
Abstract
The monocyte-derived, inflammatory dendritic cell subset plays an important role during immune responses against infections. This review will focus on the complex, changing role of this subset during mycobacterial infection. Studies demonstrate that in addition to sustaining a systemic anti-mycobacterial response, the inflammatory dendritic cell subset present in Mycobacterium-induced granulomas also influences local immune regulation within the granuloma over the course of infection. This review will also survey the literature on how similar and different inflammatory dendritic cell subsets during other infections.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
The discovery of dendritic cells (DCs) nearly four decades ago has provided immunologist with the missing link to efficiently bridge the innate and adaptive immune response (Steinman et al. 1973, 1974a, b). DCs are comprised of heterogeneous populations, with each subset uniquely qualified to support either immunity during infection and tolerance in the face of autoimmunity. Under steady-state conditions, conventional DCs make up a very small percentage of tissue and lymphoid-resident cells. In the face of infection the immune system must respond quickly. Circulating, peripheral blood monocytes provide a powerful reservoir of immune artillery, as they can efficiently give rise to DCs under infectious conditions. This monocyte-derived DC (moDCs) lineage that arises during infection shares many functional and phenotypic similarities to classical DCs, and are therefore termed ‘inflammatory DCs’ (inflamDCs). There has been a recent surge in data that irrefutably demonstrates the importance of this subset during infection by a wide array of pathogens (viruses, bacteria, fungi, protozoan, etc.). This review will discuss the origin and phenotype of moDCs, and their involvement during infection by various pathogens. The newly immerging role inflamDCs play during Mycobacterium infection will be specifically highlighted in this review.
2 Development and Phenotype
DC precursors, both classical and monocyte-derived inflammatory, originate in the bone marrow from hematopoietic stem cells. Both subsets share the same myeloid precursor that gives rise to monocyte/macrophage and DC lineages, known as the macrophage-DC precursor (MDP) (Fogg et al. 2006). However, the classical DC and monocyte developmental pathway diverges henceforth (Liu et al. 2009). Although in vitro culture systems can generate a large number of DCs by culturing monocytes with cytokine granulocyte-macrophage colony-stimulating factor (GM-CSF) (Inaba et al. 1992; Sallusto and Lanzavecchia 1994), and early in vivo data suggested the generation of a DC-like population from both murine and human monocytes (Randolph et al. 1998, 1999), recent in vivo data strongly supports the idea that monocytes do not give rise to classical lymphoid organ DCs during steady-state conditions. in vivo experiments that utilize either adoptive cell transfer models or GFP reporter mice driven by the monocyte-specific lysozyme M promoter, all demonstrate that monocytes do not have classical DC progenitor activity or are able to reconstitute classical lymphoid organ DCs during steady-state conditions (Fogg et al. 2006; Naik et al. 2006; Varol et al. 2007; Jakubzick et al. 2008; Liu et al. 2009). However, monocytes have been shown to replenish some DC populations in peripheral organs, such as the intestines, lung, and skin (Holt et al. 1994; Ginhoux et al. 2006; Varol et al. 2007). MDP exposure to fms-like tyrosine kinase 3 ligand (Flt3L) (McKenna et al. 2000; Waskow et al. 2008) or GM-CSF (Dai et al. 2002) in the bone marrow results in the divergence of conventional DC and monocyte developmental pathways, respectively. Conventional DC (cDC) development is reviewed in depth elsewhere (Liu and Nussenzweig 2010). In addition to early exposure of the growth factor GM-CSF, the monocyte lineage is further selected for at the transcriptional level. Increased expression of transcription factor PU.1 has been shown to favor monocyte lineage development (Dahl et al. 2003). Interestingly, continued expression of PU.1 further promotes monocyte differentiation into DCs by suppressing the macrophage-inducing transcription factor MafB, and vice versa (Bakri et al. 2005). Conversely, the transcription factor STAT3 sustains cDC Flt3-dependent differentiation (Laouar et al. 2003). Table 1 summarizes this and other defining characteristics between cDCs and monocyte-derived inflamDCs.
From the bone marrow, monocytes can be classified into two subpopulations based on high- or low-surface expression of Ly6C. Ly6Chigh and Ly6Clow populations were originally and often times currently used interchangeably with Gr-1high and Gr-1low, respectively. However, in addition to recognizing Ly6C, the monoclonal antibody Gr-1 (RB6-8C5) also recognizes Ly6G, a marker not expressed on monocytes (Fleming et al. 1993). Therefore, Ly6C expression is the more accurate description of murine monocyte subsets. The Ly6Chigh and Ly6Clow monocyte populations in mice are believed to correspond to the human CD14highCD16− and CD14intCD16+ populations, respectively (Randolph et al. 2008). The two murine Ly6Chigh and Ly6Clow monocyte populations can be further phenotyped by expression of different chemokine and adhesion molecules.
The conventional Ly6Chigh monocytes are also CCR2pos, CD62Lpos, CCR5neg, CD11cneg and CX3CR1low, and the Ly6Clow population is CX3CR1high, CCR2neg, CD62Lneg, CX3CR1pos, and CD11cint (Geissmann et al. 2003; Sunderkotter et al. 2004; Tacke et al. 2007). These two populations are not independently derived as the Ly6Chigh population has been shown to give rise to the Ly6Clow population, both in bone marrow and in the periphery (Sunderkotter et al. 2004; Tacke and Randolph 2006; Arnold et al. 2007; Nahrendorf et al. 2007; Varol et al. 2007). Monocyte emigration from the bone marrow is controlled by chemokine receptors CCR2 on the Ly6Chigh population, and CX3CR1 and CCR5 on the Ly6Clow counterpart, but the mechanisms behind this control are not known (Combadiere et al. 2008). Once within the blood, the Ly6ClowCX3CR1high population has been elegantly imaged crawling along vessels under steady-state conditions regardless of the direction of flow, but rapidly extravasated into tissue in the presence of infection (Auffray et al. 2007). A recent study from the Steinman group definitively demonstrated the differentiation of monocytes into DCs in vivo in response to microbial stimuli (Cheong et al. 2010). This study demonstrated the inflammation -induced recruitment of moDCs into the T cell areas of the affected lymph node and subsequent expression of DC-SIGN (CD209a). The moDC mobilization they observed was dependent on CCR7, TLR4, CD14 , and Trif expression on the DC. In spite of the many characteristics moDCs share with cDCs, antigen presentation is not one of them. A recent, definitive report by Kamphorst et al. exhaustively compared the antigen presentation capacity of cDCs and moDCs following several mechanisms of antigen capture (Kamphorst et al. 2010). This study demonstrated that while moDCs are efficient at priming CD8+ T cells through cross presentation, the cDC compartment was much more effective at presenting antigen to MHCII-restricted CD4+ T cells . While the inflamDCs may appear very similar to cDCs in terms of costimulatory molecules, MHCII expression and morphology, they may differ internally upon acquisition of antigen. A recent study by McCurley and Mellman demonstrated that the moDC subset in humans had a higher protease content, and as a result, had higher lysosomal proteolysis compared to cDCs (McCurley and Mellman 2010). This high lysosomal proteolytic activity results in quick degradation of intracellular antigen and may explain the poor ability of inflamDCs to present MHCII peptide.
