
Abstract
During spermatogenesis, pluripotent germ cells differentiate to become efficient delivery vehicles to the oocyte of paternal DNA. Though male and female germ cells both undergo meiosis to produce haploid complements of DNA, at the same time they also each undergo distinct differentiation processes that result in either sperm or oocytes. This review will discuss our current understanding of mechanisms of sperm formation and differentiation in Caenorhabditis elegans gained from studies that employ a combination of molecular, transcriptomic, and cell biological approaches. Many of these processes also occur during spermatogenesis in other organisms but with differences in timing, molecular machinery, and morphology. In C. elegans, sperm differentiation is implemented by varied modes of gene regulation, including the genomic organization of genes important for sperm formation, the generation of sperm-specific small RNAs, and the interplay of specific transcriptional activators. As sperm formation progresses, chromatin is systematically remodeled to allow first for the implementation of differentiation programs, then for sperm-specific DNA packaging required for transit of paternal genetic and epigenetic information. Sperm also exhibit distinctive features of meiotic progression, including the formation of a unique karyosome state and the centrosomal-based segregation of chromosomes during symmetric meiotic divisions. Sperm-specific organelles are also assembled and remodeled as cells complete meiosis and individualize in preparation for activation, morphogenesis, and the acquisition of motility. Finally, in addition to DNA, sperm contribute specific cellular factors that contribute to successful embryogenesis.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Differentiation
- Chromosome segregation
- Paternal epigenetics
- Meiosis
- Organelle morphogenesis
- Spermiogenesis
- Sperm maturation
- Transcriptional regulation
7.1 Introduction
Although sperm and oocytes share a mission to contribute a haploid genome to the new embryo, each has specialized functions. Small sperm are streamlined to efficiently navigate the female reproductive tract whereas large oocytes are packed with maternal materials necessary for embryonic development. Thus, the production of functionally and morphologically distinct gametes requires a precise interplay between shared and distinctive aspects of their differentiation programs. In this review we will discuss recent advances that inform our current understanding of processes required for sperm formation, particularly the spermatogenesis-specific features of germ cell formation. Our discussion will generally follow the temporal progression of spermatogenesis, while highlighting how individual processes overlap and influence one another. In particular, this chapter will emphasize the varied modes of gene regulation that underlie sperm differentiation, the systematic remodeling of chromatin required for differentiation and delivery of paternal epigenetic information, the sperm-specific features of meiotic progression, the assembly and remodeling of sperm-specific organelles, the regulated process of sperm activation, and the acquisition of cell motility. Many of these processes also occur during spermatogenesis in other organisms but with differences in timing, molecular machinery, and morphology that will be discussed as relevant.
The rapidly expanding toolkit for studying Caenorhabditis elegans has uncovered a broad range of molecular mechanisms important for sperm formation. Early ultrastructural studies of the male gonad and isolation of spermatogenesis-defective genetic mutants helped define the stages of sperm formation and laid a foundation for recent genomic and transcriptomic approaches. Technological advances in RNA interference (RNAi) and reverse genetic deletion mutant screening methods are now facilitating the analysis of candidate gene function in various stages of sperm formation and fertilization. Likewise, cytological studies of gamete formation have benefited from advances in imaging techniques and the ability to make transgenic animals that express fluorescently labeled fusion proteins. Unlike previous reviews of C. elegans spermatogenesis that have focused on insights gained from genetic mutants (L’Hernault 2006; Nelson et al. 1982), this review will integrate what has been learned about molecular mechanisms of C. elegans spermatogenesis using a myriad of experimental approaches.
7.2 Brief Overview of Sperm Formation in C. elegans
C. elegans has many features that make it an ideal model for studying spermatogenesis. Approximately half the adult body mass of both males and hermaphrodites is devoted to germ cell formation and both sexes make sperm. Hermaphrodites generate approximately 300 sperm during the last larval stage before switching to exclusively producing oocytes. Hermaphrodites are self-fertile; sperm that they make during their last larval stage are stored within one of two spermathecae, where they fertilize passing oocytes during each ovulation event. In contrast, males continue to make sperm, which they transfer to hermaphrodites through mating (Ward and Carrel 1979). Relative to hermaphrodite sperm, male sperm are both larger and more competitive (LaMunyon and Ward 1995, 1998); however, the information provided in this review will apply to sperm formation in both sexes unless otherwise noted.
Germ cell formation can be easily observed through the transparent cuticle of both sexes in fixed and live samples (L’Hernault 2006; McCarter et al. 1999; Shakes et al. 2009). Because gamete formation occurs in a linear progression along the length of the gonad (Seydoux and Schedl 2001), germ cells can be staged cytologically by their location within the gonad arm, their nuclear morphology, and the presence of distinct marker proteins, which can be assessed by immunostaining (Figs. 7.1a and 7.2). In the subsections below we will give a brief overview of the key events that occur within each of these zones shown in Figs. 7.1a and 7.2. These subsections are intended to place the events of spermatogenesis in context of other chapters in this book and to introduce readers to the key events of spermatogenesis before devoting the remainder of the chapter to an in-depth discussion of many of these sperm-specific features.

The progression of spermatid formation and pseudopod assembly. (a) Changes in nuclear morphology during spermatid formation. A single-armed wild-type male gonad visualized using DAPI and fluorescence microscopy. Regions of the gonad are labeled: mitotic, meiotic entry, and the transition zone (blue) and pachytene, the condensation zone, meiotic division zone, and haploid spermatids (purple). A nucleus exhibiting the karysome morphology is indicated in yellow. Scale bar represent 50 μm. (b) Key stages in FB-MO and MSP dynamics. The process of spermiogenesis includes budding, maturation, and activation. (1) A spermatocyte in diakinesis (before nuclear envelope breakdown) contains multiple, fully mature fibrous body (FB)–membranous organelle (MO) complexes. The major sperm protein (MSP) (green) is assembled into FBs that are enveloped by the arms of the MO. The MO head is the vesicle below the electron-dense collar (two dark bars). (2) After the MI and MII divisions the late-stage budding spermatid is fully polarized with the FB-MOs and chromatin masses partitioned to the spermatids and the intact spindle microtubules partitioned to the central residual body. (3) In an early maturing spermatid, the arms of the MO retract as the FBs are released into the cytoplasm and begin to disassemble. (4) A late-stage quiescent spermatid in which the MOs are docked and MSP is cytosolic. (5) Upon exposure to an activator, spermatids initially form microspikes as the MOs begin to fuse at the collar with the plasma membrane. (6) Motile spermatozoon with a distinct cell body containing fused MOs and MSP-filled pseudopod

Overview of the overlapping events that occur during late meiotic prophase of sperm cell formation. (a) Schematic of the progression of male germline cells (blue) during spermatogenesis. Cells are attached to the rachis through karyosome formation, then bud off of the rachis to undergo meiotic divisions. After anaphase II, haploid cells bud from residual bodies to form spermatids. (b) The corresponding chromatin morphology of cells highlighted in (a). DAPI-stained and schematic drawings (red) of the nuclear morphology of cells in the stages of late meiotic prophase indicated. (c) Staging of sperm cells can also be monitored by the presence of specific cell structures, organelles, and macromolecules, which are represented as blue bars
7.2.1 Mitotic Region
Somatic cells, called distal tip cells, define the most distal end of the gonad (Fig. 7.1a). In hermaphrodites, one distal tip cell is positioned at each end of the two gonad arms while males have both distal tip cells positioned at the end of the single-armed gonad (Kimble and Crittenden 2007; Byrd and Kimble 2009). Adjacent to the distal tip cell(s), sexually uncommitted germ cells undergo repeated rounds of mitotic duplication. The molecular mechanisms responsible for specifying the size and extent of this mitotic proliferation zone are covered in Chap. 4 (Hansen and Schedl 2012). Surprisingly, recent studies reveal that mitotically proliferating germ cells divide more rapidly in male gonads than in hermaphrodite gonads, an observation that suggests that the germ cells are sexually dimorphic even before they fully commit to an oocyte or sperm fate (Morgan et al. 2010).
7.2.2 Meiotic Entry, the Sperm/Oocyte Switch, and Early Meiotic Prophase
Exit from the mitotic region (Fig. 7.1a) is tightly coordinated with two events that are covered in other chapters of this series: transition to meiosis in Chap. 4 (Hansen and Schedl 2012) and sex determination in Chap. 3 (Zanetti and Puoti 2012). The molecular events that drive the early stages of meiotic development, including chromosome pairing, synapsis, and recombination are thought to occur similarly in both males and hermaphrodites and are described in Chap. 6 (Lui and Colaiácovo 2012). Conversely, the programs of C. elegans oogenesis and spermatogenesis differ in that spermatocytes progress through meiotic prophase at a faster rate (20–24 h rather than 54–60 h) (Jaramillo-Lambert et al. 2007) and, unlike oocytes, are not subjected to a checkpoint for DNA damage and meiotic recombination errors that results in damaged cells being removal by apoptosis (Gartner et al. 2000; Jaramillo-Lambert et al. 2010). However, to date, the earliest marker of spermatogenesis is the recently discovered transcriptional regulator, SPE-44, which can first be observed on the chromatin of sperm but not oocyte producing germ lines during early pachytene (Figs. 7.2c and 7.4) and will be discussed in Sect. 7.3.3.
7.2.3 Late Meiotic Prophase and the Meiotic Divisions
By late meiotic prophase, several features distinguish sperm- and oocyte-producing germ lines. During late pachytene and diplotene, spermatogenesis-enriched genes are being robustly transcribed and sperm-specific structures are being assembled within cells (Figs. 7.1a and 7.2). Then, immediately prior to the meiotic divisions, the spermatocytes become transcriptionally inactive and enter a sperm-specific karyosome stage, which is described in more detail in Sect. 7.4.1 (Fig. 7.1a) (Shakes et al. 2009).
During this time, the chromosomes of both oocytes and spermatocytes resolve and condense in preparation for meiotic divisions. However several aspects of these events differ in oocytes and spermatocytes, particularly in regard to events in late pachytene through the condensation zone (Figs. 7.1a and 7.2). First, shortly before initiation of the meiotic divisions, the spermatocyte chromatin becomes differentially structured through the incorporation of sperm-specific nuclear basic proteins (SNBPs) (Chu et al. 2006). Second, in contrast to oocytes that mature in response to extracellular signals (Kim et al. 2012), spermatocytes proceed directly from meiotic prophase into the meiotic divisions. Third, unlike developing oocytes, which lose their centrioles during meiotic prophase and subsequently segregate their chromosomes on an acentriolar spindle (Albertson and Thomson 1993; Kim and Roy 2006; Peters et al. 2010; Wignall and Villeneuve 2009), developing spermatocytes retain their centrioles and segregate their chromosomes on centriole-based spindles. Since the spermatocyte spindles are nucleated by centrosomes rather than through a chromatin-mediated mechanism, spindle assembly can initiate earlier in spermatocytes, before the breakdown of the nuclear envelope (Shakes et al. 2009). Lastly, in oocytes, these divisions are asymmetric and yield a single gamete whereas in spermatocytes, the divisions are symmetric and yield four equally sized gametes (L’Hernault 2006).
7.2.4 Spermiogenesis and Motility
After the completion of the meiotic divisions, spermatids develop into motile spermatozoa, a process referred to as spermiogenesis (Fig. 7.1b). We define this process as beginning immediately after anaphase II and involving three major phases. In an initial budding phase, a combination of polarization and spermatid budding mechanisms result in the separation of individual spermatids from a central residual body, in which materials that are unnecessary for the subsequent development and function of the sperm are discarded. In the subsequent, poorly characterized maturation phase, the sperm chromatin becomes surrounded by an electron-dense, RNA-enriched perinuclear halo (Ward et al. 1981). Sperm-specific complexes and organelles, which house components required for sperm motility and fertilization, also remodel in preparation for an extended quiescent state. In a final activation phase, quiescent, spherical sperm transform into bipolar and motile spermatozoa upon exposure to sex-specific extracellular signaling molecules. Motility of the crawling spermatozoa is driven by the regulated assembly and disassembly of a nematode-specific cytoskeletal protein known as the Major Sperm Protein (MSP).
