
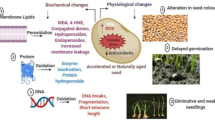
Abstract
In the perspective of seed technology, physical methods that were generally used to increase the plant production provide advantages as compared to conventional methods that use chemical substances. The impacts of physical treatments for invigoration in seeds tended to be addressed at different levels that include changes in the gene expression, the structural and morphological aspects, and metabolite or protein accumulation. The physical methods used for the pre-sowing treatments are irradiation with microwaves (MWs), magneto-priming, and ionizing radiations (IRs). “Magneto-priming” depends upon the use of magnetic fields and portrayed as a cheap, noninvasive, and eco-friendly technique with demonstrated gainful impacts on vigor, crop yield, and seed germination. Radiation such as gamma rays and X-rays has been generally viewed as a powerful contrivance in food technology and agricultural sciences. Positive impacts on seedling vigor, health, and germination were aroused by ultraviolet radiations (UV) that include UV-C and UV-A. Among all the identified physical treatments, a connected and basic research is yet expected to characterize the genotype, optimal dose required, and irradiation conditions that all depend on environment. Electron spin resonance has a huge prospective in regard to seed technology not correctly scrutinized to a point in time to end the treatment. Physical methods for the seed invigoration were described in the current chapter as well as emphasized to address the needs of seed technologists, trade makers, and producers by using the physical methods.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
The greatest difficulties that humanity is confronting are enhancing the supportability of agriculture while decreasing its ecological effect, to meet the food demand of the developing worldwide population (Edmondson et al. 2014). The impression of agricultural manageability depends on creative innovations and new methods with no antagonistic consequences on nonmarket goods.
Seeds with high vigor are intermediary of sustainable productivity and establishment of crop. The estimated seed market of the European Union in 2012 stretched up to 7 billion Euros as compared to 20% of world market, positioning third after China and the USA. Chemical compounds were generally used in modern agriculture, while the utilization of physical methods may speak in this monetarily focused on setting, inventive biotech/molecular tools, products, and treatments which are critical to accelerate the consolidation procedure of the seed industry.
Despite the extensive role of chemical methods in modern agriculture, the physical methods can also be able to play an alternative role to boost up the production in agriculture and can also be used for the storage and plant protection (Aladjadjiyan 2012). More benefits were obtained by adopting physical methods for seed invigoration as compared to the extensively used chemical treatments that involve the use of chemical substances. One of the advantages of physical methods is that they decrease the use of fertilizers, thereby lessening the contamination of the raw materials produced on the farm. Another benefit provided by the physical method is the disinfection of the seed during storage and before sowing (Aladjadjiyan 2012). Conceivable methodologies include ionizing radiation (IR), electromagnetic waves (EW), ultrasound waves (UW), and magnetic fields (MF). Extensively studied topic of the twentieth century is the treatment of the food using irradiation to ensure the greater shelflife and microbiological security; as a result, there is more concern towards the use of irradiation for the security of the food supply (Farkas and Mohácsi-Farkas 2011). The space research has shown great interest in studying the potential effect of irradiation on living organisms. Experiments revealed that Gamma rays and X-rays impede the growth of crop plant (Wolff et al. 2014; Arena et al. 2014). Because of such restriction, the cultivation of higher plants in space was impossible without new agriculture-related technologies that help to understand the possible effect of space factors on the environment but also on the development of greenhouse (De Micco et al. 2014). There is also a need to address the problems related to the development and the growth of plants in such extraterrestrial environment (Wolff et al. 2014).
Effect of physical treatments for invigoration in seeds tended to be addressed at different levels that include changes in the gene expression. Plant system serves as exceptional representations of the biological response to various factors related to environment that include the mutation in the cell due to certain chemical and physical substances (Macovei et al. 2014; Zaka et al. 2002; Confalonieri et al. 2014; Balestrazzi et al. 2011).
Effect of physical treatment on plants depends upon various factors such as MF or radiation (complete dose, its rate and type) and plant characteristics, i.e., cultivar, species, age, ploidy, and complexity of the target tissue or organ (De Micco et al. 2014). Recently, many interesting reports were published in toxological sciences about the usage of very minute concentation of toxins for the purpose of stimulation which is called hormesis (Belz and Piepho 2012; Calabrese and Baldwin 2000). It has been stated that germination, production of the photosynthetic pigment, and plant growth were stimulated by doing seed irradiation (Macovei et al. 2014; Kovács and Keresztes 2002; De Micco et al. 2014). However, the reports also stated the disastrous impacts of these irradiations were also seen in case of long exposure.
Priming is a well-recognized treatment that enhances the quality of the seed by temporarily stimulating the pre-germinative metabolism that in turn activates the processes related to DNA repair and antioxidant functions (Paparella et al. 2015). Priming of seed has arisen as an effective method to increase seed germination and seed vigor; seed priming also proved to be an effective method during field establishment and for seed growth during adversative environmental condition (Hussain et al. 2015; Ventura et al. 2012). By using treatments like osmopriming and hydropriming, the low vigor seed can be cured. In the perspective of seed technology, physical methods were proved to be more efficient when compared to the methods that were conventionally used (Bilalis et al. 2012). Extensively used physical method in the field of agriculture for pre-sowing treatment of seed was the usage of magnetic field (Aguilar et al. 2009). Magnetic field technique was generally noninvasive, cheap, and eco-friendly (Efthimiadou et al. 2014; Bilalis et al. 2012). This review will also address other physical treatments like UV radiation, microwave radiation, gamma rays, and X-rays.
The current manuscript addresses some of the extensively used physical methods of seed treatment and highlighted the possible advantages and disadvantages of these physical methods. Also, the future perception related to the use of the technique will also be discussed to meet the needs of the trade makers, consumers, and seed producers.
2 Magneto-Priming: An Appropriate Invigoration Protocol
The effects produced by the magnetic field in living organism are extensively studied. Magnetic field of the Earth is (50 μT) as stated by Belyavskaya (2004). The unit of magnetic flux density 1 tesla (T) is equal to 1 kg s−2 A−1; here, A stands for Ampere that is defined as electric current flowing with a charge of 1 coulomb per second. For the first time, the impacts of magnetic field on plants were described by Krylov and Tarakanova (1960); the phenomenon of magnetotropism was described as the auxin-like impact produced on the germinating seed by magnetic field. The techniques used in agriculture for the purpose of seed priming are electromagnetic field (EMF) and static magnetic field (SMF) that produced positive effects on crop yield, seed vigor, and seed germination (Baby et al. 2011). Static magnetic field (SMF) is considerably different from electromagnetic field (EMF). Static magnetic field can only be produced by a permanent magnet as produced by earth magnetic field or by the help of industrial processes; electromagnetic field is produced by electrically charged entities being extended indeterminately in space (Mitchell and Cambrosio 1997).
