
Abstract
In most cases, enzymes, antifungal microbial, and various bio-sourced methods were effective for the control of mycotoxin contaminations, while it has a common deficiency that not all of the mycotoxins were desirably controlled when treating with these abovementioned means. Herein, we concluded multiple physical and chemical agents to control mycotoxins and also the most widely used way in the factory production. All of the physical and part of the chemical treatments aimed at the reduction of the mycotoxins themselves, which means destroying or transformation of their molecular structures. Most chemical reagents were used in the field experiments where they aimed at the resources of these mycotoxins, i.e., fungal pathogens. Of course, some antifungal reagents are especially absorbed or combined with these mycotoxins just like multiple bio-resourced agents. After all, these two methods played an important role in the front of food processing procedures, due to their low cost, simplicity, high efficiency, and sufficient universality.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Physical Treatments for Reducing Mycotoxin Contaminations
Before the appearance of variable chemical reagents and specific biogenic enzymes, physical processing was the most effective way to remove mycotoxins during the production practice, and it is still under extensive uses till now. Among these, thermo-treatment is seemed to be the simplest and operable method. Gbashi et al. (2019) suggested parameters of 210.85 °C/54.71 min for mycotoxin-contaminated maize flour as the optimum degradation conditions. Siciliano et al. (2017) also tested the heat exposure of 140 °C for 40 min, and it was effective for detoxification of aflatoxin with no effects on lipid profile and the nutritional quality in foodstuffs. Furthermore, during food processing at high temperature, ochratoxin A (OTA) could be turned to its thermal isomerization product 2′R-Ochratoxin A, showing different metabolic characteristics in human body (Sueck et al. 2018) (Fig. 9.1).

The hypothetical molecular structures of ochratoxin A (OTA) and its thermal isomer 2′R-OTA. Important interacting residues are shown as sticks, OTA can form salt-bridge interactions with R218, R222, and K195; 2′R-OTA can only form these interactions with R222. (Sueck F et al. 2018)
Besides heat, plasma and various waves were also attempted to be widely applied for mycotoxin control. The treatment of Penicillium verrucosum on barley after an incubation of 5 days using a CO2 + O2 plasma and air plasma resulted in a reduction of the ochratoxin A (OTA) content from 49.0 (untreated) to 27.5 (1 min) and 25.7 (1 min), respectively (Durek et al. 2018). In addition, Shi et al. (Shi 2017) confirmed the disappearance of the C8═C9 double bond in the furofuran ring of aflatoxin B1 (AFB1) after treatment of high-voltage atmospheric cold plasma (Shi 2017). Very recently, the cold atmospheric pressure plasma may overcome multiple weaknesses associated with the classical methods, and most mycotoxins exposed to this plasma were degraded almost completely within 60 s (Hojnik et al. 2017; Ten Bosch et al. 2017). Hernández et al. compared three treatments during milk storage, non-treatment, pasteurization, and thermoultrasound (Hernandez-Falcon et al. 2018), suggesting that 10 min-thermoultrasound unhomogenized milk 1 day after storage kept the lowest aflatoxin M1 (AFM1) level of 0.15 ± 0.05 pg/mL. Similarly, oven roasting and microwaving of chicken breast muscles can reduce the content of ten prevalent mycotoxins, reducing their increased bioaccessibility (Sobral et al. 2019) (Fig. 9.2).

Chromatograms of AFB1 untreated (5 μg/mL) in 50% ethanol solution (a) and AFB1 sample treated by HVACP in ambient air for 5 min (b). (Shi et al. 2017)
Interestingly, the light might be connected with mycotoxin production. As demonstrated, the optimized combinational treatments of radiation and essential oil were very effective in reducing the fungal growth and mycotoxins content (Kalagatur et al. 2018a). Also, Wang et al. found that high intensity of blue light decreased the accumulation of citrinin (CIT) production in Monascus ruber, providing a unique mentality of mycotoxin reduction in the field production (Wang et al. 2016) (Fig. 9.3).

The principles of light affect the production of citrinin (CIT). (Wang L et al. 2016)
Specially for mycotoxins of Alternaria toxins, improved food processing methods, such as milling process (Janic Hajnal et al. 2019) and extrusion processing (Janic Hajnal et al. 2016), can affect the residue of tenuazonic acid (TeA), alternariol (AOH), and alternariol monomethyl ether (AME), suggesting the demands of high-grade raw-food material and optimized machine process. Not just Alternaria, various physical postharvest treatments were deadly for pathogenic fungi, which obviously cause oxidative injury, protein impairment, and cell wall degradation, thus reduced mycotoxin contaminations (Liu et al. 2018; Karlovsky et al. 2016), for instance, extrusion and alkaline cooking of corn kernels reduced the fumonisin toxicity for mice (Voss and Ryu 2017). Also, processing methods such as color sorting reduced the levels of aflatoxin and can be practically utilized to monitor the occurrence of aflatoxins in some commercial products (Zivoli et al. 2016) (Fig. 9.4).

Optimum region obtained after superimposing the contour plots of the system response. (Janic Hajnal et al. 2016)
2 Chemical Treatments for Reducing Mycotoxin Contaminations
With the rapid development of traditional inorganic and synthetic chemistry, structural biologists applied the known interplay between multiple mycotoxins and diverse structures, eliminating mycotoxins from the molecular levels. The common removers normally include various acids, fungicides, organic oils, and little parts of other materials, while all of the compounds can be divided into two parts, some are the inhibitors of fungal growth, especially the essential metabolic pathways of mycotoxin synthesis, and the others act on the mycotoxins themselves, removing them by modification or completely degradation.
