
Abstract
Increased use of nitrogenous (N) fertilizers in agriculture has significantly altered the global N-cycle because they release nitrogenous gases of environmental concerns. The emission of nitrous oxide (N2O) contributes to the global greenhouse gas accumulation and the stratospheric ozone depletion. In addition, it causes nitrate leaching problem deteriorating ground water quality. The nitrate toxicity has been reported in a number of studies showing the health hazards like methemoglobinemia in infants and is a potent cause of cancer. Despite these evident negative environmental as well as health impacts, consumption of N fertilizer cannot be reduced in view of the food security for the teeming growing world population. Various agronomic and genetic modifications have been practiced to tackle this problem. Some agronomic techniques adopted include split application of N, use of slow-release fertilizers, nitrification inhibitors and encouraging the use of organic manure over chemical fertilizers. As a matter of fact, the use of chemical means to remediate nitrate from the environment is very difficult and costly. Particularly, removal of nitrate from water is difficult task because it is chemically non-reactive in dilute aqueous solutions. Hence, the use of biological means for nitrate remediation offers a promising strategy to minimize the ill effects of nitrates and nitrites. One of the important goals to reduce N-fertilizer application can be effectively achieved by choosing N-efficient genotypes. This will ensure the optimum uptake of applied N in a balanced manner and exploring the molecular mechanisms for their uptake as well as metabolism in assimilatory pathways. The objectives of this paper are to evaluate the interrelations which exist in the terrestrial ecosystems between the plant type and characteristics of nutrient uptake and analyze the global consumption and demand for fertilizer nitrogen in relation to cereal production, evaluate the various methods used to determine nitrogen use efficincy (NUE), determine NUE for the major cereals grown across large agroclimatic regions, determine the key factors that control NUE, and finally analyze various strategies available to improve the use efficiency of fertilizer nitrogen.
The original version of this chapter was revised. An erratum to the chapter can be found at DOI 10.1007/978-3-319-51243-3_15
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
- Environment
- Phytoremediation
- Nitrate pollution
- Nitrogen oxides
- Ozone depletion
- Nitrogen use efficiency
- Nitrate reductase
- Nitrite reductase
- Glutamine synthase
- Nitrogen emission
- Nitrous oxide
- Environmental pollution
- N-efficient genotypes
- Methemoglobinemia
- Transcription factors
- Nitrogen metabolism pathway
- Nitrate toxicity
- Slow-release fertilizers
- Nitrate uptake
- Nitrate assimilation
- Transgenic plants
- Global warming
- Fertilizer use efficiency
- Nitrosamines
- Global climate change
1 Introduction
Nitrogen (N) is a vital element for life and an essential constituent of proteins, DNA, energy compounds like ATP, chlorophyll, vitamins and hormones (Galloway et al. 2002; Ohyama 2010; Abrol et al. 2012; Santiago-Antonio et al. 2014). In terrestrial and aquatic ecosystems, N supply determines the nature and diversity of plants, the population dynamics of herbivores, and vital ecological processes such as plant productivity and the cycling of carbon and soil minerals (Cassman et al. 2002; Dobermann and Cassman 2004; Boyer et al. 2004; Ladha et al. 2005a; Abrol et al. 2012; Soussana and Lemairec 2014).
The rigorous use of N fertilizers is leading towards a detrimental impact on environment in the agricultural production systems (Xu et al. 2000; Smil 2002; Zu et al. 2002; Killebrew and Wolff 2010). Toxicity due to nitrates can be either acute or chronic (Hill 1999; Bjorne et al. 2004). Acute toxicity is expressed when nitrite enters the bloodstream in large quantities and oxidizes the ferrous iron of hemoglobin to the ferric form producing methemoglobin, which cannot transport oxygen and results in methemoglobinemia (Fewtrell 2004). The reports show that over 80 % of the total nitrate intake in humans originates from vegetables and some part from the drinking water (Shoji et al. 2001; Croen et al. 2001; Anjana Umar and Iqbal 2007; Anjana Umar et al. 2007).
Although the risks have not been fully evaluated, several developed countries have established maximum permissible levels for nitrate concentration in vegetable food crops, and other countries are considering the establishment of similar standards. As a result of increasing pollution of sources adjacent to agricultural activities, the World Health Organization has set a limit of 10 mg of nitrate nitrogen per litre in domestic water supplies (Pionke et al. 1990; McPharlin and Aylam 1995). Research has shown that high concentrations of nitrate in plant food can decrease the level of ascorbic acid, a known inhibitor of the formation of carcinogenic compounds that contain nitrite.
Nitrogen deficiencies lead to chlorosis and reduced photosynthesis, which ultimately result in lower yields, and at the same time a significant reduction in the levels of phosphoenol pyruvate carboxylase, pyruvate orthophosphate dikinase, and Rubisco and a concomitant decrease in the level of their respective mRNAs in crop plants (Dordas and Sioulas 2008; Hakeem et al. 2014). The extensive use of nitrogenous fertilizers has ensured food security to developing nations, as the cereal production has kept pace with its ever-increasing population. Application of N fertilizers has increased global food production three times over the past 50 years (Rosegrant et al. 2002; Tilman et al. 2011).
Since 1962, the commercial production of N fertilizer has increased many folds to meet the growing requirements in the agriculture sector. It has been reported that doubling the agricultural food production worldwide over the past four decades has been associated with a seven folds increase in the use of nitrogenous (N) fertilizers (Hirel et al. 2007; Santiago-Antonio et al. 2014). However, it is excruciating to know that utilization of applied fertilizer nitrogen in the field by most cereal crops does not exceed 50 %. Among these, approximately 70 % of the total nitrogenous fertilizer is applied to rice and wheat cultivation (Pathak and Nedwell 2001; Norse 2003; Mosier et al. 2004a, b; Palm et al. 2004; Abrol et al. 2012; Hakeem et al. 2012a).
Such a poor N-use efficiency is a matter of great environmental concern. Even if the efficiency of nitrogenous fertilizers remains constant at the current levels, the N-losses will increase enormously as their consumption is expected to be doubled within the next few decades. Recent and future prospects of intensive N fertilizer use will result in detrimental impacts on the diversity and functioning of the variety of ecosystems (Tilman et al. 2011; Abrol et al. 2012). Manufacture of these fertilizers involves high-cost technology that requires a whole range of feed stocks and naphtha, a petroleum product. The environmental problems associated with it include NO3− pollution of the ground and surface waters and emission of oxides of N (N2O, NO, NO2−) which have positive radioactive forcing characteristics. Among these, NO2− is involved in stratospheric ozone depletion as well. Emission of unused nitrogen from agricultural fields in the form of NOx contributes to the global warming. NO2 has 298 times more global warming effect than CO2 (Signor and Cerri 2013). Their excessive and injudicious use, as reported from a number of regions, further accentuates the environmental degradation besides affecting the quality of crops, human and animal health, and causes lodging in cereals which may affect crop yields and quality. In surface water, presence of high N results in growth of algae and plants, thus accelerating eutrophication, and consequently affects water quality and usage (EPA 2007; Shen et al. 2011). The incidence of stomach cancer in humans, particularly in infants, and of non-Hodgkin’s lymphoma due to intake of water contaminated with nitrate has been reported (Mueller et al. 2001; Ikehata et al. 2010; Inoue-Choi et al. 2012). Nitrosamines produced from nitrite are reported to be carcinogenic; and NH3 gas is a pollutant because of its corrosive nature due to the formation of ammonium salts. Applied nitrogen not taken up by crops, thus constitutes a huge financial loss and a severe environmental risk. It has been estimated that by a 10 % increase in NUE would result in, the annual saving of US $ 5 billion and also substantial considerable improvement in environmental condition and soil quality (Gupta and Khosla 2012). However, Prasad (2013) stated that 1 % increase in NUE in India will account for saving of 0.17 million tonnes of N or 0.4 million tonnes of urea, which is equal to the annual production of four urea plants producing 1 lakh tonnes of urea per annum.
2 Historical Perspectives of N Use in Agriculture
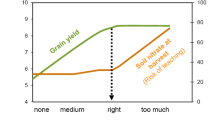
Nitrogen is the most crucial element which is essential for life (Abrol et al. 2012). Application of N fertilizers for improving crop yields and economic returns is historically recognized (Keeney and Hatfield 2008). N has been used for maximizing cereal grain yields in the twentieth century and cereal grain production has witnessed 40 percent increase per capita (Mosier et al. 2001). Nitrogen fertilizer use has increased tremendously over the last 50 years (Fig. 1). The major fraction of global N use is for increasing cereal grains production and N use will continue to increase by 15 % till the year 2015 (Mosier et al. 2004a, b). This unprecedented increase in the crop yield in the twenty-first century is possible only by using synthetic N fertilizers. The commercial production of synthetic N fertilizers has become possible due to the discovery of the Haber–Bosch process. However, increased use of N fertilizers has also led to increased N losses from agro-ecosystems and recovery of N in global crop production system (Millar et al. 2010; Subbarao et al. 2013) and rest may be lost into the atmosphere, accumulate in the soil or enter surface or subsurface water bodies through leaching or runoff. Nitrogen is lost from the soil-plant system through denitrification in the form of oxides of N like dinitrogen, nitrous oxides, nitric oxide, in the form of ammonia through volatilization, leaching of nitrates, run off and erosion (van Cleemput and Boeckx 2005; Hijleh 2014).

Histroical and predicted nitrogen fertilizer consumption in global agriculture (fertilizer.org/ifa/ifadata/results and FAO 2011)
3 Nitrogen Losses Through Oxides and Nitrates
3.1 Agriculture
The role of agricultural practices on environmental quality has been examined in the past two decades because of the links between increased agricultural productions accompanied by deterioration of environmental quality. In developing countries, intensive agricultural production systems have increased the use of N fertilizer as an effort to produce and sustain high crop yields (Fageria 2009). Consequently, N losses have also increased with passage of time (Fig. 2). Low recovery of N in annual crops is associated with its loss by volatilization, leaching, surface runoff and denitrification, which not only leads to high cost of crop production but also lead to environmental pollution (Jiao et al. 2012; Tilman et al. 2011).

Historical and predicted emission of anthropogenic reactive nitrogen in the global environment (fertilizer.org/ifa/ifadata/results and Zheng et al. 2002)
3.2 Emission of N from Fertilizers
Inorganic N applied to the soil can be absorbed by the plants, utilized by microorganism, subjected to nitrification, denitrification or volatilization (Vymazal 2007; Hakeem et al. 2012a). This represents an important source of N loss in agricultural soils under favorable conditions. Due to extensive use of N fertilizers and nitrogenous wastes, the amount of N available to plants significantly exceeds the N returned to the atmosphere by gaseous losses of N through volatilization and denitrification (Hakeem et al. 2012a). It has been reported that the concentration of N2O in environment has increased by 16.0 percent since 1750 (Godish et al. 2015). By passing each year an average of about 0.25 percent has been added and presently, the N2O concentrations are highest of the past 1000 years (Godish et al. 2015). A portion of this excess N has leached to ground waters as NO3− or has been carried in runoff waters. These are conducive conditions for N losses in agricultural soils which decrease the nitrogen use efficiency (NUE) (Hakeem et al. 2012a). Nitrate leaching to the ground water or run off into surface water bodies could lead to the levels beyond those acceptable for human consumption. Nitrate leaching into groundwater in arid regions and those containing sandy soils have been reported (Paramasivam et al. 2002; Hakeem et al. 2012a). Losses of N by leaching are affected by local variation in rainfall, water-holding capacity of soil, soil-drainage features and rates of mineralization of soil organic N (Subbarao et al. 2013). Processes such as adsorption, fixation, immobilization and microbial assimilation of added NH4-N in soils are of great importance as they affect nitrogen use efficiency and have the corresponding environmental repercussions (Kissel et al. 2004; Hakeem et al. 2012a). The immobilization and mineralization by microorganisms are most rapid when soils are warm and moist, but not saturated with water. The quantity of inorganic nitrogen available for crop use often depends on the amount of mineralization occurring, and the balance between mineralization and immobilization (Guleryuz et al. 2010). It influences the yield and quality of the crops but has a great impact on our food security, economy, development and environment (Walley et al. 2001, 2002; Tilman et al. 2002; Wood et al. 2004). It has been reported that human activities accounted for 53 % of total annual N input into Baltic Sea (HELCOM 2015). Granstedt (2000) reported that inputs in inorganic fertilizer were related to outputs of agricultural products and concluded that decreased input of fertilizers into system could help in decrease N losses from agriculture and increased recycling within the agricultural system.