Exciting advances in our understanding of some of the molecular mechanisms required for the specification of sperm formation during each of these phases are summarized in Fig. 7.2.
7.3 Genomics and Transcriptomics: Programming the Genome for Sperm Differentiation
Our understanding of how germline stem cells differentiate into motile spermatozoa has benefited from diverse experimental approaches. “Systems-wide” approaches have not only identified a comprehensive list of genes whose expression is “spermatogenesis-enriched,” but are also enabling the exploration of how their global transcriptional profiles are modified under different experiment conditions or between mutant backgrounds. In one key approach (Reinke et al. 2000, 2004), factors that contribute to sperm formation were identified through pair-wise comparisons of the expression profiles of mutant hermaphrodites that produce only oocytes (fem-1 lf), only sperm (fem-3 gf), or completely lack a germ line (glp-4(bn2)) (Barton et al. 1987; Beanan and Strome 1992; Nelson et al. 1978). In another, sperm-enriched factors were identified by comparing protein or RNA profiles from mutants that produce only sperm or only oocytes (Chu et al. 2006; Han et al. 2009). These studies are revealing the variety of molecular mechanisms that regulate expression of spermatogenesis-enriched genes. In this section, we will discuss four levels of regulation: (1) the organization and genetic structure of spermatogenesis-enriched genes within the genome, (2) sperm-specific histones and other basic nuclear proteins that modify the structure of the sperm chromatin, (3) regulatory transcription factors that govern the large-scale implementation of the sperm differentiation program, and (4) sperm-specific small RNAs at work during sperm differentiation.
7.3.1 Organization and Genetic Structure of Spermatogenesis-Enriched Genes
Several genome-wide expression studies have identified genes whose transcription is differentially regulated during spermatogenesis (Reinke et al. 2000, 2004; Bamps and Hope 2008; Maeda et al. 2001; Wang et al. 2009). These microarray studies distinguished genes with sperm-enriched expression from others, including germ line-intrinsic, oocyte-enriched, and somatic-enriched. Though all germ cell-specific classes have commonalities, sperm genes exhibit several distinct features. For example, the X chromosome contains relatively few genes that are expressed in either the germ line or the developing gametes, chromosome IV contains an abundance of sperm-enriched genes, and chromosome I contains an over-representation of germline-enriched genes. These finding suggests that genes may exhibit large- and small-scale clustering patterns within the genome that aid in their coordinated regulation. Supporting this hypothesis, an integrated analysis of germline microarray and in situ expression data revealed that many germline and oocyte genes are tightly clustered in small groups within operons (Reinke and Cutter 2009). In contrast, genes expressed during spermatogenesis were largely excluded from operons, indicating they do not exhibit small-scale clustering. Interestingly, germ line-intrinsic and oocyte-enriched genes exhibit numerous similarities in temporal regulation, response to RNAi, and the functional classes of their protein products, whereas sperm enriched genes have distinct temporal expression profiles, are generally refractory to RNAi, and encode many novel protein products. Taken together, these studies suggest that the gene expression program necessary for the execution of the sperm fate involves large-scale changes in chromatin that are aided, in part, by the genomic organization of spermatogenesis-enriched genes.
7.3.2 Chromatin Organization and Global Transcription Activation Are Coupled
Throughout spermatogenesis, the processes of meiosis, sperm differentiation, and chromatin remodeling are intimately intertwined. One distinctive feature of chromatin remodeling during spermatogenesis is the introduction of SNBPs that regulate transcription (Bettegowda and Wilkinson 2011). For example, in mammals, high levels of transcription are promoted in meiotic cells by histone acetylation and incorporation of histone variants (Lewis et al. 2003; Sassone-Corsi 2002). During this time, sperm-specific transcriptional regulatory factors must access DNA in order to implement specific differentiation programs (Kimmins et al. 2004). Once meiosis is completed, somatic histones are replaced in a stepwise fashion: first by other sperm-specific histone variants, then by transition proteins, and ultimately by protamine proteins (Kimmins and Sassone-Corsi 2005; Govin et al. 2004; Braun 2001). As a result, transcription becomes globally repressed as DNA becomes increasingly compacted. The repackaging of sperm DNA is hypothesized to streamline the sperm DNA cargo for efficient mobility, protect the DNA package from potential environmental damage, and carry potential paternal epigenetic information to the nascent embryo (Caron et al. 2005; Miller et al. 2010; Wu and Chu 2008).
In C. elegans, candidate SNBPs have been identified through comparisons of the proteomic profiles of sperm and oocyte chromatin (Chu et al. 2006). These include a sperm-specific histone H2A variant called HTAS-1 (H Two A Sperm) and three putative protamines called SPCH-1, 2, and 3 (SPerm CHromatin), all of which initially incorporate into chromosomes during late pachytene and remain associated with mature sperm chromatin (Chu et al. 2006) (Figs. 7.2 and 7.3a). Overall, the incorporation of these SNBPs correlates with global transcriptional down-regulation; levels of RNA polymerase II actively engaged in transcriptional elongation remain elevated through mid-pachytene but then decrease significantly as chromosomes condense in preparation for meiotic divisions (Fig. 7.3b) (Shakes et al. 2009). However, it has yet to be determined the extent to which subsets of genes required for sperm formation may escape transcriptional down-regulation. When compared to the process in mammals, the incorporation of SNBPs into spermatocyte chromatin during meiosis may seem precocious. However, we hypothesize that this early incorporation of SNBPs may have evolved to support the comparatively rapid progression of C. elegans spermatogenesis. One consequence of early SNBP incorporation is that the meiotic chromosomes of C. elegans spermatocytes may be distinct from those of either oocytes or mitotically dividing germ cells. In addition, the shutdown of transcription prior to the meiotic divisions implies that both the progression of sperm morphogenesis and sperm activation must be driven solely by post-transcription regulatory mechanisms.

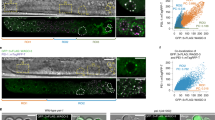
Changes in transcriptional regulation correlate with the alteration of chromatin composition. Immunolocalization of (a) the sperm-specific histone H2A variant, HTAS-1 (red) and (b) elongating RNA polymerase II (phosphorylated on the C-terminal domain on serine 2, detected using Abcam H5 antibody ab24758) (green). Regions of the male germ line are indicated. DNA is shown in blue. (a) HTAS-1 incorporates into sperm chromatin as cells condense for meiotic divisions. (b) High levels of actively elongating RNA polymerase (green) decrease dramatically as chromosomes condense for meiotic divisions, indicating global transcriptional activation is curtailed by the karyosome stage
7.3.3 Transcription Factors That Coordinate Sperm Differentiation
The role of individual transcription factors identified from the list of spermatogenesis-enriched genes has also revealed coordinated regulation of sperm formation. For example, the transcription factor SPE-44 was identified from the list of 1,343 spermatogenesis-enriched genes (Reinke 2002; Reinke et al. 2000, 2004; Kulkarni et al. 2012). A homozygous mutant of spe-44 is spermatogenesis defective (spe). Evidence suggests that SPE-44 functions as an early global transcriptional activator required for spermatocyte differentiation (Kulkarni et al. 2012). First, SPE-44 is expressed exclusively during spermatogenesis in nuclei of undifferentiated spermatocytes, well before levels of the sperm-specific protein MSP accumulate (Fig. 7.4). During pachytene, SPE-44 localizes strongly on autosomes but not the X chromosome, which is largely transcriptionally silent during spermatogenesis (Kelly et al. 2002; Reinke et al. 2000). Microarray analysis also reveals that two-thirds of the more than 500 genes whose expression is down-regulated in spe-44 mutants were previously classified as spermatogenesis-enriched genes (Reinke 2002; Reinke et al. 2000, 2004; Kulkarni et al. 2012). As a result, spe-44 mutants produce arrested spermatocytes with defects in multiple sperm-specific processes including the assembly of FBs within spermatocytes and the spermatid budding process that follows anaphase II. These results suggest that SPE-44 may function, presumably in conjunction with other transcription factors, as a key, sperm-specific transcriptional regulator of sperm differentiation.

Sperm-specific transcription factors and the expression of sperm proteins. Immuno-localization of (a) the SPE-44 transcription factor (red) and (b) the Major Sperm Protein, MSP (green) within an isolated and fixed male gonad. DNA is shown in blue. Regions of the male germ line are indicated. (a) SPE-44 (green) is expressed early in pachytene until chromosomes condense for meiotic divisions. (b) MSP is synthesized beginning in pachytene and subsequently localizes to distinct FBs, which localize as oblong stripes,within the condensation zone. MSP partitions to the spermatids in FBs but in mature, quiescent spermatids (far right) it disassembles and fills the cytoplasm
In another example, the promoters of spermatogenesis-enriched genes were compared with those of all other C. elegans genes in order to identify enriched DNA sequence motifs that could act as transcription factor binding sites (del Castillo-Olivares et al. 2009). A subset of 45 sperm-enriched genes was found to share a single bipartite consensus that was then used as a binding target in a yeast 1-hybrid screen for potential sperm-specific transcription factors (del Castillo-Olivares et al. 2009; Klass et al. 1988; Shim et al. 1995). This screen identified a single GATA-type transcription factor, ELT-1, that regulates hypodermal expression in the embryo but is also highly expressed in the male germ line (Spieth et al. 1991). In sperm producing germ lines elt-1 mRNA is present beginning from mid-pachytene; in oocyte producing germ lines, it is undetectable. Reduction of ELT-1 in either RNAi mediated knockdown or knockout mosaics resulted in defects in sperm production, including post-meiotic morphological and motility defects (del Castillo-Olivares et al. 2009). ELT-1 is also amongst the genes that are down-regulated in absence of SPE-44 (Kulkarni et al. 2012). Importantly, no phenotypes were observed in oocytes, though affected worms exhibited additional somatic defects, consistent with the role of ELT-1 in other somatic tissues (del Castillo-Olivares et al. 2009). This role for ELT-1 in C. elegans spermatogenesis is consistent with roles of GATA transcription factor family members in the regulation of mammalian sperm development (Ketola et al. 1999, 2000).
7.3.4 Regulation of Sperm Formation by Sperm-Specific Small RNAs
Studies in C. elegans of small non-coding RNAs, important regulators of mRNA degradation, translational repression, and chromatin structure, have revealed they contribute to the proper regulation of large groups of sperm genes. Three classes of small RNAs are known: microRNAs (miRNAs), Piwi-interacting RNAs (piRNAs), and endogeneous small interfering RNAs (endo-siRNAs) (Kim et al. 2009). Each class is generated by a distinct mechanism and each employs a distinct effector pathway to regulate gene expression (Suh and Blelloch 2011). Temperature-sensitive roles in C. elegans spermatogenesis have been found for specific subclasses of endo-siRNAs and piRNAs.
7.3.4.1 piRNAs
piRNAs have evolutionarily conserved functions in male fertility (Aravin et al. 2006; Houwing et al. 2007). In mammals, piRNAs are expressed in both pachytene spermatocytes and round spermatids—stages that are blocked by mutations in Piwi proteins, a distinct family of Argonaut proteins (Deng and Lin 2002; Kuramochi-Miyagawa et al. 2004). piRNAs, which do not require Dicer for their generation, are predominantly expressed in the germ line and are complementary to genomic repeat sequences (Aravin et al. 2006; Grivna et al. 2006). piRNAs in Drosophila are associated with the repression of transposable elements during spermatogenesis (Malone and Hannon 2009). Thus, a primary function ascribed to piRNAs is to guard the integrity of the genome in the germ line from foreign invaders like transposons.