In the face of infection the monocyte lineage provides a quick, profuse source of cells able to differentiate into the necessary populations of antigen presenting cells (APCs) necessary for quick combat. In this next section we will discuss their potential to differentiate into inflamDCs and the role they play during infection by bacteria, viruses, fungi and protozoa.
3 Function of Monocyte-derived Inflammatory DCs During Infection
3.1 Mycobacteria
All things considered (i.e., prevalence, new cases and deaths per year, economic burden, drug resistance, etc.) infection with Mycobacterium remains one of the most devastating diseases worldwide. The granuloma is the hallmark of mycobacterial infections, and is the interface for the host immune response and bacterial persistence. Functional granulomas protect an estimated two billion individuals from active disease by attenuating bacterial growth and dissemination (Ulrichs and Kaufmann 2006). One of the defining characteristics of Mycobacterium infection is that even in the face of an early, strong immune response, the host often fails to completely eradicate the bacteria, resulting in a long-term chronic infection. Therefore, when bearing in mind Mycobacterium infection it is best to view both the immunological and bacteriological events in two separate stages, acute and chronic. For example, granulomas formed during the acute stage of infection largely protect the host by limiting bacterial dissemination and inadvertent tissue damage. However, during chronic infection these well-formed granulomas provide the mycobacteria with a long-term survival niche (Volkman et al. 2004; Russell 2007). Understanding the role of DCs during Mycobacterium infection is still in its infancy, but the growing body of data supports the idea that they have an essential, influential role during both the acute and chronic stages. The predominating DC subset involved during both stages is monocyte-derived inflamDCs, largely characterized by Ly6CintCD11bhighCD11clow expression (Reljic et al. 2005; Humphreys et al. 2006; Wolf et al. 2007; Schreiber et al. 2010). However, the mechanisms governing monocyte recruitment, differentiation, infection, lymphatic antigen transport, and T cell priming are not equally understood for both the acute and chronic stages.There is more information regarding the role of inflamDCs during the acute phase of infection. However, recent studies are now focusing on the chronic phase. Data from these studies suggests a changing role of monocyte-derived inflamDCs during chronic Mycobacterium infection.
Recruitment of this subset to the site of acute Mycobacterium infection seems to occur in both a CCR2- and CCR5-dependent manner and not CX3CR1-dependent, as demonstrated by infection of mice with mutated expression of these chemokine receptors (Hall et al. 2009). Following Mycobacterium tuberculosis (MTb) infection, CCR2 deficient mice were highly susceptible to high doses and had a delayed immune response when challenged with low infectious doses (Peters et al. 2001; Scott and Flynn 2002). Conversely, mice over expressing CCR2 better controlled Mycobacterium bovis bacillus Calmette-Guérin (BCG ) infection by faster granuloma formation, lower bacterial burden and increased transport of bacterial antigen to the draining lymph nodes by DCs (Schreiber et al. 2008). Interestingly, humans with a single nucleotide polymorphism in the MCP-1, which resulted in increased peripheral levels of the chemokine, had increased susceptibility of MTb infection, suggesting a possible desensitization to the CCR2 ligand over time (Flores-Villanueva et al. 2005; Serbina et al. 2008). In the absence of a CCR2-expressing monocyte subset, CCR5 expression may compensate and vice versa as CCR5-deficient mice also showed no significant difference in Mtb control (Badewa et al. 2005). Studies examining the immune response in the absence of both chemokine receptors have not been done. The global requirement of CD11cpos DCs for the initiation of T cell immunity and bacterial control following MTb infection was definitively demonstrated by transiently depleting DCs using pCD11c-diphtheria toxin receptor transgenic mice (Tian et al. 2005). Depleting the CD11c population prior to infection resulted in a delayed CD4+ T cell response to MTb and impaired control of MTb replication. In back to back reports, Wolf and colleagues demonstrated that the CD11bposCD11cpos DC population recruited to the lung after MTb infection is readily infected and facilitates the transport of MTb antigen to the draining lymph node (Wolf et al. 2007). Eliminating a primary chemokine network utilized by this subset en route to lymph nodes using plt mice, which lack CCR7 ligands CCL19 and CCL21ser, they demonstrated the necessity of mycobacteria transport by CD11bposCD11cpos DCs (Wolf et al. 2007) and Mycobacterium-specific IFNγ-producing CD4+ T cells (Khader et al. 2009). Early studies that demonstrated a potent T cell stimulatory activity by lymph node DCs after BCG infection did not entirely separate lymph node-resident conventional DCs from recruited monocyte-derived inflamDCs (Jiao et al. 2002). However, recent studies specifically addressing the activation capacity of CD4+ T cells specific for Mycobacterium antigen 85B (P25) during early Mtb infection found inflamDCs to be poor T cell stimulators and elicit much less IFNγ production compared to other DC subsets (Wolf et al. 2007). As previously described for Listeria monocytogenes and Salmonella, evidence for the transfer of antigen from inflamDCs to lymph node-resident DCs also seems to be a phenomenon occurring during acute Mycobacterium infection as well. Eliminating the mechanism of inflamDC transport to the lymph nodes by using plt mice, resulted in 95% fewer bacilli reaching the lymph nodes, which effectively eliminated the source of antigen (Wolf et al. 2007). In addition to CCR7-expressing migratory DCs aiding in antigen delivery to the lymph node, it was also found that CCR7-deficient mice infected via aerosol with Mtb had decreased bacterial load in the spleen during chronic infection, demonstrating the requirement for CCR7-expressing migratory DCs for bacterial dissemination during acute Mtb infection (Kahnert et al. 2007). There are numerous studies testing the ability of DCs ex vivo, and bone marrow and moDCs in vitro, to take up mycobacteria, mature, produce cytokines and prime T cells. This is reviewed in greater detail elsewhere (Schreiber et al. 2010). Collectively, these studies yield conflicting results by demonstrating the ability of DCs to both stimulate and suppress immunity ex vivo and in vitro. Which further reaffirms the idea that DCs, particularly inflamDCs, play a multifaceted role during mycobacterial infection.