Excessive NO3− concentration (300 mg L−1) in shallow ground waters due to application N of 500 kg N ha−1 has been reported in China (Zhang et al. 1996). They concluded that NO3-N problems in the environment would increase in China because of the increasing demand for food supply.
The large increase in the size of the hypoxic zone in the Gulf of Mexico after the 1993 floods in the Midwest focused attention on the role of agriculture in nonpoint source pollution (Burkart and James 1999). Several researchers in the past concluded that mineralization of soil organic matter, commercial fertilizer and row intensity of crop production are directly related to N losses (Burkart and James 1999; Schilling and Libra 2000; Mikkelsen and Hartz 2008). However, a similar analysis of Midwestern watersheds also showed that a relationship between total N load applied and NO3-N concentrations in Iowa and Illinois watersheds was not well-defined and there was not a simple relationship between water quality and agronomic management (Hatfield et al. 2001). Nitrate-N losses from watersheds are not isolated to the Mississippi River Basin. Throughout the world, there is increasing evidence that water quality is being degraded by the movement of nutrients from agricultural lands into water bodies. Environmental quality impacts from excess nutrients have been the major cause for the development of Mineral Accounting System (MINAS) in the Netherlands. This system provides an accounting for all nutrients entering and leaving a farm to ensure a balance of nutrients on farms (Ondersteijn et al. 2002). The MINAS approach is having a positive impact and has increased farmer awareness of the impacts that farming practices have on environmental quality and the natural resources (de Koeijer et al. 2003).
Non-agricultural sources of nitrogen contribute to less than 20 percent of the N released into the environment. About 6 percent are released from point sources (basically pipes) into water bodies, while, 14 percent is deposited from atmospheric sources. The production of reactive nitrogen worldwide has more than doubled in the last century because of human activities and population growth (Fig. 3). Advances in our understanding of the nitrogen cycle and the impacts of anthropogenic activities on regional to global scales are largely hindered by the paucity of information about nitrogen inputs from human activities in fast-developing regions of the world such as the tropics. In Brazil, the production of soybean crops over an area of less than 20 million ha, was responsible for about 3.2 Tg N or close to one-third of the N inputs from anthropogenic sources. Moreover, cattle pastures account for almost 70 percent of the estimated 280 × 106 ha of agricultural land in Brazil and potentially fix significant amounts of N when well managed, further increasing the importance of biological nitrogen fixation in the nitrogen budget. Many of these anthropogenic inputs occur in the Brazilian savannah region (Cerrado), while more urbanized regions such as the state of São Paulo also have high rates of nitrogenous fertilizer inputs. In the Amazon, rates of anthropogenic nitrogen inputs are relatively low, but continuing conversion of natural forests into cattle pastures or secondary forests potentially add a significant amount of new nitrogen to Brazil. Better measurements of biological fixation rates in Brazil are necessary for improving the nitrogen budgets, especially at a more refined spatial scale (Filoso et al. 2006).

Regional contribution of reactive nitrogen release into the atmosphere (fertilizer.org/ifa/ifadata/results and Boyer et al. 2004)
In most soils, formation and emissions of N2O to the atmosphere are enhanced by an increase in available mineral nitrogen (N) through increased rates of nitrification and denitrification (Signor and Cerri 2013). Therefore, addition of N, whether in the form of organic or inorganic compounds eventually leads to enhanced N2O emissions. Global N2O emissions from agricultural systems have previously been related primarily to fertilizer N input from synthetic sources (Robertson and Vitousek 2009). Little attention has been paid to N input from other N sources or to the N2O produced from N that has moved through agricultural systems. In a new methodology used to estimate N2O emissions on the country or regional scale, the anthropogenic N input data used include synthetic fertilizer and animal waste (feces and urine) used as fertilizer. N derived from enhanced biological N-fixation through N2 fixing crops and crop residue is returned to the field. Using FAO (2011) database information which includes data on consumption of synthetic fertilizer, animal and crop production and estimates of N input from recycling of animal and crop, estimates of total N into Asian agricultural systems and resulting N2O emissions are described over the time period (from 1961 to till date) (Fig. 1).
3.3 Statistics of Nitrogen Use
The quantity and relative amounts of different types of materials applied to agricultural soils, particularly in Asia as nitrogen (N) fertilizer has changed dramatically. Using the earliest record available from the FAO database, it was observed that in 1961, out of the approximately 15.7 Tg of fertilizer N applied to agricultural fields 2.1 Tg N (13.5 % of total N applied) was from synthetic sources, approximately 6.9 Tg N from animal wastes, 1.7 Tg N from biological N-fixation, and another 5 Tg N from re-utilization of crop residue. In 1994, 40.2 Tg from synthetic fertilizer N (57.8 % of total), 14.2 Tg from animal wastes, 2.5 Tg from biological N-fixation and 12.6 Tg from crop residue totaling 69.5 Tg N were utilized within agricultural soils in all Asian countries. The increases in N utilization have increased the emission of nitrous oxide from agricultural systems. Estimated N2O from agricultural systems in Asia increased from about 0.8 Tg N2O-N in 1961 to about 2.1 in 1994. The period of time when increases in N input and resulting N2O emissions were greatest was during 1970–1990. This evaluation of N input into Asian agricultural systems and the resulting N2O emissions demonstrates the large change in global agriculture that has occurred in recent decades. Increase in N input is likely because of the increased need for food production. Although the rate of increase of N input and N2O emissions during the 1990s appears to have declined, the question arises that if this had slowed the rate of increase in a general long term trend or if global food production pressures will tend to accelerate N input demand and resulting N2O emissions as we move into the twenty-first century (Mosier and Zhaoliang 2000). However, Zheng et al. (2002) used a model (IAPN 1.0) to analyzed the anthropogenic reactive nitrogen of Asia and stated that it has been increased from ~14.4 Tg N per year in 1961 to ~67.7 Tg N per year in 2000 and most is likely to be 105.3 Tg N per year by 2030. They concluded that the lack of effective measures to improve the efficiency of N fertilizer use and prevention of NOx emissions from fossil-fuel combustion, are the key carters behind the environmental N enrichment problem. Denman et al. (2007) reported that global N2O emissions reach about 17.7 Tg N per year, share 6.7 Tg (37.8 %) from anthropogenic sources and agricultural soils 2.8 Tg N per year (15.3 %) of the total amount of emissions. Similarly, Cerri et al. (2009) reported that the agriculture sector was responsible for the about 87.2 % of N2O emissions, mainly from animal waste management and agricultural soils of Brazil. However, Brazil (2010) reported that total anthropogenic sources contribute about 0.350 Tg N per year in Brazil. Out of total, fossil fuel combustion and industrial processes share 0.020 Tg N per year, agricultural sectors 0.310 Tg N per year, land use change and forests share 0.010 and other sources contribute about 0.009 Tg N per year.
4 Implications of Nitrates in Plants
Agricultural fields are intensively fertilized with N fertilizers to realized yield potential of different field crop varieties. Although, N can be absorbed by the plants in NH4+ form, but, major fraction N is available to the plants particularly vegetables in NO3− forms. Moreover, absorption and distribution of NO3− in the crops is important with respect to environmental consequences and quality of crop produce (Chen et al. 2004). However, when the input of N exceeds the demand, plants are no longer able to absorb it, and then the N builds up in the soil (Anjana Umar and Iqbal 2007). Unutilized NO3− by the plants may be subjected to ground water pollution by leaching or surface water pollution through soil erosion or run off, which influences the nitrate content of plants particularly the leafy vegetables (NAAS 2005).
5 Sources of N Compounds in the Environment
5.1 Nitrates
Agriculture is considered to be the major sources of NO3− to the ground water as considerable amount is leached from agricultural production systems around the world. Agricultural sources of NO3− to the ground or surface water includes arable grassland, grazed pastures, arable cropping, mixed cropping with pasture leys, organic farming, horticultural systems, and forest ecosystems (Ribbe et al. 2008). Accumulation of NO3− in the soil profile coupled, with or followed by periods of high drainage are conducive conditions for leaching of NO3− to the ground water. Therefore, application of N in excessive amount or waste effluent application rates or at the wrong time (e.g. late autumn) of the year, ploughing pasture leys early in the autumn, or long periods of fallow ground, can all potentially lead to high NO3− leaching losses. Different sources follow order with respect of potential of leaching: forest < cut grassland < grazed pastures, arable cropping < ploughing of pasture < market gardens (Di and Cameron 2002). Nitrogen applied through fertilizers or manure is converted to plant-available-nitrate by bacteria living in the soil. Nitrates are subjected different fates in the soil. These may be denitrified by bacteria and release to the environment as oxides of N, may be leached to ground water and absorb by the plants (Butterbach-Bahl et al. 2013). Another source is the use of chemical fertilizers (nitrogenous fertilizers) in intensive crop producing areas and, especially when they are used in an uncontrolled manner.
Nitrates are added into the natural water bodies through numerous sources like atmosphere, geological features, anthropogenic sources, atmospheric nitrogen fixation and soil nitrogen (Gormly and Spalding 2006; Gupta and Khosla 2012). However, detailed hydrogeological investigations have indicated a heterogeneous pattern of nitrate distribution. In sandy soil with low water holding capacity and high permeability, movement of pollutants like chloride and nitrate is much quicker than in clayey soil. This is probably the main cause for high nitrates in areas with sandy soil (Khandare 2013). However, nitrates are added into ground water by different sources which include septic tanks, animal and human waste and commercial fertilizers (Gormly and Spalding 2006; Khandare 2013). Nitrogen undergoes biochemical, chemical, and physical changes in soils and waters which make it difficult or impossible to segregate different sources with any degree of certainty (Keeney and Olson 1986). Agricultural sources represent the non-point sources of NO3− pollution of ground water which include fertilizers, manure application and legume crops (Shabalala et al. 2013). Non-point sources of nitrogen from agricultural activities include fertilizers, manure application, and leguminous crops while the point sources include septic tanks and dairy lagoons (Almasri and Kaluarachchi 2004; Shabalala et al. 2013). Elevated level of nitrate in ground water are common around dairy and poultry operations, barnyards, and feedlots (Odiyo et al. 2014). However, increase in the nitrate concentration of groundwater has been of special concern because of the suspected harm to children due to nitrate if its concentration in drinking water exceeds permissible limits of 50 mg L−1 (Kapoor and Viraraghavan 1997; Thorburn et al. 2003). Elevated NO3− concentration in drinking water causes methemoglobinemia in infants and stomach cancer in adults (Almasri and Kaluarachchi 2004; Anayah and Almasri 2009). Some studies on contamination of groundwater with nitrate originating from fertilizers have shown little evidence for such an occurrence (Yang et al. 1998; Mueller et al. 2001; Ikehata et al. 2010) while others have indicated an increase in the NO3 pollution of groundwater due to fertilizer application (Ju et al. 2005; Shen et al. 2011).
5.2 Nitrous Oxide
Nitrous oxide is primarily produced in soil by the activities of microorganisms during nitrification, and denitrification processes. The ratio of N2O to N2 production depends on oxygen supply or water-filled pore space, decomposable organic carbon, N substrate supply, temperature, and pH and salinity. N2O production from soil is sporadic, both in time and space, and therefore, it is a challenge to scale up the measurements of N2O emission from a given location and time to regional and national levels (Ussiri and Lal 2013). Estimates of N2O emissions from various agricultural systems vary widely. For example, in flooded rice in the Riverina Plains, N2O emissions ranged from 0.02 % to 1.4 % of fertilizer N applied, whereas in irrigated sugarcane crops, 15.4 % of the fertilizer was lost over a 4-day period. Nitrous oxide emissions from fertilized dairy pasture soils in Victoria range from 6 to 11 kg N2O-N/ha, whereas in arable cereal cropping, N2O emissions range from <0.01 % to 9.9 % of N fertilizer applications. Nitrous oxide emissions from soil nitrite and nitrates resulting from residual fertilizer and legumes are rarely studied, but probably exceed those from fertilizers, due to frequent wetting and drying cycles over a longer period and a larger area.
