
Abstract
The application of biotechnology in cancer therapy is widespread, particularly when combined with traditional medicines. Immunotherapy the fifth pillar of cancer management is highly benefited with advancements in biotechnology over other methods such as surgery, radiation, chemotherapy, and targeted therapy. Engineered cytokines, designer vaccines, cell therapy, and gene therapy fall in immunology-based biotechnology approaches for cancer cure. These advancements along with combination therapy can be a potential remedy not only for cancer treatment but also for the current challenges of drug resistance, disease recurrence, and post treatment hazard effects.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
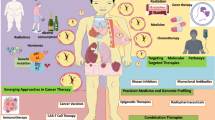
Over twenty million individuals globally were affected by cancer in 2020, and more than nine million people have died due to this malignancy(https://gco.iarc.fr/). The polyclonal growth tumor cells followed by stroma modification with the assistance of immune cells aid cancer progression (Fig. 1). Although current therapies including radiation treatment, and hormonal treatment are thought to be successful, their effectiveness is severely hampered by secondary resistance and hazard effects. In turn, this heightens the necessity for an alternate strategy in addition to traditional medications to treat the patients (Qiao et al. 2016). Recombinant DNA technology, an early kind of genetic engineering where scientists integrated genome data from many species in unique ways to aid patients in the treatment or management of specific ailments, gave rise to the older beginnings of biotechnology in the 1970s (Khan et al. 2016). Biotechnology approaches have been started to combine with conventional treatment options including chemotherapy and immunotherapy. Combining biotechnological approaches with traditional treatments improve patients' overall survival. The invention of inhibitors of the programmed cell death protein 1/programmed cell death protein ligand 1 (PD1/PDL1) and cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) for cancer therapy was recognized with Nobel Prize (Tasuku Honjo and James P. Allison) (Sato et al. 2020). Biotechnology associated immunological approaches are highly competitive with other conventional therapies considering clinical outcome. For instance, over 20 recombinant products of available cytokines can not only function as important immune response signaling transmitters but also as powerful immunotherapy candidates. The efficient and safe injection of cytokines in therapy, however, faces a range of difficulties, from reduced half-life to post-hazard effects, and pleiotropic signaling to aggressive immune functions. Chimeric antigen receptor (CAR)-T cell therapy possess the capacity to develop into immunotherapeutic agents for the treatment of hematological disorders, which present emergency health demands. To overcome challenges like hazard effects corresponding to CAR-T, CAR-T/NK cell therapies have recently gained attention as innovative treatment interventions. Although vaccines possess the potential to benefit patients who are unresponsive to existing standard-of-care immunotherapies it has yet to be considered as an oncologic treatment.

a Polyclonal growth of cancer b immune cells and cytokines involved in immune evasion
In this chapter, current cancer epidemiology and the significance of biotechnology integrated immunotherapy studies using the data curated from The Cancer Genome Atlas (TCGA)-cBioportal is discussed. The clinical advancements of CAR T cell therapy, monoclonal antibody (mAb), stem cell therapy, engineered cytokines, and various types of designer vaccines are also discussed together with the necessity of biotechnology advancements to integrate with combinatorial approaches over other conventional therapies.
2 Current Cancer Epidemiology
With over 8.8 million fatalities globally and 14 million new occurrences of cancer identified every year, cancer is among the most common diseases with high mortality. Monitoring epidemiologic data is essential because it offers crucial details on the cancer's current status statistical, biological, and geographical viewpoints, enabling the creation of suitable medical interventions (Montagnana and Lippi 2017). The most and least common cancers based on latest update of Global Cancer Observatory (GLOBOCAN)-2020 along with their percent increase or decrease comparing GLOBOCAN-2018 is as follows: prostate (9.7%), lung (4.6%) and colorectal cancer (3.7%) are the common cancer in male while Kaposi sarcoma (−3.4%), mesothelioma (−0.4%) and salivary gland tumors (1.4%) are the least. Females possess higher incidence of breast cancer (7.6%), lung cancer (5.8%) and colorectal cancer (4.8%), while it is lowest in Kaposi sarcoma (−24%) (https://gco.iarc.fr/).
A rise in cancer mortality rate even overtaking ischemic heart disease is predicted by WHO 2016–2060 projection data, and the prediction matches with GLOBOCAN 2020 update (https://gco.iarc.fr/). The immunogenomic studies reported in cBioportal based on their potential biotechnology approaches used was curated. Studies entitled Glioblastoma (Columbia: 42 samples), Metastatic melanoma (DFCI: 110 samples, MSKCC: 64 samples, UCLA: 38 samples), non-small cell lung cancer (NSCLC) (MSK: 75 and 16 samples), TMB and immunotherapy (MSKCC: 1661 samples), and Clear cell renal cell carcinoma (DFCI: 35 samples) were selected. Somatic mutations and copy number variations contributed to greater than 95% of genomic profiles in these studies. The KM plot for overall survival after pooling all the samples have shown a significant fall within a year after availing various cancer therapies. The five-year overall survival rate has shown to be less than 20%, which indicate the need for improved therapeutic approaches in cancer cure.
TCGA-cBioportal based 213 non-redundant studies were queried for PDCD1, CD274, and CTLA4 gene (Table 1), which are greatly exploited in immunology-based biotechnology approaches including immune checkpoint inhibitors (ICIs), mAb, and vaccines (Fig. 2), and their mutual exclusivity were analyzed (Table 1). PDCD1 and CTLA4 genes were shown Log2 Odds ratio of greater than three (p<0.001), which indicate their combined roles in cancer progression.

TCGA cBioportal based data representing mutational burden of PDCD1, CD274, and CTLA4. Data source https://www.cbioportal.org/
3 Biotechnology in Cancer Therapy
3.1 Monoclonal Antibody
Although immune checkpoint inhibitors (ICIs) are thought of a type of mAb, they differ in their mode of action. mAbs can locate, bind, and interfere with neo-antigens present on cancer cells (Buchbinder and Desai 2016). CD38, CCR4, PDGFR, Nectin4, TROP2, CD3, CD20, CD79B, HER2, GD2 and SLAMF7 are few of neo-antigens mAbs target in malignant cells (Zahavi and Weiner 2020). They either hijack growth factor assisted signaling via receptor ligand complex inhibition, subsequently inhibiting tumor progression (Li et al. 2005). Cetuximab, an epidermal growth factor receptor (EGFR) mAb, inhibit receptor dimerization and ligand binding followed by neoplastic cell growth inhibition (Patel et al. 2009). Trastuzumab is the pioneer drug approved by Food and Drug Administration (FDA) against Erbb2 receptor tyrosine kinase (HER)-2 which in HER2 positive breast cancer inhibit receptor internalization (Wang and Xu n.d.; Chen et al. 2003). B cells corresponding to Non-lymphoma Hodgkin possess high level of CD20 expression on their surface, in contrast to normal, embryonic B cells. Consequently, a mAb therapy that specifically aim CD20 may kill the malignant cells while leaving behind embryonic B cells to restore the body's pool of normal tissues. As a result, CD20 was chosen as the primary target for mAb treatment, and rituximab (an anti-CD20 mAb) was the first mAb to get approval for cancer therapy (Maloney et al. 1997). mAbs other than ICIs such as elotuzumab, mogamulizumab, ramucirumab, pertuzumab, enfortumab vedotin, sacituzumab, and govitecan are used to treat bladder cancer, multiple myeloma, sarcoma, gastric cancer, breast cancer, cutaneous T cell lymphoma, and triple negative cancer respectively (Anand 2019).
To determine the impact of mAbs in various cancers, multiple clinical trials are now being conducted. A phase II study (NCT04895137) combining mFOLFOX6 with PD1 and bevacizumab is being conducted to analyze tolerability and effectiveness of mAb conjunctions in people tested positive for colorectal cancer. The effectiveness of rituximab with ixazomib in treating mantle cell lymphoma has evaluated in a phase II study (NCT04047797). Rituximab possesses the capacity impair the competence of tumor cells and to alter the immune cells, therefore slowing the spread of the disease. The results indicates, patients with mantle cell lymphoma may benefit from using rituximab with ixazomib in addition to rituximab monotherapy (Vose et al. 2012).
3.2 Stem Cell Therapy
A promising approach in cancer therapeutics is stem cell treatment, which includes all conventional techniques using stem cells. Due to its greater focus on tumors and resulting reduction in target events, it can enhance the clinical effectiveness of other medicines. There are now several stem cells assisted cancer therapy methods being researched in preclinical studies, and they provide both enormous opportunities and concerns (Gomes et al.2017) (Table 2). A revolution in molecular genetics occurred in 2006 with the discovery of Yamanaka factors, which allowed transition of somatic cells to become pluripotent stem cells (iPSCs) (Takahashi and Yamanaka 2006). These iPSCs exhibit the same traits as embryonic stem cells (ESCs) while avoiding the moral dilemma of embryo killing. As of present, the creation of cancer vaccinations (Kooreman et al. 2018; Ouyang et al. 2019) and effector NK and T cells’ activation rely on iPSCs and hESCs as significant sources of material. Many kinds of specialized cells for the tissue and organ may be produced by adult stem cells (ASCs). For cancer therapy, mesenchymal stem cells (MSCs), neural stem cells (NSCs), and hematopoietic stem cells (HSCs). Notch, hedgehog, PI3K/PTEN, NF-kB, Wnt/β-catenin, and JAK/STAT based regulation of typical stem cell growth is well known. Cancer cells and CSCs will develop as a result of the ongoing alteration in the signaling pathways, and crosstalks (Matsui 2016). CSCs possess a strong ability to differentiate and self-renew, which helps with tumor development, invasion and recurrence (Jordan et al. 2006; O’Brien et al. 2010). Additionally, such cells cause malignancies to resist standard treatment (Cojoc et al. 2015; Chang 2016; Batlle and Clevers 2017). To create an effective medicine for the cure of cancer, investigation into CSCs is crucial. Leukemia, gastrointestinal, lung, brain, and breast malignancies are just a few of the tumor types where CSCs have been shown to occur. These cells are routinely separated and recognized utilizing a number of techniques, such as surface protein markers, signaling components, transcriptional candidates, and metabolic or functional characteristics (Codd et al. 2018; Toledo-Guzmán et al. 2018). Key traits and biochemical mechanisms of CSCs have some resemblance to regular stem cells, from whence they originated. Many surface markers, including CD133 HSC marker (Prominin-1), leucine-rich repeat-containing G-protein-coupled receptor (LGR)-5, CD24, CD44, epithelial cell adhesion molecule (EpCAM) are explored in distinguishing CSCs from extremely diverse cell types in malignancies, despite the fact that they may also be found in normal cells (Codd et al. 2018).
HSC transfusion has largely utilized as a primary therapy for leukemia, multiple myeloma, and lymphomas following multiple cycles of radiation or chemotherapy. Additionally, this method is now thoroughly explored in clinical trials of brain tumors (NCT00528437), breast cancer (NCT01807468), neuroblastoma, and sarcomas. Employing heterologous sources of HSCs, graft-versus-host disease (GVHD), which is frequently treated with immunoinhibitory medications with substantial adverse effects and reduced efficacy, is still a problem (Copelan 2006).
In individuals with persistent GVHD, MSCs with immune-educating properties may successfully lessen pronounced immunological reactions. Upon the injection of MSCs and HSCs together, clinical studies reported positive results without any associated negative effects. Mesenchymal-angioblast derived MSC transfusion is being evaluated for tolerability, durability, and effectiveness in people with steroid-resistant GVHD in active multi-center experiment (NCT02923375). MSCs can speed up the healing of damaged organs and may help the body tolerate high dosage chemotherapy, which will have a greater impact on tumor-destroying outcomes (Lee et al. 2011).
Stem cells as possible therapeutic carriers can enhance shelf life of enclosed cargo by protecting from degradation, enhance target specificity and reduce systemic hazard effects. Nanoparticle carrying stem cells being a long running candidate in the field of cancer therapeutics, their lack of proper targeting potential, uncontrollable cellular uptake, and fall in rapid secretion from body is still challenging (Rosenblum et al. 2018). Nanoparticles may be internally absorbed passively or actively by endocytosis, depending on their surface properties, size, processing period, and quantity (Behzadi et al. 2017). The key issues are medication dosage management and probable cell carrier hazard effects. Furthermore, the cells' quick nanoparticle exocytosis might result in the uncontrolled secretion of therapeutic medications into places that aren't intended for them. Despite affecting cell survival or functioning, Roger et al., demonstrated efficient MSC based internalization of PLA and lipid nanoparticles (Roger et al. 2010). Following their immediate tumoral infusion, MSCs transported these nanoparticles into brain tumors in the glioma mouse model. In a different investigation, intravenous injection of MSCs packed with paclitaxel-laden nanoparticles (PTX-NPs) produced drug deposits and increased nanoparticle localization in mice that established orthotopic lung tumors (Layek et al. 2018). Albeit the overall dosages of PTX-NPs used in these nanoparticle encapsulated MSCs were substantially lower than those used in PTX solution/PTX-NPs alone, it is worth noting that they greatly reduced tumor development and improved mouse longevity. MSCs confined in the lung parenchyma but later moved to tumor sites because of their cancer-tropic effect (Lee et al. 2009; Wang et al. 2019a, b). The method that promotes cellular nanoparticle absorption may enhance clinical benefits. Trans activator of transcription (TAT), for instance, can increase the uptake of poly (lactic-co-glycolic acid) (PLGA) NPs into MSCs (Moku et al. 2019).