In C. elegans, piRNAs function in both sperm and oocyte-producing germ lines. Slightly shorter than the typical 25–33 nucleotide piRNAs in other organisms, C. elegans piRNAs are 21 base pairs in length, begin with U, and physically associate with the Piwi protein PRG-1 (Han et al. 2009; Kato et al. 2009; Ruby et al. 2006; Batista et al. 2008; Das et al. 2008). The expression of these 21U piRNAs is restricted to the germ line, and strains with null mutations in either of the two Piwi homologs, prg-1 and prg-2, result in temperature-sensitive mitotic and meiotic germline defects. Thus the Piwi protein PRG-1 is important for robust thermotolerance during spermatogenesis (Batista et al. 2008; Wang and Reinke 2008). At 25°C, prg-1 males produce spermatocytes but very few spermatids, and the spermatids that are produced are defective in sperm activation (Wang and Reinke 2008). In the male germ line, PRG-1 localizes in a perinuclear fashion to the germline P-granules in developing spermatocytes but disappears as the chromatin condenses following pachytene (Batista et al. 2008). This localization to P-granules is intriguing as other P-granule associated factors have been reported to exhibit analogous, maternal-specific, temperature-sensitive, null-phenotypes (Smith et al. 2002; Spike et al. 2008).
Microarray analysis of prg-1 single mutants shows that a subset of sperm-enriched transcripts are affected by lack of PRG-1; though studies conflict as to whether PRG-1 upregulates or represses these genes (Batista et al. 2008; Wang and Reinke 2008). Targets of piRNAs include the Tc3 transposon, whose transposition is elevated at least a 100-fold in prg-1; prg-2 double mutants (Batista et al. 2008; Das et al. 2008). This repression of transposon activity suggests piRNAs may function in maintaining the genomic stability of germ cells, a role consistent with that characterized for piRNAs in other organisms. The mechanism for how piRNAs inhibit transposition in any organism is not yet understood.
C. elegans piRNAs have distinct features from those in other organisms. Besides being shorter, they have an upstream eight nucleotide core sequence, suggesting they are individually transcribed, not processed from a transcript of clustered piRNAs genes as seen in other organisms (Das et al. 2008). C. elegans piRNAs also lack any overlap, suggesting a different mode of amplification than that proposed in other organisms (Das et al. 2008; Aravin et al. 2007; Brennecke et al. 2007; Gunawardane et al. 2007). Thus mechanisms of small RNA regulation for germline function may have both conserved and adaptive features.
7.3.4.2 Endo-siRNAs
Characterization of siRNA function in the C. elegans germ line revealed an important role for small RNAs not only in response to foreign DNA but also in endogeneous gene regulation. Endo-siRNAs are generated from long dsRNAs that either base pair with complementary dsRNAs or form long hairpins (Okamura and Lai 2008; Golden et al. 2008). In C. elegans, response to long double-stranded RNAs involves two phases. First primary siRNAs are generated via cleavage by the endonuclease Dicer (Bernstein et al. 2001; Grishok et al. 2001; Ketting et al. 2001; Knight and Bass 2001). Then secondary siRNAs are produced by RNA-directed RNA polymerases (RdRPs), which amplify the silencing response (Aoki et al. 2007; Sijen et al. 2001). Post-transcriptional silencing is achieved as siRNAs partner with specific Argonaute proteins as part of the RISC complex, which degrades target mRNAs. The mechanism of silencing depends on the complementarity between the endo-siRNA and target mRNA: perfect base-pairing to the target mRNA results in mRNA cleavage while imperfect base-pairing causes silencing by an unknown mechanism (Zeng and Cullen 2003).
Sperm and oocytes have distinct classes of endo-siRNAs (Han et al. 2009). Deep sequencing of purified sperm, oocytes, and embryos in C. elegans revealed sperm and oocyte-specific classes of 26 nucleotide long small RNAs with a strong bias for a G as the first nucleotide (Han et al. 2009). Both classes of 26G RNAs are exact complements to target mRNAs and their generation requires members of the C. elegans RISC complex, including Dicer (DCR-1), an RdRP called RRF-3, and Dicer associated factors called ERI-1 and ERI-3 (Pavelec et al. 2009; Gent et al. 2009; Han et al. 2009). Oocyte-specific 26G RNAs are maternally inherited and silence gene expression during zygotic development (Han et al. 2009). In contrast, targets of the sperm-specific 26G RNAs include the sperm-enriched transcripts identified by previous germline microarray data (Reinke et al. 2000, 2004; Gent et al. 2009; Han et al. 2009). This suggests that sperm-specific 26G RNAs function to down-regulate the levels of sperm-enriched genes (Han et al. 2009). Indeed, these particular sperm genes are significantly upregulated in eri-1 and rrf-3 mutants (Gent et al. 2009; Han et al. 2009).
In an analogous fashion to the Piwi protein PRG-1, the RISC complex members eri-1, eri-3, rrf-3, and dcr-1 are required for spermatogenesis at elevated temperatures (Pavelec et al. 2009; Gent et al. 2009). At 25°C, mutant males undergo improper cytokinesis and chromosome segregation and produce misshapen spermatids that contain abnormal chromatin masses and excess tubulin. Although most of these mutant spermatids fail to activate, the motile spermatozoa that are formed can fertilize oocytes. However, the resulting embryos are nonviable (Gent et al. 2009). During oogenesis, Dicer is also required for the proper organization of the hermaphrodite germ line during the pachytene stage, and dcr-1 null hermaphrodites produce irregularly shaped, nonfunctional, endomitotic oocytes (Knight and Bass 2001). As such, unlike eri-1 and rrf-3, Dicer functions more broadly in both siRNA and miRNA pathways (Knight and Bass 2001) and has recently been shown to regulate ribonucleoprotein assembly in the hermaphrodite germ line (Beshore et al. 2011). In contrast, although eri-1 and rrf-3 are required for 26G RNA production in both spermatocytes and oocytes, loss of either eri-1 or rrf-3 causes no discernable detects in either oocytes or the resulting embryos (Pavelec et al. 2009; Gent et al. 2009). Why the loss of sperm-specific 26G RNAs results in male infertility while loss of oocyte 26Gs does not adversely affect oogenesis or embryonic development is yet unknown.
How does the RISC complex generate different sperm and oocyte targets? Distinct Argonaute protein components confer specificity to the RISC complex (Han et al. 2009). The Argonaut ERGO-1 is required for expression of oocyte but not sperm 26G RNAs; however, depletion of ERGO-1 results in near wild-type fertility. In contrast, depletion of two Argonaut proteins, T22B3.2 and ZK757.3, specifically decreases the expression of 26G RNAs in sperm but not in oocytes (Han et al. 2009; Conine et al. 2010) and results in thermosensitive male infertility. Future studies that investigate either the role of specific sperm targets or alternative functions for RISC complex members in male fertility promise to provide mechanistic insights regarding how 26G RNAs function specifically in the context of sperm development and fertility.
7.4 Meiotic Progression: Sex-Specific Features of Preparing for and Undergoing Meiotic Divisions
As summarized in the overview (Sect. 7.2.3), oocytes and spermatocytes undergo meiotic divisions that differ in the timing of production and the number and size of their end products. Underlying these difference are gamete-specific differences in chromatin condensation, cell cycle progression, and kinetochore structure.
7.4.1 Sex-Specific Differences in Preparing Chromosomes for Segregation
Though the mechanisms required to facilitate recombination appear similar in both sexes, the resolution of chromosomes in preparation for meiotic divisions differs. Following the pachytene stage, SYP-1, a component of the synaptonemal complex required for pairing, synapsis, and recombination, is removed earlier from spermatocyte chromosomes than from oocyte chromosomes (Chap. 6, Lui and Colaiácovo 2012; Shakes et al. 2009). Chromatin composition may also influence meiotic events. In other organisms, sperm chromatin is tightly compacted through the incorporation of SNBPs during a prolonged post-meiotic differentiation stage. However, in C. elegans SNBPs are incorporated as spermatocytes exit pachytene (Chu et al. 2006). This pre-loading of chromatin structural elements may allow for the efficient compaction of mature C. elegans sperm chromatin even in the absence of a prolonged post-meiotic processing period (Shakes et al. 2009).
Another striking feature is the formation of a karyosome, in which spermatocyte chromosomes retain their structural organization but come together to form a single, constricted mass (Shakes et al. 2009). In contrast, oocyte chromosomes remain as individual entities as they condense before meiotic divisions. Karyosome formation occurs in a broad range of animals; however, it more typically occurs during oogenesis with the hypothesized purpose of facilitating chromosome remodeling prior to the meiotic divisions (Gruzova and Parfenov 1993; Orr-Weaver et al. 1995; Sanyal et al. 1976). In C. elegans spermatocytes, karyosome formation occurs after desynapsis and the global down-regulation of transcription but before nuclear envelope breakdown (Shakes et al. 2009). The function of karyosome formation remains poorly understood in any organism, but is hypothesized to facilitate rapid progression to metaphase I after breakdown of the nuclear envelope.
C. elegans spermatocytes and oocytes also differ in how they transition through the cell cycle in preparation for chromosome segregation. In both, the chromatin is phosphorylated on histone H3 (HisH3-ser10) as the chromosomes condense prior to meiotic divisions. This phosphorylation is mediated by the aurora kinase (AIR-2), which regulates both kinetochores and release of chromosome cohesion during meiosis (Rogers et al. 2002; Hsu et al. 2000; McCarter et al. 1999; Burrows et al. 2006; Schumacher et al. 1998). However, during late prophase of oogenesis, AIR-2 and phosphosphorylated HisH3-ser10 can only be detected on the chromosomes of diakinetic oocytes that have received an MSP-based signal from sperm to undergo oocyte maturation. In contrast, during spermatogenesis, HisH3-ser10 phosphorylation occurs earlier, during the diplotene stage and before AIR-2 is present (Shakes et al. 2009). In addition, AIR-2 and phosphorylated HisH3-ser10 exhibit gamete-specific localization patterns during the diplotene, karyosome, and diakinesis stages (Shakes et al. 2009). These results suggest that, in diplotene spermatocytes, an additional kinase may be necessary to phosphorylate HisH3-ser10. In summary, when compared to the process during oogenesis, chromosome condensation during spermatogenesis initiates both more rapidly and, potentially, independently of an extracellular signal.
7.4.2 Sex-Specific Similarities and Differences in Kinetochore Structure
The presence of centrioles in sperm but not oocyte meiosis necessitates alterations in the mechanics of chromosome segregation. During oocyte meiosis, kinetochores assemble as cup-shaped structures, which are thought to enable the sides of tightly compacted rounded chromosomes to interface between organized microtubule bundles (Albertson and Thomson 1993; Maddox et al. 2004; Monen et al. 2005; Wignall and Villeneuve 2009; Schvarzstein et al. 2010). However, it is equally plausible that this cup-shaped morphology addresses the shared challenge of segregating homologs rather than sister chromatids during the first meiotic division. After all, despite dramatic differences in their interactions with microtubules, the kinetochores of spermatocytes are likewise cup-shaped and thus more similar in overall structure to those in oocytes (Albertson and Thomson 1993; Howe et al. 2001; Shakes et al. 2009) than to the mitotic kinetochores, which localize in a poleward fashion along the length of mitotic chromosomes (Dernburg 2001; Kitagawa 2009; Maddox et al. 2004). Importantly, oocytes and spermatocyte kinetochores do exhibit notable differences in the specific arrangement of individual inner and outer kinetochore components, including CENP-CHCP-4, HCP-1, and HCP-2 (Shakes et al. 2009). These differences may facilitate attachment to structurally distinct microtubule spindles or may be a consequence of the incorporation of SNBPs in sperm chromatin (Shakes et al. 2009). Overall, many aspects of the mechanics of chromosome segregation during sperm meiosis remain to be resolved.
