
Abstract
Fungi have been recommended as agents for the biological control of insects for over a century, but their use remains particularly limited. As with numerous microorganisms, an entomopathogenic fungus from genera Metarhizium and Verticillium produces several metabolites. While these compounds are a conspicuous feature of the biology of the producing fungi, their roles in pathogenicity and other relations with their hosts and challenging microbes are not well understood. Insect pathogenic fungi are effective in controlling the occurrence of certain populations of soil pests with varied type of action and virulence. The fungi have evolved mechanisms for adhesion and recognition of host surface cues that help in direct adaptive response including the production of hydrolytic, assimilatory and detoxifying enzymes and additional metabolites that facilitate infection in insect pests. However, entomopathogenic fungi are a chief component of integrated pest management and form an integrated part of mycoinsecticide in agriculture. Entomopathogenic fungi are considered as an effective biocontrol agent against various plant pests. Therefore, in this book chapter, we discussed the role of entomopathogenic fungi Metarhizium and Verticillium for the management of the insect pests in agricultural ecosystem.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
The practice of cultivation soil and growing of the crops has been one of the major reasons for the adoption of the civilized lifestyle of humans. The practice of agriculture has always been a source of food production as well as a livelihood means. It is estimated that almost one-third of the total crop yield is lost due to the infestation of crop pests, the infection of the pathogens and the competition from the weeds. With the advent of the synthetic pesticides, the loss of the total crop yield was reduced, and the agricultural productivity was increased as it provided protection to the crops against the pests and the diseases. Although the haphazardous use of these synthetic pesticides has led to serious problems such their persistance in environment, residual effects in the food products and development of resistance in pests (Shelton et al. 2002). Over the past few years, there has been an increased concern in the people about the potential adverse effects which are associated with the imperceptive use of the synthetic pesticides, which has, in turn, led to the urge for development of an alternative method for the control of the crop pests (Keswani et al. 2016; Mishra et al. 2015). In this context, microbial secondary metabolites of the reported entomopathogenic fungi are deemed to be employed as one of the finest alternatives.
In general, all microorganisms produce a variety of compound which are structurally related but are found in the different magnitude relatively and are classified as the primary or the secondary metabolites (Singh et al. 2016, 2017). The primary metabolites are the microbial products which are made during the log or exponential phase of the growth and whose synthesis is an integral part of the normal growth process on a microbe. These include the end products and the intermediaries of the anabolic metabolism, which are used by the cell as the building blocks for essential macromolecules (such as amino acids, nucleotides) or are converted to coenzymes (such as vitamins). They also include the resultant products from the catabolic metabolism, which leads to the production of energy and utilization of the substrate and thus ultimately to the growth (Sanchez and Demain 2008). The secondary metabolites, on the other hand, are products of the secondary metabolism which are diverse in nature and don’t have a role in the basic life processes. They are not involved in either cell metabolism or in the growth of the microorganism. They are produced at the stationary phase of the microbial growth stage and are the source of the therapeutics, insecticides, drugs, flavours and fragrances (Donadio et al. 2002). The concept of the secondary metabolism was first introduced by Kossel in 1891 (Hartmann 1985; Haslam 1986; Seigler 1998; Turner 1971). The application of the secondary metabolites as the insecticide against the crop pests has emerged to be advantageous to other alternatives as they are biodegradable, non-toxic to nontarget organisms and highly selective and have low resistance development in the target pest population (Deepa et al. 2014; Keswani 2015a, b).
A key pest can be described on the basis of economic injury level (EIL), general equilibrium population (GEP) and damage boundary (DB). Pests whose GEP always lies above EIL are persistent, severly damaging and the spray of the insecticides brings their population below EIL. The estimated annual crop loss in India by insect pests is Rs. 29,240 crores (Dhaliwal and Arora 1996).
Most of the studies on the entomophagous fungi are on Metarhizium anisopliae and Beauveria bassiana and less on equally important species of commercial importance such as Verticillium lecanii, Paecilomyces fumosoroseus, Tolypocladium spp. and Hirsutella spp. These fungi produce an array of secondary metabolites, of which some are restricted to specific genera, while others are more ubiquitous (Keswani et al. 2013). These secondary metabolites originate as a derivative from various intermediaries in primary metabolism. In general, most of the secondary metabolites emerge from the five metabolic sources, viz.:
-
(i)
Amino acids
-
(ii)
The shikimic acid pathway for the biosynthesis of aromatic amino acids
-
(iii)
The polyketide biosynthesis pathway from acetyl coenzyme A (CoA)
-
(iv)
The mevalonic acid pathway from acetyl coenzyme A
-
(v)
Polysaccharides and peptido-polysaccharides (Griffin 1994)
This chapter reviews about the different secondary metabolites secreted by the fungi Metarhizium spp. and Verticillium spp. that act against the crop’s insect pests.
2 The Hypothesis Suggesting the Role of Secondary Metabolites
There are mainly three hypotheses which suggest the role of the secondary metabolites in organisms which are as follows:
2.1 The “Waste Product” Hypothesis
The role of secondary metabolites has been rather uncertain and was initially thought to be just the waste materials. The relatively large number and amount of secondary metabolites which are observed in nature and the concept that these compounds have arisen from the “errors” in the primary metabolisms in plants led to the idea that the secondary metabolic compounds originate and accumulate as “waste products”. Although taking into consideration their nonmotility and lack of sophisticated immune system, plants have to develop their own defence system against the pathogens and predators along with the systems to lure the motile organisms, for fertilization and dissemination (Luckner 1972, 1990; Mothes 1976; Seigler 1998).
2.2 The Shunt or Overflow Hypothesis
For some, secondary metabolites are envisaged as the shunt metabolic compounds which are produced in a state of unbalanced growth for reducing the abnormal concentration of the normal cellular constituents. The synthesis of enzymes designed to carry out the secondary metabolism allows the primary metabolic enzymes to continue to function until such time as circumstances are propitious for renewed metabolic activity and growth. However, this could be linked to the depletion of nutrients such as phosphorous or nitrogen (Bu’Lock 1980; Haslam 1986).
2.3 The Increased Fit Hypothesis
The hypothesis takes into account that many natural products trigger very specific physiological responses in other organisms and in many cases bind to the receptors which have a remarkable complementarity, which means that the natural products may aid in an organism’s survival in the absence of an immune system. This fact, in turn, supports the hypothesis that the secondary metabolites increase the fitness of those individuals which possess them and they are favoured in the process of natural selection. The secondary metabolites thus have an important ecological role in the interaction with the environment and act like the communication interface between the plants and its friends and foes in the environment (Harborne 1986; Rosenthal and Janzen, 1979; Swain 1977; Torssell 1997).
Many of those secondary metabolites are fungicidal, bactericidal, repellent or poisonous to insect pests and the herbivores. The flower pigments give attracting colours for insects that help in fertilization or warning colours against the predators. Some of the secondary compounds also perform in signalling pathway as plant hormones (Haslam 1985). In addition to these, many of them are initially meant for defence against herbivores such as insect pests which would soon come up with the metabolic pathways to detoxify and even use these defence compounds.
3 The Secondary Metabolites of Metarhizium Species
3.1 Destruxins
They are a class of cyclic hexapeptides that were originally isolated from the entomophagous fungus, Metarhizium anisoplae (Kodaira 1961a, b, 1962; Roberts 1966, 1969). The discussions of the secondary metabolites of Metarhizium start and stop with the destruxins. After their first report as insect toxins (Kodaira 1961a, b), several papers and reviews describing their chemistry and biological activities have been published (Hu and Dong 2015; Liu and Tzeng 2012; Pedras et al. 2002).
They are composed of five amino acid residues and a single α-hydroxycarboxylic acid moiety (Suzuki et al. 1970; Suzuki and Tamura 1972; Pais et al. 1981), whose exact nature differentiates the major destruxins into subclasses A to F. The two of the five amino acids are N-methylated amino acids: N-methyl-L-alanine (replaced by L-alanine in protodestruxin) and N-methyl-L-valine (replaced by L-valine in desmethyldestruxin B and protodestruxin). The remaining three amino acids are β-alanine, L-leucine (e.g. in destruxin A and A1 but is replaced by L-valine in destruxin A2) and L-proline (e.g. in destruxin A and A2 but is replaced by L-pipecolic acid in destruxin A1).
The variable structural residue of destruxins is the α-hydroxycarboxylic acid unit. For example, in destruxin A it is 2-hydroxy-4-pentanoic acid; in destruxin B it is 2-hydroxy-4-methylpentanoic acid; in destruxin E it is 2-hydroxy-4,5-methylpentanoic acid; and in destruxin F it is 2,4-dihydroxy pentanoic acid (Wahlman and Davidson 1993). The destruxin analogues obtained from other fungi include destruxins A4 and A5 and homodestruxin B from an entomophagous fungus Aschersonia sp. (Krasnoff et al. 1996); roseocardin and roseotoxin B from a plant pathogenic fungus Trichothecium roseum (Springer et al. 1984; Tsunoo et al. 1997); bursaphelocides from a Mycelia sterilia (Kawazu et al. 1993); pseudodestruxins A and B from a coprophilous fungus Nigrosalbulum globosum (Che et al. 2001) and Beauveria felina (Lira et al. 2006); and isaridins A and B from an undescribed Isaria strain isolated from rat dung (Ravindra et al. 2004; Sabareesh et al. 2007).
Destruxin exhibits an array of amazing biological properties which include insecticidal activity, cytotoxic activity and moderate antibiotic (antituberculotic) activity (Pedras et al. 2002). Apart from these, destruxins have also been shown to possess immunodepressant activity in insect model systems (Vey et al. 1985; Huxham et al. 1989; Cerenius et al. 1990). They cause membrane depolarization by opening calcium channel leading to the tetanic paralysis in the insects (Samuels et al. 1988). Destruxin E seemed to be the most potent destruxin with the insecticidal activity having repellent and aphicidal properties (Robert and Riba 1989), contact insecticidal activity (Poprawski et al. 1994) and antifeedant properties (Amiri et al. 1999).
3.2 Serinocyclins
The serinocyclins were first identified from the conidia of fungus Metarhizium robertsii ARSEF 2575, and its structure was elucidated from the isolates obtained from Metarhizium acridum (Krasnoff et al. 2007). They are the cyclic heptapeptides which feature many non-proteinogenic amino acids and composed of 1-aminocyclopropane-1-carboxylic acid (ACC) which acylates 4-hydroxyproline followed by the amidation of 1-aminocyclopropane-1-carboxylic acid with L-serine, D-4-hydroxylysine, β-alanine, D-serine and L-serine to form a 22-membered macrocycle. Serinocyclin B has D-lysine in place of D-4-hydroxylysine.
Serinocyclin A showed entomophagous activity as the exposed mosquito larvae to this compound exhibited abnormal swimming as they were unable to control the position of their heads (Krasnoff et al. 2007). The compound is believed to have a neurophysiological effect on the hair tufts which are used as the rudders to adjust the head position while swimming (Brackenbury 1999, 2001). A virtual docking study in 2014 has suggested that the serinocyclin binds to glutathione S-transferase (Sanivada and Challa 2014).