In the perspective of seed technology, the possible effects exerted by magnetic field were studied in order to derive the full potential of the magnetic field for the new seed treatments. The beneficial impacts produced by the magnetic field in order to improve the accumulation of the biomass and germination parameters were discussed for a wide range of plants by Teixeira da Silva and Dobránszki (2015). The current studies tested range of the magnetic field from 0 to 300 mT. Seeds that were primed with magnets show improved vigor, germination rates, root development, and seedling biomass. Tolerance to abiotic and biotic stress was increased by using magnetic field treatment (De Souza et al. 2006; Anand et al. 2012). Bhardwaj et al. (2012) described the enhanced antioxidant activities of catalase (CAT), glutathione reductase (GR), and superoxide dismutase (SOD) in magneto-primed cucumber (Cucumis sativus L.) seeds. Soybean (Glycine max (L.) Merr. var. Js-335) seeds that were magneto-primed produce lower amount of superoxide radicals (\( {\mathrm{O}}_2^{\bullet \hbox{--} } \)) (Baby et al. 2011). Therefore, magnetic field treatments have the added potentiality to be used for diminishing the disease-induced or drought effects on the productivity of crop.
On the basis of underlying assumptions, a tremendous interest was generated to study the biochemical, physiological, and molecular mechanisms involved to gradually improve the performance of the plants with magneto-primed seed. (Baby et al. 2011; Vashisth and Nagarajan 2010; Anand et al. 2012; Javed et al. 2011; Table 1) like sunflower (Helianthus annuus L.) seeds. Magnetic field acts as a priming agent in such a way to enhance the performance of the seeds. Seeds that were treated with magnetic field show enhanced protease, dehydrogenase, and alpha-amylase activities as compared to nontreated seeds. However, higher activity level of hydrolyzing enzymes was seen in seeds treated with static magnetic field (SMF) that related to the seedling vigor, germination, and rooting traits.
A very little information is available about the mechanism of plant’s responce against magnetic field and their signal transduction pathway. Blue light photoreceptors–cryptochromes were responsible for the signaling of magnetic field in plants (Ahmad et al., 2007). However, there is still a need to fully understand the mechanisms involved in magnetobiology and the possible genotoxic impacts involved in the usage of magnetic fields (Ghodbane et al. 2013). In order to increase the seed vigor, seed germination, and photosynthetic capacity of plants primed with magnetic fields, there is still the need to fully understand the mechanisms lying behind the process.
In horticultural crop, environmental factors like humidity, light, and temperature are associated with good performance of seed, but the impact produced upon the application of environmental factors with magnetic field still remains unclear. By using the resulting knowledge, new seed treatments that were also locally adapted can be developed. Poinapen et al. (2013) exhibited that how the collective effect of magnetic field and environmental factors affect seed performance and viability in tomato (Solanum lycopersicum L.) in laboratory. It was shown that, during early stages of seed germination, relative humidity played a crucial role in modulating the performance of magneto-primed seeds.
3 Ionizing Radiation Treatments
3.1 Gamma Radiation, Most Promising Technique
Recent studies are usually based on ionizing radiation in agriculture for food security. Gamma (γ) radiation are irradiation of high energy type, so it penetrates into plant and foods easily (Jayawardena and Peiris 1988). Application of γ-rays is done usually using cobalt-60 (Moussa 2006) in the form of Gray (Gy), i.e., 1 Gy is equal to the amount of absorbed irradiation per kilogram. But when biological material was used for treatment then amount of irradiation count in sievert units. Dose rate can also be used as starting treatments. For high sterilization conditions, γ-rays are used such as microbial experiments or insect-pest pathogenic treatments. They are also used as pest prevention agent. In food security, γ-rays help to cease sprouting of vegetables and delay ripening of fruits by stopping the important enzymes and, thus, increase shelf life (Moussa 2006; Mokobia and Anomohanran 2005). Furthermore, γ-rays also cause mutation and are helpful tools in plant breeding for the purpose of new traits and varieties’ improvement (Irfaq and Nawab 2001).
Among the diverse radiobiology features, the topic that is gaining remarkable attention is the depiction of the gamma rays’ effects on seeds. Commencing study mainly focused on the impact of low-dose rate on seeds and their germination percentage. Gamma rays directly affect the proteins, nucleic acid, and cell membranes at any stage (Kovács and Keresztes 2002). However, an unintended action is described by the production of reactive oxygen species (ROS) as of water radiolysis (Esnault et al. 2010; Borzouei et al. 2010). Reactive oxygen species have the ability to damage and diffuse the cellular organelles and macromolecules. Nonetheless, strength of γ-rays depends upon dose rate, duration, and intensity. Gamma rays when delivered enhance the establishment of seedling and germination percentage and thus act as a priming agent.
In seed technology field, gamma irradiations are used for seed invigoration and as dynamic force for inspecting the molecular mechanism which involves the activation of seed response to the available physical treatment. After that, the effect of gamma rays was studied on seeds of Arabidopsis thaliana for evaluating the biological responses (Qi et al. 2015). Germination index was stimulated when seed is irradiated with total doses that were lower than 100 Gy. Similarly, the growth rate of seedling, root length, and weight were found to be significantly increased as compared to control. Among the tested doses of irradiation, A. thaliana seeds with 50 Gy dose showed the high positive impacts in all growth parameters. With γ-rays, it was also proved that it enhances all the seedling parameters along with germination performance in crops as well as woody plants. Maity et al. (2005) examined γ-ray-induced impacts on Phaseolus mungo L. and Oryza sativa L. cv-2233 seeds. Range of the radiation applied is 50–350 Gy dose. Irradiation with less doses helps to enhance the morphological traits, i.e., shoot number, number of seed per panicle, plant height, and panicle length. However, high-dose exposure is reported to bring negative effects on the seedling growth parameters. For example, in rice, the greater stimulatory dose matched to 50 Gy, while P. mungo showed improvement with 200 Gy radiations. Their effects on maize were also considered (Marcu et al. 2013).
A test was conducted to compare radiation sensitivity of plant germination rate and photosynthetic pigment contents among different seeds conditions. The triggering effects of gamma rays were also observed at 2–30 Gy as low doses and 70 Gy as high doses presented to be detrimental to plant response. Through all research, it was clear that low dose of irradiation has a constructive output on yield traits in crops, as well on the seedling establishment and germination. Two pre-sowing treatments, i.e., gamma radiation (300, 400, and 500 Gy) and simple hydropriming, were applied on okra and activated the metabolic reaction. Both treatments were applied for the improvement of seedling conditions and germination rate. Application of γ-rays with 400 Gy showed superior results in germination, quality of seeds, and yield along with photosynthetic capacity (Hegazi and Hamideldin 2010). Hence, results showed that the efficacy of dose and radiation on germination and yield traits relents on seed species.