Immune cells are capable to quickly identifying and eliminating bare oncolytic viruses from the body. Interestingly, to preserve and transport oncolytic viruses to tumor locations, stem cells may be used as a potential carrier. For instance, hNSC line transfected with CRAd-Survivin-pk7 oncolytic virus, in conjunction using temozolomide and ionizing radiation, might boost cytotoxicity in vitro in glioma cells and prolong the lifespan of mice suffering glioblastoma multiforme (Tobias et al. 2013). Additionally, it was shown that MSCs could successfully carry modified oncolytic herpes simplex virus (HSV) and oncolytic measles virus (OMV) and to inhibit the formation of glioblastoma and hepatocellular carcinoma, respectively (Duebgen et al. 2014; Ong et al. 2013).
Furthermore, stem cell derived small extracellular vesicles (sEV) can be used to load cargo molecules such as miRNA, small drugs or proteins. Compared to other manufactured nanoparticles, these endogenous carriers have a number of advantages, such as exceptional intestinal absorption, durability, cargo loading efficiency, biocompatibility, and improved internalization into tumor cells (Fuhrmann et al. 2015). Additionally, they are readily functionalized by adding receptors or corresponding ligands to enhance targeted actions in tumor microenvironment (TME) (Smyth et al. 2014; Kooijmans et al. 2016; Wang et al. 2017). The conventional transfection method was effectively used to encapsulate genetic resources, such as anti-tumor siRNAs/miRNAs, into sEVs generated from stem cells. sEVs from bone stromal cells that expressed miR-146b have shown direct infusion into tumors in a brain tumor model resulted in a notable slowdown of the development of the glioma xenograft (Katakowski et al. 2013). Further research found that sorafenib's anticancer effects on a hepatocellular carcinoma tumor model were markedly improved by sEVs released by MSCs that contain the miR-122 gene (Lou et al. 2015). sEVs made from MSCs also successfully transferred siRNA in order to silence the polo-like kinase 1 gene in bladder cancer (Greco et al. 2016).
3.3 Gene Therapy
It was initially proposed in 1966, subsequently this approach used viruses as the carriers of transforming genetic material in 1968. Gene replacement therapy followed by gene therapy trials, X-SKIDS, oncolytic virus, a gene-based medicines was approved in the cancer therapeutic milieu. Numerous potential approaches are currently ongoing for using gene therapy to treat cancer. It include: (a) use of wild type tumor inhibiting gene; (b) preventing the oncogene expression using an antisense nucleotide approach; (c) expressing apoptosis inducing gene or increase tumor responsiveness to traditional therapy or drugs; and (d) improving the immunogenicity to promote immune cell recognition (Das et al. 2015).
Stem cell-based gene therapy is a recent advancement in cancer cure. Gene directed enzyme prodrug therapy (GDEPT) employ modification of soluble factors such as tumor associated cytokines or prodrug converting enzymes (Malekshah et al. 2016; Sage et al. 2016). A healthy stem cell's enzyme composition can convert prodrugs into bioactive components that are more harmful to tumor tissues. For instance, 5-fluorocytosine is successfully transformed into the tumor-toxic compound 5-fluororacil after being infused with MSCs/NSCs that display the cytosine deaminase enzyme (Malekshah et al. 2016; Lee et al. 2013). Similarly, irinotecan, relatively less powerful prodrug, can be converted into SN-38, a molecule that is thousand times more poisonous, when carboxylesterase is present. When carboxylesterase secreting NSCs and irinotecan were administered together rather than separately, neuroblastoma mouse xenograft model responded more effectively (Choi et al. 2016; Gutova et al. 2017).
3.4 CAR-T/NK Cell Therapy
Expression of CAR by genetically altering NK and T cells, may recognize TAA with precision. “Classical” CARs are made up of an extracellular domain (scFv) that is typically obtained from a mAb fragment and connected to ICD of T-cell receptor. T cell get activated upon tumor antigen coupling with scFv, in a way that is irrespective of MHC, followed by a cytotoxic effect (Hartmann et al. 2017). New CAR constructions have constantly been created, some of which may have modified cytosolic co-stimulatory sequences or targeting sequences. The targeting domain may be made up of other entities rather than scFvs, such as designed ankyrin repeat proteins (DARPins), nanobodies, or ligands (Balakrishnan et al. 2019; Duan et al. 2019; You et al. 2019; Zhylko et al. 2020). Separating antigen detection followed by CAR-cell stimulation has also led to the development of adapter CARs. Using CAR-cells that identify specific adapter molecules for tumor antigens, site specific and time-limited treatment is possible. Multiple antigens may be targeted simultaneously as a result, and the treatment can be modified if tumor types that lack antigens are discovered. This strategy also offers the opportunity to stop the immune reaction if serious adverse effects occur (Lee et al. 2019).
NK or T cells from peripheral blood are initially extracted for the CAR cell production. CAR-encoding nucleotides are then introduced into cells using viral vectors. To combat cancerous cells, CAR-modified cells are multiplied until an adequate cell count is reached and then administered by adoptive transfer to the recipient. Most treatment scenarios include lymphodepletion before the CAR-modified cells are infused to promote effective cell regeneration (Neelapu 2019). It is crucial that allogeneic CAR-T cell treatments are now being developed (Depil et al. 2020; Jamali et al. 2020; Müller et al. 2020; Reindl et al. 2020). FDA and the European Commission have approved three medications for the management of refractory or relapsed hematological malignancies using CAR-T cells during 2017–2021 (I; U.S; Detela and Lodge 2019). A new CART cell approach, which the FDA just authorized (U.S), is now being evaluated in Europe. Following on such pinnacle results, other CAR-T cell treatments are presently being evaluated globally. Around 500 clinical studies looking into CAR-T cells for cancer care have reported. Of them, the most trials are being conducted in East Asia, followed by the US and Europe. In particular, CAR-NK cell therapies are increasingly being used in place of CAR-T cell therapies because they have promising benefits over CAR-T cells, including an inherent ability to destroy cancerous tissues and a limited number of hazard effects after infusion (Yoon et al. 2010; Rubnitz et al. 2010; Moretta et al. 2011). Although there are several CAR-T cell treatments available, only a small number of CAR-NK cell studies are being carried out globally. On clinicaltrials.gov, nineteen studies using CAR-NK cells for solid tumor treatment as well as hematological malignancies are officially enrolled. There are now three CAR-NK cell studies active in the China and US, and just one study being undertaken in Europe. Just few studies are also being conducted right now that focus on CAR-NK/T cell products and CAR-modified cytokine-based killer cells.
3.5 Engineered Cytokines
Cytokines impart various biological processes involved in cancer progression through acting as signal mediators in immune cells and cancer stroma. Structurally modified cytokines are potential immunotherapeutic candidates in cancer management, in which more than twenty modified cytokines are FDA approved for various diseases including cancer. Systematic hazards, off-target effects, poor circulation and reduced target specificity add to the limitations of cytokine-based immunotherapies. Modified cytokines regarding size, modifications in single amino acids (muteins), polymer conjugates and biomaterial implants correspond to engineering concepts of cytokines (Uricoli et al. 2021) (Table 3). The engineered immunostimulatory cytokines fall to size range of nanometer to millimeters for muteins to hydrogel implants respectively. IFNα was the first approved designer cytokine in 1986 which enhance the clinical efficacy of hairy cell leukemia by promoting apoptosis (Berraondo et al. 2019), followed by the approval of IL2 in 1992 for metastatic renal cancer. Both of these cytokines showed a favorable result in a small subset of sample groups while keeping a high degree of hazard effects (Golomb et al. 1986; Sleijfer et al. 2005) including cardiotoxicity, treatment related death and neurotoxicity (Rosenberg et al. 1989, 1998). Nonetheless, IL15 mediated NK cells and CD8+ T cells activation also had similar outcomes (Conlon et al. 2015).
Muteins, designed by single amino acid substitution, are relatively efficient in target based immune activation. IL2 can be considered as a potential pharmacological agent due to their pleotropic effects in vivo. IL2/IL2R complex is specific on CD8+T cells and NK cells than their homology counterpart (CD25) on Treg immunosuppressive cells, which substantiate tumor suppressive environment (Liao et al. 2013). For instance, IL2 superkine, mutated IL2 anywhere as R81D, I86V, I92F, L80F, and L85V, shows high binding affinity with IL2Rβ followed by activation of STAT5 and TGFβ signaling, which further promote tumor inhibiting crosstalk. Moreover, these muteins show competitive binding affinity comparing wildtype IL2/IL2R coupling, also IL2/mAb complex to an extent (Siegel and Puri 1991). Furthermore, two protein engineering method possess significant clinical potential by recombining IL2 superkine with EGFR for tumor cell targeting, and with Fc protein to prolong circulation and to enhance shelf life (Sun et al. 2019). This recombinant protein shows durable tumor suppressive effects both in monotherapy and in combination with chemotherapy. Muteins are also efficient in activating synthetic receptors on CAR-T cells (Sockolosky et al. 2018). IL2/IL2R complex crystal structures were used to create a double mutant IL2R (Y135F, H134D) which lack the ability to bind with wildtype cytokines, and then performed a yeast display-based evolution to enhance the affinity of IL2 to complex with double mutant receptor. Adoptively transferring CD4+/CD8+ T cells expressing adjuvant orthoIL2 or orthoIL2R therapy in immunocompetent mice showed enhanced expression of modified T cell population without any significant hazard effects. Similar effects were reported between wild type and engineered IL2 systems after transferring them into a syngeneic B16F10 mouse melanoma model, highlighting the possibilities of designer cytokine-based immunotherapies to work in conjunction with currently available cancer therapeutic options like CAR-T therapy (Sockolosky et al. 2018; June et al. 2018).
3.6 Strategies for Cytokine Engineering
PEGylation (protein modification using PEG) was first employed in 1970 contribute to efficient mode of cytokine chemical modification strategy (Alconcel et al. 2011; Ekladious et al. 2018). It enhances drug circulation, conformational flexibility, shelf life, drug activity, solubility and reduce nonspecific binding, enzymatic degradation, and opsonization (Naing et al. 2019). PEG modified on lysine residues of IL2 and IL2R on Treg cells improve sustained cytokine delivery in TME and peripheral blood (Charych et al. 2016). Bempegaldesleukin, a PEG modified cytokine, has improved antitumor immunity in syngeneic mouse models of colon and breast carcinoma when performed in conjunction with CAR-T cell therapy (Charych et al. 2016; Parisi et al. 2020) the drug gave promising result in clinical trial (phase I) with 35% tumor regression and 53.8% disease stabilization. Urotheial cancer, advanced melanoma, and muscle invasive bladder cancer is chosen as the disease arm (NCT04209114, NCT03635983, NCT03729245) (Bentebibel et al. 2019). Pegilodecakin, PEG modified IL10, promote oligoclonal T cell activation especially CD8+ Tc cells (Naing et al. 2019). Their phase I monotherapy trial showed 27% OS in renal cell carcinoma, but in combination with PDL1 antibodies showed 40% OS in renal ell carcinoma and 43% in NSCLC. Nonetheless, pegilodecakin has discontinued from phase II and III trials due to their challenging overall survival rates while comparing with FOLFOX therapy (Tsai et al. 2016) or antiPD1 therapy (Sun et al. 2020).
Photolabile polymers can enhance the target specificity of recombinant cytokines by masking them in conjugated form and restoring upon light exposure (Perdue et al. 2020). It can potentially enhance the activation strength, temporal control, and time scale of cytokine signals. PEG modified IL15, IL12, and IL2 linked with o-nitro benzyl linkers precisely retain protein activity upon the exposure of blue LED light. Modifications using photolabile molecules specifically enhanced IL12 half-life in C57BL/6 mice 16-fold by biasing IL2/IL2R binding, while monochromatic light exposure restored IL2 mediated T cell growth and JAK/STAT cascade.
Cytokine hybridization with ECM components is one of the potential strategies to target cytokine response in TME (Xu et al. 2019). Collagen can act as potential tumor agonistic-targeting candidate. IL2 and IL12 fused to collagen type I/IV-binding protein (lumican) significantly improved tolerability and clinical responses in melanoma mouse models, as compared to monotherapy. It further shows therapeutic potential while combining with checkpoint blockade immunotherapy and CAR-T cell therapy. A3 domain of collagen binding domain was fused with IL2 and immune checkpoint blocking antibodies like PDL1 and CTLA4. They reported total remission in 9 out of 13 animals getting combination treatment after systemic therapy in xenograft mouse models of melanoma. With regard to 1 out of 13 mice who received an unaltered combination treatment of collagen binding domain-checkpoint inhibitor and collagen binding domain-IL2, who achieved full remission, monotherapy with CBD-IL-2 did not show significant response (Ishihara et al. 2019).
