
Abstract
Diversity, dietary, and body mass analyses suggest that the early Eocene represents the major radiation event in South America metatherian evolutionary history. During this period, representatives of all orders typical of the Paleogene reached their greatest diversity (i.e., “basal ameridelphians”; Polydolopimorphia Polydolopiformes, and Bonapartheriiformes Bonapartherioidea); frugivory was the dominant trophic niche. By the middle late Eocene occurs a functional and taxonomic turnover. Among the Polydolopimorphia, frugivore types declined and were replaced by larger-sized frugivores/folivores (Polydolopiformes) and smaller-sized granivores (Bonapartheriiformes). The Sparassodonta showed a diversity increase and occupied the large-sized hypercarnivore niches. The Eocene–Oligocene boundary constitutes another extinction and turnover event marked by the disappearance of “basal ameridelphians”, the Polydolopiformes and Bonapartheriiformes Bonapartherioidea. Lineages that survive into the Deseadan are the Sparassodonta, Paucituberculata, Microbiotheria, and Bonapartheriiformes Argyrolagoidea. Dominant trophic types were those of carnivores and granivores. Environmental factors probably modeled the Paleogene metatherian faunal dynamics in South America. Mean annual temperatures (MAT) and precipitations seem the main factors modeling the taxonomic and trophic diversity, respectively. The adaptive radiation of the early Eocene seems associated with the maximum thermal event of the late Paleocene-early Eocene. The turnover event of the late Eocene seems associated with a sharp drop in the rainfall regime. The extinction and turnover event of the Eocene–Oligocene boundary also seem associated with a strong drop in ambient temperatures. The diversity in evolution of Paleogene metatherians shows a pattern similar to that of living marsupials at the latitudinal level. For a given mean temperature, the number of species in extinct associations is very close to that of the living ones.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
6.1 Introduction
Most of our understanding about the paleobiology of South American metatherians is based on the information provided by molar morphology. Although several postcranial bones (and even complete skeletons) are known for a few species (Argot 2001, 2003, 2004; Abello and Candela 2010; Argot and Babot 2011), they are rare compared with the overwhelming record of metatherians based almost exclusively on dental remains. The use of molar morphology as a paleodietary indicator has a long tradition in mammalian paleobiology (Kay and Hylander 1978; Butler 1983, 1985; Sanson 1989, 1996). All aspects of tooth shape contain information about its function: the overall size and shape of the teeth, the form and topographic location of cusps and valleys, the relative development of crests, the enamel thickness, the development of notches along crests, or the differential arrays of crests or lophs, are all informative about the physical properties of food particles processed by the teeth under study (Lucas 1984, 1994; Lucas and Luke 1984; Hiiemae 2000; Lucas and Peters 2000; Lucas et al. 2000, 2002; Spears and Crompton 1995; Strait and Vincent 1998). Some authors have questioned the use of molar morphology as the single source for paleodietary reconstructions (e.g., Sanson 1996). These authors point out that the mastication is only the first stage in the digestive process and many animals compensate their dental adaptations with modifications of their soft digestive systems. Although this is evident, it has also been well demonstrated that the morphology of masticatory systems is responsible for the acquisition and mechanical processing of foods, while the soft digestive anatomy is mostly related with the chemical digestion process (see Chivers and Langer 1994; Hiiemae 2000). This is why the real diet of mammalian species is close to that one inferred from their molar morphology (see, e.g., Walker et al. 1978; Janis 1984, 1988; Solounias et al. 1988).
Traditionally, paleodietary reconstructions of South American metatherians have been based on the qualitative analysis of molar morphology and the masticatory apparatus. However, these inferences were made in a general context of taxonomical studies rather than in strictly paleobiological ones. Examples of this can be found in the literature for almost all taxonomic groups. The Sparassodonta, for instance, have been commonly categorized as carnivorous metatherians (Marshall 1977, 1978, 1980a; Bond and Pascual 1983; Babot et al. 2002, etc.); the Polydolopimorphia have been treated as frugivorous and insectivorous-frugivorous (Pascual 1980; Marshall 1982a; Goin and Candela 2004; Goin et al. 1999, 2010a, b, etc.); the Paucituberculata have been considered as insectivorous -frugivorous and sometimes folivorous (Marshall 1980b; Bown and Fleagle 1993; Dumont et al. 2000); the Microbiotheria have been included in the niche of insectivorous-frugivorous marsupials (Marshall 1982b; Goin 1997; Goin and Abello 2013; Goin et al. 2007, 2010a, b); the Didelphimorphia have been classified as omnivorous with a variable trend towards carnivory or insectivory (Goin 1991, Goin et al. 1992, 2009); finally, the several families included in the “Ameridelphia” (basal metatherians which do not comprise a natural group; see Chap. 5) have been referred to the carnivorous (Mayulestidae and Herpethotheriidae; de Muizon 1998; Goin and Candela 2004), frugivorous (Protodidelphidae and Pucadelphidae; de Muizon 1998) and insectivorous niches (Jaskhadelphyidae and Derorhynchydae; de Muizon 1991; Goin et al. 1999).
A detailed analysis of the feeding ecology of Paleogene metatherians was recently carried out by Zimicz (2012), who applied the principles and methods of ecomorphology to reconstruct the paleodiet of 97 % species of Paleogene South American metatherians. Her study evaluated the dental morphology from both qualitative and quantitative perspectives, the functional morphology of molar teeth by means of the analysis of wear occlusal patterns and the body mass inferred from dental measurements. The dietary inferences emerged from the integration of the information provided by such different sources. Additionally, she analyzed the diversity patterns at different hierarchical levels, relating them with the evolution of main climatic parameters during Paleogene (MAT and MAP; see below). The present chapter summarizes Zimicz’s (2012) results and main conclusions. It should be noted that the Paleogene period is crucial for the understanding of many biological parameters developed by mammals throughout the world. This is so because of the distinct climate phases that transpired during this time interval. Briefly, Paleogene times saw the climax of Cenozoic’s Greenhouse World (the Early Eocene Climatic Optimum , or EECO), as well as the beginning of the Icehouse World (with the isotopic event known as “Oi1 ” by the Eocene–Oligocene boundary).
6.2 Metatherian Diversity and Climates During the Paleogene in South America
Metatherian diversity throughout the Paleogene shows a main radiation episode by the early Eocene , followed by a fall and stabilization periods in species richness (Fig. 6.1). The first event coincided with the LPTM (Late Paleocene Thermal Maximum) and EECO climatic events (Zachos et al. 2001). These warm episodes were characterized by mean annual temperatures (MAT) above 20 °C, and, in southern South America , by mean annual precipitations (MAP ) higher than 200 cm (Hinojosa 2005). Extremely high values of species richness are recorded in this period: 56 species recovered from the Itaboraian localities of Las Flores (Patagonia) and Itaboraí (Brazil ). This diversity is comprised within no less than six orders that include all Cenozoic lineages (Fig. 6.1). Diversity declines from the late early Eocene (Ypresian –Lutetian boundary) to the middle Eocene with 40 species recovered at the “Sapoan ” localities of Paso del Sapo (Patagonia ), and La Meseta (Antarctic Peninsula) , concomitantly with a progressive fall in MAT values which were established around 17 °C (Wilf et al. 2005). For southern South America, this interval is regarded as one of the most humid of the entire Cenozoic Era, with an estimated MAP of 400 cm (Wilf et al. 2009). By the late middle Eocene diversity equilibrates to around 20 species (during the Barrancan Subage of the Casamayoran SALMA ), and remains stable through the late Eocene–early Oligocene (Mustersan and Tinguirirican SALMAs). Climatic parameters by this interval attest, at least for Patagonia, to a strong reduction in MAP, down to 100 cm or less with relatively stable values of MAT around 17 °C (Fig. 6.2) both inferred from foliar physiognomy of the Río Turbio floral association (Hinojosa 2005). Later, the fossil record indicates a sharp reduction in species richness (only ten recorded species) by the early Oligocene La Cantera Patagonian local fauna (Fig. 6.2). From a paleoclimatic perspective, this fauna represents the span immediately after the Oi1 global cooling event (Zachos et al. 2001). Temperatures fall to lowest values of the Cenozoic with an estimated MAT of about 14.5 °C (Middle Ñirihuau Fm. Flora; Hinojosa 2005). This fall in global temperatures correlates with several oceanographic events that took place at the Eocene–Oligocene boundary, such as the strong drop in eustatic level (Haq et al. 1987), the opening of the Drake Passage , and the establishment of the Antarctic Circumpolar Current (ACC; Lagabrielle et al. 2009; see Chap. 3). The slight increase in the species richness recorded during the late Oligocene (Deseadan SALMA; 14 species; see Fig. 6.2) also correlates with a transient rise in global temperatures (an event known as Late Oligocene Warming , or LOW) and MAT (Hinojosa 2005).

