
Abstract
The use of microbial consortia and biofertilizers has emerged as a promising approach for enhancing the yield and quality of essential oils in aromatic plants. This strategy leverages the benefits of microorganisms such as arbuscular mycorrhizal fungi (AMF) and plant growth-promoting rhizobacteria (PGPRs) to improve plant growth and the production of secondary metabolites. However, achieving optimal results also requires consideration of environmental factors such as light, temperature, humidity, and soil fertility, as well as appropriate cultivation techniques. Innovative systems such as aeroponic and hydroponic systems, as well as micropropagation, can provide even more favorable growth conditions for medicinal plants, resulting in the production of high-quality bioactive substances. Implementation of this approach has significant implications for the pharmaceutical and herbal industries, as well as for overall health and well-being. By leveraging the benefits of microbial consortia and biofertilizers, we can enhance the sustainability and productivity of agricultural practices while also promoting a healthier and more natural approach to medicine.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
9.1 Introduction
Throughout history, medicinal plants have been an important source of therapeutic agents for treating various ailments and diseases, including diarrhea, fever, colds, and malaria. This has been highlighted by several studies, such as Dambisya and Tindimwebwa (2003); Ghiaee et al. (2014), Mathens and Bellanger (2010), and Titanji et al. (2008). The increase in population and the consequent rise in human needs to meet their needs has caused humans to exploit resources and facilities during the last century with the help of new technologies and relying on low-cost natural resources. The world’s growing population will lead to several advancements, resulting in increase in food production during recent decades. Meanwhile, plant nutrition plays a key role in significantly increasing food supply (Mukhopadhyay et al. 2021). Increasing plant production has become possible through the use of man-made commercial fertilizers. Sustainable agriculture is a system that, while enjoying economic dynamism, can improve the environment and optimal use of available resources, and also play a significant role in providing human food needs and improving the quality of life of human societies. In addition, sustainable agriculture by observing the principles of ecology can increase the efficiency of resource use while creating a balance in the environment and provide a basis for productivity for a longer period of time (Polukhin and Panarina 2022). In sustainable agriculture, in addition to emphasizing on reducing the consumption of synthetic chemicals and pesticides, crop rotation, organic fertilizers, plant residues and agricultural wastes are used as a substitute for some of the chemical fertilizers, and the use of fertilizers is also optimal according to the plant’s needs and production potential (Mukhopadhyay et al. 2021). In this way, the amount of damage or negative effects for natural resources and the environment will be reduced to a minimum. Long-term studies show that excessive use of chemical fertilizers reduces the performance of agricultural and medicinal plants. This reduction is due to soil acidification, reduction of soil biological activities, loss of soil physical properties, and absence of micronutrients in K, P, N fertilizers. In many cases, the use of chemical fertilizers causes environmental pollution and ecological damage, which also increases the cost of production. In order to reduce these risks, resources and institutions should be used that, in addition to providing the current needs of the plant, also ensure the sustainability of agricultural systems in the long term.
By producing humus, organic fertilizers reduce the adverse effects of chemical fertilizers and increase the efficiency of fertilizer use (Khosropour et al. 2023). Sustainable agriculture in the form of combined use of chemical and organic fertilizers as a suitable solution for the development of sustainable agriculture during the transition period from conventional agriculture to sustainable agriculture is effective for producing agricultural products and maintaining plant performance at an acceptable level (Khosropour et al. 2023). Organic materials are one of the important elements of soil fertility due to their beneficial effects on the physical, chemical, biological, and fertility properties of the soil. Organic fertilizers increase the organic matter and pH of the soil, and due to the improvement of the chemical properties of the soil, such as cation exchange capacity and increasing the activity of microorganisms and the amount of access to nutrients, they increase the fertility of the soil (Mukhopadhyay et al. 2021).
Plants in nature are associated with a vast number of beneficial microorganisms such as endophytic or symbiotic bacteria and fungi. These microorganisms have a crucial role in maintaining the health, development, and productivity of plants, and also play a significant part in regulating the synthesis of various metabolites. The importance of this relationship has been emphasized in several studies, including Ezzati Lotfabadi et al. (2021), Panke-Buisse et al. (2015), Castrillo et al. (2017), de Vries et al. (2020), Brader et al. (2014), and Compant et al. (2021).
Bioactive compounds, including carotenoids, essential oils, antioxidants, and flavors, are often incorporated into food products to enhance their flavor, aroma, texture, and overall appeal while also improving their nutritional and health benefits. Recent research has suggested that the inoculation of arbuscular mycorrhizal fungi (AMF) can help optimize the production of these important biomolecules (Hazzoumi et al. 2015; Oliveira et al. 2015). The symbiotic relationship between plants and AMF has been shown to boost the production of secondary compounds, which can increase the value of the plant’s phytomass and its potential medicinal properties (Oliveira et al. 2013; Zitter-Eglseer et al. 2015).
This chapter aims to provide an overview of the current knowledge on the inoculation of AMF and PGPRs, their mechanisms, and their effects on the production of essential oils and bioactive compounds in aromatic plants. Additionally, this chapter highlights the benefits of using microbial consortia and biofertilizer management for enhancing the yield and quality of essential oils. Finally, the chapter identifies areas where further research is needed to gain a better understanding of the microbial communities and their mechanisms.
9.2 Biofertilizers
Fertilizers are materials that are used to achieve maximum production per unit area. However, the use of fertilizers should be able to improve the quality of agricultural products in addition to increasing production, and while not polluting the environment, it should reduce the accumulation of nitrate pollutants in the consuming organs of crops to the minimum possible, so as to ensure the health of humans and animals (Kumar et al. 2022). In the last few decades, the use of chemical inputs in agricultural lands has caused many environmental problems, including the pollution of water sources, the decline in the quality of agricultural products and food, and the reduction of soil fertility. Based on this, paying special attention to agriculture from the point of view of sustainability in order to preserve the environment and meet the needs of society in terms of food is necessary and unavoidable, and paying attention to alternative systems has recently attracted a large number of studies (Fasusi et al. 2021; Kumar et al. 2022).