Cytokines can also be combined with their corresponding receptor fragments to sterically hinder their binding affinity with specific immune cells. ALT-803 is an agonist complex with N72D mutation in IL15. It possesses significant binding potential with IL15Rβ while fused with fragments of IgG1 Fc and IL15Rα. The IL15/IL15R binding affinity caused a 150-fold increase both in NK and T cells, promising enhanced survival in MOPC-315P and 5T33P multiple myeloma cancer models in an IFNγ independent mechanism. Moreover, CD44++ Tc cell proliferation was coupled with NKG2D upregulation without PD1 expression. This validates an innate-like nonspecific tumor suppression (Xu et al. 2013). ALT803 and IL15 in CT26 and B16F10 cancer models showed a competing advantage of anti-tumor activity achieved by 20 fold higher in vivo half-life. ALT803 phase I trial (NCT01885897) in lymphoma and leukemia cases showing relapse after hematopoietic cell transfer no hazard effects and 96h constant serum concentration. Furthermore, 19% of patients have shown measurable Tc cell and NK cell proliferation without Treg proliferation (Liu et al. 2016; Margolin et al. 2018). Combination of rituximab (anti-CD20) with ALT803 in preclinical studies has increased NK mediated granzyme secretion and IFNγ production in primary B-cell lymphoma and follicular lymphoma cells (Rosario et al. 2014). Nivolumab along with ALT803 is ongoing in a phase II trial focusing grade 3 tumors (NCT02523469) (Wrangle et al. 2018).
Antibody-cytokine complexes can further therapeutic potential of cytokine monotherapy. IgG recombined with IL21, IL10, IL12, IL4, IL2, and TNFα showed improved circulation and tissue specificity in clinical trials and murine models (Hutmacher et al. 2019). The whole antibodies or paratope fragments such as scFv, nanobody, Fab, and diabody domains can be linked with cytokines using the principles of affinity binding or using flexible peptide linkers. Antibody complexes can be used to bias cytokine site specific binding, for instance, IL2-S4B6 murine anti-IL2mAb complexes bind with IL2Rβ expressing Tc cells and NK cells but can bind with Treg cells while substituting this antibody with JES61 (Létourneau et al. 2010). The same effects were reported in vivo. Hu14.18-IL2 complex are made using antibody clone 14.18 binding Fc fragment of GD2, which is a disialoganglioside reported in NET like melanoma and neuroblastoma (Neal et al. 2004). Phase I/II clinical trials of Hu14.18-IL2 complex showed clinical outcomes when performed along with additional therapies. Hu14.18-IL2 along with GM-CSF showed 16.1% objective response, but 76% partial response with combination therapy (Shusterman et al. 2019). Clinical studies are now being conducted with Hu.14.18-IL2 for the treatment of Stage IV unresectable melanoma in conjunction with ipilimumab, nivolumab, and radiation therapy (NCT03958383), additionally for the ex vivo growth of functionalised NK cells in neuroblastoma (NCT03209869) (Albertini et al. 2012). Moreover, immunocytokines like L19, F8, F16 can also target neo vasculature or ECM constituents such as extra domain A/B of fibronectin (Villa et al. 2008). F8-IL2 complex specifically deliver IL2 to neo vasculatures, and showed improved clinical efficacy while treated with cytarabine in acute myeloid leukemia (Gutbrodt et al. 2013).
3.7 Methods of Cytokine Delivery
Genetic, particle and chemical-based approaches
IL10 is released by immune system cells, both adaptive and innate, to control the action of proinflammatory cytokines. Evidence to justify the use of autocrine IL10 signaling in cancer immunotherapy, particularly when combined with immune checkpoint suppression are reported. This signaling may extend CD8+ T cell activity (Trinchieri 2007). Utilizing IL-10, nanoparticle systems show tumor suppressive responses via the activation of Th17 cells in colon and lung cancer (Li et al. 2018). Polylactic acid (PLA) microspheres produced using phase inversion nanoencapsulation were used to administer IL-10 orally, and the colon cancer model showed improved survival after dosing. This remarkably effective treatment outcomes were linked to a decrease in pro-tumorigenic Tregs (Foxp3+ CD4+ RORγt-) and Th17 (RORγt+ CD4+ IL-17+) cells, as well as an increase in Tc cells (Gu et al. 2017). Intratracheal delivery of PLA IL10 in the LSL-K-rasG12D genetic model of NSCLC likewise resulted in the restoration of Th17 axis dysfunction and decreased tumor development, which further the potential of particle-based cytokine delivery (Li et al. 2018).
TRAIL can modulate innate immune systems. Lipopolysaccharide (LPS) and IFNβ stimulation induce TRAIL upregulation in monocytes and macrophages, as well IFNγ induce similar effects in dendritic cells and NK cells (Falschlehner et al. 2009). TRAIL/death receptor (DR)-4 and -5 coupling induce apoptotic effects in various cancers, and TRAIL-based immunotherapy demonstrated therapeutic benefits in at least six types of cancers. Their poor circulation and limited target specificity causes modest clinical responses (Soria et al. 2011). To manage, gold, lipid, DNA, and polymer-based structure formulations in nanoparticles are under consideration. TRAIL liposomes can effectively target the tissue sites and can benefit from membrane based presentations (de Miguel et al. 2015). For instance, TRAIL-functionalized liposomes in colon cancer xenograft models enhance ligand-mediated apoptosis through DR5 activation (de Miguel et al. 2015). More sophisticated method such as TRAIL and R8H3 based liposomes modified with hyaluronic acid gel (gel-lipid nanostructure) consisting of doxorubicin can deliver both TRAIL and doxorubicin with improved specificity achieving tumor suppressing potential in breast cancer xenograft model (Jiang et al. 2014). As well, PLA2 degradable POPC liposome shell surrounded by a DNA nanocore can efficiently transfer TRAIL to cancer sites (Sun et al. 2014). This complex achieved tumor cell apoptosis and leukocyte adherence, respectively, by using TRAIL and E-selectin dual conjugated nanoparticle. Xenograft mouse model corresponding to prostate cancer has shown enhanced apoptosis of circulating tumor cells (CTC) and inhibition of metastasis by integrating TRAIL based nanoparticles with leukocytes in vivo (Ming et al. 2017).
IFNγ can enhance effector activity of Tc cell, M1 macrophages, and dendritic cells. IFNγ-engineered delivery systems have the potency to improve the tumor site’s localization prior to clearance to reduce the effects on systemic dosing. IFNγ encapsulated within liposomes can be a method in melanoma vaccination and has experimented in melanoma induced mice models (van Slooten et al. 2000). External magnetic field directed IFNγ-encapsulated nanoparticle enhance tumor infiltration of macrophages and Tc cells in comparison to nondirected nanoparticles in Pan02 pancreatic cancer model (Mejías et al. 2011). IFNγ based nanoparticle consisting of poly lactic-co-glycolic acid (PLGA) and polyvinyl alcohol (PVA) layer for structural support and cytokine secretion, respectively, can adhere to macrophages in vitro while hindering phagocytosis (Wyatt Shields et al. 2020). Comparing naked IFNγ treatment, this complex enhanced M1 polarization and improved OS in 4T1 mammary tumor models.
TNFα assisted isolated limb perfusion (ILP) in soft tissue sarcoma can be competitively benefited with gold nanoparticles due to their potency to deliver rhTNFα in vivo. The particles can be stabilized using Au-S bond and surface conjugation of PEG-SH. Moreover, rhTNFα has three-fold tolerative dose comparing cytimmune (CYT-6091) with no dose-limiting hazard effects (Libutti et al. 2010). Zr/IFNγ and TNFα complexed colloidal gold (CYT-IFNg and CYT-Z-TNF respectively) are also considered (Jiang et al. 2014).
The poor circulation and notable hazard effects of engineered IL12 cause fall in phase III clinical trials (Berraondo et al. 2019). In hypoxic tumor microenvironments, Poly (β-amino ester) consisting of 2-(4-imidazolyl) ethylamine, 1,6-hexanediol diacrylate, and amino-terminal PEG disintegrate, therefore preferentially release IL-12 cargo. In a B16-F10 xenograft mouse model, this system has reported to be durable at physiological pH, significantly enhance intratumoral IL-12, suppress carcinogenesis, and alter TAM phenotype without exhibiting any harmful effects (Overwijk and Restifo 2001).
Hydrogel and implants-based approach
Cytokines can be encapsulated within macroscale matrices or implants to overcome systemic immunotherapy challenges (Chao et al. 2020). Degradable (lactate-)hydroxyethyl methacrylate dextran (dex-lactate-HEMA) hydrogels and nondegradable dextran methacrylated (dex-MA) hydrogels enable for the regulated and adjustable delivery of IL2 (Cadée et al. 2002). Similar clinical efficacy to exogenous IL2 administration and refusal of tumor cell reactivation was obtained after implanting IL2-based hydrogels in mice models of metastatic lymphosarcoma (Bos et al. 2004). Poly(ethyl-l-glutamate)-poly (ethylene glycol)-poly(ethyl-l-glutamate) hydrogels (PELG-PEG-PELG) were loaded with IL-2, IFN, and doxorubicin in B16-F10 melanoma mice model and has shown two-fold tumor inhibition capacity. Chemotherapy-immunotherapy combination approach has also shown similar results using cisplatin and IL15 encapsulated within mPEG-b-PELG assisted hydrogels (Wu et al. 2017). Moreover, IFNα loaded hydrogels hinder tumor proliferation in hepatic xenograft mice models. Hyaluronic acid-based hydrogel loaded with PEGylated TRAIL significantly reduce tumor burden in pancreatic xenograft mice models (Wang et al. 2022).
3.8 Cancer Vaccines
One of the biggest therapeutic breakthroughs of 20th century was the development of vaccinations to combat contagious illnesses, however the vaccination strategies go further than disease prevention (Lin et al. 2022). mAb trials was challenging two decades back before the success of nivolumab, rituximab, CAR-T cell and antiPD1 antibody (Berger 2008). The vaccines can generally mimic the structure of antigens such as tumor cells, neoantigens, long and short peptides, and RNA/DNA, while immune cells such as dendritic cells, carrier proteins, CD40L, and TLR agonists act as adjuvants (Melief et al. 2015). Antigens in vaccines may be either predetermined (known) or anonymous (unknown). The former contains predetermined common antigens (expressed in most patients) or predetermined individualized antigens (specific for individual patient). Ex vivo (in a lab) or in situ (in the tumor site) colocalization of anonymous antigen vaccines with APCs are under consideration (Lin et al. 2022).
3.9 Shared Vaccines
Shared antigens are displayed in a high enough percentage of cancer patients so that vaccine developers may focus on these patient populations. Shared antigens can target both tumor specific antigens (TSA) and tumor associated antigens (TAA), and can be tested using standard tests like IHC, flow cytometry and cytology. They are the major focus of preclinical and clinical studies since 1990 (Lin et al. 2022). Neo-epitope TSA EGFRvIII is exhibited in around 25% of EGFR-overexpressing glioblastomas (Katanasaka et al. 2013), while viral TSA HPV E7 and E6, and TAA Wilms' tumor protein are upregulated in most breast malignancies, Wilm's tumor, and acute myeloid leukemia (AML) (Maslak et al. 2018). These vaccines are beneficial in terms of time and resource comparing personalized vaccines. TSAs of EBV virus encode various neoantigens such as latent membrane protein (LMP)-1 and -2 in NKT cell lymphoma and nasopharyngeal carcinoma (Tsao et al. 2002).
Modified vaccinia Ankara (MVA) virus with positive for EBNA-LMP2 fusion protein enhanced T cell response (Taylor et al. 2014). P53 and RB proteins can be sequestrated by HPV E7 and E6 TSAs promoting squamous carcinoma progression. ISA101, a synthetic long peptide vaccine induces tumor regression through activating T cell response in intraepithelial neoplasia (Kenter et al. 2009). Moreover, LCMVi vectors with E7+ induce E7 specific T cell activation. Overexpressed EGFRvIII are mutant self-proteins widely reported in NSCLC and glioblastoma. Glioblastoma patients previously undertaken CAR-T cell therapy have shown promising anti-EGFRvIII outcomes (O’Rourke et al. 2017). Phase II clinical trial using rindopepimut, an EGFRvIII peptide vaccine, together with temozolomide and GM-CSF activate humoral responses (Schuster et al. 2015) albeit the inability of the trial to show significant clinical outcome (Weller et al. 2017).
TAAs are associated with either tumor site, tumor development, or tumor abundant proteins while comparing with TSAs. Wilm’s tumor protein is a development-based TF that assist tumor progression (Cheever et al. 2009). WT1 peptide vaccines with higher HLA affinity enhance clinical responses in acute myeloid leukemia cases and prompted to phase III trials now (NCT04229979) (Maslak et al. 2018). Melanoma-associated antigen 3 (MAGE-A3) is a widely used biomarker in NSCLC, melanoma, and myeloma wherein it induces anti-apoptotic effect. TLR4-agonist MAGE-A3 vaccine, AS02B, as well show humoral tumor suppressive effects without significant clinical benefits (Vansteenkiste et al. 2013). However, AS15, another TLR4 agonist shows competing clinical benefits over AS02B, and is in phase II clinical trials now (Kruit et al. 2013).