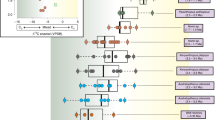
Paleogene chronostratigraphic and biochronologic chart with the biochrons and relative diversity of each of South American metatherian lineages, and the evolution of climatic parameters. References T°, global temperatures (from Zachos et al. 2001); MAP, Mean Annual Precipitation (from Hinojosa 2005); SALMAs, South American Land-Mammal Ages

Diversity and climatic parameters during the Paleogene for South American metatherians. Metatherian species richness is expressed as a function of a Mean Annual Temperatures (TMA; mainly from Hinojosa 2005), b Mean Annual Precipitation (MAP). References Bar, Barrancan Subage (Casamayoran SALMA); Can, La Cantera fauna; Des, Deseadan SALMA; Ita, Itaboraian SALMA; Mus, Mustersan SALMA; Rio, Riochican SALMA; Sap, Paso del Sapo fauna (also named as “Sapoan” in the text); Tin, Tinguirirican SALMA; Tiu, Tiupampan SALMA; Vac, Vacan Subage of the Casamayoran SALMA
The climatic evolution of South America during the Paleogene was accompanied by a succession of floristic assemblages strongly influenced by the evolution of MAT and MAP (Woodburne et al. 2013). During most of the Paleocene and early middle Eocene, megathermal forests dominate the South American landscapes. These forests were composed of Neotropical elements (Romero 1986; Hinojosa et al. 2005; Barreda and Palazzesi 2007). From the late middle Eocene (Barrancan Subage ) to the early Oligocene (Tinguirirican SALMA) there was a remodeling in the floristic associations, which is evidenced by the presence of the austral microthermal genus Nothofagus and other meso- and micro-thermal angiosperms in Patagonian landscapes (Barreda and Palazzesi 2007). The development of savanna-like environments is supported by the record of paleosoils that attest to a transition from humid or subhumid to arid or semiarid conditions during the middle Eocene–early Oligocene (Bellosi and González 2010). Additionally, the late Eocene signals the disappearance from Patagonia of megathermal taxa, which were replaced by micro- and meso-thermal ones (Barreda and Palazzesi 2007). Finally, the Oligocene–Miocene transition in Patagonia verifies a new expansion of mega- and meso-thermal elements and a structural change in the landscapes that show a mixture of forests and grasslands biomes (Barreda and Palazzesi 2007).
It is noteworthy that metatherian diversity throughout the Paleogene shows clear similarities to that of extant South American marsupials when species richness is analyzed as a function of MAT. Birney and Monjeau (2003) concluded that the diversity of living South American marsupials is a function of the latitudinal gradient at a continental scale. For each latitudinal band they tabulated the MAT and MAP , plus other climatic variables. Comparing the species richness of fossil and living faunas of equivalent values of MAT, the result is surprisingly consistent: past diversity is quite similar to the present, both fitting well into a logarithmic curve (Fig. 6.3). Interestingly, fossil assemblages are richer than the living ones for each temperature value. Taking into account the extreme cases: the species richness of the tropical Las Flores association (Itaboraian SALMA) is 36 while any of its living, Neotropical equivalents (latitudinal bands between 10°N and 20°S) have maximum values of 30 species (Zimicz 2012). On the contrary, the cooler La Cantera Patagonian local fauna includes 10 metatherian species, while their living Patagonian equivalent that occurs between 35°S and 40°S has 7 species. This pattern is verified at virtually each latitudinal band and well-known fossil faunas, thus suggesting that the strong influence of temperatures on metatherian biology can be traced to the origin of the group, at least in South America (Zimicz 2012; see also Chap. 2).

Evolution of several trophic indexes during the Paleogene among South American metatherians. a All Patagonian frugivores, b only Patagonian strict frugivores, c all Patagonian carnivores, d all Patagonian granivores, e all South American folivores, f all South American strict insectivores
6.3 Feeding Ecology
6.3.1 Insectivory
Mammals that feed on invertebrate prey are regarded as insectivorous (Feldhamer et al. 2004). Invertebrates are a resource rich in high digestible nutrients such as fats and proteins, in addition to their proportionally high water contents (Hume 1999). Many invertebrates have gregarious habits; this minimizes the energy expenditure that a predator must invest in locating them. However, some restrictions exist in the feeding on invertebrate prey, restrictions which are mostly related to body mass and the input/output energy rate (Schmidt-Nielsen 1975). A medium- or large-sized mammal would require a large amount of invertebrates for satisfying its high-energy requirements, something that implies the investment of many active hours in searching for food. This would obviously unbalance the metabolic equation for a large mammal. On the contrary, a small mammal can exploit advantageously the invertebrate resource because it satisfies its requirements while exclusively feeding on one patch.
From a mechanical perspective, invertebrates are highly variable in their physical properties (Evans 2003). Taking into account exclusively the hardness of the exoskeleton, invertebrates can be subdivided into five categories, from soft to hard (Freeman 1981). This variability in the mechanical food properties has a correlate in the tooth morphology of the predator. In this sense, insectivorous mammals show a great diversity in crown morphology that follows some morphofunctional principles (Lucas 1994). The relative development of trigon and talonid basins and the roundness of cusps and crests reflect the relative hardness of the processed material. The crushing activity needed to comminute hard particles takes place on the molar basins; in turn, soft insect materials do not require large amount of crushing, but instead shearing activity prevails. The relief between the molar trigonid and talonid is also variable, depending of the hardness of the food particle: low relief is indicative of hard invertebrate processing because it is able to generate large crushing stresses (Dewar 2003). On the contrary, high relief is suggestive of soft invertebrate comminution because of the resulting shearing stress needed to reduce them.
Living marsupials that exploit the insectivorous niche in any of their multiple combinations are numerous; some examples are the caenolestids Lestoros , Ryncholestes and Caenolestes ; the microbioterid Dromiciops gliroides, as well as several didelphids as Thylamys , Marmosa , Monodelphis , and Lestodelphys (Hume 1999; Gardner 2007; see Chap. 2). Extinct metatherians that have an inferred insectivorous diet can be found in many different lineages. Almost all Paleogene orders include at least some insectivorous taxa. Among the “Ameridelphia” , main exponents of insectivorous feeding habits are the Derorhynchidae and Jaskhadelphidae, which probably fed strictly on invertebrate resources. Several combinations of insectivory with other dietary categories are exemplified by the Peradectidae, Sternbergiidae and Pucadelphyidae . Among the Polydolopimorphia, only one Bonapartheriiformes ( Praedens aberrans) was strictly insectivorous ; nevertheless, insectivorous habits were widespread when combined with other feeding categories such as frugivory or granivory (e.g., Glasbiidae , Polydolopidae , Bonapartheriidae , and Prepidolopidae). Extinct Paucituberculata had several strictly insectivorous members (e.g., Bardalestes , Evolestes ), as well as many insectivorous-frugivorous types (e.g., Pilchenia , Parabderites , Carlothentes ). Microbiotherians exploited the insectivory niche, mostly related with frugivory through their entire biochron ; some examples are Microbiotherium , Kirutherium , and Mirandatherium . Finally, Paleogene Didelphimorphia exploited insectivory in a strict sense (e.g., Peradectes ), as well as combined with frugivory (e.g., Caroloameghinia , Procaroloameghinia ).