Today, with the increase in agricultural and horticultural production to meet the growing needs of the expanding population, concern has been raised about the future of providing food and green space for the people (Mahapatra et al. 2022). Pollution of water, soil, air, and soil erosion, resistance of pests to poisons, and spread of chemical fertilizers caused us to go back to the past and industrial crops in order to preserve resources. So, to produce healthy and clean products and as a result, healthy and cheerful people, we have no other way than biological agriculture (Dehsheikh et al. 2020). Considering the increasing demand for the consumption of biological agricultural products, which is based on the proper management of the soil and the growth environment of plants and trees, it is done in such a way that the balance between the required elements in the soil is not disturbed in the nutrition of plants and trees. During growth, there is no need to use poisons and pesticides, and instead of using chemical fertilizers, natural fertilizers such as leaf soil, algae, and animal and biological fertilizers should be used to feed the agricultural soil (Fasusi et al. 2021; Gurikar et al. 2022). If there is a need to fight against pests, instead of using chemical pesticides and poisons, biological methods such as effective microorganisms, ladybugs, bees and bacteria, or pest-resistant cultivars are used in cultivation, and in this type of agriculture, modified seeds are used (Kumar et al. 2022). Genetically modified and exposed to radiation is not used. From this point of view, the final product that reaches the consumer will be free of toxic and chemical residues and preservatives. On the other hand, quality food products, which are the product of biofertilizers, not only satisfy consumers, but also provide and guarantee their physical health (Gurikar et al. 2022). Advantages of using biological fertilizer are as follows:
-
1.
Increasing the amount of nutrients and soil organic matter,
-
2.
Reducing the need to use chemical fertilizers,
-
3.
Increasing the absorption power of food by plants,
-
4.
Prevent soil erosion,
-
5.
The sulfur in the fertilizer improves soil salinity (Chojnacka et al. 2020).
Classification of biofertilizers according to the type of microorganisms:
-
1.
Bacterial biofertilizers (rhizobium-azotobacter-azospirlium),
-
2.
Fungal biofertilizers (mycorrhiza),
-
3.
Algal biofertilizers (blue-green algae and Azolla),
-
4.
Actinomycetes biofertilizers (Frankia).
9.3 Arbuscular Mycorrhizal Fungi (AMF)
One of the most intricate ecosystems on earth is the soil. Fungus is an essential biotic element of the soil that also keeps the plant-soil healthy. Fungi and vascular plants may cooperate to improve soil quality, according to paleontological and evolutionary evidence. Paleobotanists have demonstrated that these symbiotic connections before the development of terrestrial plants. They evolved alongside arbuscules in the Devonian period (400 Mya), hyphae and spores in the Ordovician period (460 Mya), and glomeromycota in the Ediacaran period (600 Mya). Primary associations in the soil environment are vesicular-arbuscular mycorrhizae (VAM), ectomycorrhizae (ECM), orchid mycorrhizae, ericoid mycorrhizae, ectendomycorrhizae (arbutoid), centianoid mycorrhizae, and monotropoid mycorrhiza. The endomycorrhizal association, one of the seven major types of mycorrhizae, is the most widespread and may be found in all kinds of soil (Gujre et al. 2021). VAM are known as arbuscular mycorrhizal fungi, or AMF, because they produce arbuscules and vesicles. AMF assists in nutrient balancing and enhances the efficiency of the soil system. It reproduces by means of small, multinucleate, asexual spores, the quantity of which is greatly enhanced by the mycelium. With the aid of AMF diversity, biodiversity, ecosystem variability, and productivity can all be preserved. Over 85% of plant families worldwide, across all sorts of environments, are colonized by mycorrhiza. Although they are not limited to it and can extend to 70 cm or even more, AMF typically appear in the topsoil zone of 0–20 cm (Sangwan and Prasanna 2022).
The population of AMF is at its peak in the spring, and the soil has an average (Bhardwaj et al. 2023). The diverse agricultural systems and related practices control the population of AMF, which are detritivores closely linked to organic carbon. Nonetheless, there is a notable rise in the AMF population with crops like sorghum, chickpeas, and maize, but oats, barley, and wheat only develop slowly. By destroying the soil aggregates and upsetting the mycelia network, practices like tillage, crop rotation, residual crop burning, inadequate drainage, fungicide sprays, and waterlogging have a detrimental impact on AMF. AMF can trap a considerable quantity of carbon through its exudations and turnover rates. Around the roots, a dense network of fungi creates an advantageous soil structure with a binding effect. Abiotic stress like heavy metals and water stress can be reduced by the fungus network inside the soil. With the help of the viscous glycoprotein glomalin, AMF promotes soil aggregation in the pedosphere. Its color is reddish-brown, and it has a powerful reinforcing impact that improves the general soil quality. Moreover, the main element of AMF called glomalin interacts with ions in the rhizosphere to increase metal binding.
9.4 Essential Oils
Due to rising consumer demand and interest in these plants for therapeutic purposes, the appeal of aromatic and medicinal plants continues to expand applications in the culinary and other human-made fields. Consumers are becoming more aware of the possibilities and advantages of aromatic and medicinal plants and their metabolites as they become more informed about issues related to food, nutrition, and health (Hanif et al. 2019). These plants generate a variety of secondary metabolites, including essential oils (EOs). EOs have a highly complex chemical makeup. EO constituents individually have beneficial uses in a variety of industries, including agriculture, the environment, and human health. EOs are effective complements to synthetic compounds which are widely utilized in the chemical industry. The term “essential oil” dates back to the sixteenth century and derives from the drug Quinta Essentia, named by Paracelsus von Hohenheim of Switzerland (Brenner 1993). EOs are the substances obtained from a vegetable raw material, either by steam distillation or by mechanical processes from the epicarp of citrus, or “dry”distillation (Hanif et al. 2019). Inorganic solvents like water do not dissolve EOs; however, organic solvents do (ether, alcohol, and fixed oils). With the exception of vetiver, sassafras, and cinnamon, they are volatile liquids with a distinct smell and a density below unity. They are widely utilized in the cosmetics, aromatherapy, and perfume industries (Falleh et al. 2020). Aromatherapy is a therapeutic treatment that uses essential oils in baths, massages, and inhalations (volatile oils). A plant can govern and regulate its surroundings (play an ecological function) by using EOs, which can operate as chemical messages to deter predators, draw pollinating insects, prevent seed germination, and communicate with other plants. Moreover, EOs have insecticidal, deterring, and antifungal properties (Hanif et al. 2019).