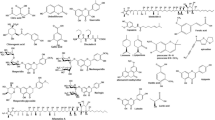
Over the past decades, some scientists have found plenty of antifungal agents to control mycotoxin contamination. Moon et al. (2017) tested 38 coumarins and found that 4-hydroxy-7-methyl-3-phenyl coumarin was the most effective controller of aflatoxin for its downregulated function of aflD, aflK, aflQ, and aflR in the pathways of aflatoxin biosynthesis. Furthermore, coumarins can prevent the aflatoxin B1 (AFB1)-induced hepatotoxic effects via increasing antioxidant capacities and inhibiting the CYP450 isozyme-mediated activation (Zhang et al. 2016). One of the anthocyanins, delphinidin, showed great protection effects of gut from Alternaria-induced genotoxicity (Aichinger et al. 2018). Also, grape seed proanthocyanidin extract was found to be related to the activation of the Nrf2/ARE signaling pathway, which mRNA expression levels can be downregulated by zearalenone (ZEN), protecting the ZEN-induced hepatic injury (Long et al. 2016). Besides, more and more active substances, such as rutin and its derivatives (Huang et al. 2019), tannis (Peng et al. 2017) and validamycin (Li et al. 2019), were revealed to be connected with biocontrol efficiency and mycotoxin production of filamentous fungi. A similar group of biocontrol substances also includes piperine, and the main purpose was aimed at AFB1. Results showed that piperine inhibits AFB1 production while positively modulating fungal antioxidant status in Aspergillus flavus (Caceres et al. 2017), and thus it was demonstrated useful of its antigenotoxic effect in broiler chickens (da Silva Cardoso et al. 2016). Furthermore, researchers assessed several piperine-like synthetic compounds, in which 1-(2-methylpiperidin-1-yl)-3-phenylprop-2-en-1-one showed the most potential of antifungal and antiaflatoxigenic effects against Aspergillus flavus (Moon et al. 2016) (Fig. 9.5).

The proposed working model of how validamycin affects the pathogen F. graminearum and the host wheat. (Li et al. 2019)
Fortunately, we can also see some valuable investigations aimed at the detoxification of mycotoxin in vitro. With the development of new macromolecular inorganic carbon matters such as activated carbon (Kalagatur et al. 2017), activated carbon fibers (El Khoury et al. 2018) make the mycotoxin absorption available. Similar eliminated artificial adsorbents include varied clays or bentonites. Cetylpyridinium chloride ions modified around organozeolites prepared by clinoptilolite and phillipsite are responsible for ZEN absorption (Markovic et al. 2016). Moreover, it was identified from two human cells that bentonite modified with zinc enhanced the antiaflatoxigenic ability (Nones et al. 2017). Researchers also evaluated the detoxification potential of both ZEN and AFB via multiple montmorillonites (Wang et al. 2018a, 2019b) and surfactants (Wang et al. 2018b), plus, Kang et al. (2016) used the combined batch experiment with computational models, indicating that adsorption affinity of mycotoxins is dependent on the surfaces of clay aggregates. With the aid of micropore structure in chitosan derivatives (Yang et al. 2017), shrimp shells (Assaf et al. 2018), natural zeolites (Eroglu et al. 2017), and bentonite clay (Shannon et al. 2016), small molecules of mycotoxins were all sedimented and aggregated to the fixed microporous structure, reducing their toxicities to variable target organisms (Fig. 9.6).

There is also one part of important mycotoxin absorbents or antifungal agents, nanocomposites. Gao et al. (2019) synthesized an organic-inorganic P(DMDAAC-AGE)/Ag/ZnO composite for the usage of antibacterial cotton fabrics with laundering durability, showing more than 99.00% of anti-mildew performance even after 11 washing cycles. A kind of surface active maghemite nanoparticles was made to bind citrinin; mass spectroscopy and magnetization measurements showed that the material was an efficient and reliable mean for citrinin removal (Magro et al. 2016). Ag doped hollow TiO2 nanoparticles were identified to be effective in protecting farm products affected by Fusarium solani or other fungi, where the Ag promotes the formation of stable Ag-S and disulfide bonds (R-S-S-R) in cellular protein, leading to cell damage (Boxi et al. 2016). Furthermore, nanocomposites mixed with other agents can significantly increase their antifungal property, such as chitosan nanoparticles encapsulated with essential oil (Kalagatur et al. 2018b), pyraclostrobin solid nanodispersion by self-emulsifying technique (Wang et al. 2019a).
Ozone treatment was also effective for pathogen prevention and mycotoxin control. It has been concluded that ozone has great potential to improve the functionalities of grain products while ensuring food safety (Zhu 2018). After treatment of ozone at 1.10 mg/L for 120 min, Fusarium rot development and neosolaniol accumulation in fruits were discriminately controlled (Hua-Li et al. 2018) (Fig. 9.7).

The proposed mechanism of the reaction of ozone to the mycotoxin NEO. (Hua-Li et al. 2018)
Interestingly, potassium may also relate to the antifungal attempt. Penicillium expansum under 5 mM potassium phosphite stress presented yield reduction of patulin and lower infectivity to apple fruit (Lai et al. 2017). Moreover, Chang et al. demonstrated that addition of triphenylphosphonium cation can enhance or at least restore compounds’ antifungal activity, resulting in mitochondrial dysfunction and increased levels of intracellular reactive oxygen species (Chang et al. 2018).
Besides, other common chemical treatments to remove mycotoxins were varied organic active substances; among these organic acids, essential oils, aldehyde derivatives, organism extracts, and artificial fungicides were the substances with great representations.