HER2 is upregulated in 30% breast carcinoma but a relatively smaller percentage in ovarian and gastrointestinal tumors, and can get bound by anti-HER2 mAb. Nelipepimut-S, HLA-I-restricted peptide vaccine, can induce Tc cell response but with reduced clinical benefits (Mittendorf et al. 2016), and is similar to AE37 (HLA-II-restricted) (Mittendorf et al. 2019). Interestingly, a multi-epitope-based HLA-I or II restricted targeting showed Tc cell response durable for a year (Knutson et al. 2001).
3.10 Personalized Vaccines
Personalized vaccines are designed specific to cancer patients and are designed using high-throughput nucleic acid sequencing methods. Germline nucleic acid extraction, sequencing, and HLA typing are employed upstream to personalized vaccine designing. For instance, neo-epitopes of TSA KRAS G12D, which has reduced frequency in terms of oncogene, is employed. This method puts the responsibility on the vaccinator to repeatedly identify the significant epitopes, however it also enables the immune system to approach malignancies without recognized common antigens. The avidity between HLA and TCR ensures clinical efficiency of vaccine as a whole (Lin et al. 2022). When combined to checkpoint blockage, targeting customized antigens releases patients' broad T cell responsiveness is increased by T cells that evade thymic −ve determination(Sahin). Putative neo-epitopes are selected from somatic mutations using techniques like NetMHC algorithm based on their affinity for the patient's HLA alleles (Nielsen et al. 2003). After prioritizing neo-epitopes based on analysis of tumoral transcriptome data, It is customary to choose up to 20 neo-epitopes, and then produce RNA, neo-epitope viral vectors, or peptides that adhere to good manufacturing practice (GMP). Neo-epitopes may promote APC activation or keep APC uptake stable to help them be more immunogenic.
Patients diagnosed with advanced melanoma responded anecdotally to an earlier customized vaccination employing synthetic RNA vaccine to generate 10 neo-epitope potential targets, primarily CD8+ and CD4+ neo-epitope-specific T cell activity (Nielsen et al. 2003). These poly-specific processes, which might be made more potent by inhibiting PD-1 or rendered useless by reducing tumor cell HLA class I, most certainly had a key role in the dramatic decline in long-term metastatic episodes. Accordingly, another study using 13–20 long fragments of projected NEO-PV-01 (neo-epitope) generated higher CD4+ over CD8+ T cells particular to altered sequence (Ott 2017).
Another tumor-specific alteration is the distinctive Ig or TCR idiotype, which results via somatic hypermutation and locus gene recombination. These mutations are often retained in tumor tissues and is reported in lymphomas, myelomas, and hematologic malignancies. In the Genitope and Favrille phase III studies, lymphoma patients who had received rituximab or chemotherapy but had an idiotype associated to KLH delivered with GM-CSF were given the vaccine. Neither research demonstrated a therapeutic advantage over a placebo. When given to patients in standard therapy following chemotherapy, a different NCI-Bioves trial (phase III), using the same vaccine approach showed a substantial DFS; however, the relevance of the consequence was obscured by the high rate of patient dropout prior to vaccination. Flt3L has shown advantage over GM-CSF based APC activation, substantially with more efficient adaptive immune response (2016).
3.11 Ex Vivo Vaccines
APC colocalized or ex vivo include tumor cell isolation and colocalization with APC in order to enhance their antigenic effects. Ex vivo vaccines in patient body can express antigens to T cells. HSP such as HSP70, gp96, and HSP110-based ex vivo vaccines has shown to cause tumor suppressive responses (Moseley 2000). HSP-gp96 complex (HSPPC-96) could not report significant survival benefits in renal cell carcinoma and melanoma (Testori et al. 2008; Wood et al. 2008). Glioblastoma patients receiving this vaccination showed inverse correlation between PDL1 expression and survival, suggesting combinatorial approaches using antiPDL1 and HSSPC-96(NCT03018288) (Bloch et al. 2017). TAAs can be enriched in ex vivo vaccines focusing KRAS, P53, and EGFR. Canvaxin, prostate GVAX, melacine, and lucaxin are few ex-vivo vaccines but with limited survival values (Kozłowska et al. 2013). Tumor cells expressed with GM-CSF (GVAX) in acute myeloid leukemia patients did not show clinical benefits (Ho et al. 2022), however their combination with anti-furin shRNA to inhibit TGFβ production shows clinical benefits in Ewing’s sarcoma (Ghisoli 2016).
3.12 In Situ Vaccines
In situ vaccines present APC to stimulate tumor loading antigen, followed by T cell activation. They are benefited for their ability to present wide ranges of TSA while comparing with ex vivo. DCs which are usually inactivated in TME can be administered ex vivo to stabilize intratumoral cytokine levels including IL12, TNF and IL8 (Subbiah et al. 2018). Despite minimal radiotherapy, embryonic dendritic cells with improved phagocytic capability can be infused with GMCSF and rituximab (Kolstad et al. 2015). The importance of vigilant immune surveillance is highlighted by periodic T cell regressions and activation at nearby and distant malignancies. Similar research without radiation produced lymphoma-specific Tc and Th activation as well as tumor regressions in untreated malignancies (Cox et al. 2019). CC chemokine ligand (CCL)-1 expressing DCs injected in NSCLC patients induced PDL1 expression and Tc cell infiltration, indicating their anti-tumoral effects (Lee et al. 2017).
Flt3L, cross-presenting fraction cDC1, is the main hematopoietic progenitor development and differentiation component that mobilizes dendritic cells. Therefore, administering Flt3L rather than their direct administration could be a more feasible way to replace tumor tissue dendritic cells. Nine out of 29 vaccinated NSCLC patients who received targeted irradiation with Flt3L infusion had corresponding responses (Cox et al. 2019). B cell lymphoma patients who received intratumoral Flt3L, low-dose radiation and poly-ICLC, as part of a phase I-II study had early signs of memory Tc cell infiltration to untreated tumor locations linked to overall tumor shrinkage, a few of them lasted from months to years (Hammerich et al. 2019), however their follow up trial targeting PD1 in situ vaccination in breast, head and neck cancer is ongoing (NCT03789097).
Dendritic cells like plasmacytoid (pDC) express TLR9 receptors but not in cDC1. TLR9 agonists include hypomethylated CpG islands especially CpG-A, -B, or -C that can cause IFN production that is pro-inflammatory. Phase III trial using CpG-B tilsotolimod and ipilimumab showed ORR of 9% (Haymaker), which is similar to ipilimumab monotherapy. CMP-001 (CpG-A) vaccine in melanoma patients 28% ORR while administered in combination with pembrolizumab albeit CMP-001 monotherapy causes systemic regression (Milhem et al. 2020). TLR3 expressed on cDC1 can activate nuclear factor (NF)-kB pathway. Hiltonol, a TLR3 agonist, can control tumor growth and prolonged survival in head and neck cancer, and liver cancer (Torre 2017; Kyi et al. 2018).
Oncolytic bacteria and viruses that are intratumorally injected can be a subtype of in situ vaccination. The possibility for systemic vaccination following intratumoral delivery of oncolytic viruses might lead to a variety of therapeutic strategies. The only oncogenic virus recognised by the FDA is talimogene laherparepvec (TVEC), a transgenic GM-CSF-releasing herpes simplex virus (HSV)-1 that has increased tumour shrinkage and survival in non-injected areas (Andtbacka 2015; Kaufman et al. 2016).
3.13 Vaccine Delivery Vehicles
Biotechnology has the potency to answer limitations of cancer vaccines especially regarding toxicity, shelf life, tumor penetration, opsonization, and off-target effects by designing vaccine delivery vehicles (Rosenblum et al. 2018; Kudling et al. 2022). Nanoparticle based delivery systems can carry antagonists or agonists to modulate tumor immune system. It includes liposomes, extracellular vesicles, dendrimers, inorganic nanocarriers, and more (Li et al. 2022; Reda et al. 2022). It can be exploited well in combination therapy, for instance administration of cytokines along with chemotherapeutic drugs. Magnetic nanoparticles (Feridex, EndoremVR, GastromarkVR), virus derived, taxane based, and polymer nanoparticles can incorporate pharma compounds to the target sites (Carrasco-Esteban et al. 2021). Extracellular vesicles are nanosized membrane enclosed structures naturally derived from cells and consist of a wide spectrum of their parent cells (Wiklander et al. 2019). They are involved in normal physiology as well in pathology through transferring cargo (Yang et al. 2021). Engineered EVs derived from fibroblast like mesenchymal cells, for instance, target KRAS pathway with the help of their cargo such as siRNA and shRNA (Ma et al. 2021). Additionally, because they lack the carriers recommended for the treatment of cancer, pharmaceuticals themselves may function as nanoscale therapies. Antibody-drug conjugates (ADCs), drug nanocrystals, drug-drug conjugate nanoparticles, and prodrug self-assembled nanoparticles are examples of nanoparticles devoid of carrier molecules. For instance, the drug SN38 (7-ethyl-10-hydroxycamptothecin) has indeed been coupled with the PEG-CH=N-Doxorubicin precursor (pH-responsive) to increase drug release to tumors and destroy both CSC and non-CSCs (Carrasco-Esteban et al. 2021).
4 Biotechnology Approaches Over Traditional Therapy
Surgery, radiation therapy, chemotherapy, targeted therapy, and immunotherapy are the five conventional cancer care modalities (Siamof et al. 2020). With unique shortcomings in these therapeutic approaches are now being addressed by designer, specially formulated drugs designed to increase survival rates and reduce adverse effects, clinical cancer treatment has undergone a fundamental revolution. Following surgery, the whole lung tumor and any nearby lymph nodes must be eliminated (Hoy et al. 2019). Radiation treatment cannot be performed in aggressive situations. It destroys malignant cells that are allowed for radiation. It also damages the normal tissue that are located in its exposure (Bogart et al. 2022). Chemotherapy stops malignant cells from growing and dividing, multiplying, and generating additional cells (Nagasaka and Gadgeel 2018). Immunotherapy aim to improve the body's natural defenses fight cancer. For the purpose of re-establishing immune response functionality, it uses substances that may be created in vivo or in vitro. Patients with NSCLC may get immunotherapy alone, or in conjunction with other chemotherapy medicines. When targeted treatment is not an option, immunotherapy or their combinational methods may be used (American Society of Clinical Oncology (ASCO) 2020).
ICIs, mAb, vaccines, and adoptive cancer therapy (ACT) are the recent biotechnology advancements in cancer immunotherapy. The biotechnology applications though ranges from nucleic acid-protein sequencing, drug discovery, vaccine designing, cancer model preparations, it is widely benefited in chemotherapy, immunotherapy, and combination therapy. ICIs are generally designed to target PD1/PDL1, and CTLA4, and the former ones are FDA approved (Jain 2018; Sharon 2014) (Fig. 3). ICIs mainly function by inhibiting the immune suppressive signal crosstalk that is triggered by the interaction between PD1 and PDL1, regaining T lymphocytes' basic ability to eradicate tumor cells (Sławiński et al. 2020).

Data source https://www.rcsb.org/
RCSB-PDB structures of PD1 and CTLA4 complexed with various ligand or drug molecules. Green color represents PD1 and CTLA4, while further colors denote heavy and light chains of bound molecules. a 6UMT: PD1/PDL2 (Tang and Kim 2019) b 7VUX: PD1/609A Fab (Zhao et al. 2022) c 5WT9: PD1/nivolumab d 7CU5: PD1/camrelizumab (Liu et al. 2020) e 6JBT: PD1/toripalimab (Liu et al. 2019) f 7E9B: PD1/ HLX10 (anti-PD1 antibody) (Issafras et al. 2021) g 1I85: CTLA4/B7 (Schwartz et al. 2001) h 5GGV: CTLA4/tremelimumab (Lee et al. 2016) i 5TRU: CTLA4/ipilimumab (Ramagopal et al. 2017).
IgG1 and IgG4 antibodies, such as avelumab, atezolizumab, cemiplimab, durvalumab, pembrolizumab, and nivolumab, may hijack PD1/PDL1 axis in NSCLC in combinatorial first-line treatment. They can elicit tumor inhibiting effects via adaptive and innate immune system pathways (Chen 2017; Mezquita and Planchard 2018; Yuan et al. 2019; Akinleye and Rasool 2019). A range of disorders, particularly cancers with different origin, are now being treated using ICIs, especially blocking via mAbs. Integrated treatment, which employs mAbs as well as other drugs at a reasonable dosage, may increase patients' chances of surviving NSCLC (Rosenblatt and Avigan 2017).
The effectiveness of ipilimumab (CTLA-4 inhibitor) in ES-SCLC in conjunction with the first treatment was investigated in the CA184-041, ICE, and CA184-156 phase 2 and 3 investigations. 42 participants were included in the ICE study and administered ipilimumab along with etoposide and carboplatin. PFS was indeed the study's primary goal, however it wasn't effectively attained (PFS 6.9 months) (Antonia et al. 2016). CTLA4 based therapy require combinatorial approaches for better clinical benefits.