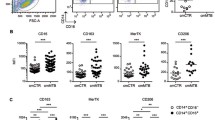
Recent data from our group has supported this multifunctional role for inflamDCs during Mycobacterium infection, particularly within the granuloma. Using in vivo imaging, a recent study by Egen and colleagues demonstrated the intense movement of T cells within the acute granuloma (Egen et al. 2008). Their data showed that newly arrived T cells efficiently scan granuloma-resident APCs. This scanning is necessary, as newly primed T cells require a second antigen encounter at the site of inflammation in order to rapidly secrete cytokines (Mohrs et al. 2005). By phenotyping monocyte-derived inflamDCs in both acute and chronic BCG -induced granulomas, we found that their phenotype varied greatly from the progression of acute to chronic infection. InflamDCs found in acute granulomas had high expression of MHCII, CD40, CD80 and CD86, and were able to support IFNγ production from newly arrived primed Mycobacterium-specific P25 CD4+ T cells (Schreiber et al. 2010). However, as infection progressed the inflamDCs within the chronic granuloma had decreased expression of MHCII and T cell costimulatory molecules, and had a significant increase in T cell inhibitory molecules PD-L1 and PD-L2. These DCs could no longer support IFNγ production from newly arrived Mycobacterium-specific P25 CD4+ T cells . This inability was in part dependent on PD-L1/2:PD-1 signaling, as blockade of this pathway partially restored IFNγ production. Monocyte-derived inflamDCs, by affecting local IFNγ availability, may contribute to granuloma maturation and bacterial persistence (Fig. 1).

Changing role of inflamDCs in Mycobacterium-induced granulomas over the course of infection
Enabling bacterial persistence during mycobacterial infection has partially led to the belief that chronic granulomas are ‘walled-off’ structures, inaccessible to immune surveillance. One defining feature of moDCs is their profound ability to migrate. Therefore, we asked the question of whether chronic granulomas were assessable to inflamDCs, and if so, were they able to then leave the granuloma with antigenic cargo en route to the draining lymph nodes? Using a kidney capsule transplant model, which consists of grafting granuloma-containing tissue underneath the kidney capsule of uninfected recipients, we were able to study CD11cpos cellular migration into and out of both acute and chronic Mycobacterium-induced granulomas using CD11c enhanced yellow fluorescent protein (CD11c-EYFP) mice with ubiquitously fluorescing DCs (Schreiber et al. 2011. Manuscript Under Review)(Lindquist et al. 2004). This study demonstrated that inflamDCs had considerable access to enter and exit chronic granulomas, even more so than acute lesions, with a turnover rate of ~60% and 30%, respectively, by one week. Determining the life span of inflamDCs within the granuloma is complicated. Circulating monocytes have a relatively short half-life, as do migrating DCs; however, monocytes have also been shown to live for months after differentiation (Whitelaw 1966; Kamath et al. 2002; Liu et al. 2007; Gonzalez-Mejia and Doseff 2009). By staining acute and chronic granulomatous lesions with a marker for apoptosis and CD11c, the DCs present do not appear to be dying locally within the lesions during either acute or chronic infection (Fig. 2). It is difficult to discern whether the change in DC flux throughout the course of infection is due to a general shrinking of the lesions or potential differences in DC expansion locally due to M-CSF availability. InflamDCs leaving granulomas migrated not only to the draining lymph nodes, but also to other systemic lymphoid organs. This systemic migration supported priming of P25 CD4+ T cells during both acute and chronic infection , albeit to a lesser extent in the latter. Grafting both acute and chronic granulomas into MHCII−/− recipients resulted in total abrogation of P25 CD4+ T cell activation, demonstrating the necessity for recipient MHCIIpos cells. Eliminating the mechanism used by inflamDCs to migrate from the granuloma to the draining renal lymph node by transplanting granulomas from CCR7−/− mice, abrogated P25 CD4+ T cells activation during acute infection. Collectively demonstrating the role of inflamDCs during T cell priming as antigenic ferries (Randolph 2006).

Tunel staining on sections from 3- and 10-week BCG -infected mice. White dashed line indicates periphery of granuloma lesions. Staining with CD11c (green) and apoptosis indicator, Tunel (red), demonstrates that DCs are not undergoing apoptosis within the granuloma throughout chronic infection
It is evident that the changing phenotype and function of inflamDCs in mycobacterial granulomas plays an important role, for better or worse, in the disease course. Better understanding this cellular compartment during both acute and chronic Mycobacterium infection will inevitably lead to a better understanding of the disease and potential targets for therapeutic intervention.
3.2 Other Bacteria
Pioneering work on inflamDCs largely came from studies done by the groups of Pamer and Leenen on Listeria monocytogenes (Auffray et al. 2009). A Gram-positive intracellular bacterium, L. monocytogenes got its species name ‘monocytogenes’ after the observation of increased monocytes in infected rabbits (Murray et al. 1926; Serbina et al. 2008). The early requirement for monocyte-derived Ly6C-expressing cells was further confirmed by depleting mice of this population using the RB6-8C5 monoclonal antibody within 24 h of L. monocytogenes infection, resulting in death within six days (Czuprynski et al. 1994). Upon infection with L. monocytogenes, Ly6Chigh cells egress from the bone marrow in considerable numbers by CCR2-mediated signals largely in response to monocyte chemoattractant protein-1 (MCP-1) (Kurihara et al. 1997; Sunderkotter et al. 2004; Serbina et al. 2006; Tsou et al. 2007). Induction of MCP-1 expression during L. monocytogenes infection is dependent on cytosolic recognition of the bacterium and independent of the MyD88 signaling pathway (Serbina et al. 2003). MCP-1 is also significantly induced after infection by Toxoplasma gondii, Aspergillus fumigatus and Mycobacterium tuberculosis (Lin et al. 1998; Blease et al. 2001; Robben et al. 2005). Mice lacking CCR2 succumb to fatal infection by L. monocytogenes and lack the CD11bintCD11cint population that is morphologically similar to the splenic CD11chigh population (i.e., pleomorphic nuclei, dendrites and numerous vacuoles) (Kurihara et al. 1997; Serbina et al. 2006). This CD11bintCD11cint population present during L. monocytogenes infection also produced TNF- and iNOS (Tip), thereafter referred to as TipDCs (Serbina et al. 2003). Kang et al. recently demonstrated that the differentiation of TipDCs from the recruited Ly6Chigh monocytes was dependent on the ability of splenic CD11cpos cells to induce IFNγ production from NK cells (Kang et al. 2008). In addition to CCR2, Auffrey and colleagues demonstrated a role for chemokine receptor CX3CR1 on the recruitment of Ly6Cpos(GR1pos)CD115pos moDC precursors to the spleen during L. monocytogenes infection (Auffray et al. 2009). A follow up study by this group used Cx 3 cr1 gfp/ + mice to elegantly image in vivo monocyte recruitment during L. monocytogenes infection (Auffray et al. 2007). Here, the GR1negCX3CR1pos monocytes observed patrolling the vasculature rapidly extravasated into inflamed tissue and initiated a macrophage-like transcriptional program by up regulating cMaf and MafB. Conversely, the GR1negCX3CR1pos subset took on a DC transcriptional profile by up regulation of Pu.1 and RelB genes within 2 hours post infection. Once recruited to the site of infection both differentiation of monocytes into TipDCs and production of IL-12 wash shown to be MyD88 dependent (Serbina et al. 2003; Zhan et al. 2010). CX3CR1 is also required by moDCs recruited to the intestinal lamina propria during Salmonella typhimurium infection in order to extend their dendrites into the intestinal lumen and transport antigen to the lymph node (Niess et al. 2005). MoDCs are among the better migratory DC populations; however, their ability to directly prime the immune system has been called into question in infectious models across the board (Randolph et al. 2008). Using a Salmonella typhimurium bacterial infection model, Ravindran and colleagues demonstrated that although a recruited Ly6Chigh DC-like population was required for induction of an anti-Salmonella CD4+ T cell response, the Ly6Chigh phagocytes themselves were not directly responsible for this induction (Ravindran et al. 2007). Rather, the Ly6Chigh population was necessary for transporting the antigen from the site of infection to the draining lymph nodes. Rydstrom and colleagues also demonstrated the inability of a monocyte-derived TipDC-like population to activate antigen-specific T cells following Salmonella infection (Rydstrom and Wick 2007).