3.3 Metachelins
It is a group of coprogen-type hydroxamate siderophores that were isolated first from Metarhizium robertsii ARSEF 2575 when it was grown in iron-exploited medium (Krasnoff et al. 2014). The isolated medium included N α-dimethyl coprogen and dimerumic acid which were known earlier to be obtained from Alternaria longipes, Fusarium dimerum (Jalal et al. 1988), Alternaria brassicicola (Oide et al. 2006), Verticillium dahliae (Harrington and Neilands, 1982) and Gliocladium virens (Jalal et al. 1986), respectively. Apart from these known compounds, four novel siderophores were also reported from M. robertsii.
Dimerumic acid is synthesized by the condensation of two molecules of 5-anhydromevalonyl-N-5-hydroxyornithine to form a diketopiperazine ring, and further N α-dimethyl coprogen is synthesized by the head-to-tail esterification of the third molecule of 5-anhydromevalonyl-N-5-hydroxyornithine to one of the terminal hydroxyl group. One of the four novel siderophores which is also the major component of the mixture, metachelin A, is derived from N-dimethyl coprogen molecule after the glycosylation of both terminal hydroxyl groups by D-mannose and N-oxidation of the dimethyl nitrogen.
Metachelin forms the hexadentate chelating complexes with Fe+3 and other trivalent metal cations like Al+3 and Ga+3. Metachelins and related compounds from Me. robertsii showed approximately equal activity to that of the bacterial siderophore, ferrioxamine, in a CAS plate assay (Krasnof et al. 2014).
3.4 Ferricrocin
It is an intracellular hexapeptide of the ferrichrome-type siderophore that was produced in its ferrated form by M. robertsii 2575 (Jalal et al. 1988). It was first reported in Aspergillus spp. (Zähner et al. 1963). It is presumed to receive environmental iron scavenged by extracellular siderophores and to transport it to its target sites in the cell (Wallner et al. 2009). Ferricrocin has the sequence Ser-Ser-Gly-Orn1-Orn2-Orn3-Orn4 where the Orn units are all Nδ-acetyl-Nδ-hydroxyornithines.
3.5 Tyrosine Betaine
It was isolated and characterized from Metarhizium anisopliae var. anisopliae strain ESALQ 1037 (Carollo et al. 2010). It is a dipeptide molecule having a molecule of betaine that is conjugated with tyrosine whose structure is identified as 2-{[1-carboxy-2-(4-hydroxyphenyl)ethyl] amino}-N,N,N-trimethyl-2-oxoethanammonium (Carollo et al. 2010). It was then also identified in an HPLC screening of the conidial extracts of Metarhizium acridum (ARSEF 324, ARSEF 3391 and ARSEF 7486) and Metarhizium brunneum (ARSEF 1095, ARSEF 5626 and ARSEF 5749) (Carollo et al. 2010). It is also observed to co-occur with serinocyclins and ferricrocin in extracts of conidia of Metarhizium guizhouense (ARSEF 683), Metarhizium pingshaense (ARSEF 2106 and ARSEF 5197) and M. robertsii (ARSEF 2575 and ARSEF 4123) during mass spectrometric analysis (Donzelli and Krasnoff 2016). The biological activity of this compound has not been reported yet.
3.6 Metacytofilin
It is an immunosuppressive compound that was obtained from Metarhizium spp. TA2759 which is a two-residue depsipeptide having the structure 3α-hydroxy-6β-methylamino-6α-(−methyl propyl)-3β-phenylmethyl-4H-2,3,5,6-tetrahydro-1,4-oxazine-2,5-dione (Iijima et al. 1992).
3.7 Fungerins
Isolated from Metarhizium spp. FKI-1079, fungerin, which was initially identified from a Fusarium spp. (Singh et al. 2001), along with two novel analogues, namely, hydroxyfungerin A and its regioisomer hydroxyfungerin B, has an imidazole core (Uchida et al. 2005). The potentiality of the new compound which is unique to the Metarhizium strain was 1/12 in acute toxicity assay against brine shrimp (Artemia) and was inactive at 10 μg/disk against Caenorhabditis elegans or against a panel of microbe which included nine bacteria and five fungi as compared to fungerin (Donzelli and Krasnoff 2016).
3.8 Aurovertins
In 2008, three new analogues of aurovertins (F–H) were isolated along with previously described aurovertin D from M. anisopliae HF260 (Azumi et al. 2008). Aurovertins were isolated first from Calcarisporium arbuscula with the structural elucidation first for aurovertin B (Mulheirn et al. 1974). They are known to inhibit the mitochondrial, bacterial and chloroplast ATPases (F1) and so are used for probing these critical enzymes (Donzelli and Krasnoff 2016).
3.9 Metacridamides
The two compounds, metacridamides A and B, were isolated from spores of Metarhizium acridum ARSEF 3341 composed of 19-membered macrocyclic lactones (Krasnoff et al. 2012). They neither showed the insecticidal activity nor the antimicrobial activity (Donzelli and Krasnoff 2016).
3.10 JBIRs
The compounds JBIR-19 and JBIR-20 were isolated from Metarhizium spp. fE61 having two 24-membered macrolides differing from each other by one hydroxyl substitution (Kozone et al. 2009). JBIR-19 showed weak antimicrobial activity against Saccharomyces cerevisiae at MICs of 200 μM, but JBIR-20 did not show any antimicrobial activity at this concentration, although both of them induced cell elongation of the same at the concentrations of 3.1 μM and 13 μM, respectively (Kozone et al. 2009).
3.11 Helvolic Acid
Helvolic acid was isolated from M. anisopliae, and its 6-deacetyl analogue, helvolinic acid, was isolated from M. anisopliae, Metarhizium brunneum and eight other fungi (Turner and Aldridge 1983). It was originally isolated as an antibacterial “fumigacin”, from Aspergillus fumigatus and Aspergillus clavatus, but was not structurally elucidated (Waksman et al. 1943). The full structure was finally solved in 1970 as a fusidane similar to fusidic acid which is built on the skeleton of cyclopentanoperhydrophenanthrene (Iwasaki et al. 1970; Okuda et al. 1964, 1967). Helvolic acid along with its 1,2-hydro analogue isolated from M. anisopliae strain HF293 was shown to have antibacterial activity against Staphylococcus aureus (Lee et al. 2008).
3.12 Metarhizins
Metarhizins A and B are the two functionalized diterpenes which are produced by Metarhizium flavoviride and are similar to viridoxins (Donzelli and Krasnoff 2016). Metarhizin A has (2R, 3S)-2-hydroxy-3-methylpentanoate at C3 as in viridoxin A, but metarhizin B has (R)-2-hydroxy-3-methylbutanoic acid (deaminated Val) (Kikuchi et al. 2009).
3.13 Ovalicins
The type of compound of this group, ovalicin, was isolated from Pseudeurotium ovalis (Sigg and Weber 1968). Its hydroxylated analogue, Mer-f3 or 12-hydroxy-ovalicin, was obtained from Metarhizium spp. f3 (Kuboki et al. 1999). The ovalicins are monocyclic sesquiterpenoids having highly oxygenated cyclohexane ring and two epoxide groups (Donzelli and Krasnoff 2016). 12-Hydroxy-ovalicin showed immunosuppressive activity in a mixed lymphocyte culture reaction assay and leukaemia cells of L-1210 mouse (Kuboki et al. 1999). It has also shown potent cytotoxicity against four human cancer cell lines and human umbilical vein endothelial cells (Donzelli and Krasnoff 2016).
3.14 Taxanes
The overwhelmingly effective chemotherapeutic to cancer, placitaxel, and the other related taxanes were isolated originally from various species of yew trees’ bark. Subsequently, placitaxel was reported from an endophyte, Taxomyces andreanae, living on Pacific yew (Taxus brevifolia) (Stierle et al. 1993). Among the more than 200 reported placitaxel-producing endophytic fungi, the highest yield is obtained from M. anisopliae (H-27 Accession FJ375161) (Donzelli and Krasnoff 2016). A controversy attached to the compound is that whether it is indeed a product of fungi at all (Heinig et al. 2013) and, if so, whether it is the result of a fungal version of the accepted plant pathway (Croteau et al. 2006).
3.15 Cytochalasins
These molecules are the subset of the “cytochalasans” which were thoroughly reviewed by Scherlach et al. (2010). The first cytochalasans which were described in 1966 are cytochalasins A and B that were obtained from Phoma strains S 298 (Rothweiler and Tamm 1966) and Helminthosporium dematioideum (Aldridge et al. 1967), respectively, and later cytochalasins C and D were isolated from the cultures of M. anisopliae (Roberts 1981). Cytochalasin D is also known to additionally occur in the fungi Zygosporangium mansonii and Helminthosporium species (Zimmermann 2007). In subsequent years many subclasses of compounds have been put together under the cytochalasins which include scoparisins, chaetoglobosins, penochalasins, aspochalsins, phomacins and alachalasins (Scherlach et al. 2010). In 2000, two new cytochalasin analogues were isolated from M. anisopliae in a screen for plant growth retardants, viz. diacetyl-cytochalasin C and an unnamed isomer (Fujii et al. 2000).
Cytochalasans constitute a perhydro-isoindolone molecule which is fused typically with a macrocyclic ring which may be a carbocycle, a lactone or a cyclic carbonate. The cytochalasins bear a benzyl group to the hydrogenated isoindolone moiety. The cytochalasins act as the inhibitors of the actin-cofilin interaction (Roberts, 1981; Strasser et al. 2000). When the plasmatocytes of greater wax moth (Galleria melanoleuca) were treated with the cytochalasins obtained from M. anisopliae, it was found that it caused the inhibition of attachment and also showed morphological alterations to the untreated ones (Vilcinskas et al. 1997a, b). This inhibition indicates the impairment in the plasmatocytes of the greater wax moth to perform the cell movements required for proper functioning of the cytoskeleton. Despite the basic biological activities of the cytochalasins, they are overshadowed by the destruxins in the collective effort of the secondary metabolites against insects.
3.16 Swainsonine
The compound was discovered after the observations of neurological symptoms and weight loss in livestock feeding on Swainsona spp. (Family Fabaceae), in Swainsona canescens, which inhibited lysosomal α-mannosidase (Dorling et al. 1978). The compound was named swainsonine, and its structure was elucidated as indolizidine-1,2,8-triol (Colegate et al. 1979). It was revealed to be isolated first from the fungus Rhizoctonia leguminicola and not from the plant after the complete structural elucidation of a compound that was previously obtained from the aforementioned fungus identical to swainsonine (Guengerich et al. 1973). Swainsonine was then subsequently isolated from M. anisopliae F-3622 (Hino et al. 1985). It is an indolizidine alkaloid moiety containing a fused piperidine and pyrrolidine ring system. They act as an aphid-feeding deterrent (Dreyer et al. 1985).
3.17 Viridoxins
Isolated from M. flavoviride (ARSEF 2133), viridoxins A and B are composed of a diterpenoid core with a 6-methoxy-2,3-dimethyl-γ-pyrone moiety that is attached to the 19th carbon and with (2R,3S)-2-hydroxy-3-methyl pentanoate and (R)-2-hydroxy-4-methyl pentanoate, respectively, at the third carbon (Gupta et al. 1993). They have shown insecticidal activity against the Colorado potato beetle (Leptinotarsa decemlineata) as leaf contamination (Gupta et al. 1993).