γ-rays treatments could not only be used for crops species but also for the conservation of native species. For this purpose, seeds of Moluccella laevis L. were tested on seedling stage (Minisi et al. 2013). Corresponding to previous explanations, γ-rays treated at 5 Gy as minute dose increase the percentage of germination than untreated samples or those treated with up to 20 Gy. Low-dose γ-rays also have the ability to stimulate seed vigor, germination, and growth of seedling in deadly nightshade (Abdel-Hady et al. 2008), wild oat (Avena fatua L.) (Maherchandani 1975; Majeed et al. 2010), garden cress (Lepidium sativum L.) (Dubey et al. 2007), and okra (Abelmoschus esculentus L. Monch.) (Moussa 2006). Collective evidences were similar in all these works that the less number of doses of γ-rays resulted in advantageous action in seeds that are physically treated, which hysterics the definition of hormesis (Luckey 1980, 2006). The threshold of low dose is typically placed just above the background of natural radiation levels (2.4 millisievert per year) and demarcated by the border between bionegative and biopositive effects. Hormesis signifies an adaptive retort of organisms and cells to intermittent and moderate stresses, thus provoking the initiation of repair mechanism and cellular defense against the stress-causing agent, or else missing in the lack of stress (Calabrese and Baldwin 2000; Mattson 2008).
Molecular and biological processes which can be benifited by radiation hormesis are still under argued. Various researchers pointed out reactive oxygen species production in seed in response to γ-rays, subsequently they acted as signaling molecules and antioxidant responses in stress. Resultantly, radiation-treated plant can tolerate the common stresses, i.e., water loss, temperature variation, and light intensity during growth (Gicquel et al. 2012). Abdel-Hady et al. (2008) and Sjodin (1962) appealed that low-dose gamma irradiation enhanced enzyme activity as well as proteins and nucleic acid synthesis in seeds.
Constructed on this, it is clearly described that significant increase is achieved in seed metabolism which further breaks dormancy and improves germination of seeds. Study was also conducted for the evaluation of chronic and severe gamma irradiation effects on genetic stability and expression in seeds (Kovalchuk et al. 2007). In retort to acute radiation, the plant metabolism is moved to activation of mechanisms for pro-survival, immediate repair of the damage, and perhaps the inhibition of cell differentiation/cell division. In distinction, chronic irradiation activated an alternative response, based on gene expression that involved in general stress and metabolism of nucleic is motivated. Furthermore, the output of the following study also expresses the variation in methylation pattern and chromatin modification, which are interlinked by chronic radiation.
Through all these researches, it was obvious that efficient results were attained by low-dose treatment of γ radiations in seed technology with low-dose treatment which gives efficient results. Nonetheless, additional studies are desirable to comprehend the molecular base evaluation, essential in the enhanced growth/response detected in exposed seeds. In spite of the advantageous features cited, γ-irradiation treatments in a seed equipment perception are subject to the proper formation of γ radiation conveniences able to graft at an industrial scale. Solicitation of this seed invigoration method still involves a widespread research imperative to describe the optimum conditions (dose rate, total dose) which should be contingent to genotype, crop species, and environmental background.
3.2 How Are Seedling Development and Seed Germination Affected by X-Rays
Impact of X-rays on living organisms has not yet been completely assumed. The wavelength of X-rays from 0.01–10nm in range of electromagnetic spectrum and carry energies between 120 eV-120 KeV. In agriculture, Lenient X-rays from 0.12 to 12 keV are considered best for agriculture products (Kotwaliwale et al. 2014). In contrast, in humans, X-rays have broad-spectrum use in medical as well as in research on plants (Beard et al. 1958).
According to De Micco et al. (2014) and Einset and Collins (2015) after the 1960s, various researches have been published on the seed treated with X-ray radiations. Cumulative X-ray irradiation doses have been found to have a negative impact on root growth and germination of date palm seeds (Al-Enezi et al. 2012). On the contrary, researchers also found out stimulatory impact on growth of leaf of x-ray irradiation with 0.56 Gy radiation level. However, in tomato, with treatments of X-rays on seeds, significant increases in anatomical and ecophysiological characteristics are reported (De Micco et al. 2014). The complete results presented that development and germination of functional leaves were not dependent on doses.
In addition, during seed treatment with high dose of X-ray radiation, there are minor changes in leaves structure that structurally resulted in poor photosynthetic efficiency. The documentation of candidate species that were radiation-resistant can be a significant achievement in agricultural research (Arena et al. 2014).
Previously, very little information have been presented about the effects of X-rays on crops due to focusing only on plant physiology. New evidences related to physiological and molecular mechanisms essential for the resistance of plant tissues to X-rays could be revealed by using profiling techniques that were globally used (e.g., Omics).
3.3 Seeds Treatment with Ultraviolet Radiation
With the diminution of stratosphere ozone layer, assessment of solar ultraviolet (UV) radiation to the earth is more. Numerous studies showed the effect of UV radiation on plant and ecosystem (Kovács and Keresztes 2002; Heisler et al. 2003). A few studies reported the impact of UV radiation on seed germination and its biology (Table 2).
Since radiation is exceptionally destructive to organisms, it is not recommended for an organism under natural conditions (Hollósy 2002). Furthermore, this UV-C radiation is able to penetrate rapidly into plant tissues and is nonionizing, which maintains its prospective as a germicidal agent. Cabbage is treated with low dose of UV-C and resisted to black rot (Brown et al. 2001). Again, UV-C also has great impact on growth parameters and quality in green house. Pre-sowing treatment of lettuce with UV-C resulted in high salt stress. So, the consequences exhibited that seedlings treated with UV-C were competent to alleviate the effect of extreme salinity; it might be possible due to the enhanced free radical foraging activity in leaf tissues (Ouhibi et al. 2014).
In spite of UV-B radiation containing only 1.5% of the total spectrum, physiology of crops is also affected by radiation, including damaging of DNA, protein, and membranes resulting in restriction of growth of plant and thus reduction in photosynthesis (Choudhary and Agrawal 2014; Hideg et al. 2013). Their impacts are studied in mash bean for seedling growth, germination, and development (Shaukat et al. 2013). In contrast, authors described improved results in germination rate with UV-B light but not for germination percentage which remained same as earlier. Besides, many harmful results are clear using UV-B light, i.e., decreased length of root and shoot. Biochemically, it increases the amount of total soluble phenols and activity of tyrosine ammonia lyase.