9.5 The Effects of AMF on Essential Oil
The positive role of AMF in improving EO content and EO profile of various plants species has been addressed. The AMF can modify the biosynthesis pathway of EO production in plants and lead to significant changes in yield and compounds. The improvement of EO content by AMF has been presented in Table 9.1. In this regard, the improvement of EO content has been reported on Mentha piperita (Zare et al. 2023), Lavandula angustifolia (Pirsarandib et al. 2022), and Cymbopogon citratus (de Souza et al. 2022; Eke et al. 2020). Khalediyan et al. (2021) has shown that MF and PGPR can increase linalol, methyl chavicol, trans-geraniol, camphor, and limonene concentrations in basil EO and carvacrol, thymol, p-cymene, α-terpinene, and γ-terpinene in satureja EO components.
Several medicinal plants, including Chlorophytum borivilianum, Dioscorea spp., Gymnema sylvestre, Glycyrrhiza uralensis, Libidibia ferrea, Ocimum basilicum, Satureja macrostema, and Salvia miltiorrhiza, have shown a positive correlation between the biomass of arbuscular mycorrhizal fungi (AMF)-colonized plants and the concentration of secondary metabolites. Studies by Dave et al. (2011), Lu et al. (2015), Zimare et al. (2013), Chen et al. (2017), Silvia et al. (2014), Zolfaghari et al. (2013), Carreón-Abud et al. (2015), and Yang et al. (2017) have highlighted this relationship. However, for Cynara cardunculus colonized by R. intraradices and F. mosseae, a significant increase in yield was observed, but the concentrations of phenolics decreased (Colonna et al. 2016).
AMF can be beneficial for plants in various ways, including improving their nutrient uptake and indirectly affecting the concentration of secondary metabolites. For example, when Glycyrrhiza uralensis was grown under nutrient-deficient conditions, F. mosseae helped to increase its shoot and root biomass, root system architecture, and flavonoid accumulation (Chen et al. 2017). Furthermore, AMF can help micropropagated medicinal plants to survive and grow better when transferred from in vitro to ex vivo conditions. Studies have shown that when Spilanthes acmella and Glycyrrhiza glabra plantlets were inoculated with F. mosseae, their survival rate was 100% and they exhibited improved growth and development in greenhouse and glasshouse conditions, whereas without AMF, the survival rate was only 60–70% (Yadav et al. 2012, 2013). Similarly, Scutelleria integrifolia seedlings inoculated with C. etunicatum had significantly increased height and fresh weight of shoots, roots, and seeds following micropropagation (Joshee et al. 2007).
According to Zeng et al. (2013), it is commonly believed that the increased levels of various secondary metabolites found in AMF-colonized plants are a result of several defense response pathways that are triggered. These pathways include the carotenoid pathway, the phenylpropanoid pathway, and alkaloid synthesis, which are known to promote signaling, stress tolerance, nutrient uptake, and resistance against biotic and abiotic stresses (Kaur and Suseela 2020). Despite this, the exact mechanisms through which AMF induce changes in the concentration of phytochemicals in plant tissues are not yet fully understood (Toussaint et al. 2007).
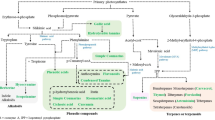
The production of terpenoids, phenolic compounds, and alkaloids in plants and how they are influenced by AMF has been the subject of many studies. Terpenoids, which are synthesized from isoprene units, are produced through two distinct pathways: the mevalonic acid (MVA) pathway and the methyleritrophosphate (MEP) pathway (Zhi et al. 2007). Various nutritional and non-nutritional factors have been suggested to account for the higher production of secondary metabolites in plants colonized by AMF, as outlined in Fig. 9.1, in studies by Kapoor et al. (2017), Sharma et al. (2017), and Dos Santos et al. (2021).

The production of secondary metabolites, such as terpenoids, phenolics, and flavonoids, is affected by both nutritional and non-nutritional factors in plants that have been colonized by AMF and PGPRs
AMF and PGPRs can impact the production of secondary metabolites in medicinal plants through both nutritional and non-nutritional means. Nutritional factors such as the availability of essential nutrients and micronutrients can influence the growth and secondary metabolite production in plants. AMF can enhance nutrient uptake in plants, resulting in increased secondary metabolite production. Similarly, PGPRs can promote plant growth and improve nutrient uptake by fixing atmospheric nitrogen, solubilizing phosphorus, and producing plant growth-promoting hormones.
Non-nutritional factors such as light, temperature, humidity, and soil pH can also influence secondary metabolite production in plants by altering gene expression and enzyme activity. High light intensity, for instance, can stimulate the production of certain secondary metabolites, while low humidity can decrease plant growth and secondary metabolite production. In addition, stress conditions like drought or pathogen attack can induce the production of specific secondary metabolites in plants as a defense mechanism. To optimize the production of secondary metabolites in medicinal plants using AMF and PGPRs, it is important to consider both nutritional and non-nutritional factors. By understanding and manipulating these factors, we can potentially increase the yield and quality of bioactive compounds in medicinal plants, which can have significant implications for the pharmaceutical and herbal industries.
Initially, the increase in production of secondary metabolites in AMF-colonized plants was believed to be due to improved nutrient uptake (Lima et al. 2015; Oliveira et al. 2015) as a nutritional factor. For example, phosphorus is essential for the synthesis of terpenoid precursors via the MVA and MEP pathways, by increasing the concentration of high-energy pyrophosphate compounds like IPP and DMAPP (Kapoor et al. 2002, 2004; Zubek et al. 2010). However, Khaosaad et al. (2006) discovered that the concentration of essential oils significantly increased in two Origanum sp. genotypes colonized by F. mosseae, while the levels of essential oils remained unchanged in plants treated with P. This suggests that the elevated production of essential oils in AMF-colonized Origanum sp. plants may depend directly on the association with the fungus rather than just improved nutrient uptake as a nutritional factor.