In ley cropping systems, significant N2O losses could occur, from the accumulation of mainly nitrate-N, following mineralization of organic N from legume-based pastures. Extensive grazed pastures and rangelands contribute annually about 0.2 kg N/ha as N2O (93 kg/ha per year CO2-equivalent). Tropical savannas probably contribute an order of magnitude more, including that from frequent fires. Unfertilized forestry systems may emit less, but the fertilized plantations emit more N2O than the extensive grazed pastures. However, currently there are limited data to quantify N2O losses in systems under ley cropping, tropical savannas, and forestry in Australia. Overall, there is a need to examine the emission factors used in estimating national N2O emissions; for example, 1.25 % of fertilizer or animal-excreted N appearing as N2O.
The primary consideration for mitigating N2O emissions from agricultural lands is to match the supply of mineral N (from fertilizer applications, legume-fixed N, organic matter, or manures) to its spatial and temporal needs by crops/pastures/trees. Thus, when appropriate, mineral N supply should be regulated through slow-release (urease and/or nitrification inhibitors, physical coatings, or high C/N ratio materials) or split fertilizer application. Also, N use could be maximized by balancing other nutrient supplies to plants. Moreover, non-legume cover crops could be used to take up residual mineral N following N-fertilized main crops or mineral N accumulated following legume leys. For manure management, the most effective practice is the early application and immediate incorporation of manure into soil to reduce direct N2O emissions as well as secondary emissions from deposition of ammonia volatilized from manure and urine.
Current models such as DNDC (DeNitrification-DeComposition) and DAYCENT (Daily Century) can be used to simulate N2O production from soil after parameterization with the local data, and appropriate modification and verification against the measured N2O emissions under different management practices. The DNDC model was developed to assess N2O, NO, N2 and CO2emissions from agricultural soils (Li et al. 2001). Recently, DAYCENT and DNDC model simulations of N2O fluxes were used for the assessment of the impacts of climate change on the gas flux and biomass production from a humid pasture (Abdalla et al. 2011). Similarly, DAYCENT model was used to quantify the on-farm GHG emissions and N dynamics of land use conversion to N-managed switchgrass in the Southern U.S (Jim et al. 2011).
In summary, improved estimates of N2O emission from agricultural lands and mitigation options can be achieved by a directed national research program that is of considerable duration, covers sampling season and climate, and combines different techniques (chamber and micrometeorological) using high precision analytical instruments and simulation modelling, under a range of strategic activities in the agriculture sector (Dalal et al. 2003).
5.3 Emissions During Crop Residues/Organic Matter Decomposition
Crop residue serves as the basic source for N in the agricultural practices during the process of nitrification and denitrification (Signor and Cerri 2013). It has been evident from the results of Gomes et al. (2009) that crop residues added to soil has high denitrification activity and N2O fluxes compared to field without crops. This may lead an increase in the soil aggregation, moisture retention capacity compared to the topsoil in ecological horizon. Thus, the incorporation of residues caused an accelerated effect on the NO emissions rate and the effects of tillage on mineralization of soil organic matter directly influence the rate of NO and N2O emissions from soils. Some earlier investigations in the past by researchers have observed that higher N2O losses are due to high denitrification activity for no-tillage systems compared to conventional tillage (Toma and Hatano 2007). This may be related to the higher denitrification activity. In a no-tillage crop system, soil N2O emissions were lower (Escobar et al. 2010). However, N2O losses from no-tillage systems may be lower than from tilled soils where fields remained uncultivated for a number of years. Some studies report an increase in NO emission caused by ploughing in a temperate climate and under tropical conditions.
5.4 Non-agricultural Sources
The nutrients present in sewage and industrial effluent can be effectively used as a supplement to the chemical fertilizer if managed properly (Emongor and Ramolemana 2004). Direct discharge of septic tanks, sewage and industrial effluent causes pollution of surface water and ground water through percolation. In groundwater recharge areas with large portions of agricultural land, the nitrate concentration of well water has shown rising trends in many countries within the last two- to three decades. Nitrate leaching from agricultural land must be considered as an important non-point source for nitrate contamination of the groundwater. In developing countries, most of the urban areas have not well developed system for collection and dumping of domestic and industrial wastes. In most areas, sewerage system may be non-functional or non-existent which led to disposal of human fecal matters and industrial wastes directly into the surface drains. In other cases, most of houses have septic-soil absorption system for wastewater disposal which could lead to the contamination of ground water with NO3− (Ramaraju et al. 1999).
6 Atmospheric Deposition
Nitrogen is atmospherically deposited to surface water bodies downwind of anthropogenic emissions. Atmospheric deposition accounted for 10 % to over 40 % of new N loading to estuaries in eastern U.S. coast and eastern Gulf of Mexico (Paerl et al. 2002). Historically, N deposition due to urbanization (NOx, peroxyacetyl nitrate or PAN) and agricultural expansion (NH4+ and possibly organic N) has increased in coastal airsheds. Ammonium is dominant form of atmospherically deposited N in western Europe due to abundance of livestock operations while in United States, dominant form of atmospheric N is oxides of N (NOx) emitted from fossil fuel combustion (Paerl et al. 2002). These quantitative and qualitative aspects of atmospherically deposited N may promote biotic changes now apparent in estuarine and coastal waters, including the proliferation of harmful algal blooms, with cascading impacts on water quality and fisheries. In addition to these sources, oxides of N are generated through lightening and with rain water these oxides of nitrogen are available in water (Schumann and Huntrieser 2007). The atmospheric contribution was supposed to be the smallest but recent reports on dry and wet depositions of pollutants suggested a contribution to the extent of 25 % of the total load of nitrate (Gupta et al. 2007).
7 Health Risks of Nitrate and Nitrogen Oxides
The use of nitrogenous fertilizer is reported to be the major source of NO3− contamination of groundwater (Nolan and Stoner 2000), and if this contaminated groundwater is pumped for drinking purpose and may have a direct effect on human health. The risk of ground-water contamination by nitrogenous sources depends on the input of nitrogen input to the land surface and the degree of leaching and accumulation. Recently, an agricultural nonpoint source is held responsible for the pollution of rivers and lakes. The impact of agricultural practices on groundwater quality is of particular concern as in many cases, ground water level is shallow and thus are vulnerable to water pollution, particularly from NO3−. Use of high NO3− irrigation water could also lead to toxic levels of NO3− in leafy vegetables. Although current epidemiological data provide conflicting evidence regarding the potential long-term health risks of nitrate levels encountered in the diet, it is widely accepted that the reduction of dietary nitrate is a desirable preventive measure, as it may pose serious health hazards. The toxic effects of nitrate are due to its endogenous conversion to nitrite and this ion has been implicated in the occurrence of methaemoglobinemia, gastric cancer and many other diseases (Anjana Umar et al. 2007). Initially, the risk of methaemoglobinemia was limited to the infants only (Greer and Shannon 2005), but later, Gupta et al. (2008) have reported it in all age groups with high nitrate ingestion, with the infants and higher age groups (>45 years) being the most susceptible to nitrate toxicity. Apart from this, effect of nitrate consumption on human health has been studied in detail by Gupta et al. (2008). Although attention has been paid to the level of nitrate in drinking water and the extent of nitrate pollution of groundwater bodies in India due to heavy nitrogen use in India, there is a gap in our knowledge regarding the concentration of nitrate in leafy vegetables consumed regularly.
The quality of drinking water is harmfully influenced by nitrate through the possible formations of nitrite causing methemoglobinemia in babies and cancer in older persons due to the formation of nitrosamines in the digestive tract (Ward 2009). The World Health Organization (WHO) recommends that the safe level of NO3-N in drinking water is 11.3 mg NO3-N dm−3, the unsafe level is set at 22.6 mg NO3-N dm−3. Besides the concern for the environment, the use of high amounts of fertilizer N is also prohibitive because of its high cost. The applied NO3-N not taken up by the plant can be leached into the groundwater. It depends on soil texture, rainfall, N fertilization and type of crop (Liu et al. 2012). Several research results indicated that the fertilizer N management has a great influence on groundwater contamination, and it is suggested that with proper N management, groundwater pollution as well as economic loss can be avoided (Liu et al. 2012; Hijleh 2014).
8 Acute Toxic Effects
Exposure to high nitrate may cause acute toxicity due to acute exposure.
8.1 In Human
Nitrate toxicity is mainly caused due to reduction of nitrates to nitrites after ingestion. The health hazards is directly caused by nitrite that oxidizes the hemoglobin to methemoglobin (Manassaram et al. 2007; Tables 1, 2, and 3). Drinking water is the major source of nitrates intake into human body thereby contributing about < 3–21 % of the average adult intake of nitrate besides other sources like vegetables and meat (WHO 2011). The potential health hazard related to dietary nitrate is formation of N-nitroso compounds from nitrite. Nitrates are also suspected to be involved in some other human diseases. These diseases include cancer due to production of N-nitroso compounds, hypertension, increased infant mortality, central nervous system birth defects, diabetes, spontaneous abortions, respiratory tract infections, and changes to the immune system (Fewtrell 2004; Chiu and Tsai 2007). Methemoglobin (MetHb) is produced due to the oxidation of ferrous (Fe2+) iron in hemoglobin to ferric (Fe3+) which affects oxygen binding capacity of MetHb. This results in expression of cyanosis, stupor, and cerebral anoxia (Fewtrell 2004). Human lethal doses of 4–50 g NO3− (equivalent to 67–833 mgNO3−/kg bw) have been reported. Toxic doses with methemoglobin formation as a criterion for toxicity-ranged from 2 to 5 g of NO3 (WHO 2011). These values are equivalent to 33–83 and 100–150 mg NO3−/kg body weight respectively.
8.2 In Animals
Acute nitrate toxicity to animals varies according to species. Generally, ruminants suffer from methemoglobinemia while monogastric animals may face severe gastritis. Ingestion of nitrate also cause impaired functioning of thyroid, decreased feed consumption, and interference with vitamin A and E metabolism (Bruning-Fann and Kaneene 1993; McDowell 2012). Hematologic changes caused due to high nitrate exposure include both compensatory increases in red blood cells and anemia, along with increased neutrophils and eosinophils. Unlike nitrate, nitrite is capable of inducing methemoglobinemia in a wide range of species, i.e. cattle, sheep, swine, dogs, guinea pigs, rats, chickens and turkeys. In rats, chronic nitrite exposure causes pathologic changes in a variety of tissues, alterations in motor activity and brain electrical activity, and alters gastric mucosal absorption (Bruning-Fann and Kaneene 1993; WHO 2011). Nitrite affects the metabolism of sulfonamide drugs in animals such as the pig, guinea pig, and rat. The N-nitroso compound di-methylnitrosamine causes toxic hepatosis in cattle, sheep, mink, and fox. Nitrosamines have been reported in cow’s milk and been found to pass into the milk of goats under experimental conditions (WHO 2011).
The studies relating to acute exposure of nitrate were conducted in animals. The acute oral toxicity of nitrate to laboratory animals is low to moderate. LD50 values of 1600–9000 mg of sodium nitrate per kg of body weight have been reported in mice, rats, and rabbits. Ruminants are more sensitive to the effects of nitrate as a result of high nitrate reduction in the rumen; the LD50 for cows was 450 mg of sodium nitrate per kg of body weight. Nitrite is more toxic than nitrate: LD50 values of 85–220 mg of sodium nitrite per kg of body weight have been reported for mice and rats (WHO 2011).
8.3 Mechanism of Methemoglobin Formation
The essential action in the formation of methemoglobin is an oxidation of the ferrous to the ferric ion. This oxidation may be brought about in one of the following ways (Sharma et al. 2013): by the direct action of the oxidants, the action of hydrogen donors in the presence of oxygen, and by auto oxidation. In the presence of nitrites, the ferrous ion of hemoglobin gets directly oxidized to the ferric state. Normally, the methemoglobin that is formed is reduced by the following reaction.
Oxidized cytochrome b5 (Red Cyt b5) is regenerated by the enzyme cytochrome b5 reductase:
Thus, the enzyme cytochrome b5 reductase plays a vital role in counteracting the effects of nitrate ingestion. However, permissible concentration of nitrate (50 mg L−1) normally present in water or food do not cause any risk associated with the human.