6.3.2 Carnivory
The term refers to the intake of animal tissue as the principal dietary item (Feldhamer et al. 2004). Some authors (Chivers and Langer 1994) used the word “faunivorous ” to refer those animals that feed on other animals. Here, the term “carnivory ” is exclusively employed for those animals that feed exclusively on vertebrate tissues. Although vertebrate meat is a material with highly digestible nutrients, prey acquisition usually involves a great amount of energy expenditure by means of active hunting. This entails some metabolic constraints that are reflected in the body mass of the carnivorous species, which have normally intermediate sizes between the (smaller) insectivorous, granivorous , or frugivorous types on the one hand, and the (larger) herbivorous grazers on the other hand. However, the size range occupied by them is quite large, and size classification requires a particular scale. Consequently, small species are regarded as those weighing less than 2 kg; medium species are those whose body mass range from 2 to 10 kg, while large species are those weighing more than 10 kg.
Carnivorous mammals display a set of unique morphological specializations in the dentition for vertebrate meat comminution, in such a way that they can be studied as a separate guild with respect to the remaining dietary categories. The functional classification used here follows that of Van Valkenburgh (1988) in which hypercarnivore species are those whose diet is exclusively composed by vertebrate meat or bones; mesocarnivore species are those that eat mainly vertebrate meat but can include other items as invertebrates; finally, the hypocarnivore species are those whose diet is mainly composed of nonvertebrate prey such as invertebrates and fruits. Each of the above categories has a morphological correlate that can be measured by a set of morphometric parameters (Van Valkenburgh 1991). Five variables were used for discriminating between the three carnivory subcategories. The relative grinding area (RGA) is the most powerful of them and allows comparison of the relative development of the talonid —which is null in hypercarnivore species and large in hypocarnivore ones. The relative shape of the largest lower premolar (RPS ) indicates the use of this tooth as a crushing or shearing tool, being a sharp crest in hypercarnivore species (except for bone eaters) and a crushing tool in the remaining subcategories. The relative size of the largest lower premolar (PS ), indicates the potential use of the same as the principal element that supports the main forces during the power stroke. The relative height of the dentary or jaw depth (JD ), below the carnassial tooth, is a measure of the load that the shearing complex can support during power stroke; it varies across the subcategories. The shape of the talonid is a qualitative variable that describes the relative development of this structure as well as the number of cusps carried by it. Hypercarnivore species have no talonid in their lower molars; mesocarnivore species have a moderate talonid with one cusp; hypocarnivore species have large talonids with two cusps.
Among living marsupials, only the reddish opossum , Lutreolina crassicaudata (Fig. 2.2b) can be regarded as a mesocarnivore species, while the remaining large didelphids are either hypocarnivore or omnivore species (e.g., Didelphis , Chironectes , Philander ; see Fig. 2.2c–e, respectively). In sharp contrast, Paleogene metatherians show a great diversity of carnivorous species, most of which are referable to the Sparassodonta ; a few other Paleogene lineages also exploited the carnivory niche to some extent (e.g., Herpetotheriidae , Mayulestidae ). Most carnivorous taxa were hypercarnivores and, to a lesser degree, mesocarnivore. Among the Sparassodonta (undoubtedly, the most carnivorous of all metatherian lineages) the mesocarnivory niche was filled by the basal genus Patene . Hypercarnivory was largely occupied by sparassodonts of medium (e.g., Nemolestes ) to large size (e.g., Proborhyaenidae , basal Borhyaenoidea , and Borhyaenidae; although the latter were almost exclusively of Neogene times). Among the wide variety of hypercarnivores, can be mentioned the large predaceous Arminiheringia (middle to late Eocene; Barrancan Subage ) and the large predaceous Pharsophorus and Plesiofelis . A special mention is deserved by the giant proborhyaenids Proborhyaena gigantea and Paraborhyaena boliviana; the large sizes reached by these metatherians (93 and 76 kg each) were comparable to the largest Australasian predaceous marsupials, the Thylacoleonidae (Wroe 2003, 2004). Particularly interesting is the feeding ecology inferred for P. gigantea, whose morphometric indexes suggest hypercarnivorous as well as scavenger habits. Although the large size of P. gigantea prevented the use of bone as a food resource, at least in the context of the Optimal Foraging Theory (Charnov 1976), the widely documented co-existence with large predaceous birds of the family Phorusrhacidae , suggests a positive interaction between both, being birds responsible for the capture and killing of the prey, consuming only meat tissues, while the large proborhyaenids could have taken advantage of the carcasses. This type of interaction is well documented in living and extinct faunas (Palmqvist et al. 2011).
6.3.3 Frugivory
Mammals that feed on reproductive structures of plants are considered frugivorous (Feldhamer et al. 2004). Fruits are variable in their consistency as well as in their chemical composition. The diversity of fruits is reflected in the wide variety of morphotypes among frugivorous mammals. From a mechanical perspective, fruits can be classified as soft and juicy or hard and brittle (see Crompton and Hiiemae 1985; Lucas et al. 2002; Lucas and Peters 2000). Here, hard and brittle fruits are included in the granivory category because they share most physical properties with seeds. Soft fruits do not offer a great amount of resistance to comminution and this is reflected in the tooth morphology of frugivorous mammals. Mainly due to the fact that only lower compressive forces are required for dividing a fruit, frugivorous molars exhibit lower crowns, wide and shallow basins (which are the more prominent character of the molars), lower and rounded cups and crests, and low or null relief between talonids and trigonids in the lower molars. Sometimes, enamel crenulations are visible on the basin of the frugivorous molars .
Living South American marsupials that feed mainly on fruits are the species of the genus Caluromys , Glironia , and Metachirus (see Fig. 2.2a, f). Among the extinct taxa, with the single exception of the Sparassodonta, all remaining lineages of Paleogene metatherians had at least one frugivorous taxon referable to them. Among the “Ameridelphia” the most frugivorous trend was observed in the Protodidelphidae , which exhibited a wide range of body masses, from medium to large. The Polydolopimorphia exploited intensively the frugivory strategy in all combinations. The Glasbiidae (basal Polydolopimorphia) has been identified as frugivorous-insectivorous (e.g. Palangania , Bobbschaefferia ). Among Polydolopidae polydolopiforms there have been recorded some strictly frugivorous types ( Pliodolops ), several frugivorous-insectivorous (e.g., Archaeodolops , Polydolops , Pseudolops ) as well as several frugivorous-folivorous types (e.g., Kramadolops , Amphidolops ). The Bonapartheriidae bonapartheriiforms shows some of its members feeding on fruits and insect resources (i.e., Bonapartherium , Incadolops ). Paucituberculata was a lineage that exploited extensively the strict frugivory (e.g., Pilchenia , Sasawatsu , Perulestes ) as well as combined with insectivory (e.g., Carlothentes , Parabderites ). The Microbiotheria exploited strict frugivorous habits in their medium to large-sized members (i.e., Woodburnodontidae ), as well as the frugivorous -insectivorous niche in small- and medium-sized forms (i.e., Microbiotheriidae , as Microbiotherium and Clenia ).
6.3.4 Granivory
Animals that feed mainly on seeds and nuts are named granivorous (Feldhamer et al. 2004). Seeds have a high nutritional value because of their proportionally large fat contents. Fats are the principal energy reserve in mammals because they provide twice the energy of carbohydrates per weight unit (Randall et al. 1997) and are a food resource widely used by small mammals. From a mechanical point of view, seeds and nuts are hard and brittle materials (see Crompton and Hiiemae 1985; Lucas et al. 2002; Lucas and Peters 2000) that require great compressive loads in order to fracture them. Molars adapted to the processing of seeds and nuts have low crowns, low relief between talonids and trigonids, thick enamel layers, and short, rounded crests. Additionally, other dental elements (incisors and premolars ) commonly show some specializations to granivory , such as their differential enamel distribution or the development of striated crests (Lucas and Peters 2000; Krause 1982).