3.3 Viruses
In addition to bacteria, studies using viral infection models have significantly broadened our understanding of inflamDCs. Under steady state conditions, few monocytes are recruited to the vaginal epithelium. Although, following intravaginal infection with herpes simplex virus-2 (HSV-2), recruited GR-1pos monocytes readily gave rise to vaginal epithelial DCs, including CD11cposMHCIIpos subsets (Iijima et al. 2007; Eidsmo et al. 2009). Similar to the granuloma kidney capsule transplant model previously discussed, Wakim and colleagues grafted HSV-infected dorsal root ganglia under the kidney capsule of recipient naïve mice, which resulted in viral reactivation (Wakim et al. 2008). They sophisticatedly demonstrated that the recruited monocyte-derived Ly6Chigh DCs were able to restimulate memory viral-specific T cells within the graft. In addition to HSV, monocyte-derived inflamDCs are also readily involved in influenza infection. Upon infection with the virulent influenza A virus CD11cposCD11bhighLy6Cpos inflamDCs are recruited in a CCR2-dependent manner directly into the lymph nodes, as their recruitment was not hindered in neither plt nor CCR7−/− mice (Nakano et al. 2009). When purified and stimulated ex vivo, these LN recruited inflamDCs produced IL-12(p70) and had the ability to stimulate OTII CD4+ T cells . A study by Aldridge et al. also found CCR2-dependent recruitment of Ly6cposCD11bpos TipDCs into the lungs following influenza A infection (Aldridge et al. 2009). Interestingly, they found that these TipDCs presented antigen to cytotoxic CD8+ T cells in the lung and limiting the number of DCs recruited using pioglitazone, a synthetic pharmacological agent that suppressed CCL2 secretion, they could dampen inflammation -induced pathology. That indicated that too many TipDCs are not additive in protection and could even be lethal if not controlled. Viral models have also supported evidence for the dual requirement of inflamDCs migrating from the site of infection and lymph node-resident DCs in the initiation of T cell immunity. Belz and colleagues demonstrated that during both HSV and influenza infection, efficient antigen-specific T cell priming required the collaboration of both lymph node-residing DCs and DCs originating from the site of infection (Belz et al. 2004). Using an HSV skin infection model, Allan and colleagues further demonstrated that skin-originating DCs were required to shuttle antigen to the draining lymph node, where lymph node-resident CD8α pos DCs were principally responsible for initiating the T cell response (Allan et al. 2006).
3.4 Fungi
The role of inflamDCs during fungal infection remains one of the less understood areas. Aspergillus fumigatus was previously shown to recruit monocytes in a CCR2-dependent mechanism in mice (Blease et al. 2000). A recent study from the Pamer group reconfirmed Ly6C-expressing monocyte recruitment using CCR2 and demonstrated that this cellular compartment was responsible for transporting Aspergillus antigen from the lung to the draining lymph node for CD4+ T cell priming (Hohl et al. 2009).A more recent study demonstrated that A. fumigatus infection of neutropenic hosts resulted in an even more robust recruitment of inflamDCs, suggestive of a novel defense mechanism to circumvent loss of neutrophils (Park et al. 2010). Antigenic hand-off was also demonstrated in a recent study by Erlsand and colleagues, whom investigated the immune response to an attenuated vaccine strain of Blastomyces dermatitidis (Ersland et al. 2010). They found that monocyte-derived inflamDCs were quickly recruited to the subcutaneous immunization site and along with skin-migratory DCs, shuttled antigen to the lymph nodes; however, lymph node-resident DCs were responsible for priming CD4+ T cells .
3.5 Protozoan
More so than other infectious models, the recruitment and differentiation of inflamDCs from recruited monocytes was definitively demonstrated by Leon and Ardavin using the protozoan parasite Leishmania major (Leon et al. 2007). Mice infected with L. major received an adoptive transfer of purified monocytes, traceable by the Ly5.1/5.2 congenic markers. After 72 hours post transfer, two DC lineages arose from the transferred monocytes: CD11clowLy-6ChighMHCIIint and CD11cintLy-6Cint-highMHC IIint-high populations. The moDCs were also the predominate cellular compartment infected and the only cells presenting L. major peptide LACK in the context of MHCII, suggesting that they contribute to anti-parasitic CD4+ T cell response. In a follow up study, this same group demonstrated that during L. major infection monocytes were recruited through inflamed dermal venules by binding of PSGL-1 with P/E-selectin and L-selectin with PNAd, and migration through high endothelial venules relied only the latter interaction (Leon and Ardavin 2008). A recent study also confirmed that the same monocyte-derived inflamDC ingesting the parasite L. major in mice was the same counter subset in humans (Zhan et al. 2010). Monocyte-derived inflamDCs have also been studied in the protozoan pathogen, Toxoplasma gondii. During infection, GR-1posCCR2posCX3CR1low monocytes are recruited to inflamed tissue in a CCR2-dependent manner, demonstrated by lethality observed in CCR2 or MCP-1 deficient mice (Robben et al. 2005). Adoptive transfer of inflammatory monocytes into CCR2−/− T. gondii infected mice restored protection (Dunay et al. 2008).
4 Conclusions
The dynamic, crucial role inflamDCs play during infection is only beginning to be fully understood and appreciated. From our discussion here, inflamDCs represent a common thread that can be drawn between the various infectious models. This monocyte-derived lineage is rapidly recruited to the site of infection, be it liver, lung, spleen or skin, within hours. The TNF and iNOS-producing capacity of these cells in the face of various pathogens represents a common line of innate defense. One of the most significant commonalities of inflamDCs shared between the various pathogens is their role in shuttling antigen from the site of infection to the respective draining lymph nodes and transferring this precious cargo to lymph node-resident DCs, which relies on their professional migratory abilities. The mechanism (s) behind this antigenic transfer are currently not known. Based on the current literature, a recent review by Randolph et al. speculates that this transfer may be occurring by two potential mechanisms (Randolph et al. 2008). The first scenario involves the moDCs dying upon arrival to the lymph node and lymph node-resident DCs reprocessing the dead DC and its antigenic cargo. The second model suggests that moDCs transfer intact portions of their plasma membrane that contain MHC/peptide complexes to lymph node-resident DCs. Better understanding the mechanism behind antigenic exchange will increase our understanding of the initiation of adaptive immunity during infection and may potentially provide a target for immune intervention.