9.6 Plant Growth-Promoting Rhizobacteria
Plant-microbe coevolution has led to some of the bacteria becoming facultative intracellular endophytes (Bulgarelli et al. 2013). Among these free-living bacteria are PGPRs that exert beneficial effects on plants through direct and indirect mechanisms. Beneficial rhizobacteria have been utilized to improve water and nutrient uptake, abiotic and biotic stress tolerance. Even though numerous soil bacteria have been reported to promote plant growth and development, the mode(s) of action by which the bacteria exhibit beneficial activities are often not well understood. The molecular basis of plant-bacteria interaction mechanisms responsible for the physiological changes are beginning to be discerned, mainly due to the emerging “omics” approaches. PGPRs can affect the physiological and biochemical attributes in plants. The changes in secondary metabolite like essential oils is eminent in the interaction of plants and PGPRs. Table 9.1 shows the positive effect of PGPRs in improving essential oil quality and quantity.
9.6.1 Azotobacter
PGPR helps to replace chemical fertilizer for the sustainable agriculture production by fixing the atmospheric nitrogen and producing growth-promoting substances. Among the PGPR group, Azotobacter are ubiquitous, aerobic, free-living, and N2-fixing bacteria commonly living in soil, water, and sediments. Being the major group of soilborne bacteria, Azotobacter plays different beneficial roles and is known to produce varieties of vitamins, amino acids, plant growth hormones, antifungal substances, hydrogen cyanide, and siderophores. The growth-promoting substances, such as indoleacetic acid, gibberellic acid, and arginine, produced by Azotobacter have direct influence on shoot and root length as well as seed germination of several agricultural crops. Azotobacter species are efficient in fixation of highest amount of nitrogen, production of indoleacetic acid and gibberellic acid, and formation of larger phosphate-solubilizing zone. Many species of Pseudomonas, Bacillus, and Azotobacter can grow and survive at extreme environmental conditions, namely, tolerant to higher salt concentration, pH values, and even at dry soils with maximum temperature. Different factors affect Azotobacter population in soil such as pH, phosphorus content, soil aeration, and moisture contents. A. chroococcum found tolerant to a maximum NaCl concentration of 6% with a temperature of 45 °C and also up to pH of 8. Azotobacter species such as A. vinelandii, A. chroococcum, A. salinestris, A. tropicalis, and A. nigricans are able to produce antimicrobial compounds which inhibit the growth of common plant pathogens, viz., Fusarium, Aspergillus, Alternaria, Curvularia, and Rhizoctonia species. Pesticides used to control pests, insects, and phytopathogens are known to cause direct effect on soil microbiological aspects, environmental pollution, and health hazards in all living beings of the soil ecosystem. The species of Azotobacter are known to tolerate up to 5% pesticide concentration and also to degrade heavy metals and pesticides. A. chroococcum and A. vinelandii proved their biodegradation efficiency of many commonly used pesticides, viz., endosulfan, chlorpyrifos, pendimethalin, phorate, glyphosate, and carbendazim.
9.6.2 Pseudomonas Sp.
Pseudomonas sp. is an aerobic, gram-negative, ubiquitous organism present in agricultural soils and well adapted to grow in the rhizosphere. This rhizobacterium possesses many traits to act as a biocontrol agent and to promote the plant growth ability. It grows rapidly in vitro and can be mass-produced. It rapidly utilizes seed and root exudates and colonizes and multiplies in the rhizosphere and spermosphere environments. In the plant rhizosphere, it produces a wide spectrum of bioactive metabolites, that is, antibiotics, siderophores, volatiles, and growth-promoting substances; competes aggressively with other microorganisms; and adapts to environmental stresses. In addition, pseudomonads are responsible for the natural suppressiveness of some soilborne pathogens. It suppresses the growth of pathogenic microorganisms by various mechanisms, namely, production of antibiotics, bacteriocins, siderophores, hydrolytic enzymes such as β-1,3-glucanase and chitinases, and other metabolites such as phytoalexins and induction of systemic resistance. In this chapter, the characteristics of Pseudomonas sp., plant growth-promoting properties, mechanisms of plant growth promotion, and induction of systemic resistance by plant growth-promoting rhizobacterium (PGPR) against diseases and insect and nematode pests have been reviewed. PGPR strains initiating induced systemic resistance against a wide array of plant pathogens causing fungal, bacterial, and viral diseases and insect and nematode pests are discussed. Synergistic effects of PGPR strain mixtures and PGPRs as endophytes are brought out. Modes of action of Pseudomonas against fungal pathogens have been explained. Plant-disease controls by P. fluorescens have been elaborated. Interaction of P. fluorescens with pesticides has been indicated. Formulation characteristics, its approved uses in India, methods of application, and data requirements for registration particularly in India are discussed. Factors affecting growth of biopesticides and future issues and research needs in biopesticides are discussed.
PGPRs can exert both direct and indirect effects on plants, as shown in Fig. 9.2. Direct effects involve the PGPRs’ direct interactions with the plants, such as the production of plant growth hormones like auxins, cytokinins, and gibberellins, which can enhance plant growth and development. Additionally, PGPRs can secrete siderophores, iron-chelating compounds that facilitate iron uptake by plants, especially in iron-limited soils.

Direct and indirect effects of PGPRs on plants
Indirect effects refer to the influence of PGPRs on other soil organisms, which subsequently affect plant growth and development. For example, PGPRs can stimulate the activity of other soil microorganisms, including mycorrhizal fungi, which can form mutually beneficial associations with plants to enhance nutrient uptake. PGPRs can also trigger systemic resistance in plants, making them more resistant to pathogens and pests. The direct and indirect effects of PGPRs on plants can significantly enhance plant growth, nutrient uptake, and overall health, which can have vital implications for agriculture and sustainable crop production.
9.7 The Effects of PGPRs on Essential Oil
The positive role of PGPRs in improving EO content and EO profile of some medicinal plants has been reported. The PGPRs can modify the biosynthesis pathway of EO production in plants and lead to significant changes in yield and compounds. The improvement of EO content by PGPRs has been presented in Table 9.2. Accordingly, Yilmaz & Karik reported that PGPRs inoculation under low drought stress-enhanced Trachyspermum ammi seeds’ EO bioactivity. Sammak showed that the synergistic effect of AMF and Pseudomonas fluorescens improved growth and yield of Thymus kotschyanus EOs. Amini et al. (2020) noted that PGPRs improved EO yield and composition of Dracocephalum moldavica as affected by inoculation treatments under drought stress condition.