3.18 N-(Methyl-3-Oxodec-6-Enoyl)-2-Pyrroline and N-(Methyl-3-Oxodecanoyl)-2-Pyrroline
These are the two substituted pyrrolines that were reported from Metarhizium flavoviride HF698 as a weak plant pathogenic oomycete inhibitor (Putri et al. 2014). They were previously reported from Penicillium brevicompactum as the juvenile hormone inhibitors and also showed insecticidal activity against Oncopeltus fasciatus (Cantin Sanz et al. 1999; Moya et al. 1998) (Table 2.1).
4 The Secondary Metabolites of Verticillium Species
4.1 Bassianolide
It is a toxic metabolite which is obtained from Beauveria bassiana and Verticillium lecanii (Suzuki et al. 1977), and it was originally isolated from both fungi which were entomophagous on the cadavers of Bombyx mori pupae (Murakoshi et al. 1978). The bassianolide is an octadepsipeptide with a 24-membered macrolactone ring which is formed as the cyclic tetrameric ester of the dipeptidol monomer D-hydroxyisovaleric acid-N-methylleucine (Xu et al. 2008). The insecticidal activity of bassianolide was shown by Suzuki et al. in 1977, and it also inhibits acetylcholine-induced smooth muscle contraction (Nakajyo et al. 1983). They are proven to induce atony to the Helicoverpa (Heliothis) zea larvae (Champlin and Grula 1979).
4.2 Cyclosporines
They are also called as cyclosporines and were discovered in the 1970s obtained from Trichoderma polysporum and Cylindrocarpon lucidum (Borel et al. 1977; Dreyfuss et al. 1976). They are a series of cyclo-undecapeptide that were also reported to be produced by the Verticillium species by Jegorov and Weiser in 1990. They have insecticidal activities and were reported effective against larvae of mosquito (Matha et al. 1988; Podsiadlowski et al. 1998). Apart from that, cyclosporin A has the immunosuppressive effect on insect humoral immune response (Fiolka 2008) and cellular immune response (Vilcinskas et al. 1999).
4.3 Enniatins
They were first discovered in the 1940s (Gäumann et al. 1947). The analogues of enniatin are produced by various species of fungi including Verticillium (Herrmann et al. 1996; Supothina et al. 2004). Enniatin molecule is an N-methylated cyclohexadepsipeptides which comprise of three units each of N-methylated branched-chain L-amino acid and D-2-hydroxy acid that are arranged in an alternate fashion (Firakova et al. 2007). They are reported to inhibit ABC transporters (Hiraga et al. 2005), act as ionophores (Levy et al. 1995; Doebler 2000) and suppress acyl-CoA: cholesterol acyltransferase (Tomoda et al. 1992). They have the insecticidal properties (Monma et al. 2006) and are shown to act against the larvae of spruce budworm (Choristoneura fumiferana) (Strongman et al. 1988), Galleria mellonella (Mule et al. 1992) and adult of the blowfly (Calliphora erythrocephala) (Grove and Pople 1980).
4.4 Dipicolinic Acid
It is chemically known as pyridine-2,6-dicarboxylic acid. It is the metabolic product of several entomophagous fungi including Verticillium spp. (Shima, 1955). Dipicolinic acid was shown to have the insecticidal properties against blowfly (Calliphora erythrocephala) (Claydon and Grove 1982).
4.5 Verticilides
It was first isolated from the fungus Verticillium spp. FK-1033 (Omura et al. 2004). The compound is composed of a 24-membered ring cyclic depsipeptide containing a sequence of cyclo-[(2R)-2-hydroxyheptanonyl-N-methyl-L-alanyl] (Omura et al. 2004; Monma et al. 2006). The verticilides are shown to inhibit the ryanodine binding to the ryanodine receptors in cockroach and mouse (Monma et al. 2006; Shiomi et al. 2010).
4.6 Enalin
An analogue of enalin A, a coumaranone from the mangrove fungus Verrucukina enalia (Lin et al. 2002); 2,6-dihydroxy-2-methyl-7-(prop-1E-enyl)-1-benzofuran-3(2H)-one was obtained as one of the three compounds from Verticillium spp. isolated from the roots of wild Rehmannia glutinosa (You et al. 2009). Enalin A is widely distributed from microorganisms to higher plants and is known to have antimicrobial, antifungal, phytotoxic (Furumoto et al. 1997) and antidiabetic (Manickam et al. 1997) activities. The analogue of enalin A obtained from Verticillium spp. exhibited antibiotic activity against Septoria spp. and Fusarium spp. and also inhibited the growth of itself to some extent (You et al. 2009).
4.7 Massariphenone
It was originally reported from the marine-derived fungus Massarina spp. (Abdel-Wahab et al. 2007). Massariphenone was obtained as one of the three compounds from Verticillium spp. isolated from the roots of wild Rehmannia glutinosa (You et al. 2009). The chemical formula of the compound was as C10H12O3 by a high-resolution mass spectrometric data, and NMR spectrum of the compound showed signals of a 1,2,4-tri-substituted benzene ring, an aryl methyl group and an OCHCH3 unit (Abdel-Wahab et al. 2007). It has slight antibiotic activity as it inhibited the growth of Septoria spp. and Fusarium spp. only slightly (You et al. 2009).
4.8 Ergosterol Peroxide
It is reported from a wide range of fungal species and was first obtained from Cordyceps sinensis as an antitumor sterol (Bok et al. 1999). It is chemically 5α,8α-epidioxy-24(R)-methylcholesta-6,22-dien-3β-ol and was also obtained as one of the three compounds from Verticillium spp. isolated from the roots of wild Rehmannia glutinosa along with massariphenone and analogue of enalin A. It significantly inhibited biomass accumulation of Septoria spp., Fusarium spp. and Rhizoctonia spp. at a low concentration of 0.97 μg/ml in liquid culture (You et al. 2009).
4.9 Radicicol (Monorden)
Radicicol was isolated from Verticillium chlamydosporium (= Pochonia chlamydosporia) in search for the nematicidal mechanisms from nematophagous fungi (Khambay et al. 2000). It was originally found as an antifungal compound by Delmotte and Delmotte-Plaquee in 1953. Monorden E and an analogue of radicicol, monorden analogue-1, were purified from the fungus Pochonia chlamydosporia var. chlamydosporia strain TF-0480 (Shinonaga et al. 2009a, b). Radicicol and monorden E were originally obtained from a mycoparasite Humicola spp. FO-2942 that produced amidepsines, diacylglycerol acyltransferase inhibitors (Niu 2017). They have antifungal activity only against Aspergillus niger (Arai et al. 2003; Yamamoto et al. 2003). Radicicol does not have any nematicidal activity against root-knot nematode Meloidogyne incognita (Niu 2017), although it possess antiviral activity against herpes simplex virus (Hellwig et al. 2003).
4.10 Pochonins
Pochonins were all isolated first from the strains of V.chlamydosporium (= P. chlamydosporia) (Hellwig et al. 2003; Shinonaga et al. 2009a, b). Pochonins A–F were isolated from Pochonia chlamydosporia var. catenula strain P 0297 (Hellwig et al. 2003), and pochonins G–P along with pochonins B, D, E and F were isolated and characterized from P. chlamydosporia strain TF-0480 (Shinonaga et al. 2009a, b). Except pochonins F and J, all are chlorine-containing resorcylic acid lactones. Pochonins G and H are the first two compounds in the radicicol family to possess a furan ring, and pochonins L–N are the first three analogues of radicicol with an E-configuration of a double bond at C5–C6. Pochonin K is a 14-aldofuranose radicicol derivative, and pochonin I has a single benzene moiety in the macrolide ring (Niu 2017). Pochonins A–F except for pochonin D showed inhibitory action against herpes simplex virus 1 (Hellwig et al. 2003).
4.11 Monocillins
Monocillins I–IV along with radicicol were isolated originally from the fungus Monocillium nordinii, a mycoparasite of pine-pine gall rust Endocronartium harknessii (Ayer et al. 1980; Ayer and Peña-Rodriguez, 1987). Monocillins are the non-chlorine-containing resorcylic acid lactones. Monocillins II–III along with radicicol, pochonin F and a novel monocillin II glycoside were isolated from P. chlamydosporia var. catenulata strain P 0297 (Hellwig et al. 2003). All the four monocillins were later isolated from P. chlamydosporia strain TF-0480 (Shinonaga et al. 2009a, b). Monocillin III is a dechloro analogue of pochonin A showing potent inhibitory activity against herpes simplex virus 1, and monocillin II is same of pochonin D but with no inhibitory activity to the same virus (Hellwig et al. 2003). Monocillin I has antifungal activities against a wide variety of fungi including Phycomyces blaksleeanus, Pythium debaryanum, Ceratocystis ulmi as the cause of Dutch elm disease and Phellinus pini pointing towards the nonspecific nature. However, monocillins II–IV don’t show the same antifungal activities (Ayer et al. 1980; Aver and Peña-Rodriguez 1987).
4.12 Phomalactones
In 2000, the first study on isolation of phomalactone was reported from the fungus V. chlamydosporium (= P. chlamydosporia) in a bioassay against the root-knot nematode Meloidogyne incognita as a nematicidal metabolite (Niu 2017). It was first isolated from the phytopathogenic fungus, Nigrospora spp. (Evans et al. 1969), and was later purified from Phoma minispora (Yamamoto et al. 1970; Yamano et al. 1971), Hirsutella thompsonii var. synnematosa (Krasnoff and Gupta 1994), Paecilomyces cateniobliquus (= Isaria cateniobliqua) YMF1.01799 (Wu et al. 2012) and Nigrospora sphaerica (= Khuskia oryzae) (Kim et al. 2001). It has shown nematicidal action against M. incognita; dose-dependent insecticidal activity against apple maggot flies, Rhagoletis pomonella; and mild toxicity to tephritid fruit flies. Apart from that, it has also shown inhibitory actions to spores of Beauveria bassiana and M. anisopliae (Krasnoff and Gupta 1994). The growth inhibition of a wide range of microorganisms including fungi, bacteria and a protozoan is shown by phomalactone (Niu 2017).
4.13 Aurovertins
Aurovertins D, E, F and I were first isolated from the parasitic fungus of root-knot nematode, P. chlamydosporia strain YMF 1.00613 (Niu et al. 2009). Aurovertin D is toxic to the free-living nematode, Panagrellus redivivus (Niu 2017).
4.14 Pseurotin A
It was originally isolated from Pseudeurotium ovalis (Bloch et al. 1976) and is a spirocyclic alkaloid containing oxygen and nitrogen atoms. It was reported as the main metabolite from most of the isolates of P. chlamydosporia propagated in Q6 medium (Hellwig et al. 2003). It acts as a chitin synthase inhibitor (Wenke et al. 1993) and has a moderate effect on phytopathogenic bacteria Erwinia carotovora and Pseudomonas syringae (Niu 2017).
4.15 Oosporein
It was originally obtained from a fungus Oospora colorans as a red-coloured pigment in 1944 (Niu 2017). It was later obtained from Verticillium psalliotae (= Lecanicillium psalliotae) that was selected as an antagonist against fungus causing late blight of tomato, P. infestans (Wainwright et al. 1986). It has a strong inhibitory action especially against Phytophthora infestans (Niu 2017).