7.5 The Assembly and Function of Sperm-Specific Organelles: Packing for the Trip
In all species, sperm cells change morphology during the later stages of spermatogenesis. In mammals, this differentiation process is supported by a post-meiotic burst of sperm-specific transcription and translation. In contrast, in C. elegans global transcriptional activation ceases shortly before the meiotic divisions (Shakes et al. 2009). Furthermore, translation largely halts after the meiotic divisions when translational machinery is discarded in the residual body (Ward et al. 1981). As a result, C. elegans sperm assemble modular, “pre-fabricated” sperm-specific organelles prior to the meiotic divisions, which are subsequently modified and regulated during the course of differentiation by post-translational modifications.
Striking features of developing C. elegans spermatocytes are the sperm-specific FB–MO complexes (Roberts et al. 1986; Ward et al. 1981; Wolf et al. 1978). FB–MO complexes can first be observed during late pachytene, expanding in size through the meiotic divisions (Figs. 7.1b, 7.4b and 7.5b). The FBs sequester newly synthesized filaments of the MSP in a paracrystaline-like state. The MO component is a Golgi-derived organelle with three regions: a glycoprotein-filled “head” portion, a body with a highly convoluted membrane morphology that presumably maximizes its surface area, and an electron-dense collar that separates these two domains (Fig. 7.1b). In developing spermatocytes, the FBs and MOs are intimately associated; the microvillus-like arms of the MO body envelope the growing FBs. Following anaphase II, FB–MO complexes then partition to the budding spermatids. After the spermatids detach from the residual body, the MO membranes retract from and release the FB. At that point the detached MOs dock with the plasma membrane and the FBs disassemble.

Comparison of spermiogenesis in (a) vertebrates and (b) C. elegans highlighting analogous events. (a) In vertebrates, following anaphase II, spermatocytes undergo incomplete cytokinesis to generate four, interconnected haploid spermatids. These spermatids then undergo a multi-week maturation process of spermiogenesis that includes the following events: a burst of sperm-specific transcription and translation, the formation of a mature acrosome, the mature flagellum, and the compaction and reshaping of the nucleus. Materials unneeded by the spermatozoon are then partitioned into a residual body (RB) as the spermatozoon completes cellularization. Sperm activation causes a spermatozoon to become fully motile. (b) In C. elegans, following anaphase II, spermatocytes initiate a cleavage furrow that regresses and morphs into a polarization and budding process during which time unneeded materials are partitioned away from the differentiating sperm and left in a central residual body as spermatids detach. During a short (minutes-long) maturation step the MOs mature and dock, the FBs disassociate and subsequently disassemble, and an RNA-enriched perinuclear halo forms around the compact chromatin mass. Male spermatids are stored in this quiescent state until stimulated by extracellular signals to active and form bipolar, motile spermatozoa. Both hermaphrodite and male sperm activation occurs in less than 10 minutes
Our understanding of the FBs is expanding with the discovery of genes required for FB assembly/disassembly dynamics. To date, the assembly of MSP into FBs is known to require both a member of the casein kinase I superfamily, spe-6 (Muhlrad and Ward 2002; Varkey et al. 1993) and SPE-7, a spermatogenesis-enriched cytosolic protein which seems to function as an essential structural and scaffolding component (M. Presler, K. Messina, and D. Shakes, unpublished data). In terms of FB disassembly, it has previously been thought MSP is released from FBs immediately upon the detachment of the haploid sperm from the residual body (L’Hernault 2006; Roberts et al. 1986). However, others have observed a sizable and distinct population of spermatids in which FBs have released from the MOs but have not fully disassembled (Wu et al. 2012, D. Greenstein, and D. Shakes, unpublished data). This suggests that the disassembly of MSP into individual dimers and their dispersal throughout the cytoplasm involves a more gradual “maturation” process. Complete disassembly of FBs is not essential for subsequent pseudopod formation; however, mutants that fail to fully disassemble their FBs have small and inefficient pseudopods (Ward et al. 1981). In a potential counterbalance to SPE-6, two paralogs of PP1 phosphatase (GSP-3 and GSP-4) have recently been shown to be essential for FB disassembly (Wu et al. 2012). Thus, the emerging picture is that FB assembly and disassembly may be regulated by a combination of scaffolding proteins such as SPE-7 and post-translational modifiers like SPE-6 and GSP-3/4.
Within C. elegans spermatocytes, the assembly of MOs likely involves homotypic vacuolar fusions, since MOs fail to form in spermatocytes that lack the HOPS complex protein, SPE-39 (Zhu and L’Hernault 2003; Zhu et al. 2009). In the absence of SPE-39, spermatocytes fill with large numbers of small vesicles and “naked” FBs. Although most spe-39 spermatocytes attempt both the two meiotic divisions, they fail to undergo cytokinesis after meiosis I. After anaphase II, they attempt a normal budding division but fail to polarize and are only able to form abnormally small spermatid buds.
Analysis of specific spe mutants suggests that close association of the MOs and FBs facilitates the partitioning of the MSP-rich FBs into spermatids and away from the residual body. For example, in spe-39 mutants, defects in MO assembly result in the formation of “naked” FBs that, in the absence of associated MOs, fail to properly partition to spermatids during the budding division (Zhu and L’Hernault 2003). Similar defects are observed when FB–MO complexes disassociate prematurely as in spe-4 (Arduengo et al. 1998) and spe-10 mutants (Gleason et al. 2006; Shakes and Ward 1989). SPE-4 encodes a member of the presenilin family and is thought to function in the regulated cleavage of adjacent integral membrane proteins, as presenilin does for the amyloid precursor protein, APP. In spe-4 mutants, the cleavage of specific integral membrane proteins is presumably dysregulated. As a result, spe-4 spermatocytes can undergo normal meiotic chromosome segregation but are unable to either polarize or initiate the budding division following anaphase II (Arduengo et al. 1998; L’Hernault and Arduengo 1992). In the absence of the MO-localized, transmembrane palmitoyl transferase protein SPE-10, the MOs effectively partition to spermatids; however, the “naked” FBs are left behind in the residual body where they either remain or bud off directly as small cytoplasts (Gleason et al. 2006; Shakes and Ward 1989). Studies of sperm from other nematode species may also differentiate the functions of FB from MOs. For example, ultrastructural studies (Justine 2002; Justine and Jamieson 2000) reveal that spermatocytes in certain other nematode species either exhibit morphologically distinct MO-structures (Turpeenniemi 1998; Shepherd and Clark 1983) or lack MOs altogether (Lee and Anya 1967; Shepherd and Clark 1983; Yushin and Commans 2005).
Further studies in both C. elegans and other nematode species are needed to better understand the intricacies of FB assembly and disassembly, as well as the seemingly parallel process of MSP assembly and disassembly within the pseudopod of crawling spermatozoa. In particular, there is evidence in C. elegans that genes required for FB assembly and disassembly, like spe-6 and the PP1 phosphatases gsp-3 and gsp-4, function again later in both sperm activation and MSP-based pseudopod motility (Muhlrad and Ward 2002; Wu et al. 2012). Furthermore, independent biochemical studies of Ascaris spermatozoa have convincingly shown that the rapid assembly of MSP complexes at the leading edge of the pseudopod is mediated by a combination of integral membrane phosphoproteins, cytosolic proteins, and the casein I kinase-related protein called MSP polymerization-activating kinase (MPAK) (Chap. 11, Marcello et al. 2012; Buttery et al. 2003; Italiano et al. 1996; LeClaire et al. 2003; Aitken and De Iuliis 2007), while the disassembly of MSP at the base of the pseudopod is mediated by a PP2 phosphatase (Yi et al. 2007).
7.6 Spermiogenesis
Spermiogenesis refers to the post-meiotic processes that convert sessile haploid spermatids into motile spermatozoa. In both insects and vertebrates, spermiogenesis involves the formation of an acrosome, hypercondensation of the chromatin, construction of the flagella, elimination of excess cytoplasm, and separation of individual sperm from the larger syncytium. In most organisms, these processes take days to weeks to complete and are supported by a final post-meiotic burst of sperm-specific transcription and an extended period of protein synthesis. In C. elegans, the term “spermiogenesis” has previously been used in reference to the 10-min transformation of fully cellularized spherical spermatids into bipolar crawling spermatozoa (L’Hernault 2006; Shakes and Ward 1989; Ward et al. 1983). However, in order to draw more informative parallels between the developmental events in C. elegans and other organisms, including vertebrates, here we define spermiogenesis as including three phases (Fig. 7.5). After anaphase II, spermatids first individualize by budding and forming a residual body. The second phase, which we term “spermatid maturation,” directly follows anaphase II and includes polarization of cellular components, spermatid budding, and maturation of nuclear and cytoplasmic components. The later phase, which we refer to as “sperm activation,” includes the conversion of quiescent, spherical spermatids into motile, bipolar spermatozoa.
7.6.1 Polarization and Budding
At the completion of anaphase II, a shallow cleavage furrow initiates at the mid-point between the two haploid nuclei that have segregated to opposite poles via centrosome-organized spindles (Fig. 7.5b) (Shakes et al. 2009, 2011; Ward et al. 1981). However this transient furrow rapidly regresses as the cells bud from a central residual body. Cellular components are then differentially partitioned into either the bud or residual body. Actin, endoplasmic reticulum, and ribosomes accumulate within the residual body while the FB–MO complexes and mitochondria partition along with the nuclei into the budding spermatids (Ward 1986). Towards the end of this polarization process, the microtubules detach from the centrioles and move into the residual body. We propose that this polarization is analogous to the cytoplasmic shedding events that typically occur at the end of spermiogenesis as individual spermatids detach from the syncytium in other organisms (Fig. 7.5a, b).
The molecular mechanisms that underlie the spermatid budding division and the associated polarization events remain poorly understood. Analysis of spe mutants suggests that the two events are regulated by distinct actin-mediated processes. Both are disrupted in spermatocytes deficient in the actin-binding, kelch-related protein SPE-26 (Varkey et al. 1995), whereas spermatocytes that lack SPE-15 (myosin VI) can bud but are unable to differentially partition their cellular components (Kelleher et al. 2000). The constriction forces associated with spermatid bud formation remain unknown; ultrastructural studies indicate that the constriction points between the buds and residual body are devoid of either an actin/myosin ring or microtubules (Ward et al. 1983). Centrosomes and microtubules may set up the initial patterning of the budding division as the number of spermatid buds correlates with the number of centrosomes in spermatocytes that over-replicate their centrosomes (Peters et al. 2010). Interestingly, both polarization and spermatid budding can be uncoupled from cell cycle progression beyond metaphase I (Golden et al. 2000). Analysis of mutants with temperature-sensitive defects in the anaphase-promoting complex (APC/C) revealed that budding can proceed despite the presence of a stabilized arrested metaphase I spindle (Golden et al. 2000). Such mutant budding yield residual bodies that contain an intact metaphase spindle (Golden et al. 2000), and spermatids that lack chromatin but can nevertheless activate, crawl, and fertilize oocytes (Sadler and Shakes 2000). It has yet to be determined whether either spermatid budding or polarization can occur in the absence of assembled microtubules. Although much remains to be learned about both the polarization and budding events, their striking physical and functional similarities to the process of “sperm individualization” in other organisms suggest that comparative studies between the two will be informative for both (Fig. 7.5a, b).
7.6.2 Spermatid Maturation
By analogy to the events of spermiogenesis in other organisms, the “maturation” events of C. elegans spermiogenesis include the remodeling and docking of the MOs, the disassembly of MSP from the FBs, the final remodeling of chromatin, and the formation of an RNA-enriched perinuclear halo (Fig. 7.5b). Although spermatid maturation has not been a widely used term in the C. elegans literature, we suggest that it provides a useful term to describe the numerous cellular changes that occur in spermatids from the time that they separate from the residual body until they are either stored in a quiescent state within the male seminal vesicle or are signaled by sperm activators to undergo morphogenesis.