UV-A radiation signifies about 6.3% of the received solar radiation and is considered as less dangerous (Hollósy 2002). Hamid and Jawaid (2011) examined the impacts of UV-A radiation on the seeds of mung bean and the results were auspicious and exhibited the improved results in germination rate and seedling performance with root–shoots length and leaf area. This is the only study which is accessible to the researcher for further research. However, a deeper insight is still requested to elucidate about the molecular mechanisms underlying the improvements occurring in the UV-treated seeds.
3.4 Microwaves Potentialities in Seed Technology
The main component of electromagnetic spectrum is microwave having frequency ranging from 300 MHz-300 GHz. So the electromagnetic radiation has both thermal and nonthermal effects in biological systems (Banik et al. 2003). Furthermore, different biological effects are caused by microwave such as wave forms, frequencies, duration of exposures, and modulation (Vian et al. 2006). Meanwhile, various studies show that a very limited literature is available on the effects of microwaves on humans and animals (Jayasanka and Asaeda 2014). When living tissue absorbed MW radiation, it brings dipole rotation in electron orbit and, hence, leads to the fast and selective heating. Interestingly, present studies focus on 2.45 Ghz radiations, which is occupied by molecule of water in living cells (Iuliana et al. 2013).
In seed technology before sowing and storing of seed, nonlethal MWs treatments have been widely used (Knox et al. 2013). Remarkably, growth of weeds can be prevented by MWs treatments (Sahin 2014). So, MW treatment is an alternative way of chemical control of weeds because it has no hazardous effect on environment; therefore, it is valid on greenhouse of horticulture and some ornamentals nurseries.
Nowadays, big problem in the field of agriculture is poor seed germination, but the seed invigoration treatment is the best method for enhancement rate of germination (Table 3). Effects of wireless routers, MWs, and mobile devices, were also studied in three aromatic species, i.e., Parsley, celery, and dill (Soran et al. 2014). So the seeds of these three species were bare and used to assess the effects on leaf anatomy, volatile emissions, and essential oil content. Results revealed effect of MWs on three species-induced chemical and structural modifications.
4 Effects of Physical Seed Invigoration Studies by Physical Methods
Through electron paramagnetic resonance (EPR), which analysis of short lived species, i.e., reactive oxygen species for their quantitative and qualitative, can be measured? This potent tool, presently known as one of the most sensitive and specific, upswings from the novel studies in quantum mechanics supported by Zeeman (1897). That becomes necessary to the research based on biological system with transition metals and organic radicals (Sahu et al. 2013).
Electron paramagnetic resonance active system comprises a molecular orbit containing single unpaired electron. In the absenteeism of MF, same energy is possessed by the two quantum states. When a magnetic field is activated, increase in the energy state with decrease in the energy of −1/2 state depends upon the magnetic field strength. Single unpaired electrons were able to change their orbit state, and thus, actions related to the energy absorption are supervised and then converted into a spectrum (Weil and Bolton 2006). By using trace molecules, i.e., spin probes, EPR resolution can be further improved, which are artificially presented in the biological systems that are targeted. With the ability of stable diamagnetic tracer, molecules are capable to fix the momentary reactive oxygen species and instigate long-lived paramagnetic spin adducts, i.e., radical species. The second gather in cells during hours or minutes enables detection through EPR (Sahu et al. 2013). Electron paramagnetic resonance spectroscopy is presently used in planta to dichotomize and comprehend the varieties of functions played by reactive oxygen species in stress response and plant growth regulation mechanism (Steffen-Heins and Steffens 2015).
The prospective of this instrument for the purpose of high-resolution summarizing of radicals in seeds is still abundantly exploited. EPR is ominously added to comprehend the role of free radicals in seed weakening. In soybean seeds, an unknown organic radical was identified through electron paramagnetic resonance and then further confirmed and dated in dried seeds of maize (Leprince et al. 1995). Primarily at the seed level, effect of temperature and oxygen on reactive oxygen species (ROS)-induced wound was also exposed by EPR spectra. A fascinating feature of seed practicability discovered by electron paramagnetic resonance is connected to variations in thickness and movement of molecules stirring when the cytoplasm arrives in a shiny state. Meanwhile, when seeds are stored under low temperature, the distinctiveness takes place and it has been supervised by the help of spin probes. The molecular mobility of spin probes can be slow or fast reliant on their dissemination among different seed constituents. Nitroxide spin probes, undeniably, were effectively used to inaugurate an association between occurrence of intracellular glasses, molecular mobility, and stability of seed storage (Hoekstra and Leprince 1998; Buitink et al. 1998, 1999). Temperature-dependent and moisture changes stirring in pea and Impatiens seeds are connected through rate of aging and that is verified in cytosol because of the revolving motion of a spin probe (Buitink et al. 2000).
Mostly, scientists used a nitroxide spin probe in wheat embryos to check if solid EPR was efficaciously used for efficient examination of seeds for plasma membranes reliability (Smirnov et al. 1992). So, result revealed greater than 13% moisture in the semipermeable cell wall of seed. Furthermore, study on seeds of wheat also validated the impacts of aging of the embryo (Golovina et al. 1997). Also, it reveals that permeability of plasma membrane improved more quickly during seed aging. The strategic role of permeability of membrane precision during imbibition of seeds of neem (Azadirachta indica L.) was also examined by spin probe technique using nitroxyl (Sacandè et al. 2001).
Species profiles of paramagnetic are developing as favorable to screen against stress tolerance in grains. Large values of semiquinone with carbohydrate radicals were associated with seeds that are water stress sensitive and also linked with greater starch contents by biochemical tests. Researches also revealed that stress tolerance can be identified by EPR radical profiles. Likewise, Kurdziel et al. (2015) used EPR to examine wheat grains of four genotypes through free radical profiles in embryo, endosperm, and seed coat and found various levels of water stress tolerance. Drought-sensitive genotypes having more carbohydrates are identified as selectable marker under water stress tolerance. EPR was also used to examine the effect of ozone application on barley, oat, and wheat grains against oxidative stress tolerance (Labanowska et al. 2016). Electron paramagnetic resonance exposed that the number of paramagnetic species like Cu (II), Fe (III), and Mn (II), altered upon exposure of ozone, is also dependent on the tolerance behavior of particular genotype. Furthermore, the patterns of organic radicals that are stable (carbohydrate semiquinone and phenoxyl) suggestively changed ozone treatment as given, presenting improvement of paramagnetic species of the cultivars that are tolerant. The information assimilated over EPR constructed studies in seeds or food processing as revealed previously, sustenance the method to inspect the impacts of physical invigoration methods for seed treatments. Marcu et al. (2013) used EPR to study the biochemical changes induced in maize seeds when irradiated with cumulative doses of γ-radiation.