Our discussion on the role of inflamDCs during Mycobacterium infection demonstrates the plasticity this monocyte-derived linage can have during a single infection over time. Figure 1 illustrates this changing role. During acute infection inflamDCs facilitate an anti-Mycobacterium T cell response by reboosting newly recruited CD4+ T cells within the granuloma to produce IFNγ, an essential cytokine to promote macrophage killing of intracellular bacteria. As infection progresses, traffic of inflamDCs into and out of chronic granulomas supports Mycobacterium-specific T cell priming in the lymph nodes by providing a continuous source of granuloma-residing antigen. However, once these newly primed T cells reach the granuloma, the phenotype of the local inflamDCs does not support IFNγ production, thereby contributing to bacterial persistence.
In closing, this review demonstrates that the monocyte-derived inflamDCs are often the dominant DC subset present at sites of inflammation induced by a variety of infectious agents. DCs are a necessary element of both the innate and adaptive immune response; therefore, making it important to increase our understanding of their function . At the site of infection inflamDCs have been shown to both induce and suppress the immune response, and can also critically effect the host-pathogen interaction. Many questions still remain regarding this DC subset. Some of them include: How does the inflamDC regulate components of the inflammatory response? What is their expected lifespan within an inflammatory lesion? What is the extent of their traffic to and from the inflammatory lesions? And are they able to sustain systemic immunity? In regards to Mycobacterium-host interactions and the anatomical structure of the granuloma, which is required for mycobacterial containment, the inflamDC plays a critical role in the pathogenesis. They facilitate dissemination of bacteria and bacterial antigens, which drives long-term immunity, and regulate intragranuloma processes; thereby representing an important part of the puzzle of long-term bacterial containment
References
Aldridge, J. R., Jr., C. E. Moseley, D. A. Boltz, N. J. Negovetich, C. Reynolds, J. Franks, S. A. Brown, P. C. Doherty, R. G. Webster and P. G. Thomas. (2009) TNF/iNOS-producing dendritic cells are the necessary evil of lethal influenza virus infection. Proc Natl Acad Sci U S A. 106, 5306-5311
Allan, R. S., J. Waithman, S. Bedoui, C. M. Jones, J. A. Villadangos, Y. Zhan, A. M. Lew, K. Shortman, W. R. Heath and F. R. Carbone. (2006) Migratory dendritic cells transfer antigen to a lymph node-resident dendritic cell population for efficient CTL priming. Immunity. 25, 153-162
Arnold, L., A. Henry, F. Poron, Y. Baba-Amer, N. van Rooijen, A. Plonquet, R. K. Gherardi and B. Chazaud. (2007) Inflammatory monocytes recruited after skeletal muscle injury switch into antiinflammatory macrophages to support myogenesis. J Exp Med. 204, 1057-1069
Auffray, C., D. Fogg, M. Garfa, G. Elain, O. Join-Lambert, S. Kayal, S. Sarnacki, A. Cumano, G. Lauvau and F. Geissmann. (2007) Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science. 317, 666-670
Auffray, C., D. K. Fogg, E. Narni-Mancinelli, B. Senechal, C. Trouillet, N. Saederup, J. Leemput, K. Bigot, L. Campisi, M. Abitbol, T. Molina, I. Charo, D. A. Hume, A. Cumano, G. Lauvau and F. Geissmann. (2009) CX3CR1+ CD115+ CD135+ common macrophage/DC precursors and the role of CX3CR1 in their response to inflammation. J Exp Med. 206, 595-606
Auffray, C., M. H. Sieweke and F. Geissmann. (2009) Blood monocytes: development, heterogeneity, and relationship with dendritic cells. Annu Rev Immunol. 27, 669-692
Badewa, A. P., L. J. Quinton, J. E. Shellito and C. M. Mason. (2005) Chemokine receptor 5 and its ligands in the immune response to murine tuberculosis. Tuberculosis (Edinb). 85, 185-195
Bakri, Y., S. Sarrazin, U. P. Mayer, S. Tillmanns, C. Nerlov, A. Boned and M. H. Sieweke. (2005) Balance of MafB and PU.1 specifies alternative macrophage or dendritic cell fate. Blood. 105, 2707-2716
Belz, G. T., C. M. Smith, L. Kleinert, P. Reading, A. Brooks, K. Shortman, F. R. Carbone and W. R. Heath. (2004) Distinct migrating and nonmigrating dendritic cell populations are involved in MHC class I-restricted antigen presentation after lung infection with virus. Proc Natl Acad Sci U S A. 101, 8670-8675
Blease, K., B. Mehrad, N. W. Lukacs, S. L. Kunkel, T. J. Standiford and C. M. Hogaboam. (2001) Antifungal and airway remodeling roles for murine monocyte chemoattractant protein-1/CCL2 during pulmonary exposure to Asperigillus fumigatus conidia. J Immunol. 166, 1832-1842
Blease, K., B. Mehrad, T. J. Standiford, N. W. Lukacs, J. Gosling, L. Boring, I. F. Charo, S. L. Kunkel and C. M. Hogaboam. (2000) Enhanced pulmonary allergic responses to Aspergillus in CCR2−/− mice. J Immunol. 165, 2603-2611
Cheong, C., I. Matos, J. H. Choi, D. B. Dandamudi, E. Shrestha, M. P. Longhi, K. L. Jeffrey, R. M. Anthony, C. Kluger, G. Nchinda, H. Koh, A. Rodriguez, J. Idoyaga, M. Pack, K. Velinzon, C. G. Park and R. M. Steinman. (2010) Microbial stimulation fully differentiates monocytes to DC-SIGN/CD209(+) dendritic cells for immune T cell areas. Cell. 143, 416-429
Combadiere, C., S. Potteaux, M. Rodero, T. Simon, A. Pezard, B. Esposito, R. Merval, A. Proudfoot, A. Tedgui and Z. Mallat. (2008) Combined inhibition of CCL2, CX3CR1, and CCR5 abrogates Ly6C(hi) and Ly6C(lo) monocytosis and almost abolishes atherosclerosis in hypercholesterolemic mice. Circulation. 117, 1649-1657
Czuprynski, C. J., J. F. Brown, N. Maroushek, R. D. Wagner and H. Steinberg. (1994) Administration of anti-granulocyte mAb RB6-8C5 impairs the resistance of mice to Listeria monocytogenes infection. J Immunol. 152, 1836-1846