8.4 Water Bodies
A considerable amount of N is deposited into estuarine and coastal waters due to atmospheric deposition downwind of anthropogenic emissions. Approximately, 40 % of total N added to the eastern U.S. coast and eastern Gulf of Mexico is contributed by atmospheric deposition. In receiving estuarine and coastal waters, phytoplankton community structural and functional changes, associated water quality, and trophic and biogeochemical alterations (i.e. algal blooms, hypoxia, food web, and fisheries habitat disruption) are frequent consequences of N-driven eutrophication. Increases in and changing the proportions of various new N sources regulate phytoplankton competitive interactions, dominance, and successional patterns. These quantitative and qualitative aspects of AD-N and other atmospheric nutrient sources (e.g. iron) may promote biotic changes now apparent in estuarine and coastal waters, including the proliferation of harmful algal blooms, with cascading impacts on water quality and fisheries (Paerl et al. 2002).
8.5 Ozone Depletion
Among the various factors, N2O is the most important factor responsible for ozone-depletion and it will continue to be on the top of all the factors in twenty-first century. Decreasing the emission of N2O would help in amelioration of ozone layer depletion (Ravishankara et al. 2009). Ozone layer in stratosphere is depleted by different chemicals of anthropogenic origin which is a serious environmental threat of this era. Oxides of N are major factors which are responsible for the depletion of the ozone layer. Depletion of the ozone layer by oxides of N is explained by the following reactions;
The main cause behind the depletion of the ozone layer is the stockpile of halocarbons in the stratosphere through various heterogeneous chemical reactions with aerosol particles (Solomon 1999). Amongst the various ozone depleting gases, N2O is the most predominant gases emitted mainly by human activities (Ravishankara et al. 2009), and the emission of N2O causes maximum hazards to the environment when released either singly or in combination with other hydrocarbon (Daniel et al. 2010). Moreover, change in the concentration of N2O may also be directly responsible for the chemical and radioactive changes in the stratospheres (Portmann et al. 2012).
9 Remediation Measures
In the present scenario a wide range of remedial measures are available for the treatment of water bodies. Blending low-nitrate in high-nitrate water is one of the cheapest methods, but it depends upon the availability of low-nitrate water supplies in the surrounding area. In this regards, other methods such as biological denitrification, ion exchange, reverse osmosis and electro-dialysis are the most feasible alternatives. Previously researchers concluded that biological denitrification and ion exchange as the most striking options for the treatment of water bodies in the UK followed by reservoir storage, lining of boreholes and relocation of boreholes.
9.1 Concepts of Nitrogen Use Efficiency (NUE)
The term NUE has two basic components; (1) Nitrogen uptake, recovery or acquisition efficiency and (2) Nitrogen use, physiological N use, or internal N use efficiency. The terms NUE is a ratio between an output (i.e., grain yield, dry matter yield, N accumulation in grain, or N accumulation in total plant dry matter) and input (i.e. total N supply, soil N supply or fertilizer N supply). NUE is based on different parameters of efficiency, including N uptake, utilization or acquisition efficiency, and N-use (physiological/internal) efficiency. Nitrogen use efficiency and crop response to applied N are important for evaluation of N requirements for crop to achieve maximum economic yield. Globally, recovery of N in crop plants is generally < 50 % and remaining is lost due to volatilization, leaching, surface runoff, denitrification, and plant canopy (Fageria 2009). Low recovery of N leads to higher cost of crop production and environmental pollution. Under such conditions, enhancing NUE will improve crop yields, reducing cost of production, and maintaining environmental quality. NUE can be classified into agronomic, recovery and physiological efficiency.
Agronomic efficiency of nitrogen measures is the overall efficiency or an integrative index of total economic outputs relative to the use of all sources of N (indigenous soil N and applied fertilizer N). Nitrogen recovery efficiency considers the efficiency of the plant to take up N and physiological N use efficiency is the efficiency with which the plant uses N from acquired available N to produce grain or total plant dry matter. To calculate the efficiency of soil vis-a-vis fertilizer N, the output from a non-fertilizer or control plot where the source of N is only soil should be determined. The average agronomic N efficiency had a relatively narrow compared to applied N applied, and amongst the three cultivates crops it was recorded least in maize and the maximum in rice crops. These differences in agronomic efficiency indicate that maize and rice produce larger economic outputs than wheat in terms of the use of all N sources (soil and fertilizer). The average estimates of N recovery efficiency based on grain N were similar among the three crops and were highest (44 %) in maize and lowest (35 %) in wheat. The average physiological efficiency of N ranged from 35 to 69 kg grain increase per kg of N uptake. The main reason for large physiological efficiency of N values with rice was the crop’s low grain-N concentration (9–12 g kg−1), especially when compared with that of maize (13–14 g kg−1) and wheat (16–18 g kg−1). Rice is also known to be highly efficient in mobilizing N from senescing leaves and clumps to grain as indicated by its large N-harvest index (Ladha et al. 2005a) suggested that the mode of photosynthesis (C3 or C4 photosynthetic pathway) and grain-N concentrations govern PEN.
In general, grain yields increase with an increase in total N uptake and some genotypes produce different grain yields with the same amount of N uptake. Differences in the efficiency of physiological N use may arise from differences in (1) internal N requirement for plant growth, (2) ability of the plant to translocate, distribute and mobilize absorbed N to and from various organs, (3) flag leaf N import/export and leaf senescence pattern and (4) plant's efficiency in converting CO2 to carbohydrate (Ladha et al. 2005a).
Proper N-application timing and rates are critical for meeting plant needs and improving NUE. In addition, the growth stage of plants at the time of fertilizer application also determines NUE, with significant genotypic variations (Hirel et al. 2007; Hakeem et al. 2011). It was opined that the timing of fertilizer N applications has a significant effect on the uptake of fertilizer N by the crop and the resulting partitioning of added N between soil and plant. While the amount of N available to the plant can be improved by using sustained-release fertilizers, split applications, minimizing fertilizer losses and other nutrient management and crop management strategies, the inherent efficiency of the plant to utilize available N for higher productivity needs to be tackled biologically (Abrol et al. 1999). Nitrogen applications in split doses have been found to increase yield, NUE, and N uptake efficiency (NUpE), compared with fall application in hard red winter wheat under temperate conditions (Abrol et al. 1999; Sowers et al. 1994).
Nitrogen use efficiency (NUE) in the context of photosynthesis is called as photosynthetic nitrogen use efficiency (PNUE), which is determined by the rate of carbon assimilation per unit leaf nitrogen (Kumar et al. 2001). As Ribulose bisphosphate carboxylase/oxygenase (Rubisco) is a major sink for nitrogen supplied in the form of fertilizers, manipulation of crops to improve NUE would have economic and environmental benefits, reducing the pollution of water by nitrates.
9.2 Transgenomic Approaches
9.2.1 Nitrogen Use Efficiency
Various approaches have been used for the improvement of N-use efficiency (NUE) among crop plants and also to reduce N losses from agricultural fields. Application of the same amount of N in more than two splits under field conditions increases the nitrogen availability at later stages of growth to exploit the sub-optimal activity of the upper leaf laminae (Abrol et al. 1999). Differential regulations of various nitrates and ammonium transporters in plants are well known (von Wirén et al. 2000; Bu et al. 2011), however, studies regarding the effect of over expression of genes of these transporters are very rare. Studies involving transgenic over expression of a chl1 cDNA (representing the constitutive high-affinity nitrate transporter) driven by the cauliflower mosaic virus 35S promoter in a chl1 mutant have revealed effective recovery of the nitrate uptake defect for the constitutive phase (Liu and Tsay 2003). This was however not reflected in the case of induced phase, which was consistent with the constitutive level of chl1 expression in the transgenic plant. In another experiment involving transgenic tobacco plants expressing the NpNRT2.1 gene (encoding high affinity nitrate transporter), a steady-state increase in its mRNA levels accompanied by an increase in the NO3− influx, but the NO3− contents were remarkably similar in wild-type and transgenic plants has been reported (Fraisier et al. 2000). These findings indicate that increasing the uptake of nitrate by genetic manipulation may not necessarily lead to a concomitant improvement in nitrate utilization or NUE, though it remains to be seen whether different plants respond differently to the overexpression of different transporters. Ammonium transporters are very well characterized in plant systems (Glass 2003; Loqué and von Wirén 2004), but the effect of their over expression on plant growth and development is yet to be elucidated.
9.2.2 Nitrate Reductase (NR)
Nitrate reductase (NR) has long been considered to be the rate-limiting step in nitrate assimilation, but transgenic manipulation of NR expression in Nicotiana spp. indicated the importance of other steps (Stitt 1999). Constitutive NR expression led to a twofold increase in NR activity and a 20 % decrease in foliar nitrate content along with an increase in the total amino acid contents, but without any changes in the total N, soluble sugars, starch and productivity parameters (Masclaux et al. 2001). While the NR double mutant Nia30 does not show detectable NR activity, when transformed with the nia2-cDNA, it showed decreased NR activity with higher levels of nitrate accumulation (Hänsch et al. 2001). Transformed Nicotiana plumbaginifolia plants constitutively expressing nitrate reductase (NR) show temporarily delayed drought-induced losses in NR activity, thereby allowing for the more rapid recovery of N assimilation following a short-term water deficit. Deregulation of NR gene expression by constitutive expression in transgenic plants resulted in reduced nitrate levels in the tissues of tobacco (Quilleré et al. 1994). Although other factors such as NO3− availability regulate the flux through the pathway of N assimilation, the NR transformants were better equipped in terms of available NR protein, which rapidly restore N assimilation. While no tangible effects on biomass accumulation could be attributed in the short term, under field conditions of fluctuating water availability, constitutive NR expression was able to confer a physiological advantage by preventing slowly reversible losses in N-assimilation capacity (Ferrario-Méry et al. 1998). Since NR is post-translationally regulated by phosphorylation and binding of 14-3-3 proteins, various attempts have been made to counter this inhibitory effect on NR regulation. Deletion of 56 amino acids in the amino-terminal domain of NR was previously shown to impair this type of regulation in Nicotiana plumbaginifolia (Provan et al. 2000). Additional reports of over-expression of NR genes from various plants have been accumulating over the last decade (Ferrario-Méry et al. 2001; Lillo et al. 2004) but none of these has had any major implication for improving the nitrogen-use efficiency.
9.2.3 Nitrite Reductase (NiR)
Overexpression of NiR genes in Arabidopsis and tobacco resulted in increased NiR transcript levels but decreased enzyme activity levels, which were attributed to post translational modifications (Takahashi et al. 2001). There is no evidence yet of any benefit of NiR over-expression in terms of plant NUE.
9.2.4 Glutamine Synthetase (GS)
Transgenic tobacco plants enriched or reduced in plastidic glutamine synthetase (GS2, a key enzyme in photorespiration) were constructed (Kozaki and Takeba 1996). Those transgenic plants having twice the normal amount of GS2 had an improved capacity for photorespiration and an increased tolerance to high-intensity light, whereas those with a reduced amount of GS2 had a diminished capacity for photorespiration and were photo-inhibited more severely by high-intensity light compared with the control plants. Reassimilation of ammonia in transformed tobacco was also studied by Ferrario-Méry et al. (2001), while, Hoshida et al. (2000) investigated the role of constructs for the over expression of GS2 in rice plants (Hoshida et al. 2000). Ectopic expression of GS1 has been shown to alter plant growth (Fuentes-Ramírez et al. 2001) and the over expression of GS1 in transgenic plants could cause the enhancement of photosynthetic rates, higher rates of photorespiration and enhanced resistance to water stress (El-Khatib et al. 2004; Fuentes-Ramírez et al. 2001). The overexpression of soybean cytosolic GS1 in the shoots of Lotus corniculatus was reported to accelerate plant development, leading to early senescence and premature flowering, particularly when plants were grown under conditions of high ammonium (Vincent et al. 1997). Ectopic expression of pea GS1 in tobacco leaves was suggested to provide an additional or alternative route for the reassimilation of photorespiratory ammonium, resulting in increased efficiency of N assimilation and enhanced plant growth (Oliveira et al. 2002). Some additional empirical evidence for enhanced nitrogen-assimilation efficiency in GS1 transgenic lines were also provided (Man et al. 2005). However, differences in the degree of ectopic GS1 expression have been reported (Fuentes et al. 2001) and attributed to positional effects, effectiveness of chimeric constructs, or differences in growth conditions. These differences could account for the lack of correlation between the enhanced expression of GS1 and concomitant growth (Ortega et al. 2001; Vincent et al. 1997). Transgenic overexpression and antisense technology have been employed recently to modulate the expression of NADH-GOGAT in alfalfa and rice plants (Schoenbeck et al. 2000; Yamaya et al. 2002). The studies on transgenic rice plants expressing antisense RNA for either GS1 or NADH-GOGAT point towards the possible involvement of GS1 in the export of N via phloem in senescing leaves. On the other hand, in case of developing leaf blades and spikelets, NADH-GOGAT was implicated in the utilization of glutamine transported from senescing organs (Yamaya et al. 2002). While these genes appear to be good candidates for improving NUE in the short run, the degree of improvement may vary with the crop and cropping conditions.
