
Abstract
With more than 100 species, living South American marsupials (Mammalia, Metatheria) give only a glimpse of the much higher taxonomic and ecological diversity acquired by metatherians throughout the Cenozoic Era. The term Metatheria designs a taxon within Mammalia that includes not only Marsupialia but also all therian mammals more related to Marsupialia than to Eutheria. Several features (e.g., epipubic ones) formerly considered as diagnostic of Metatheria are now regarded either a primitive condition or not present in all members of this group. Other derived features, such as the presence of a shelf-like, inflected angular process in the lower jaw, are consistently present in all metatherians. A brief characterization of all major South American, Cenozoic metatherian lineages is given: “basal ameridelphians,” Sparassodonta, Didelphimorphia, Paucituberculata, Microbiotheria, and Polydolopimorphia (the latter including Polydolopiformes and Bonapartheriiformes). Three periods can be distinguished in the history of our knowledge of Cenezoic South American Metatheria: the first one (1878–1930) is intimately linked to Florentino Ameghino, Argentina’s most notable paleontologist; much of our knowledge on extinct metatherians from South America was elaborated by him. The second period (1930–1977) occurred under the influence of George Gaylord Simpson’s ideas. Bryan Patterson and Rosendo Pascual also had an important imprint in South America’s Mammalian Paleontology. The third period (1977-present) is currently evolving under new phylogenetic, taxonomic, and paleobiogeographic paradigms; influences are multiple and major reviews of specific lineages are currently in the making. A final note on the incompleteness of Cenozoic South America’s fossil record is made: only the mid to high latitudes, basically in the Southern Cone, are moderately well-sampled in their terrestrial fossil record.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Figure 1.1 shows a young (subadult ) specimen of the Pampean , short-tailed opossum , Monodelphis dimidiata, in several postures. The specimen was captured at the basal slopes of Sierras Bayas , near the town of Balcarce, in central Buenos Aires Province, Argentina. It is a quite small individual, less than 20 g in body mass and a head and body length of 95 mm. Throughout the Cenozoic Era, many of the metatherians that populated South America were of comparably small size, an inconspicuous aspect, and generalized morphology. Most of them weighed less than 1 kg. Their probably nocturnal or crepuscular habits and, for many, arboreal biotopes, made them even more inconspicuous. This is clearly the case of the single living group of metatherians in South America, the Marsupialia (see below for a characterization of Metatheria and Marsupialia). Australasian marsupials , on the contrary, have developed a much wider variety of taxonomic, morphological, and adaptive types.

A single, subadult specimen of Monodelphis dimidiata from the Balcarce area in Buenos Aires Province, central Argentina. The specimen is shown in frontal (a–c, e), ventral (d), and dorsal (f) views. Photographs by Bruno Pianzola
When considering the diversity of all Cenozoic metatherians in South America, the panorama widens considerably. This is so even taking in account that metatherians were just one of the three major lineages of therian mammals that evolved in this continent since early Paleogene times—the remaining two being the Xenarthra (armadillos, glyptodonts, sloths, ant-eaters, and related taxa), and the South American native ungulates (abbreviated SANU ), a diverse array of medium- to very large-sized herbivores: Litopterna , Notoungulata , Xenungulata , Astrapotheria , and Pyrotheria (no SANU survived to the Recent ). Both Xenarthrans and SANU developed numerous types of browsing and grazing herbivores of middle to very large size; that is the main reason why metatherians did not exploit those niches, especially the grazing ones (see, e.g., Simpson 1950; Patterson and Pascual 1968; Pascual 2006). Notwithstanding, the taxonomic, morphological, and adaptive types developed by South American Cenozoic metatherians is still impressive. An idea of this diversity is suggested by the fact that they are currently referred to 33 families and no less than five orders (see Table 5.1).
1.1 Abbreviations and Conventions
The following abbreviations and conventions are used throughout the whole volume.
1.1.1 Abbreviations
- AAG:
-
Australo-Antarctic Gulf
- ACC:
-
Antarctic Circumpolar Current
- BAT:
-
Brown adipose tissue
- BMR:
-
Basal metabolic rate
- °C:
-
Degree Celsius or centigrade
- CTM:
-
Cretaceous Thermal Maximum
- CT scan:
-
Computed Tomography Scan
- EECO:
-
Early Eocene Climatic Optimum
- EOB:
-
Eocene–Oligocene boundary
- FABI:
-
First American Biotic Interchange
- FMR:
-
Field metabolic rate
- g:
-
Grame
- GABI:
-
Great American Biotic Interchange
- GHG:
-
Greenhouse gases
- JD:
-
Relative height of the dentary (or jaw depth)
- K-Ar:
-
(40K–40Ar) Potassium–Argon radiometric dating
- kg:
-
Kilogram
- K/Pg:
-
Cretaceous–Paleogene (boundary)
- ky:
-
One thousand years (see Chap. 3 for a definition)
- LGM:
-
Last Glacial Maximum
- LOW:
-
Late Oligocene Warming
- LPTM:
-
Latest Paleocene Thermal Maximum (now regarded as PETM)
- Ma:
-
Megannum (see Chap. 3)
- MAP:
-
Mean annual precipitation
- MAT:
-
Mean annual temperature
- MECO:
-
Middle Eocene Climatic Optimum
- MLP:
-
Museo de La Plata
- mm:
-
Millimeter
- MMCO:
-
Middle Miocene Climatic Optimum
- MNI:
-
Minimal number of individuals
- my:
-
One million years (see Chap. 3)
- NALMA:
-
North American Land Mammal Age
- ODP:
-
Ocean Drilling Program
- Oi1:
-
A basal Oligocene event, associated with extreme cooling, in the oxygen isotope values of marine carbonates
- PETM:
-
Palecene–Eocene Thermal Maximum
- PS:
-
Relative size of the largest lower premolar
- RGA:
-
Relative grinding area
- RPS:
-
Relative shape of the largest lower premolar
- SALMA:
-
South American Land-Mammal Age
- SANU:
-
South American Native Ungulates
- SEG:
-
Southeastern Gondwana
- SEM:
-
Scanning Electron Microscopy
- SST:
-
Sea Surface Temperature
- STR:
-
South Tasman Rise
- SWG:
-
Southwestern Gondwana
- TLV:
-
Thoracolumbar vertebrae
- y.b.p.:
-
Years before present
1.1.2 Dental Nomenclature
The generalized dental formula for metatherians is: I/i 5/4, C/c 1/1, P/p 3/3, M/m 4/4, being I, upper incisor ; I, lower incisor; C, upper canine ; c, lower canine ; P, upper premolar ; p, lower premolar; M, upper molar; m, lower molar. Figure 1.2 shows a lower (a) and an upper (b) molar of a generalized “opossum-like ” metatherian , with the nomenclature used here for cusps (in upper case letters), crests, cinguli and basins.

Molar nomenclature used in this work. a Lower left molar; b upper left molar. In (b) the centrocrista includes the postparacrista + the premetacrista
1.1.3 Chronology
Figure 1.3 illustrates South America’s Cenozoic biochronological succession. We follow the standard chronological scale of Gradstein et al. (2012). South America’s biochronological scheme largely follows that of Goin et al. (2012), excepting the ages of the Peligran SALMA and the Carodnia Zone , for which we follow Clyde et al. (2014). Late Cenozoic biochronology (latest Miocene onward) follows Tomassini et al. (2013). Late Pleistocene–Holocene chronology follows Tonni (2009).

South America’s Cenozoic biochronological succession. After Clyde et al. (2014, early Paleocene), Woodburne et al. (2014a, b, early-middle Eocene ), Dunn et al. (2013, late Eocene-early Miocene), Tomassini et al. (2013, late Miocene-Pliocene), Tonni (2009, late Pleistocene-Holocene). The “ Carodnia Zone ,” “Sapoan ,” “Pinturan,” “Colloncuran,” and “Mayoan” have not been formalized as SALMAs. Vacan and Barrancan are subages of the Casamayoran SALMA. For Paleogene biochronological units, see a review in Woodburne et al. (2014a)
1.1.4 Classification
Table 5.1 shows the classification of South American metatherians, above the generic level, used in this study. Families including living representatives are marked in bold. See below and Chap. 5 for a brief characterization of extinct groups.
1.2 Metatheria and Marsupialia
1.2.1 Fossils and Radiations
The oldest metatherian known up to now is the Early Cretaceous (ca. 125 Ma) Sinodelphys szalayi, from the Yixian Fm of Liaoning Province, northeastern China (Luo et al. 2003). It was a small-sized creature (around 30 g of body mass) with arboreal capabilities. Based on this discovery, Luo et al. (2003) argued in favor of the hypothesis that Asia was the likely center for diversification of metatherians (and of eutherians as well). In turn, Cifelli and Davis (2003) noted striking differences in the patterns of therian diversification between Asia and North America : “Eutherians dominated in Eurasia throughout the Cretaceous , but were absent from North America through most of the Late Cretaceous and did not attain appreciable diversity there until the last ~10 million years of the period” (Cifelli and Davis 2003: 1900). To these authors, this pattern constitutes a puzzle that remains to be solved.
According to Luo et al. (2003), the metatherian fossil record and phylogeny suggest a series of successive, major episodes of diversification: (1) the divergence of metatherians and eutherians in Asia occurred no later than 125 Ma in the Early Cretaceous ; (2) it was followed by the evolution of deltatheroidan-like metatherians both in Asia and North America during the late Early Cretaceous (120 to 100 Ma); (3) then, a major diversification episode occurred in North America by the early Late Cretaceous (around 100 Ma); and finally (4) the Paleocene diversification of proximal relatives to crown marsupials occurred in South America . (As discussed in Chaps. 3 and 7, it is more probable that this event happened by the Late Cretaceous). To these, we could add a fifth major episode: (5) the (possibly, latest Cretaceous ; see Chaps. 3, 4, and 7) for the origin and later radiation of Australidelphians and their dispersal throughout southern South America, Antarctica , and Australia . The history of Southern Hemisphere metatherians involves the last two steps of this succession.