Dahl, R., J. C. Walsh, D. Lancki, P. Laslo, S. R. Iyer, H. Singh and M. C. Simon. (2003) Regulation of macrophage and neutrophil cell fates by the PU. 1:C/EBPalpha ratio and granulocyte colony-stimulating factor. Nat Immunol. 4, 1029-1036
Dai, X. M., G. R. Ryan, A. J. Hapel, M. G. Dominguez, R. G. Russell, S. Kapp, V. Sylvestre and E. R. Stanley. (2002) Targeted disruption of the mouse colony-stimulating factor 1 receptor gene results in osteopetrosis, mononuclear phagocyte deficiency, increased primitive progenitor cell frequencies, and reproductive defects. Blood. 99, 111-120
Dunay, I. R., R. A. Damatta, B. Fux, R. Presti, S. Greco, M. Colonna and L. D. Sibley. (2008) Gr1(+) inflammatory monocytes are required for mucosal resistance to the pathogen Toxoplasma gondii. Immunity. 29, 306-317
Egen, J. G., A. G. Rothfuchs, C. G. Feng, N. Winter, A. Sher and R. N. Germain. (2008) Macrophage and T cell dynamics during the development and disintegration of mycobacterial granulomas. Immunity. 28, 271-284
Eidsmo, L., R. Allan, I. Caminschi, N. van Rooijen, W. R. Heath and F. R. Carbone. (2009) Differential migration of epidermal and dermal dendritic cells during skin infection. J Immunol. 182, 3165-3172
Ersland, K., M. Wuthrich and B. S. Klein. (2010) Dynamic interplay among monocyte-derived, dermal, and resident lymph node dendritic cells during the generation of vaccine immunity to fungi. Cell Host Microbe. 7, 474-487
Fleming, T. J., M. L. Fleming and T. R. Malek. (1993) Selective expression of Ly-6G on myeloid lineage cells in mouse bone marrow. RB6-8C5 mAb to granulocyte-differentiation antigen (Gr-1) detects members of the Ly-6 family. J Immunol. 151, 2399-2408
Flores-Villanueva, P. O., J. A. Ruiz-Morales, C. H. Song, L. M. Flores, E. K. Jo, M. Montano, P. F. Barnes, M. Selman and J. Granados. (2005) A functional promoter polymorphism in monocyte chemoattractant protein-1 is associated with increased susceptibility to pulmonary tuberculosis. J Exp Med. 202, 1649-1658
Fogg, D. K., C. Sibon, C. Miled, S. Jung, P. Aucouturier, D. R. Littman, A. Cumano and F. Geissmann. (2006) A clonogenic bone marrow progenitor specific for macrophages and dendritic cells. Science. 311, 83-87
Geissmann, F., S. Jung and D. R. Littman. (2003) Blood monocytes consist of two principal subsets with distinct migratory properties. Immunity. 19, 71-82
Ginhoux, F., F. Tacke, V. Angeli, M. Bogunovic, M. Loubeau, X. M. Dai, E. R. Stanley, G. J. Randolph and M. Merad. (2006) Langerhans cells arise from monocytes in vivo. Nat Immunol. 7, 265-273
Gonzalez-Mejia, M. E. and A. I. Doseff. (2009) Regulation of monocytes and macrophages cell fate. Front Biosci. 14, 2413-2431
Hall, J. D., S. L. Kurtz, N. W. Rigel, B. M. Gunn, S. Taft-Benz, J. P. Morrison, A. M. Fong, D. D. Patel, M. Braunstein and T. H. Kawula. (2009) The impact of chemokine receptor CX3CR1 deficiency during respiratory infections with Mycobacterium tuberculosis or Francisella tularensis. Clin Exp Immunol. 156, 278-284
Hohl, T. M., A. Rivera, L. Lipuma, A. Gallegos, C. Shi, M. Mack and E. G. Pamer. (2009) Inflammatory monocytes facilitate adaptive CD4 T cell responses during respiratory fungal infection. Cell Host Microbe. 6, 470-481
Holt, P. G., S. Haining, D. J. Nelson and J. D. Sedgwick. (1994) Origin and steady-state turnover of class II MHC-bearing dendritic cells in the epithelium of the conducting airways. J Immunol. 153, 256-261
Humphreys, I. R., G. R. Stewart, D. J. Turner, J. Patel, D. Karamanou, R. J. Snelgrove and D. B. Young. (2006) A role for dendritic cells in the dissemination of mycobacterial infection. Microbes Infect. 8, 1339-1346
Iijima, N., M. M. Linehan, S. Saeland and A. Iwasaki. (2007) Vaginal epithelial dendritic cells renew from bone marrow precursors. Proc Natl Acad Sci U S A. 104, 19061-19066
Inaba, K., M. Inaba, N. Romani, H. Aya, M. Deguchi, S. Ikehara, S. Muramatsu and R. M. Steinman. (1992) Generation of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony-stimulating factor. J Exp Med. 176, 1693-1702
Jakubzick, C., F. Tacke, F. Ginhoux, A. J. Wagers, N. van Rooijen, M. Mack, M. Merad and G. J. Randolph. (2008) Blood monocyte subsets differentially give rise to CD103+ and CD103- pulmonary dendritic cell populations. J Immunol. 180, 3019-3027
Jiao, X., R. Lo-Man, P. Guermonprez, L. Fiette, E. Deriaud, S. Burgaud, B. Gicquel, N. Winter and C. Leclerc. (2002) Dendritic cells are host cells for mycobacteria in vivo that trigger innate and acquired immunity. J Immunol. 168, 1294-1301
Kahnert, A., U. E. Hopken, M. Stein, S. Bandermann, M. Lipp and S. H. Kaufmann. (2007) Mycobacterium tuberculosis triggers formation of lymphoid structure in murine lungs. J Infect Dis. 195, 46-54
Kamath, A. T., S. Henri, F. Battye, D. F. Tough and K. Shortman. (2002) Developmental kinetics and lifespan of dendritic cells in mouse lymphoid organs. Blood. 100, 1734-1741
Kamphorst, A. O., P. Guermonprez, D. Dudziak and M. C. Nussenzweig. (2010) Route of antigen uptake differentially impacts presentation by dendritic cells and activated monocytes. J Immunol. 185, 3426-3435
Kang, S. J., H. E. Liang, B. Reizis and R. M. Locksley. (2008) Regulation of hierarchical clustering and activation of innate immune cells by dendritic cells. Immunity. 29, 819-833
Khader, S. A., J. Rangel-Moreno, J. J. Fountain, C. A. Martino, W. W. Reiley, J. E. Pearl, G. M. Winslow, D. L. Woodland, T. D. Randall and A. M. Cooper. (2009) In a murine tuberculosis model, the absence of homeostatic chemokines delays granuloma formation and protective immunity. J Immunol. 183, 8004-8014