PGPRs have been found to positively affect the essential oil (EO) production of medicinal plants through various mechanisms.
One of the main mechanisms is the ability of PGPRs to improve nutrient acquisition in plants, particularly the uptake of nitrogen, phosphorus, and other essential minerals. PGPRs can achieve this by solubilizing insoluble minerals, chelating minerals, and producing enzymes that facilitate nutrient uptake. By improving nutrient availability, PGPRs can support the growth and development of medicinal plants, leading to an increase in EO production.
PGPRs can also modulate plant hormone levels, including the production of phytohormones such as auxins, cytokinins, and gibberellins. These hormones are crucial in plant growth and development, and can influence the production of secondary metabolites such as EOs. For example, auxins have been shown to promote the biosynthesis of several EO components, while cytokinins can enhance the synthesis of certain aromatic compounds.
Furthermore, PGPRs can help to mitigate abiotic and biotic stress in medicinal plants. Stresses such as drought, salinity, and pathogen attack can negatively impact plant growth and development, as well as EO production. PGPRs can alleviate these stresses by inducing the expression of stress-responsive genes and producing compounds such as osmolytes, antioxidants, and siderophores that protect the plant from damage. In addition, PGPRs can interact with other microorganisms in the rhizosphere, including mycorrhizal fungi and other beneficial bacteria. These interactions can promote synergistic effects on plant growth and EO production, by improving nutrient acquisition, hormone levels, and stress tolerance.
Overall, the effects of PGPRs on EO production in medicinal plants are complex and multifaceted. The ability of PGPRs to improve nutrient uptake, induce hormone production, and modulate stress responses can enhance the growth and development of medicinal plants, leading to an increase in EO production and quality. Further research is needed to fully understand the mechanisms behind the effects of PGPRs on EO production, as well as to identify the optimal PGPR strains and application methods for different medicinal plant species.
9.8 Conclusion
In conclusion, the use of biofertilizers and microbial consortia is a promising and sustainable approach to enhance the growth and secondary metabolite production of medicinal plants. By harnessing the beneficial effects of microorganisms like AMF and PGPRs, we can improve plant resilience and increase the accumulation of active compounds. However, to achieve optimal results, it is crucial to also consider environmental factors such as light, temperature, humidity, and soil fertility, as well as cultivation techniques. By using innovative substrate-based and substrate-free systems like aeroponic and hydroponic systems, and micropropagation, we can create ideal growth conditions for medicinal plants and ensure the production of high-quality bioactive substances. This can have significant implications for the pharmaceutical and herbal industries, as well as for the overall health and well-being of society.
References
Amini R, Zafarani-Moattar P, Shakiba MR, Sarikhani MR (2020) Essential oil yield and composition of Moldavian balm (Dracocephalum moldavica L.) as affected by inoculation treatments under drought stress condition. J Essent Oil Bear Plants 23(4):728–742. https://doi.org/10.1080/0972060X.2020.1765851
Arpanahi AA, Feizian M (2019) Arbuscular mycorrhizae alleviate mild to moderate water stress and improve essential oil yield in thyme. Rhizosphere 9:93–96. https://doi.org/10.1016/j.rhisph.2019.05.007
Arpanahi AA, Feizian M, Mehdipourian G, Khojasteh DN (2020) Arbuscular mycorrhizal fungi inoculation improve essential oil and physiological parameters and nutritional values of thymus daenensis Celak and Thymus vulgaris L. under normal and drought stress conditions. Eur J Soil Biol 100:103217. https://doi.org/10.1016/j.ejsobi.2020.103217
Azizi S, Kouchaksaraei MT, Hadian J, Abad ARFN, Sanavi SAMM, Ammer C, Bader MKF (2021) Dual inoculations of arbuscular mycorrhizal fungi and plant growth-promoting rhizobacteria boost drought resistance and essential oil yield of common myrtle. For Ecol Manag 497:119478. https://doi.org/10.1016/j.foreco.2021.119478
Bhardwaj AK, Chandra KK, Kumar R (2023) Water stress changes on AMF colonization, stomatal conductance and photosynthesis of Dalbergia sissoo seedlings grown in entisol soil under nursery condition. For Sci Technol 19:1–13
Brader G, Compant S, Mitter B, Trognitz F (2014) Metabolic potential of endophytic bacteria. Curr Opin Biotechnol 27:30–37
Brenner DM (1993) Perilla: botany, uses and genetic resources. In: Janick J, Simon JE (eds) New crops. Wiley, New York, pp 322–328
Bulgarelli D, Schlaeppi K, Spaepen S, van Themaat EVL, Schulze-Lefert P (2013) Structure and functions of the bacterial microbiota of plants. Annu Rev Plant Biol 64:807–838
Carreón-Abud Y, Torres-Martínez R, Farfán-Soto B, Hernández-García A, Ríos-Chávez P, Bello-González MA, Martínez-Trujillo M, Salgado-Garciglia R (2015) Arbuscular mycorrhizal symbiosis increases the content of volatile terpenes and plant performance in Satureja macrostema (Benth.) Briq. Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas 14(4):273–279. https://doi.org/10.13140/RG.2.1.3233.9449
Castrillo G, Teixeira PJ, Paredes SH, Law TF, de Lorenzo L, Feltcher ME, Finkel OM, Breakfield NW, Mieczkowski P, Jones CD, Paz-Ares J, Dangl JL (2017) Root microbiota drive direct integration of phosphate stress and immunity. Nature 543:513–518