5 Combination Therapy Involving Biotechnology
Patients with NSCLC or aggressive melanoma respond well to ICI, but B cell acute lymphoblastic leukemia cases have reported with outstanding results in CAR T-cell therapy. Understanding why particular medicines have been extremely effective in treating certain cancers but less effective in treating others might aid in the more rational design of clinical trials to test therapy for additional malignancies (Khalil et al. 2016). Immune hijacking adopted by TME can suppress endogenous TILs and CAR-T cells from producing a potent antitumor response (John 2013; Moon et al. 2014). Immune modifying mAbs, including those that provide checkpoint blockage, would probably be required for the formation of immune based antitumor treatments for tumors with significant neoantigen expressing capability in a immunosensitive milieu. Evolutionarily conserved T cells may be oblivious to cancers with reduced neoantigen-presenting ability, like those with fewer possibly immunostimulatory genetic abnormalities such as B cell acute lymphoblastic leukemia (Alexandrov 2013) or those other ways, do not display neoantigens due to reduced antigen processing, demonstration, or HLA expression. It is possible that in an immunoinhibitory milieu, these cancer kinds did not face the urge to co-evolve. mAbs individually would be fewer likely to induce a potent antitumor activity in this circumstance since TIL and antigen presentation load are likely to be modest. In contrast, CAR-T cells does not get inhibited by such inhibitors, and as shown for CD19 targeted CAR-T cell treatment for B-ALL, may cause fast full cures in approximately to 90% of participants with this cancer type, which has a modest mutational rate (Alexandrov 2013). Combination treatment will exert the efficient anticancer impact when PD1 axis and CTLA4 are blocked. Combined anti-PD1 t and anti-CTLA-4 treatment has reported a great deal of potential (Postow 2015; Chapman et al. 2015; Larkin 2015). For instance, individuals with metastatic melanoma (phase III) showed a remarkable 58% RR when treated using this strategy. Again, for treatment of malignancy, medicines hitherto believed to not work via immune regulation as well as combinations with other types of immune regulation are being extensively researched (Larkin 2015).
ISA101 (LSP vaccine) and anti-PD-1 medication were combined in research that showed improved therapeutic benefits compared to either treatment used alone, including in PD-L1 positive malignancies (Quezada et al. 2006). Higher humoral effects as well as an OS advantage as a supplementary, inadequate objective were seen in randomized phase II study of EGFRvIII vaccine along with bevacizumab. The findings imply that tumor inhibitory humoral mechanisms could not be enough and that selecting the best possible combination therapy may determine effectiveness of vaccines (Postow 2015). Neoantigen-based therapeutic benefits and T cell responses may be stronger comparing those anticipated with anti-PD1 monotherapy, according to a wider trial that included anti-PD-1 and neo-epitope vaccination in 60 melanoma cases, bladder cancer and NSCLC (Lozano et al. 2012). In a pilot investigation, anti-neo-epitope and tumor inhibitory effects by T cells connected to long-term survival were induced in 25 ovarian cancer cases when autologous dendritic cells with oxidized autologous tumor cell homogenate were injected alone or in combination with chemotherapy and anti-VEGF mAb (Wang et al. 2019a, b).
6 Conclusion and Future Perspectives
Although years of research in cancer have yielded many therapeutic strategies, disease resistance, recurrence, and post-medication hazard effects are still a major concern. The PDCD1, CD274, and CTLA4 gene which are widely exploited in mAb designing and ICIs, were curated from TCGA-cBioportal and have shown significant co-occurrence and expression overlaps. This indicates the need for a combined approach in designing ICIs and mAbs. Moreover, the exact regulation or controlling engineered cytokines or their delivery vehicles' activity is a significant barrier to the efficiency of cancer immunotherapy. As an alternative approach, ongoing research examines the role of photolabile cytokines on adoptively transplanted T cell durability and specificity, and the application of photocages with enhanced local penetrance. The addition of cytokines with IgG motifs can significantly change target specificity as well as the eventual safety and effectiveness the otherwise poorly tolerated or ineffective cytokine treatment. Continued studies on TME, novel antibody architectures, and cytokines in innate immunity may contribute to the designing of antibody-cytokine combinations and hybrids and, consequently, the therapeutic outcomes for patients receiving this potential category of immunotherapy. T cell responses corresponding to neo-antigens and clinical responses that may have exceeded those anticipated with anti-PD1 monoclonal therapy were found in larger research involving multiple solid tumors (Ott et al. 2020). Understanding the initial cancer mutational burden of each patient before the application of conventional therapeutic approaches and combining these advancements may be a potential strategy for cancer cure.
References
Adapter chimeric antigen receptor (aCAR)-engineered NK-92 cells: an off-the-shelf cellular therapeutic for universal tumor targeting. Blood 1
Akinleye A, Rasool Z (2019) Immune checkpoint inhibitors of PD-L1 as cancer therapeutics. J Hematol Oncol 12. https://doi.org/10.1186/S13045-019-0779-5
Albertini MR, Hank JA, Gadbaw B et al (2012) Phase II trial of hul4.18-IL2 for patients with metastatic melanoma. Cancer Immunol Immunother 61:2261. https://doi.org/10.1007/S00262-012-1286-5
Alconcel SNS, Baas AS, Maynard HD (2011) FDA-approved poly(ethylene glycol)–protein conjugate drugs. Polym Chem 2:1442–1448. https://doi.org/10.1039/C1PY00034A
Alexandrov, (2013) LB Signatures of mutational processes in human cancer. Nature 500:415–421
American Society of Clinical Oncology (ASCO) (2020) Lung Cancer—Non-Small Cell: Types of Treatment | Cancer.Net. In: Cancer.Net. https://www.cancer.net/cancer-types/lung-cancer-non-small-cell/types-treatment. Accessed 10 Oct 2022
Anand R (2019) Combination of CTLA-4 and PD-1 blockers for treatment of cancer. J Exp Clin Cancer Res 38:1–12
Andtbacka RHI (2015) Talimogene laherparepvec improves durable response rate in patients with advanced melanoma. J Clin Oncol 33:2780–2788
Antonia SJ, Lopez-Martin JA, Bendell J (2016) Nivolumab aloneand nivolumab plus ipilimumab in recurrent small-cell lung cancer (CheckMate 032): a multicentre, open-label, phase 1/2 trial. Lancet Oncol 17:10 1016 1470–2045 16 30098–5
Balakrishnan A, Rajan A, Salter AI et al (2019) Multispecific targeting with synthetic ankyrin repeat motif chimeric antigen receptors. Clin Cancer Res 25:7506–7516. https://doi.org/10.1158/1078-0432.CCR-19-1479
Batlle E, Clevers H (2017) Cancer stem cells revisited. Nat Med 23:1124–1134. https://doi.org/10.1038/nm.4409
Behzadi S, Serpooshan V, Tao W et al (2017) Cellular uptake of nanoparticles: journey inside the cell. Chem Soc Rev 46:4218–4244. https://doi.org/10.1039/c6cs00636a
Bentebibel SE, Hurwitz ME, Bernatchez C et al (2019) A first-in-human study and biomarker analysis of NKTR-214, a novel IL2Rβγ-biased cytokine, in patients with advanced or metastatic solid tumors. Cancer Discov 9:711–721. https://doi.org/10.1158/2159-8290.CD-18-1495
Berger R (2008) Phase I safety and pharmacokinetic study of CT-011, a humanized antibody interacting with PD-1, in patients with advance hematologic malignancies. Clin Cancer Res 14:3044–3051
Berraondo P, Sanmamed MF, Ochoa MC et al (2019) Cytokines in clinical cancer immunotherapy. Br J Cancer 120:6–15. https://doi.org/10.1038/S41416-018-0328-Y
Bloch O, Lim M, Sughrue ME et al (2017) Autologous heat shock protein peptide vaccination for newly diagnosed glioblastoma: impact of peripheral PD-L1 expression on response to therapy. Clin Cancer Res 23:3575–3584. https://doi.org/10.1158/1078-0432.CCR-16-1369
Bogart JA, Waqar SN, Mix MD (2022) Radiation and systemic therapy for limited-stage small-cell lung cancer. J Clin Oncol 40:661–670. https://doi.org/10.1200/JCO.21.01639
Bos GW, Jacobs JJL, Koten JW et al (2004) In situ crosslinked biodegradable hydrogels loaded with IL-2 are effective tools for local IL-2 therapy. Eur J Pharm Sci 21:561–567. https://doi.org/10.1016/j.ejps.2003.12.007
Buchbinder EI, Desai A (2016) CTLA-4 and PD-1 pathways similarities, differences, and implications of their inhibition. Am J Clin Oncol: Cancer Clin Trials 39:98–106. https://doi.org/10.1097/COC.0000000000000239
Cadée JA, de Groot CJ, Jiskoot W et al (2002) Release of recombinant human interleukin-2 from dextran-based hydrogels. J Control Release 78:1–13. https://doi.org/10.1016/S0168-3659(01)00483-7
Carrasco-Esteban E, Domínguez-Rullán JA, Barrionuevo-Castillo P et al (2021) Current role of nanoparticles in the treatment of lung cancer. J Clin Transl Res 7:140. https://doi.org/10.18053/jctres.07.202102.005
Chang JC (2016) Cancer stem cells: Role in tumor growth, recurrence, metastasis, and treatment resistance. Medicine (United States) 95:S20–S25. https://doi.org/10.1097/MD.0000000000004766
Chao Y, Chen Q, Liu Z (2020) Smart injectable hydrogels for cancer immunotherapy. Adv Funct Mater 30:1902785. https://doi.org/10.1002/ADFM.201902785
Chapman PB, D’Angelo SP, Wolchok JD (2015) Rapid eradication of a bulky melanoma mass with one dose of immunotherapy. N Engl J Med 372:2073–2074. https://doi.org/10.1056/nejmc1501894
Charych DH, Hoch U, Langowski JL et al (2016) NKTR-214, an engineered cytokine with biased IL2 receptor binding, increased tumor exposure, and marked efficacy in mouse tumor models. Clin Cancer Res 22:680–690. https://doi.org/10.1158/1078-0432.CCR-15-1631
Cheever MA, Allison JP, Ferris AS et al (2009) The prioritization of cancer antigens: a national cancer institute pilot project for the acceleration of translational research. Clin Cancer Res 15:5323–5337. https://doi.org/10.1158/1078-0432.CCR-09-0737
Chen YM (2017) Immune checkpoint inhibitors for nonsmall cell lung cancer treatment. J Chin Med Assoc 80:7–14. https://doi.org/10.1016/J.JCMA.2016.08.005
Chen JS, Lan K, Hung MC (2003) Strategies to target HER2/neu overexpression for cancer therapy. Drug Resist Updates 6:129–136. https://doi.org/10.1016/S1368-7646(03)00040-2
Choi SS, Yoon K, Choi SA et al (2016) Tumor-specific gene therapy for pancreatic cancer using human neural stem cells encoding carboxylesterase. Oncotarget 7:75319–75327. https://doi.org/10.18632/oncotarget.12173
Codd AS, Kanaseki T, Torigo T, Tabi Z (2018) Cancer stem cells as targets for immunotherapy. Immunology 153:304–314. https://doi.org/10.1111/imm.12866
Cojoc M, Mäbert K, Muders MH, Dubrovska A (2015) A role for cancer stem cells in therapy resistance: cellular and molecular mechanisms. Semin Cancer Biol 31:16–27. https://doi.org/10.1016/j.semcancer.2014.06.004
Conlon KC, Lugli E, Welles HC et al (2015) Redistribution, hyperproliferation, activation of natural killer cells and CD8 T cells, and cytokine production during first-in-human clinical trial of recombinant human interleukin-15 in patients with cancer. J Clin Oncol 33:74–82. https://doi.org/10.1200/JCO.2014.57.3329
Copelan EA (2006) Hematopoietic stem-cell transplantation. N Engl J Med 354:1813–1826