Kurihara, T., G. Warr, J. Loy and R. Bravo. (1997) Defects in macrophage recruitment and host defense in mice lacking the CCR2 chemokine receptor. J Exp Med. 186, 1757-1762
Laouar, Y., T. Welte, X. Y. Fu and R. A. Flavell. (2003) STAT3 is required for Flt3L-dependent dendritic cell differentiation. Immunity. 19, 903-912
Leon, B. and C. Ardavin. (2008) Monocyte migration to inflamed skin and lymph nodes is differentially controlled by L-selectin and PSGL-1. Blood. 111, 3126-3130
Leon, B., M. Lopez-Bravo and C. Ardavin. (2007) Monocyte-derived dendritic cells formed at the infection site control the induction of protective T helper 1 responses against Leishmania. Immunity. 26, 519-531
Lin, Y., J. Gong, M. Zhang, W. Xue and P. F. Barnes. (1998) Production of monocyte chemoattractant protein 1 in tuberculosis patients. Infect Immun. 66, 2319-2322
Lindquist, R. L., G. Shakhar, D. Dudziak, H. Wardemann, T. Eisenreich, M. L. Dustin and M. C. Nussenzweig. (2004) Visualizing dendritic cell networks in vivo. Nat Immunol. 5, 1243-1250
Liu, K. and M. C. Nussenzweig. (2010) Origin and development of dendritic cells. Immunol Rev. 234, 45-54
Liu, K., G. D. Victora, T. A. Schwickert, P. Guermonprez, M. M. Meredith, K. Yao, F. F. Chu, G. J. Randolph, A. Y. Rudensky and M. Nussenzweig. (2009) In vivo analysis of dendritic cell development and homeostasis. Science. 324, 392-397
Liu, K., C. Waskow, X. Liu, K. Yao, J. Hoh and M. Nussenzweig. (2007) Origin of dendritic cells in peripheral lymphoid organs of mice. Nat Immunol. 8, 578-583
McCurley, N. and I. Mellman. (2010) Monocyte-derived dendritic cells exhibit increased levels of lysosomal proteolysis as compared to other human dendritic cell populations. PLoS One. 5, e11949
McKenna, H. J., K. L. Stocking, R. E. Miller, K. Brasel, T. De Smedt, E. Maraskovsky, C. R. Maliszewski, D. H. Lynch, J. Smith, B. Pulendran, E. R. Roux, M. Teepe, S. D. Lyman and J. J. Peschon. (2000) Mice lacking flt3 ligand have deficient hematopoiesis affecting hematopoietic progenitor cells, dendritic cells, and natural killer cells. Blood. 95, 3489-3497
Mohrs, K., A. E. Wakil, N. Killeen, R. M. Locksley and M. Mohrs. (2005) A two-step process for cytokine production revealed by IL-4 dual-reporter mice. Immunity. 23, 419-429
Murray, E., R. Webb and M. Swann. (1926) A disease of rabbits characterized by a large mononuclear monocytosis, caused by a hitherto undescribed bacillus Bacterium monocytogenes. J Pathol Bacteriol. 29, 407-439
Nahrendorf, M., F. K. Swirski, E. Aikawa, L. Stangenberg, T. Wurdinger, J. L. Figueiredo, P. Libby, R. Weissleder and M. J. Pittet. (2007) The healing myocardium sequentially mobilizes two monocyte subsets with divergent and complementary functions. J Exp Med. 204, 3037-3047
Naik, S. H., D. Metcalf, A. van Nieuwenhuijze, I. Wicks, L. Wu, M. O’Keeffe and K. Shortman. (2006) Intrasplenic steady-state dendritic cell precursors that are distinct from monocytes. Nat Immunol. 7, 663-671
Nakano, H., K. L. Lin, M. Yanagita, C. Charbonneau, D. N. Cook, T. Kakiuchi and M. D. Gunn. (2009) Blood-derived inflammatory dendritic cells in lymph nodes stimulate acute T helper type 1 immune responses. Nat Immunol. 10, 394-402
Niess, J. H., S. Brand, X. Gu, L. Landsman, S. Jung, B. A. McCormick, J. M. Vyas, M. Boes, H. L. Ploegh, J. G. Fox, D. R. Littman and H. C. Reinecker. (2005) CX3CR1-mediated dendritic cell access to the intestinal lumen and bacterial clearance. Science. 307, 254-258
Park, S. J., M. D. Burdick, W. K. Brix, M. H. Stoler, D. S. Askew, R. M. Strieter and B. Mehrad. (2010) Neutropenia Enhances Lung Dendritic Cell Recruitment in Response to Aspergillus via a Cytokine-to-Chemokine Amplification Loop. J Immunol.
Peters, W., H. M. Scott, H. F. Chambers, J. L. Flynn, I. F. Charo and J. D. Ernst. (2001) Chemokine receptor 2 serves an early and essential role in resistance to Mycobacterium tuberculosis. Proc Natl Acad Sci U S A. 98, 7958-7963
Randolph, G. J. (2006) Migratory dendritic cells: sometimes simply ferries? Immunity. 25, 15-18
Randolph, G. J., S. Beaulieu, S. Lebecque, R. M. Steinman and W. A. Muller. (1998) Differentiation of monocytes into dendritic cells in a model of transendothelial trafficking. Science. 282, 480-483
Randolph, G. J., K. Inaba, D. F. Robbiani, R. M. Steinman and W. A. Muller. (1999) Differentiation of phagocytic monocytes into lymph node dendritic cells in vivo. Immunity. 11, 753-761
Randolph, G. J., C. Jakubzick and C. Qu. (2008) Antigen presentation by monocytes and monocyte-derived cells. Curr Opin Immunol. 20, 52-60
Ravindran, R., L. Rusch, A. Itano, M. K. Jenkins and S. J. McSorley. (2007) CCR6-dependent recruitment of blood phagocytes is necessary for rapid CD4 T cell responses to local bacterial infection. Proc Natl Acad Sci U S A. 104, 12075-12080
Reljic, R., C. Di Sano, C. Crawford, F. Dieli, S. Challacombe and J. Ivanyi. (2005) Time course of mycobacterial infection of dendritic cells in the lungs of intranasally infected mice. Tuberculosis (Edinb). 85, 81-88
Robben, P. M., M. LaRegina, W. A. Kuziel and L. D. Sibley. (2005) Recruitment of Gr-1+ monocytes is essential for control of acute toxoplasmosis. J Exp Med. 201, 1761-1769
Russell, D. G. (2007) Who puts the tubercle in tuberculosis? Nat Rev Microbiol. 5, 39-47
Rydstrom, A. and M. J. Wick. (2007) Monocyte recruitment, activation, and function in the gut-associated lymphoid tissue during oral Salmonella infection. J Immunol. 178, 5789-5801
Sallusto, F. and A. Lanzavecchia. (1994) Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J Exp Med. 179, 1109-1118
Schreiber, H. A. and M. Sandor. (2010) The role of dendritic cells in mycobacterium-induced granulomas. Immunol Lett. 130, 26-31