Chen ML, Yang G, Sheng Y, Li PY, Qiu HY, Zhou XT, Huang LQ, Chao Z (2017) Glomus mosseae inoculation improves the root system architecture, photosynthetic efficiency and flavonoids accumulation of liquorice under nutrient stress. Front Plant Sci 8:931. https://doi.org/10.3389/fpls.2017.00931
Chojnacka K, Moustakas K, Witek-Krowiak A (2020) Bio-based fertilizers: a practical approach towards circular economy. Bioresour Technol 295:122223
Colonna E, Rouphael Y, De Pascale S, Barbieri G (2016) Effects of mycorrhiza and plant growth promoting rhizobacteria on yield and quality of artichoke. Acta Hortic 1147:43–50. https://doi.org/10.17660/ActaHortic.2016.1147.7
Compant S, Cambon MC, Vacher C, Mitter B, Samad A, Sessitsch A (2021) The plant endosphere world–bacterial life within plants. Environ Microbiol 23(4):1812–1829
Dambisya YM, Tindimwebwa G (2003) Traditional remedies in children around Eastern Cape. South Africa East Afr Med J 80(8):402–405
Darakeh SASS, Weisany W, Abdul-Razzak Tahir N, Schenk PM (2022) Physiological and biochemical responses of black cumin to vermicompost and plant biostimulants: arbuscular mycorrhizal and plant growth-promoting rhizobacteria. Ind Crop Prod 188:115557
Dave S, Das J, Tarafdar JC (2011) Effect of vesicular arbuscular mycorrhizae on growth and saponin accumulation in Chlorophytum borivilianum. Science Asia 37(2):165–169. https://doi.org/10.2306/scienceasia1513-1874.2011.37.165
de Souza BC, da Cruz RMS, Lourenço ELB, Pinc MM, Dalmagro M, da Silva C, Alberton O et al (2022) Inoculation of lemongrass with arbuscular mycorrhizal fungi and rhizobacteria alters plant growth and essential oil production. Rhizosphere 22:100514. https://doi.org/10.1016/j.rhisph.2021.100514
de Vries FT, Griffiths RI, Knight CG, Nicolitch O, Williams A (2020) Harnessing rhizosphere microbiomes for drought-resilient crop production. Science 368:270–274
Dehsheikh AB, Sourestani MM, Zolfaghari M, Enayatizamir N (2020) Changes in soil microbial activity, essential oil quantity, and quality of Thai basil as response to biofertilizers and humic acid. J Clean Prod 256:120439. https://doi.org/10.1016/j.jclepro.2020.120439
Dos Santos EL, Falcão EL, Barbosa da Silva FS (2021) Mycorrhizal technology as a bioinsumption to produce phenolic compounds of importance to the herbal medicine industry. Res Soc Dev 10(2). https://doi.org/10.33448/rsd-v10i2.15891
Eke P, Adamou S, Fokom R, Nya VD, Fokou PVT, Wakam LN et al (2020) Arbuscular mycorrhizal fungi alter antifungal potential of lemongrass essential oil against fusarium solani, causing root rot in common bean (Phaseolus vulgaris L.). Heliyon 6(12):e05737. https://doi.org/10.1016/j.heliyon.2020.e05737
Eshaghi Gorgi O, Fallah H, Niknejad Y, Barari Tari D (2022) Effect of plant growth promoting rhizobacteria (PGPR) and mycorrhizal fungi inoculations on essential oil in Melissa officinalis L. under drought stress. Biologia 77:11–20. https://doi.org/10.1007/s11756-021-00761-x
Ezzati Z, Weisany W, Abdul-razzak Tahir N (2021) Arbuscular mycorrhizal fungi species improve the fatty acids profile and nutrients status of soybean cultivars grown under drought stress. J Appl Microbiol 132:2177–2188
Falleh H, Jemaa MB, Saada M, Ksouri R (2020) Essential oils: A promising eco-friendly food preservative. Food Chem 330:127268. https://doi.org/10.1016/j.foodchem.2020.127268
Fasusi OA, Cruz C, Babalola OO (2021) Agricultural sustainability: microbial biofertilizers in rhizosphere management. Agriculture 11(2):163. https://doi.org/10.3390/agriculture11020163
Ghiaee A, Naghibi F, Esmaeili S, Mosaddegh M (2014) Herbal remedies connected to malaria like fever in Iranian ancient medicinal books. Iran J Parasitol 9(4):553–559
Golubkina N, Logvinenko L, Novitsky M, Zamana S, Sokolov S, Molchanova A, Caruso G et al (2020) Yield, essential oil and quality performances of Artemisia dracunculus, Hyssopus officinalis and Lavandula angustifolia as affected by arbuscular mycorrhizal fungi under organic management. Plan Theory 9(3):375. https://doi.org/10.3390/plants9030375
Gujre N, Soni A, Rangan L, Tsang DC, Mitra S (2021) Sustainable improvement of soil health utilizing biochar and arbuscular mycorrhizal fungi: A review. Environ Pollut 268:115549. https://doi.org/10.1016/j.envpol.2020.115549
Gurikar C, Sreenivasa MY, Gowda NN, Lokesh AC (2022) Azotobacter—A potential symbiotic rhizosphere engineer. In: Rhizosphere engineering. Academic Press, pp 97–112. https://doi.org/10.1016/B978-0-12-824508-6.00006-4
Hanif MA, Nisar S, Khan GS, Mushtaq Z, Zubair M (2019) Essential oils. In: Essential oil research: trends in biosynthesis, analytics, industrial applications and biotechnological production, vol 3-17. Springer, Cham. https://doi.org/10.1016/B978-0-12-818765-3.00001-1
Hatami M, Khanizadeh P, Bovand F, Aghaee A (2021) Silicon nanoparticle-mediated seed priming and pseudomonas spp. inoculation augment growth, physiology and antioxidant metabolic status in Melissa officinalis L. plants. Ind Crop Prod 162:113238. https://doi.org/10.1016/j.indcrop.2021.113238
Hazzoumi Z, Moustakime Y, Elharchli EH, Joutei KA (2015) Effect of arbuscular mycorrhizal fungi (AMF) and water stress on growth, phenolic compounds, glandular hairs, and yield of essential oil in basil (Ocimum gratissimum L.). Chem Biol Technol Agric 2:2–11. https://doi.org/10.1186/s40538-015-0033-6
Joshee N, Yadav AK, Mentreddy R (2007) Mycorrhizal fungi and growth and development of micropropagated Scutellaria integrifolia plants. Ind Crop Prod 25(2):169–177. https://doi.org/10.1016/j.indcrop.2006.09.005