Paleogene metatherians widely exploited the granivory niche in diverse combinations and with a variety of molar morphologies, always associated to small body masses. Among the Polydolopimorphia , the Bonapartheriiformes developed a variety of granivorous morphotypes. The Prepidolopidae includes several granivorous-insectivorous species (i.e., Prepidolops , Punadolops ). The Rosendolopidae strongly exploited seed resources, all being members of the family strictly granivorous (i.e., Rosendolops , Hondonadia ). All Argyrolagoidea show a trend towards granivory, mostly associated with folivory except for a single insectivorous species ( Praedens aberrans). Both the Groeberiidae and the Argyrolagidae , as well as basal argyrolagoids, display an extraordinary combination of morphological characters that allowed them to exploit alternatively seeds as well as foliage resources.
6.3.5 Folivory
Mammals that feed on leaves and stems of dicot plants are named folivorous (Feldhamer et al. 2004). The foliage consumption requires several digestive adaptations for the processing of a food item characterized by its low digestibility and nutritional content. This requires that the folivorous mammal must devote much time to food intake, develop highly specialized digestive modifications in order to maximize the energy extraction, or both. Living Australasian folivorous marsupials (e.g., Phascolarctos , Pseudocheirus ) use the digestive strategy of hindgut fermenters, in which the caecum and sometimes the proximal colon are strongly enlarged and modified for the digestion of leaves (Chivers and Langer 1994). However, the digestion in the gut is maximized by the reduction of the food particle that is carried out by molars during mastication (Hume 1999). The highly specialized molar teeth of koalas are subselenodont in structure and are composed of a series of curved blades and additionally accessory crests that provide extra cutting edges. Koalas are oblique-chewers, their main chewing activity taking place along all crests, and there is a continuous contact between opposing blades, so that mastication occurs in the horizontal plane (Hume 1999). Paleogene Bonapartheriiformes of the family Gashterniidae display a quite similar pattern, although more simple, than that of Australian folivorous marsupials. Bonapartheriiform molars are composed of a series of curved blades that suggest an oblique movement of the mandible during the power stroke. Molar enamel shows relicts of several crenulations directed at the same angle that the main molar crests. The wear pattern of gashterniids is quite similar to that of Phascolarctos and Pseudocheirus in the relative development of Phase I and Phase II facets during mastication (Zimicz 2012). The strong morphological similarities mentioned above suggest that gashterniids were the most obvious South American folivorous marsupials; their inferred body mass, although much smaller than that of a koala, for instance, lies within the range of folivorous mammals (see Zimicz 2012).
6.4 Trophic Index and Paleogene Climates
The trophic index is a rate that quantifies the relative proportion of some trophic category over the whole fauna or taxonomic association (Zimicz 2012).The frugivory index, for example, measured in an assemblage referable to a particular time span, is the number of frugivorous species over the total species number for that age. Throughout the Paleogene , two trophic indexes show a strong relation with climatic variables among South American metatherian associations: the frugivory and carnivory indexes.
The frugivory index shows a positive correlation with MAT (Zimicz 2012) which is particularly evident during the middle Eocene , when temperatures began to decrease (Fig. 6.4a, b). Frugivorous types dominate the late Paleocene-early Eocene tropical associations; their decline is closely correlated with the fall in average temperatures. On the contrary, the carnivory index shows a negative correlation with MAT (Zimicz 2012). It increases while MAT falls from the middle Eocene to the middle Oligocene (Fig. 6.4c). This association between frugivorous and carnivorous types with MAT also occurs in living faunas of South American marsupials (Birney and Monjeau 2003). The granivory index, although does not show a linear correlation with MAT, is particularly informative because it increases by the middle Eocene, when MAP falls around or below 100 cm (Fig. 6.4d). From the middle Eocene to the late Oligocene, granivorous types dominate the Patagonian metatherian associations together with carnivorous types (Zimicz 2012). Other trophic indexes, such as the insectivorous and folivorous ones, are not correlated with specific climatic variables, and show similar values during most of the Paleogene, independently of MAT or MAP fluctuations (Fig. 6.4e, f).

A comparison between species richness of fossil (black line) and living (gray line) South American metatherian assemblages as a function of the mean annual temperatures (MAT)
6.5 Body Mass
Body size is a measure of the spatial dimension of an organism and is the most important attribute of a biological entity (Damuth and McFadden 1990). It is traditionally measured by mean body mass, a physical quantity whose unit is the kilogram (kg), and it can be inferred for extinct mammals. Traditionally, body size of extinct mammals was inferred from the linear measurement of dental elements, which are highly correlated with body mass (Damuth and McFadden 1990). The information on body mass of extinct species is of great utility in order to understand their ecological role, as well as for the comprehension of evolutionary events that have affected different lineages. This is why the study of the evolution of body mass has a long tradition virtually in all branches of the tree of life (McNab 1971; Stanley 1973; McKinney 1990a, b). Two main theories (rules) about changes in body mass have focused researchers’ attention during most of the last six decades: Cope’s Rule and Bergman’s Rule .
Cope’s Rule describes the widespread trend of animal groups to evolve toward larger physical sizes (Cope 1887). Even though this rule was derived primarily from mammalian studies, other animal groups have been shown as examples of size increase within lineages (Stanley 1973). Although the validity of Cope’s concept has been reputed several times because of its numerous exceptions, its recognition as a rule acknowledges the great number of confirmed cases of evolution towards larger sizes. Several advantages of evolutionary size increases are recognized, the most salient being: improved capacity to capture prey or ward off predators; increased home range and, consequently, a larger availability of foods resources; extended longevity; reduced mortality; greater reproductive success; finally, an increased heat retention per volume unit (Stanley 1973). However, as pointed by Stanley, the intrinsic advantage is a largely oversimplified concept. Small animals are well adapted to their own conditions as are larger ones, and the selective factors that influence the size direction are mostly related to the ancestral size and the limits that the lineage can reach—which are mainly determined by body design as well as phylogenetic or environmental constraints (Mckinney 1990a). The generalized trend to size increase in mammals is, then, the result of a probabilistic process, because small animals are much more common than larger ones (McKinney 1990b). Therefore, the ancestral size of any clade is more probably smaller than larger, in such a way that the only possible trend is to increase or keep stable the body mass. Gould (1988) named this pattern as “asymmetrical expansion of variance around founding lineages.” The asymmetrical nature of this pattern is caused by the placing of the ancestral size near the lower limit of size variation. The same logic is applied to the inverse pattern in which the ancestral size is near the upper limit of size variation, so that the expansion of variance occurs toward smaller sizes (McKinney 1990a).
The geographic variation of body mass within animal groups sometimes follows a pattern toward an increase in size with latitude. This pattern was originally described at the species level (Mayr 1963) and was named as the Bergman’s Rule . Briefly, this rule states that “the races from cooler climates tend to be larger than the same races living at warm climates” (Mayr 1963), at least for warm-blooded vertebrates. The basic assumption of this rule is that the increase in body mass has an energetic advantage in cold environments because the surface/volume ratio is lower in larger animals than in smaller ones (McNab 1971). A consequence of this is that the cost of thermoregulation that depends on the thermal gradient between the animal and the environment diminishes with size increase. Like Cope’s, Bergman’s Rule cannot apply to all animal groups and is mostly restricted to some groups of homeothermic animals, marsupials among them.