3 Organic Acids
Acids play a great role in control of pathogens and mycotoxins; single or multiple combined acids were proved effective in the food or food products protection. Researchers purified cuminic acid from the seeds of Cuminum cyminum L., which significantly distorted the mycelial morphology of Fusarium oxysporum; more importantly, synthetic genes of bikaverin (Bike1, Bike2, and Bike3) and fusaric acid (FUB1, FUB2, FUB3 and FUB4) both were downregulated (Sun et al. 2017). Also, p-aminobenzoic acid was present in the secretions of rhizobacterium Lysobacter to show a broad spectrum of antifungal activities, which inhibits septation during cell division (Laborda et al. 2019). Most acids targeted in the membrane of fungal hyphae, ethyl p-coumarate, can cause severe lipid peroxidation and heavy oxidative damage to the cellular membranes and organelles of pathogens (Li et al. 2018). Chlorogenic and caffeic acids were also groups of antifungal micromolecular acids; both can be degraded into protocatechuic acid in the Fusarium graminearum and dramatically enhanced the counteraction of the fungal ability of mycotoxin production (Gauthier et al. 2016). Similar compounds including I-pyroglutamic acid (Bilska et al. 2018), sinapic acid (Kulik et al. 2017a), fluorinated dihydroguaiaretic acid (Yamauchi et al. 2017), etc. were all with antifungal potential. Meanwhile, gene expression revealed that Tri genes responsible for trichothecene biosynthesis were downregulated after acid treatment (Fig. 9.8).

Effect of cuminic acid on mycelia morphology of Fusarium oxysporum f. sp. niveum. (d–f) Untreated plates; (a–c) plates treated with cuminic acid at EC50 value (22.53 μg/mL). Values are means and standard errors. (Sun Y et al. 2017)
Combination of multiple or even unknown acids seems like to have synergistic effects for the antifungal property. For example, the cell-free supernatant of Lactobacillus plantarum inhibits the production of aflatoxins by 91%, where lactic acid, phenyllactic acid, hydroxyphenyllacetic acid, and indole lactic acid were detected in the metabolites and all were tested separately with antiaflatoxigenic property (Guimaraes et al. 2018). Combination of exogenous abscisic acid and gibberellic acid can also reduce head blight severity, as phytohormones, combining fungicidal agents and these acids may also be useful for the management of head blight infection (Buhrow et al. 2016). Phenolic acids were a group of organic matters with a phenolic hydroxyl group; caffeic, chlorogenic, ferulic, and p-coumaric acid were all tested. All those have somewhat variable effects on fungal growth and mycotoxin production, depending on the host strain and the concentration (Ferruz et al. 2016). Moreover, Ferruz et al. also proved that phenolic acids have the antifungal property and upregulated ergosterol biosynthesis by Fusaria (Kulik et al. 2017b).
4 Fungicides
Traditional fungicides were mostly aimed at pathogen growth inhibition, thus leading to the reduction of multiple mycotoxins. Wayne et al. (Jurick et al. 2019) tested the sensitivity of Penicillium spp. to difenoconazole and suggested a minimum limitation of 5 μg/ml to fruit quality and mycotoxin control. A field experiment showed that prothioconazole could significantly reduce the total deoxynivalenol and zearalenone contents by applications at 18th leaf and R2 stages, while not visibly changing other mycotoxin contents (Limay-Rios and Schaafsma 2018).
And tebuconazole was also one efficient fungicide, the addition of tebuconazole significantly decreases the DON production, and meanwhile, TRI5 and TRI6 of biosynthetic genes were downregulated (Diao et al. 2018). Similarly, epoxiconazole (Duan et al. 2018), triticonazole (Zhang et al. 2018), arylpyrazole (Zhang et al. 2017), and varied triazoles (Hellin et al. 2017) were all verified to be effective to the pathogen prevention and mycotoxin control (Fig. 9.9).

Frequency distribution of isolates in fungicide concentration that inhibit mycelial growth by 50% for technical grade difenoconazole for Penicillium isolates. (Jurick et al. 2019)
As a powerful and simple method to control fungal pathogens, fungicides were always compared with other emerging antifungal agents, but no doubt, fungicides were still the most widely used field method. Scaglioni et al. (Scaglioni and Blandino 2018) compared the inhibition effects of fumonisin production between tebuconazole and microalgal phenolic extracts, and both showed antifungal potential, while either would directly control the fumonisin contents. Moreover, a direct comparison of the synthetic commercial antifungal compound and the antifungal protein PgAFP showed that both were with fungal growth inhibition. Furthermore, the presence of PgAFP produced a significant reduction in mycotoxins accumulation (da Cruz Cabral et al. 2019). Additionally, researchers investigated the effects of two fungicides, quinone outside inhibitors and demethylation inhibitors, on Fusarium head blight and deoxynivalenol production, and results suggested that the combination effect of two fungicides depended on the active ingredients and whether the two were applied as a mixture at heading or sequentially (Paul and Bradley 2018). Anyway, new emerging antifungal agents were always tested by making comparison with these commercial fungicides, which represents a dominance of fungicides in fungal pathogens and mycotoxins control in a very long period (Fig. 9.10).

The changes on the colonies of Alternaria tenuissima sp. grp. grown on wheat media in the presence of two antifungal treatments (PgAFP and F + M) at two water activity (aw) levels, 0.95 and 0.98 after 6 days of incubation at 25 °C. (da Cruz Cabral L et al. 2019)
5 Aldehydes, Essential Oils, and Organism Extracts
Besides organic acids, aldehydes and their derivatives were also popular for mycotoxin control, and most of them were derived from plants, especially herbs. Harohally et al. (2017) tried to enhanced the antiaflatoxigenic activity of 2-hydroxy-4-methoxybenzaldehyde, cinnamaldehyde and the molecules with similar chemical structures, and the productions, Schiff bases, which showed great commendable antiaflatoxigenic activity. Plus, the antiaflatoxigenic potential of cinnamaldehyde was attributed to alleviate oxidative stress, which was possibly induced by modifications of cellular structure (Sun et al. 2016). Meanwhile, cuminaldehyde thiosemicarbazone was structurally modified, and then it was testified to be more special for aflatoxin control via yeast experimental model (Degola et al. 2017).