Kapoor R, Anand G, Gupta P, Mandal S (2017) Insight into the mechanisms of enhanced production of valuable terpenoids by arbuscular mycorrhiza. Phytochem Rev 16:677–692. https://doi.org/10.1007/s11101-017-9508-5
Kapoor R, Giri B, Mukerji KG (2002) Glomus macrocarpum: a potential bioinoculant to improve essential oil quality and concentration in dill (Anethum graveolens L.) and Carum (Trachyspermum ammi (Linn.) Sprague). World J Microb Biot 18:459–463. https://doi.org/10.1023/A:1015262525430
Kapoor R, Giri B, Mukerji KG (2004) Improved growth and essential oil yield and quality in Foeniculum vulgare mill on mycorrhizal inoculation supplemented with P-fertilizer. Bioresour Technol 93(3):307–311. https://doi.org/10.1016/j.biortech.2003.10.016
Kaur S, Suseela V (2020) Unraveling arbuscular mycorrhiza-induced changes in plant primary and secondary metabolome. Meta 10(8):335. https://doi.org/10.3390/metabo10080335
Khalediyan N, Weisany W, Schenk PM (2021) Arbuscular mycorrhizae and rhizobacteria improve growth, nutritional status and essential oil production in Ocimum basilicum and Satureja hortensis. Ind Crop Prod 160:113163. https://doi.org/10.1016/j.indcrop.2020.113163
Khaosaad T, Vierheilig H, Nell M, Zitterl-Eglseer K, Novak J (2006) Arbuscular mycorrhiza alter the concentration of essential oils in oregano (Origanum sp., Lamiaceae). Mycorrhiza 16(6):443–446. https://doi.org/10.1007/s00572-006-0043-8
Khosropour E, Hakimi L, Weisany W (2023) Role of engineered nanomaterials in biotic stress management. Plant Biology, sustainability and climate change, Engineered Nanomaterials for Sustainable Agricultural Production, Soil Improvement and Stress Management. Elsevier, pp 257–272
Kordi S, Salmasi SZ, Kolvanagh JS, Weisany W, Shannon DA (2020) Intercropping system and N2 fixing bacteria can increase land use efficiency and improve the essential oil quantity and quality of sweet basil (Ocimum basilicum L.). Front Plant Sci 11:610026. https://doi.org/10.3389/fpls.2020.610026
Kumar S, Sindhu SS, Kumar R (2022) Biofertilizers: an ecofriendly technology for nutrient recycling and environmental sustainability. Curr Res Microb Sci 3:100094. https://doi.org/10.1016/j.crmicr.2022.100094
Lima KB, Riter Netto AF, Martins MA, Freitas MSM (2015) Crescimento, acúmulo de nutrientes e fenóis totais de mudas de cedro australiano (Toona ciliata) inoculadas com fungos micorrízicos. Ciência Florestal 25:853–862. https://doi.org/10.5902/1980509819663
Lu FC, Lee CY, Wang CL (2015) The influence of arbuscular mycorrhizal fungi inoculation on yam (Dioscorea spp.) tuber weights and secondary metabolite content. PeerJ 3:e1266. https://doi.org/10.7717/peerj.1266
Mahapatra DM, Satapathy KC, Panda B (2022) Biofertilizers and nanofertilizers for sustainable agriculture: Phycoprospects and challenges. Sci Total Environ 803:149990. https://doi.org/10.1016/j.scitotenv.2021.149990
Mathens A, Bellanger R (2010) Herbs and other dietary supplements: current regulations and recommendations for use to maintain health in the management of the common cold or other related infectious respiratory illnesses. J Pharm Pract 23:117–127
Mirzaei M, Ladan Moghadam A, Hakimi L, Danaee E (2020) Plant growth promoting rhizobacteria (PGPR) improve plant growth, antioxidant capacity, and essential oil properties of lemongrass (Cymbopogon citratus) under water stress. Iranian J Plant Physiol 10(2):3155–3166. https://doi.org/10.22034/ijpp.2020.1881571.1183
Mukhopadhyay R, Sarkar B, Jat HS, Sharma PC, Bolan NS (2021) Soil salinity under climate change: challenges for sustainable agriculture and food security. J Environ Manag 280:111736. https://doi.org/10.1016/j.jenvman.2020.111736
Oliveira MS, Campos MAS, Silva FSB (2015) Arbuscular mycorrhizal fungi and vermicompost to maximize the production of foliar biomolecules in Passiflora alata Curtis seedlings. J Sci Food Agric 95:522–528. https://doi.org/10.1002/jsfa.6777
Oliveira MS, Campos MA, Albuquerque UP, Silva FS (2013) Arbuscular mycorrhizal fungi (AMF) affect biomolecules content in Myracrodruon urundeuva seedlings. Ind Crop Prod 50:244–247. https://doi.org/10.1016/j.indcrop.2013.07.049
Pankaj U, Verma RS, Yadav A, Verma RK (2019) Effect of arbuscular mycorrhizae species on essential oil yield and chemical composition of palmarosa (Cymbopogon martinii) varieties grown under salinity stress soil. J Essent Oil Res 31(2):145–153. https://doi.org/10.1080/10412905.2018.1558801
Panke-Buisse K, Poole A, Goodrich J, Ley RE, Kao-Kniffin J (2015) Selection on soil microbiomes reveals reproducible impacts on plant function. ISME J 9:980–989
Pirsarandib Y, Hassanpouraghdam MB, Rasouli F, Aazami MA, Puglisi I, Baglieri A (2022) Phytoremediation of soil contaminated with heavy metals via arbuscular mycorrhiza (Funneliformis mosseae) inoculation ameliorates the growth responses and essential oil content in lavender (lavandula angustifolia L.). Agronomy 12(5):1221. https://doi.org/10.3390/agronomy12051221
Polukhin AA, Panarina VI (2022) Financial risk management for sustainable agricultural development based on corporate social responsibility in the interests of food security. Risks 10(1):17. https://doi.org/10.3390/risks10010017
Sadegh Sadat Darakeh SA, Weisany W, Diyanat M, Ebrahimi R (2021) Bio-organic fertilizers induce biochemical changes and affect seed oil fatty acids composition in black cumin (Nigella sativa Linn). Ind Crop Prod 164:113383. https://doi.org/10.1016/j.indcrop.2021.113383