4.16 Pyrenocines
The first pyrenocines to be described are pyrenocines A and B which were isolated as the phytotoxic metabolites of Pyrenochaeta terrestris (= Setophoma terrestris) causing pink root disease of onions (Sato et al. 1981). They both were isolated from entomophagous pathogen Verticillium hemipterigenum BCC 1449 (Nilanonta et al. 2003). They have several biological activities, reportedly showing phytotoxicity, cytotoxicity and antifungal, antibacterial, antimalarial and antitrypanosomal activity (Sparace et al. 1987; Krohn et al. 2008).
4.17 Vertinoids
It is a group of compounds that are obtained from Verticillium intertextum ATCC 46284 (Trifonov et al. 1983, 1986). The three secondary metabolites, viz. the hexaketide yellow sorbillin, its derivative 2′,3′-dihydrosorbicillin and the dimeric hexaketide yellow bisvertinoquinol, were reported in 1983, and four new dimeric hexaketides were reported in 1986, viz. bisvertinol, dihydrobisvertinol, isodihydrobisvertinol and bisvertinolone. All the compounds are hexaketide-derived secondary metabolites having two additional methyl groups, one at C2 and the other at C4 of the C12 chain (Niu 2017).
4.18 Vertinolide
It was obtained as a new tetronic acid derivative from the fungus Verticillium intertextum ATCC 46284 (Trifonov et al. 1982). It contains a 4-hydroxy-3,5-dimethyl-2(5H)-furanone-5-yl and an (E,E)-2,4-hexadienon-1-yl substructures with a dimethylene bridge in between.
4.19 Lowdenic Acid
It was isolated from non-sporulating cultures of an undescribed fungus Verticillium spp. (MYC-406 = NRRL 29280 = CBS 102427) (Angawi et al. 2003). It has shown antifungal activity against A. flavus (NRRL 6541), Candida albicans ATCC 90029, Staphylococcus aureus ATCC 29213 and Bacillus subtilis ATCC 6051 (Niu 2017). Lowdenic acid possesses an unusual bicyclic structure containing a furylidene ring which is linked via a C=C double bond to a tetrahydrofurandione ring (Angawi et al. 2003).
4.20 Asteltoxins
They have a trienic α-pyrone structure and are related to citreoviridin and aurovertins. Asteltoxin was the first identified compound as a mycotoxin metabolite of the fungus Aspergillus stellatus (Kruger et al. 1979). Later on, four asteltoxin-type metabolites along with two new asteltoxins were isolated from the fungus Pochonia bulbillosa (=Metapochonia bulbilosa) 8-H-28, which was obtained from the fruiting body of Elaphocordyceps capitata (= Tolypocladium capitatum) (Adachi et al. 2015). Asteltoxin has shown inhibitory action against Escherichia coli BF1-ATPase (Satre 1981).
4.21 Bigutol
Bigutol along with its derivative methylbigutol was isolated from the mycoparasitic fungus Verticillium biguttutum (Morris et al. 1995). They both have prenylated 4-(hydroxymethyl)benzene-1,2-diol moiety in their structure. Bigutol and methylbigutol both inhibit the growth of Rhizoctonia solani and other plant pathogenic fungi (Morris et al. 1995).
4.22 Ascochlorin
Ascochlorin-type compounds were first isolated from the fungus Ascochyta viciae (= Septoria viciae) (Tamura et al. 1968), and later on it has been purified from an array of fungus which includes Fusarium sp. LL-Z1272 (Ellestad et al. 1969), Cylindrocladium ilicicola (= Calonectria pyrochroa) MFC-870 (Hayakawa et al. 1971; Minato et al. 1972), Nectria coccinea (= Neonectria coccinea) (Aldridge et al. 1972), Colletotrichum nicotianae (= Colletotrichum tabacum) (Kosuge et al. 1973), Ascochyta viciae (Sasaki et al. 1974), Acremonium luzulae (= Gliomastix luzulae) (Cagnoli-Bellavita et al. 1975), Cephalosporium diospyri (= Nalanthamala diospyri) IFO 6118 (Kawagishi et al. 1984), Cylindrocarpon lucidum (= Thelonectria lucida) (Singh et al. 1996), a sponge-derived fungus Acremonium sp. (Zhang et al. 2009) and a leafhopper pathogenic fungus, Microcera sp. BCC 17074 (Isaka et al. 2015). The metabolites are a class of a 2,4-dihydroxy-5-chloro-6-methylbenzaldehyde (or 5-chloroorclaldehyde) having a sesquiterpene side chain at C5. In 1994, a series of ascochlorin-type compounds which included a new ascochlorin, 8′,9′-dehydroascochiorin, and five known ascochlorins were identified from Verticillium spp. FO-2787 (Takamatsu et al. 1994). Again in 2004, a new ascochlorin, 8′-hydroxyascochlorin, and a novel ascochlorin glycoside, vertihemipterin A, together with six known ascochlorins, were isolated from the entomophagous fungus Verticillium hemipterigenum (= Torrubiella hemipterigena) BCC 2370 (Seephonkai et al. 2004). The members of ascochlorin-type compounds are known to exhibit antifungal activity (Bal Tembe et al. 1999), antiviral activity and antitumour activity (Takatsuki et al. 1969) (Table 2.2).
5 The Fate of Secondary Metabolites of Metarhizium spp. and Verticillium spp.
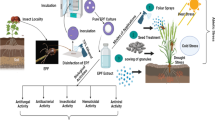
The two fungi are accessible in the market in both solid and liquid formulations containing the spore and the mycelium of the fungus. When these two entomophagous fungi are applied to a crop ecosystem, it comes in direct contact with humans and target insects and to the crop on which it is applied. The indirect interaction of these fungi happens to occur by drifting to soil, water and atmosphere (Hu et al. 2016). Humans are the first ones to come into contact with the cultures of these fungal entomopathogens whether be it the people who are producing the formulation or the people who are applying it on to their field. There are reports of fungal spore allergy caused by the entomopathogenic fungi including M. anisopliae to the workers producing it (Zimmermann 2007), although there are no reports of any sort of things because of the secondary metabolites.
When applied to the crop, the two fungi reach to the proximity of the target insect pests. The spores and mycelium of the fungi adhere to the external surface of insect and then start its infection process. The fungi penetrate through the cuticle of the insect, and various metabolites including various enzymes such as cutinases produced by the fungal mycelia or spores are known to aid in this step of the process. After the penetration, the fungi proliferate itself inside the target insect pest body and then carry on with its life cycle and in the process produce various primary and secondary metabolites. After a successful establishment, the fungi are known to produce the secondary metabolites of which many have the insecticidal effect on the insect pests that are still living up to this stage of fungal infection cycle. So at last, the fungal establishment and its secondary metabolites along with the cadavers of the target insect pests enter the environment. Till now, estimating the number of the fungal metabolites getting released in the environment is very difficult (Hu et al. 2016), although a few of the research point out that the number of metabolites of the entomopathogenic fungi reaching the environment is scarce. As, for example, destruxins, a secondary metabolite of Metarhizium spp., targets insect pests, as the compound decomposes shortly after the death of the host insect pest. The decomposition of destruxin is presumed to be due to the activity of the hydrolytic enzymes in the cadaver, being independent of host or soil and biota, apparently. Thus, destruxins are restricted essentially to the pathogen and the target host and are unlikely to contaminate the environment or enter the food chain (Skrobek et al. 2008).
The target crop along with the weeds also comes in direct contact with the applied entomopathogenic fungus. As the entomopathogenic species of these two fungi are not phytopathogenic, the fungal mycelial and spores in the suspension just only get deposited on the applied plant’s surface but not in those cases where the species of the Metarhizium have been shown to possess endophytic characteristics (Mantzoukas et al. 2015).
Next in the line is the indirect interaction that is caused by the drifting of entomopathogenic fungal formulation to the soil, water and atmosphere while applying. The soil is believed to be the reservoir of the microorganisms including the entomopathogenic fungi. The fungus can survive in the soil as a spore or mycelium and/or in form of any dormant or active structures. Drift from the application and the dropping from the target pest cadavers, fungal spores, mycelia and the metabolites can reach the soil system, but there are no such reports of metabolites of fungal entomopathogen being detected in soil (Hu et al. 2016). Although beauvericins have been detected in drainage water after Fusarium spp. was inoculated on the wheat plants (Schenzel et al. 2012), there are no reports showing metabolites from these two entomopathogenic fungi reaching the water system, and neither are there reports of the same in the atmosphere (Hu et al. 2016).
6 Secondary Metabolites of Metarhizium spp. and Verticillium spp. as Potent Insecticidal Agents
As discussed earlier, the secondary metabolites are the organic compounds which do not play a direct role in organisms’ growth and metabolism (Andersson 2012). Various species of the entomopathogenic Metarhizium and Verticillium fungi along with the other entomopathogenic fungi have been investigated as a source of a wide range of secondary metabolites which possess bioactivities against a broad range of the insect pests. As a result, diversified metabolites have been reported that display insecticidal properties against insect pests (Khan et al. 2012). Destruxins (A and B) (Kodaira 1961a, b), serinocyclin A (Krasnoff et al. 2007), cytochalasins (Vilcinskas et al. 1997a, b), swainsonine (Dreyer et al. 1985) and viridoxins (Gupta et al. 1993) produced from Metarhizium spp. and bassianolides (Champlin and Grula 1979), cyclosporines (Matha et al. 1988; Podsiadlowski et al. 1998), enniatins (Grove and Pople 1980; Strongman et al. 1988; Mule et al. 1992; Monma et al. 2006), dipicolinic acid (Claydon and Grove 1982), verticilides (Monma et al. 2006), phomalactones (Krasnoff and Gupta 1994) and oosporein (Eyal et al. 1994; Wilson 1971) produced from the Verticillium spp. are the secondary metabolites that have shown the insecticidal properties apart from the metabolites of other entomopathogenic fungi that have also shown the insecticidal properties. Secondly, there are also certain extracellular enzymes that are produced by Metarhizium spp. and Verticillium spp. such as chitinase, protease and lipases that also possess certain insecticidal properties (da Silva et al. 2010). So, proper attention is given on the isolation and purification of such enzymes from their producing entomopathogenic fungal species and their utilization in formulations of biopesticides. Some of these formulations also have been patented by their inventors such as the following: an enzyme preparation composed of at least one protease derived from Metarhizium, Beauveria, Verticillium and Aschersonia was formulated and patented (US4987077) (Charnley et al. 1991) and a technology of controlling insect pest prepared with chitinolytic enzymes was patented (US6069299) (Broadway et al. 2000). Similarly, the formulations of the secondary metabolites of Metarhizium spp. and Verticillium spp. that have shown the insecticidal properties can be engineered and used as target-specific green pesticides.