Similarly, essential oils were mainly separated from medical plants, different from other antifungal agents; most of these oils were mixtures with complicated composition. As reported, the bioactive ethylene-vinyl alcohol copolymer (EVOH) films, containing cinnamaldehyde, linalool, isoeugenol, or citral, showed strong inhibition of OTA biosynthesis in corn at very low concentrations (Tarazona et al. 2018). And Mateo et al. (2017) compared several oils contained in EVOH films; the result showed that EVOH-cinnamaldehyde film showed the most inhibition of fungal growth and aflatoxin control. Natural essential oil derived from turmeric was used as an eco-friendly antifungal agent. Moreover, expression of mycotoxin genes in aflatoxin biosynthetic pathway was relatively downregulated (Hu et al. 2017). The essential oil of rosemary was also applied to the investigation of multiple pathogens, including Alternaria spp., Fusarium spp., Colletotrichum spp., and Penicillium spp., all with good control practice (Waithaka et al. 2017). Moreover, similar essential oils with antifungal properties were extracted from other plants, such as Hedychium spicatum L. (Kalagatur et al. 2018a), Mentha cardiaca L. (Dwivedy et al. 2017), and so on. Clove oil-in-water nanoemulsions were one new material with different oil composition, and addition of corn oil or triacylglycerol would reduce its antifungal potential to this material, while the inhibition activity would be enhanced when clove oil was encapsulated in nanoemulsions (Wan et al. 2018).
Actually, all the pure antifungal agents were from multiple organisms; as a result, extractions from these can also be of antifungal potential. Researchers investigated the reduction rate of grape pomace to varied mycotoxins, and it can reduce the gastrointestinal absorption of mycotoxins, as well as be a natural binder for multiple mycotoxins (Gambacorta et al. 2016). In the extraction of peanut, peanut stilbenoids could provide the new strategies for preventing plant invasion and production of aflatoxin (Sobolev et al. 2018). Furthermore, the fermentation liquid of Bacillus subtilis also showed great growth inhibition to varied fungal pathogens, for its volatile organic compounds in the metabolites (Gao et al. 2018) (Fig. 9.11).

Initial apricot kernels (a), final product (b) and reject product (c) obtained by e-sorting and discolored apricot kernel (a, b) and healthy apricot kernels. (Zivoli et al. 2016)
6 Conclusion
To sum up, physical treatments for control of mycotoxin contamination were aimed at the molecular structure of mycotoxins; change of multiple physical parameters leads to the destruction or modification of their carbon skeleton, resulting in the removal of origin mycotoxins. On the other hand, for chemical treatments, most detoxifiers were aimed at the antifungal property, thus leading to the reduction of mycotoxins. Chemical reagents are too large of a group; some are broad antifungal agents, and some are specific, also with little certain microstructures to absorb mycotoxins. Unlike other emerging bio-sourced enzymes, physical and chemical detoxifiers were always the direct, simple, and broad ways to remove mycotoxins, or we can conclude that pretreatment of foodstuffs would always be these traditional ways to remove mycotoxins and protect food safety. Industrial production should be a processing chain, from treatment to the early generation and then to the final package and delivery; among these, before treatment, either the physical or chemical pretreatments show their indispensability in practical uses.
References
Aichinger G, Puntscher H, Beisl J, Kutt ML, Warth B, Marko D (2018) Delphinidin protects colon carcinoma cells against the genotoxic effects of the mycotoxin altertoxin II. Toxicol Lett 284:136–142. https://doi.org/10.1016/j.toxlet.2017.12.002
Assaf JC, El Khoury A, Atoui A, Louka N, Chokr A (2018) A novel technique for aflatoxin M1 detoxification using chitin or treated shrimp shells: in vitro effect of physical and kinetic parameters on the binding stability. Appl Microbiol Biotechnol 102(15):6687–6697. https://doi.org/10.1007/s00253-018-9124-0
Bilska K, Stuper-Szablewska K, Kulik T (2018) Resistance-related l-pyroglutamic acid affects the biosynthesis of trichothecenes and phenylpropanoids by F. graminearum sensu stricto. Toxins 10(12). https://doi.org/10.3390/toxins10120492
Boxi SS, Mukherjee K, Paria S (2016) Ag doped hollow TiO2 nanoparticles as an effective green fungicide against Fusarium solani and Venturia inaequalis phytopathogens. Nanotechnology 27(8):085103. https://doi.org/10.1088/0957-4484/27/8/085103
Buhrow LM, Cram D, Tulpan D, Foroud NA, Loewen MC (2016) Exogenous abscisic acid and gibberellic acid elicit opposing effects on Fusarium graminearum infection in wheat. Phytopathology 106(9):986–996. https://doi.org/10.1094/phyto-01-16-0033-r
Caceres I, El Khoury R, Bailly S, Oswald IP, Puel O, Bailly JD (2017) Piperine inhibits aflatoxin B1 production in Aspergillus flavus by modulating fungal oxidative stress response. Fungal Genet Biol 107:77–85. https://doi.org/10.1016/j.fgb.2017.08.005