5 Conclusion
Seeds with extraordinary potency are substitution of crop formation and regular output. By using physical methods, seed vigor and germination are enhanced because physical method is eco-friendly and provides benefit on large scale. The presence of suitable services to achieve the physical treatment on the preemergence metabolism is obstructing its efficiency on priming treatment applications. It is also exhibited during evaluation that radiation treatment has a very little effect on biochemical changes or gene expression. Hence, the deficiency of biomarkers related to appropriate time or radiation dose restricts the application of seed procedures at large scale. Nonetheless, scientists from industry and academia are presently attentive on incapacitating these problems. Recent improvements on high-output methods (e.g., Omics) as compared with latest discharge of genomic properties on crops are predictable to lead. Additional studies are looked for to identify the molecular players generated during the riposte of seed invigoration treatments, specifically in radio-tolerant cultivars or species. In comparison, by intensifying the total cultivars analyzed with each changed method, it will be conceivable to recognize best appropriate goals for a physical treatment that is specific in stopping harmful conditions. The environmental situations in moderating the reaction of seed to the given radioactivity behaviors could not be deserted, establishing itself as an auspicious research opportunity. This information is vital to improve new plans to enterprise new treatments to modify and progress seed invigoration and germination. A combined method is required to speed up translational and basic research related to seed expertise, thus, at the end, fabricating strategies for the seed operatives.
References
Abdel-Hady MS, Okasha EM, Soliman SSA, Talaat M (2008) Effect of gamma radiation and gibberellic acid on germination and alkaloid production in Atropa belladonna L. Aust J Basic Appl Sci 2:401–405
Afzal I, Mukhtar K, Qasim M, Basra SMA, Shahid M, Haq Z (2012) Magnetic stimulation of marigold seed. Int Agrophys 26:335–339. https://doi.org/10.2478/v10247-012-0047-1
Aguilar CH, Dominguez-Pacheco A, Carballo AC, Cruz-Orea A, Ivanov R, Bonilla JLL, Montañez JPV (2009) Alternating magnetic field irradiation effects. Acta Agrophysica 14:7–17
Ahmad M, Galland P, Ritz T, Wiltschko R, Wiltschko W (2007) Magnetic intensity affects cryptochrome-dependent responses in Arabidopsis thaliana. Planta 225:615–624. https://doi.org/10.1007/s00425-006-0383
Al-Enezi N, Al-Bahrany A, Al-Khayri J (2012) Effect of X-irradiation on date palm seed germination and seedling growth. Emir J Food Agric 24:415–424
Aladjadjiyan A (2010) Effect of microwave irradiation on seeds of lentils (Lens Culinaris, Med.). Rom J Biophys 20:213–221
Aladjadjiyan A (2012) Physical factors for plant growth stimulation improve food quality. In: Aladjadjiyan A (ed) Food production – approaches, challenges and tasks. InTech, Rijeka, pp 145–168
Anand A, Nagarajan S, Verma APS, Joshi DK, Pathak PC, Bhardwaj J (2012) Pre-treatment of seeds with static magnetic field ameliorates soil water stress in seedlings of maize (Zea mays L.). Indian J Biochem Biophys 49:63–70
Arena C, De Micco V, Macaeva E, Quintens R (2014) Space radiation effects on plant and mammalian cells. Acta Astronaut 104:419–431. https://doi.org/10.1016/j.actaastro.2014.05.005
Baby SM, Narayanaswamy GK, Anand A (2011) Superoxide radical production and performance index of photosystem II in leaves from magnetoprimed soybean seeds. Plant Signal Behav 6:1635–1637. https://doi.org/10.4161/psb.6.11.17720
Balakhnina T, Bulak P, Nosalewicz M, Pietruszewski S, Włodarczyk T (2015) The influence of wheat Triticum aestivum L. seed pre-sowing treatment with magnetic fields on germination, seedling growth, and antioxidant potential under optimal soil watering and flooding. Acta Physiol Plant 37:59. https://doi.org/10.1007/s11738-015-1802-2
Balestrazzi A, Confalonieri M, Macovei A, Carbonera D (2011) Seed imbibition in Medicago truncatula Gaertn: expression profiles of DNA repair genes in relation to PEG-mediated stress. J Plant Physiol 168:706–713. https://doi.org/10.1016/j.jplph.2010.10.008
Banik S, Bandyopadhyay S, Ganguly S (2003) Bioeffects of microwave – a brief review. Bioresour Technol 87:155–159. https://doi.org/10.1016/S0960-8524(02)00169-4
Beard BH, Haskins FA, Gardner CO (1958) Comparison of effects of X-rays and thermal neutrons on dormant seeds of barley, maize, mustard, and safflower. Genetics 43:728–736
Belyavskaya NA (2004) Biological effects due to weak magnetic field on plants. Adv Space Res 34:1566–1574. https://doi.org/10.1016/j.asr.2004.01.021
Belz RG, Piepho H-P (2012) Modeling effective dosages in hormetic dose-response studies. PLoS One 7:e33432. https://doi.org/10.1371/journal.pone.0033432
Bhardwaj J, Anand A, Nagarajan S (2012) Biochemical and biophysical changes associated with magnetopriming in germinating cucumber seeds. Plant Physiol Biochem 57:67–73. https://doi.org/10.1016/j.plaphy.2012.05.008
Bilalis DJ, Katsenios N, Efthimiadou A, Karkanis A, Efthimiadis P (2012) Investigation of pulsed electromagnetic field as a novel organic pre-sowing method on germination and initial growth stages of cotton. Electromagn Biol Med 31:143–150. https://doi.org/10.3109/15368378.2011.624660
Borzouei A, Naseriyan B, Majdabadi A, Kafi M, Khazaei H (2010) Effects of gamma radiation on germination and physiological aspects of wheat (Triticum aestivum L.) seedlings. Pak J Bot 42:2281–2290