7.6.3 Sperm Activation and Cellular Morphogenesis
Unlike other events in spermatogenesis, sperm activation is triggered by extracellular signals, which have yet to be identified. When activated in vitro spermatids can be observed to first develop several long, thin microspikes around the spherical cell before fully polarizing to form a distinct pseudopod and cell body (Shakes and Ward 1989; Ward et al. 1983; Fig. 7.1b). During this time, the docked MOs fuse with the plasma membrane in the cell body in a manner that releases soluble glycoproteins from the head while the electron-dense collar of the MO establishes a permanent fusion pore with the plasma membrane that opens into the invaginated, pocket-like membrane of the MO body (Fig. 7.1b) (Nelson and Ward 1980; Wolf et al. 1978).
To date, the extracellular activators and underlying the cellular response machinery remain incompletely understood. Early studies revealed that spermatids isolated from C. elegans males could be activated in vitro by the addition of either the protease Pronase or substances that elevated the intracellular pH (triethanolamine or the N + K + ionophore monesin) (Nelson and Ward 1980; Shakes and Ward 1989). More recently, wortmannin was identified as another potent activator, suggesting a potential role for PI(3,4,5)P3 signaling in the process (Bae et al. 2009). Although all of these in vitro activators, particularly Pronase, continue to be used in an experimental context, their in vivo molecular targets remain undefined.
Genetic screens have revealed sex-specific activation mechanisms (L’Hernault et al. 1988; Minniti et al. 1996; Nance et al. 1999, 2000; Shakes and Ward 1989; Geldziler et al. 2005). For example, mutations in the so-called “spe-8 class” genes disrupt sperm activation in affected hermaphrodites but not in their male siblings. These mutants share complex phenotypic traits with two features. First, mutant hermaphrodites are self-sterile because the sperm they produce do not activate unless exposed to and activated “in trans” by male seminal fluid from either wild-type or spe-8 class mutant males. Second, the mutant males are fertile and their sperm activate normally both in vivo and in response to activators that raise the intracellular pH; however, their sperm arrest with microspikes when exposed to Pronase. The “spe-8 class” of genes encodes diverse proteins: SPE-8 is a non-receptor tyrosine kinase with an SH2 domain, SPE-27 is a soluble protein and SPE-12, SPE-19, and SPE-29 are all transmembrane proteins. These initial studies suggested that male and hermaphrodite sperm were normally activated by distinct sex-specific activators but that both male and hermaphrodite sperm redundantly express the cellular machinery required to respond to either activator (Fig. 7.6a).

Signaling and response elements involved in sperm activation. (a) Diagram of a two-pathway model for sperm activation. The SWM-1 protease inhibitor is present in both males and hermaphrodites, but its only known target to date is the male-specific protease TRY-5. The action of TRY-5 as a component of a male-specific activation pathway may be direct or indirect. The cellular response pathway downstream of TRY-5 (Pathway “X”) is present in both male and hermaphrodite sperm. The SPE-8 group components comprise a second cellular response pathway, which is also present in both male and hermaphrodite sperm. The in vivo activator of this SPE-8 pathway has yet to be either molecularly or mutationally identified. This unknown activator is definitely present in hermaphrodites, and it may or may not be redundantly present in males. The in vitro activator Pronase activates sperm via the SPE-8 pathway. In vitro activation by weak bases bypasses the absence of either TRY-5 or SPE-8 group components. The two cellular response pathways converge to inhibit the SPE-6 kinase. (b) Hypothetical model for sperm activation events (adopted from Gosney et al. 2008). Close-up views of the region indicated by the gray boxes are shown on the schematic of docked or fused MOs below. In quiescent spermatids, cytosolic SPE-6 actively phosphorylates and thus inhibits the MO membrane protein SPE-4. Upon sperm activation, signaling components activate to inhibit SPE-6. As a result, SPE-4 becomes active and can cleave FER-1. The proteolytically processed form of FER-1 promotes MO fusion with the plasma membrane
In an independent study, Stanfield and Villeneuve identified a sperm activation mutant in which only the males were infertile (Stanfield and Villeneuve 2006). In this case, the males were infertile because their sperm activated precociously within the seminal vesicle, a situation that proved to be incompatible with successful sperm transfer. Notably, swm-1 encodes an extracellular protease inhibitor, implying that a protease target of SWM-1 either serves directly as an in vivo activator or indirectly as part of an activation cascade. Conversely since swm-1 is normally expressed in both males and hermaphrodites, either hermaphrodite fertility is unaffected by precocious sperm activation or hermaphrodites lack one or more downstream elements of this SWM-1 regulated activation cascade.
In fact, one key target of SWM-1 is a male-specific, seminal fluid protease called TRY-5 (Smith and Stanfield 2011). Analysis of GFP fusion constructs revealed that TRY-5 is expressed and stored in the valve and vas deferens cells of the male gonad until it is secreted and transferred to the hermaphrodite during ejaculation. Notably, Smith and Stanfield found that males that only lack TRY-5 are fertile, whereas males that lack both TRY-5 and SPE-8 are infertile. This observation is consistent with a model in which male sperm can be activated through either of two distinct and redundant pathways. However, it leaves open the question of whether the sperm of try-5 males is being activated “in trans” by a hermaphrodite-specific activator within the uterus or whether males redundantly express activators for both the spe-8 and pathway “X” response elements (Fig. 7.6a). A key distinction between these two models is whether the activator of the spe-8 pathway will prove to be hermaphrodite specific in its expression.
Intriguingly, these activation pathways also link back to SPE-6 (casein kinase I) and SPE-4 (presenilin), which were discussed earlier in the context of FB assembly and the regulated disassociation the FB from the MO. Specific non-null alleles of both spe-6 and spe-4 both genetically suppress the self-sterility of spe-8 class hermaphrodites and cause precocious sperm activation in males (Gosney et al. 2008; Muhlrad and Ward 2002). These results suggest that SPE-6 and SPE-4 may function as shared elements in the both the TRY-5 and SPE-8 activation pathways (Gosney et al. 2008; Smith and Stanfield 2011). In one scenario (Fig. 7.6b), soluble SPE-6 within unactivated sperm phosphorylates and thus inhibits the function of SPE-4 within the membrane of unfused MOs (Gosney et al. 2008). However sperm activation by either sperm activation pathway inhibits SPE-6, enabling SPE-4 to become active and cleave adjacent transmembrane proteins (Gosney et al. 2008).
One proposed target of SPE-4 is FER-1 (Fig. 7.6b). FER-1 is a multi-pass transmembrane protein that localizes to the MOs of unactivated spermatids but, upon MO fusion, distributes to both MO body and the plasma membrane. Multiple forms of FER-1 are produced from proteolytically processing during sperm activation (Achanzar and Ward 1997; Washington and Ward 2006). When sperm from fer-1 mutants are exposed to activators, they fail in MO fusion. fer-1 sperm are able to form pseudopods; however, they are abnormally short and fertilization incompetent. This is because proteins that are essential for the sperm–oocyte interactions, like the tetraspanin protein SPE-38, likely remain sequestered in the unfused MOs (Chatterjee et al. 2005) (Chap. 11, Marcello et al. 2012). FER-1 is suspected to be a direct participant in the fusion event since other members of the ferlin superfamily are also associated with membrane fusion events.
7.7 The Deliverables
The ultimate goal of spermatogenesis is to generate haploid spermatozoa that can find and fertilize oocytes to contribute genetically to the subsequent generation. While the details of MSP-based motility and fertilization will be covered in Chap. 11 (Marcello et al. 2012), we briefly consider here the contributions that sperm make to the embryo.
-
1.
A haploid complement of DNA with epigenetic information. A conserved feature of spermatogenesis in all organisms involves the large-scale but incomplete replacement of histones by protamines within sperm chromatin. This change accommodates a tighter packing state and protects the chromatin from genotoxic factors (Miller et al. 2009). Recent studies suggest that the remaining histones, modified in the context of sex-specific differences in gene transcription within the germ line, retain epigenetic information that continues to pattern gene transcription in both the gametes and the early embryo (Sha and Fire 2005; Arico et al. 2011).
-
2.
A centriole pair that is required to generate the first mitotic spindle. Whereas the centrosomes of oocytes are lost during oogenesis (Kim and Roy 2006), each haploid sperm inherits a single pair of centrioles (Albertson 1984; Albertson and Thomson 1993). After fertilization, this centriole pair duplicates and combines with maternal components to generate the two active centrosomes that establish the two poles of the embryo’s first mitotic spindle (Dammermann et al. 2008; Pelletier et al. 2006).
-
3.
A cue that specifies the anterior–posterior axis of the embryo. In C. elegans, the sperm cues the anterior–posterior axis. Interestingly, this cue does not reflect either the point of sperm entry (Goldstein and Hird 1996) or the paternally contributed DNA (Sadler and Shakes 2000). Instead, multiple lines of evidence suggest that polarity is specified by the paternally contributed centrosome and their nucleation of a microtubule aster on one side of the embryo (O’Connell et al. 2000; Cowan and Hyman 2004; Hamill et al. 2002; Wallenfang and Seydoux 2000). In addition, the sperm has been implicated breaking the symmetry of the acto-myosin network and initiating a Rho-mediated cortical flow by delivering a localized bolus of CYK-4, a sperm-enriched Rho-GAP (Jenkins et al. 2006), while also contributing to the localized depletion of the Rho-GEF ECT-2 in the immediate vicinity of the centrosome (Motegi and Sugimoto 2006).
-
4.
SPE-11, a sperm-supplied factor that is required for egg activation. In C. elegans, the oocyte chromosomes complete their meiotic divisions and form an impermeable eggshell only after fertilization. When wild-type oocytes are fertilized by sperm that lack SPE-11, the resulting zygotes fail to either produce polar bodies or construct a functional eggshell (Browning and Strome 1996; Hill et al. 1989; McNally and McNally 2005; Johnston et al. 2010). Within spermatozoa, the SPE-11 protein localizes to the perinuclear halo (Browning and Strome 1996), and within spe-11 mutant sperm, the perinuclear halo is structurally aberrant (Hill et al. 1989). Despite these defects, oocytes fertilized by spe-11 mutant sperm produce viable embryos if functional SPE-11 is expressed in the oocytes (Browning and Strome 1996).
-
5.
Paternal RNAs. Early cytological studies suggest that the perinuclear halo may be enriched in RNA (Ward et al. 1981). Small RNAs are part of the paternal cargo, as sperm-specific small RNAs, like the 26G endo-siRNAs described in Sect. 7.3.4.2, were identified from purified sperm (Han et al. 2009). It is yet unclear if a specific cadre of mRNAs is also carried over to the new embryo upon fertilization. Future studies are necessary to reveal the extent to which the paternal RNA component influences zygotic development.
7.8 Summary and Future Perspectives
Overall, a big picture view of what it takes to generate motile spermatozoa reveals numerous mechanisms that overlap and intertwine after cells commit to spermatogenesis.
At the level of gene expression, implementation of the spermatocyte fate likely requires a team of transcriptional regulators, including both spermatogenesis-specific genes like spe-44 and non-cell specific factors like elt-1 acting on genes whose genomic organization is distinct from that of either germline-specific or oocyte-specific genes. Equally important, the spermatogenesis program uses mechanisms to subsequently turn off global gene expression as the chromatin is packaged for long-term protection. These changes are mediated by a combination of post-translational histone modifications and the incorporation of various sperm basic nuclear proteins (SBNPs). Such changes place epigenetic marks on the chromatin that can affect gene expression of paternally inherited chromatin within in the embryo. In addition, both piRNAs and endo-siRNAs appear to play a back-up role in facilitating the appropriate repression of gene expression, particularly under conditions of stress.