The evolution of metatherian body mass during the Paleogene shows that most of the lineages had a body mass below 1000 g; however, during some time intervals this barrier was widely overcome (Fig. 6.5). The early Eocene Itaboraian SALMA constitutes the first episode of a size range expansion toward larger sizes. The large frugivorous Protodidelphidae and the large folivorous Gashterniidae are responsible for this range variation. During the middle to the late Eocene (up to the Barrancan Subage ) there is a new size expansion toward larger sizes, this time staged by the hypercarnivorous Sparassodonta which are also involved in the late Oligocene (Deseadan SALMA) range expansion. In general, throughout the Paleogene, larger sizes were associated to frugivory, folivory, and carnivory , with other dietary categories mostly restricted to sizes below 1 kg. During the Paleogene, the Cope’s Rule is verified at different hierarchical levels among the Polydolopiformes . The same pattern occurs at the family and genus level (i.e., Kramadolops). Polydolopiformes occupied a range from small to large sizes. An inverse pattern to Cope’s Rule is verified in the Microbiotheria and bonapartheriiform polydolopimorphians. The first shows a general trend towards size decrease, which is independent of temperatures (Zimicz in prep.). Microbiotheres were confined mostly to the smaller size ranges (<100 g) but some members reached the medium (100–1000 g) and large (>1000 g) sizes (e.g. Pachybiotherium and Woodburnodontidae ). Most bonapartheriiforms were confined to the smaller size ranges below 100 g (Zimicz 2012) and only some Bonapartheriidae and Gashterniidae reached medium (100–1000 g) and large (>1000 g) sizes respectively.

Evolution of South American metatherians body mass throughout the Paleogene. Boxes represents 50 % of the species in each category; the plus sign (+) represents the value of the median; bars represents the variation range. Reference bm, body mass, estimated from molar measurements using the least square regression equations of Gordon (2003) and Zimicz (2004)
Paleogene metatherians verify Bergman’s Rule , Cope’s Rule and in several cases a pattern of inverse trends to these. The Polydolopiformes polydolopimorphians and the Paucituberculata show several cases of adjustment to Bergman’s Rule at different hierarchical levels. Among the Polydolopiformes, size increase as a function of temperature drop is a process verified at the subordinal, as well as at the family and genus ( Kramadolops and Amphidolops ) levels. Different families of Paucituberculata show clinal increases in body mass as a consequence of temperature drops (Abderitidae and Palaeothentidae ; Zimicz 2004). Woodburnodontid microbiotherians shows an increase in body mass according to temperature fall, an opposite trend to that of the whole order, in which the average body mass diminishes through time (Zimicz 2004). At the subfamily level, both bonapartheriiform polydolopimorphians and the Sparassodonta show an apparent adjustment to Bergman’s Rule in some Lazarous genus or species ( Gashternia and Pharsophorus , respectively). However, in the case of Bonapartheriiformes , although the pattern is latitudinal, it doesn’t correlate with any fall in temperatures because, by the early Eocene , a strong latitudinal gradient in temperatures was not yet established. In the case of the Sparassodonta, the size increase seems to be associated with the evolution of hypercarnivory. By the late Oligocene occurs the giant proborhyanid Proborhyaena gigantea, of about 93 kg (Zimicz 2012). The total size range within the Sparassodonta extends from small (<2 kg) to large (>10 kg) sizes; the largest amplitude of this range was reached between the late Oligocene and the middle Miocene (Zimicz, in preparation).
6.6 Paleogene Macroevolutionary Events
Throughout the Paleogene , overall diversity patterns in all lineages of metatherians exhibit some critical intervals that characterize major biotic events:
Basal ameridelphians were especially abundant during the early Eocene (Itaboraian SALMA). They filled the niches of medium- to large-sized frugivorous, small-sized insectivorous , and small mesocarnivorous types . After the early middle Eocene (“Sapoan”), “ameridelphians” show a strong reduction in their species richness and continue to decline until their extinction in late Eocene.
Polydolopiform polydolopimorphians exhibit a radiation process during the early Eocene (Itaboraian SALMA ) reaching their acme during the early middle Eocene (“Sapoan” age). By this time, the ecological niches filled by the group were mainly frugivorous and insectivorus of small and medium sizes. The middle Eocene is a span of stable diversity for Polydolopiformes ; by the late Eocene their species richness begins to decline. Finally, the last record of a representative of this suborder occurs at the La Cantera local fauna in Patagonia . This final interval (from the Barrancan Subage to the La Cantera times) is ecologically characterized by the dominance of large-sized frugivorous-folivorous types and the virtually absence of small- and medium-sized frugivorous ones.
Among the bonapartheriiform polydolopimorphians, the Bonapartheriioidea shows a similar pattern: a strong radiation process during the early Eocene (Itaboraian SALMA) followed by a stable interval during the middle to late Eocene (Barrancan Subage to Tinguirirican SALMA ). The last record for bonapartheriids transpired at the La Cantera local fauna. The Bonapartheriidae of the early Eocene was large-sized folivorous and medium-sized insectivorous-frugivorous forms. After the middle late Eocene (Barrancan Subage) a strong change towards small-sized granivorous types was recorded for most Bonapartheriioidea (Zimicz 2012). The Argyrolagoidea have their first record during the middle Eocene (Vacan Subage ) and continues at low diversity values (3–4 species) during the remaining Paleogene and Neogene times. They were mainly specialized in the small-sized granivorous and folivorous niches. Their last-known record occurs by the late Pliocene (Marplatan Subage).
Microbiotherians are first recorded by the early Eocene (Itaboraian SALMA; Khasia is regarded here as a probable pedyiomiid; consequently, it is excluded from the Microbiotheria; see Chap. 5). Microbiothere diversity was relatively stable and low, during Paleogene times; by the early Miocene they reached their acme (Goin and Abello 2013) and their record continues through the Neogene up to the recent . By the early middle Eocene, two families of microbiotheres, with different ecological roles, are recorded: Woodburnodontidae (medium- and large-sized frugivorous) and Microbiotheriidae (small-sized insectivorous -frugivorous types). Thereafter, a size constraint toward small sizes occurs with a few exceptions (i.e., Pachybiotherium ) during the Neogene .
Sparassodonta diversity is low until the middle late Eocene (Barrancan Subage ) when several medium- and large-sized hypercanivore types make their appearance. A decline in species richness occurs after the middle Eocene, while a new increase followed by the acme of the group took place during the late Oligocene–early Miocene (from the Deseadan SALMA to the Santacrucian SALMA).
The Paucituberculata shows a low diversity during the Paleogene. The order is recorded since the early Eocene (Itaboraian SALMA); it remained scarce throughout the whole Paleogene . In sharp contrast, the acme of the group took place by the beginning of the Neogene, during the early Miocene (Colhuehuapian and Santacrucian SALMAs ). From an ecological perspective, the niches filled by them during the Paleogene were those of small- to large-sized insectivorous and frugivorous .
The above summary led us to identify three main Paleogene episodes in the evolution of metatherians in South America . The late Paleocene-early Eocene constitutes the main radiation event of the Paleogene and, moreover, of the entire Cenozoic . All Cenozoic lineages of South American metatherians were already present from the early Eocene (Itaboraian SALMA); the main ecological niche filled by them was that of the various frugivorous types. This diversity peak and the domain of frugivorous types indicated by the trophic index correlates with the hyperthermal events LPTM and EECO (Zachos et al. 2001) and the high precipitation regime that configured South America’s tropical landscapes. After the early Eocene, global temperatures experienced a progressive drop only interrupted by the transient warming event of the middle Eocene (MECO ).
The most conspicuous environmental change that took place in southern South America during the early middle Eocene was the abrupt fall in precipitation evidenced in the MAP variation from 400 cm (Laguna del Hunco Flora; Wilf et al. 2009) to 150 cm (Río Turbio Flora; Hinojosa 2005). Metatherian assemblages went with this climatic deterioration declining in species richness; they also experienced several functional and taxonomic turnovers. This biotic event is particularly evident, within the Polydolopidae , in the replacement of small- and medium-sized insectivorous-frugivorous types by the large frugivorous-folivorous ones. The replacement of folivorous and frugivorous bonapartheriids by their granivorous counterparts, as well as the advent of the large- and medium-sized hypercarnivore sparassodonts, constitute additional evidence in favor of the existence of more open (less forested) environments. The Barrancan Subage not only marks the initial radiation of open landscape indicators but also shows the declining and subsequent extinction of groups that could not adapt to the changing conditions (e.g., “basal ameridelphians”). This relatively warm but arid interval extends from the early late Eocene (Barrancan Subage) to the Tinguirirican SALMA .