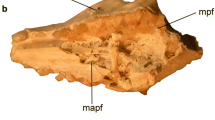
The oldest record of a therian mammal in South America is that of Cocatherium lefipanum, from the earliest Paleocene of western Patagonia (Paso del Sapo area, Chubut Province; Goin et al. 2006). It consists of an isolated, worn lower molar (Fig. 1.4) recovered from levels of the Maastrichtian-Danian Lefipán Fm, and referred by Goin et al. (2006) to the Polydolopimorphia (Marsupialia ). A striking aspect of Cocatherium is that, even though it is the oldest South American metatherian, it is not generalized but instead quite derived in its molar pattern . For instance, (1) it is bunodont , (2) the talonid is relatively short, (3) the labial face of the molar doubles the height of the lingual face, (4) the paraconid is reduced and set close to the metaconid , (5) the protoconid is robust but low, probably of the same height as the also robust metaconid, and finally (6) the talonid has a tall, laterally compressed entoconid . Taking in account that the specimen was found less than 5 m above the inferred Cretaceous -Tertiary boundary at the Lefipán Fm, the degree of specialization shown by Cocatherium is suggestive of an earlier (Late Cretaceous) evolution of this and other metatherian lineages. This agrees with the idea of a Late Cretaceous arrival of the first immigrant lineage (lineages? see below) from North America. As suggested in Chap. 3, this arrival could have taken place either by Campanian or by Maastrichtian times. In addition, Case et al. (2005) argued that at least “didelphimorphians” (now regarded as basal ameridelphians) and polydolopimorphians (typical South American metatherians), had their origins in North America, probably in southern latitudes of that continent, and then dispersed into South America by the end of the Cretaceous. Forasiepi (2009) also suggested that the Sparassodonta had a Late Cretaceous, North American origin, previous to their dispersal into South America.

Cocatherium lefipanum (Metatheria, Polydolopimorphia), the oldest Cenozoic therian mammal known up to now from South America. Specimen LIEB-PV 1001 , an isolated, lower right molar in labial (a) and occlusal views. Abbreviations Pa, paraconid ; Me, metaconid ; Pr, protoconid; Hy, hypoconid ; En, entoconid; Hyl, hypoconulid. Modified after Goin et al. (2006)
In his thorough description of the early Paleocene mammalian assemblage of Tiupampa (Bolivia ; Tiupampan SALMA ), Muizon (1991) noted that the Tiupampan eutherians show much closer affinities to those of North America (Puercan NALMA ) than those shown by metatherians. He concluded that placental mammals arrived in South America in a dispersal event that was not synchronous with, but subsequent to that of metatherians. Other authors agreed with this (e.g., Pascual and Ortiz Jaureguizar 1991) and suggested that during the latest Cretaceous and the earliest Paleocene , there probably were several dispersal events between the Americas, the earliest of which may have involved the metatherian immigration into South America (see Chap. 3). More recently Goin et al. (2012) named this inferred dispersal event as FABI (First American Biotic Interchange) , a term that is reminiscent of the complexities and sequential aspect of the late Tertiary GABI, the Great American Biotic Interchange (see Chap. 4).
1.2.2 Definitions and Characteristic Features
Since the last decades of the twentieth century, the term Marsupialia has acquired a more restricted sense than in previous interpretations. Today it is regarded as the crown group including all extant marsupials, their common ancestor, and all of their descendants Rowe (1998). In turn, Metatheria is a more comprehensive term for designing a taxon within Mammalia that includes not only Marsupialia but also all therian mammals more related to Marsupialia (i.e., their stem relatives) than to Eutheria (Rougier et al. 1998; Kielan-Jaworowska et al. 2004; Williamson et al. 2014). Hence, by definition, all non-marsupial metatherians are extinct—as well as many Cenozoic marsupials. Because of this distinction, it is quite difficult to extend to all metatherians the series of features of soft anatomy, reproduction, and physiology that characterize living marsupials (see Chap. 2 for a detailed characterization of these features among living South American marsupials). Notwithstanding, it is reasonable to assume that many of these characters were also present in non-marsupial metatherians. Several features of metatherian physiology (as, for instance, the lack of Brown Adipose Tissue, or BAT) are a potential source of biases in the evolution of the group (see Chap. 2; Sánchez-Villagra 2012).
Several features of the hard parts (skeleton and teeth) of metatherians that have been considered diagnostic of this group are not exclusive of them. This is the case, for instance, of the presence of epipubic bones in the pelvic girdle , a primitive condition as they have also been found in early eutherians (Kielan-Jaworowska 1975). The presence of an alisphenoid bulla is a derived feature within several lineages rather than a metatherian synapomorphy (see, e.g., Kielan-Jaworowska et al. 2004). In the middle ear , the absence of a groove for the stapedial artery on the promontorium is also present in eutherians and Vincelestes , though a few others, such as the lack of a foramen for the superior ramus of the stapedial artery, absence of the related ascending canal, the posterolateral placement of the transverse sinus with respect to the subarcuate fossa , and the separation of the jugular foramen from the inferior petrosal opening, have been referred to as metatherian synapomorphies (Rougier et al. 1998). The presence of an angular process in the rear part of the dentary is also frequently cited as a synapomorphy of Metatheria (it is absent in Sinodelphys ). Even though this feature is also present in a few Cretaceous eutherians, its shelf-like aspect is exclusive of the group (Averianov and Kielan-Jaworowska 1999).
Luo et al. (2003) listed several derived features of the postcranial skeleton that are quite constant among metatherians: (1) Manus : carpals with a hypertrophied hamate (relative to the capitate and trapezoid ), an enlarged triquetrum (relative to the lunate and distal ulna ), and an enlarged scaphoid (relative to the lunate and/or trapezium). (2) Pes: tarsals have a transversely broad but anteroposteriorly short navicular ; the navicular facet on the astragalar head is spread medially along the length of the neck, such that the head with its navicular facet is asymmetrical with regard to the main axis of the astragalar neck; the calcaneocuboid facet is obliquely oriented with respect to the length of the calcaneus , and is buttressed by a large anteroventral tubercle; the base of the peroneal process is level with the cuboid facet or anterior to it; the sustentacular process forms a pointed triangle. (3) Typical metatherian features in the forelimb are: much wider supraspinous fossa than infraspinous fossa at midlength of the scapula ; the cranial border of the scapula has a strongly sigmoidal profile, ending anteriorly in a pronounced supraspinous incisure ; the ectepicondylar region of the humerus has a shelf-like supinator crest with a sigmoidal profile.
Several dental features are also common among metatherians but not exclusive of the group: a generalized dental formula of I/i 5/4, C/c 1/1, P/p 3/3, M/m 4/4, molars with a typical tribosphenic aspect, or the sharp morphological break between the premolar and molar series. Kielan-Jaworowska et al. (2004) list two features as synapomorphies of Metatheria : the reduction in the premolar number (from four or five in the ancestral boreosphenidans to three in metatherians), and the reduction of diphyodonty (i.e., the presence of two successive sets of teeth, deciduous and permanent), restricted to the third premolar (Luckett 1993). Some other features listed by Luo et al. (2003; see also O’Leary et al. 2013) as synapomorphies of Metatheria are the presence of an entoconid which is close, or twinned to the hypoconulid in the lower molars, presence of a stylar cusp B (StB, opposite the paracone ) at least on the upper second molar; and presence of a stylar cusp D (StD, opposite the metacone ) at least on the penultimate upper molar.
In their extensive morphological analysis of 229 dental, cranial, postcranial, and soft tissue characters and selected mammalian taxa, Horovitz and Sánchez-Villagra (2003) listed the following unambiguous apomorphies as diagnostic of Metatheria: four upper molars; staggered third lower incisor ; marsupial pattern of dental replacement; angular process of the dentary medially inflected; the palatal process of the premaxilla reaches the canine alveolus or is immediately posterior to it, and absence of the stapedial artery sulcus on the petrosal . In turn, they regarded the following unambiguous apomorphies as diagnostic of Marsupialia: humerus with equal proximal extension of capitulum and trochlea ; spherical distal process of ulna , contributing to a ball and socket articulation; posterior shelf of the tibia present and extending posteriorly beyond the medial astragalotibial facet; dorsal mesiolateral orientation of the calcaneal sustentacular facet; poorly developed alisphenoid tympanic wing ; transverse canal foramen anterior to the carotid foramen , and palatal vacuities present in both maxillary and palatine bones.
1.3 South American Metatherians, Living and Extinct
With slightly more than 100 species (Brown 2004; Voss and Jansa 2009; Wilson and Mittermeier 2015), living, South American marsupials account for around 10 % of the terrestrial mammals fauna of South America (i.e., excluding volant and marine mammals; Goin et al. in press). They are currently grouped in three orders: Didelphimorphia , Paucituberculata , and Microbiotheria —several didelphid didelphimorphians are also distributed in Central and North America. Living didelphimorphians are grouped in the family Didelphidae (Didelphoidea) ; they are, by far, the largest group of living marsupials in the New World (more than 90 species). According to recent systematic accounts, they are classified in four subfamilies: Glironiinae , Caluromyinae , Hyladelphinae , and Didelphinae (Voss and Jansa 2009; see Chap. 2). Extinct didelphimorphians include the Sparassocynidae (also referable to the Didelphoidea), a clade of mesocarnivorous didelphoids around the size of the large living didelphines (500–600 g; Zimicz 2014), as well as the Peradectoidea (Horovitz et al. 2009) with two families: Peradectidae (but see Williamson et al. 2012) and Caroloameghiniidae , the latter bearing several convergent features with small-sized primates (Goin 2006).
Living paucituberculatans are restricted to one family, the relatively generalized, shrew-like Caenolestidae , with around half a dozen species. Extinct paucituberculatans (with around 50 species; Abello 2007) are included in this and three additional families: Pichilipidae , Palaeothentidae , and Abderitidae , the latter bearing some of the most derived dental adaptations for the whole order, as a plagiaulacoid -like first molar (not premolar; Marshall 1980). The earliest record of the Paucituberculata dates back to the early Eocene (Itaboraian SALMA). However, the paucituberculatan record is scarce throughout most of the Eocene. They reached their climax by the early-middle Miocene , when representatives of the Palaeothentidae and Abderitidae underwent a rapid radiation. A recent analysis of the paleoecology of 10 species of Patagonian paucituberculatans of Santacrucian age (late early Miocene) led Abello et al. (2012) to recognize a variety of frugivorous , insectivorous , and a combination of both feeding habits, a relative wide variety of body masses and curso-saltatorial , scansorial , and arboreal locomotor behavior.
Living microbiotherians are restricted to a single species belonging to the family Microbiotheriidae : the highly arboreal Dromiciops gliroides, restricted to the Valdivian forests of the Southern Andes (see Chap. 2; Martin 2010 ) . Similarly, most extinct microbiotherians inhabited in the southernmost portion of South America , and in the Antarctic Peninsula as well. They were taxonomically varied, with several representatives of the family Microbiotheriidae flourishing during the early Miocene (Abello et al. 2012, Chornogubsky and Kramarz 2012; Goin and Abello 2013). A less varied, older, larger-sized, and more generalized group is that of the Woodburnodontidae (Goin et al. 2007), from the early Eocene of Patagonia and the early-middle Eocene of Antarctica . Earlier claims (e.g., Muizon 1991) that Khasia cordillerensis, from the early Paleocene of Tiupampa (Bolivia ) is a microbiotherian, are not followed here (see Chap. 5; Woodburne et al. 2013: Table 1).