7 Conclusion and Future Perspectives
In this chapter, the secondary metabolites of various Metarhizium spp. and Vetricillium spp. have been exemplified. Most of these compounds exhibited a profound range of biological activities including antifungal, antibacterial, antitumoral, insecticidal and enzyme-inhibiting abilities. In many cases of the secondary metabolites from these two entomopathogenic fungi, only a superficial research is done except for a few of the metabolites. As the new facets of the secondary metabolites are yet to be explored, there is a wide scope of discovering many new compounds as well as the biological activities of the already discovered compounds. There is also a whole new area of using the secondary metabolites of these two fungi along with the other entomopathogenic fungi as pesticide formulations as only few of them are presently available in the market. Although entomopathogenic fungal formulations are present in the market as suspension of mycelia and spore, there is no any prominent product that uses a secondary metabolite as a pest control. There is a need of a full-fledged research that is focused on finding the novel secondary metabolites, proving the different biological activities of the metabolites, standardization of the effective quantity of the metabolites in their biological activities, finding ways to use the metabolites for human welfare, demarcation of the metabolite use in diverse field of science and technology and adverse effect that may occur due to the metabolites. The major problem of the natural product research is its randomness which is obligated to be rectified. Many new technologies in this particular area are waiting to be revealed and put to use in agricultural and agri-allied sectors and also in other sectors. The specificity in the mode of action of these metabolites makes them eco-friendly and, thus, helps in the sustainable development. With the use of these products, we can maintain the balance in nature while meeting the human demands. It is time we should take keen interest in identification of the natural products of the entomopathogenic fungi as the promising new source of bioactive natural compounds because the time has never been more suitable to do so as now we have all the analytical and the molecular tools at our disposal.
References
Abdel-Wahab MA, Asolkar RN, Inderbitzin P, Fenical W (2007) Secondary metabolite chemistry of the marine-derived fungus Massarina sp. strain CNT 016. Phytochemistry 68(8):1212–1218
Adachi H, Doi H, Kasahara Y, Sawa R, Nakajima K, Kubota Y, Nomoto A (2015) Asteltoxins from the entomopathogenic fungus Pochonia bulbillosa 8-H-28. J Nat Prod 78(7):1730–1734
Aldridge DC, Armstrong JJ, Speake RN, Turner WB (1967) The cytochalasins, a new class of biologically active mould metabolites. Chem Commun (Camb) 1:26–27
Aldridge DC, Borrow A, Foster RG, Large MS, Spencer H, Turner WB (1972) Metabolites of Nectria coccinea. J Chem Soc Perkin Trans 1:2136–2141
Amiri B, Ibrahim L, Butt TM (1999) Antifeedant properties of destruxins and their potential use with the entomogenous fungus Metarhizium anisopliae for improved control of crucifer pests. Biocontrol Sci Tech 9(4):487–498
Andersson PF (2012) Secondary metabolites associated with plant disease, plant defense and biocontrol (Vol. 2012, No. 52)
Angawi RF, Swenson DC, Gloer JB, Wicklow DT (2003) Lowdenic acid: a new antifungal polyketide-derived metabolite from a new fungicolous Verticillium sp. J Nat Prod 66(9):1259–1262
Arai M, Yamamoto K, Namatame I, Tomoda H, Omura S (2003) New monordens produced by amidepsine-producing fungus Humicola sp. FO-2942. J Antibiol 56(6):526–532
Aver WA, Peña-Rodriguez L (1987) Minor metabolites of Monocillium nordinii. Phytochemistry 26(5):1353–1355
Ayer WA, Lee SP, Tsuneda A, Hiratsuka Y (1980) The isolation, identification, and bioassay of the antifungal metabolites produced by Monocillium nordinii. Can J Microbiol 26(7):766–773
Azumi M, Ishidoh KI, Kinoshita H, Nihira T, Ihara F, Fujita T, Igarashi Y (2008) Aurovertins F–H from the entomopathogenic fungus Metarhizium anisopliae. J Nat Prod 71(2):278–280
Bal Tembe S, Kundu S, Roy K, Hiremath CP, Gole G, de Souza EP, Pillmoor JB (1999) Activity of the ilicicolins against plant pathogenic fungi. Pest Manag Sci 55(6):645–647
Bloch P, Tamm C, Bollinger P, Petcher TJ, Weber HP (1976) Pseurotin, a new metabolite of Pseudeurotium ovalis stolk having an unusual hetero-spirocyclic system. Helv Chim Acta 59(1):133–137
Bok JW, Lermer L, Chilton J, Klingeman HG, Towers GN (1999) Antitumor sterols from the mycelia of Cordyceps sinensis. Phytochemistry 51(7):891–898
Borel JF, Feurer C, Magnee C, Stähelin H (1977) Effects of the new anti-lymphocytic peptide cyclosporin A in animals. Immunology 32(6):1017
Brackenbury J (1999) Regulation of swimming in the Culex pipiens (Diptera, Culicidae) pupa: kinematics and locomotory trajectories. J Exp Biol 202:2521
Brackenbury J (2001) The vortex wake of the free-swimming larva and pupa of Culex pipiens (Diptera). J Exp Biol 204:1855–1867
Broadway RM, Gary EH, Cornell Research Foundation, Inc (2000) Fungus and insect control with chitinolytic enzymes. US Patent 6,069,299
Bu’Lock JD (1980) Mycotoxins as secondary metabolites. In: Steyn PS (ed) The biosynthesis of mycotoxins. Academic, New York, pp 1–16
Cagnoli-Bellavita N, Ceccherelli P, Fringuelli R (1975) Ascochlorin: a terpenoid metabolite from Acremonium luzulae. Phytochemistry 14:807
Cantin Sanz A, Sanz M, Del Pilar M, Castillo López M, Primo Millo J, Miranda Alonso MÁ, Primo Yufera E (1999) Isolation and synthesis of N-(2-methyl-3-oxodec-8-enoyl)-2-pyrroline and 2-(hept-5-enyl)-3-methyl-4-oxo-6, 7, 8, 8a-tetrahydro-4H-pyrrolo (2, 1-b)-3-oxazine, two new fungal metabolites with in vivo antijuvenile hormone and insecticidal activity. Eur J Org Chem 1:221–226
Carollo CA, Calil ALA, Schiave LA, Guaratini T, Roberts DW, Lopes NP, Braga GU (2010) Fungal tyrosine betaine, a novel secondary metabolite from conidia of entomopathogenic Metarhizium spp. fungi. Fungal Biol 114(5–6):473–480
Cerenius L, Trörnqvist PO, Vey A, Johansson MW, Söderhäll K (1990) The effect of the fungal toxin destruxin E on isolated crayfish haemocytes. J Insect Physiol 36(10):785–789
Champlin FR, Grula EA (1979) Noninvolvement of beauvericin in the entomopathogenicity of Beauveria bassiana. Appl Environ Microbiol 37(6):1122–1126
Charnley K, Richard MC, St. Leger RJ, Agriculture Genetics Company Ltd (1991) Preparations of protease enzymes derived from entomopathogenic fungi. US Patent 4,987,077
Che Y, Swenson DC, Gloer JB, Koster B, Malloch D (2001) Pseudodestruxins A and B: new cyclic depsipeptides from the coprophilous fungus Nigrosabulum globosum. J Nat Prod 64(5):555–558
Claydon N, Grove JF (1982) Insecticidal secondary metabolic products from the entomogenous fungus Verticillium lecanii. J Invertebr Pathol 40(3):413–418
Colegate SM, Dorling PR, Huxtable CR (1979) A spectroscopic investigation of swainsonine: an α-mannosidase inhibitor isolated from Swainsona canescens. Aust J Chem 32(10):2257–2264
Croteau R, Ketchum RE, Long RM, Kaspera R, Wildung MR (2006) Taxol biosynthesis and molecular genetics. Phytochem Rev 5(1):75–97
Da Silva WOB, Santi L, Scharank A, Vainstein MH (2010) Metarhizium anisopliae lipolytic activity plays a pivotal role in Rhipicephalus (Boophilus) microplus infection. Fungal Biol 144:10–15. https://doi.org/10.1016/j.mycres.2009.08.003
Deepa R, Manjunatha H, Krishna V, Kumara Swamy BE (2014) Evaluation of antimicrobial activity and antioxidant activity by electrochemical method of ethanolic extract of Pterocarpus marsupium Roxb Bark. J Biotechnol Biomater 4:166. https://doi.org/10.4172/2155-952X.1000166
Delmotte P, Delmotte-Plaquee J (1953) A new antifungal substance of fungal origin. Nature 171(4347):344
Dhaliwal GS, Arora R (1996) Principles of insect pest management. National Agricultural Technology Information Centre, Ludhiana
Doebler JA (2000) Effects of neutral ionophores on membrane electrical characteristics of NG108-15 cells. Toxicol Lett 114(1–3):27–38
Donadio S, Monciardini P, Alduina R, Mazza P, Chiocchini C, Cavaletti L, Sosio M, Puglia AM (2002) Microbial technologies for the discovery of novel bioactive metabolites. J Biotechnol 99:187–198
Donzelli BGG, Krasnoff SB (2016) Molecular genetics of secondary chemistry in Metarhizium fungi. In: Advances in genetics, vol 94. Academic, Amsterdam, pp 365–436
Dorling PR, Huxtable CR, Vogel P (1978) Lysosomal storage in Swainsona spp. toxicosis: an induced mannosidosis. Neuropathol Appl Neurobiol 4(4):285–295
Dreyer DL, Jones KC, Molyneux RJ (1985) Feeding deterrency of some pyrrolizidine, indolizidine, and quinolizidine alkaloids towards pea aphid (Acyrthosiphon pisum) and evidence for phloem transport of indolizidine alkaloid swainsonine. J Chem Ecol 11(8):1045–1051
Dreyfuss M, Härri E, Hofmann HEA, Kobel H, Pache W, Tscherter H (1976) Cyclosporin A and C. Eur J Appl Microbiol Biotechnol 3(2):125–133
Ellestad GA, Evans RH Jr, Kunstmann MP (1969) Some new terpenoid metabolites from an unidentified Fusarium species. Tetrahedron 25(6):1323–1334
Espada A, Dreyfuss MM (1997) Effect of the cyclopeptolide 90-215 on the production of destruxins and helvolic acid by Metarhizium anisopliae. J Ind Microbiol Biotechnol 19(1):7–11
Evans RH, Ellested GA, Kunstmann MP (1969) Two new metabolites from an unidentified Nigrospora species. Tetrahedron Lett 10:1791–1794
Eyal J, Mabud MA, Fischbein KL, Walter JF, Osborne LS, Landa Z (1994) Assessment of Beauveria bassiana Nov. EO-1 strain, which produces a red pigment for microbial control. Appl Biochem Biotechnol 44(1):65–80