In parallel, spermatocytes must accomplish meiotic chromosome segregation. A better understanding of the mechanistic differences between the meiotic divisions of spermatocytes and oocytes will help us distinguish the features that specifically facilitate the segregation of homologs in meiosis I from those that accommodate the distinct microtubule structures of meiotically dividing oocytes and spermatocytes. For example, in spermatocytes, core machinery, like centrosomes and kinetochores, are utilized differentially from oocytes. Progression of spermatocytes from meiotic prophase into M-phase is likewise distinct as it features a unique karyosome state and distinctions in the timing of desynapsis.
A fresh perspective on the events following anaphase II suggests interesting and informative analogies between the spermiogenesis programs of C. elegans amoeboid sperm and the flagellated spermatozoa of vertebrates and Drosophila. Although differentially ordered, both types include distinct phases of polarization/budding, spermatid maturation, and sperm activation that may involve a subset of analogous molecular mechanisms (Fig. 7.5). One distinction to note is the fate of centrosomes. During nematode spermiogenesis, the centrosome is specifically turned off and the microtubules are discarded whereas in vertebrates and Drosophila, the centrosome is converted into a basal body that facilitates assembly of the flagella (Li et al. 1998). Continuing investigations of centrosome dynamics and fate should provide important new insights regarding their function.
Like sperm, many other cell types, including spores and pluripotent stems cells, undergo extended periods of quiescence. Recent studies of C. elegans sperm activation suggest that a quiescent state for C. elegans sperm is essential for efficient sperm transfer and appropriate activation within hermaphrodites. Future investigations of underlying molecular mechanisms may provide important insights into the spatial and temporal regulation of differentiation and morphogenesis.
Although sperm from different organisms can be morphologically distinct, studies defining both conserved and adapted features between organisms can help identify key components required for spermatogenesis. As a result, the application of C. elegans as a model system has great potential to make further significant contributions to our understanding of the processes of chromosome segregation, differentiation, morphogenesis, and motility acquisition.
References
Achanzar WE, Ward S (1997) A nematode gene required for sperm vesicle fusion. J Cell Sci 110(Pt 9):1073–1081
Aitken RJ, De Iuliis GN (2007) Origins and consequences of DNA damage in male germ cells. Reprod Biomed Online 14(6):727–733
Albertson DG (1984) Formation of the first cleavage spindle in nematode embryos. Dev Biol 101(1):61–72
Albertson DG, Thomson JN (1993) Segregation of holocentric chromosomes at meiosis in the nematode, Caenorhabditis elegans. Chromosome Res 1(1):15–26
Aoki K, Moriguchi H, Yoshioka T, Okawa K, Tabara H (2007) In vitro analyses of the production and activity of secondary small interfering RNAs in C. elegans. EMBO J 26(24):5007–5019
Aravin A, Gaidatzis D, Pfeffer S, Lagos-Quintana M, Landgraf P, Iovino N, Morris P, Brownstein MJ, Kuramochi-Miyagawa S, Nakano T, Chien M, Russo JJ, Ju J, Sheridan R, Sander C, Zavolan M, Tuschl T (2006) A novel class of small RNAs bind to MILI protein in mouse testes. Nature 442(7099):203–207
Aravin AA, Hannon GJ, Brennecke J (2007) The Piwi-piRNA pathway provides an adaptive defense in the transposon arms race. Science 318(5851):761–764
Arduengo PM, Appleberry OK, Chuang P, L’Hernault SW (1998) The presenilin protein family member SPE-4 localizes to an ER/Golgi derived organelle and is required for proper cytoplasmic partitioning during Caenorhabditis elegans spermatogenesis. J Cell Sci 111(Pt 24):3645–3654
Arico JK, Katz DJ, van der Vlag J, Kelly WG (2011) Epigenetic patterns maintained in early Caenorhabditis elegans embryos can be established by gene activity in the parental germ cells. PLoS Genet 7(6):e1001391
Bae YK, Kim E, L’Hernault SW, Barr MM (2009) The CIL-1 PI 5-phosphatase localizes TRP Polycystins to cilia and activates sperm in C. elegans. Curr Biol 19(19):1599–1607
Bamps S, Hope IA (2008) Large-scale gene expression pattern analysis, in situ, in Caenorhabditis elegans. Brief Funct Genomic Proteomic 7(3):175–183
Barton MK, Schedl TB, Kimble J (1987) Gain-of-function mutations of fem-3, a sex-determination gene in Caenorhabditis elegans. Genetics 115(1):107–119
Batista PJ, Ruby JG, Claycomb JM, Chiang R, Fahlgren N, Kasschau KD, Chaves DA, Gu W, Vasale JJ, Duan S, Conte D Jr, Luo S, Schroth GP, Carrington JC, Bartel DP, Mello CC (2008) PRG-1 and 21U-RNAs interact to form the piRNA complex required for fertility in C. elegans. Mol Cell 31(1):67–78
Beanan MJ, Strome S (1992) Characterization of a germ-line proliferation mutation in C. elegans. Development 116(3):755–766
Bernstein E, Caudy AA, Hammond SM, Hannon GJ (2001) Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 409(6818):363–366
Beshore EL, McEwen TJ, Jud MC, Marshall JK, Schisa JA, Bennett KL (2011) C. elegans dicer interacts with the P-granule component GLH-1 and both regulate germline RNPs. Dev Biol 350(2):370–381
Bettegowda A, Wilkinson MF (2011) Transcription and post-transcriptional regulation of spermatogenesis. Philos Trans R Soc Lond B Biol Sci 365(1546):1637–1651
Braun RE (2001) Packaging paternal chromosomes with protamine. Nat Genet 28(1):10–12
Brennecke J, Aravin AA, Stark A, Dus M, Kellis M, Sachidanandam R, Hannon GJ (2007) Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 128(6):1089–1103
Browning H, Strome S (1996) A sperm-supplied factor required for embryogenesis in C. elegans. Development 122(1):391–404
Burrows AE, Sceurman BK, Kosinski ME, Richie CT, Sadler PL, Schumacher JM, Golden A (2006) The C. elegans Myt1 ortholog is required for the proper timing of oocyte maturation. Development 133(4):697–709
Buttery SM, Ekman GC, Seavy M, Stewart M, Roberts TM (2003) Dissection of the Ascaris sperm motility machinery identifies key proteins involved in major sperm protein-based amoeboid locomotion. Mol Biol Cell 14(12):5082–5088
Byrd DT, Kimble J (2009) Scratching the niche that controls Caenorhabditis elegans germline stem cells. Semin Cell Dev Biol 20(9):1107–1113
Caron C, Govin J, Rousseaux S, Khochbin S (2005) How to pack the genome for a safe trip. Prog Mol Subcell Biol 38:65–89
Chatterjee I, Richmond A, Putiri E, Shakes DC, Singson A (2005) The Caenorhabditis elegans spe-38 gene encodes a novel four-pass integral membrane protein required for sperm function at fertilization. Development 132(12):2795–2808
Chu D, Liu H, Nix P, Wu T, Ralston E, Yates J, Meyer B (2006) Sperm chromatin proteomics identifies evolutionarily conserved fertility factors. Nature 443(7107):101–105
Conine CC, Batista PJ, Gu W, Claycomb JM, Chaves DA, Shirayama M, Mello CC (2010) Argonautes ALG-3 and ALG-4 are required for spermatogenesis-specific 26 G-RNAs and thermotolerant sperm in Caenorhabditis elegans. Proc Natl Acad Sci USA 107(8):3588–3593
Cowan CR, Hyman AA (2004) Centrosomes direct cell polarity independently of microtubule assembly in C. elegans embryos. Nature 431(7004):92–96
Dammermann A, Maddox PS, Desai A, Oegema K (2008) SAS-4 is recruited to a dynamic structure in newly forming centrioles that is stabilized by the gamma-tubulin-mediated addition of centriolar microtubules. J Cell Biol 180(4):771–785
Das PP, Bagijn MP, Goldstein LD, Woolford JR, Lehrbach NJ, Sapetschnig A, Buhecha HR, Gilchrist MJ, Howe KL, Stark R, Matthews N, Berezikov E, Ketting RF, Tavare S, Miska EA (2008) Piwi and piRNAs act upstream of an endogenous siRNA pathway to suppress Tc3 transposon mobility in the Caenorhabditis elegans germline. Mol Cell 31(1):79–90
del Castillo-Olivares A, Kulkarni M, Smith HE (2009) Regulation of sperm gene expression by the GATA factor ELT-1. Dev Biol 333(2):397–408
Deng W, Lin H (2002) miwi, a murine homolog of piwi, encodes a cytoplasmic protein essential for spermatogenesis. Dev Cell 2(6):819–830
Dernburg AF (2001) Here, there, and everywhere: kinetochore function on holocentric chromosomes. J Cell Biol 153(6):F33–F38
Gartner A, Milstein S, Ahmed S, Hodgkin J, Hengartner MO (2000) A conserved checkpoint pathway mediates DNA damage—induced apoptosis and cell cycle arrest in C. elegans. Mol Cell 5(3):435–443
Geldziler B, Chatterjee I, Singson A (2005) The genetic and molecular analysis of spe-19, a gene required for sperm activation in Caenorhabditis elegans. Dev Biol 283(2):424–436
Gent JI, Schvarzstein M, Villeneuve AM, Gu SG, Jantsch V, Fire AZ, Baudrimont A (2009) A Caenorhabditis elegans RNA-directed RNA polymerase in sperm development and endogenous RNAi. Genetics 183(4):1297–1314
Gleason EJ, Lindsey WC, Kroft TL, Singson AW, L’Hernault SW (2006) spe-10 encodes a DHHC-CRD zinc-finger membrane protein required for endoplasmic reticulum/Golgi membrane morphogenesis during Caenorhabditis elegans spermatogenesis. Genetics 172(1):145–158
Golden A, Sadler PL, Wallenfang MR, Schumacher JM, Hamill DR, Bates G, Bowerman B, Seydoux G, Shakes DC (2000) Metaphase to anaphase (mat) transition-defective mutants in Caenorhabditis elegans. J Cell Biol 151(7):1469–1482
Golden DE, Gerbasi VR, Sontheimer EJ (2008) An inside job for siRNAs. Mol Cell 31(3):309–312
Goldstein B, Hird SN (1996) Specification of the anteroposterior axis in Caenorhabditis elegans. Development 122(5):1467–1474
Gosney R, Liau WS, Lamunyon CW (2008) A novel function for the presenilin family member spe-4: inhibition of spermatid activation in Caenorhabditis elegans. BMC Dev Biol 8:44
Govin J, Caron C, Lestrat C, Rousseaux S, Khochbin S (2004) The role of histones in chromatin remodelling during mammalian spermiogenesis. Eur J Biochem 271(17):3459–3469
Grishok A, Pasquinelli AE, Conte D, Li N, Parrish S, Ha I, Baillie DL, Fire A, Ruvkun G, Mello CC (2001) Genes and mechanisms related to RNA interference regulate expression of the small temporal RNAs that control C. elegans developmental timing. Cell 106(1):23–34
Grivna ST, Beyret E, Wang Z, Lin H (2006) A novel class of small RNAs in mouse spermatogenic cells. Genes Dev 20(13):1709–1714
Gruzova MN, Parfenov VN (1993) Karyosphere in oogenesis and intranuclear morphogenesis. Int Rev Cytol 144:1–52