Cox MC, Castiello L, Mattei M et al (2019) Clinical and antitumor immune responses in relapsed/refractory follicular lymphoma patients after intranodal injections of IFNα-Dendritic cells and rituximab: a phase i clinical trial. Clin Cancer Res 25:5231–5241. https://doi.org/10.1158/1078-0432.CCR-19-0709
Das SK, Menezes ME, Bhatia S et al (2015) Gene therapies for cancer: strategies, challenges and successes. J Cell Physiol 230:259. https://doi.org/10.1002/JCP.24791
Depil S, Duchateau P, Grupp SA et al (2020) Off-theshelf’ allogeneic CAR T cells: development and challenges. Nat Rev Drug Discov 19:185–199
Detela G, Lodge A (2019) EU regulatory pathways for ATMPs: standard, accelerated and adaptive pathways to marketing authorisation. Mol Ther Methods Clin Dev 13:205–232. https://doi.org/10.1016/j.omtm.2019.01.010
Duan H, Huang H, Jing G (2019) An antibody fab fragment-based chimeric antigen receptor could efficiently eliminate human thyroid cancer cells. J Cancer 10:1890–1895. https://doi.org/10.7150/jca.30163
Duebgen M, Martinez-Quintanilla J, Tamura K, et al (2014) Stem cells loadedwith multimechanistic oncolytic herpes simplex virus variants for brain tumor therapy. J Natl Cancer Inst 106
Ekladious I, Colson YL, Grinstaff MW (2018) Polymer–drug conjugate therapeutics: advances, insights and prospects. Nat Rev Drug Discov 18:4 18:273–294. https://doi.org/10.1038/s41573-018-0005-0
Falschlehner C, Schaefer U, Walczak H (2009) Following TRAIL’s path in the immune system. Immunology 127:145–154. https://doi.org/10.1111/J.1365-2567.2009.03058.X
Fuhrmann G, Serio A, Mazo M et al (2015) Active loading into extracellular vesicles significantly improves the cellular uptake and photodynamic effect of porphyrins. J Control Release 205:35–44. https://doi.org/10.1016/j.jconrel.2014.11.029
Ghisoli M (2016) Three-year follow up of GMCSF/bi-shRNAfurin DNA-transfected autologous tumor immunotherapy (Vigil) in metastatic advanced Ewing’s sarcoma. Mol Ther 24:1478–1483
Golomb HM, Jacobs A, Fefer A et al (1986) Alpha-2 interferon therapy of hairy-cell leukemia: a multicenter study of 64 patients. J Clin Oncol 4:900–905. https://doi.org/10.1200/JCO.1986.4.6.900
Gomes JP, Assoni AF, Pelatti M et al (2017) Deepening a simple question: can MSCs be used to treat cancer? Anticancer Res 37:4747–4758
Greco KA, Franzen C, Foreman KE et al (2016) PLK-1 silencing in bladder cancer by siRNA delivered with exosomes. Urology 91:241 1–241 7
Gu T, de Jesus M, Gallagher HC et al (2017) Oral IL-10 suppresses colon carcinogenesis via elimination of pathogenicCD4+ T-cells and induction of antitumor CD8+ T-cell activity. Oncoimmunology 6. https://doi.org/10.1080/2162402X.2017.1319027/SUPPL_FILE/KONI_A_1319027_SM1530.ZIP
Gutbrodt KL, Schliemann C, Giovannoni L et al (2013) Antibody-based delivery of interleukin-2 to neovasculature has potent activity against acute myeloid leukemia. Sci Transl Med 5. https://doi.org/10.1126/SCITRANSLMED.3006221
Gutova M, Goldstein L, Metz M et al (2017) Optimization of a neural stem-cell-mediated carboxylesterase/irinotecan gene therapy for metastatic neuroblastoma. Mol Ther Oncolytics 4:67–76. https://doi.org/10.1016/j.omto.2016.11.004
Hammerich L, Marron TU, Upadhyay R et al (2019) Systemic clinical tumor regressions and potentiation of PD1 blockade with in situ vaccination. Nat Med 25:814–824. https://doi.org/10.1038/s41591-019-0410-x
Hartmann J, Schüßler‐Lenz M, Bondanza A, Buchholz CJ (2017) Clinical development of CAR T cells—challenges and opportunities in translating innovative treatment concepts. EMBO Mol Med 9:1183–1197. https://doi.org/10.15252/emmm.201607485
Haymaker C (n.d.) Tilsotolimod with ipilimumab drives tumor responses in anti-PD-1 refractory melanoma. Cancer Discov 11:1996–2013
Ho VT, Kim HT, Brock J et al (2022) GM-CSF secreting leukemia cell vaccination for MDS/AML after allogeneic HSCT: a randomized, double-blinded, phase 2 trial. Blood Adv 6:2183–2194. https://doi.org/10.1182/bloodadvances.2021006255
Hoy H, Lynch T, Beck M (2019) Surgical treatment of lung cancer. Crit Care Nurs Clin North Am 31:303–313. https://doi.org/10.1016/J.CNC.2019.05.002
Hutmacher C, Nuñez NG, Liuzzi AR et al (2019) Targeted delivery of IL2 to the tumor stroma potentiates the action of immune checkpoint inhibitors by preferential activation of NK and CD8+ T Cells. Cancer Immunol Res 7:572–583. https://doi.org/10.1158/2326-6066.CIR-18-0566
I GS Kite’s TecartusTM (KTE-X19) Granted Conditional Marketing Authorization for the Treatment of Relapsed or Refractory Mantle Cell Lymphoma in Europe
Ishihara J, Ishihara A, Sasaki K et al (2019) Targeted antibody and cytokine cancer immunotherapies through collagen affinity. Sci Transl Med 11. https://doi.org/10.1126/SCITRANSLMED.AAU3259
Issafras H, Fan S, Tseng CL et al (2021) Structural basis of HLX10 PD-1 receptor recognition, a promising anti-PD-1 antibody clinical candidate for cancer immunotherapy. PLoS One 16. https://doi.org/10.1371/JOURNAL.PONE.0257972
JR (n.d.) Serotherapy of acute lymphoblastic leukemia with monoclonal antibody. Blood 58:141–152
Jain P (2018) Role of immune-checkpoint inhibitors in lung cancer. Ther Adv Respir Dis 12:1753465817750075
Jamali A, Hadjati J, Madjd Z et al (2020) Highly efficient generation of transgenically augmented CAR NK cells overexpressing CXCR4. Front Immunol 11. https://doi.org/10.3389/fimmu.2020.02028
Jiang T, Mo R, Bellotti A et al (2014) Gel–liposome-mediated co-delivery of anticancer membrane-associated proteins and small-molecule drugs for enhanced therapeutic efficacy. Adv Funct Mater 24:2295–2304. https://doi.org/10.1002/ADFM.201303222
John LB (2013) Anti PD 1 antibody therapy potently enhances the eradication of established tumors by gene-modified T cells. Clin Cancer Res 19:5636–5646
Jordan CT, Guzman ML, Noble M (2006) Cancer stem cells. N Engl J Med 355:1253–1261
June CH, O’Connor RS, Kawalekar OU et al (2018) CAR T cell immunotherapy for human cancer. Science 359:1361–1365. https://doi.org/10.1126/SCIENCE.AAR6711
Katakowski M, Buller B, Zheng X et al (2013) Exosomes from marrow stromal cells expressing miR-146b inhibit glioma growth. Cancer Lett 335:201–204. https://doi.org/10.1016/j.canlet.2013.02.019
Katanasaka Y, Kodera Y, Kitamura Y et al (2013) Epidermal growth factor receptor variant type III markedly accelerates angiogenesis and tumor growth via inducing c-myc mediated angiopoietin-like 4 expression in malignant glioma. Mol Cancer 12:31. https://doi.org/10.1186/1476-4598-12-31
Kaufman HL, Amatruda T, Reid T et al (2016) Systemic versus local responses in melanoma patients treated with talimogene laherparepvec from a multi-institutional phase II study. J Immunother Cancer 4:12. https://doi.org/10.1186/s40425-016-0116-2
Kenter GG (2009) Vaccination against HPV-16 oncoproteins for vulvar intraepithelial neoplasia. N Engl J Med 361:1838–1847
Khalil DN, Smith EL, Brentjens RJ, Wolchok JD (2016) The future of cancer treatment: immunomodulation, CARs and combination immunotherapy. Nat Rev Clin Oncol 13:273–290. https://doi.org/10.1038/NRCLINONC.2016.25
Khan S, Ullah MW, Siddique R et al (2016) Role of recombinant DNA technology to improve life. Int J Genomics. https://doi.org/10.1155/2016/2405954
Knutson KL, Schiffman K, Disis ML (2001) Immunization with a HER-2/neu helper peptide vaccine generates HER-2/neu CD8 T-cell immunity in cancer patients. J Clin Investig 107:477–484. https://doi.org/10.1172/JCI11752
Kolstad A, Kumari S, Walczak M et al (2015) Sequential intranodal immunotherapy induces antitumor immunity and correlated regression of disseminated follicular lymphoma. Blood 125:82–89. https://doi.org/10.1182/blood-2014-07-592162
Kooijmans SAA, Schiffelers RM, Zarovni N, Vago R (2016) Modulation of tissue tropism and biological activity of exosomes and other extracellular vesicles: new nanotools for cancer treatment. Pharmacol Res 111:487–500. https://doi.org/10.1016/j.phrs.2016.07.006
Kooreman NG, Kim Y, de Almeida PE et al (2018) Autologous iPSC-based vaccines elicit anti-tumor responses in vivo. Cell Stem Cell 22:501-513.e7. https://doi.org/10.1016/j.stem.2018.01.016
Kozłowska A, Mackiewicz J, Mackiewicz A (2013) Therapeutic gene modified cell based cancer vaccines. Gene 525:200–207. https://doi.org/10.1016/j.gene.2013.03.056
Kruit WHJ, Suciu S, Dreno B et al (2013) Selection of immunostimulant AS15 for active immunization with MAGE-A3 protein: results of a randomized phase II study of the European organisation for research and treatment of cancer melanoma group in metastatic melanoma. J Clin Oncol 31:2413–2420. https://doi.org/10.1200/JCO.2012.43.7111
Kudling TV, Clubb JHA, Quixabeira DCA et al (2022) Local delivery of interleukin 7 with an oncolytic adenovirus activates tumor-infiltrating lymphocytes and causes tumor regression. Oncoimmunology 11:20965–20972. https://doi.org/10.1080/2162402X.2022.2096572
Kyi C, Roudko V, Sabado R et al (2018) Therapeutic immune modulation against solid cancers with intratumoral poly-ICLC: A pilot trial. Clin Cancer Res 24:4937–4948. https://doi.org/10.1158/1078-0432.CCR-17-1866
Larkin J (2015) Combined nivolumab and ipilimumab or monotherapy in untreated melanoma. N Engl J Med 373:1270–1271. https://doi.org/10.1056/nejmc1509660
Layek B, Sadhukha T, Panyam J, Prabha S (2018) Nano-engineered mesenchymal stem cells increase therapeutic E_cacy of anticancer drug through true active tumor targeting. Mol Cancer Ther 17:1196–1206
Lee HJ, Doo SW, Kim DH et al (2013) Cytosine deaminase-expressing human neural stem cells inhibit tumor growth in prostate cancer-bearing mice. Cancer Lett 335:58–65. https://doi.org/10.1016/j.canlet.2013.01.048
Lee JM, Lee MH, Garon E et al (2017) Phase I trial of intratumoral injection of CCL21 gene–modified dendritic cells in lung cancer elicits tumor-specific immune responses and CD8+ T-cell infiltration. Clin Cancer Res 23:4556–4568. https://doi.org/10.1158/1078-0432.CCR-16-2821
Lee YG, Marks I, Srinivasarao M et al (2019) Use of a single CAR T cell and several bispecific adapters facilitates eradication of multiple antigenically different solid tumors. Cancer Res 79:387–396. https://doi.org/10.1158/0008-5472.CAN-18-1834
Lee RH, Oh JY, Choi H, Bazhanov N (2011) Therapeutic factors secreted by mesenchymal stromal cells and tissue repair. J Cell Biochem 112:3073–3078. https://doi.org/10.1002/jcb.23250
Lee RH, Pulin A, Seo MJ et al (2009) Intravenous hMSCs improve myocardial infarction in mice because cells embolized in lung are activated to secrete the anti-inflammatory protein TSG-6. Cell Stem Cell 5:54–63
Lee JY, Lee HT, Shin W et al (2016) Structural basis of checkpoint blockade by monoclonal antibodies in cancer immunotherapy. Nat Commun 7. https://doi.org/10.1038/NCOMMS13354
Li Q, Anderson CD, Egilmez NK (2018) Inhaled IL-10 Suppresses Lung Tumorigenesis via Abrogation of Inflammatory Macrophage-Th17 Cell Axis. J Immunol 201:2842–2850. https://doi.org/10.4049/JIMMUNOL.1800141
Li S, Schmitz KR, Jeffrey PD et al (2005) Structural basis for inhibition of the epidermal growth factor receptor by cetuximab. Cancer Cell 7:301–311. https://doi.org/10.1016/j.ccr.2005.03.003
Li L, Zhang Y, Zhou Y et al (2022) Quaternary nanoparticles enable sustained release of bortezomib for hepatocellular carcinoma. Hepatology. https://doi.org/10.1002/hep.32584
Liao W, Lin JX, Leonard WJ (2013) Interleukin-2 at the crossroads of effector responses, tolerance, and immunotherapy. Immunity 38:13–25. https://doi.org/10.1016/J.IMMUNI.2013.01.004