9.2.5 Transcription Factors
While, identification of global nitrate-induced transcription factors in higher plants has not been successful, a recent study suggests that targeting other transcription factors may help improve N assimilation and NUE. Yanagisawa (2004) generated transgenic Arabidopsis lines overexpressing Dof1, a maize protein that belongs to the Dof family of plant-specific transcription factors known to activate the expression of several C-metabolizing genes associated with organic acid metabolism. The transformants showed up to 30 % higher N content, higher levels of amino acids, better growth under low-nitrogen conditions and higher levels of mRNAs and enzyme activities for PEP carboxylase and pyruvate kinase, without any reduction of NR, GS and GOGAT RNAs. The genes up regulated by Dof1 overexpression clearly belong to the list of known nitrate-responsive genes, though it is not clear whether Dof1 is inducible by nitrate. If Dof1 is not nitrate-inducible, it means that multiple transcription factors may be involved in the coordinated expression of the N and C metabolizing genes. A few other attempts to manipulate signaling/regulatory proteins have been made, but did not yield any significant advantage in terms of NUE. Castaings et al. (2011) has recently reviewed the roles of nitrate transporters, transcription factors, and kinases in their association with hormones or N-containing molecules in the regulation of N assimilation.
9.2.6 Omics Approach in Searching the Candidate Genes/Proteins to Enhance Nitrogen Use Efficiency
Ove the last decade, omics approach has been used to understand the mechanism of physiological characteristics and to search the possible candidate genes/proteins responsible to various physiological processes in plants. However, It has been found that these physiological processes are showing a multitude of complex networks and interdependent pathways involving many genes, proteins, enzymes, and metabolites, than to a single pathway (Fernie and Stitt 2012).
Recently, Simons et al. (2014) reviewed the role of omics studies in understanding NUE in Maize. There are a few proteomics studies avaliable in various crop plants viz., rice (Hakeem et al. 2012b, c), wheat (Bahrmam et al. 2004; Bahrman et al. 2005), maize (Amiour et al. 2012) to investigate into the response of a plant to N limitation. Similarly, metabolomics approach has been valuable in improving our understanding of the interactions between C and N metabolism (Fait et al. 2011). however, these studies does not show any direct relationship with transcriptome or proteome studies. Hence, it is suggested that when integrated with other ‘omics’ data and with information from quantitative genetics, transcriptomics and interactomics, we shall be able to understanding the complex regulatory networks underlying important phenotypic traits such as yield and nutrient perception and utilization (Verma et al. 2013).
10 Conclusion and Future Prospectives
Nitrate is one of the most extensively studied anion in water because of its chronic toxic manifestations. It is one of the common pollutants of drinking water all over the world. Despite the availability of the vast literature on nitrate toxicity, the studies of its impacts on the human populations are still limited because of ethical and other considerations, and, the most toxic manifestations are available in animal studies only. Nitrogen oxides stay more than 100 years in the atmosphere, and each molecule of nitrogen oxide absorbs 200 times more radiation than carbon dioxide. This will also play an important role in global climate change (Ozturk et al. 2013). Numerous epidemiological studies have also been conducted to establish certain cause-effect relationships for many clinical manifestations of nitrate toxicity and the results of such studies are normally inconclusive or the correlations are statistically non-significant (Cristina et al. 2014). This has led to a very insufficient knowledge generation among researchers and practicing doctors about the preventive and curative aspects of different manifestations. The farming community should be made aware of the harmful effects of N2O emission from agriculture and should feel the need to mitigate its emission through optimum crop management practices. Inexpensive soil and plant tissue testing facilities should be made available to farmers.
The world, has been caught to the desirability of nitrogen fertilizers very quickly, and faced with its bad consequences before thinking and doing research on the effects on environmental health and the possible influence on climate. As a remedy to this dangerous trend, two focal points should be considered: first, we should encourage organic farming, and reduce the use of artificial fertilizers and pesticides.
Nitrate-containing forages may be used with proper precautions. Problems with nitrate toxicity can be avoided with proper management of forage and livestock. Drought-stricken forages may contain toxic amounts of nitrates for cattle, and prevention of problems is essentially the only valid approach to avoiding economic loss. Suspected forages should be tested and results used in the feed management. Limiting forage intake and nitrate-free feeds and preventing animals from eating nitrate-containing forages, are important in avoiding cattle losses, silages suspected of toxic levels of nitrate should be allowed to aerate overnight before feeding (Ozturk et al. 2013). Ammonia, butyric acid and non-protein nitrogen levels of the silage increase when nitrates are present in the silage crop. Enterobacteria are mainly responsible for the degradation of nitrate in silage, Clostridia can reduce nitrite to ammonia, Lactobacilli reduce nitrate. The type of plant material and nutritional value of forage crops is closely related with the quality. There is a positive correlation between nitrogen fertilization of plants and their crude protein content, 1000 ppm or more nitrate (upto 2000 ppm) in dry matter content has been reported to be an acceptable criterion for the quality of herbs (Ozturk et al. 2013).
There is an immediate need for considering the estimation of the amount of nitrogen fertilizers to be applied to the fields or under glasshouses as kg N/ha rather than the amount of fertilizer per ha. At the same time fertilizer application and irrigation practices reducing nitrogen losses in runoff should be used. The importance of environmental education comes to the forefront here. With an increase in the level of education of the users, the importance given to the analysis increases. Such people avoid unnecessary and excessive fertilizer usage due to environmental awareness. The soil nitrogen supply status can be deduced from the previous cropping, and manurial history of the soil, and should be categorised into an index system.
The recommendations for nitrate containing fodder use are: feed containing 1000 ppm nitrate could be used securely; feeds with nitrates between 1000 and 2000 ppm are slightly risky; forages containing nitrate between 2000 and 3400 ppm should not be given to animals, if given, the ratio between non nitrate to nitrate feeds should not exceed 50 percent; forages containing nitrate between 3400 and 4000 ppm should be kept away from the pregnant animals, If given, ratio between non nitrate to nitrate feeds should not exceed 25 percent; feeds containing more than 4000 ppm nitrate forage are poisonous and should not be given to the animals; high nitrate fodders must be given at 2–3 week intervals. The nitrate levels in the forage crops growing on dry and wet soils could be reduced by late harvesting. Animals should not consume high nitrate feed when they are hungry. Qualified researchers should guide the producers about the use of fertilizers in vegetable farming in greenhouses. Testing soils for nitrogen has been a useful practice in the drier regions for many years, and in such regions fertilizer rates can be adjusted to account for NO3− found in the soil prior to planting. The producers should get the scientific analysis of soil and plant samples by the experts in the field and greenhouses for a sustainable production and prevention of soil pollution through irrigation (Guo et al. 2006).
Effective means of minimizing the accumulation of nitrates in vegetables include identification of low nitrate accumulating genotypes, manipulation of fertilizer regimes, exploiting natural diurnal fluctuations in nitrate concentration, and use of nitrification inhibitors.
Rates needed to achieve different yields with different crops vary, nitrate consumption through vegetables can be kept low by harvesting them at the proper time. Some crops should be harvested in the afternoon of a sunny day when nitrate concentration in the leaves is low. It has been found that the leaf nitrate concentration is at its lowest at 1:00 p.m., when the sunlight is maximum (Anjana Umar and Iqbal 2007).
Only nitrous oxide is of concern from the perspective of agroindustry. This gas may account for up to 15 percent of the total global warming potential. Its global warming potential is estimated to be more than 250 times that of carbon dioxide. Moreover, its lifetime residency in the atmosphere is estimated to be about 130 years. Nitrous oxide also leads to ozone depletion. Globally, anthropogenic activity accounts for 64 percent of the total nitrous oxide emissions of which agriculture accounts for 92 percent.
Emissions of ammonia and methane relate primarily to animal production systems. It is this addition of nitrogen to natural ecosystems, which is of environmental concern both in the terms of its implications for biodiversity and the acidification of soil and aquatic systems. For this purpose, correct nutrient application is very important.
Improving NUE in major food crops requires collaboration among farmers, agronomists, soil scientists, agricultural economists, ecologists, and policymakers (Dobermann and Cassman 2004; Galloway et al. 2002). Agriculture can contribute significantly to reducing the global reactive N load by enhancing the indigenous N supply, which, in turn, will require innovative crop and soil management practices.
The expansion and intensification of agricultural production to meet the needs of a burgeoning population have brought vast land areas under cultivation. Continued growth in cultivated areas, however, is very unlikely since in the future there will be more competition for domestic and industrial uses. This means that crop productivity has to be increased, for which fertilizer, particularly N, plays the most crucial role. The global demand for N fertilizer is increasing day by day, driven mainly by cereal production. The world presently uses around 83 Mt of N, which is about a 100‐fold increase over the last 100 years. About 60 % of global N fertilizer consumption has been used to produce the world’s three major cereals, rice, maize, and wheat. The projections are that 50–70 % more cereal grain would be required by 2050 to feed 9.3 billion people. This will require an increased use of N fertilizer of a similar magnitude (50–70 %) with the current state of NUE. Moreover, there will be enormous environmental penalties for the use of such a large amount of inorganic, reactive N. Therefore, to supply sufficient N to meet increased food demand and maintain a safe environment, fertilizer N must be used efficiently and prudently, that is, N use efficiency must be improved. This chapter evaluates NUE for the major cereals grown across large agroclimatic regions and analyzes various strategies available to improve NUE.
Fertilizer N use efficiency is a complex term with many components. To quantify NUE, the term most widely used is a ratio with output (biological yield or economic yield in terms of biomass or N content) as the numerator and input (N supply from soil, fertilizer, or soil plus fertilizer) as the denominator. Based on a large number of studies conducted in research fields from across the globe in a wide diversity of agroecologies, 44–55 % of the applied fertilizer N was recovered in the first crop. When fertilizer N recovery in plant and soil during succeeding seasons was considered, an additional recovery of about 7 % was estimated. A N recovery efficiency of 40 % or more has been obtained with improved N management practices.
To achieve an N use efficiency of near 100 %, similar to that of symbiotic N2 fixing systems such as legume rhizobium, will remain a huge challenge and currently not within reach. In symbiotic N2 fixing systems, the N requirement and supply are highly synchronized and almost no leakage (excluding N leakage through root exudation and NH3 volatilization from leaves) of N occurs. Absolute synchrony similar to that of legumes will not be attainable in fertilizer dependent cereal production systems. Nonetheless, efficiency levels close to 70–90 % obtained in several well managed controlled experiments can be targeted, at least in well managed irrigated cereal systems.
Fertilizer NUE is governed by three major factors: N uptake by the crop, N supply from soil and fertilizer, and N losses from soil–plant systems. The crop N requirement is the most important factor influencing NUE. Much research has been conducted during the past 50 years to improve NUE by trying to develop better fertilizers or improved N management practices, based mainly on a better synchronization between the supply and the uptake of N by the crop. There is significant potential to increase NUE in cereals.