Chang W, Liu J, Zhang M, Shi H, Zheng S, Jin X, Gao Y, Wang S, Ji A, Lou H (2018) Efflux pump-mediated resistance to antifungal compounds can be prevented by conjugation with triphenylphosphonium cation. Nat Commun 9(1):5102. https://doi.org/10.1038/s41467-018-07633-9
da Cruz CL, Delgado J, Patriarca A, Rodriguez A (2019) Differential response to synthetic and natural antifungals by Alternaria tenuissima in wheat simulating media: growth, mycotoxin production and expression of a gene related to cell wall integrity. Int J Food Microbiol 292:48–55. https://doi.org/10.1016/j.ijfoodmicro.2018.12.005
da Silva Cardoso V, Vermelho AB, Ribeiro de Lima CA, Mendes de Oliveira J, Freire de Lima ME, Pinto da Silva LH, Direito GM, Miranda Danelli MD (2016) Antigenotoxic effect of piperine in broiler chickens intoxicated with aflatoxin B1. Toxins 8(11). https://doi.org/10.3390/toxins8110316
Degola F, Bisceglie F, Pioli M, Palmano S, Elviri L, Pelosi G, Lodi T, Restivo FM (2017) Structural modification of cuminaldehyde thiosemicarbazone increases inhibition specificity toward aflatoxin biosynthesis and sclerotia development in Aspergillus flavus. Appl Microbiol Biotechnol. https://doi.org/10.1007/s00253-017-8426-y
Diao X, Han Y, Liu C (2018) The fungicidal activity of Tebuconazole enantiomers against Fusarium graminearum and its selective effect on DON production under different conditions. J Agric Food Chem 66(14):3637–3643. https://doi.org/10.1021/acs.jafc.7b05483
Duan Y, Xiao X, Li T, Chen W, Wang J, Fraaije BA, Zhou M (2018) Impact of epoxiconazole on Fusarium head blight control, grain yield and deoxynivalenol accumulation in wheat. Pestic Biochem Physiol 152:138–147. https://doi.org/10.1016/j.pestbp.2018.09.012
Durek J, Schluter O, Roscher A, Durek P, Frohling A (2018) Inhibition or stimulation of ochratoxin a synthesis on inoculated barley triggered by diffuse coplanar surface barrier discharge plasma. Front Microbiol 9:2782. https://doi.org/10.3389/fmicb.2018.02782
Dwivedy AK, Prakash B, Chanotiya CS, Bisht D, Dubey NK (2017) Chemically characterized Mentha cardiaca L. essential oil as plant based preservative in view of efficacy against biodeteriorating fungi of dry fruits, aflatoxin secretion, lipid peroxidation and safety profile assessment. Food Chem Toxicol: Int J Published Br Ind Biol Res Assoc 106(Pt A):175–184. https://doi.org/10.1016/j.fct.2017.05.043
El Khoury R, Choque E, El Khoury A, Snini SP (2018) OTA prevention and detoxification by actinobacterial strains and activated carbon fibers: preliminary results. Toxins 10(4). https://doi.org/10.3390/toxins10040137
Eroglu N, Emekci M, Athanassiou CG (2017) Applications of natural zeolites on agriculture and food production. J Sci Food Agric 97(11):3487–3499. https://doi.org/10.1002/jsfa.8312
Ferruz E, Atanasova-Penichon V, Bonnin-Verdal MN, Marchegay G, Pinson-Gadais L, Ducos C, Loran S, Arino A, Barreau C, Richard-Forget F (2016) Effects of phenolic acids on the growth and production of T-2 and HT-2 toxins by Fusarium langsethiae and F. sporotrichioides. Molecules (Basel, Switzerland) 21(4):449. https://doi.org/10.3390/molecules21040449
Gambacorta L, Pinton P, Avantaggiato G, Oswald IP, Solfrizzo M (2016) Grape pomace, an agricultural byproduct reducing mycotoxins absorption: in vivo assessment in pig using urinary biomarkers. J Agric Food Chem. https://doi.org/10.1021/acs.jafc.6b02146
Gao H, Li P, Xu X, Zeng Q, Guan W (2018) Research on volatile organic compounds from Bacillus subtilis CF-3: biocontrol effects on fruit fungal pathogens and dynamic changes during fermentation. Front Microbiol 9:456. https://doi.org/10.3389/fmicb.2018.00456
Gao D, Li Y, Lyu B, Lyu L, Chen S, Ma J (2019) Construction of durable antibacterial and anti-mildew cotton fabric based on P(DMDAAC-AGE)/Ag/ZnO composites. Carbohydr Polym 204:161–169. https://doi.org/10.1016/j.carbpol.2018.09.087
Gauthier L, Bonnin-Verdal MN, Marchegay G, Pinson-Gadais L, Ducos C, Richard-Forget F, Atanasova-Penichon V (2016) Fungal biotransformation of chlorogenic and caffeic acids by Fusarium graminearum: new insights in the contribution of phenolic acids to resistance to deoxynivalenol accumulation in cereals. Int J Food Microbiol 221:61–68. https://doi.org/10.1016/j.ijfoodmicro.2016.01.005
Gbashi S, Madala NE, De Saeger S, De Boevre M, Njobeh PB (2019) Numerical optimization of temperature-time degradation of multiple mycotoxins. Food Chem Toxicol 125:289–304. https://doi.org/10.1016/j.fct.2019.01.009
Guimaraes A, Santiago A, Teixeira JA, Venancio A, Abrunhosa L (2018) Anti-aflatoxigenic effect of organic acids produced by Lactobacillus plantarum. Int J Food Microbiol 264:31–38. https://doi.org/10.1016/j.ijfoodmicro.2017.10.025
Harohally NV, Cherita C, Bhatt P, Anu Appaiah KA (2017) Antiaflatoxigenic and antimicrobial activities of Schiff bases of 2-hydroxy-4-methoxybenzaldehyde, cinnamaldehyde, and similar aldehydes. J Agric Food Chem 65(40):8773–8778. https://doi.org/10.1021/acs.jafc.7b02576