Libutti SK, Paciotti GF, Byrnes AA et al (2010) Phase I and pharmacokinetic studies of CYT-6091, a novel PEGylated colloidal Gold-rhTNF nanomedicine. Clin Cancer Res 16:6139. https://doi.org/10.1158/1078-0432.CCR-10-0978
Lin MJ, Svensson-Arvelund J, Lubitz GS et al (2022) Cancer vaccines: the next immunotherapy frontier. Nat Cancer 3:8 3:911–926. https://doi.org/10.1038/s43018-022-00418-6
Liu H, Guo L, Zhang J et al (2019) Glycosylation-independent binding of monoclonal antibody toripalimab to FG loop of PD-1 for tumor immune checkpoint therapy. Mabs 11:681–690. https://doi.org/10.1080/19420862.2019.1596513
Liu B, Kong L, Han K et al (2016) A novel fusion of ALT-803 (Interleukin (IL)-15 Superagonist) with an antibody demonstrates antigen-specific antitumor responses. J Biol Chem 291:23869. https://doi.org/10.1074/JBC.M116.733600
Liu K, Tan S, Jin W et al (2020) N‐glycosylation of PD‐1 promotes binding of camrelizumab. EMBO Rep 21. https://doi.org/10.15252/EMBR.202051444
Lou G, Song X, Yang F et al (2015) Exosomes derived from miR-122-modifiedadipose tissue-derived MSCs increase chemosensitivity of hepatocellular carcinoma. J Hematol Oncol 122
Lozano E, Dominguez-Villar M, Kuchroo V, Hafler DA (2012) The TIGIT/CD226 axis regulates human T cell function. J Immunol 188:3869–3875. https://doi.org/10.4049/jimmunol.1103627
Létourneau S, van Leeuwen EMM, Krieg C et al (2010) IL-2/anti-IL-2 antibody complexes show strong biological activity by avoiding interaction with IL-2 receptor alpha subunit CD25. Proc Natl Acad Sci U S A 107:2171–2176. https://doi.org/10.1073/PNAS.0909384107
M N’diaye, (2016) Rat bone marrow-derived dendritic cells generated with GM-CSF/IL-4 or FLT3L exhibit distinct phenotypical and functional characteristics. J Leukoc Biol 99:437–446
Ma Y, Dong S, Li X et al (2021) Extracellular vesicles: an emerging nanoplatform for cancer therapy. Front Oncol 10:606906. https://doi.org/10.3389/fonc.2020.606906
Malekshah OM, Chen X, Nomani A et al (2016) Enzyme/prodrug systems for cancer genetherapy. Curr Pharmacol Rep 2:299–308
Maloney DG, Grillo-López AJ, White CA et al (1997) IDEC-C2B8 (rituximab) anti-CD20 monoclonal antibody therapy in patients with relapsed low-grade non-Hodgkin’s lymphoma. Blood 90:2188–2195. https://doi.org/10.1182/blood.v90.6.2188.2188_2188_2195
Margolin K, Morishima C, Velcheti V et al (2018) Phase I trial of Alt-803, A novel recombinant Interleukin-15 complex, in patients with advanced solid tumors. Clin Cancer Res 24:5552. https://doi.org/10.1158/1078-0432.CCR-18-0945
Maslak PG, Dao T, Bernal Y et al (2018) Phase 2 trial of a multivalent WT1 peptide vaccine (galinpepimut-S) in acute myeloid leukemia. Blood Adv 2:224–234. https://doi.org/10.1182/bloodadvances.2017014175
Matsui WH (2016) Cancer stem cell signaling pathways. Medicine (United States) 95:S8–S19. https://doi.org/10.1097/MD.0000000000004765
Mejías R, Pérez-Yagüe S, Gutiérrez L et al (2011) Dimercaptosuccinic acid-coated magnetite nanoparticles for magnetically guided in vivo delivery of interferon gamma for cancer immunotherapy. Biomaterials 32:2938–2952. https://doi.org/10.1016/J.BIOMATERIALS.2011.01.008
Melief CJ, Hall T, Arens R et al (2015) Therapeutic cancer vaccines. J Clin Invest 125:3401–3412
Mezquita L, Planchard D (2018) Durvalumab for the treatment of non-small cell lung cancer. Expert Rev Respir Med 12:627–639. https://doi.org/10.1080/17476348.2018.1494575
de Miguel D, Gallego-Lleyda A, Anel A, Martinez-Lostao L (2015) Liposome-bound TRAIL induces superior DR5 clustering and enhanced DISC recruitment in histiocytic lymphoma U937 cells. Leuk Res 39:657–666. https://doi.org/10.1016/J.LEUKRES.2015.03.019
Milhem M, Zakharia Y, Davar D et al (2020) 304 Intratumoral injection of CMP-001, a toll-like receptor 9 (TLR9) agonist, in combination with pembrolizumab reversed programmed death receptor 1 (PD-1) blockade resistance in advanced melanoma. J Immunother Cancer 8:A186–A187. https://doi.org/10.1136/jitc-2020-sitc2020.0304
Ming Y, Li Y, Xing H et al (2017) Circulating tumor cells: From theory to nanotechnology-based detection. Front Pharmacol 8:35. https://doi.org/10.3389/FPHAR.2017.00035/BIBTEX
Mittendorf EA, Lu B, Melisko M et al (2019) Efficacy and safety analysis of nelipepimut-S vaccine to prevent breast cancer recurrence: a randomized, multicenter, phase III clinical trial. Clin Cancer Res 25:4248–4254. https://doi.org/10.1158/1078-0432.CCR-18-2867
Mittendorf EA (2016) Primary analysis of a prospective, randomized, single-blinded phase II trial evaluating the HER2 peptide AE37 vaccine in breast cancer patients to prevent recurrence. Ann Oncol 27:1241–1248
Moku G, Layek B, Trautman L et al (2019) Improving payload capacity and Anti-Tumor E_cacy of Mesenchymal stem cells using TAT peptide functionalized polymeric nanoparticles. Cancers 11:491
Montagnana M, Lippi G (2017) Cancer diagnostics: current concepts and future perspectives. Ann Transl Med 5:268
Moon EK, Wang LC, Dolfi DV et al (2014) Multifactorial T-cell hypofunction that is reversible can limit the efficacy of chimeric antigen receptor-transduced human T cells in solid tumors. Clin Cancer Res 20:4262–4273. https://doi.org/10.1158/1078-0432.CCR-13-2627
Moretta L, Locatelli F, Pende D et al (2011) Killer Ig-like receptor-mediated control of natural killer cell alloreactivity in haploidentical hematopoietic stem cell transplantation. Blood 117:764–771. https://doi.org/10.1182/blood-2010-08-264085
Moseley P (2000) Stress proteins and the immune response. Immunopharmacology 48:299–302. https://doi.org/10.1016/S0162-3109(00)00227-7
Müller S, Bexte T, Gebel V et al (2020) High cytotoxic efficiency of lentivirally and alpharetrovirally engineered CD19-Specific chimeric antigen receptor natural killer cells against acute Lymphoblastic Leukemia. Front Immunol 10. https://doi.org/10.3389/fimmu.2019.03123
Nagasaka M, Gadgeel SM (2018) Role of chemotherapy and targeted therapy in early-stage non-small cell lung cancer. Expert Rev Anticancer Ther 18:63–70. https://doi.org/10.1080/14737140.2018.1409624
Naing A, Wong DJ, Infante JR et al (2019) Pegilodecakin combined with pembrolizumab or nivolumab for patients with advanced solid tumours (IVY): a multicentre, multicohort, open-label, phase 1b trial. Lancet Oncol 20:1544–1555. https://doi.org/10.1016/S1470-2045(19)30514-5
Neal ZC, Yang JC, Rakhmilevich AL et al (2004) Enhanced activity of hu14.18-IL2 immunocytokine against murine NXS2 neuroblastoma when combined with interleukin 2 therapy. Clin Cancer Res 10:4839–4847. https://doi.org/10.1158/1078-0432.CCR-03-0799
Neelapu SS (2019) CAR-T efficacy: Is conditioning the key? Blood 133:1799–1800. https://doi.org/10.1182/blood-2019-03-900928
Nielsen M, Lundegaard C, Worning P et al (2003) Reliable prediction of T-cell epitopes using neural networks with novel sequence representations. Protein Sci 12:1007–1017. https://doi.org/10.1110/ps.0239403
Ong HT, Federspiel MJ, Guo CM et al (2013) Systemically delivered measles virus-infected mesenchymal stem cells can evade host immunity to inhibit liver cancer growth. J Hepatol 59:999–1006. https://doi.org/10.1016/j.jhep.2013.07.010
Ott PA (2017) An immunogenic personal neoantigen vaccine for patients with melanoma. Nature 547:217–221
Ott PA, Hu-Lieskovan S, Chmielowski B et al (2020) A phase Ib trial of personalized neoantigen therapy plus anti-PD-1 in Patients with advanced melanoma, non-small cell lung cancer, or bladder cancer. Cell 183:347-362.e24. https://doi.org/10.1016/j.cell.2020.08.053
Ouyang X, Telli ML, Wu JC (2019) Induced pluripotent stem cell-based cancer vaccines. Front Immunol 10. https://doi.org/10.3389/fimmu.2019.01510
Overwijk WW, Restifo NP (2001) B16 as a Mouse Model for Human Melanoma. In: Coligan JE et al (eds) Current protocols in immunology. CHAPTER: Unit. https://doi.org/10.1002/0471142735.IM2001S39
O’Brien C, Kreso A, Jamieson CHM (2010) Cancer stem cells and self-renewal. Clin Cancer Res 16:3113–3120
O’Rourke DM A single dose of peripherally infused EGFRvIIIdirected CAR T cells mediates antigen loss and induces adaptive resistance in patients with recurrent glioblastoma. Sci Transl Med 9:984
Parisi G, Saco JD, Salazar FB et al (2020) Persistence of adoptively transferred T cells with a kinetically engineered IL-2 receptor agonist. Nat Commun 11:1 11:1–12. https://doi.org/10.1038/s41467-019-12901-3
Patel D, Bassi R, Hooper A et al (2009) Anti-epidermal growth factor receptor monoclonal antibody cetuximab inhibits EGFR/HER-2 heterodimerization and activation. Int J Oncol 34:25–32. https://doi.org/10.3892/ijo_00000125
Perdue LA, Do P, David C et al (2020) Optical control of cytokine signaling via bioinspired, polymer-Induced latency. Biomacromol 21:2635–2644. https://doi.org/10.1021/ACS.BIOMAC.0C00264/SUPPL_FILE/BM0C00264_SI_001.PDF
Postow MA (2015) Nivolumab and ipilimumab versus ipilimumab in untreated melanoma. N Engl J Med 372:2006–2017
Qiao J, Liu Z, Fu YX (2016) Adapting conventional cancer treatment for immunotherapy. J Mol Med 94:489–495. https://doi.org/10.1007/s00109-016-1393-4
Quezada SA, Peggs KS, Curran MA, Allison JP (2006) CTLA4 blockade and GM-CSF combination immunotherapy alters the intratumor balance of effector and regulatory T cells. J Clin Investig 116:1935–1945. https://doi.org/10.1172/JCI27745
Ramagopal UA, Liu W, Garrett-Thomson SC et al (2017) Structural basis for cancer immunotherapy by the first-in-class checkpoint inhibitor ipilimumab. Proc Natl Acad Sci U S A 114:E4223–E4232. https://doi.org/10.1073/PNAS.1617941114
Reda M, Ngamcherdtrakul W, Nelson MA et al (2022) Development of a nanoparticle-based immunotherapy targeting PD-L1 and PLK1 for lung cancer treatment. Nat Commun 13:4261. https://doi.org/10.1038/s41467-022-31926-9
Reindl LM, Albinger N, Bexte T et al (2020) Immunotherapy with NK cells: recent developments in gene modification open up new avenues. Oncoimmunology 9:1–12. https://doi.org/10.1080/2162402X.2020.1777651
Roger M, Clavreul A, Venier-Julienne MC et al (2010) Mesenchymal stem cells as cellular vehicles for delivery of nanoparticles to brain tumors. Biomaterials 31:8393–8401. https://doi.org/10.1016/j.biomaterials.2010.07.048
Rosario M, Liu B, Kong L et al (2014) The IL-15 superagonist ALT-803 enhances anti-CD20 antibody-directed NK cell ADCC and in vivo clearance of B cell lymphomas. J Immunother Cancer 2:P168. https://doi.org/10.1186/2051-1426-2-S3-P168
Rosenberg SA, Lotze MT, Yang JC et al (1989) Experience with the use of high-dose interleukin-2 in the treatment of 652 cancer patients. Ann Surg 210:474–485. https://doi.org/10.1097/00000658-198910000-00008
Rosenberg SA, Yang JC, White DE, Steinberg SM (1998) Durability of complete responses in patients with metastatic cancer treated with high-dose interleukin-2: identification of the antigens mediating response. Ann Surg 228:307. https://doi.org/10.1097/00000658-199809000-00004
Rosenblatt J, Avigan D (2017) Targeting the PD-1/PD-L1 axis in multiple myeloma: a dream or a reality? Blood 129:275–279. https://doi.org/10.1182/BLOOD-2016-08-731885
Rosenblum D, Joshi N, Tao W et al (2018) Progress and challenges towards targeted delivery of cancer therapeutics. Nat Commun 9:9. https://doi.org/10.1038/s41467-018-03705-y
Rubnitz JE, Inaba H, Ribeiro RC et al (2010) NKAML: a pilot study to determine the safety and feasibility of haploidentical natural killer cell transplantation in childhood acute myeloid leukemia. J Clin Oncol 28:955–959. https://doi.org/10.1200/JCO.2009.24.4590