The most abrupt, drastic episode in Cenozoic metatherian evolution is the Eocene–Oligocene transition which is recorded as a sudden drop in species richness accompanied by the extinction of several lineages and functional turnovers in many of the survivor lineages. This extinction event was global in its impact and has been variously termed in different regions and continents: Grande Coupure in Europe (Sthelin 1910), Terminal Eocene Event in North America (Prothero 1994) or Mongolian Remodeling in Asia (Meng and McKenna 1998). In the context of South American mammals, the extinction event of the Eocene–Oligocene boundary was named the Patagonian Hinge ( Bisagra Patagónica ; Goin et al. 2010a, b). From a taxonomic perspective, the early Oligocene La Cantera local fauna verifies the last record of Polydolopiformes and Bonapartheriiformes Bonapartheriioidea, as well as that of “basal ameridelphians.” From an ecological perspective, this brief interval attests to several adjustments to Bergman’s Rule (i.e., Sparassodonta, Polydolopiformes, Microbiotheria), the radiation of carnivorous and granivorous types, and the virtual absence of frugivorous species. This event was crucial for the future development of metatherian associations. The Sparassodonta, Paucituberculata, Bonapartheriiformes Argyrolagoidea, Didelphimorphia , and Microbiotheria were the only survivors to this “hinge,” and they characterized a new, quite different history of metatherian mammals in South America during the Neogene .
All Paleogene biotic events that affected metatherian faunas in South America can be included in a model of Turnover-pulses (Vrba 1985) or Court Jester hypothesis (Barnosky 2001) in which the climatic factors are the main regulators of the successive mammalian associations. In the case of South American metatherians, both temperatures (global and MAT) and rainfall (MAP) were the factors that mostly influenced their evolutionary history throughout these times—and, quite probably, during the whole Cenozoic .
6.7 Thermal Biology: Key for the Understanding of Metatherian Evolution
Traditionally, ambient temperatures have been considered the main factor that regulates the distribution, diversity and other variables of the life history of living marsupials. The underlying reason for this is the high cost of thermoregulation that prevents marsupials from inhabiting cold environments (McNab 2005). Marsupials have a low ability to survive in cold regions because they allocate virtually all energy to maintain their body temperature; this process depends on the temperature gradient between the individual and its environment (McNab 1986, 2005; see also Chap. 2). Consequently, the energy needed to maintain homeostasis is high, and marsupials must employ diverse strategies for surviving in cold areas. Some of these strategies are the caudal fat storage and hibernation or daily torpor capacity (Geiser 2001; Birney and Monjeau 2003; Bozinovic et al. 2005). Although the effects of temperature on the biology of marsupials are widely known, the impact of precipitation was poorly considered. Fisher et al. (2001) pointed out that the rainfall regime has not influenced the life history variables of marsupials. However, other authors have provided information about the effect of rainfall on many variables of marsupial life strategies (see a review in Lee and Cockburn 1985). Among the most important life history parameters, lactation has a strong impact on the survivorship of the breeding marsupial , a fact that is highly correlated with the seasonal variation in rainfall (Green 1997). It is important to note that the gestation period in marsupials is short, and the young ones are born in a completely altricial condition (Tyndale-Biscoe 1987). Marsupial females perform the highest energy expenditure of the reproductive process during lactation to ensure the complete development of their altricial young (Lee and Cockburn 1985). The lactation period is strongly synchronized with water and food availability (Shaw 2006; see Chap. 2).
The impact of precipitation on the geographic distribution of living South American marsupials has been documented by Birney and Monjeau (2003) and Martin (2008). Other authors have argued that hibernation is triggered by a sustained scarcity of food and water availability in the South American microbiotherian Dromiciops gliroides (Bozinovic et al. 2004; Körtner and Geiser 1998) and other small Australian marsupials (Geiser 2001; Geiser and Ferguson 2001). Food availability is directly linked with rainfall, especially for herbivorous mammals. The successive frugivory indexes emerging from metatherian associations throughout the Paleogene strongly support this assertion. MAP had an impact on Paleogene mammal communities because of the restructuring of landscapes as well as its impact in food availability at the lowest levels of the food web. The result can be observed in the evolution of ecological and taxonomic diversity which, in metatherians, is reflected as biotic events of different intensity.
As summarized above, temperatures as well as precipitation were the most important factors that regulated the life history of Paleogene metatherians. Temperatures are the main regulator of taxonomical diversity; precipitations constitute a complementary factor whose additive effects impact on the ecological configuration of faunas, promoting functional turnovers at different scales.
References
Abello MA, Candela AM (2010) Postcranial skeleton of the Miocene marsupial Palaeothentes (Paucituberculata, Palaeothentidae): paleobiology and phylogeny. J Vert Paleontol 5:1515–1527
Argot C (2001) Functional-adaptive anatomy of the forelimb in the didelphidae, and the paleobiology of the Paleocene marsupials Mayulestes ferox and Pucadelphys andinus. J Morphol 247(1):51–79
Argot C (2003) Functional adaptations of the postcranial skeleton of two Miocene borhyaenoids (Mammalia, Metatheria) Borhyaena and Prothylacynus, from South America. Palaeontol 46:1213–1267
Argot C (2004) Evolution of South American mammalian predators (Borhyaenoidea): anatomical and palaeobiological implications. Zool J Linn Soc 140:487–521
Argot C, Babot J (2011) Postcranial morphology, functional adaptations and paleobiology of Callistoe vincei, a predaceous metatherian from the Eocene of Salta, North-western Argentina. Paleontol 54(2):447–480
Babot MJ, Powell JE, de Muizon C (2002) Callistoe vincei, a new Proborhyaenidae (Borhyaenoidea, Metatheria, Mammalia) from the Early Eocene of Argentina. Geobios 35:615–629
Barnosky AD (2001) Distinguishing the effects of the red queen and court jester on Miocene mammal evolution in the northern Rocky Mountains. J Vertebr Paleontol 21:172–185
Barreda V, Palazzesi I (2007) Patagonian vegetation turnovers during the Paleogene-early Neogene: origin of arid-adapted floras. Botan Rev 73:31–50
Bellosi ES, Gonzales P (2010) Paleosols of the middle Cenozoic Sarmiento Formation, central Patagonia. In: Madden RH, Carlini AA, Vucetich MG, Kay RF (eds) The paleontology of Gran Barranca: evolution and environmental change through the Middle Cenozoic of Patagonia. Cambridge University Press, New York
Birney EC, Monjeau JA (2003) Latitudinal variation in South American marsupial biology. In: Menna J, Dickman C, Archer M (eds) Predators with pouches. The biology of carnivorous marsupials. Csiro Publishing, Colingwood
Bond M, Pascual R (1983) Nuevos y elocuentes restos craneanos de Proborhyaena gigantea Ameghino, 1897 (Marsupialia, Borhyaenidae, Proborhyaeninae) de la edad Deseadense. Un ejemplo de coevolución. Ameghiniana 10(1–2):47–60