Many of the strategies needed to achieve such increases have already been developed. The use of an integrated crop management strategy comprising optimal soil, water, and crop management under good climatic conditions could attain a large NUE value. In addition, cereal genotypes with a large harvest index must be used to obtain high REN and PEN because the harvest index is tightly linked to NUE. Many approaches have been suggested for increasing NUE, for example, optimal time, rate, and methods of application for matching N supply with crop demand; the use of specially formulated forms of fertilizer, including those with urease and nitrification inhibitors; the integrated use of fertilizer, manures, and = or crop residues; and optimizing irrigation management. In addition, some modern tools such as precision farming technologies, simulation modeling, decision support systems, and resource‐conserving technologies also help to improve NUE.
Importantly, some of these techniques are being adopted on a large scale by many farmers. Two success stories based on crop demand based N management warrant mention. In large scale agriculture practiced in developed countries, precision farming studies have demonstrated that variablerate N fertilizer application has the potential to significantly reduce the N rate required to achieve yields similar to those obtained with Standard uniform management practices. In agriculture with small to medium farm size in developing countries, the use of a simple and inexpensive leaf color chart assists farmers in applying N when the plant needs it. The use of this simple tool, currently practiced by about 350,000 farmers globally, has reduced fertilizer N use by about 25 %.
Because some of the main factors, such as climate controlling crop needs for N, are largely outside farmer control, it will remain difficult to predict the amount of N to apply for optimum growth. Along similar lines, the amount of N that becomes available for crop uptake through net soil N mineralization varies across the landscape and is diYcult to predict. Therefore, because of those uncertainties related to the demand for and supply of N to the crop, it will always remain a daunting task to achieve significantly higher fertilizer N use efficiencies. However, our study indicated that farmers can increase NUE substantially in well managed agricultural systems by adopting appropriate technologies and carefully crafted local policies that contribute to improved N management. In any event, improved farmer knowledge regarding N management techniques is essential. A key challenge is to ensure that such techniques are cost effective and user friendly so that they provide attractive options for adoption by farmers.
Change history
12 October 2017
An erratum has been published.
References
Abdalla M, Jones M, Yeluripati J, Smith P, Burke J, Williams M (2011) Testing DayCent and DNDC model simulations of N2O fluxes and assessing the impacts of climate change on the gas flux and biomass production from a humid pasture. Atmos Environ 44:2961–2970
Abrol YP, Chatterjee SR, Kumar PA, Jain V (1999) Improvement in nitrogen use efficiency: physiological and molecular approaches. Curr Sci 76:1357–1364
Abrol YP, Pandey R, Raghuram N, Ahmad A (2012) Nitrogen cycle sustainability and sustainable technologies for nitrogen fertilizer and energy management. J Indian Inst Sci 92:17–36
Almasri MN, Kaluarachchi JJ (2004) Assessment and management of long-term nitrate pollution of ground water in agriculture-dominated watersheds. J Hydrol 295:225–245
Amiour N, Imbaud S, Clement G et al (2012) The use of metabolomics integrated with transcriptomic and proteomic studies for identifying key steps involved in the control of nitrogen metabolism in crops such as maize. J Exp Bot 63:5017–5033
Anayah FM, Almasri MN (2009) Trends and occurrences of nitrate in the groundwater of the West Bank, Palestine. Appl Geogr 29:588–601
Anjana Umar S, Iqbal M (2007) Nitrate accumulation in plants, factors affecting the process, and human health implications. A review. Agron Sustain Dev 27:45–57
Anjana Umar S, Iqbal M, Abrol YP (2007) Are nitrate concentrations in leafy vegetables within safe limits. Curr Sci 92:355–360
Bahrmam N, Gouis J, Negroni L, Amilhat L, Leroy P, Laine AL, Jamion O (2004) Differential protein expression assessed by two-dimensional gel electrophoresis for two wheat varieties grown at four nitrogen levels. Proteomics 4:709–719
Bahrman N, Gouy A, Devienne-Barret F, Hirel B, Vedele F, Le Gouis J (2005) Differential change in root protein patterns of two wheat varieties under high and low nitrogen nutrition levels. Plant Sci 168:81–87
Bjorne H, Petersson J, Phillipson M, Weitzberg E, Holm L, Lundberg JO (2004) Nitrate in saliva increases gastric mucosal blood flow and mucus thickness. J Clin Invest 113:106–114
Boyer E, Howarth RH, Galloway JN, Dentener FJ, Cleveland C, Asner GP, Green P, Vorosmarty C (2004) Current nitrogen inputs to world regions. In: Mosier AR, Syers JK, Freney JR (eds) Agriculture and the nitrogen cycle: assessing the ımpacts of fertilizer use on food production and the environment. SCOPE 65, Paris, France, pp 221–230
Brasil (2010) Ministério da Ciência e Tecnologia. Coordenação Geral de Mudanças Globais do Clima. Segunda comunicação nacional do Brasil à convenção-quadro das Nações Unidas sobre mudança do clima. Ministério da Ciência e Tecnologia, DF, Brasília
Bruning-Fann CS, Kaneene J (1993) The effects of nitrate, nitrite, and N-nitroso compounds on animal health. Vet Hum Toxicol 35:237–253
Bu Y, Takano T, Nemoto K, Liu S (2011) Research progress of ammonium transporter in rice plants. Genom Appl Biol 2:19–23
Burkart MR, James DE (1999) Agricultural-nitrogen contributions to hypoxia in the Gulf of Mexico. J Environ Qual 28:850–859
Butterbach-Bahl K, Baggs EM, Dannenmann M, Kiese R, Zechmeister-Boltenstern S (2013) Nitrous oxide emissions from soils: how well do we understand the processes and their controls. Philos Trans R Soc Lond B Biol Sci 368:20130122
Cassman KG, Dobermann A, Walters D (2002) Agroecosystems, nitrogen‐use eYciency, and nitrogen management. Ambio 31:132–140
Castaings L, Marchive C, Meyer C, Krapp A (2011) Nitrogen signalling in Arabidopsis: how to obtain insights into a complex signalling network. J Exp Bot 62:1391–2011
Cerri CC, Ferreira Maia SM, Galdos MV, Cerri CEP, Feigl JB, Bernoux M (2009) Brazilian greenhouse gas emissions: the importance of agriculture and livestock. Sci Agr Piracicaba, Braz 66:831–843
Chen BM, Wang ZH, Li SX, Wang GX, Song HX, Wang XN (2004) Effects of nitrate supply on plant growth, nitrate accumulation, metabolic nitrate concentration and nitrate reductase activity in three leafy vegetables. Plant Sci 167:635–643
Chiu HF, Tsai SS (2007) Nitrate in drinking water and risk of death from bladder cancer: an ecological case-control study in Taiwan. J Toxicol Environ Health A 70:1000–1004
Cristina MV, Manolis K, Sylvaine C, Michael RT, Roel V, John RN, Mark JN, Patrick L (2014) Assessing exposure and health consequences of chemicals in drinking water: current state of knowledge and research needs. Environ Health Perspect 122(3):213–221
Dalal RC, Wang W, Robertson GP, Parton WJ (2003) Nitrous oxide emission from Australian agricultural lands and mitigation options: a review. Aust J Soil Res 41:165–195
Daniel JS, Fleming EL, Portmann RW, Velders GJM, Jackman CH, Ravishankara AR (2010) Options to accelerate ozone recovery: ozone and climate benefits. Atmos Chem Phys 10:7697–7707
de Koeijer TJ, Wossink GAA, Smit AB, Janssens SRM, Renkema JA, Struik PC (2003) Assessment of the quality of farmers’ environmental management and its effects on resource use efficiency: a Dutch case study. Agric Sys 78:85–103
Denman KL, Brasseur G, Chidthaisong A, Ciais P, Cox PM, Dickinson RE, Hauglustaine D, Heinze C, Holland E, Jacob D, Lohmann U, Ramachandran S, da Silva Dias PL, Wofsy SC, Zhang X (2007) Couplings between changes in the climate system and biogeochemistry. In: Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M, Miller HL (eds) Climate change 2007: the physical science basis contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, pp 499–588
Di HJ, Cameron KC (2002) Nitrate leaching in temperate agroecosystems: sources, factors and mitigating strategies. Nutr Cycl Agroecosyst 46:237–256
Dobermann A, Cassman KG (2004) Environmental dimensions of fertilizer nitrogen: what can be done to increase nitrogen use eYciency and ensure global food security? In: Mosier AR, Syers JK, Freney JR (eds) Agriculture and the nitrogen cycle: assessing the ımpacts of fertilizer use on food production and the environment. SCOPE 65, Paris, France, pp 261–278
Dordas CA, Sioulas C (2008) Safflower yield, chlorophyll content, photosynthesis, and water use efficiency response to nitrogen fertilization under rainfed conditions. Ind Crop Prod 27:75–85
El-Khatib RT, Hamerlynck EP, Gallardo F, Kirby EG (2004) Transgenic poplar characterized by ectopic expression of a pine cytosolic glutamine synthetase gene exhibits enhanced tolerance to water stress. Tree Physiol 24:729–736
Emongor VE, Ramolemana GM (2004) Treated sewage effluent (water) potential to be used for horticultural production in Botswana. Phys Chem Earth 29:1101–1108
EPA (2007) Hypoxia in the northern gulf of Mexico: an update by the environmental protection agency science advisory board. Environmental Protection Agency, Washington, USA, Report no. EPA-SAB-08-003
Escobar LF et al (2010) Postharvest nitrous oxide emissions from a subtropical oxisol as influenced by summer crop residues and their management. Revista Brasileira de Ciência do Solo 34:507–516
Fageria NK (2009) Mineral nutrition versus yield of field crops. In: Frageria NK (ed) The use of nutrients in crop plants. CRC Press, Bocca Rotan, FL, pp 1–29
FAO (2011) Current world fertilizer trends and outlook to 2015. FAO, Rome, Italy
Fait A, Nesi AN, Angelovici R, Lehmann M, Pham PA, Song L et al (2011) Targeted enhancement of glutamate-to-gamma-aminobutyrate conversion in Arabidopsis seeds affects carbon-nitrogen balance and storage reserves in a development-dependent manner. Plant Physiol 157:1026–1042
Ferrario-Méry S, Valadier MH, Foyer CH (1998) Overexpression of nitrate reductase in tobacco delays drought-induced decreases in nitrate reductase activity and mRNA. Plant Physiol 117:293–302
Ferrario-Méry S, Masclaux C, Suzuki A, Valadier MH, Hirel B, Foyer CH (2001) Glutamine and a-ketoglutarate are metabolite signals involved in nitrate reductase gene transcription in untransformed and transformed tobacco plants deficient in ferredoxin-glutamine-a-ketoglutarate. Planta 213:265–271
Fewtrell L (2004) Drinking-water nitrate, methemoglobinemia, and global burden of disease: a discussion. Environ Health Perspect 112:1371–1374
Filoso S, Martinelli LA, Howarth RW, Boyer EW, Dentener F (2006) Human activities changing the nitrogen cycle in Brazil. Biogeochemistry 79:61–89
Fraisier V, Gojon A, Tillard P, Daniel‐Vedele F (2000) Constitutive expression of a putative high‐affinity nitrate transporter in Nicotiana plumbaginifolia: evidence for post‐transcriptional regulation by a reduced nitrogen source. Plant J 23:489–496
