouwer CD, de Meer K (2004) Homocysteine clearance and methylation flux rates in health and end-stage renal disease: association with S-adenosylhomocysteine. Am J Phys 287:F215–F223
Stead LM, Brosnan JT, Brosnan ME, Vance DE, Jacobs RL (2006) Is it time to reevaluate methyl balance in humans? Am J Clin Nutr 83:5–10
Stühlinger MC, Tsao PS, Her JH, Kimoto M, Balint RF, Cooke JP (2001) Homocysteine impairs the nitric oxide synthase pathway: role of asymmetric dimethylarginine. Circulation 104:2569–2575
Stumvoll M, Meyer C, Mitrakou A, Nadkarni V, Gerich JE (1997) Renal glucose production and utilization: new aspects in humans. Diabetologia 40:749–757
Stumvoll M, Meyer C, Perriello G, Kreider M, Welle S, Gerich J (1998) Human kidney and liver gluconeogenesis: evidence for organ substrate selectivity. Am J Phys 274:E817–E826
Stumvoll M, Perriello G, Meyer C, Gerich J (1999) Role of glutamine in human carbohydrate metabolism in kidney and other tissues. Kidney Int 55:778–792
Sun MS, Pan CJ, Shieh JJ, Ghosh A, Chen LY, Mansfield BC, Ward JM, Byrne BJ, Chou JY (2002) Sustained hepatic and renal glucose-6-phosphatase expression corrects glycogen storage disease type la in mice. Hum Mol Genet 11:2155–2164
Tangri N, Stevens LA, Griffith J, Tighiouart H, Djurdjev O, Naimark D, Levin A, Levey AS (2011) A predictive model for progression of chronic kidney disease to kidney failure. JAMA 305:1553–1559
Tinker DA, Brosnan JT, Herzberg GR (1986) Interorgan metabolism of amino acids, glucose, lactate, glycerol and uric acid in the domestic fowl (Gallus domesticus). Biochem J 240:829–836
Tizianello A, Deferrari G, Garibotto G, Robaudo C, Lutman M, Passerone G, Bruzzone M (1983) Branched-chain amino acid metabolism in chronic renal failure. Kidney Int 16:S17–S22
Trachtman H, Futterweit S, Maesaka J, Ma C, Valderrama E, Fuchs A, Tarectecan AA, Rao PS, Sturman JA, Boles TH (1995) Taurine ameliorates chronic streptozocin-induced diabetic nephropathy in rats. Am J Phys 269:F429–F438
Triscari J, Stern JS, Johnson PR, Sullivan AC (1979) Carbohydrate metabolism in lean and obese Zucker rats. Metabolism 28:183–189
Tsikas D, Wu G (2015) Homoarginine, arginine, and relatives: analysis, metabolism, transport, physiology, and pathology. Amino Acids 47:1697–1702
Tsikas D, Bollenbach A, Hanff E, Kayacelebi AA (2018) Asymmetric dimethylarginine (ADMA), symmetric dimethylarginine (SDMA) and homoarginine (hArg): the ADMA, SDMA and hArg paradoxes. Cardiovasc Diabetol 17:1
Upadhyay K, Silverstein DM (2014) Renal development: a complex process dependent on inductive interaction. Curr Pediatr Rev 10:107–114
Van De Poll MC, Soeters PB, Deutz NE, Fearon KC, Dejong CH (2004) Renal metabolism of amino acids: its role in interorgan amino acid exchange. Am J Clin Nutr 79:185–197
Vercoutère B, Durozard D, Baverel G, Martin G (2004) Complexity of glutamine metabolism in kidney tubules from fed and fasted rats. Biochem J 378:485–495
Verrey F, Ristic Z, Romeo E, Ramadan T, Makrides V, Dave MH, Wagner CA, Camargo SM (2005) Novel renal amino acid transporters. Annu Rev Physiol 67:557–572
Verrey F, Singer D, Ramadan T, Vuille-dit-Bille RN, Mariotta L, Camargo SM (2009) Kidney amino acid transport. Pflugers Arch 458:53–60
Wang W, Wu Z, Dai Z, Yang Y, Wang J, Wu G (2013) Glycine metabolism in animals and humans: implications for nutrition and health. Amino Acids 45:463–477
Wang W, Wu Z, Lin G, Hu S, Wang B, Dai Z, Wu G (2014a) Glycine stimulates protein synthesis and inhibits oxidative stress in pig small intestinal epithelial cells. J Nutr 144:1540–1548
Wang W, Dai Z, Wu Z, Lin G, Jia S, Hu S, Dahanayaka S, Wu G (2014b) Glycine is a nutritionally essential amino acid for maximal growth of milk-fed young pigs. Amino Acids 46:2037–2045
Watford M, Hod Y, Chiao YB, Utter MF, Hanson RW (1981) The unique role of the kidney in gluconeogenesis in the chicken. The significance of a cytosolic form of phosphoenolpyruvate carboxykinase. J Biol Chem 256:10023–10027
Watowich SS (2011) The erythropoietin receptor: molecular structure and hematopoietic signaling pathways. J Investig Med 59:1067–1072
Webster AC, Nagler EV, Morton RL, Masson P (2017) Chronic kidney disease. Lancet 389:1238–1252