Gunawardane LS, Saito K, Nishida KM, Miyoshi K, Kawamura Y, Nagami T, Siomi H, Siomi MC (2007) A slicer-mediated mechanism for repeat-associated siRNA 5′ end formation in Drosophila. Science 315(5818):1587–1590
Hamill DR, Severson AF, Carter JC, Bowerman B (2002) Centrosome maturation and mitotic spindle assembly in C. elegans require SPD-5, a protein with multiple coiled-coil domains. Dev Cell 3(5):673–684
Han T, Manoharan AP, Harkins TT, Bouffard P, Fitzpatrick C, Chu DS, Thierry-Mieg D, Thierry-Mieg J, Kim JK (2009) 26G endo-siRNAs regulate spermatogenic and zygotic gene expression in Caenorhabditis elegans. Proc Natl Acad Sci USA 106(44):18674–18679
Hansen D, Schedl T (2012) Stem cell proliferation versus meiotic fate decision in C. elegans. Advances in Experimental Medicine and Biology 757:71–99. (Chap. 4, this volume) Springer, New York
Hill DP, Shakes DC, Ward S, Strome S (1989) A sperm-supplied product essential for initiation of normal embryogenesis in Caenorhabditis elegans is encoded by the paternal-effect embryonic-lethal gene, spe-11. Dev Biol 136(1):154–166
Houwing S, Kamminga LM, Berezikov E, Cronembold D, Girard A, van den Elst H, Filippov DV, Blaser H, Raz E, Moens CB, Plasterk RH, Hannon GJ, Draper BW, Ketting RF (2007) A role for Piwi and piRNAs in germ cell maintenance and transposon silencing in Zebrafish. Cell 129(1):69–82
Howe M, McDonald KL, Albertson DG, Meyer BJ (2001) HIM-10 is required for kinetochore structure and function on Caenorhabditis elegans holocentric chromosomes. J Cell Biol 153(6):1227–1238
Hsu J, Sun Z, Li X, Reuben M, Tatchell K, Bishop D, Grushcow J, Brame C, Caldwell J, Hunt D, Lin R, Smith M, Allis C (2000) Mitotic phosphorylation of histone H3 is governed by Ipl1/aurora kinase and Glc7/PP1 phosphatase in budding yeast and nematodes. Cell 102(3):279–291
Italiano JE Jr, Roberts TM, Stewart M, Fontana CA (1996) Reconstitution in vitro of the motile apparatus from the amoeboid sperm of Ascaris shows that filament assembly and bundling move membranes. Cell 84(1):105–114
Jaramillo-Lambert A, Ellefson M, Villeneuve AM, Engebrecht J (2007) Differential timing of S phases, X chromosome replication, and meiotic prophase in the C. elegans germ line. Dev Biol 308(1):206–221
Jaramillo-Lambert A, Harigaya Y, Vitt J, Villeneuve A, Engebrecht J (2010) Meiotic errors activate checkpoints that improve gamete quality without triggering apoptosis in male germ cells. Curr Biol 20(23):2078–2089
Jenkins N, Saam JR, Mango SE (2006) CYK-4/GAP provides a localized cue to initiate anteroposterior polarity upon fertilization. Science 313(5791):1298–1301
Johnston WL, Krizus A, Dennis JW (2010) Eggshell chitin and chitin-interacting proteins prevent polyspermy in C. elegans. Curr Biol 20(21):1932–1937
Justine JL (2002) Male and female gametes and fertilization. In: Biology of nematodes. Taylor & Francis, London
Justine JL, Jamieson BGM (2000) Nematode, vol IX, part B. Progress in male gamete ultrastructure and phylogeny. Reproductive biology of invertebrates, vol IX, part B. Wiley, Chichester
Kato M, de Lencastre A, Pincus Z, Slack FJ (2009) Dynamic expression of small non-coding RNAs, including novel microRNAs and piRNAs/21U-RNAs, during Caenorhabditis elegans development. Genome Biol 10(5):R54
Kelleher JF, Mandell MA, Moulder G, Hill KL, L’Hernault SW, Barstead R, Titus MA (2000) Myosin VI is required for asymmetric segregation of cellular components during C. elegans spermatogenesis. Curr Biol 10(23):1489–1496
Kelly WG, Schaner CE, Dernburg AF, Lee M-H, Kim SK, Villeneuve AM, Reinke V (2002) X chromosome silencing in the germline of C. elegans. Development 129(2):479–492
Ketola I, Rahman N, Toppari J, Bielinska M, Porter-Tinge SB, Tapanainen JS, Huhtaniemi IT, Wilson DB, Heikinheimo M (1999) Expression and regulation of transcription factors GATA-4 and GATA-6 in developing mouse testis. Endocrinology 140(3):1470–1480
Ketola I, Pentikainen V, Vaskivuo T, Ilvesmaki V, Herva R, Dunkel L, Tapanainen JS, Toppari J, Heikinheimo M (2000) Expression of transcription factor GATA-4 during human testicular development and disease. J Clin Endocrinol Metab 85(10):3925–3931
Ketting RF, Fischer SE, Bernstein E, Sijen T, Hannon GJ, Plasterk RH (2001) Dicer functions in RNA interference and in synthesis of small RNA involved in developmental timing in C. elegans. Genes Dev 15(20):2654–2659
Kim DY, Roy R (2006) Cell cycle regulators control centrosome elimination during oogenesis in Caenorhabditis elegans. J Cell Biol 174(6):751–757
Kim VN, Han J, Siomi MC (2009) Biogenesis of small RNAs in animals. Nat Rev Mol Cell Biol 10(2):126–139
Kim S, Spike CA, Greenstein D (2012) Control of oocyte growth and meiotic maturation in C. elegans. Advances in Experimental Medicine and Biology 757:277–320. (Chap. 10, this volume) Springer, New York
Kimble J, Crittenden SL (2007) Control of germline stem cells, entry into meiosis, and the sperm/oocyte decision in C. elegans. Annu Rev Cell Dev Biol 23:405–433
Kimmins S, Sassone-Corsi P (2005) Chromatin remodelling and epigenetic features of germ cells. Nature 434(7033):583–589
Kimmins S, Kotaja N, Davidson I, Sassone-Corsi P (2004) Testis-specific transcription mechanisms promoting male germ-cell differentiation. Reproduction 128(1):5–12
Kitagawa R (2009) Key players in chromosome segregation in Caenorhabditis elegans. Front Biosci 14:1529–1557
Klass M, Ammons D, Ward S (1988) Conservation in the 5′ flanking sequences of transcribed members of the Caenorhabditis elegans major sperm protein gene family. J Mol Biol 199(1):15–22
Knight SW, Bass BL (2001) A role for the RNase III enzyme DCR-1 in RNA interference and germ line development in Caenorhabditis elegans. Science 293(5538):2269–2271
Kulkarni M, Shakes DC, Guevel K, Smith HE (2012) SPE-44 implements sperm cell fate. PLoS Genet. 8(4):e1002678
Kuramochi-Miyagawa S, Kimura T, Ijiri TW, Isobe T, Asada N, Fujita Y, Ikawa M, Iwai N, Okabe M, Deng W, Lin H, Matsuda Y, Nakano T (2004) Mili, a mammalian member of piwi family gene, is essential for spermatogenesis. Development 131(4):839–849
LaMunyon CW, Ward S (1995) Sperm precedence in a hermaphroditic nematode (Caenorhabditis elegans) is due to competitive superiority of male sperm. Experientia 51(8):817–823
LaMunyon CW, Ward S (1998) Larger sperm outcompete smaller sperm in the nematode Caenorhabditis elegans. Proc R Soc Lond B Biol Sci 265(1409):1997–2002
LeClaire LL 3rd, Stewart M, Roberts TM (2003) A 48 kDa integral membrane phosphoprotein orchestrates the cytoskeletal dynamics that generate amoeboid cell motility in Ascaris sperm. J Cell Sci 116(Pt 13):2655–2663
Lee DL, Anya AO (1967) The structure and development of the spermatozoon of Aspiculuris tetraptera (Nematoda). J Cell Sci 2(4):537–544
Lewis JD, Abbott DW, Ausio J (2003) A haploid affair: core histone transitions during spermatogenesis. Biochem Cell Biol 81(3):131–140
L’Hernault SW (2006) Spermatogenesis. WormBook:1–14
L’Hernault SW, Arduengo PM (1992) Mutation of a putative sperm membrane protein in Caenorhabditis elegans prevents sperm differentiation but not its associated meiotic divisions. J Cell Biol 119(1):55–68
L’Hernault SW, Shakes DC, Ward S (1988) Developmental genetics of chromosome I spermatogenesis-defective mutants in the nematode Caenorhabditis elegans. Genetics 120(2):435–452
Li K, Xu EY, Cecil JK, Turner FR, Megraw TL, Kaufman TC (1998) Drosophila centrosomin protein is required for male meiosis and assembly of the flagellar axoneme. J Cell Biol 141(2):455–467
Lui DY, Colaiácovo MP (2012) Meiotic development in C. elegans. In: Schedl T (ed) Advances in experimental medicine and biology, Chap. 6. Springer, Boston
Maddox PS, Oegema K, Desai A, Cheeseman IM (2004) “Holo”er than thou: chromosome segregation and kinetochore function in C. elegans. Chromosome Res 12(6):641–653
Maeda I, Kohara Y, Yamamoto M, Sugimoto A (2001) Large-scale analysis of gene function in Caenorhabditis elegans by high-throughput RNAi. Curr Biol 11(3):171–176
Malone CD, Hannon GJ (2009) Small RNAs as guardians of the genome. Cell 136(4):656–668
Marcello MR, Singaravelu G, Singson A (2012) Fertilization. Advances in Experimental Medicine and Biology 757:321–350. (Chap. 11, this volume) Springer, New York
McCarter J, Bartlett B, Dang T, Schedl T (1999) On the control of oocyte meiotic maturation and ovulation in Caenorhabditis elegans. Dev Biol 205(1):111–128
McNally KL, McNally FJ (2005) Fertilization initiates the transition from anaphase I to metaphase II during female meiosis in C. elegans. Dev Biol 282(1):218–230
Miller D, Brinkworth M, Iles D (2009) Paternal DNA packaging in spermatozoa: more than the sum of its parts? DNA, histones, protamines and epigenetics. Reproduction 139(2):287–301
Miller D, Brinkworth M, Iles D (2010) Paternal DNA packaging in spermatozoa: more than the sum of its parts? DNA, histones, protamines and epigenetics. Reproduction 139(2):287–301
Minniti AN, Sadler C, Ward S (1996) Genetic and molecular analysis of spe-27, a gene required for spermiogenesis in Caenorhabditis elegans hermaphrodites. Genetics 143(1):213–223
Monen J, Maddox PS, Hyndman F, Oegema K, Desai A (2005) Differential role of CENP-A in the segregation of holocentric C. elegans chromosomes during meiosis and mitosis. Nat Cell Biol 7(12):1248–1255
Morgan DE, Crittenden SL, Kimble J (2010) The C. elegans adult male germline: stem cells and sexual dimorphism. Dev Biol 346(2):204–214
Motegi F, Sugimoto A (2006) Sequential functioning of the ECT-2 RhoGEF, RHO-1 and CDC-42 establishes cell polarity in Caenorhabditis elegans embryos. Nat Cell Biol 8(9):978–985
Muhlrad PJ, Ward S (2002) Spermiogenesis initiation in Caenorhabditis elegans involves a casein kinase 1 encoded by the spe-6 gene. Genetics 161(1):143–155
Nance J, Minniti AN, Sadler C, Ward S (1999) spe-12 encodes a sperm cell surface protein that promotes spermiogenesis in Caenorhabditis elegans. Genetics 152(1):209–220
Nance J, Davis EB, Ward S (2000) spe-29 encodes a small predicted membrane protein required for the initiation of sperm activation in Caenorhabditis elegans. Genetics 156(4):1623–1633
Nelson GA, Ward S (1980) Vesicle fusion, pseudopod extension and amoeboid motility are induced in nematode spermatids by the ionophore monensin. Cell 19(2):457–464