Fernie AR, Stitt M (2012) On the discordance of metabolomics with proteomics and transcriptomics: coping with increasing complexity in logic, chemistry, and network interactions scientific correspondence. Plant Physiol 158:1139–1145
Fuentes SI, Allen DJ, Ortiz‐Lopez A, Hernández G (2001) Over‐expression of cytosolic glutamine synthetase increases photosynthesis and growth at low nitrogen concentrations. J Exp Bot 52:1071–1081
Fuentes-Ramírez LE, Bustillos-Cristales R, Tapia-Hernández A, Jiménez-Salgado T, Wang ET, Martínez-Romero E, Caballero-Mellado J (2001) Novel nitrogen-fixing acetic acid bacteria, Gluconacetobacter johannae sp. nov. and Gluconacetobacter azotocaptans sp. nov., associated with coffee plants. Int J Syst Evol Microbiol 51:1305–1314
Galloway JN, Cowling EB, Seitzinger SP, Socolow RH (2002) Reactive nitrogen: too much of a good thing? Ambio 31:60–71
Glass ADM (2003) Nitrogen use efficiency of crop plants: physiological constraints upon nitrogen absorption. Crit Rev Plant Sci 22:453–470
Godish T, Davis WT, Fu JS (2015) Air quality, 5th edn. CRC Press, Boca Raton, FL
Gomes RL, Scrimshaw MD, Lester JN (2009) Fate of conjugated natural and synthetic steroid estrogens in crude sewage and activated sludge batch studies. Environ Sci Technol 43:3612–3618
Gormly J, Spalding RF (2006) Sources and concentrations of nitrate‐nitrogen in ground water of the central platte region, Nebraskaa. Ground Water 17:291–301
Granstedt A (2000) Increasing the efficiency of plant nutrient recycling within the agricultural system as a way of reducing the load to the environment–experience from Sweden and Finland. Agric Ecosyst Environ 80:169–185
Greer FR, Shannon M (2005) Infant methemoglobinemia: the role of dietary nitrate in food and water. Pediatrics 116:784–786
Guleryuz G, Gucel S, Ozturk M (2010) Nitrogen mineralization in a high altitude ecosystem in the Mediterranean phytogeographical region of Turkey. J Environ Biol 31(4):503–514
Guo M, Li H, Zhang Y, Zhang X, Lu A (2006) Effects of water table and fertilization management on nitrogen loading to groundwater. Agric Water Manag 82:86–98, ISSN 0378–3774
Gupta ML, Khosla R (2012) Precision nitrogen management and global nitrogen use efficiency. In: Harald K, Butron GMP (eds) Proceedings of the 11th international conference on precision agriculture. Indianapolis, USA
Gupta SK, Gupta RC, Chhabra SK, Eskiocak S, Gupta AB, Gupta R (2008) Health issues related to N pollution in water and air. Indian Agric Environ Health 94:1469–1477
Gupta S, Gupta RC, Gupta AB (2007) Nitrate toxicity and human health. In: Abrol YP, Raghuram N, Sachdev MS (eds) Agricultural nitrogen uses and its environmental implications. I.K. International Publication House Pvt Ltd, New Delhi, India, pp 517–547
Hakeem KR, Ahmad A, Iqbal M, Gucel S, Ozturk M (2011) Nitrogen-efficient cultivars can reduce nitrate pollution. Environ Sci Pollut Res 18:1184–1193
Hakeem KR, Chandna R, Ahmad A, Iqbal M (2012a) Reactive nitrogen inflows use efficiency in agriculture: an environment perspective. In: Ahmad A, Prasad MNV (eds) Environment adaptation and stress tolerance of plants in the era of climate change. Springer, New York, pp 217–232
Hakeem KR, Chandna R, Ahmad A, Iqbal M (2012b) Physiological and molecular analysis of applied nitrogen in rice (Oryza sativa L.) genotypes. Rice Sci 19(3):213–222
Hakeem KR, Chandna R, Ahmad A, Iqbal M (2012c) Physiological studies and proteomic analysis for differentially expressed proteins and their possible role in the root of N-efficient rice (Oryza sativa L.). Mol Breed 32(4):785–798
Hakeem KR, Sabir M, Khan F, Rehman RU (2014) Nitrogen regulation and signalling in plants. In: Hakeem KR, Rehman RU, Tahir I (eds) Plant signaling: understanding the molecular crosstalk. Springer, India, pp 117–132
Hänsch R, Fessel DG, Witt C, Hesberg C, Hoffmann G, Walch‐Liu P, Engels C, Kruse J, Rennenberg H, Kaiser WM, Mendel RR (2001) Tobacco plants that lack expression of functional nitrate reductase in roots show changes in growth rates and metabolite accumulation. J Exp Bot 52:1251–1258
Hatfield JL, Sauer TJ, Prueger JH (2001) Managing soils to achieve greater water use efficiency. Agron J 93:271–280
HELCOM (2015) Updated Fifth Baltic Sea pollution load compilation (PLC-5.5). Baltic Sea Environment proceedings no. 145, pp 143
Hijleh RJA (2014) Chemical and microbial risk management assessment of drinking water in faria catchment. M.Sc thesis, An-Najah National University, Nablus, Palestine
Hill MJ (1999) Nitrate toxicity: myth or reality. Brit J Nutr 81:343–344
Hirel B, Le Gouis J, Ney B, Gallais A (2007) The challenge of improving nitrogen use efficiency in crop plants: towards a more central role for genetic variability and quantitative genetics within integrated approaches. J Exp Bot 58:2369–2387
Hoshida H, Tanaka Y, Hibino T, Hayashi Y, Tanaka A, Takabe T, Takabe T (2000) Enhanced tolerance to salt stress in transgenic rice that overexpresses chloroplast glutamine synthetase. Plant Mol Biol 43:103–111
Ikehata K, Murphy RR, Liu Y, Sun RN, Nessl MB (2010) Health effects associated with wastewater treatment, reuse, and disposal. Water Environ Res 82:2047–2066
Inoue-Choi M, Ward MH, Cerhan JR, Weyer PJ, Anderson KE, Robin K (2012) Interaction of nitrate and folate on the risk of breast cancer among postmenopausal women. Nutr Cancer 64:685–694
Jiao P, Xu D, Wang S, Wang Y, Liu K, Tang G (2012) Nitrogen loss by surface runoff from different cropping systems. Soil Res 50:58–66
Jim FC, Shelie AM, James RF (2011) Using DAYCENT to quantify on-farm GHG emissions and N dynamics of land use conversion to N-managed switchgrass in the Southern U.S. Agri Ecosys Environ 141(3-4):332–341
Ju XT, Kou CL, Zhang FS, Christie P (2005) Nitrogen balance and groundwater nitrate contamination: comparison among three intensive cropping systems on the north China plain. Environ Pollut 143:117–125
Kapoor A, Viraraghavan T (1997) Nitrate removal from drinking water-review. J Environ Eng 123:371–380
Keeney DR, Hatfield JL (2008) The nitrogen cycle historical perspective and current and potential future concerns. In: Hatfield JL, Follett RF (eds) Nitrogen in the environment: sources problems and management. Academic, Elsevier, pp 1–18
Keeney D, Olson RA (1986) Sources of nitrate to ground water. Crit Rev Environ Sci Technol 16:257–304
Khandare HW (2013) Scenario of nitrate contamination in groundwater: its causes and prevention. Int J ChemTech Res 5:1921–1926
Killebrew K, Wolff H (2010) Environmental impacts of agricultural technologies. Agricultural Policy and Statistics Team of the Bill and Melinda Gates Foundation, EPAR brief no. 65, pp 1–18
Kissel D, Cabrera M, Vaio N, Craig J, Rema J, Morris L (2004) Rainfall timing and ammonia loss from urea in a loblolly pine plantation. Soil Sci Soc Am J 68:1744–1750
Kozaki A, Takeba G (1996) Photorespiration protects C3 plants from photooxidation. Nature 384:557–560
Kumar S, Tamura K, Jakobsen IB, Nei M (2001) MEGA2: molecular evolutionary genetics analysis software. Bioinformatics 17:1244–1245
Ladha JK, Pathak H, Krupnik TJ, Six J, van Kessel C (2005) Efficiency of fertilizer nitrogen in cereal production: retrospects and prospects. Adv Agron 87:85–156
Li C, Zhuang Y, Cao M, Crill P, Dai Z, Frolking S, Moore B, Salas W, Song W, Wang X (2001) Comparing a process-based agro ecosystem model to the IPCC methodology for developing a national inventory of N2O emissions from arable lands in China. Nut Cycl Agroecosys 60:1–3
Lillo C, Meyer C, Lea US, Provan F, Oltedal S (2004) Mechanism and importance of post‐translational regulation of nitrate reductase. J Exp Bot 55:1275–1282
Liu KH, Tsay YF (2003) Switching between the two action modes of the dual-affinity nitrate transporter CHL1 by phosphorylation. EMBO J 22:1005–1013
Liu Z, Song X, Jiang L, Lin H, Xu Y, Gao X, Zheng F, Tan D, Wang M, Jing S, Shen Y (2012) Strategies for managing soil nitrogen to prevent nitrate-N leaching in intensive agriculture system. In: Hernandez Soriano MC (ed) Soil health and land use management. InTech, pp 133–154. http://www.intechopen.com/books/soil-health-and-land-usemanagement/strategies-for-managing-soil-nitrogen-to-prevent-nitrate-n-leaching-in-intensive-agriculturesystem. Accessed 18 Apr 2015
Loqué D, von Wirén N (2004) Regulatory levels for the transport of ammonium in plant roots. J Exp Bot 55:1293–1305
Man HM, Boriel R, El‐Khatib R, Kirby EG (2005) Characterization of transgenic poplar with ectopic expression of pine cytosolic glutamine synthetase under conditions of varying nitrogen availability. New Phytol 167:31–39
Manassaram DM, Backer LC, Moll DM (2007) A review of nitrates in drinking water: maternal exposure and adverse reproductive and developmental outcomes. Environ Health Prospect 114:320–327
Masclaux C, Quillere I, Gallais A, Hirel B (2001) The challenge of remobilisation in plant nitrogen economy. A survey of physio‐agronomic and molecular approaches. Ann Appl Biol 138:69–81
McPharlin IR, Aylam PM (1995) Nitrogen requirements of lettuce under sprinkler irrigation and fertilization on spearwood sand. J Plant Nutr 18:219–241
McDowell LR (2012) Vitamins in animal nutrition: comparative aspects to human nutrition. Academic, London
Mikkelsen M, Hartz TK (2008) Nitrogen sources for organic crop production. Better Crop 92:16–19
Millar N, Robertson GP, Grace PR, Gehl RJ, Hoben JP (2010) Nitrogen fertilizer management for nitrous oxide (N2O) mitigation in intensive corn (Maize) production: an emissions reduction protocol for US Midwest agriculture. Mitig Adapt Strat Glob Change 15:185–204
Mosier AR, Zhaoliang Z (2000) Changes in patterns of fertilizer nitrogen use in Asia and its consequences for N2O emissions from agricultural systems. Nutr Cycl Agroecosyst 57:107–117
Mosier AR, Bleken M, Chaiwanakupt P, Ellis EC, Freney JR, Howarth RB, Matson PA, Minami K, Naylor R, Weeks KN (2001) Policy implications of human-accelerated nitrogen cycling. Biogeochemistry 52:281–320
Mosier AR, Syers JK, Freney JR (2004a) Agriculture and the nitrogen cycle: assessing the impacts of fertilizer use on food production and the environment, 2nd edn. Island Press, Washington, DC
Mosier AR, Syers JK, Freney JR (2004b) Nitrogen fertilizer: an essential component of increased food, feed, and fiber production. In: Mosier AR, Syers JK, Freney JR (eds) Agriculture and the nitrogen cycle: assessing the ımpacts of fertilizer use on food production and the environment. SCOPE 65, Paris, France, pp 3–15
Mueller BA, Newton K, Holly EA, Preston-Martin S (2001) Residential water source and the risk of childhood brain tumors. Environ Health Perspect 109:551–556
NAAS (2005) Policy options for efficient nitrogen use, policy paper no. 33. National Academy of Agricultural Sciences, New Delhi, pp 1–4
Norse D (2003) Fertilizers and world food demand implications for environmental stresses. Paper presented at the IFA-FAO agriculture conference on global food security and the role of sustainable fertilization, FAO, Rome, Italy, 26–28 Mar 2003
Nolan BT, Stoner JD (2000) Nutrients in ground waters of the conterminous United States, 1992–1995. Environ Sci Technol 34:1156–1165
Odiyo JO, Makungo R, Muhlarhi TG (2014) The impacts of geochemistry and agricultural activities on groundwater quality in the Soutpansberg fractured aquifers. In: Rebbia CA (ed) Water pollution XII. WIT Press, Ashurst, Southampton, UK, pp 121–132