Hellin P, Scauflaire J, Van Hese V, Munaut F, Legreve A (2017) Sensitivity of Fusarium culmorum to triazoles: impact of trichothecene chemotypes, oxidative stress response and genetic diversity. Pest Manag Sci 73(6):1244–1252. https://doi.org/10.1002/ps.4450
Hernandez-Falcon TA, Monter-Arciniega A, Cruz-Cansino NDS, Alanis-Garcia E, Rodriguez-Serrano GM, Castaneda-Ovando A, Garcia-Garibay M, Ramirez-Moreno E, Jaimez-Ordaz J (2018) Effect of thermoultrasound on aflatoxin M1 levels, physicochemical and microbiological properties of milk during storage. Ultrason Sonochem 48:396–403. https://doi.org/10.1016/j.ultsonch.2018.06.018
Hojnik N, Cvelbar U, Tavcar-Kalcher G, Walsh JL, Krizaj I (2017) Mycotoxin decontamination of food: cold atmospheric pressure plasma versus “classic” decontamination. Toxins 9(5). https://doi.org/10.3390/toxins9050151
Hu Y, Zhang J, Kong W, Zhao G, Yang M (2017) Mechanisms of antifungal and anti-aflatoxigenic properties of essential oil derived from turmeric (Curcuma longa L.) on Aspergillus flavus. Food Chem 220:1–8. https://doi.org/10.1016/j.foodchem.2016.09.179
Hua-Li X, Yang B, Raza H, Hu-Jun W, Lu-Mei P, Mi-Na N, Xiao-Yan C, Yi W, Yong-Cai L (2018) Detection of NEO in muskmelon fruits inoculated with Fusarium sulphureum and its control by postharvest ozone treatment. Food Chem 254:193–200. https://doi.org/10.1016/j.foodchem.2018.01.149
Huang Z, Zhang L, Wang Y, Gao H, Li X, Huang X, Huang T (2019) Effects of rutin and its derivatives on citrinin production by Monascus aurantiacus Li AS3.4384 in liquid fermentation using different types of media. Food Chem 284:205–212. https://doi.org/10.1016/j.foodchem.2019.01.109
Janic Hajnal E, Colovic R, Pezo L, Orcic D, Vukmirovic D, Mastilovic J (2016) Possibility of Alternaria toxins reduction by extrusion processing of whole wheat flour. Food Chem 213:784–790. https://doi.org/10.1016/j.foodchem.2016.07.019
Janic Hajnal E, Mastilovic J, Bagi F, Orcic D, Budakov D, Kos J, Savic Z (2019) Effect of wheat milling process on the distribution of Alternaria toxins. Toxins 11(3):139. https://doi.org/10.3390/toxins11030139
Jurick WM 2nd, Macarisin O, Gaskins VL, Janisiewicz WJ, Peter KA, Cox KD (2019) Baseline sensitivity of Penicillium spp. to difenoconazole. Plant Dis 103(2):331–337. https://doi.org/10.1094/pdis-05-18-0860-re
Kalagatur NK, Karthick K, Allen JA, Nirmal Ghosh OS, Chandranayaka S, Gupta VK, Krishna K, Mudili V (2017) Application of activated carbon derived from seed shells of Jatropha curcas for decontamination of zearalenone mycotoxin. Front Pharmacol 8:760. https://doi.org/10.3389/fphar.2017.00760
Kalagatur NK, Kamasani JR, Siddaiah C, Gupta VK, Krishna K, Mudili V (2018a) Combinational inhibitory action of Hedychium spicatum L. essential oil and gamma-radiation on growth rate and mycotoxins content of Fusarium graminearum in maize: response surface methodology. Front Microbiol 9:1511. https://doi.org/10.3389/fmicb.2018.01511
Kalagatur NK, Nirmal Ghosh OS, Sundararaj N, Mudili V (2018b) Antifungal activity of chitosan nanoparticles encapsulated with Cymbopogon martinii essential oil on plant pathogenic Fungi Fusarium graminearum. Front Pharmacol 9:610. https://doi.org/10.3389/fphar.2018.00610
Kang F, Ge Y, Hu X, Goikavi C, Waigi MG, Gao Y, Ling W (2016) Understanding the sorption mechanisms of aflatoxin B1 to kaolinite, illite, and smectite clays via a comparative computational study. J Hazard Mater 320:80–87. https://doi.org/10.1016/j.jhazmat.2016.08.006
Karlovsky P, Suman M, Berthiller F, De Meester J, Eisenbrand G, Perrin I, Oswald IP, Speijers G, Chiodini A, Recker T, Dussort P (2016) Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res 32(4):179–205. https://doi.org/10.1007/s12550-016-0257-7
Kulik T, Stuper-Szablewska K, Bilska K, Busko M, Ostrowska-Kolodziejczak A, Zaluski D, Perkowski J (2017a) Sinapic acid affects phenolic and Trichothecene profiles of F. culmorum and F. graminearum Sensu Stricto. Toxins 9(9). https://doi.org/10.3390/toxins9090264
Kulik T, Stuper-Szablewska K, Bilska K, Busko M, Ostrowska-Kolodziejczak A, Zaluski D, Perkowski J (2017b) trans-cinnamic and chlorogenic acids affect the secondary metabolic profiles and ergosterol biosynthesis by Fusarium culmorum and F. graminearum sensu stricto. Toxins 9(7). https://doi.org/10.3390/toxins9070198
Laborda P, Li C, Zhao Y, Tang B, Ling J, He F, Liu F (2019) Antifungal metabolite p-aminobenzoic acid (pABA): mechanism of action and efficacy for the biocontrol of pear bitter rot disease. J Agric Food Chem 67(8):2157–2165. https://doi.org/10.1021/acs.jafc.8b05618
Lai T, Wang Y, Fan Y, Zhou Y, Bao Y, Zhou T (2017) The response of growth and patulin production of postharvest pathogen Penicillium expansum to exogenous potassium phosphite treatment. Int J Food Microbiol 244:1–10. https://doi.org/10.1016/j.ijfoodmicro.2016.12.017
Li W, Yuan S, Sun J, Li Q, Jiang W, Cao J (2018) Ethyl p-coumarate exerts antifungal activity in vitro and in vivo against fruit Alternaria alternata via membrane-targeted mechanism. Int J Food Microbiol 278:26–35. https://doi.org/10.1016/j.ijfoodmicro.2018.04.024