Bozinovic F, Ruiz G, Rosenmann M (2004) Energetics and torpor of a South American living fossil, the microbiotheriid Dromiciops gliroides. J Comp Physiol B 174:293–297
Bozinovic F, Ruiz G, Cortés A, Rosenmann M (2005) Energetics, thermoregulation and torpor in the Chilean mouseopossum Thylamys elegans (Didelphidae). Rev Chilena Hist Nat 78:199–206
Bown TM, Fleagle JG (1993) Systematics, Biostratigraphy, and dental evolution of the Palaeothentidae, later Oligocene to earlymiddle Miocene (Deseadan-Santacrucian) caenolestoid marsupials of South America. Mem Paleont Soc 29:1–76
Butler PM (1983) Molar wear facets of early tertiary North American primates. In: Symposium of the 4 international congress of primatology, vol 3, pp 1–27
Butler PM (1985) Homologies of molar cusps and crests, and their bearing on assessments of rodent phylogeny. In: Luckett WP, Hartenberger J-L (eds) Evolutionary relationships among rodents. A multidisciplinary analysis. Plenum Press, New York
Charnov EL (1976) Optimal foraging: attack strategy of a mantid. Am Nat 110(971):141–151
Chivers DJ, Langer P (1994) The digestive system in mammals. Cambridge University Press, Cambridge
Cope ED (1887) The origin of the fittest. D. Appleton & Co, New York
Damuth J, MacFadden BJ (1990) Body size in mammalian paleobiology: estimation and biological implications. Cambridge University Press, Cambridge
de Muizon C (1991) La fauna de mamíferos de Tiupampa (Paleoceno Inferior, Formación Santa Lucía) Bolivia. In: Suarez-Soruco R (ed) Fósiles y Facies de Bolivia, Vol. I: Vertebrados. Rev Técn Yacimientos Petrolíferos Fiscales de Bolivia 12:575–624
de Muizon C (1998) A new carnivorous marsupial from the Paleocene of Bolivia and the problem of the marsupial monophyly. Nature 370:208–211
Dewar EW (2003) Functional diversity within the Littleton fauna (early Paleocene) Colorado: evidence from body mass, tooth structure and tooth wear. Paleobios 23(1):1–19
Dumont ER, Strait SG, Friscia AR (2000) Abderitid marsupials from the Miocene of Patagonia: an assessment of form, function, and evolution. J Paleontol 74(6):1161–1172
Evans AR (2003) Functional dental morphology of insectivorous microchiropterans: spatial modelling and functional analysis of tooth form and the influence of tooth wear and dietary properties. PhD thesis, School of Biological Sciences, Monash University
Feldhamer GA (2004) Mammalogy: adaptation, diversity, and ecology. McGraw-Hill, San Francisco
Fisher DO, Owens IPF, Johnson C (2001) The ecological basis of life history variation in marsupials. Ecol 82(12):3531–3540
Freeman PW (1981) Correspondence of food habits and morphology in insectivorous bats. J Mammal 62(1):166–173
Gardner AL (2007) Mammals of South America, Volume 1: Marsupials, Xenarthrans, Shrews and bats. University of Chicago Press, Chicago
Geiser F (2001) Hibernation: endotherms. In: Enciclopedia of life sciences. Wiley, Chichester
Geiser F, Ferguson C (2001) Intraspecific differences in behaviour and physiology: effects of captive breeding on patterns of torpor in feathertail gliders. J Comp Physiol B 171:569–576
Goin FJ (1991) Los Didelphoidea (Mammalia, Marsupialia, Didelphimorphia) del Cenozoico tardío de la Región Pampeana. PhD thesis, Univ Nac La Plata
Goin FJ (1997) New clues for understanding Neogene marsupial radiation. In: Kay RF, Madden RH, Cifelli RL, Flynn JF (eds) Vertebrate paleontology in the neotropics. The Miocene fauna of La Venta, Colombia. Smithsonian Institution Press, Washington
Goin FJ, Abello MA (2013) Los Metatheria sudamericanos de comienzos del Neógeno (Mioceno temprano Edad Mamífero Colhuehuapense): Microbiotheria y Polydolopimorphia. Ameghiniana 50(I):51–78
Goin FJ, Candela A (2004) New Paleogene Marsupials from the Amazon Basin of Eastern Perú. In: Campbell KE Jr (ed) The Paleogene Mammalian Fauna of Santa Rosa, Amazonian Perú. Nat Hist Mus Los Angeles County, Science Series, vol 40, pp 15–60
Goin FJ, Velázquez C, Scaglia O (1992) Orientación de las crestas cortantes en el molar tribosfénico. Sus implicancias funcionales en didelfoideos (Marsupialia) fósiles y vivientes. Rev Mus La Plata, ns, Paleontol 9(57):183–198
Goin FJ, Woodburne MO, Case JA, Vizcaíno SF, Reguero MA (1999) New discoveries of “opossum-like” marsupials from Antartica (Seymour Island, Middle Eocene). J Mammal Evol 6(4):335–365
Goin FJ, Zimicz AN, Reguero MA, Santillana S, Marenssi SA, Moly JJ (2007) A new Mammal from the Eocene of Antarctica, and the origins of the microbiotheria. Rev Asoc Geol Arg 62(4):597–603
Goin FJ, Candela AM, Abello MA, Oliveira EO (2009) Earliest South American Paucituberculatans and their significance in the understanding of “pseudodiprotodont” marsupial radiations. Zool J Linn Soc 155:867–884
Goin FJ, Abello MA, Chornogubsky L (2010a) Middle Tertiary marsupials from central Patagonia (early Oligocene of Gran Barranca): understanding South America’s Grande Coupure. In: Madden RH, Carlini AA, Vucetich MG, Kay RF (eds) The paleontology of Gran Barranca: evolution and environmental change through the Middle Cenozoic of Patagonia. Cambridge University Press, New York
Goin FJ, Tejedor MF, Abello MA, Martin GM (2010b) Un nuevo microbiotérido (Mammalia, Marsupialia, Microbiotheria) de la Formación Pinturas (Mioceno temprano) de la provincia de Santa Cruz. Ameghiniana 47(1):117–122
Goin FJ, Zimicz AN, Forasiepi AM, Chornogubsky L, Abello MA (2013) The rise and fall of South American metatherians: contexts, adaptations, radiations, and extinctions. In: Rosenberger AL, Tejedor MF (eds) Origins and evolution of cenozoic South American Mammals. Vertebrate Paleobiology and Paleoanthropology Series. Springer, Berlin
Gould SJ (1988) Trends as changes in variances: a new slant on progress and directionality in evolution. J Paleontol 62:319–329
Gordon CL (2003) A first look at estimating body size in dentally conservative marsupials. J Mammal Evol 10(1/2):1–21
Green B (1997) Field energetics and water flux in marsupials. In: Saunders NR, Hinds LA (eds) Marsupial biology: recent research, new perspectives. University of New South Wales Press, Sydney
Haq BU, Handerbol J, Vail PR (1987) Chronology of fluctuating sea levels since the triassic. Science 235:1156–1167
Hiiemae KM (2000) Feeding in mammals. In: Schwenk K (ed) Feeding, form, function and evolution in tetrapod vertebrates. Academic Press, San Diego
Hiiemae K, Crompton AW (1985) Mastication, food transport and swallowing. In: Hildebrand M, Bramble B, Liem K, Wake D (eds) Functional vertebrate morphology. Belknapp Press-Harvard University Press, Cambridge
Hinojosa LF (2005) Cambios climáticos y vegetacionales inferidos a partir de paleofloras cenozoicas del sur de Sudamérica. Rev Chilena Hist Nat 32:95–115
Hume ID (1999) Marsupial nutrition. Cambridge University Press, Cambridge
Janis CM (1984) The use of fossil ungulate communities as indicators of climate and environment. In: Brenchley P (ed) Fossils and climate. Wiley, Chichester
Janis CM (1988) An estimation of tooth volume and hypsodonty indices in ungulate mammals; and the correlation of these factors with dietary preferences. In: Proceedings of the VIIth international symposium on dental morphology. Mem Mus nat Hist Nat 53:367–387
Kay RF, Hylander WL (1978) The dental structure of mammalian folivores with special reference to Primates and Phalangeroidea (Marsupialia). In: Montgomery G (ed) The ecology of arboreal folivores. Smithsonian Institution Press, Washington
Körtner G, Geiser F (1998) Ecology of natural hibernation in the marsupial mountain pygmy-possum (Burramys parvus). Oecologia 113:170–178