Sammak AS, Anvari M, Matinizadeh M, Mirza M (2020) The synergistic effect of arbuscular mycorrhizal fungi and Pseudomonas fluorescens on growth and qualitative and quantitative yield of Thymus kotschyanus essential oil. J Essent Oil Bear Plants 23(3):532–547. https://doi.org/10.1080/0972060X.2020.1773586
Sangwan S, Prasanna R (2022) Mycorrhizae helper bacteria: unlocking their potential as bioenhancers of plant–arbuscular mycorrhizal fungal associations. Microb Ecol 84(1):1–10. https://doi.org/10.1007/s00248-021-01728-1
Sharma E, Anand G, Kapoor R (2017) Terpenoids in plant and arbuscular mycorrhiza-reinforced defense against herbivorous insects. Ann Bot 119:791–801. https://doi.org/10.1093/aob/mcw218
Silvia FA, Silva FSB, Maia LC (2014) Biotechnical application of arbuscular mycorrhizal fungi used in the production of foliar biomolecules in ironwood seedlings [Libidibia ferrea (Mart. Ex Tul.) L.P. Queiroz var. ferrea]. J Med Plant Res 8(20):814–819. https://doi.org/10.5897/JMPR2014.5483
Titanji VP, Zofou D, Ngemenya MN (2008) The antimalarial potential of medicinal plants used for the treatment of malaria in Cameroonian folk medicine. Afr J Tradit Complement Altern Med 5(3):302–321
Toussaint JP, Smith FA, Smith SE (2007) Arbuscular mycorrhizal fungi can induce the production of phytochemicals in sweet basil irrespective of phosphorus nutrition. Mycorrhiza 17(4):291–297. https://doi.org/10.1007/s00572-007-0123-1
Weisany W, Raei Y, Ghasemi-Golezani K (2016b) Funneliformis mosseae alters seed essential oil content and composition of dill in intercropping with common bean. Ind Crop Prod 79:29–38. https://doi.org/10.1016/j.indcrop.2015.10.041
Weisany W, Raei Y, Pertot I (2015) Changes in the essential oil yield and composition of dill (Anethum graveolens L.) as response to arbuscular mycorrhiza colonization and cropping system. Ind Crop Prod 77:295–306. https://doi.org/10.1016/j.indcrop.2015.09.027
Weisany W, Raei Y, Salmasi SZ, Sohrabi Y, Ghassemi-Golezani K (2016a) Arbuscular mycorrhizal fungi induced changes in rhizosphere, essential oil and mineral nutrients uptake in dill/common bean intercropping system. Ann Appl Biol 169(3):384–397. https://doi.org/10.1111/aab.12309
Weisany W, Tahir NAR, Schenk PM (2021) Coriander/soybean intercropping and mycorrhizae application lead to overyielding and changes in essential oil profiles. Eur J Agron 126:126283. https://doi.org/10.1016/j.eja.2021.126283
Yadav K, Aggarwal A, Singh N (2013) Arbuscular mycorrhizal fungi induced acclimatization and growth enhancement of Glycyrrhiza glabra L.: a potential medicinal plant. Agric Res 2:43–47. https://doi.org/10.1007/s40003-012-0026-8
Yadav K, Singh N, Aggarwal A (2012) Arbuscular mycorrhizal technology for the growth enhancement of micropropagated Spilanthes acmella Murr. Plant Prot Sci 48:31–36. https://doi.org/10.17221/60/2011-PPS
Yang Y, Ou XH, Yang G, Xia YS, Chen ML, Guo LP, Liu DH (2017) Arbuscular mycorrhizal fungi regulate the growth and phyto-active compound of salvia miltiorrhiza seedlings. Appl Sci 7(1):68. https://doi.org/10.3390/app7010068
Yilmaz A, Karik Ü (2022) AMF and PGPR enhance yield and secondary metabolite profile of basil (Ocimum basilicum L.). Ind Crop Prod 176:114327. https://doi.org/10.1016/j.indcrop.2021.114327
Zare AS, Ganjeali A, Kakhki MRV, Mashreghi M, Cheniany M (2023) Additive effects of arbuscular mycorrhizae and TiO2 nanoparticles on growth and essential oils enhancement of peppermint. Rhizosphere 25:100659. https://doi.org/10.1016/j.rhisph.2022.100659
Zeng Y, Guo LP, Chen BD, Hao ZP, Wang JY, Huang LQ, Yang G, Cui XM, Yang L, Wu ZX, Chen ML, Zhang Y (2013) Arbuscular mycorrhizal symbiosis and active ingredients of medicinal plants: current research status and perspectives. Mycorrhiza 23(4):253–265. https://doi.org/10.1007/s00572-013-0486-4
Zhi LY, Chuan CD, Lian QC (2007) Regulation and accumulation of secondary metabolites in plant-fungus symbiotic system. Afr J Biotechnol 6:1266–1271. https://doi.org/10.5897/AJB2007.000-2415
Zimare S, Borde M, Jite PK, Malpathak N (2013) Effect of AM fungi (gf, gm) on biomass and gymnemic acid content of Gymnema sylvestre (Retz.). Biol Fertil Soils 49(3):439–445. https://doi.org/10.1007/s00374-012-0734-x
Zitter-Eglseer K, Nell M, Lamien-Meda A, Steinkellner S, Wawrosch C, Kopp B, Novak J (2015) Effects of root colonization by symbiotic arbuscular mycorrhizal fungi on the yield of pharmacologically active compounds in Angelica archangelica L. Acta Physiol Plant 37:2–11. https://doi.org/10.1007/s11738-014-1695-5
Zolfaghari M, Nazeri V, Sefdkon F, Rejali F (2013) Effect of arbuscular mycorrhizal fungi on plant growth and essential oil content and composition of Ocimum basilicum L. Iranian J Plant Physiol 3:643–650
Zubek S, Stojakowska A, Anielska T, Turnau K (2010) Arbuscular mycorrhizal fungi alter thymol derivative contents of Inula ensifolia L. Mycorrhiza 20(7):497–504. https://doi.org/10.1007/s00572-010-0316-1
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2024 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Weisany, W., Khosropour, E., Alavian, A. (2024). Application of Microbial Consortia and Biofertilizer to Improve the Quality and Yield of Essential Oils in Aromatic Plants. In: Prakash, B., Dubey, N.K., Freitas Brilhante de São José, J. (eds) Plant Essential Oils. Springer, Singapore. https://doi.org/10.1007/978-981-99-4370-8_9
Download citation
DOI: https://doi.org/10.1007/978-981-99-4370-8_9
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-99-4369-2
Online ISBN: 978-981-99-4370-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)