Li J, Duan Y, Bian C, Pan X, Yao C, Wang J, Zhou M (2019) Effects of validamycin in controlling Fusarium head blight caused by Fusarium graminearum: inhibition of DON biosynthesis and induction of host resistance. Pestic Biochem Physiol 153:152–160. https://doi.org/10.1016/j.pestbp.2018.11.012
Limay-Rios V, Schaafsma AW (2018) Effect of prothioconazole application timing on Fusarium mycotoxin content in maize grain. J Agric Food Chem 66(19):4809–4819. https://doi.org/10.1021/acs.jafc.8b00788
Liu J, Sui Y, Wisniewski M, Xie Z, Liu Y, You Y, Zhang X, Sun Z, Li W, Li Y, Wang Q (2018) The impact of the postharvest environment on the viability and virulence of decay fungi. Crit Rev Food Sci Nutr 58(10):1681–1687. https://doi.org/10.1080/10408398.2017.1279122
Long M, Yang SH, Han JX, Li P, Zhang Y, Dong S, Chen X, Guo J, Wang J, He JB (2016) The protective effect of grape-seed proanthocyanidin extract on oxidative damage induced by Zearalenone in Kunming mice liver. Int J Mol Sci 17(6). https://doi.org/10.3390/ijms17060808
Magro M, Moritz DE, Bonaiuto E, Baratella D, Terzo M, Jakubec P, Malina O, Cepe K, Aragao GMF, Zboril R, Vianello F (2016) Citrinin mycotoxin recognition and removal by naked magnetic nanoparticles. Food Chem 203:505–512. https://doi.org/10.1016/j.foodchem.2016.01.147
Markovic M, Dakovic A, Rottinghaus GE, Kragovic M, Petkovic A, Krajisnik D, Milic J, Mercurio M, de Gennaro B (2016) Adsorption of the mycotoxin zearalenone by clinoptilolite and phillipsite zeolites treated with cetylpyridinium surfactant. Colloids Surf B Biointerfaces 151:324–332. https://doi.org/10.1016/j.colsurfb.2016.12.033
Mateo EM, Gomez JV, Dominguez I, Gimeno-Adelantado JV, Mateo-Castro R, Gavara R, Jimenez M (2017) Impact of bioactive packaging systems based on EVOH films and essential oils in the control of aflatoxigenic fungi and aflatoxin production in maize. Int J Food Microbiol 254:36–46. https://doi.org/10.1016/j.ijfoodmicro.2017.05.007
Moon YS, Choi WS, Park ES, Bae IK, Choi SD, Paek O, Kim SH, Chun HS, Lee SE (2016) Antifungal and antiaflatoxigenic methylenedioxy-containing compounds and piperine-like synthetic compounds. Toxins 8(8). https://doi.org/10.3390/toxins8080240
Moon YS, Kim L, Chun HS, Lee SE (2017) 4-Hydroxy-7-methyl-3-phenylcoumarin suppresses aflatoxin biosynthesis via downregulation of aflK expressing versicolorin B synthase in Aspergillus flavus. Molecules 22(5):712. https://doi.org/10.3390/molecules22050712
Nones J, Solhaug A, Eriksen GS, Macuvele DLP, Poli A, Soares C, Trentin AG, Riella HG, Nones J (2017) Bentonite modified with zinc enhances aflatoxin B1 adsorption and increase survival of fibroblasts (3T3) and epithelial colorectal adenocarcinoma cells (Caco-2). J Hazard Mater 337:80–89. https://doi.org/10.1016/j.jhazmat.2017.04.068
Paul PA, Bradley CA (2018) Meta-analysis of the effects of QoI and DMI fungicide combinations on Fusarium head blight and deoxynivalenol in wheat. Plant Dis 102(12):2602–2615. https://doi.org/10.1094/pdis-02-18-0211-re
Peng K, Jin L, Niu YD, Huang Q, McAllister TA, Yang HE, Denise H, Xu Z, Acharya S, Wang S, Wang Y (2017) Condensed tannins affect bacterial and fungal microbiomes and mycotoxin production during ensiling and upon aerobic exposure. Appl Environ Microbiol. https://doi.org/10.1128/aem.02274-17
Scaglioni PT, Blandino M (2018) Application of fungicides and microalgal phenolic extracts for the direct control of Fumonisin contamination in maize. J Agric Food Chem 66(19):4835–4841. https://doi.org/10.1021/acs.jafc.8b00540
Shannon TA, Ledoux DR, Rottinghaus GE, Shaw DP, Dakovic A, Markovic M (2016) The efficacy of raw and concentrated bentonite clay in reducing the toxic effects of aflatoxin in broiler chicks. Poult Sci. https://doi.org/10.3382/ps/pew408
Shi H (2017) Structures of degradation products and degradation pathways of aflatoxin B1 by high-voltage atmospheric cold plasma (HVACP) treatment. J Agric Food Chem 65(30):6222–6230. https://doi.org/10.1021/acs.jafc.7b01604
Siciliano I, Dal Bello B, Zeppa G, Spadaro D, Gullino ML (2017) Static hot air and infrared rays roasting are efficient methods for aflatoxin decontamination on hazelnuts. Toxins 9(2). https://doi.org/10.3390/toxins9020072
Sobolev V, Arias R, Goodman K, Walk T, Orner V, Faustinelli P, Massa A (2018) Suppression of aflatoxin production in Aspergillus species by selected peanut (Arachis hypogaea) stilbenoids. J Agric Food Chem 66(1):118–126. https://doi.org/10.1021/acs.jafc.7b04542
Sobral MMC, Cunha SC, Faria MA, Martins ZE, Ferreira I (2019) Influence of oven and microwave cooking with the addition of herbs on the exposure to multi-mycotoxins from chicken breast muscle. Food Chem 276:274–284. https://doi.org/10.1016/j.foodchem.2018.10.021
Sueck F, Poor M, Faisal Z, Gertzen CGW, Cramer B (2018) Interaction of ochratoxin a and its thermal degradation product 2′R-ochratoxin a with human serum albumin. Toxins 10(7). https://doi.org/10.3390/toxins10070256