Krause DW (1982) Jaw movement, dental function, and diet in the Paelocene multituberculate Ptilodus. Paleobiol 8:265–281
Lagabrielle Y, Godderis Y, Donnadieu Y, Mallabieille J, Suarez M (2009) The tectonic history of Drake Passage and its possible impacts on global climate. Earth Planet Sci Lett 279:197–211
Lee AK, Cockburn A (1985) Evolutionary ecology of marsupials. Cambridge University Press, Cambridge
Lucas PW (1994) Categorization to food items relevant to oral processing. In: Chivers DJ, Langer P (eds) The digestive system in mammals: food, form and function. Cambridge University Press, Cambridge
Lucas PW, Luke DA (1984) Chewing it over: basic principles of food breakdown. In: Chivers DJ, Wood BA, Bilsborough A (eds) Food acquisition and processing in primates. Plenum Press, New York
Lucas PW, Peters CR (2000) Function of postcanine tooth crown shape in mammals. In: Teadford MF, Smith MM, Ferguson MWJ (eds) Development, function and evolution of teeth. Cambridge University Press, Cambridge
Lucas PW, Prinz JF, Agrawal KR, Bruce IC (2002) Food physics and oral physiology. Food Qual Prefer 13:203–213
McKinney ML (1990a) Clasifying and analysing evolutionary trends. In: McNamara KJ (ed) Evolutionary trends. Belhaven Press, London
McKinney ML (1990b) Trends in body-size evolution. In: McNamara KJ (ed) Evolutionary trends. Belhaven Press, London
McNab BK (1971) On the ecological significance of Bergmann’s rule. Ecology 52:845–854
McNab BK (1986) Food habits, energetics and the reproduction of marsupials. J Zool London A 208:595–614
McNab BK (2005) Uniformity in the basal metabolic rate of marsupials: its causes and consequences. Rev Chilena Hist Nat 78:183–198
Marshall LG (1977) A new species of Lycopsis (Borhyaenidae: Marsupialia) from the La Venta Fauna (Late Miocene) of Colombia, South America. J Paleontol 51(3):633–642
Marshall LG (1978) Evolution of the Borhyaenidae, extinct South American predaceous marsupials. Univ California Publ Geol Sci 117:1–89
Marshall LG (1980a) Review of the Hathlyacyninae, an extinct subfamily of South American “dog-like” marsupials. Fieldiana Geol NS 7:1–132
Marshall LG (1980b) Systematics of the South American marsupial family Caenolestidae. Fieldiana Geol NS 5:1–145
Marshall LG (1982a) Systematics of the South American marsupial family Microbiotheriidae. Fieldiana Geol NS 10:1–75
Marshall LG (1982b) Systematics of the extinct South American marsupial family Polydolopidae. Fieldiana Geol NS 12:1–109
Martin GM (2008) Sistemática, distribución y adaptaciones de los marsupiales patagónicos. PhD thesis, Univ Nac La Plata
Mayr E (1963) Animal species and evolution. Belknap Press, Cambridge
Meng J, Mckenna MC (1998) Faunal turnovers of Paleogene mammals from the Mongolian Plateau. Nature 394:364–367
Palmqvist P, Martínez-Navarro B, Pérez-Claros JA, Torregrosa V, Figuerido B, Jiménez Arenas JM, Patrocinio Espigares M, Ros-Montoya S, De Renzi M (2011) The giant hyena Pachycrocuta brevirostris: modelling the bone-cracking behavior of an extinct carnivore. Quatern Int 12:1–19
Pascual R (1980) Nuevos y singulares tipos ecológicos de marsupiales extinguidos de América del Sur (Paleoceno tardío o Eoceno temprano) del noroeste argentino. Actas 2 Congr Arg Paleontol Bioestrat 2:151–173
Prothero DR (1994) The late Eocene-Oligocene extinctions. Ann Rev Earth Planet Sci 22:145–165
Randall D, Burggren W, French K (1997) Eckert. Fisiología animal. Mecanismos y adaptaciones. McGraw-Hill Interamericana, Madrid
Romero EJ (1986) Paleogene Phytogeography and Climatology of South America. Ann Missouri Bot Gard 73(2):449–461
Sanson GD (1989) Morphological adaptations of theeth to diets and feeding in the Macropodoidea. In: Grigg G, Jarman P, Hume I (eds) Kangaroos, Wallabies and Rat-kangaroos. Surrey Beatty and Sons Pty. Limited, New South Wales
Sanson GD (1996) Predicting the diet of fossil mammals. In: Vickers-Rich P, Monaghan JM, Baird RF, Rich TH (eds) Vertebrate Palaeontology of Australasia. Monash University of Publishing Committee, Melbourne
Schmidt-Nielsen K (1975) Scaling in biology: the consequences of size. J Exp Zool 194:287–307
Shaw G (2006) Reproduction. In: Armati PJ, Dickman CRY, Hume ID (eds) Marsupials. Cambridge University Press, Cambridge
Solounias N, Teaford M, Walker A (1988) Interpreting the diet of extinct ruminants: the case of a non-browsing giraffid. Paleobiology 14(3):287–300
Spears RI, Crompton RH (1995) The mechanical significance of the occlusal geometry of great ape molars in food breakdown. J Human Evol 31:517–535
Stanley MS (1973) An explanation for Cope’s rule. Evol 27(1):1–26
Stehlin HG (1910) Remarques sur les faunules de Mammifères des couches Éocènes et Oligocènes du Bassin de Paris. Bull Soc Géol France 9:488–520
Strait SG, Vincent JFV (1998) Primate Faunivores: physical properties of prey items. Int J Primatol 19(5):867–878
Tyndale-Biscoe CH, Renfree MB (1987) Reproductive physiology of Marsupials. Cambridge University Press, Cambridge
Van Valkenburgh B (1988) Diversity in past and present guilds of large predatory mammals. Paleobiology 14(2):155–173
Van Valkenburgh B (1991) Iterative evolution of hipercarnivory in canids (Mammalia, Carnivora): evolutionary interactions among sympatric predators. Paleobiology 17(4):340–362
Vrba ES (1985) Environment and evolution: alternative causes of the temporal distribution of evolutionary events. S Afr J Sci 81:229–236
Walker A, Hoeck HN, Pérez L (1978) Microwear of mammalian teeth as indicator of diet. Science 201(4359):908–910
Wilf P, Johnson KR, Cuneo R, Smith ME, Singer BS, Gandolfo MA (2005) Eocene plant diversity at Laguna del Hunco and Río Pichileufú, Patagonia, Argentina. Am Nat 165(6):10–44
Wilf P, Little SA, Iglesias A, Zamaloa M, Gandolfo MA, Cúneo NR, Johnson KR (2009) Papuacedrus (Cupressaceae) in Eocene Patagonia: a new fossil link to Australasian rainforests. Am J Bot 96(11):2031–2047
Woodburne MO, Goin FJ, Bond M, Carlini AA, Gelfo JN, López GM, Iglesias A, Zimicz AN (2013) Paleogene Land Mammal Faunas of South America; a response to global climatic changes and indigenous floral diversity. J Mamm Evol. doi:10.1007/s10914-012-9222-1
Wroe S, Myers T, Seebacher F, Kear B, Gillespie A, Crowther M, Salisbury S (2003) An alternative method for predicting body mass: the case of the Pleistocene marsupial lion. Palaeobiology 29(3):403–411
Wroe S, Argot C, Dickman C (2004) On the rarity of big fierce carnivores and primacy of isolation and area: tracking large mammalian carnivore diversity on two isolated continents. Proc R Soc London (B) 271:1203–1211
Zachos J, Pagani M, Sloan L, Thomas E, Billups K (2001) Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292:686–693
Zimicz AN (2004) Paleoecología y extinción de los marsupiales con dentición plagiaulacoide de América del Sur: un estudio basado en los cambios en el tamaño corporal. Lic thesis, Univ Nac Patagonia S J Bosco
Zimicz AN (2012) Ecomorfología de los marsupiales paleógenos de América del Sur. PhD thesis, Univ Nac La Plata
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Copyright information
© 2016 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Goin, F.J., Woodburne, M.O., Zimicz, A.N., Martin, G.M., Chornogubsky, L. (2016). Paleobiology and Adaptations of Paleogene Metatherians. In: A Brief History of South American Metatherians. Springer Earth System Sciences. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-7420-8_6
Download citation
DOI: https://doi.org/10.1007/978-94-017-7420-8_6
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-017-7418-5
Online ISBN: 978-94-017-7420-8
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)