Microbiotherians have been subject of intensive research in the last three decades. Following initial studies by Szalay (1982) they are currently grouped in the Cohort Australidelphia, a clade that includes microbiotherians plus the four extant Australasian marsupial orders (Dasyuromorphia , Peramelemorphia , Notoryctemorphia , and Diprotodontia ). Numerous studies based on a variety of character systems have supported the monophyly of this clade, although the precise affiliation of the Microbiotheria within it has been matter of debate. Several authors (e.g., Amrine-Madsen et al. 2003) argued that microbiotherians are the sister-group of all Australasian marsupials , while others (e.g., Kirsch et al. 1997) supported a scheme where microbiotherians lie within that group.
The above-mentioned taxa do not exhaust the variety of lineages that inhabited South America during Cenozoic times (see Fig. 1.5). As “basal ameridelphians,” we refer here to a series of non-marsupial, non-sparassodontan metatherians that were especially abundant in the Late Cretaceous of North America as well as in the Paleogene of South America. The Pucadelphyidae , Jaskhadelphyidae , and Mayulestidae include several of the earliest Cenozoic metatherians, most of them recovered from Tiupampa (early Paleocene) of Bolivia (see Chap. 3). They were small to very small-sized, mostly of insectivorous habits, and already displayed several specialized features both skeletal and dental (Muizon and Céspedes in press). Several other, more specialized “basal ameridelphians” are known from the late Paleocene-early Eocene of Itaboraí , Brazil : Protodidelphidae , Derorhynchidae , Sternbergiidae (Oliveira 1998; Oliveira and Goin 2011).

Extinct South American diversity of metatherian mammals. Leonardo da Vinci’s “Vitruvian Man ” (1) serves as a parameter for size comparisons. Depicted are the living Didelphis albiventris (2), a female specimen with six pouch-young individuals attached to its back, and a hanging subadult ), and the extinct Bonapartherium hinakusijum (3), Minusculodelphis minimus (4), Derorhynchus singularis (5), Argyrolagus scagliai (6), Groeberia minoprioi (7), Patagonia peregrina (8), Abderites meridionalis (9), Caroloameghinia mater (10), Kramadolops maximus (11), Proborhyaena gigantea (12), and Thylacosmilus atrox (13). The phylogenetic affinities of Necrolestes patagonensis (14) are still unclear. Goin et al. (2007) regarded it as a Mammalia incertae sedis (for a more recent review, see Rougier et al. 2012). Drawings and composition by Manuel Sosa
Representatives of the Order Polydolopimorphia were quite abundant throughout the Paleogene Period, though much more restricted in the Neogene up to the Pliocene Epoch. Two major lineages (Suborders) compose this group: the Bonapartheriiformes on one hand, and the Polydolopiformes on the other. Representatives of this order were the first to develop a molar pattern of quadrangular shape, apt for the comminution of plant materials of a variety of shapes, sizes, and hardness. It has been suggested that the Polydolopimorphia were derived from the Microbiotheria (Goin et al. 2009a), thus their tentative inclusion among the Australidelphian marsupials in this book (Table 5.1). The Bonapartheriiformes include two major groups: The (earlier) Bonapartherioidea include the families Prepidolopidae , Rosendolopidae , Bonapartheriidae , and Gashterniidae . The (younger) Argyrolagoidea include the distinctly derived Groeberiidae , Patagoniidae , and Argyrolagidae . The Polydolopiformes include a stem lineage represented by Roberthoffstetteria nationalgeographica, plus the Polydolopidae , possibly the most highly derived metatherians ever developed in the New World . Three taxa from the Late Cretaceous of North America, Glasbius , Hatcheritherium , and Ectocentrocristus , have been mentioned as basal either to the whole order (the former two) or to the Polydolopiformes (Ectocentrocristus; but see Williamson et al. 2012; Case et al. 2005). This implies that stem polydolopimorphians were already developed in North America before their arrival in South America (Case et al. 2005). Actually, the oldest known South American metatherian, Cocatherium lefipanum from the earliest Paleocene of Patagonia, has been regarded as a member of this order (see above and Goin et al. 2006).
The Sparassodonta is another order of strictly South American metatherians that become extinct before the end of the Cenozoic . They occupied a wide variety of hypo-, meso-, and hypercarnivorous niches. Almost 60 species referable to this lineage have been described from Colombia , Brazil , Chile , Uruguay , Bolivia , and Argentina (Forasiepi 2009). The early Paleocene Allqokirus australis, from the early Danian (Puercan equivalent; see Gelfo et al. 2009) of Tiupampa , Bolivia, has been referred as a basal Sparassodonta (Muizon 1991). Thus, the Sparassodonta, together with the Polydolopimorphia and several basal “ameridelphians”, can be regarded as the oldest metatherians so far known in this continent.
Five families of Sparassodonta have been recognized up to now. The generalized Hondadelphidae are restricted to the South American tropics (Middle Miocene of Colombia , Marshall 1976). The Hathliacynidae range from Patagonia to Colombia and are mostly Neogene in age; they were relatively small carnivores, several of which had opportunistic feeding habits. The larger Borhyaenidae (up to the size of a modern puma) were also widespread in South America , ranging from the middle Eocene to the Pliocene . Proborhyaenids include the largest known New World metatherian so far known, Proborhyaena gigantea, with about 100 kg of body mass (Zimicz 2014). They are known from a few sites in southern and northern Argentina, and Bolivia , of late Eocene-Oligocene age. Finally, the extremely specialized, sabre-toothed Thylacosmilidae are known from Neogene sites of Argentina , Uruguay , and Colombia (Forasiepi 2009).
1.4 A Historical Perspective
The first metatherians known to Western eyes were, of course, not extinct but living. In 1500, the seafarer Vicente Yáñez Pinzón presented to the Catholic Monarchs three juvenile opossums (probably referable to some species of Didelphis ), as well as the skins of their mother and an additional juvenile, both dead during their trip from Brazil to Spain (Cabrera and Yepes 1960). Since then, many historians, chroniclers, explorers, and naturalists of the sixteenth, seventeenth, and eighteenth centuries mentioned these animals under a variety of names, many of which were derived from aboriginal names. The first Western graphic representation of an opossum is that of the Waldseemüller map of 1516, the “Carta Marina” (see, e.g., http://myloc.gov/exhibitions/earlyamericas/interactives/maps/html/cartamarina1516/view1516.html). There, a rather strange marsupial is depicted within a poorly recognizable South American contour, and near a region labeled as “Cannibal Land” (Terra Canibalor, present Brazil ). Other representations must have excited the imagination of many naturalists during these early centuries. An outstanding example is that of Albertus Seba’s “Cabinet of animal curiosities,” one of the most prized natural history books of all times (Seba 1734; Fig. 1.6). Carl Linnaeus’ 10th edition of the Systema Naturae already incorporates American opossums, all species known by then included in the genus Didelphis (Order Bestiae ; Linnaeus 1758). From the nineteenth century onward, an enormous corps of the literature has been devoted to the taxonomy, anatomy, natural history, physiology , ecology, and paleontology of New World marsupials (for the latter, see below).

Some of the oldest representations ever made of American marsupials by Western naturalists. Both drawings were included in Albertus Seba’s (1734–1765) Locupletissimi Rerum Naturalium Thesaurus. The upper figure is from Vol. 1 (1734), Plate XXXVI, while the lower one is from the same volume, Plate XXXVIII. Note the stress made by the artist on the pouch and pouch young of the depicted marsupials (most probably referable to an undetermined species of the four-eyed opossum Philander ). Images courtesy of the Missouri Botanical Garden, http://www.botanicus.org
Less known or studied are the numerous graphic, sculptural, and oral narrative references to American opossums among the native archaeological remains throughout the whole continent (López Austin 1990). Figure 1.7 shows three of them, coming from quite different archaeological context and cultures. From southern North America to central Argentina, opossums have played an important role in creation myths , legends , as well as symbols of specific meaning. In rural areas of northwestern Argentina , for instance, it is not infrequent to see an opossum skin (e.g., Didelphis albiventris) close to the beds of a woman in labor due to the small size of pouch young in all marsupial species, it is regarded as an omen of painless birth (Contreras 1983). Several myths throughout Central and South America give a fire-gifting role for the opossum , quite similar to that of Prometheus in Greek mythology (López Austin 1990). Finally, aboriginal medicine has usually a number of recipes that include parts of opossums (typically, the tail) for therapeutic purposes (Hartmann 1952).

a–c Schematic drawings of three opossum representations in archaeological objects belonging to different Central and South American cultures. a An opossum carved in a stone throne at Complejo Arqueológico Ventarrón-Collud , in northern Perú (see Alva Meneses 2008); b schematic drawing of a sculpture in stone of an opossum, belonging to the Tumaco-Tolita culture in the northern Andes , Ecuador (specimen Nr. LT-36-48-70, Museo Banco Central del Ecuador, Esmeraldas ; after a photograph in Ugalde 2006); c depiction of an opossum in a Mayan Code (Dresden Code ; after Gates 1932: Pl. 29). All drawings by Marcela Tomeo
The history of our scientific knowledge of extinct South American metatherians dates back to the last quarter of the nineteenth century. Paleontological discoveries in the Americas predate by far this date; however, most of these records were largely incidental, as well as related to other lineages of (usually very large) South American mammals: xenarthrans, SANU , proboscideans, etc. One of the earliest records of fossil mammals in this continent is attributed to a Jesuit priest, Father Guevara, who was probably the first one to infer the extinct condition of the “Carcarañá giants” (after Carcarañá River in Santa Fe Province, Argentina) : “…These giants, formidable towers of flesh, whose single name awakens the horrors and astonishment of peoples [and] attracts our attention, do not exist in present times; nevertheless, their old traces that from time to time are discovered by the Carcarañá and elsewhere, are evidence of their existence in past times” (Celaya 2005: 7; original in Spanish).
Following a “kuhnean ” approach (e.g., Kuhn 1970), Goin (1991) distinguished three periods in our knowledge of South American extinct metatherians. Each of them was characterized, and influenced, by distinct phylogenetic paradigms, available research technologies, and involved scholars. Each period included moments of theoretical effervescence followed by longer, mostly descriptive phases that were conceptually hegemonized by previously established paradigms. Finally, each period was influenced by the historic, political, cultural, and technological avatars of our contemporary history. Boundary dates for each of these periods, especially the latter one, are somewhat arbitrary, as it is hard to separate a process of successive knowledge achievements that were characterized, notably in South America , by their continuity.