Sage EK, Thakrar RM, Janes SM (2016) Genetically modified mesenchymal stromal cells in cancer therapy. Cytotherapy 18:1435–1445
Sahin U Personalized RNA mutanome vaccines mobilize polyspecific therapeutic immunity against cancer. Nature 547:222–226
Sato H, Okonogi N, Nakano T (2020) Rationale of combination of anti-PD-1/PD-L1 antibody therapy and radiotherapy for cancer treatment. Int J Clin Oncol 25:801–809. https://doi.org/10.1007/s10147-020-01666-1
Schuster J, Lai RK, Recht LD et al (2015) A phase II, multicenter trial of rindopepimut (CDX-110) in newly diagnosed glioblastoma: the ACT III study. Neuro Oncol 17:854–861. https://doi.org/10.1093/neuonc/nou348
Schwartz JCD, Zhang X, Fedorov AA et al (2001) Structural basis for co-stimulation by the human CTLA-4/B7-2 complex. Nature 410:604–608. https://doi.org/10.1038/35069112
Sharon E (2014) Immune checkpoint inhibitors in clinical trials. Chin J Cancer 33:434–444
Shusterman S, Naranjo A, van Ryn C et al (2019) Antitumor activity and tolerability of hu14.18-IL2 with GMCSF and isotretinoin in recurrent or refractory neuroblastoma: a children’s oncology group phase II study. Clin Cancer Res 25:6044–6051. https://doi.org/10.1158/1078-0432.CCR-19-0798
Siamof CM, Goel S, Cai W (2020) Moving beyond the pillars of cancer treatment: perspectives from nanotechnology. Front Chem 8:598100. https://doi.org/10.3389/fchem.2020.598100
Siegel JP, Puri RK (1991) Interleukin-2 toxicity. J Clin Oncol 9:694–704. https://doi.org/10.1200/JCO.1991.9.4.694
Sleijfer S, Bannink M, van Gool AR et al (2005) Side effects of interferon-alpha therapy. Pharm World Sci 27:423–431. https://doi.org/10.1007/S11096-005-1319-7
Smyth T, Petrova K, Payton NM et al (2014) Surface functionalization of exosomes using click chemistry. Bioconjug Chem 25:1777–1784. https://doi.org/10.1021/bc500291r
Sockolosky JT, Trotta E, Parisi G et al (2018) Selective targeting of engineered T cells using orthogonal IL-2 cytokine-receptor complexes. Science 359:1037–1042. https://doi.org/10.1126/SCIENCE.AAR3246
Soria JC, Márk Z, Zatloukal P et al (2011) Randomized phase II study of dulanermin in combination with paclitaxel, carboplatin, and bevacizumab in advanced non-small-cell lung cancer. J Clin Oncol 29:4442–4451. https://doi.org/10.1200/JCO.2011.37.2623
Subbiah V, Murthy R, Hong DS et al (2018) Cytokines produced by dendritic cells administered intratumorally correlate with clinical outcome in patients with diverse cancers. Clin Cancer Res 24:3845–3856. https://doi.org/10.1158/1078-0432.CCR-17-2707
Sun W, Jiang T, Lu Y et al (2014) Cocoon-like self-degradable DNA nanoclew for anticancer drug delivery. J Am Chem Soc 136:14722–14725. https://doi.org/10.1021/JA5088024/SUPPL_FILE/JA5088024_SI_001.PDF
Sun JY, Zhang D, Wu S et al (2020) Resistance to PD-1/PD-L1 blockade cancer immunotherapy: mechanisms, predictive factors, and future perspectives. Biomark Res 8:1–10. https://doi.org/10.1186/S40364-020-00212-5/FIGURES/1
Sun Z, Ren Z, Yang K et al (2019) A next-generation tumor-targeting IL-2 preferentially promotes tumor-infiltrating CD8+ T-cell response and effective tumor control. Nat Commun 10:1 10:1–12. https://doi.org/10.1038/s41467-019-11782-w
Sławiński G, Wrona A, Dabrowska-Kugacka A et al (2020) Immune checkpoint inhibitors and cardiac toxicity in patients treated for non-small lung cancer: a review. Int J Mol Sci 21:1–19. https://doi.org/10.3390/IJMS21197195
Takahashi K, Yamanaka S (2006) Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126:663–676. https://doi.org/10.1016/j.cell.2006.07.024
Tang S, Kim PS (2019) A high-affinity human PD-1/PD-L2 complex informs avenues for small-molecule immune checkpoint drug discovery. Proc Natl Acad Sci U S A 116:24500–24506. https://doi.org/10.1073/PNAS.1916916116
Taylor GS, Jia H, Harrington K et al (2014) A recombinant modified vaccinia ankara vaccine encoding Epstein-Barr Virus (EBV) target antigens: a phase I trial in UK patients with EBV-positive cancer. Clin Cancer Res 20:5009–5022. https://doi.org/10.1158/1078-0432.CCR-14-1122-T
Testori A, Richards J, Whitman E et al (2008) Phase III comparison of vitespen, an autologous tumor-derived heat shock protein gp96 peptide complex vaccine, with physician’s choice of treatment for stage IV melanoma: The C-100-21 study group. J Clin Oncol 26:955–962. https://doi.org/10.1200/JCO.2007.11.9941
The growth of brain tumors can be suppressed by multiple transplantation of mesenchymal stem cellsexpressing cytosine deaminase. Int J Cancer 127:1975–1983
Tobias AL, Thaci B, Auffinger B et al (2013) The timing of neural stem cell-based virotherapy is critical for optimal therapeutic efficacy when applied with radiation and chemotherapy for the treatment of glioblastoma. Stem Cells Transl Med 2:655–666. https://doi.org/10.5966/sctm.2013-0039
Toledo-Guzmán ME, Bigoni-Ordóñez GD, Hernández MI, Ortiz-Sánchez E (2018) Cancer stem cell impact on clinical oncology. World J Stem Cells 10:183–195. https://doi.org/10.4252/wjsc.v10.i12.183
Torre AN (2017) A phase I trial using local regional treatment, nonlethal irradiation, intratumoral and systemic polyinosinic-polycytidylic acid polylysine carboxymethylcellulose to treat liver cancer: in search of the abscopal effect. J Hepatocell Carcinoma 4:111–121
Trinchieri G (2007) Interleukin-10 production by effector T cells: Th1 cells show self control. J Exp Med 204:239. https://doi.org/10.1084/JEM.20070104
Tsai YJ, Lin JK, Chen WS et al (2016) Adjuvant FOLFOX treatment for stage III colon cancer: how many cycles are enough? Springerplus 5:1318. https://doi.org/10.1186/S40064-016-2976-9
Tsao SW, Tramoutanis G, Dawson CW et al (2002) The significance of LMP1 expression in nasopharyngeal carcinoma. Semin Cancer Biol 12:473–487. https://doi.org/10.1016/S1044579X02000901
U.S BMS Food and Drug Administration Approves Bristol Myers Squibb’s Breyanzi (lisocabtagene maraleucel), a New CAR T Cell Therapy for Adults with Relapsed or Refractory Large B-cell Lymphoma
U.S GSI FDA Approves Kite’s TecartusTM, the first and only CAR T treatment for relapsed or refractory mantle cell lymphoma
Uricoli B, Birnbaum LA, Do P et al (2021) Engineered cytokines for cancer and autoimmune disease immunotherapy. Adv Healthc Mater 10:2002214. https://doi.org/10.1002/ADHM.202002214
Vansteenkiste J, Zielinski M, Linder A et al (2013) Adjuvant MAGE-A3 immunotherapy in resected non–small-cell lung cancer: phase II randomized study results. J Clin Oncol 31:2396–2403. https://doi.org/10.1200/JCO.2012.43.7103
Villa A, Trachsel E, Kaspar M et al (2008) A high-affinity human monoclonal antibody specific to the alternatively spliced EDA domain of fibronectin efficiently targets tumor neo-vasculature in vivo. Int J Cancer 122:2405–2413. https://doi.org/10.1002/IJC.23408
Wang J, Li W, Zhang L et al (2017) Chemically edited exosomes with dual ligand purified by microfluidic device for active targeted drug delivery to tumor cells. ACS Appl Mater Interfaces 9:27441–27452. https://doi.org/10.1021/acsami.7b06464
Wang L, Zhang D, Ren Y et al (2022) Injectable hyaluronic acid hydrogel loaded with BMSC and NGF for traumatic brain injury treatment. Mater Today Bio 13:100201. https://doi.org/10.1016/J.MTBIO.2021.100201
Wang J, Xu B (n.d.) Targeted therapeutic options and future perspectives for HER2-positive breast cancer. Signal Transduct Target Ther 4:1–22
Wang W, Liu JR, Zou W (2019a) Immunotherapy in ovarian cancer. Surg Oncol Clin N Am 28:447. https://doi.org/10.1016/J.SOC.2019.02.002
Wang X, Chen H, Zeng X et al (2019b) Efficient lung cancer-targeted drug delivery via a nanoparticle/MSC system. Acta Pharm Sin B 9:167–176
Weller M (2017) Rindopepimut with temozolomide for patients with newly diagnosed, EGFRvIII-expressing glioblastoma (ACT IV): a randomised, double-blind, international phase 3 trial. Lancet Oncol 18:1373–1385
Wiklander OPB, Brennan M, Lötvall J et al (2019) Advances in therapeutic applications of extracellular vesicles. Sci Transl Med 11:8521. https://doi.org/10.1126/scitranslmed.aav8521
Wood C, Srivastava P, Bukowski R et al (2008) An adjuvant autologous therapeutic vaccine (HSPPC-96; vitespen) versus observation alone for patients at high risk of recurrence after nephrectomy for renal cell carcinoma: a multicentre, open-label, randomised phase III trial. The Lancet 372:145–154. https://doi.org/10.1016/S0140-6736(08)60697-2
Wrangle JM, Velcheti V, Patel MR et al (2018) ALT-803, an IL-15 superagonist, in combination with nivolumab in patients with metastatic non-small cell lung cancer: a non-randomised, open-label, phase 1b trial. Lancet Oncol 19:694–704. https://doi.org/10.1016/S1470-2045(18)30148-7
Wu X, Wu Y, Ye H et al (2017) Interleukin-15 and cisplatin co-encapsulated thermosensitive polypeptide hydrogels for combined immuno-chemotherapy. J Control Release 255:81–93. https://doi.org/10.1016/J.JCONREL.2017.04.011
Wyatt Shields C, Evans MA, Wang LLW et al (2020) Cellular backpacks for macrophage immunotherapy. Sci Adv 6. https://doi.org/10.1126/SCIADV.AAZ6579/SUPPL_FILE/AAZ6579_SM.PDF
Xu W, Jones M, Liu B et al (2013) Efficacy and mechanism-of-action of a novel superagonist interleukin-15: interleukin-15 receptor αSu/Fc fusion complex in syngeneic murine models of multiple myeloma. Cancer Res 73:3075–3086. https://doi.org/10.1158/0008-5472.CAN-12-2357
Xu S, Xu H, Wang W et al (2019) The role of collagen in cancer: from bench to bedside. J Transl Med 17:1 17:1–22. https://doi.org/10.1186/S12967-019-2058-1
Yang P, Peng Y, Feng Y et al (2021) Immune cell-derived extracellular vesicles—new strategies in cancer immunotherapy. Front Immunol 12(12):771551. https://doi.org/10.3389/fimmu.2021.771551
Yoon SR, Lee YS, Yang SH, Ahn KH et al. (2010) Generation of donor natural killer cells from CD34(+) progenitor cells and subsequent infusion after HLA-mismatched allogeneic hematopoietic cell transplantation: a feasibility study. Bone Marrow Transplant 45:1038–1046
You F, Wang Y, Jiang L et al (2019) A novel CD7 chimeric antigen receptor-modified NK-92MI cell line targeting T-cell acute lymphoblastic leukemia. Am J Cancer Res 9:64–78
Yuan M, Huang LL, Chen JH et al (2019) The emerging treatment landscape of targeted therapy in non-small-cell lung cancer. Signal Transduct Target Ther 4. https://doi.org/10.1038/S41392-019-0099-9
Zahavi D, Weiner L (2020) Monoclonal antibodies in cancer therapy. Antibodies 9:1–20. https://doi.org/10.3390/antib9030034
Zhao J, Jiang L, Yang H et al (2022) A strategy for the efficient construction of anti-PD1-based bispecific antibodies with desired IgG-like properties. MAbs 14. https://doi.org/10.1080/19420862.2022.2044435
Zhylko A, Winiarska M, Graczyk-Jarzynka A (2020) The great war of today: modifications of car-t cells to effectively combat malignancies. Cancers (Basel) 12:1–29. https://doi.org/10.3390/cancers12082030
van Slooten ML, Storm G, Zoephel A et al (2000) Liposomes containing interferon-gamma as adjuvant in tumor cell vaccines. Pharm Res 17:1 17:42–48. https://doi.org/10.1023/A:1007514424253
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
George, A., Varghese, J., Padinharayil, H. (2023). Potential of Biotechnology in Cancer Management. In: Kulkarni, S., Haghi, A.K., Manwatkar, S. (eds) Novel Technologies in Biosystems, Biomedical & Drug Delivery. Springer, Singapore. https://doi.org/10.1007/978-981-99-5281-6_2
Download citation
DOI: https://doi.org/10.1007/978-981-99-5281-6_2
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-99-5280-9
Online ISBN: 978-981-99-5281-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)