Schreiber, H. A., P. D. Hulseberg, J. Lee, J. Prechl, P. Barta, N. Szlavik, J. S. Harding, Z. Fabry and M. Sandor. (2011) Dendritic cells in chronic mycobacterial granulomas restrict local anti-bacterial T cell response in a murine model. PLoS One. 5, e11453
Schreiber, O., K. Steinwede, N. Ding, M. Srivastava, R. Maus, F. Langer, J. Prokein, S. Ehlers, T. Welte, M. D. Gunn and U. A. Maus. (2008) Mice that overexpress CC chemokine ligand 2 in their lungs show increased protective immunity to infection with Mycobacterium bovis bacille Calmette-Guerin. J Infect Dis. 198, 1044-1054
Scott, H. M. and J. L. Flynn. (2002) Mycobacterium tuberculosis in chemokine receptor 2-deficient mice: influence of dose on disease progression. Infect Immun. 70, 5946-5954
Serbina, N. V. and E. G. Pamer. (2006) Monocyte emigration from bone marrow during bacterial infection requires signals mediated by chemokine receptor CCR2. Nat Immunol. 7, 311-317
Serbina, N. V., T. Jia, T. M. Hohl and E. G. Pamer. (2008) Monocyte-mediated defense against microbial pathogens. Annu Rev Immunol. 26, 421-452
Serbina, N. V., W. Kuziel, R. Flavell, S. Akira, B. Rollins and E. G. Pamer. (2003a) Sequential MyD88-independent and -dependent activation of innate immune responses to intracellular bacterial infection. Immunity. 19, 891-901
Serbina, N. V., T. P. Salazar-Mather, C. A. Biron, W. A. Kuziel and E. G. Pamer. (2003b) TNF/iNOS-producing dendritic cells mediate innate immune defense against bacterial infection. Immunity. 19, 59-70
Steinman, R. M. and Z. A. Cohn. (1973) Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J Exp Med. 137, 1142-1162
Steinman, R. M. and Z. A. Cohn. (1974a) Identification of a novel cell type in peripheral lymphoid organs of mice. II. Functional properties in vitro. J Exp Med. 139, 380-397
Steinman, R. M., D. S. Lustig and Z. A. Cohn. (1974b) Identification of a novel cell type in peripheral lymphoid organs of mice. 3. Functional properties in vivo. J Exp Med. 139, 1431-1445
Sunderkotter, C., T. Nikolic, M. J. Dillon, N. Van Rooijen, M. Stehling, D. A. Drevets and P. J. Leenen. (2004) Subpopulations of mouse blood monocytes differ in maturation stage and inflammatory response. J Immunol. 172, 4410-4417
Tacke, F., D. Alvarez, T. J. Kaplan, C. Jakubzick, R. Spanbroek, J. Llodra, A. Garin, J. Liu, M. Mack, N. van Rooijen, S. A. Lira, A. J. Habenicht and G. J. Randolph. (2007) Monocyte subsets differentially employ CCR2, CCR5, and CX3CR1 to accumulate within atherosclerotic plaques. J Clin Invest. 117, 185-194
Tacke, F. and G. J. Randolph. (2006) Migratory fate and differentiation of blood monocyte subsets. Immunobiology. 211, 609-618
Tian, T., J. Woodworth, M. Skold and S. M. Behar. (2005) In vivo depletion of CD11c+ cells delays the CD4 +T cell response to Mycobacterium tuberculosis and exacerbates the outcome of infection. J Immunol. 175, 3268-3272
Tsou, C. L., W. Peters, Y. Si, S. Slaymaker, A. M. Aslanian, S. P. Weisberg, M. Mack and I. F. Charo. (2007) Critical roles for CCR2 and MCP-3 in monocyte mobilization from bone marrow and recruitment to inflammatory sites. J Clin Invest. 117, 902-909
Ulrichs, T. and S. H. Kaufmann. (2006) New insights into the function of granulomas in human tuberculosis. J Pathol. 208, 261-269
Varol, C., L. Landsman, D. K. Fogg, L. Greenshtein, B. Gildor, R. Margalit, V. Kalchenko, F. Geissmann and S. Jung. (2007) Monocytes give rise to mucosal, but not splenic, conventional dendritic cells. J Exp Med. 204, 171-180
Volkman, H. E., H. Clay, D. Beery, J. C. Chang, D. R. Sherman and L. Ramakrishnan. (2004) Tuberculous granuloma formation is enhanced by a mycobacterium virulence determinant. PLoS Biol. 2, e367
Wakim, L. M., J. Waithman, N. van Rooijen, W. R. Heath and F. R. Carbone. (2008) Dendritic cell-induced memory T cell activation in nonlymphoid tissues. Science. 319, 198-202
Waskow, C., K. Liu, G. Darrasse-Jeze, P. Guermonprez, F. Ginhoux, M. Merad, T. Shengelia, K. Yao and M. Nussenzweig. (2008) The receptor tyrosine kinase Flt3 is required for dendritic cell development in peripheral lymphoid tissues. Nat Immunol. 9, 676-683
Whitelaw, D. M. (1966) The intravascular lifespan of monocytes. Blood. 28, 455-464
Wolf, A. J., L. Desvignes, B. Linas, N. Banaiee, T. Tamura, K. Takatsu and J. D. Ernst. (2007a) Initiation of the adaptive immune response to Mycobacterium tuberculosis depends on antigen production in the local lymph node, not the lungs. J Exp Med.
Wolf, A. J., B. Linas, G. J. Trevejo-Nunez, E. Kincaid, T. Tamura, K. Takatsu and J. D. Ernst. (2007b) Mycobacterium tuberculosis infects dendritic cells with high frequency and impairs their function in vivo. J Immunol. 179, 2509-2519
Zhan, Y., Y. Xu, S. Seah, J. L. Brady, E. M. Carrington, C. Cheers, B. A. Croker, L. Wu, J. A. Villadangos and A. M. Lew. (2010) Resident and monocyte-derived dendritic cells become dominant IL-12 producers under different conditions and signaling pathways. J Immunol. 185, 2125-2133
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer Science+Business Media, LLC
About this chapter
Cite this chapter
Schreiber, H.A., Sandor, M. (2012). Monocyte-derived Inflammatory Dendritic Cells in the Granuloma During Mycobacterial Infection. In: Lambris, J., Hajishengallis, G. (eds) Current Topics in Innate Immunity II. Advances in Experimental Medicine and Biology, vol 946. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-0106-3_16
Download citation
DOI: https://doi.org/10.1007/978-1-4614-0106-3_16
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4614-0105-6
Online ISBN: 978-1-4614-0106-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)