Nelson GA, Lew KK, Ward S (1978) Intersex, a temperature-sensitive mutant of the nematode Caenorhabditis elegans. Dev Biol 66(2):386–409
Nelson GA, Roberts TM, Ward S (1982) Caenorhabditis elegans spermatozoan locomotion: amoeboid movement with almost no actin. J Cell Biol 92(1):121–131
O’Connell KF, Maxwell KN, White JG (2000) The spd-2 gene is required for polarization of the anteroposterior axis and formation of the sperm asters in the Caenorhabditis elegans zygote. Dev Biol 222(1):55–70
Okamura K, Lai EC (2008) Endogenous small interfering RNAs in animals. Nat Rev Mol Cell Biol 9(9):673–678
Orr-Weaver TL, Parfenov VN, Dudina LM, Kostiuchek DF, Gruzova MN, Parfenov V, Potchukalina G, Dudina L, Kostyuchek D, Gruzova M, Sanyal MK, Taymor ML, Berger MJ (1995) Meiosis in Drosophila: seeing is believing. Proc Natl Acad Sci USA 92 (23):10443–10449
Pavelec DM, Lachowiec J, Duchaine TF, Smith HE, Kennedy S (2009) Requirement for ERI/DICER complex in endogenous RNAi and sperm development in Caenorhabditis elegans. Genetics 183(4):1283–1295
Pelletier L, O’Toole E, Schwager A, Hyman AA, Muller-Reichert T (2006) Centriole assembly in Caenorhabditis elegans. Nature 444(7119):619–623
Peters N, Perez DE, Song MH, Liu Y, Muller-Reichert T, Caron C, Kemphues KJ, O’Connell KF (2010) Control of mitotic and meiotic centriole duplication by the Plk4-related kinase ZYG-1. J Cell Sci 123(Pt 5):795–805
Reinke V (2002) Functional exploration of the C. elegans genome using DNA microarrays. Nat Genet 32(Suppl):541–546
Reinke V, Cutter AD (2009) Germline expression influences operon organization in the Caenorhabditis elegans genome. Genetics 181(4):1219–1228
Reinke V, Smith HE, Nance J, Wang J, Van Doren C, Begley R, Jones SJ, Davis EB, Scherer S, Ward S, Kim SK (2000) A global profile of germline gene expression in C. elegans. Mol Cell 6(3):605–616
Reinke V, Gil IS, Ward S, Kazmer K (2004) Genome wide germline enriched and sex biased expression profiles in Caenorhabditis elegans. Development 131(2):311–323
Roberts TM, Pavalko FM, Ward S (1986) Membrane and cytoplasmic proteins are transported in the same organelle complex during nematode spermatogenesis. J Cell Biol 102(5):1787–1796
Rogers E, Bishop JD, Waddle JA, Schumacher JM, Lin R (2002) The aurora kinase AIR-2 functions in the release of chromosome cohesion in Caenorhabditis elegans meiosis. J Cell Biol 157(2):219–229
Ruby JG, Jan C, Player C, Axtell MJ, Lee W, Nusbaum C, Ge H, Bartel DP (2006) Large-scale sequencing reveals 21U-RNAs and additional microRNAs and endogenous siRNAs in C. elegans. Cell 127(6):1193–1207
Sadler PL, Shakes DC (2000) Anucleate Caenorhabditis elegans sperm can crawl, fertilize oocytes and direct anterior-posterior polarization of the 1-cell embryo. Development 127(2):355–366
Sanyal MK, Taymor ML, Berger MJ (1976) Cytologic features of oocytes in the adult human ovary. Fertil Steril 27(5):501–510
Sassone-Corsi P (2002) Unique chromatin remodeling and transcriptional regulation in spermatogenesis. Science 296(5576):2176–2178
Schumacher JM, Golden A, Donovan PJ (1998) AIR-2: an Aurora/Ipl1-related protein kinase associated with chromosomes and midbody microtubules is required for polar body extrusion and cytokinesis in Caenorhabditis elegans embryos. J Cell Biol 143(6):1635–1646
Schvarzstein M, Wignall SM, Villeneuve AM (2010) Coordinating cohesion, co-orientation, and congression during meiosis: lessons from holocentric chromosomes. Genes Dev 24(3):219–228
Seydoux G, Schedl T (2001) The germline in C. elegans: origins, proliferation, and silencing. Int Rev Cytol 203:139–185
Sha K, Fire A (2005) Imprinting capacity of gamete lineages in Caenorhabditis elegans. Genetics 170(4):1633–1652
Shakes DC, Ward S (1989) Mutations that disrupt the morphogenesis and localization of a sperm-specific organelle in Caenorhabditis elegans. Dev Biol 134(2):307–316
Shakes DC, Wu JC, Sadler PL, Laprade K, Moore LL, Noritake A, Chu DS (2009) Spermatogenesis-specific features of the meiotic program in Caenorhabditis elegans. PLoS Genet 5(8):e1000611
Shakes DC, Allen AK, Albert KM, Golden A (2011) emb-1 encodes the APC16 subunit of the Caenorhabditis elegans anaphase-promoting complex. Genetics 189(2):549–560
Shepherd AM, Clark SA (1983) The structure and development of the spermatozoon of Aspicularis tetraptera (Nematoda). J Cell Sci 2:537–544
Shim YH, Bonner JJ, Blumenthal T (1995) Activity of a C. elegans GATA transcription factor, ELT-1, expressed in yeast. J Mol Biol 253(5):665–676
Sijen T, Fleenor J, Simmer F, Thijssen KL, Parrish S, Timmons L, Plasterk RH, Fire A (2001) On the role of RNA amplification in dsRNA-triggered gene silencing. Cell 107(4):465–476
Smith JR, Stanfield GM (2011) TRY-5 is a sperm-activating protease in Caenorhabditis elegans seminal fluid. PLoS Genet 7(11):e1002375
Smith P, Leung-Chiu WM, Montgomery R, Orsborn A, Kuznicki K, Gressman-Coberly E, Mutapcic L, Bennett K (2002) The GLH proteins, Caenorhabditis elegans P granule components, associate with CSN-5 and KGB-1, proteins necessary for fertility, and with ZYX-1, a predicted cytoskeletal protein. Dev Biol 251(2):333–347
Spieth J, Shim YH, Lea K, Conrad R, Blumenthal T (1991) elt-1, an embryonically expressed Caenorhabditis elegans gene homologous to the GATA transcription factor family. Mol Cell Biol 11(9):4651–4659
Spike C, Meyer N, Racen E, Orsborn A, Kirchner J, Kuznicki K, Yee C, Bennett K, Strome S (2008) Genetic analysis of the Caenorhabditis elegans GLH family of P-granule proteins. Genetics 178(4):1973–1987
Stanfield GM, Villeneuve AM (2006) Regulation of sperm activation by SWM-1 is required for reproductive success of C. elegans males. Curr Biol 16(3):252–263
Suh N, Blelloch R (2011) Small RNAs in early mammalian development: from gametes to gastrulation. Development 138(9):1653–1661
Turpeenniemi TA (1998) Ultrastructure of spermatozoa in the nematode Halalaimus dimorphus (Nemata: Oxystominidae). J Nematol 30(4):391–403
Varkey JP, Jansma PL, Minniti AN, Ward S (1993) The Caenorhabditis elegans spe-6 gene is required for major sperm protein assembly and shows second site non-complementation with an unlinked deficiency. Genetics 133(1):79–86
Varkey JP, Muhlrad PJ, Minniti AN, Do B, Ward S (1995) The Caenorhabditis elegans spe-26 gene is necessary to form spermatids and encodes a protein similar to the actin-associated proteins kelch and scruin. Genes Dev 9(9):1074–1086
Wallenfang MR, Seydoux G (2000) Polarization of the anterior-posterior axis of C. elegans is a microtubule-directed process. Nature 408(6808):89–92
Wang G, Reinke V (2008) A C. elegans Piwi, PRG-1, regulates 21U-RNAs during spermatogenesis. Curr Biol 18(12):861–867
Wang X, Zhao Y, Wong K, Ehlers P, Kohara Y, Jones SJ, Marra MA, Holt RA, Moerman DG, Hansen D (2009) Identification of genes expressed in the hermaphrodite germ line of C. elegans using SAGE. BMC Genomics 10:213
Ward S (1986) The asymmetric localization of gene products during the development of Caenorhabditis elegans spermatozoa. Gametogenesis and the early embryo. A.R. Liss, New York, pp. 55–75
Ward S, Carrel JS (1979) Fertilization and sperm competition in the nematode Caenorhabditis elegans. Dev Biol 73(2):304–321
Ward S, Argon Y, Nelson GA (1981) Sperm morphogenesis in wild-type and fertilization-defective mutants of Caenorhabditis elegans. J Cell Biol 91(1):26–44
Ward S, Hogan E, Nelson GA (1983) The initiation of spermiogenesis in the nematode Caenorhabditis elegans. Dev Biol 98(1):70–79
Washington NL, Ward S (2006) FER-1 regulates Ca2 + − mediated membrane fusion during C. elegans spermatogenesis. J Cell Sci 119 (Pt 12):2552–2562
Wignall SM, Villeneuve AM (2009) Lateral microtubule bundles promote chromosome alignment during acentrosomal oocyte meiosis. Nat Cell Biol 11(7):839–844
Wolf N, Hirsh D, McIntosh JR (1978) Spermatogenesis in males of the free-living nematode, Caenorhabditis elegans. J Ultrastruct Res 63(2):155–169
Wu TF, Chu DS (2008) Epigenetic processes implemented during spermatogenesis distinguish the paternal pronucleus in the embryo. Reprod Biomed Online 16(1):13–22
Wu JC, Go AC, Samson M, Cintra T, Mirsoian S, Wu TF, Jow MM, Routman EJ, Chu DS (2012) Sperm development and motility are regulated by PP1 phosphatases in Caenorhabditis elegans. Genetics 190(1):143–157
Yi K, Buttery SM, Stewart M, Roberts TM (2007) A Ser/Thr kinase required for membrane-associated assembly of the major sperm protein motility apparatus in the amoeboid sperm of Ascaris. Mol Biol Cell 18(5):1816–1825
Yushin VV, Commans A (2005) ltrastructure of sperm development in the free-living marine nematode Metachromadora itoi (Chromadoria, Desmodorida). Acta Zoologica 86(4):255–265
Zanetti S, Puoti A (2012) Sex determination in the C. elegans germline. Advances in Experimental Medicine and Biology 757:41–69. (Chap. 3, this volume) Springer, New York
Zeng Y, Cullen BR (2003) Sequence requirements for micro RNA processing and function in human cells. RNA 9(1):112–123
Zhu GD, L’Hernault SW (2003) The Caenorhabditis elegans spe-39 gene is required for intracellular membrane reorganization during spermatogenesis. Genetics 165(1):145–157
Zhu GD, Salazar G, Zlatic SA, Fiza B, Doucette MM, Heilman CJ, Levey AI, Faundez V, L’Hernault SW (2009) SPE-39 family proteins interact with the HOPS complex and function in lysosomal delivery. Mol Biol Cell 20(4):1223–1240
Acknowledgments
We thank David Greenstein and Harold Smith for sharing data prior to publication. We thank Meghann Shorrock for assistance with figures. We also thank Dana Byrd, Margaret Jow, and Kari Price for critical reading of this manuscript. This work was supported grants from the National Science Foundation to D.S.C. (MCB-0747515) and the National Institutes of Health to D.S.C. (R15 HD068996) and D.C.S. (R15 GM096309).
Author information
Authors and Affiliations
Corresponding authors
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media New York
About this chapter
Cite this chapter
Chu, D.S., Shakes, D.C. (2013). Spermatogenesis. In: Schedl, T. (eds) Germ Cell Development in C. elegans. Advances in Experimental Medicine and Biology, vol 757. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-4015-4_7
Download citation
DOI: https://doi.org/10.1007/978-1-4614-4015-4_7
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4614-4014-7
Online ISBN: 978-1-4614-4015-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)