Brown JE, Lu TY, Stevens C, Khan VA, Lu JY, Wilson CL, Collins DJ, Wilson MA, Lgwegbe ECK, Chalutz E, Droby S (2001) The effect of low dose ultraviolet light-C seed treatment on induced resistance in cabbage to black rot (Xanthomonas campestris pv. campestris). Crop Prot 20:873–883. https://doi.org/10.1016/S0261-2194(01)00037-0
Buitink J, Claessens MAE, Hemminga MA, Hoekstra FA (1998) Influence of water content and temperature on molecular mobility and intracellular glasses in seeds and pollen. Plant Phys 118:531–554. https://doi.org/10.1104/pp.118.2.531
Buitink J, Hemminga MA, Hoekstra FA (1999) Characterization of molecular mobility in seed tissues: an electron paramagnetic resonance spin probe study. Biophys J 76:3315–3322. https://doi.org/10.1016/S0006-3495(99)77484-9
Buitink J, Leprince O, Hemminga MA, Hoekstra FA (2000) Molecular mobility in the cytoplasm: an approach to describe and predict lifespan of dry germplasm. Proc Natl Acad Sci U S A 97:2385–2390. https://doi.org/10.1073/pnas.040554797
Calabrese EJ, Baldwin LA (2000) Radiation hormesis: its historical foundations as a biological hypothesis. Hum Exp Toxicol 19:41–75. https://doi.org/10.1191/096032700678815585
Celestino C, Picazo ML, Toribio M (2000) Influence of chronic exposure to an electromagnetic field on germination and early growth of Quercus suber seeds: preliminary study. Electromagn Biol Med 19:115–120
Confalonieri M, Faè M, Balestrazzi A, Donà M, Macovei A, Valassi A, Giraffa G, Carbonera D (2014) Enhanced osmotic stress tolerance in Medicago truncatula plants overexpressing the DNA repair gene MtTdp2α (tyrosyl-DNA phosphodiesterase 2). Plant Cell Tissue Organ Cult 116:187–203. https://doi.org/10.1007/s11240-013-0395-y
Choudhary KK, Agrawal SB (2014) Ultraviolet-B induced changes in morphological, physiological and biochemical parameters of two cultivars of pea (Pisum sativum L.). Ecotoxicol Environ Saf 100:178–187. https://doi.org/10.1016/j.ecoenv.2013.10.032
De Micco V, Paradiso R, Aronne G, De Pascale S, Quarto M, Arena C (2014) Leaf anatomy and photochemical behaviour of Solanum lycopersicum L. plants from seeds irradiated with low-LET ionising radiation. Sci World J 2014:428141. https://doi.org/10.1155/2014/428141
De Souza A, Garcí D, Sueiro L, Gilart F, Porras E, Licea L (2006) Pre-sowing magnetic treatments of tomato seeds increase the growth and yield of plants. Bioelectromagnetics 27:247–257. https://doi.org/10.1002/bem.20206
Dubey AK, Yadav JR, Singh B (2007) Studies on induced mutations by gamma irradiation in okra (Abelmoschus esculentus (L.) Monch.). Progress Agric 7:46–48
Edmondson JL, Davies ZG, Gaston KJ, Leake JR (2014) Urban cultivation in allotments maintains soil qualities adversely affected by conventional agriculture. J Appl Ecol 51:880–889. https://doi.org/10.1111/1365-2664.12254
Efthimiadou A, Katsenios N, Karkanis A, Papastylianou P, Triantafyllidis V, Travlos I, Bilalis DJ (2014) Effects of presowing pulsed electromagnetic treatment of tomato seed on growth, yield, and lycopene content. Sci World J 2014:369745. https://doi.org/10.1155/2014/369745
Einset J, Collins AR (2015) DNA repair after X-irradiation: lessons from plants. Mutagenesis 30:45–50. https://doi.org/10.1093/mutage/geu054
Esnault M-A, Legue F, Chenal C (2010) Ionizing radiation: advances in plant response. Environ Exp Bot 68:231–237. https://doi.org/10.1016/j.envexpbot.2010.01.007
Farkas J, Mohácsi-Farkas C (2011) History and future of food irradiation. Trends Food Sci Technol 22:121–126. https://doi.org/10.1016/j.tifs.2010.04.002
Farokh P, Mahmoodzadeh H, Satari T (2010) Response of seed germination of safflower to UV-B radiation. Res J Environ Sci 4:70–74. https://doi.org/10.3923/rjes.2010.70.74
Ghodbane S, Lahbib A, Sakly M, Abdelmelek H (2013) Bioeffects of static magnetic fields: oxidative stress, genotoxic effects, and cancer studies. Biomed Res Int 2013:602987. https://doi.org/10.1155/2013/602987
Gicquel M, Taconnat L, Renou JP, Esnault MA, Cabello-Hurtado F (2012) Kinetic transcriptomic approach revealed metabolic pathways and genotoxic-related changes implied in the Arabidopsis response to ionising radiations. Plant Sci 195:106–119. https://doi.org/10.1016/j.plantsci.2012.06.015
Golovina EA, Tikhonov AN, Hoekstra FA (1997) An electron paramagnetic resonance spin-probe study of membrane permeability changes with seed ageing. Plant Phys 114:383–389
Guajardo-Flores D, Serna-Guerrero D, Serna-Saldívar SO, Jacobo-Velázquez DA (2014) Effect of germination and UV-C radiation on the accumulation of flavonoids and saponins in black bean seed coats. Cereal Chem 91:276–279. https://doi.org/10.1094/CCHEM-08-13-0172-R
Hamid N, Jawaid F (2011) Influence of seed pre-treatment by UV-A and UV-C radiation on germination and growth of mung beans. Pakistan J Chem 1:164–167. https://doi.org/10.15228/2011.v01.i04.p04
Hegazi AZ, Hamideldin N (2010) The effect of gamma irradiation on enhancement of growth and seed yield of okra [Abelmoschus esculentus (L.) Monech] and associated molecular changes. J Hortic For 2:38–51
Heisler GM, Grant RH, Gao W, Slusser JR (2003) Ultraviolet radiation and its impacts on agriculture and forests. Agric For Meteorol 120:3–7. https://doi.org/10.1016/j.agrformet.2003.08.007
Hideg E, Jansen MAK, Strid A (2013) UV-B exposure, ROS, and stress: inseparable companions or loosely linked associates? Trends Plant Sci 18:107–115. https://doi.org/10.1016/j.tplants.2012.09.003
Hollósy F (2002) Effects of ultraviolet radiation on plant cells. Micron 33:179–197. https://doi.org/10.1016/S0968-4328(01)00011-7
Hussain S, Zheng M, Khan F, Khaliq A, Fahad S, Peng S, Huang J, Cui K, Nie L (2015) Benefits of rice seed priming are offset permanently by prolonged storage and the storage conditions. Sci Rep 5:8101. https://doi.org/10.1038/srep08101