Fiolka MJ (2008) Immunosuppressive effect of cyclosporin A on insect humoral immune response. J Invertebr Pathol 98(3):287–292
Firakova S, Proksa B, Šturdíková M (2007) Biosynthesis and biological activity of enniatins. Die Pharmazie-Int J Pharm 62(8):563–568
Fujii Y, Tani H, Ichinoe M, Nakajima H (2000) Zygosporin D and two new cytochalasins produced by the fungus Metarrhizium anisopliae. J Nat Prod 63(1):132–135
Furumoto T, Hamasaki T, Nakajima H (1997) Vasinfectins A and B: new phytotoxins from Neocosmospora vasinfecta. Tetrahedron Lett 38(31):5523–5524
Gäumann E, Roth S, Ettlinger L, Plattner PA, Nager U (1947) Enniatin, ein neues, gegen Mykobakterien wirksames Antibiotikum. Experientia 3(5):202–203
Griffin DH (1994) Spore dormancy and germination. Fungal physiology, 2nd edn. John Wiley & Sons, New York, pp 375–398
Grove JF, Pople M (1980) The insecticidal activity of beauvericin and the enniatin complex. Mycopathologia 70(2):103–105
Gu Y, Wang Y, Ma X, Wang C, Yue G, Zhang Y, Wen X (2015) Greater taxol yield of fungus Pestalotiopsis hainanensis from dermatitic scurf of the giant panda (Ailuropoda melanoleuca). Appl Biochem Biotechnol 175(1):155–165
Guengerich FP, DiMari SJ, Broquist HP (1973) Isolation and characterization of a l-pyridine fungal alkaloid. J Am Chem Soc 95(6):2055–2056
Gupta S, Krasnoff SB, Renwick JAA, Roberts DW, Steiner JR, Clardy J (1993) Viridoxins A and B: novel toxins from the fungus Metarhizium flavoviride. J Org Chem 58(5):1062–1067
Harborne JB (1986) Recent advances in chemical ecology. Nat Prod Rep 3:323–344
Harrington GJ, Neilands JB (1982) Isolation and characterization of dimerum acid from Verticillium dahliae. J Plant Nutr 5(4–7):675–682
Hartmann T (1985) Prinzipien des pflanzlichen Sekundärstoffwechsels. Plant Syst Evol 150:15–34
Haslam E (1985) Metabolites and metabolism, a commentary on secondary metabolism. Clarendon Press, Oxford, 161 p
Haslam E (1986) Secondary metabolism, fact or fiction. Nat Prod Rep 3:217–249
Hayakawa S, Minato H, Katagiri K (1971) The ilicicolins, antibiotics from Cylindrocladium ilicicola. J Antibiot 24(9):653–654
Heinig U, Scholz S, Jennewein S (2013) Getting to the bottom of Taxol biosynthesis by fungi. Fungal Divers 60(1):161–170
Hellwig V, Mayer-Bartschmid A, Müller H, Greif G, Kleymann G, Zitzmann W, Stadler M (2003) Pochonins A− F, new antiviral and antiparasitic resorcylic acid lactones from Pochonia chlamydosporia var. catenulata. J Nat Prod 66(6):829–837
Herrmann M, Zocher R, Haese A (1996) Enniatin production by fusarium strains and its effect on potato tuber tissue. Appl Environ Microbiol 62(2):393–398
Hino M, Nakayama O, Tsurumi Y, Adachi K, Shibata T, Terano H, Imanaka H (1985) Studies of an immunomodulator, swainsonine. J Antibiot 38(7):926–935
Hiraga K, Yamamoto S, Fukuda H, Hamanaka N, Oda K (2005) Enniatin has a new function as an inhibitor of Pdr5p, one of the ABC transporters in Saccharomyces cerevisiae. Biochem Biophys Res Commun 328(4):1119–1125
Hu Q, Dong T (2015) Non-ribosomal peptides from entomogenous fungi. In: Sree KS, Varma A (eds) Biocontrol of lepidopteran pests. Springer, Cham, pp 169–206
Hu Q, Li F, Zhang Y (2016) Risks of mycotoxins from mycoinsecticides to humans. Biomed Res Int 2016:1
Huxham IM, Lackie AM, McCorkindale NJ (1989) Inhibitory effects of cyclodepsipeptides, destruxins, from the fungus Metarhizium anisopliae, on cellular immunity in insects. J Insect Physiol 35(2):97–105
Iijima M, Masuda T, Nakamura H, Naganawa H, Kurasawa S, Okami Y, Iitaka Y (1992) Metacytofilin, a novel immunomodulator produced by Metarhizium sp. TA2759. J Antibiot 45(9):1553–1556
Isaka M, Yangchum A, Supothina S, Laksanacharoen P, Luangsa-ard JJ, Hywel-Jones NL (2015) Ascochlorin derivatives from the leafhopper pathogenic fungus Microcera sp. BCC 17074. J Antibiot 68(1):47
Iwasaki S, Sair MI, Igarashi H, Okuda S (1970) Revised structure of helvoic acid. J Chem Soc Chem Commun 17:1119–1120
Jalal MA, Love SK, van der Helm D (1986) Siderophore-mediated iron (III) uptake in Gliocladium virens: 1. Properties of cis-Fusarinine, trans-fusarmine, dimerum acid, and their ferric complexes. J Inorg Biochem 28(4):417–430
Jalal MAF, Hossain MB, van der Helm D, Barnes CL (1988) Structure of ferrichrome-type siderophores with dissimilar N δ-acyl groups: Asperchrome B1, B2, B3, D1, D2 and D3. Biol Met 1(2):77–89
Jegorov A, Weiser J (1990) Production of cyclosporins by entomopathogenic fungi. Microbios Lett 45(178):65–69
Kawagishi H, Sato H, Sakamura S, Kobayashi K, Tadao U (1984) Isolation and structure of a new diprenyl phenol, colletorin B produced by Cephalosporium diospyri. Agric Biol Chem 48(7):1903–1904
Kawazu K, Murakami T, Ono Y, Kanzaki H, Kobayashi A, Mikawa T, Yoshikawa N (1993) Isolation and characterization of two novel nematicidal depsipeptides from an imperfect fungus, strain D1084. Biosci Biotechnol Biochem 57(1):98–101
Keswani C (2015a) Proteomics studies of thermotolerant strain of Trichoderma spp. Ph.D. thesis, Banaras Hindu University, Varanasi, India
Keswani C (2015b) Ecofriendly management of plant diseases by biosynthesized secondary metabolites of Trichoderma spp. J Brief Idea. https://doi.org/10.5281/zenodo.15571
Keswani C, Singh SP, Singh HB (2013) Beauveria bassiana: status, mode of action, applications and safety issues. Biotech Today 3:16–20
Keswani C, Bisen K, Singh V, Sarma BK, Singh HB (2016) Formulation technology of biocontrol agents: present status and future prospects. In: Arora NK, Mehnaz S, Balestrini R (eds) Bioformulations: for sustainable agriculture. Springer, New Delhi, pp 35–52
Khambay BPS, Bourne JM, Cameron S, Kerry BR, Zaki MJ (2000) A nematicidal metabolite from Verticillium chlamydosporium. Pest Manag Sci 56(12):1098–1099
Khan S, Guo L, Maimaiti Y, Mijit M, Qiu D (2012) Entomopathogenic fungi as microbial biocontrol agent. Mol Plant Breed 3(1)
Kikuchi H, Hoshi T, Kitayama M, Sekiya M, Katou Y, Ueda K, Oshima Y (2009) New diterpene pyrone-type compounds, metarhizins A and B, isolated from entomopathogenic fungus, Metarhizium flavoviride and their inhibitory effects on cellular proliferation. Tetrahedron 65(2):469–477
Kim JC, Choi GJ, Park JH, Kim HT, Cho KY (2001) Activity against plant pathogenic fungi of phomalactone isolated from Nigrospora sphaerica. Pest Manag Sci 57(6):554–559
Kodaira Y (1961a) Biochemical studies on the muscardine fungi in the silkworms. J Fac Text Sci Technol Sinshu Univ Seric 5:1–68
Kodaira Y (1961b) Toxic substances to insects, produced by Aspergillus ochraceus and Oospora destructor. Agric Biol Chem 25:261–262
Kodaira Y (1962) Studies on the new toxic substances to insects, destruxin A and B, produced by Oospora destructor. Part I. Isolation and purification of destruxin A and B. Agric Biol Chem 26:36–42
Kosuge Y, Suzuki A, Hirata S, Tamura S (1973) Structure of colletochlorin from Colletotrichum nicotianae. Agric Biol Chem 37(2):455–456
Kozone I, Ueda JY, Watanabe M, Nogami S, Nagai A, Inaba S, Ohya Y, Takagi M, Shin-ya K (2009) Novel 24-membered macrolides, JBIR-19 and-20 isolated from Metarhizium sp. fE61. J Antibiot 62(3):159
Krasnoff SB, Gupta S (1994) Identification of the antibiotic phomalactone from the entomopathogenic fungus Hirsutella thompsonii var. synnematosa. J Chem Ecol 20(2):293–302
Krasnoff SB, Gibson DM, Belofsky GN, Gloer KB, Gloer JB (1996) New destruxins from the entomopathogenic fungus Aschersonia sp. J Nat Prod 59(5):485–489
Krasnoff SB, Sommers CH, Moon YS, Donzelli BG, Vandenberg JD, Churchill AC, Gibson DM (2006) Production of mutagenic metabolites by Metarhizium anisopliae. J Agric Food Chem 54(19):7083–7088
Krasnoff SB, Keresztes I, Gillilan RE, Szebenyi DME, Donzelli BGG, Churchill ACL, Gibson DM (2007) Serinocyclins A and B, cyclic heptapeptides from Metarhizium anisopliae. J Nat Prod 70:1919–1924
Krasnoff SB, Englich U, Miller PG, Shuler ML, Glahn RP, Donzelli BG, Gibson DM (2012) Metacridamides A and B, macrocycles from conidia of the entomopathogenic fungus Metarhizium acridum. J Nat Prod 75(2):175–180
Krasnoff SB, Keresztes I, Donzelli BG, Gibson DM (2014) Metachelins, mannosylated and N-oxidized coprogen-type siderophores from Metarhizium robertsii. J Nat Prod 77:1685–1692
Krohn K, Sohrab MH, Draeger S et al (2008) New pyrenocines from an endophytic fungus. Nat Prod Commun 3:1689–1692
Kruger GJ, Steyn PS, Vleggaar R, Rabie CJ (1979) X-ray crystal structure of asteltoxin, a novel mycotoxin from Aspergillus stellatus Curzi. J Chem Soc Chem Commun 10:441–442
Kuboki H, Tsuchida T, Wakazono K, Isshiki K, Kumagai H, Yoshioka T (1999) Mer-f3, 12-hydroxy-ovalicin, produced by Metarrhizium sp. f3. The J Antibiot 52(6):590–593
Kulanthaivel P, Hallock YF, Boros C, Hamilton SM, Janzen WP, Ballas LM, Katz B (1993) Balanol: a novel and potent inhibitor of protein kinase C from the fungus Verticillium balanoides. J Am Chem Soc 115(14):6452–6453
Lee SY, Kinoshita H, Ihara F, Igarashi Y, Nihira T (2008) Identification of novel derivative of helvolic acid from Metarhizium anisopliae grown in medium with insect component. J Biosci Bioeng 105(5):476–480
Levy D, Bluzat A, Seigneuret M, Rigaud JL (1995) Alkali cation transport through liposomes by the antimicrobial fusafungine and its constitutive enniatins. Biochem Pharmacol 50(12):2105–2107
Lin Y, Wu X, Deng Z, Wang J, Zhou S, Vrijmoed LLP, Jones EG (2002) The metabolites of the mangrove fungus Verruculina enalia no. 2606 from a salt lake in the Bahamas. Phytochemistry 59(4):469–471
Lira SP, Vita-Marques AM, Seleghim MH, Bugni TS, LaBarbera DV, Sette LD, Berlinck RG (2006) New destruxins from the marine-derived fungus Beauveria felina. J Antibiot 59(9):553