1.4.1 First Period (1878–1930)
This period or cycle is intimately linked to the genius and figure of Florentino Ameghino (1854–1911), the most notable South American paleontologist of all times and Argentina’s father of science. Almost a half of all the genera currently recognized of South American extinct metatherians were described by Ameghino between 1888 and 1904. Several of his genial intuitions relative to the evolutionary history of our mammals still stand as the most reasonable evolutionary hypotheses. Additionally, we owe him the first biostratigraphic scheme for South America’s Cenozoic , mostly based on his brother Carlos’ observations and collections made in Patagonia , as well as in central Argentina, especially the Pampean region, made by both of them. He wrote about and discussed most Argentinian fossil localities, biostratigraphical units (his “étagés ”), and mammalian taxa known during his lifetime.
Ameghino studied fossils by the naked eye, or at the most with the help of a simple magnifying glass (Sbarra 1963); this is remarkable taking into account the tiny size of some of the fossils he described. A passionate man, he supported some bitter, even almost violent, discussions with contemporary naturalists (e.g., Germán Burmeister, Francisco Moreno, Alcides Mercerat), generally with more abundant and better arguments. He was the largest source of information of his time concerning South American extinct mammals, and the one that elaborated many of the soundest interpretative models regarding their chronologic succession, evolution, and phylogenetic relationships. His was an age of notable scientific production in many disciplines in many South American countries. In some of them, as in Argentina , the hiring of many foreign naturalists by official institutions brought an exponential increase in our knowledge of regional geography, natural resources, as well as the cultural and natural heritage, Paleontology among them. Florentino Ameghino stands as the founder of South America’s Vertebrate Paleontology.
Notably enough, Ameghino never regarded the Metatheria as a natural group. A few years before his death, he commented: “…[T]he division of mammals in two subclasses is a great mistake, as it raises impassable barriers that prevent us to recognize the close affinities among animals of such fundamentally identical structures as those of the dog and the thylacine ” (Ameghino 1910: 41; Spanish in the original).
To give an idea of the magnitude of his production, and just regarding South American, Neogene “opossum-like ” marsupials, Ameghino contributed more than 70 % of the publications of this period (e.g., Ameghino 1882, 1884, 1885, 1888, 1889, 1891a, b, c, 1893a, b, c, 1898, 1900, 1900-1903, 1902a, b, 1904, 1906, 1907, 1908, 1910). Other researchers involved in such studies were Burmeister (1879), Winge (1893), Mercerat (1898), Sinclair (1906), Rovereto (1914), Frenguelli (1921), Cabrera (1927, 1928).
1.4.2 Second Period (1930–1977)
Since the beginnings of the third decade of last century, South American paleomammalogy greatly benefited from the contributions of several North American scientists, most notably from one in particular: George Gaylord Simpson (1902–1984). One of the main figures of the Synthetic Theory of Evolution, Simpson gave light to an impressive scientific production, both empirical and theoretical (see, e.g., Gingerich 1986). Due to several field trips to Patagonia, Simpson got involved in the study of South American extinct mammals; as a consequence, he was able to discuss many of Ameghino’s previous ideas. Overall, his thoughts on South American mammalian taxonomy, evolution, succession, biochronology, and turnovers, turned out to constitute a new paradigm. Particularly important are his contributions on the early establishment of mammalian lineages in this continent, as well as the concepts of isolation and of successive, distinct taxonomic associations (his “mammal strata”) throughout Cenozoic times (e.g., Simpson 1948, 1950, 1967). Simpson was also active in the description, allocation, and phylogenetic speculation of several major lineages of South American metatherians, most notably polydolopid polydolopimorphians and didelphoid didelphimorphians (for the latter see Goin 1991). Also relevant as major frameworks on mammalian evolution in South America, stand the contributions by Patterson and Pascual (1968, 1972).
This period also saw major improvements in the design of a biochronological scheme for the continental Cenozoic of South America . Efforts were led by Rosendo Pascual and his team at the Museo de La Plata (Pascual et al. 1965, 1966; Pascual and Odreman Rivas 1971, 1973), who refined and completed previows work by Ameghino (e.g., 1906), Kraglievich (1952). Our knowledge of Paleogene metatherians was notably expanded with the discovery and description, by Carlos de Paula Couto, of the early Eocene assemblage of Itaboraí , in southeastern Brazil , largely dominated by stem “ameridelphians” (Paula Couto 1952a, b, c, 1960, 1962, 1970; see a review in Oliveira 1998). Another interesting fauna described in this period is that of the ?late Paleocene of Laguna Umayo in southern Perú (Sigé 1971, 1972; see Sigé et al. 2004 for a reassessment of its age). By the end of this period and the beginning of the third one, the late Oligocene (Deseadan SALMA ) Salla-Luribay (Bolivia ) assemblages were first discovered and described (Hoffstetter 1968, 1969, 1976; Hoffstetter and Lavocat 1970; Hoffstetter et al. 1971; for a preliminary review on the metatherian taxa, see a review in Villarroel and Marshall 1982). Also, the extraordinary mammalian fauna of the middle Miocene of La Venta , also rich in metatherians, began to be described (e.g., Marshall 1976, 1977).
From a technological point of view, this second period was characterized by the intensive use of now standard optical instruments, as the stereomicroscope, stereoscopic photography, or the micrometric ocular and calipers, for the observation and measurement of the cranial, postcranial, and dental structures of mammals. This prompted a huge advance in our knowledge of the diversity and evolution of many extinct lineages of small-sized mammals, including those of metatherians. Field methods still kept on the long tradition of naked-eye prospecting.
Taking again the example of Neogene “opossum-like ” marsupials, there is a quite balanced production among local and foreign researchers: Rusconi (1932), Simpson (1932a, b, 1935a, b, 1938, 1948, 1972, 1974), Kraglievich (1934), Patterson (1937), Riggs and Patterson (1939), Ringuelet (1953, 1966), Reig (1952, 1955a, b, 1957a, b, 1958a, b, c), Hoffstetter (1963), Zetti (1967), Reig and Simpson (1972), Marshall (1976, 1977), Del Corro (1977).
1.4.3 Third Period (1977–Present)
Even though somewhat arbitrary, the date chosen as the beginning of this third period is coincidental with the appearance, within a short period of five years, of a series of highly influential works by two North American researchers. The first one was that of John A.W. Kirsch: “The comparative serology of Marsupialia, and a Classification of Marsupials” (Kirsch 1977). In it, the author introduced a new methodology in the analysis of living metatherians (he compared around a hundred species), and tested his results with those of more traditional, morphological taxonomy. Even though he accepted, with slight modifications, Ride’s (1964) multiordinal scheme for the arrangement of marsupials, he recognized the uniqueness of microbiotheriids among the South American metatherians. We owe the second major contribution to Frederick Szalay, who a few years later (e.g., Szalay 1982; see also Szalay 1994) would promote a drastic re-arrangement of marsupial affinities based on their tarsal morphology. He recognized the Australidelphia , including microbiotherians and all Australasian lineages, as a natural group.
Regarding South American mammalian evolution as a whole, the theoretical framework during this period consisted of a series of attempts to relate the different phases in mammalian evolution with the biotic (floral and ecosystemic dynamics) and abiotic (tectonic, temperatures, and rainfall) factors that framed it. Again, a leading figure in this field was the late Pascual (1984, 1996, 2006; Pascual and Ortiz-Jaureguizar 1990, 1991, 2007; Pascual et al. 1996; other contributions include those of Simpson 1980; Flynn et al. 2003; Goin et al. 2012; Woodburne et al. 2013; among others). Several of these studies were triggered as a consequence of improvements in the geochronological calibration of the Cenozoic succession (e.g., Marshall and Pascual 1978; Marshall and Patterson 1981; Marshall et al. 1977, 1981, 1983a, b; Flynn and Swisher 1995; Kay et al. 1999; Flynn and Wyss 1999; Flynn et al. 2003; Tejedor et al. 2009; Ré et al. 2010; Madden et al. 2010). Preliminary attempts to specifically relate metatherian evolutionary phases with global climates were presented by Goin et al. (2010, in press).
This period can be characterized, in technological terms, by the use of a series of tools that greatly improved our capacity for anatomical and taxonomical analysis: the use of plastic casts, especially of dental specimens, for comparative purposes; the application of scanning electron microscopy (SEM) , computed tomography scan (CT-scan ), digital image managing, etc., in morphological analyses; the use of a variety of cladistic methodologies in the phylogenetic analysis; finally, information technologies hugely expanded the publishing and sharing of scientific knowledge. In the field, various methodologies have proven to be successful in fossil prospecting; regarding small vertebrates, as is the case of most metatherians, the use of dry-screening , or underwater screen-washing of fossil-bearing sediments has exponentially enlarged collections from a number of localities.
Regarding our knowledge of the evolutionary history of South American metatherians, this period is characterized by an exponential growth in available information on both extinct and extant local faunas and isolated taxa, and by the numerous proposals of alternate (and frequently antagonistic) phylogenetic schemes (for the latter, see Chap. 5). Besides numerous works describing isolated metatherian taxa, notable are the discovery of many new faunal assemblages that radically enhanced our knowledge of mammalian evolution in this continent. Among the mammalian associations discovered and/or described during this period, with a significant metatherian content, prominent are those of the early Paleocene of Tiupampa in Bolivia (Marshall et al. 1983a, 1985; Marshall and Muizon 1988; Muizon 1991, 1994, 1998; Muizon and Cifelli 2001), “medial” Paleocene of Punta Peligro in southern Argentina (e.g., Bonaparte et al. 1993; Bond et al. 1995; Gelfo et al. 2009), early to middle Eocene of Paso del Sapo in southern Argentina (Tejedor et al. 2009), ?middle Eocene of the La Meseta Fm in northernmost Antarctic Peninsula (Woodburne and Zinsmeister 1982, 1984; Goin et al. 1999; Chornogubsky et al. 2009), middle-late Eocene of Northwestern Argentina (Pascual 1980a, b, 1981, 1983; Pascual et al. 1981; Goin et al. 1986); middle to late Eocene of Contamana in northeastern Perú (Antoine et al. 2012); ?late Eocene of Santa Rosa in eastern Perú (Campbell 2004; Goin and Candela 2004), latest Eocene-earliest Oligocene of Tinguiririca in central Chile (Wyss et al. 1990, 1994; Flynn et al. 2003; Croft et al. 2008), early Oligocene faunas of La Cancha and La Cantera in southern Argentina (Goin et al. 2010); reviews of the early Miocene, Colhuehuapian SALMA metatherians from Patagonia (Goin et al. 2007; Goin and Abello 2013) and of middle Miocene of La Venta (Colombia ) metatherians (Goin 1997a); late Miocene of Cerro Azul Fm and Caleufú in Central Argentina (Goin et al. 2000; Abello et al. 2002). Among post-tertiary faunas discovered and described in this period, can be mentioned the interesting, early to late Holocene successions, including many marsupial specimens, of Tixi Cave in southeastern Pampas , Argentina (Goin 2001) and of several sites in southeastern Brazil (Hadler et al. 2009a, b; Oliveira et al. 2011).