Irfaq M, Nawab K (2001) Effect of gamma irradiation on some morphological characteristics of three wheat (Triticum aestivum L.) cultivars. J Biol Sci 1:935–937. https://doi.org/10.3923/jbs.2001.935.937
Iuliana C, Caprita R, Giancarla V, Sorina R (2013) Response of barley seedlings to microwaves at 2.45 GHz. Sci Pap Anim Sci Biotechnol 46:185–191
Jakubowski T (2010) The impact of microwave radiations at different frequencies on the weight of seed potato germs and crop of potato tubers. Agric Eng 6:57–64
Javed N, Ashraf M, Akram NA, Al-Qurainy F (2011) Alleviation of adverse effects of drought stress on growth and some potential physiological attributes in maize (Zea mays L.) by seed electromagnetic treatment. Photochem Photobiol 87:1354–1362. https://doi.org/10.1111/j.1751-1097.2011.00990.x
Jayasanka SMDH, Asaeda T (2014) The significance of microwaves in the environment and its effect on plants. Environ Rev 22:220–228. https://doi.org/10.1016/j.biortech.2015.02.055
Jayawardena SDL, Peiris R (1988) Food crops breeding in Sri Lanka - achievements and challenges. BIO News 4:22–34
Knox OGG, McHugh MJ, Fountaine JM, Havis ND (2013) Effects of microwaves on fungal pathogens of wheat seed. Crop Prot 50:12–16. https://doi.org/10.1016/j.cropro.2013.03.009
Kotwaliwale N, Singh K, Kalne A, Jha SN, Seth N, Kar A (2014) X-ray imaging methods for internal quality evaluation of agricultural produce. J Food Sci Technol 51:1–15. https://doi.org/10.1007/s13197-011-0485-y
Kovács E, Keresztes Á (2002) Effect of gamma and UV-B/C radiation on plant cells. Micron 33:199–210. https://doi.org/10.1016/S0968-4328(01)00012-9
Kovalchuk I, Molinier J, Yao Y, Arkhipov A, Kovalchuk O (2007) Transcriptome analysis reveals fundamental differences in plant response to acute and chronic exposure to ionizing radiation. Mutat Res 624:101–113. https://doi.org/10.1016/j.mrfmmm.2007.04.009
Krylov A, Tarakanova GA (1960) Magnetotropism of plants and its nature. Plant Phys 7:156–160
Kurdziel M, Dlubacz A, Weselucha-Birczynska A, Filek M, Labanowska M (2015) Stable radicals and biochemical compounds in embryos and endosperm of wheat grains differentiating sensitive and tolerant genotypes-EPR and Raman studies. J Plant Phys 183:95–107. https://doi.org/10.1016/j.jplph.2015.05.018
Labanowska M, Kurdziel M, Filek M (2016) Changes of paramagnetic species in cereal grains upon short-term ozone action as a marker of oxidative stress tolerance. J Plant Physiol 190:54–66. https://doi.org/10.1016/j.jplph.2015.10.011
Leprince O, Hoekstra FA (1998) The responses of cytochrome redox state and energy metabolism to dehydration support a role for cytoplasmic viscosity in desiccation tolerance. Plant Physiol 1181253–1181264. https://doi.org/10.1104/pp.118.4.1253
Leprince O, Vertucci CW, Hendry GAF, Atherton NM (1995) The expression of desiccation-induced damage in orthodox seeds is a function of oxygen and temperature. Physiol Plant 94:233–240. https://doi.org/10.1034/j.1399-3054.1995.940208.x
Luckey TD (1980) Hormesis with ionizing radiation. CRC Press, Boca Raton
Luckey TD (2006) Radiation hormesis: the good, the bad, and the ugly. Dose Response 4:169–190. https://doi.org/10.2203/dose-response.06-102.Luckey
Macovei A, Garg B, Raikwar S, Balestrazzi A, Carbonera D, Buttafava A (2014) Synergistic exposure of rice seeds to different doses of γ-ray and salinity stress resulted in increased antioxidant enzyme activities and gene-specific modulation of TC-NER pathway. Biomed Res Int 170:780–787. https://doi.org/10.1155/2014/676934
Maherchandani N (1975) Effects of gamma radiation on the dormant seed of Avena fatua L. Radiat Bot 15:439–443. https://doi.org/10.1016/0033-7560(75)90018-6
Maity JP, Mishra D, Chakraborty A, Saha A, Santra SC, Chanda S (2005) Modulation of some quantitative and qualitative characteristics in rice (Oryza sativa L.) and mung (Phaseolus mungo L.) by ionizing radiation. Radiat Phys Chem 74:391–394. https://doi.org/10.1016/j.radphyschem.2004.08.005
Majeed A, Ahmad H, Muhammad Z (2010) Variation in chlorophyll contents and grain yield of Lepidium sativum L as induced by gamma irradiation. Int J Biol Sci Eng 1:147–151
Marcu D, Damian G, Cosma C, Cristea V (2013) Gamma radiation effects on seed germination, growth and pigment content, and ESR study of induced free radicals in maize (Zea mays). J Biol Phys 39:625–634. https://doi.org/10.1007/s10867-013-9322-z
Mattson MP (2008) Hormesis defined. Ageing Res Rev 7:1–7. https://doi.org/10.1016/j.arr.2007.08.007
Minisi FA, El-Mahrouk ME, Rida ME-DF, Nasr MN (2013) Effects of gamma radiation on germination, growth characteristics and morphological variations of Moluccella laevis L. Am Eurasian J Agric Environ Sci 13:696–704
Mitchell LM, Cambrosio A (1997) The invisible topography of power: electromagnetic fields, bodies and the environment. Soc Stud Sci 27:221–271. https://doi.org/10.1177/030631297027002002
Mokobia CE, Anomohanran O (2005) The effect of gamma irradiation on the germination and growth of certain Nigerian agricultural crops. J Radiol Prot 25:181–188. https://doi.org/10.1088/0952-4746/25/2/006
Moussa HR (2006) Role of gamma irradiation in regulation of NO3 level in rocket (Eruca vesicaria subsp. sativa) plants. Russ J Plant Physiol 53:193–197. https://doi.org/10.1134/S1021443706020075
Ouhibi C, Attia H, Rebah F, Msilini N, Chebbi M, Aarrouf J, Urban L, Lachaal M (2014) Salt stress mitigation by seed priming with UV-C in lettuce plants, growth, antioxidant activity and phenolic compounds. Plant Phys Biochem 83:126–133. https://doi.org/10.1016/j.plaphy.2014.07.019
Paparella S, Araújo SS, Rossi G, Wijayasinghe M, Carbonera D, Balestrazzi A (2015) Seed priming: state of the art and new perspectives. Plant Cell Rep 34:1281–1293. https://doi.org/10.1007/s00299-015-1784-y