Liu BL, Tzeng YM (2012) Development and applications of destruxins: a review. Biotechnol Adv 30:1242–1254
Luckner M (1972) Secondary metabolism in plants and animals. Chapman and Hall, London
Luckner M (1990) Secondary metabolism in plants and animals, 3rd edn. Springer, Berlin
Manickam M, Ramanathan M, Farboodniay Jahromi MA, Chansouria JPN, Ray AB (1997) Antihyperglycemic activity of phenolics from Pterocarpus marsupium. J Nat Prod 60(6):609–610
Mantzoukas S, Chondrogiannis C, Grammatikopoulos G (2015) Effects of three endophytic entomopathogens on sweet sorghum and on the larvae of the stalk borer Sesamia nonagrioides. Entomol Exp Appl 154(1):78–87
Matha V, Weiser J, Olejnicek J (1988) The effect of tolypin in Tolypocladiumniveum crude extract against mosquito and blackfly larvae in the laboratory. Folia Parasitol 35(4):379–381
Minato H, Katayama T, Hayakawa S, Katagiri K (1972) Identification of iligigolins with asgochlorin and LL-Z 1272. J Antibiot 25(5):315–316
Minato H, Matsumoto M, Katayama T (1973) Studies on the metabolites of Verticillium sp. structures of verticillins A, B, and C. J Chem Soc Perkin Trans 1:1819–1825
Mishra S, Singh A, Keswani C, Saxena A, Sarma BK, Singh HB (2015) Harnessing plant-microbe interactions for enhanced protection against phytopathogens. In: Arora NK (ed) Plant microbe symbiosis– applied facets. Springer, New Delhi, pp 111–125
Monma S, Sunazuka T, Nagai K, Arai T, Shiomi K, Matsui R, Ōmura S (2006) Verticilide: elucidation of absolute configuration and total synthesis. Org Lett 8(24):5601–5604
Morris RA, Ewing DF, Whipps JM, Coley-Smith JR (1995) Antifungal hydroxymethyl-phenols from the mycoparasite Verticillium biguttatum. Phytochemistry 39(5):1043–1048
Mothes K (1976) Secondary plant substances as materials for chemical high quality breeding in higher plants. In: Mansell RL, Wallace J (eds) Biochemical interaction between plants and insects, Recent advances in phytochemistry, vol 10. Plenum, New York, pp 385–405
Moya P, Cantín Á, Castillo MA, Primo J, Miranda MA, Primo-Yúfera E (1998) Isolation, structural assignment, and synthesis of N-(2-Methyl-3-oxodecanoyl)-2-pyrroline, a new natural product from Penicillium brevicompactum with in vivo anti-juvenile hormone activity. J Org Chem 63(23):8530–8535
Mule G, D’Ambrosio A, Logrieco A, Bottalico A (1992) Toxicity of mycotoxins of fusarium sambucinum for feeding in Galleria mellonella. Entomol Exp Appl 62(1):17–22
Mulheirn LJ, Beechey RB, Leworthy DP, Osselton MD (1974) Aurovertin B, a metabolite of Calcarisporium arbuscula. J Chem Soc Chem Commun 21:874–876
Murakoshi S, Ichinoe M, Suzuki A, Kanaoka M, Isogai A, Tamura S (1978) Presence of toxic substance in fungus bodies of the entomopathogenic fungi, Beauveria bassiana and Verticillium lecanii. Appl Entomol Zool 13(2):97–102
Nakajyo S, Shimizu K, Kometani A, Suzuki A, Ozaki H, Urakawa N (1983) On the inhibitory mechanism of bassianolide, a cyclodepsipeptide, in acetylcholine-induced contraction in Guinea-pig Taenia coli. Jpn J Pharmacol 33(3):573–582
Nilanonta C, Isaka M, Chanphen R, Thong-orn N, Tanticharoen M, Thebtaranonth Y (2003) Unusual enniatins produced by the insect pathogenic fungus Verticillium hemipterigenum: isolation and studies on precursor-directed biosynthesis. Tetrahedron 59(7):1015–1020
Niu XM (2017) Secondary metabolites from Pochonia chlamydosporia and other species of Pochonia. In: Perspectives in sustainable nematode management through Pochonia chlamydosporia applications for root and rhizosphere health. Springer, Cham, pp 131–168
Niu XM, Wang YL, Chu YS, Xue HX, Li N, Wei LX, Zhang KQ (2009) Nematodetoxic aurovertin-type metabolites from a root-knot nematode parasitic fungus Pochonia chlamydosporia. J Agric Food Chem 58(2):828–834
Oide S, Moeder W, Krasnoff S, Gibson D, Haas H, Yoshioka K, Turgeon BG (2006) NPS6, encoding a nonribosomal peptide synthetase involved in siderophore-mediated iron metabolism, is a conserved virulence determinant of plant pathogenic ascomycetes. Plant Cell 18(10):2836–2853
Okuda S, Iwasaki S, Tsuda K, Sano Y, Hata T, Udagawa S, Yamaguchi H (1964) The structure of helvolic acid. Chem Pharm Bull 12(1):121–124
Okuda S, Iwasaki S, Sair MI, Machida Y, Inoue A, Tsuda K (1967) Stereochemistry of helvolic acid. Tetrahedron Lett 24:2295–2302
Pais M, Das BC, Ferron P (1981) Depsipeptides from Metarhizium anisopliae. Phytochemistry 20(4):715–723
Pedras MSC, Zaharia IL, Ward DE (2002) The destruxins: synthesis, biosynthesis, biotransformation, and biological activity. Phytochemistry 59:579–596
Podsiadlowski L, Matha V, Vilcinskas A (1998) Detection of a P-glycoprotein related pump in Chironomus larvae and its inhibition by verapamil and cyclosporin A. Comp Biochem Physiol B: Biochem Mol Biol 121(4):443–450
Poprawski TJ, Robert PH, Maniania NK (1994) Contact toxicity of the mycotoxin destruxin E to Empoasca vitis (Göthe) (Hom., Cicadellidae). J Appl Entomol 117(1–5):135–143
Putri SP, Ishido KI, Kinoshita H, Kitani S, Ihara F, Sakihama Y, Nihira T (2014) Production of antioomycete compounds active against the phytopathogens Phytophthora sojae and Aphanomyces cochlioides by clavicipitoid entomopathogenic fungi. J Biosci Bioeng 117(5):557–562
Ravindra G, Ranganayaki R, Raghothama S, Srinivasan MC, Gilardi RD, Karle IL, Balaram P (2004) Two novel hexadepsipeptides with several modified amino acid residues isolated from the fungus Isaria. Chem Biodivers 1(3):489–504
Robert P, Riba G (1989) Toxic and repulsive effects of spray, ‘peros’ and systemic applications of destruxin E to aphids. Mycopathologia 108(3):179–183
Roberts DW (1966) Toxins from the entomogenous fungus Metarrhizium anisopliae: I. Production in submerged and surface cultures, and in inorganic and organic nitrogen media. J Invertebr Pathol 8:212–221
Roberts DW (1969) Toxins from the entomogenous fungus Metarrhizium anisopliae: isolation of destruxins from submerged cultures. J Invertebr Pathol 14:82–88
Roberts DW (1981) Toxins of entomopathogenic fungi. In: Burges HD (ed) Microbial control of pests. Academic, London, pp 441–464
Rosenthal GA, Janzen DH (1979) Herbivores. Academic, New York
Rothweiler W, Tamm C (1966) Isolation and structure of phomin. Cell Mol Life Sci 22(11):750–752
Sabareesh V, Ranganayaki RS, Raghothama S, Bopanna MP, Balaram H, Srinivasan MC, Balaram P (2007) Identification and characterization of a library of microheterogeneous cyclohexadepsipeptides from the fungus Isaria. J Nat Prod 70:715
Samuels RI, Charnley AK, Reynolds SE (1988) The role of destruxins in the pathogenicity of 3 strains of Metarhizium anisopliae for the tobacco hornworm Manduca sexta. Mycopathologia 104(1):51–58
Sanchez S, Demain AL (2008) Metabolic regulation and overproduction of primary metabolites. Microb Biotechnol 1:283–319
Sanivada SK, Challa MM (2014) Computational interaction of entomopathogenic fungal secondary metabolites with proteins involved in human xenobiotic detoxification. Int J Pharm Pharm Sci 6:312
Sasaki H, Hosokawa T, Nawata Y, Ando K (1974) Isolation and structure of ascochlorin and its analogs. Agric Biol Chem 38(8):1463–1466
Sato H, Konoma K, Sakamura S, Furusaki A, Matsumoto T, Matsuzaki T (1981) X-ray crystal structure of pyrenocine A, a phytotoxin from Pyrenochaeta terrestris. Agric Biol Chem 45(3):795–797
Satre M (1981) The effect of asteltoxin and citreomontanine, two polyenic α-pyrone mycotoxins, on Escherichia coli adenosine triphosphatase. Biochem Biophys Res Commun 100(1):267–274
Schenzel J, Forrer HR, Vogelgsang S, Hungerbühler K, Bucheli TD (2012) Mycotoxins in the environment: I. Production and emission from an agricultural test field. Environ Sci Technol 46(24):13067–13075
Scherlach K, Boettger D, Remme N, Hertweck C (2010) The chemistry and biology of cytochalasans. Nat Prod Rep 27(6):869–886
Seephonkai P, Isaka M, Kittakoop P, Boonudomlap U, Thebtaranonth Y (2004) A novel ascochlorin glycoside from the insect pathogenic fungus Verticillium hemipterigenum BCC 2370. J Antibiot 57(1):10–16
Seigler DS (1998) Plant secondary metabolism. Springer US, Boston. https://doi.org/10.1007/978-1-4615-4913-018
Shelton AM, Zhao JZ, Roush RT (2002) Economic, ecological, food safety, and social consequences of the deployment of Bt transgenic plants. Annu Rev Entomol 47:845–881
Shima M (1955) On the metabolic products of the silkworm muscardines. Bull Sericult Exp Stat 14:427–449
Shinonaga H, Kawamura Y, Ikeda A, Aoki M, Sakai N, Fujimoto N, Kawashima A (2009a) The search for a hair-growth stimulant: new radicicol analogues as WNT-5A expression inhibitors from Pochonia chlamydosporia var. chlamydosporia. Tetrahedron Lett 50(1):108–110
Shinonaga H, Kawamura Y, Ikeda A, Aoki M, Sakai N, Fujimoto N, Kawashima A (2009b) Pochonins K–P: new radicicol analogues from Pochonia chlamydosporia var. chlamydosporia and their WNT-5A expression inhibitory activities. Tetrahedron 65(17):3446–3453
Shiomi K, Matsui R, Kakei A, Yamaguchi Y, Masuma R, Hatano H, Turberg A (2010) Verticilide, a new ryanodine-binding inhibitor, produced by Verticillium sp. FKI-1033. J Antibiot 63(2):77
Sigg HP, Weber HP (1968) Isolierung und Strukturaufklärung von Ovalicin. Helv Chim Acta 51(6):1395–1408
Singh SB, Ball RG, Bills GF, Cascales C, Gibbs JB, Goetz MA, Silverman KC (1996) Chemistry and biology of cylindrols: novel inhibitors of Ras farnesyl-protein transferase from Cylindrocarpon lucidum. J Org Chem 61(22):7727–7737
Singh SB, Zink DL, Dombrowski AW, Dezeny G, Bills GF, Felix JP, Goetz MA (2001) Candelalides A−C: novel diterpenoid pyrones from fermentations of sesquicillium c andelabrum as blockers of the voltage-gated potassium channel Kv1. 3. Org Lett 3(2):247–250