This was also a period of several major reviews of metatherian lineages. Beginning in 1976, Larry G. Marshall would accomplish a remarkable series of reviews of most South American groups known up to then (Marshall 1976b, 1977, 1979, 1980, 1981a, b, 1982a, b, 1987; Marshall et al. 1990). More recent reviews, including the addition of newly described taxa, include those of Goin (1991), Sánchez-Villagra et al. (2000; see also Sánchez-Villagra 2001; Sánchez-Villagra and Kay 1997), Forasiepi (2007), Abello (2007, 2012, 2013), Chornogubsky (2010), Chornogubsky et al. (2009), Goin et al. (2009a, b).
Finally, and taking again the case of Neogene opossum -like marsupials, works in this period show a consolidated production by South American researchers (e.g., Marshall 1976; Villarroel and Marshall 1983; Prado et al. 1985; Goin 1997b, c, 2001; Goin and De Los Reyes 2012; Goin and Montalvo 1989; Goin and Pardiñas 1996; Goin et al. 2009b; Cozzuol et al. 2006; Forasiepi et al. 2009; Oliveira et al. 2011; Abello et al. in press).
1.5 The Incompleteness of the Fossil Record
A final note should be made on the biased, incomplete fossil record of South American metatherians—and of other terrestrial vertebrates as well. Figure 1.8 shows a chart of the first half of the Cenozoic Era, relating periods, epochs, SALMAs , and known extinct South American and Antarctic terrestrial faunas. The latter have been related to their latitude. As can be appreciated, the best known areas are those of mid- to high latitudes up to 50°N. South and north of them, the fossil record is almost a black hole. This lack of records is particularly relevant for metatherians, as they are usually distributed in tropical -subtropical areas of the continent. As argued in Chaps. 4 and 7, metatherian radiations seem to have been triggered by phases of global warming (i.e., the climatic optima) not only in Cenozoic times but also during the Cretaceous .

“Black holes” in our knowledge of metatherian faunas in South America. The time span considered here is the first half of the Cenozoic (i.e., Paleocene-Eocene). Areas in black refer to the still unsampled regions of South America for this time span. See Chap. 4 for details on South America’s biogeographic regions and kingdoms
The best known tropical, extinct metatherian assemblage is, up to now, that of the middle Miocene of La Venta in Colombia (see Marshall 1976a; Goin 1997a). Among the thylacosmilid sparassodonts , which are well represented in specimens and taxa, noteworthy is the persistence of several taxa which are generalized with respect to even earlier assemblages in higher latitudes (e.g., Patagonia). The earliest record of a thylacosmilid sparassodont is that of an isolated upper molar recovered from early Miocene levels of Colhuehuapian age at Gran Barranca, in southern Chubut Province, Patagonia (Goin et al. 2007). Even though it comes from levels at least five million years older than those of La Venta, it already shows morphological specializations as advanced, or even more so, than those of the tropical, middle Miocene Anachlysictis gracilis—comparisons have to be made on indirect grounds, as the latter was based on a lower jaw. Moreover, an additional specimen from La Venta, originally referred by Goin (1997a, p. 203) to a “Thylacosmilidae ?”, could represent the most generalized thylacosmilid so far known. The persistence of generalized types in the tropics, as is the case of the La Venta thylacosmilids, suggests that the lack of records from tropical areas in South America still precludes our better understanding of metatherian’s phylogenetic relationships. Many “missing links” are still hiding in the tropics, maybe for ever.
References
Abello MA (2007) Sistemática y bioestratigrafía de los Paucituberculata (Mammalia, Marsupialia) del Cenozoico de América del Sur. Unpubl. thesis, Univ Nacional de La Plata
Abello MA (2013) Analysis of dental homologies and phylogeny of Paucituberculata (Mammalia: Marsupialia). Biol J Linn Soc 109:441–465
Abello MA, Montalvo CI, Goin FJ (2002) Marsupiales del Mioceno Superior de Caleufú (La Pampa, Argentina). Ameghiniana 39(4):433–442
Abello MA, Ortiz-Jaureguizar EO, Candela AM (2012) Paleoecology of the Paucituberculata and Microbiotheria (Mammalia, Marsupialia) from the late Early Miocene of Patagonia. In: Vizcaino SF, Kay RF, Bargo MS (eds) Early Miocene Paleobiology in Patagonia. High-latitude paleocommunities of the Santa Cruz Formation. Cambridge University Press, Cambridge
Abello MA, De los Reyes M, Voglino C, Candela AM, Coira B, Galli CI (in press) Primer registro de Marsupialia (Mammalia, Metatheria) para la Formación Maimará (Mioceno Tardío), Quebrada de Humahuaca (Provincia de Jujuy, Argentina). Ameghiniana
Alva Meneses I (2008) Los complejos de Cerro Ventarrón y Collud-Zarpán: del Precerámico al Formativo en el valle de Lambayeque. Bol Arqueol, Pontif Univ Catól Perú 12:97–117
Ameghino F (1882) Catálogo explicativo de las colecciones de antropología prehistórica y de Paleontología, Florentino Ameghino. In: Catálogo de la sección de la Provincia de Buenos Aires en la Exposición Continental Sudamérica. Anexo A, Buenos Aires
Ameghino F (1884) Excursiones geológicas y paleontológicas en la Provincia de Buenos Aires. Bol Acad Nac Cs Córdoba 6:161–257
Ameghino F (1885) Informe sobre el Museo Antropológico y Paleontológico de la Universidad Nacional de Córdoba durante el año 1885. Bol Acad Nac. Cs. Córdoba 8:347–360
Ameghino F (1888) Lista de las especies de mamíferos fósiles del Mioceno Superior de Monte Hermoso hasta ahora conocidos. PE Coni, Buenos Aires
Ameghino F (1889) Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas Acad Nac Cs Córdoba, Vol 4, Buens Aires
Ameghino F (1891a) Las antiguas conexiones del continente sudamericano y la fauna eocena argentina. Rev Arg Hist Nat 1:123–216
Ameghino F (1891b) Burmeister: adiciones al examen crítico de los mamíferos fósiles tratados en el “Examen crítico de los mamíferos y reptiles fósiles, etc”. por A. Bravard. Rev Arg Hist Nat 1:259–290
Ameghino F (1891c) Sobre la supuesta presencia de creodontes en el Mioceno Superior de Monte Hermoso. Rev Arg Hist Nat 1:437
Ameghino F (1893a) Les mammifères fossiles de La Patagonie Australe. Rev Sci 51:13–17
Ameghino F (1893b) New discoveries of a fosil Mammalia of Southern Patagonia. Amer Nat 27:445
Ameghino F (1893c) Les premieres mammiferes. Relation entre les mammiféres diprotodontes eocenes de l´Amerique du Nord et deux de la Republique Argentine. Rev Gen Sci Pures et Appliq 4(3):77
Ameghino F (1898) Sinopsis geológico-paleontológica de la República Argentina. In: Segundo Censo de la República Argentina, Vol 1, La Plata
Ameghino F (1900) Presencia de mamíferos diprotodontes en los depósitos terciarios de Panamá. Anal Soc Cient Arg 49:235
Ameghino F (1900–1903) L´age des formations sedimentaires de Patagonie. Anal Soc Cient Arg 50:109–130
Ameghino F (1902a) Línea filogenética de los Proboscídios. Anal Mus Nac Buenos Aires, Ser 3(1):1–2
Ameghino F (1902b) Premiere contribution a laconnaissance de la faune mammalogique des couches a Colpodon. Bol Acad Nac Cs Córdoba 17:71–140
Ameghino F (1904) Nuevas especies de mamíferos cretáceos y terciarios de la República Argentina. Anal Soc Cient Arg 57:225–291
Ameghino F (1906) Les formations sedimentaires du Crétacé Supèrieur et du Tertiaire de Patagonie, avec un paralieleentre leurs faunes mammalogiques et celles de l´ancien continente. Anal Mus Nac Hist Nat Buenos Aires 3(8):1–568
Ameghino F (1907) Nota sobre una pequeña colección de huesos de mamíferos, procedente de las grutas calcáreas de Iporanga (Sao Paulo, Brazil). Rev. Mus Paulista 7:59–124
Ameghino F (1908) Las formaciones sedimentarias de la Región litoral de Mar del Plata y Chapadmalal. Anal Mus Nac Hist Nat Buenos Aires 3(10):43–428
Ameghino F (1910) Paleontología Argentina: relaciones filogenéticas y geográficas. Anal Inst Enseñ Gen 1:11–84
Amrine-Madsen H, Scally M, Westerman M, Stanhope MJ, Krajewski C, Springer MS (2003) Nuclear gene sequence provides evidence for the monophyly of Australidelphian marsupials. Mol Phylogenet Evol 28:186–196
Antoine P-O, Marivaux L, Croft DA, Billet G, Ganerød M, Jaramillo C, Martin T, Orliac MJ, Tejada J, Altamirano AJ, Duranthon F, Fanjat G, Rousse S, Salas Gismondi R (2012) Middle Eocene rodents from Peruvian Amazonia reveal the pattern and timing of caviomorph origins and biogeography. Proc R Soc B 279(1732):1319–1326
Averianov AO, Kielan-Jaworowska Z (1999) Marsupials from the Late Cretaceous of Uzbekistan. Acta Palaeontol Pol 44:71–81
Bonaparte JF, Van Valen LM, Kramartz A (1993) La fauna local de Punta Peligro, Paleoceno Inferior, de la Provincia del Chubut, Patagonia, Argentina. Evol Monogr 14:1–61
Bond M, Carlini AA, Goin FJ, Legarreta L, Ortiz-Jaureguizar E, Pascual R, Uliana MA (1995) Episodes in South American land mammal evolution and sedimentation: testing their apparent concomitance in a Palaeocene succession from Central Patagonia. VI Congr Arg Paleont Bioestrat Trelew, Actas, pp 47–58
Brown B (2004) Atlas of New World Marsupials. Fieldiana Zool (NS) 102:1–308
Burmeister G (1879) Description Physique de la Republique Argentine. II Animaux Vertebres, Premiere partie. PE Coni, Buenos Aires
Cabrera A (1927) Datos para el conocimiento de los Dasiuroideos fósiles argentinos. Rev Mus La Plata 30:271–315
Cabrera A (1928) Un nuevo marsupial Plioceno de Monte Hermoso. Rev Mus La Plata 31:335–337
Cabrera A, Yepes J (1960) Mamíferos Sudamericanos, vol 1. Ediar, Buenos Aires
Campbell KE Jr (ed) (2004) The Paleogene mammalian fauna of Santa Rosa, Amazonian Perú. Nat Hist Mus Los Angeles County, Sci Ser 40, Los Angeles
Case JA, Goin FJ, Woodburne MO (2005) “South American” marsupials from the Late Cretaceous of North America and the origin of marsupial cohorts. J Mammal Evol 12:461–494
Celaya RA (2005) Orígenes e inicios de la localidad de Aldao (Depto. San Lorenzo). Congreso Argentino de Inmigración y IV Congreso de la Historia de los Pueblos de la Provincia de Santa Fe. Asoc Amigos Arch Gen Prov, Esperanza, Santa Fe. Unpubl. https://www.google.com.ar/#bav=on.2,or.r_qf.&fp=ef598c4af7830a47&q=+Ricardo+Alberto+Celaya.