Weiner ID, Mitch WE, Sands JM (2015) Urea and ammonia metabolism and the control of renal nitrogen excretion. Clin J Am Soc Nephrol 10:1444–1458
Weinstein JR, Anderson S (2010) The aging kidney: physiological changes. Adv Chronic Kidney Dis 17:302–307
Welbourne TC (1974) Ammonia production and pathways of glutamine metabolism in the isolated perfused rat kidney. Am J Phys 226:544–548
Welbourne TC (1987) Interorgan glutamine flow in metabolic acidosis. Am J Phys 253:F1069–F1076
Wetzel MD, Gao T, Venkatachalam M, Morris SM Jr, Awad AS (2019) L-Homoarginine supplementation prevents diabetic kidney damage. Physiol Rep 7:e14235
Wieczorek-Surdacka E, Hanff E, Chyrchel B, Kuźniewski M, Surdacki A, Tsikas D (2019) Distinct associations between plasma osteoprotegerin, homoarginine and asymmetric dimethylarginine in chronic kidney disease male patients with coronary artery disease. Amino Acids 51:977–982
Wirthensohn GA, Guder WG (1986) Renal substrate metabolism. Physiol Rev 66:469–497
Wu G, Knabe DA (1995) Arginine synthesis in enterocytes of neonatal pigs. Am J Physiol 269:R621-R629
Wu G (1997) Synthesis of citrulline and arginine from proline in enterocytes of postnatal pigs. Am J Phys 272:G1382–G1390
Wu G (2013a) Amino acids: biochemistry and nutrition. CRC Press, Boca Raton
Wu G (2013b) Functional amino acids in nutrition and health. Amino Acids 45:407–411
Wu G (2016) Dietary protein intake and human health. Food Funct 7:1251–1265
Wu G (2020a) Management of metabolic disorders (including metabolic diseases) in ruminant and nonruminant animals. In: Bazer FW, Lamb GC, Wu G (eds) Animal agriculture: challenges, innovations, and sustainability. Elsevier, New York, pp 471–492
Wu G (2020b) Important roles of dietary taurine, creatine, carnosine, anserine and hydroxyproline in human nutrition and health. Amino Acids 52:329-360
Wu G, Morris SM (1998) Arginine metabolism: nitric oxide and beyond. Biochem J 336:1–17
Wu G, Meininger CJ, Knabe DA, Baze FW, Rhoads MJ (2000) Arginine nutrition in development, health and disease. Curr Opin Clin Nutr Metab Care 3:59–66
Wu G, Bazer FW, Davis TA, Kim SW, Li P, Rhoads JM, Satterfield MC, Smith SB, Spencer TE, Yin Y (2009) Arginine metabolism and nutrition in growth, health and disease. Amino Acids 37:153–168
Wu ZL, Hou YQ, Hu SD, Bazer FW, Meininger CJ, McNeal CJ, Wu G (2016) Catabolism and safety of supplemental L-arginine in animals. Amino Acids 48:1541–1552
Wu G (2018) Principles of Animal Nutrition. CRC Press, Boca Raton, Florid
Wu ZL, Hou YQ, Dai ZL, Hu CA, Wu G (2019) Metabolism, nutrition, and redox signaling of hydroxyproline. Antioxid Redox Signal 30:674–682
Wyss M, Kaddurah-Daouk R (2000) Creatine and creatinine metabolism. Physiol Rev 80:1107–1213
Yánez AJ, Nualart F, Droppelmann C, Bertinat R, Brito M, Concha II, Slebe JC (2003) Broad expression of fructose-1, 6-bisphosphatase and phosphoenolpyruvate carboxykinase provide evidence for gluconeogenesis in human tissues other than liver and kidney. J Cell Physiol 197:189–197
Yin M, Zhong Z, Connor HD, Bunzendahl H, Finn WF, Rusyn I, Li X, Raleigh JA, Mason RP, Thurman RG (2002) Protective effect of glycine on renal injury induced by ischemia-reperfusion in vivo. Am J Phys 282:F417–F423
Young GA (1991) Amino acids and the kidney. Amino Acids 1:183–192
Zhou CP, Song F, Wu G (2018) Catabolism of branched-chain amino acids in tissues of hybrid striped bass (Morone chrysops x M. Saxatilis). Aquaculture America Annual Meeting, Las Vegas, Feb. 19–22, 2018
Acknowledgments
We thank students and research assistants in our laboratory for helpful discussions. Financial support by Guangdong Yeuhai Feeds Group Co., Ltd. is gratefully appreciated.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Li, X., Zheng, S., Wu, G. (2020). Amino Acid Metabolism in the Kidneys: Nutritional and Physiological Significance. In: Wu, G. (eds) Amino Acids in Nutrition and Health. Advances in Experimental Medicine and Biology, vol 1265. Springer, Cham. https://doi.org/10.1007/978-3-030-45328-2_5
Download citation
DOI: https://doi.org/10.1007/978-3-030-45328-2_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-45327-5
Online ISBN: 978-3-030-45328-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)