Singh HB, Sarma BK, Keswani C (eds) (2016) Agriculturally important microorganisms: commercialization and regulatory requirements in Asia. Springer, Singapore
Singh HB, Sarma BK, Keswani C (eds) (2017) Advances in PGPR. CABI, Wallingford
Skrobek A, Shah FA, Butt TM (2008) Destruxin production by the entomogenous fungus Metarhizium anisopliae in insects and factors influencing their degradation. BioControl 53(2):361–373
Sparace SA, Reeleder RD, Khanizadeh S (1987) Antibiotic activity of the pyrenocines. Can J Microbiol 33(4):327–330
Springer JP, Cole RJ, Dorner JW, Cox RH, Richard JL, Barnes CL, Van der Helm D (1984) Structure and conformation of roseotoxin B. J Am Chem Soc 106(8):2388–2392
Stierle A, Strobel G, Stierle D (1993) Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 260(5105):214–216
Strasser H, Vey A, Butt TM (2000) Are there any risks in using entomopathogenic fungi for pest control, with particular reference to the bioactive metabolites of Metarhizium, Tolypocladium and Beauveria species. Biocontrol Sci Tech 10(6):717–735
Strongman DB, Strunz GM, Giguere P, Yu CM, Calhoun L (1988) Enniatins from Fusarium avenaceum isolated from balsam fir foliage and their toxicity to spruce budworm larvae, Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae). J Chem Ecol 14(3):753–764
Supothina S, Isaka M, Kirtikara K, Tanticharoen M, Thebtaranonth Y (2004) Enniatin production by the entomopathogenic fungus Verticillium hemipterigenum BCC 1449. J Antibiot 57(11):732–738
Suzuki A, Tamura S (1972) Isolation and structure of protodestruxin from Metarrhizium anisopliae. Agric Biol Chem 36(5):896–898
Suzuki A, Takahashi N, Tamura S (1970) Mass spectrometry of destruxins A and B, insecticidal cyclodepsipeptides produced by Metarrhizium anisopliae. J Mass Spectrom 4(S1):175–180
Suzuki A, Kanaoka M, Isogai A, Tamura S, Murakoshi S, Ichinoe M (1977) Bassianolide, a new insecticidal cyclodepsipeptide from Beauveria bassiana and Verticillium lecanii. Tetrahedron Lett 18(25):2167–2170
Swain T (1977) Secondary compounds as protective agents. Annu Rev Plant Physiol 28:479–501
Takamatsu S, Rho MC, Masuma R, Hayashi M, Komiyama K, Tanaka H, Omura S (1994) A novel testosterone 5α-reductase inhibitor, 8′, 9′-dehydroascochlorin produced by Verticillium sp. FO-2787. Chem Pharm Bull 42(4):953–956
Takatsuki A, Tamura G, Arima K (1969) Antiviral and antitumor antibiotics. XIV. Effects of ascochlorin and other respiration inhibitors on multiplication of Newcastle disease virus in cultured cells. Appl Microbiol 17(6):825–829
Tamura G, Suzuki S, Takatsuki A, Ando K, Arima K (1968) Ascochlorin, a new antibiotic, found by paper-disc agar-diffusion method. I. J Anti Biot 21(9):539–544
Toki S, Ando K, Yoshida M, Kawamoto I, Sano H, Matsuda Y (1992a) ES-242-1, a novel compound from Verticillium sp., binds to a site on N-methyl-D-aspartate receptor that is coupled to the channel domain. J Antibiot 45(1):88–93
Toki S, Ando K, Kawamoto I, Sano H, Yoshida M, Matsuda Y (1992b) ES-242-2,-3,-4,-5,-6,-7, and-8, novel bioxanthracenes produced by Verticillium sp., which act on the N-methyl-D-aspartate receptor. J Antibiot 45(7):1047–1054
Tomoda H, Nishida H, Huang XH, Masuma R, Kim YK, Omura S (1992) New cyclodepsipeptides, enniatins D, E and f produced by Fusarium sp. FO-1305Áü. J Antibiot 45(8):1207–1215
Torssell KBG (1997) Natural products chemistry. A mechanistic, biosynthetic and ecological approach, 2nd edn. Apotekarsocieteten, Stockholm, p 480
Trifonov L, Bieri JH, Prewo R, Dreiding AS, Rast DM, Hoesch L (1982) The constitution of vertinolide, a new derivative of tetronic acid, produced by Verticillium intertextum. Tetrahedron 38(3):397–403
Trifonov LS, Bieri JH, Prewo R, Dreiding AS, Hoesch L, Rast DM (1983) Isolation and structure elucidation of three metabolites from Verticillium intertextum: sorbicillin, dihydrosorbicillin and bisvertinoquinol. Tetrahedron 39(24):4243–4256
Trifonov LS, Hilpert H, Floersheim P, Dreiding AS, Rast DM, Skrivanova R, Hoesch L (1986) Bisvertinols: a new group of dimeric vertinoids from Verticillium intertextum. Tetrahedron 42(12):3157–3179
Tsunoo A, Kamijo M, Taketomo N, Sato Y, Ajisaka K (1997) Roseocardin, a novel cardiotonic cyclodepsipeptide from Trichothecium roseum TT103. J Antibiot 50(12):1007–1013
Turner WB (1971) Fungal metabolites. Academic, London, p 446
Turner W, Aldridge D (1983) Fungal metabolites II. Academic, London
Uchida R, Imasato R, Yamaguchi Y, Masuma R, Shiomi K, Tomoda H, Ōmura S (2005) New insecticidal antibiotics, hydroxyfungerins A and B, produced by Metarhizium sp. FKI-1079. J Antibiot 58(12):804
Usuki H, Toyo-oka M, Kanzaki H, Okuda T, Nitoda T (2009) Pochonicine, a polyhydroxylated pyrrolizidine alkaloid from fungus Pochonia suchlasporia var. suchlasporia TAMA 87 as a potent β-N-acetylglucosaminidase inhibitor. Bioorg Med Chem 17(20):7248–7253
Valadon LRG, Mummery RS (1977) Natural beta apo 4’carotenoic acid methyl ester in the fungus Verticillium agaricinum. Phytochemistry
Vey A, Quiot JM and Vago C (1985) Immunodepressive effect of fungal toxins: inhibition of the reaction of multicellular encapsulation by the destruxins [infection with Aspergillus niger in Galleria mellonella; biological control]. Comptes Rendus de l’Academie des Sciences Serie 3 Sciences de la Vie
Vilcinskas A, Matha V, Götz P (1997a) Effects of the entomopathogenic fungus Metarhizium anisopliae and its secondary metabolites on morphology and cytoskeleton of plasmatocytes isolated from the greater wax moth, Galleria mellonella. J Insect Physiol 43(12):1149–1159
Vilcinskas A, Matha V, Götz P (1997b) Inhibition of phagocytic activity of plasmatocytes isolated from Galleria mellonella by entomogenous fungi and their secondary metabolites. J Insect Physiol 43(5):475–483
Vilcinskas A, Jegorov A, Landa Z, Götz P, Matha V (1999) Effects of beauverolide L and cyclosporin A on humoral and cellular immune response of the greater wax moth, Galleria mellonella. Comp Biochem Physiol C: Pharmacol Toxicol Endocrinol 122(1):83–92
Wahlman M, Davidson BS (1993) New destruxins from the entomopathogenic fungus Metarhizium anisopliae. J Nat Prod 56(4):643–647
Wainwright M, Betts RP, Teale DM (1986) Antibiotic activity of oosporein from Verticillium psalliotae. Trans Br Mycol Soc 86:168–170. https://doi.org/10.1016/S0007-1536(86)80133-4
Waksman SA, Horning ES, Spencer EL (1943) Two antagonistic fungi, Aspergillus fumigatus and Aspergillus clavatus, and their antibiotic substances. J Bacterial 45(3):233
Wallner A, Blatzer M, Schrettl M, Sarg B, Lindner H, Haas H (2009) Ferricrocin, a siderophore involved in intra-and transcellular iron distribution in Aspergillus fumigatus. Appl Environ Microbiol 75(12):4194–4196
Wenke J, Anke H, Sterner O (1993) Pseurotin A and 8-O-demethylpseurotin A from Aspergillus fumigatus and their inhibitory activities on chitin synthase. Biosci Biotechnol Biochem 57(6):961–964
Wilson BJ (1971) Miscellaneous Aspergillus goxins. In: Ciegler A, Kadis S, Aje SJ (eds) Microbial toxins. A comprehensive treatise, vol 6. Academic, New York, pp 288–289
Wu FB, Li TX, Yang MH, Kong LY (2016) Guanacastane-type diterpenoids from the insect-associated fungus Verticillium dahliae. J Asian Nat Prod Res 18(2):117–124
Wu HY, Wang YL, Tan JL, Zhu CY, Li DX, Huang R, Niu XM (2012) Regulation of the growth of cotton bollworms by metabolites from an entomopathogenic fungus Paecilomyces cateniobliquus. J Agric Food Chem 60(22):5604–5608
Xu Y, Orozco R, Wijeratne EK, Gunatilaka AL, Stock SP, Molnár I (2008) Biosynthesis of the cyclooligomer depsipeptide beauvericin, a virulence factor of the entomopathogenic fungus Beauveria bassiana. Chem Biol 15(9):898–907
Yamamoto I, Suide H, Henmi T et al (1970) Antimicrobial α/β-unsaturated δ-lactones from fungi. Takeda Kenkyusho Ho 29:1–10
Yamamoto K, Hatano H, Arai M, Shiomi K, Tomoda H, Omura S (2003) Structure elucidation of new monordens produced by Humicola sp. FO-2942. J Antibiot 56(6):533–538
Yamano T, Hemmi S, Yamamoto I et al (1971) Fermentative production of antibiotic phomalactone. Patent report, Japan. 71 32,800 (Ct. C 12d, A 61k, C 07g). Takeda Chemical Industries Ltd
You F, Han T, Wu JZ, Huang BK, Qin LP (2009) Antifungal secondary metabolites from endophytic Verticillium sp. Biochem Syst Ecol 37(3):162–165
Zähner H, Keller-Schierlein W, Hütter R, Hess-Leisinger K, Deer A (1963) Stoffwechselprodukte von Mikroorganismen. ArchivfürMikrobiologie 45(2):119–135
Zhang P, Bao B, Dang HT, Hong J, Lee HJ, Yoo ES, Jung JH (2009) Anti-inflammatory sesquiterpenoids from a sponge-derived fungus Acremonium sp. J Nat Prod 72(2):270–275
Zimmermann G (2007) Review on safety of the entomopathogenic fungus Metarhizium anisopliae. Biocontrol Sci Tech 17(9):879–920
Acknowledgement
R.N. Yadav is highly thankful to the International Rice Research Institute India for financial support.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Yadav, R.N., Mahtab Rashid, M., Zaidi, N.W., Kumar, R., Singh, H.B. (2019). Secondary Metabolites of Metarhizium spp. and Verticillium spp. and Their Agricultural Applications. In: Singh, H., Keswani, C., Reddy, M., Sansinenea, E., García-Estrada, C. (eds) Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms. Springer, Singapore. https://doi.org/10.1007/978-981-13-5862-3_2
Download citation
DOI: https://doi.org/10.1007/978-981-13-5862-3_2
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-13-5861-6
Online ISBN: 978-981-13-5862-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)