Chornogubsky L (2010) Sistemática de la familia Polydolopidae (Mammalia, Marsupialia, Polydolopimorphia) de América del Sur y la Antártica. Unpubl thesis, Univ Buenos Aires
Chornogubsky L, Kramarz AG (2012) Nuevos hallazgos de Microbiotheriidae (Mammalia, Marsupialia) en la Formación Pinturas (Mioceno temprano, Argentina). Ameghiniana 49(4):442–450
Chornogubsky L, Goin FJ, Reguero M (2009) A reassessment of Antarctic polydolopid marsupials (Middle Eocene, La Meseta Formation). Antarct Sci 21(3):285–297
Cifelli RL, Davis BM (2003) Marsupial origins. Science 302:1899–1900
Clyde WC, Wilf P, Iglesias A, Slingerland RL, Barnum T, Bijl PK, Bralower TJ, Brinkhuis H, Comer EE, Huber BT, Ibañez-Mejia M, Jicha BR, Krause JM, Schueth JD, Singer BS, Raigemborn MS, Schmitz MD, Sluijs A, Zamaloa MC (2014) New age constraints for the Salamanca formation and lower Río Chico Group in the western San Jorge Basin, Patagonia, Argentina: implications for K/Pg extinction recovery and land mammal age correlations. Bull Geol Soc Amer
Contreras JR (1983) La comadreja overa. In: Fauna Argentina, Publ 11. Centro Editor de América Latina, Buenos Aires
Cozzuol MA, Goin FJ, de los Reyes M, Rancy A (2006) The oldest species of Didelphis (Mammalia, Marsupialia, Didelphidae), from the Late Miocene of Amazonia. J Mammal 87(4):663–667
Croft DA, Flynn JJ, Wyss AR (2008) The Tinguiririca fauna of Chile and the early stages of “modernization” of South American mammal faunas. Arq Mus Nac Rio de Janeiro 66(1):1–21
de Muizon C (1991) La fauna de mamíferos de Tiupampa (Paleoceno Inferior, Formación Santa Lucía) Bolivia. In: Suarez-Soruco R (ed) Fósiles y Facies de Bolivia, vol 1, Vertebrados. Rev Técn Yac Petrolíferos Fiscales de Bolivia 12:575–624
de Muizon C (1994) A new carnivorous marsupial from the Palaeocene of Bolivia and the problem of marsupial monophyly. Nature 370:208–211
de Muizon C (1998) Mayulestes ferox, a borhyaenoid (Metatheria, Mammalia) from the early Palaeocene of Bolivia. Phylogenetic and palaeobiologic implications. Géodiversitas 20:19–142
de Muizon C, Céspedes R (in press) The beginning of the age of therian mammals in South America: Tiupampa, a transition between Northern and Southern worlds in the basal Paleocene. In: Rosenberger AL, Tejedor MF (eds) Origins and evolution of Cenozoic South American mammals. Springer, New York
de Muizon C, Cifelli RL (2001) A new basal “didelphoid” (Marsupialia, Mammalia) from the early Paleocene of Tiupampa (Bolivia). J Vert Paleontol 21:87–97
de Paula Couto C (1952a) Fossil mammals from the begining of the Cenozoic in Brazil. Marsupialia: Polydolopidae and Borhyaenidae. Amer Mus Novitates 1559:1–27
de Paula Couto C (1952b) Fossil mammals from the beginning of the Cenozoic in Brazil. Marsupialia: Didelphidae. Amer Mus Novitates 1567:1–26
de Paula Couto C (1952c) A new name for Mirandaia ribeiroi Paula Couto, 1952. J Mamm 33:503
de Paula Couto C (1960) Marsupiais fósseis do Paleoceno do Brasil. An Acad Bras Cs 33(3–4):321–333
de Paula Couto C (1962) Didelfídeos fósiles del Paleoceno de Brasil. Rev Mus Arg Cs Nat “B. Rivadavia” 112:135–66
de Paula Couto C (1970) News on the fossil marsupials from the Riochican of Brazil. An Acad Bras Cs 42(1):19–34
Del Corro J (1977) Un nuevo microbioterio (Marsupialia) del Eoceno de Patagonia. Rev Mus Arg Cs Nat “B. Rivadavia” 2(2):31–33
Dunn RE, Madden RH, Kohn MJ, Schmitz MD, Strömberg CAE, Carlini AA, Ré GH, Crowley J (2013) A new chronology for middle Eocene–early Miocene South American Land Mammal Ages. GSA Bulletin 125(3/4):539–555
Flynn JJ, Swisher CC III (1995) Cenozoic South American land mammal ages: correlation to global geochronologies. In: Berggren WA, Kent DV, Aubry M-P, Hardenbol J (eds) Geochronology, time-scales and global stratigraphic correlation: a unified framework for an historical geology. Soc Strat Geol Spec Pub 54:317–333
Flynn JJ, Wyss AR (1999) New marsupials from the Eocene-Oligocene transition of the Andean Main Range, Chile. J Vert Paleontol 19:533–549
Flynn JJ, Wyss AR, Croft DA, Charrier R (2003) The Tinguiririca Fauna, Chile: biochronology, paleoecology, biogeography, and a new earliest Oligocene South American Land Mammal Age. Palaeogeogr Palaeoclim Palaeoecol 195:229–259
Forasiepi AM (2009) Osteology of Arctodictis sinclairi (Mammalia, Metatheria, Sparassodonta) and phylogeny of Cenozoic metatherian carnivores from South America. Monogr Mus Arg Cs Nat “B Rivadavia” NS 6:1–174
Forasiepi AM, Goin FJ, Martinelli AG (2009) Contribution to the knowledge of the Sparassocynidae (Mammalia, Metatheria, Didelphoidea), with comments on the age of the Aisol Formation (Neogene), Mendoza Province, Argentina. J Vert Paleont 29(4):1–12
Frenguelli J (1921) Los terrenos de la costa atlántica en los alrededores de Miramar (Pcia. de Buenos Aires). Bol Acad Nac Cs Córdoba 24:325–485
Gates W (1932) The Dresden Codex. Maya Society Publication 2, J Hopkins Univ, Baltimore
Gelfo JN, Goin FJ, Woodburne MO, de Muizon C (2009) Biochronological relationships of the earliest South American Paleogene mammalian faunas. Palaeontology 52:251–269
Gingerich PD (1986) George Gaylord Simpson: empirical theoretician. Contrib Geol Sp Paper 3:3–9
Goin FJ (1991) Los Didelphoidea (Mammalia, Marsupialia, Didelphimorphia) del Cenozoico tardío de la Región Pampeana. Unpubl thesis, Univ Nac La Plata
Goin FJ (1997a) New clues for understanding Neogene Marsupial radiations. In: Kay RF, Madden RH, Cifelli RL, Flynn JJ (eds) A history of the neotropical fauna. Vertebrate Paleobiology of the Miocene in Colombia. Smithsonian Institute Press, Washington
Goin FJ (1997b) Thylamys zettii, nueva especie de marmosino (Marsupialia, Didelphidae) del Cenozoico tardío de la Región Pampeana. Ameghiniana 34(4):481–484
Goin FJ (1997c) Sobre la edad y afinidades de Zygolestes paranensis Ameghino, 1898 (Marsupialia, Didelphidae, Marmosinae). Neotropica 43(109–110):15–19
Goin FJ (2001) Marsupiales (Didelphidae: Marmosinae & Didelphinae). In: Mazzanti D, Quintana C (eds) Cueva Tixi: Cazadores y recolectores de las Sierras de Tandilia Oriental. 1: Geología, Paleontología y Zooarqueología. Lab Arqueol Univ Nac Mar del Plata, Publ Esp 1, Mar del Plata
Goin FJ (2006) A review of the Caroloameghiniidae, Paleogene South American “primate-like” marsupials (?Didelphimorphia, Peradectoidea). In: Kalthoff D, Martin T, Möors T (eds) Festband für Herrn Professor Wighart v. Koenigswald anlässlich seines 65. Geburtstages. Palaeontographica Abt. A 278. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart
Goin FJ, Abello MA (2013) Los Metatheria sudamericanos de comienzos del Neógeno (Mioceno temprano, edad mamífero Colhuehuapense): Microbiotheria y Polydolopimorphia. Ameghiniana 50(1):51–78
Goin FJ, Candela AM (2004) New Paleogene marsupials from the Amazonian basin, Southeastern Perú. In: Campbell KE Jr (ed) The Paleogene mammalian fauna of Santa Rosa, Amazonian Perú. Nat Hist Mus Los Angeles County, Sci Ser 40, Los Angeles
Goin FJ, de los Reyes M (2012) Contribución al conocimiento de los representantes extintos de Lutreolina Thomas, 1910 (Mammalia, Marsupialia, Didelphidae). Rev Hist Natural Fund Félix de Azara 1(2):15–25
Goin FJ, Montalvo C (1989) Revisión sistemática y reconocimiento de una nueva especie del género Thylatheridium Reig (Marsupialia, Didelphidae). Ameghiniana 25(2):161–167
Goin FJ, Pardiñas U (1996) Revisión de las especies del género Hyperdidelphys Ameghino, 1904 (Mammalia, Marsupialia, Didelphidae). Su significación filogenética, estratigráfica y adaptativa en el Neógeno del Cono Sur Sudamericano. Estud Geol 52:327–359
Goin FJ, Palma RM, Pascual R, Powell JE (1986) Persistencia de un primitivo Borhyeanidae (Mammalia, Marsupialia) en el Eoceno temprano de Salta (Formación Lumbrera, Argentina). Aspectos geológicos y paleoambientales relacionados. Ameghiniana 23(1–2):47–56
Goin FJ, Woodburne MO, Case JA, Vizcaíno SF, Reguero M (1999) New discoveries of “opossum-like” marsupials from Antarctica (Seymour Island, Middle Eocene). J Mamm Evol 6(4):335–365
Goin FJ, Montalvo C, Visconti G (2000) Los Marsupiales (Mammalia) del Mioceno Superior de la Formación Cerro Azul (Provincia de La Pampa, Argentina). Rev Esp Geol 56:101–126
Goin FJ, Pascual R, Tejedor MF, Gelfo JN, Woodburne MO, Case JA, Reguero MA, Bond M, Cione AL, Udrizar Sauthier D, Balarino L, Scasso RA, Medina FA, Ubaldón MC (2006) The earliest Tertiary therian mammal from South America. J Vert Paleontol 26:505–510