Poinapen D, Brown DCW, Beeharry GK (2013) Seed orientation and magnetic field strength have more influence on tomato seed performance than relative humidity and duration of exposure to non-uniform static magnetic fields. J Plant Physiol 170:1251–1258. https://doi.org/10.1016/j.jplph.2013.04.016
Qi W, Zhang L, Wang L, Xu H, Jin Q, Jiao Z (2015) Pretreatment with low-dose gamma irradiation enhances tolerance to the stress of cadmium and lead in Arabidopsis thaliana seedlings. Ecotoxicol Environ Saf 115:243–249. https://doi.org/10.1016/j.ecoenv.2015.02.026
Rajendra P, Nayak HS, Rao BB, Subramanyam C, Devendranath D, Gunasekaran B, Aradhya SRS, Bhaskaran A (2005) Effects of power frequency electromagnetic fields on growth of germinating Vicia faba L., the broad bean. Electromagn Biol Med 24:39–54. https://doi.org/10.1081/JBC-200055058
Reddy MVB, Raghavan GSV, Kushalappa AC, Paulitz TC (1998) Effect of microwave treatment on quality of wheat seeds infected with Fusarium graminearum. J Agric Eng Res 71:113–117. https://doi.org/10.1006/jaer.1998.0305
Reddy KV, Reshma SR, Jareena S, Nagaraju M (2012) Exposure of greengram seeds (Vigna radiate var. radiata) to static magnetic fields: effects on germination and alfa-amylase activity. Res J Seed Sci 5:106–114. https://doi.org/10.3923/rjss.2012.106.114
Sacandè M, Golovina EA, Van Aelst AC, Hoekstra FA (2001) Viability loss of neem (Azadirachta indica) seeds associated with membrane phase behaviour. J Exp Bot 52:919–931. https://doi.org/10.1093/jexbot/52.358.919
Sahin H (2014) Effects of microwaves on the germination of weed seeds. J Biosyst Eng 39:304–309. https://doi.org/10.5307/JBE.2014.39.4.304
Sahu ID, McCarrik RM, Lorigan GA (2013) Use of electron paramagnetic resonance to solve biochemical problems. Biochemistry 52:5967–5984. https://doi.org/10.1021/bi400834a
Scialabba A, Tamburello C (2002) Microwave effects on germination and growth of radish (Raphanus sativus L.) seedlings. Acta Bot Gall 149:113–123. https://doi.org/10.1080/12538078.2002.10515947
Shaukat S, Farooq M, Siddiqui M, Zaidi S (2013) Effect of enhanced UV-B radiation on germination, seedling growth and biochemical responses of Vigna mungo (L.) Hepper. Pak J Bot 45:779–785
Shine MB, Guruprasad KN, Anand A (2011) Enhancement of germination, growth, and photosynthesis in soybean by pre-treatment of seeds with magnetic field. Bioelectromagnetics 32:474–484. https://doi.org/10.1002/bem.20656
Siddiqui A, Dawar S, Javed Zaki M, Hamid N (2011) Role of ultra violet (UV-C) radiation in the control of root infecting fungi on groundnut and mung bean. Pak J Bot 43:2221–2224
Sjodin J (1962) Some observations in X1 and X2 of Vicia faba L., after treatment with different mutagens. Hereditas 48:565–586
Smirnov AI, Golovina HA, Yakimchenko OE, Aksyonov SJ, Lebedev YS (1992) In vivo seed investigation by electron paramagnetic resonance spin probe technique. J Plant Phys 140:447–452. https://doi.org/10.1016/S0176-1617(11)80823-0
Soran M-L, Stan M, Niinemets Ü, Copolovici L (2014) Influence of microwave frequency electromagnetic radiation on terpene emission and content in aromatic plants. J Plant Physiol 171:1436–1443. https://doi.org/10.1016/j.jplph.2014.06.013
Steffen-Heins A, Steffens B (2015) EPR spectroscopy and its use in planta-a promising technique to disentangle the origin of specific ROS. Front Environ Sci 3:15. https://doi.org/10.3389/fenvs.2015.00015
Talei D, Valdiani A, Maziah M, Mohsenkhah M (2013) Germination response of MR 219 rice variety to different exposure times and periods of 2450 MHz microwave frequency. Sci World J 2013:408026. https://doi.org/10.1155/2013/408026
Teixeira da Silva JA, Dobránszki J (2015) Magnetic fields: how is plant growth and development impacted? Protoplasma 253:231–248. https://doi.org/10.1007/s00709-015-0820-7
Vashisth A, Nagarajan S (2010) Effect on germination and early growth characteristics in sunflower (Helianthus annuus) seeds exposed to static magnetic field. J Plant Physiol 167:149–156. https://doi.org/10.1016/j.jplph.2009.08.011
Ventura L, Donà M, Macovei A, Carbonera D, Buttafava A, Mondoni A, Rossi G, Balestrazzi A (2012) Understanding the molecular pathways associated with seed vigor. Plant Physiol Biochem 60:196–206. https://doi.org/10.1016/j.plaphy.2012.07.031
Vian A, Roux D, Girard S, Bonnet P, Paladian F, Davies E, Ledoigt G (2006) Microwave irradiation affects gene expression in plants. Plant Signal Behav 1:67–70. https://doi.org/10.4161/psb.1.2.2434
Weil JA, Bolton JR (2006) Electron paramagnetic resonance: elementary, theory and practical applications. John Wiley and Sons, Inc, Hoboken
Wolff SA, Coelho LH, Karoliussen I, Jost A-IK (2014) Effects of the extraterrestrial environment on plants: recommendations for future space experiments for the MELiSSA higher plant compartment. Life 4:189–204. https://doi.org/10.3390/life4020189
Zaka R, Chenal C, Misset M (2002) Study of external low irradiation dose effects on induction of chromosome aberrations in Pisum sativum root tip meristem. Mutat Res 517:87–99. https://doi.org/10.1016/S1383-5718(02)00056-6
Zeeman P (1897) The effect of magnetisation on the nature of light emitted by a substance. Nature 55:347. https://doi.org/10.1038/055347a0
Author information
Authors and Affiliations
Corresponding authors
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Rehman, A., Mazhar, K., Ijaz, M., Ali, Q., Ahmad, S. (2019). Seedling Pretreatment: Methods and Protocols. In: Hasanuzzaman, M., Fotopoulos, V. (eds) Priming and Pretreatment of Seeds and Seedlings. Springer, Singapore. https://doi.org/10.1007/978-981-13-8625-1_5
Download citation
DOI: https://doi.org/10.1007/978-981-13-8625-1_5
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-13-8624-4
Online ISBN: 978-981-13-8625-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)