Ohyama T (2010) Nitrogen as a major essential element of plants. In: Ohyama T, Sueyoshi K (eds) Nitrogen assimilation in plants. Signpost, Trivandrum, Kerala, India
Oliveira ALM, Urquiaga S, Döbereiner J, Baldani JI (2002) The effect of inoculating endophytic N2-fixing bacteria on micropropagated sugarcane plants. Plant and Soil 24:205–215
Ondersteijn C, Beldman A, Daatselaar C, Giesen G, Huirne R (2002) The Dutch mineral accounting system and the European nitrate directive: implications for N and P management and farm performance. Agric Ecosyst Environ 92:283–296
Ortega JL, Temple SJ, Sengupta-Gopalan C (2001) Constitutive overexpression of cytosolic glutamine synthetase (GS1) gene in transgenic alfalfa demonstrates that GS1 may be regulated at the level of RNA stability and protein turnover. Plant Physiol 126:109–121
Ozturk M, Gucel S, Sakcali S, Baslar S (2013) Nitrate and edible plants in the Mediterranean Region of Turkey: an overview. In: Umar S et al (eds) Nitrate in leafy vegetables—toxicity and safety measures. I.K. Intern. Publ. House Pvt. Ltd, New Delhi-Bangalore (India), pp 17–51
Paerl HW, Dennis RL, Whitall DR (2002) Atmospheric deposition of nitrogen: implications for nutrient over-enrichment of coastal waters. Estuaries 25:677–693
Palm CA, Machado PLOA, Mahmood T, Melillo J, Murrell ST, Nyamangara J, Scholes M, Sisworo E, Olesen JE, Pender J, Stewart J, Galloway JL (2004) Societal responses for addressing nitrogen fertilizer needs: balancing food production and environmental concerns. In: Mosier AR, Syers JK, Freney JR (eds) Agriculture and the nitrogen cycle assessing the ımpacts of fertilizer use on food production and the environment. SCOPE 65, Paris, France, pp 71–89
Paramasivam S, Alva AK, Fares A, Sajwan KS (2002) Fate of nitrate and bromide in an unsaturated zone of a sandy soil under citrus production. J Environ Qual 31:671–681
Pathak H, Nedwell DB (2001) Strategies to reduce nitrous oxide emission from soil with fertilizer selection and nitrification inhibitor. Water Air Soil Poll 129:217–228
Pionke HB, Pionke ML, Sharma KJ (1990) Hirschberg Impact of irrigated horticulture on nitrate concentrations in groundwater. Agr Ecosyst Environ 32:119–132
Portmann RW, Daniel JS, Ravishankara AR (2012) Stratospheric ozone depletion due to nitrous oxide: influences of other gases. Philos Trans R Soc B Sci 367:1256–1264
Prasad R (2013) Fertilizer nitrogen, food security, health and the environment. Proc Indian Nat Sci Acad 79:997–1010
Provan F, Aksland LM, Meyer C, Lillo C (2000) Deletion of the nitrate reductase N-terminal domain still allows binding of 14-3-3 proteins but affects their inhibitory properties. Plant Physiol 123:757–764
Quilleré I, Dufossé C, Roux Y, Foyer CHF, Caboche M, Morot-Gaudry JF (1994) The effects of deregulation of NR gene expression on growth and nitrogen metabolism of Nicotiana plumbaginifolia plants. J Exp Bot 45:1205–1211
Ramaraju HK, Venkatachalappa M, Ranganna G, Sadashivaiah C, Manamohan Rao N (1999) Hazards due to migration of septic tank leakages in peri-urban settlements. In: Ellis JB (ed) Impact of urban growth on surface water and groundwater quality. International Association of Hydrogeological Science, Publication no 259, pp 219–225
Ravishankara AR, Daniel JS, Portmann RW (2009) Nitrous oxide (N2O): the dominant ozone-depleting substance emitted in the 21st century. Science 326:123–125
Ribbe L, Delgado P, Salgado E, Flüge WA (2008) Nitrate pollution of surface water induced by agricultural non-point pollution in the Pocochay watershed, Chile. Desalination 226:13–20
Robertson GP, Vitousek PM (2009) Nitrogen in agriculture: balancing an essential resource. Annu Rev Energy Environ 34:97–125
Rosegrant MW, Cai X, Cline SA (2002) World water and food to 2025: dealing with scarcity. International Food Policy Research Institute (IFPRI), Washington, DC
Santiago-Antonio G, Lizama-Gasca MG, Carrillo-Pech M, Echevarría-Machado I (2014) Natural variation in response to nitrate starvation among varieties of habanero pepper (Capsicum chinense Jacq.). Aust J Crop Sci 8:523–535
Schilling KE, Libra RD (2000) The relationship of nitrate concentrations in streams to row crop land use in Iowa. J Environ Qual 29:1846–1851
Schoenbeck MA, Temple SJ, Trepp GB, Blumenthal JM, Samac DA, Gantt JS, Hernandez G, Vance CP (2000) Decreased NADH glutamate synthase activity in nodules and flowers of alfalfa (Medicago sativa L.) transformed with an antisense glutamate synthase transgene. J Exp Bot 51:29–39
Schumann U, Huntrieser H (2007) The global lightning-induced nitrogen oxides source. Atmos Chem Phys 7:2623–2818
Shabalala AN, Combrinck L, McCrindle R (2013) Effect of farming activities on seasonal variation of water quality of Bonsma Dam, KwaZulu-Natal. South Afr J Sci 109, Art. #0052, pp 7. doi:10.1590/sajs.2013/20120052
Sharma MK, Sharma H, Bapna N (2013) Toxic effects of high nitrate intake in oesophagus and stomach of rabbits. Int J Med Res Health Sci 2:407–411
Shen Y, Lei H, Yang D, Kanae S (2011) Effects of agricultural activities on nitrate contamination of groundwater in a Yellow river irrigated region. Int Assoc Hydrogeol Sci 348:73–80
Shoji S, Delgado J, Mosier A, Miura Y (2001) Use of controlled release fertilizers and nitrification inhibitors to increase nitrogen use efficiency and to conserve air and water quality. Commun Soil Sci Plant Anal 32:1051–1070
Signor D, Cerri CEP (2013) Nitrous oxide emissions in agricultural soils: a review. Pesq Agropecu Trop 43:322–338
Simons M, Saha R, Guillard L, Clément G, Armengaud P, Cañas R, Maranas CD, Lea PJ, Hirel B (2014) Nitrogen-use efficiency in maize (Zea mays L.): from ‘omics’ studies to metabolic modelling. J Exp Bot 65:5657–5671
Smil V (2002) Nitrogen and food production: proteins for human diets. Ambio 31:126–131
Solomon S (1999) Stratospheric ozone depletion: a review of concepts and history. Rev Geophys 37:275–316
Soussana JF, Lemairec G (2014) Coupling carbon and nitrogen cycles for environmentally sustainable intensification of grasslands and crop-livestock systems. Agric Ecosyst Environ 190:9–17
Sowers KE, Pan WL, Miller BC, Smith JL (1994) Nitrogen use efficiency of split nitrogen applications in soft white winter wheat. Agron J 86:942–948
Stitt M (1999) Nitrate regulation of metabolism and growth. Curr Opin Plant Biol 2:178–186
Subbarao GV, Sahrawat KL, Nakahara K, Rao IM, Ishitani M, Hash CT, Kishii M, Rasouli S, Whalen JK, Madramootoo CA (2013) Review: reducing residual soil nitrogen losses from agroecosystems for surface water protection in Quebec and Ontario, Canada: best management practices, policies and perspectives. Can J Soil Sci 94:109–127
Takahashi M, Sasaki Y, Ida S, Morikawa H (2001) Nitrite reductase gene enrichment improves assimilation of NO2 in Arabidopsis. Plant Physiol 126:731–741
Thorburn PJ, Biggs JS, Weier KL, Keating BA (2003) Nitrate in groundwaters of intensive agricultural areas in coastal Northeastern Australia. Agric Ecosyst Environ 94:49–58
Tilman D, Balzer C, Hill J, Befort BL (2011) Global food demand and the sustainable intensification of agriculture. Proc Natl Acad Sci U S A 108:20260–20264
Tilman D, Cassman KG, Matson PA, Naylor RL, Polasky S (2002) Agricultural sustainability and intensive production practices. Nature 418:671–677
Toma Y, Hatano R (2007) Effect of crop residue C: N ratio on N2O emissions from Gray Lowland soil in Mikasa, Hokkaido, Japan. Soil Sci Plant Nutr 53:198–205
Ussiri D, Lal R (2013) Soil emission of nitrous oxide and its mitigation. Springer, New York
van Cleemput O, Boeckx P (2005) Alteration of nitrogen cycling by agricultural activities, and its environmental and health consequences. Gayana Bot 62:98–109
Verma S, Nizam S, Verma PK (2013) Biotic and abiotic stress signalling in plants. In: Ahmad A, Sarwat M, Abdin MZ (eds) Stress signalling in plants: genomics and proteomics perspective, vol 1. Springer Science, New York, pp 25–49
Vincent R, Fraisier V, Chaillou S, Limami MA, Deleens E, Phillipson B, Douat C, Boutin JP, Hirel B (1997) Overexpression of a soybean gene encoding cytosolic glutamine synthetase in shoots of transgenic Lotus corniculatus L. plants triggers changes in ammonium assimilation and plant development. Planta 201:424–433
von Wirén N, Lauter FR, Ninnemann O, Gillissen B, Walch‐Liu P, Engels C, Jost W, Frommer WB (2000) Differential regulation of three functional ammonium transporter genes by nitrogen in root hairs and by light in leaves of tomato. Plant J 21:167–175
Vymazal J (2007) Removal of nutrients in various types of constructed wetlands. Sci Total Environ 380:48–65
Ward MH (2009) Too much of a good thing. Nitrate from nitrogen fertilizers and cancer. Rev Environ Health 24:357–363
WHO (2011) Nitrate and nitrite in drinking-water. World Health Organization, Avenue Appia, Geneva, Switzerland
Walley F, Pennock D, Solohub M, Hnatowich G (2001) Spring wheat (Triticum aestivum) yield and grain yield protein responses to N fertilizer in topographically defined landscape positions. Can J Soil Sci 81:505–514
Walley F, Yates T, Van Groenigen JW, Van Kessel C (2002) Relationships between soil nitrogen availability indices, yield, and nitrogen accumulation of wheat. Soil Sci Soc Am J 66:1549–1561
Wood S, Henao J, Rosegrant M (2004) The role of nitrogen in sustaining food production and estimating future nitrogen fertilizer needs to meet food demand. In: Mosier AR, Syers JK, Freney JR (eds) Agriculture and the nitrogen cycle: assessing the ımpacts of fertilizer use on food production and the environment. SCOPE 65, Paris, France, pp 245–260
Xu XK, Wang YS, Zheng XH, Wang MX, Wang ZJ, Zhou LK, Van Cleemput O (2000) Methane emission from a simulated rice field ecosystem as influenced by hydroquinone and dicyandiamide. Sci Total Environ 263:243–253
Yamaya T, Obara M, Nakajima H, Sasaki S, Hayakawa T, Sato T (2002) Genetic manipulation and quantitative‐trait loci mapping for nitrogen recycling in rice. J Exp Bot 53:917–925
Yanagisawa S (2004) Dof domain proteins: plant-specific transcription factors associated with diverse phenomena unique to plants. Plant Cell Physiol 45:386–391
Yang CY, Cheng MF, Tsai SS, Hsieh YL (1998) Calcium, magnesium, and nitrate in drinking water and gastric cancer mortality. Japan J Cancer Res 89:124–130
Zhang W, Tian Z, Zhang N, Li X (1996) Nitrate pollution of groundwater in northern China. Agric Ecosyst Environ 59:223–231
Zheng X, Fu C, Xu X, Huang Y, Han S, Hu F, Chen G (2002) The Asian nitrogen cycle case study. Ambio 31:79–87
Zu X, Boeckx P, Wang Y, Huang Y, Zheng X, Hu F, van Cleemput O (2002) Nitrous oxide and methane emissions during rice growth and through rice plants: effect of dicyandiamide and hydroquinone. Biol Fertil Soils 36:53–58
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Hakeem, K.R., Sabir, M., Ozturk, M., Akhtar, M.S., Ibrahim, F.H. (2016). Nitrate and Nitrogen Oxides: Sources, Health Effects and Their Remediation. In: de Voogt, P. (eds) Reviews of Environmental Contamination and Toxicology Volume 242. Reviews of Environmental Contamination and Toxicology, vol 242. Springer, Cham. https://doi.org/10.1007/398_2016_11
Download citation
DOI: https://doi.org/10.1007/398_2016_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-51242-6
Online ISBN: 978-3-319-51243-3
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)