Goin FJ, Zimicz AN, Reguero MA, Santillana S, Marenssi SA, Moly JJ (2007) New mammal from the Eocene of Antarctica, and the origins of the Microbiotheria. Rev Asoc Geol Arg 62:597–603
Goin FJ, Candela AM, Abello MA, Oliveira EV (2009a) Earliest South American Paucituberculatans and their significance in the understanding of “pseudodiprotodont” marsupial radiations. Zool J Linn Soc 155:867–884
Goin FJ, Zimicz N, de los Reyes M, Soibelzon L (2009b) A new large didelphid of the genus Thylophorops (Mammalia: Didelphimorphia: Didelphidae) from the Late Tertiary of the Pampean Region (Argentina). Zootaxa 2005:35–46
Goin FJ, Abello MA, Chornogubsky L (2010) Middle tertiary marsupials from central Patagonia (early Oligocene of Gran Barranca): understanding South America’s Grande Coupure. In: Madden RH, Carlini AA, Vucetich MG, Kay RF (eds) The Paleontology of Gran Barranca: evolution and environmental change through the Middle Cenozoic of Patagonia. Cambridge University Press, New York
Goin FJ, Gelfo JN, Chornogubsky L, Woodburne MO, Martin T (2012) Origins, radiations, and distribution of South American mammals: from greenhouse to icehouse worlds. In: Patterson BD, Costa LP (eds) Bones, clones, and biomes: an 80-million year history of Recent Neotropical mammals. The University of Chicago Press, Chicago
Goin FJ, Zimicz AN, Forasiepi AM, Chornogubsky LC, Abello MA (in press) The rise and fall of South American Metatherians: Contexts, Adaptations, Radiations, and Extinctions. In: Rosenberger AL, Tejedor MF (eds) Origins and evolution of Cenozoic South American Mammals. Springer, Dordrecht
Gradstein FM, Ogg JG, Schmitz MD, Ogg GM (2012) The geologic time scale 2012 (vols 1, 2). Elsevier, Amsterdam
Hadler P, Ferigolo J, Goin FJ (2009a) Mamíferos de pequeno porte (Didelphimorphia, Chiroptera e Rodentia) do Pleistoceno final/Holoceno do Brasil, con énfase o Rio Grande do Sul. In: Ribeiro AM, Bauermann SG, Saldanha Scherer S (eds) Quaternario do Rio Grande do Sul. Integrando conhecimentos. Monograf Soci Brasileira Paleontol Porto Alegre
Hadler P, Goin FJ, Ferigolo J, Ribeiro AM (2009b) Environmental change and marsupial assemblages in Holocene successions of Southern Brazil. Mamm Biol 74:87–99
Hartman C (1952) Possums. University of Texas Press, Austin
Hoffstetter R (1963) La faune Pleistocene de Tarija (Bolivia). Note prelimimaire. Bull Mus Nat d’Hist Naturelle 33(2):152–160
Hoffstetter R (1968) Un gisement de mammiferes deseadiens (Oligocene inferieur) en Bolivie. Compt Rendus Acad Sciences 267:1095–1097
Hoffstetter R (1969) Un primate de l’Oligocene interieur sud-american: Branisella boliviana gen. et sp. nov. Compt Rendus Acad Sciences 269:434–437
Hoffstetter R (1976) Rongeurs caviomorphes de l’Oligocene de Bolivie: Palaeovertebrata 7:1–14
Hoffstetter R, Lavocat R (1970) Decouverte dans le Deseadien de Bolivie des genres pentalophodontes appuyant les affinities africaines des rongeurs Caviomorphes. Compt Rendus Acad Sci 271:172–175
Hoffstetter R, Martínez R, Matteur M, Tomasi P (1971) Lacayani, un nouveau gisement bolivien de mammiferes deseadiens (Oligocene inferieur). Compt Rendus Acad Sciences 273:2215–2218
Horovitz I, Sánchez-Villagra MR (2003) A morphological analysis of marsupial mammal higher-level phylogenetic relationships. Cladistics 19:181–212
Horovitz T, Bloch MJ, Ladevèze S, Kurz C, Sánchez-Villagra MR (2009) Cranial anatomy of the earliest marsupials and the origin of opossums. PLoS ONE 4(12):e8278. doi:10.1371/journal.pone.0008278
Kay RF, Madden RH, Vucetich MG, Carlini AA, Mazzoni MM, Re GH, Heizler M, Sandeman H (1999) Revised age of the Casamayoran South American Land Mammal ‘Age’ -climatic and biotic implications. Proc Nat Acad Sci 96(23):13235–13240
Kielan-Jaworowska Z (1975) Possible occurrence of marsupial bones in Cretaceous Eutherian mammals. Nature 255:698–699
Kielan-Jaworowska Z, Cifelli RL, Luo ZX (2004) Mammals from the age of dinosaurs. Origin, evolution, and structure. Columbia University Press, New York
Kirsch JAW (1977) The comparative serology of Marsupialia, and a classification of marsupials. Australian J Zool Suppl Ser 52:1–152
Kirsch JAW, Lapointe FJ, Springer MS (1997) DNA-hybridisation studies of marsupials and their implications for metatherian classification. Australian J Zool 45:211–280
Kraglievich JL (1934) La antigüedad pliocena de las faunas de Monte Hermoso y Chapadmalal, deducidas de su comparación con las que le precedieron y sucedieron. El Siglo Ilustrado, Buenos Aires
Kraglievich JL (1952) El perfil geológico de Chapadmalal y Miramar, provincia de Buenos Aires. Rev Mus Municip Cs Nat Tradicional Mar del Plata 1:8–37
Kuhn TS (1970) The structure of scientific revolutions, 2nd edn. The University of Chicago Press, Chicago (Enlarged)
Linnaeus C (1758) Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis, 10 edn, vol 1, Holmiae. Salvius, Stockholm
López Austin A (1990) The myths of the Opossum. Pathways of Mesoamerican mythology. University of New Mexico Press, Albuquerque
Luckett WP (1993) An ontogenetic assessment of dental homologies in therian mammals. In: Szalay FS, Novacek MJ, McKenna MC (eds) Mammal phylogeny: mesozoic differentiation, multituberculates, monotremes, early therians, and marsupials. Springer, New York
Luo ZX, Ji Q, Wible JR, Yuan CX (2003) An Early Cretaceous tribosphenic mammal and metatherian evolution. Science 302:1934–1940
Madden RH, Kay RF, Vucetich MG, Carlini AA (2010) Gran Barranca: a 23 million-year record of middle Cenozoic faunal evolution in Patagonia. In: Madden RH, Carlini AA, Vucetich MG, Kay RF (eds) The Paleontology of Gran Barranca: evolution and environmental change through the Middle Cenozoic of Patagonia. Cambridge University of Press, New York
Marshall LG (1976a) New didelphine from the la Venta fauna (Miocene) of Colombia, South America. J Paleontol 50(3):402–418
Marshall LG (1976b) Evolution of the Thylacosmilidae, extinct saber-toothed marsupials of South America. PaleoBios 23:1–30
Marshall LG (1977) A new species of Lycopsis (Borhyaenidae: Marsupialia) from the La Venta Fauna (Late Miocene) of Colombia, South America. J Paleontol 51(3):633–642
Marshall LG (1979) Review of the Prothylacyninae, an extinct subfamily of South American “Dog-like” Marsupials. Fieldiana NS 3:1–50
Marshall LG (1980) Systematics of the South American Marsupial Family Caenolestidae. Fieldiana NS 5:1–145
Marshall LG (1981a) Review of the Hathlyacyninae, an extinct subfamily of South American “Dog-like” Marsupials. Fieldiana NS 7:1–120
Marshall LG (1981b) The Families and Genera of Marsupialia. Fieldiana NS 8:1–65
Marshall LG (1982a) Systematics of the South American Marsupial Family Microbiotheriidae. Fieldiana NS 10:1–75
Marshall LG (1982b) Systematics of the Extinct South American Marsupial Family Polydolopidae. Fieldiana NS 12:1–109
Marshall LG (1987) Systematics of Itaboraian (middle Paleocene) age “opossum-like” marsupials from the limestone quarry at São José de Itaboraí, Brazil. In: Archer M (ed) Possums and opossums: studies in evolution. Surrey Beatty & Sons Pty Lted, Chipping Norton (R Zool Soc New South Wales)
Marshall LG, de Muizon C (1988) The dawn of the age of mamals in South America. Nat Geogr Res 4:23–55
Marshall LG, Pascual R (1978) Una escala temporal radiométrica preliminar de las Edades -mamífero del Cenozoico medio y tardío sudamericano. In: Obra del Centenario del Museo de La Plata, vol 5. La Plata
Marshall LG, Patterson B (1981) Geology and geochronology of the mammal-bearing tertiary of the Valle de Santa Maria and Rio Corral Quemado, Catamarca Province, Argentina. Fieldiana NS 9:1–80
Marshall LG, Pascual R, Curtis GH, Drake RE (1977) South American geochronology: radiometric time scale for Middle to Late Tertiary mammal-bearing horizons in Patagonia. Science 195(4284):1325–1328
Marshall LG, Butler RF, Drake RE, Curtis DH (1981) Calibration of the beginning of the age of mammals in Patagonia. Science 212:43–45
Marshall LG, de Muizon C, Sigé B (1983a) Late Cretaceous mammals (Marsupialia) from Bolivia. Géobios 16:739–745
Marshall LG, Hoffstetter R, Pascual R (1983b) Geocronology of the continental mammal-bearing tertiary of South America. Palaeovertebrata, Mém Extraord:1–93
Marshall LG, de Muizon C, Sigé B (1985) The “Rosetta stone” for mammalian evolution in South America. Nat Geogr Res 1:274–288