Sun Q, Shang B, Wang L, Lu Z, Liu Y (2016) Cinnamaldehyde inhibits fungal growth and aflatoxin B1 biosynthesis by modulating the oxidative stress response of Aspergillus flavus. Appl Microbiol Biotechnol 100(3):1355–1364. https://doi.org/10.1007/s00253-015-7159-z
Sun Y, Wang Y, Han LR, Zhang X, Feng JT (2017) Antifungal activity and action mode of cuminic acid from the seeds of Cuminum cyminum L. against Fusarium oxysporum f. sp. Niveum (FON) causing Fusarium wilt on watermelon. Molecules 22(12). https://doi.org/10.3390/molecules22122053
Tarazona A, Gomez JV, Gavara R, Mateo-Castro R, Gimeno-Adelantado JV, Jimenez M, Mateo EM (2018) Risk management of ochratoxigenic fungi and ochratoxin A in maize grains by bioactive EVOH films containing individual components of some essential oils. Int J Food Microbiol 269:107–119. https://doi.org/10.1016/j.ijfoodmicro.2018.02.002
Ten Bosch L, Pfohl K, Avramidis G, Wieneke S, Viol W, Karlovsky P (2017) Plasma-based degradation of mycotoxins produced by Fusarium, Aspergillus and Alternaria species. Toxins 9(3). https://doi.org/10.3390/toxins9030097
Voss K, Ryu D (2017) Reduction of Fumonisin toxicity by extrusion and Nixtamalization (alkaline cooking). J Agric Food Chem 65(33):7088–7096. https://doi.org/10.1021/acs.jafc.6b05761
Waithaka PN, Gathuru EM, Githaiga BM, Kimani SN (2017) Control of passion fruit fungal diseases using essential oils extracted from rosemary (Rosmarinus officinalis) and eucalyptus (Eucalyptus agglomerata) in Egerton University Main Campus Njoro, Kenya. Int J Microbiol 2017:2814581. https://doi.org/10.1155/2017/2814581
Wan J, Zhong S, Schwarz P, Chen B, Rao J (2018) Influence of oil phase composition on the antifungal and mycotoxin inhibitory activity of clove oil nanoemulsions. Food Funct 9(5):2872–2882. https://doi.org/10.1039/c7fo02073b
Wang L, Dai Y, Chen W, Shao Y, Chen F (2016) Effects of light intensity and color on the biomass, extracellular red pigment, and citrinin production of Monascus ruber. J Agric Food Chem 64(50):9506–9514. https://doi.org/10.1021/acs.jafc.6b04056
Wang G, Lian C, Xi Y, Sun Z, Zheng S (2018a) Evaluation of nonionic surfactant modified montmorillonite as mycotoxins adsorbent for aflatoxin B1 and zearalenone. J Colloid Interface Sci 518:48–56. https://doi.org/10.1016/j.jcis.2018.02.020
Wang G, Miao Y, Sun Z, Zheng S (2018b) Simultaneous adsorption of aflatoxin B1 and zearalenone by mono- and di-alkyl cationic surfactants modified montmorillonites. J Colloid Interface Sci 511:67–76. https://doi.org/10.1016/j.jcis.2017.09.074
Wang C, Guo L, Yao J, Wang A, Gao F, Zhao X, Zeng Z, Wang Y, Sun C, Cui H, Cui B (2019a) Preparation, characterization and antifungal activity of pyraclostrobin solid nanodispersion by self-emulsifying technique. Pest Manag Sci. https://doi.org/10.1002/ps.5390
Wang G, Xi Y, Lian C, Sun Z, Zheng S (2019b) Simultaneous detoxification of polar aflatoxin B1 and weak polar zearalenone from simulated gastrointestinal tract by zwitterionic montmorillonites. J Hazard Mater 364:227–237. https://doi.org/10.1016/j.jhazmat.2018.09.071
Yamauchi S, Nishiwaki H, Nakazaki S, Akiyama K (2017) Structure-antifungal activity relationship of fluorinated dihydroguaiaretic acid derivatives and preventive activity against Alternaria alternata Japanese pear pathotype. J Agric Food Chem. https://doi.org/10.1021/acs.jafc.7b01896
Yang G, Jin Q, Xu C, Fan S, Wang C, Xie P (2017) Synthesis, characterization and antifungal activity of coumarin-functionalized chitosan derivatives. Int J Biol Macromol. https://doi.org/10.1016/j.ijbiomac.2017.08.009
Zhang NY, Qi M, Zhao L, Zhu MK, Guo J, Liu J, Gu CQ, Rajput SA, Krumm CS, Qi DS, Sun LH (2016) Curcumin prevents aflatoxin B(1) hepatoxicity by inhibition of cytochrome P450 isozymes in chick liver. Toxins 8(11). https://doi.org/10.3390/toxins8110327
Zhang J, Peng JF, Wang T, Kang Y, Jing S, Zhang ZT (2017) Synthesis and biological evaluation of arylpyrazoles as fungicides against phytopathogenic fungi. Mol Divers. https://doi.org/10.1007/s11030-017-9727-x
Zhang Q, Zhang Z, Tang B, Gao B, Tian M, Sanganyado E (2018) Mechanistic insights into stereospecific bioactivity and dissipation of chiral fungicide triticonazole in agricultural management. J Agric Food Chem 66(28):7286–7293. https://doi.org/10.1021/acs.jafc.8b01771
Zhu F (2018) Effect of ozone treatment on the quality of grain products. Food Chem 264:358–366. https://doi.org/10.1016/j.foodchem.2018.05.047
Zivoli R, Gambacorta L, Piemontese L, Solfrizzo M (2016) Reduction of aflatoxins in apricot kernels by electronic and manual color sorting. Toxins 8(1). https://doi.org/10.3390/toxins8010026
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Hu, D., Wu, A. (2019). Chemical and Physical Treatments for Reducing Mycotoxin Contaminations. In: Wu, A. (eds) Food Safety & Mycotoxins. Springer, Singapore. https://doi.org/10.1007/978-981-32-9038-9_9
Download citation
DOI: https://doi.org/10.1007/978-981-32-9038-9_9
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-32-9037-2
Online ISBN: 978-981-32-9038-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)