Marshall LG, Case JA, Woodburne MO (1990) Phylogenetic relationships of the families of marsupials. In: Genoways H (ed) Current mammalogy, vol 2. Plenum Press, New York
Martin GM (2010) Geographic distribution and historical occurrence of Dromiciops gliroides Thomas (Marsupialia, Microbiotheria). J Mamm 91:1025–1035
Mercerat A (1898) Sur de nouveaux restes fossils de Carnassiers primitifs de Monte Hermoso. An Soc Cient Arg 47:56–60
O’Leary MO, Bloch JI, Flynn JJ, Gaudin TJ, Giallombardo A, Giannini NP, Goldberg SL, Kraatz BP, Luo Z-X, Meng J, Ni X, Novacek MJ, Perini, Randall ZS, Rougier GW, Sargis EJ, Silcox MT, Simmons NB, Spaulding M, Velazco PM, Weksler M, Wible JR, Cirranello AL (2013) The placental mammal ancestor and the post–K-Pg Radiation of Placentals. Science 339 (662). doi:10.1126/science.1229237
Oliveira EV (1998) Taxonomia, filogenia e paleobiogeografia de marsupiais “poliprotodontes” do Mesopaleoceno da Bacia de Itaboraí, Rio de Janeiro, Brasil. Unpub thesis, Univ Fed Rio Grande do Sul
Oliveira EV, Goin FJ (2011) A reassesment of bunodont metatherians from the Paleogene of Itaboraí (Brazil): Systematics and age of the Itaboraian SALMA. Revista Brasileira de Paleontologia 14(2):105–136
Oliveira EV, Villa Nova P, Goin FJ, Dos Santos Avilla L (2011) A new hyladelphine marsupial (Didelphimorphia, Didelphidae) from cave deposits of northern Brazil. Zootaxa 3041:51–62
Pascual R (1980a) Prepidolopidae, nueva familia de Marsupialia Didelphoidea del Eoceno sudamericano. Ameghiniana 17(3):216–242
Pascual R (1980b) Nuevos y singulares tipos ecológicos de marsupiales extinguidos de América del Sur (Paleoceno tardío o Eoceno temprano) del Noroeste argentino. Actas 2 Congr Arg Paleontol Bioestrat, 1 Congr Latinoam Paleontol Buenos Aires 3:151–173
Pascual R (1981) Adiciones al conocimiento de Bonaparterium hinakusijum (Marsupialia, Bonapartheriidae) del Eoceno temprano del Noroeste argentino. Ann 2 Congr Latinoam Paleontol Porto Alegre 2:507–520
Pascual R (1983) Novedosos marsupiales paleógenos de la Formación Pozuelos (Grupo Pastos Grandes) de la Puna, Salta, Argentina. Ameghiniana 20(3–4):265–280
Pascual R (1984) La sucesión de las Edades-mamífero, de los climas y del diastrofismo sudamericanos durante el Cenozoico: fenómenos concurrentes. Anal Acad Nac Cs Exact Fís Nat 36:15–37
Pascual R (1996) Late Cretaceous-recent land-mammals. An approach to South American geobiotic evolution. Mastozool Neotr 3(2):133–152
Pascual R (2006) Evolution and geography: the biogeographic history of South American land mammals. Ann Missouri Bot Gard 93:209–230
Pascual R, Odreman Rivas O (1971) Evolución de las comunidades de los vertebrados del Terciario argentino, los aspectos paleozoogeográficos y paleoclimáticos relacionados. Ameghiniana 8(3–4):372–412
Pascual R, Odreman Rivas O (1973) Las unidades estratigráficas del Terciario portadoras de mamíferos. Su distribución y sus relaciones con los acontecimientos diastróficos. Acts 5 Congr Geol Arg 3:293–338
Pascual R, Ortiz-Jaureguizar E (1990) Evolving climates and mammal faunas in Cenozoic South America. J Human Evol 19(1–2):23–60
Pascual R, Ortiz-Jaureguizar E (1991) El Ciclo Faunístico Cochabambiano (Paleoceno Temprano): su incidencia en la historia biogeográfica de los mamíferos sudamericanos. In: Suáres Soruco R (ed) Fósiles y facies de Bolivia, vol 1: Vertebrados. Rev Técn Yacimientos Petrolíferos Fiscales, Bolivia 12(3–4), Santa Cruz de la Sierra
Pascual R, Ortiz-Jaureguizar E (2007) The South American mammalian history: two major unrelated episodes. J Mamm Evol 14:75–137
Pascual R, Ortega Hinojosa EJ, Gondar D, Tonni EP (1965) Las edades del Cenozoico mamalífero de la Argentina, con especial atención a aquellas del territorio bonaerense. An Com Invest Cient Prov Buenos Aires 6:165–193
Pascual R, Ortega Hinojosa EJ, Gondar D, Tonni EP (1966) Las edades del Cenozoico mamalífero de la provincia de Buenos Aires. In: Borrello AV, Pascual R (eds) Paleontografía Bonaerense, 4: Vertebrata 2. Com Invest Cien Prov Buenos Aires, La Plata, pp 3–12
Pascual R, Bond M, Vucetich MG (1981) El Subgrupo Santa Bárbara (Grupo Salta) y sus vertebrados. Cronología, paleoambientes y paleobiogeografía. Actas 8 Congr Geol Arg 3:743–758
Pascual R, Ortiz-Jaureguizar E, Prado JL (1996) Land mammals: paradigm of Cenozoic South American geobiotic evolution. In: Arratia G (ed) Contributions of Southern South America to vertebrate paleontology München Geowiss Abh (A) 30, München
Patterson (1937) Didelphines from the Pliocene of Argentine. Proc Geol Soc Amer:379
Patterson B, Pascual R (1968) Evolution of mammals in southern continents. V. Fossil mammal fauna of South America. Quart Rev Biol 43(4):409–451
Patterson B, Pascual R (1972) The fossil mammal fauna of South America. In: Keast A, Erk FC, Glass B (eds) Evolution of mammals and Southern continents. State Univesity of New York Press, Albany
Prado JL, Goin FJ, Tonni EP (1985) Lestodelphys halli (Mammalia, Didelphidae) in Holocene sediments of Southeastern Buenos Aires Province (Argentina): morphological and palaeoenvironmental considerations. Quatern South Amer Antarct Penins 3:93–107
Ré GH, Bellosi ES, Heizler M, Vilas JF, Madden RH, Carlini AA, Kay RF, Vucetich MG (2010) A geochronology for the Sarmiento Formation at Gran Barranca. In: Madden RH, Carlini AA, Vucetich MG, Kay RF (eds) The paleontology of Gran Barranca: evolution and environmental change through the Middle Cenozoic of Patagonia. Cambridge University Press, New York
Reig OA (1952) Descripción previa de nuevos ungulados y marsupiales fósiles del Plioceno y del Eocuartario argentinos. Rev Mus Mun Cs Nat Trad Mar del Plata 1:119–129
Reig OA (1955a) Noticia preliminar sobre la presencia de microbiotherinos vivientes en la fauna sudamericana. Invest Zool Chilenas 2:121–130
Reig OA (1955b) Un Nuevo género y especie de cenolestinos del Plioceno de la Provincia de Buenos Aires. Rev Asoc Geol Arg 10(1):60–71
Reig OA (1957a) Nota previa sobre los marsupiales de la formación Chasicó. Ameghiniana 1(3):27–31
Reig OA (1957b) Sobre la posición sistemática de Zygolestes paranensis Amegh. Y de Zygolestes entrerrianus Amegh., conuna reconsideración sobre la edad y la correlación del “Mesopotamiense“. Holmbergia 5:209–226
Reig OA (1958a) Comunicación preliminar sobre nuevas especies del género Thylatheridium Reig (Mammalia, Didelphidae). Neotropica 4(15):89–95
Reig OA (1958b) Notas para una actualización el conocimiento de la fauan de la formación Chapadmalal. I. Lista faunística preliminar. Acta Geol Lilloana 2:251–254
Reig OA (1958c) Notas para una actualización el conocimiento de la fauan de la formación Chapadmalal. II. Amphibia, Reptilia, Aves, Mammalia (Marsupialia; Didelphidae, Borhyaenidae). Acta Geol Lilloana 2:255–283
Reig OA, Simpson GG (1972) Sparassocynus (Marsupialia, Didelphidae), a peculiar mammal from the late Cenozoic of Argentina. J Zool London 167:511–539
Ride WDL (1964) A review of Australian fossils marsupials. J R Soc Western Australia 47:97–131
Riggs E, Patterson B (1939) Stratigraphy of late Miocene and Pliocene deposits os the Province of catamarca (Argentina), with notes on the faunae. Physis 14:143–162
Ringuelet AB (1953) Revisión de los Didélfidos fósiles argentinos. Rev Mus Cs Nat La Plata NS 2:265–308
Ringuelet AB (1966) Marsupialia. In: Borrello AV, Pascual R (eds) Paleontografía Bonaerense, Vol 4 Vertebrata. Com Invest Cient Prov Bsuenos Aires, La Plata
Rougier GW, Wible JR, Novacek MJ (1998) Implications of Deltatheridium specimens for early marsupial history. Nature 396:459–463
Rougier GW, Wible JR, Beck RMD, Apesteguía S (2012) The Miocene mammal Necrolestes demonstrates the survival of a Mesozoic nontherian lineage into the late Cenozoic of South America. PNAS 109:20053–20058
Rovereto R (1914) Los estratos araucanos y sus fósiles. Anal Mus Hist Nat Buenos Aires 25:1–249
Rowe T (1998) Definition, diagnosis, and origin of mammalia. J Vert Paleontol 8(3):241–264
Rusconi R (1932) Una nueva especie de comadreja fósil Didelphys seneti n. sp. An Soc Cient Arg 113:213–215
Sánchez-Villagra MR (2001) The phylogenetic relationships of argyrolagid marsupials. Zool J Linn Soc 131:481–496
Sánchez-Villagra MR (2012) Why are there fewer marsupials than placentals? On the relevance of geography and physiology to evolutionary patterns of mammalian diversity and disparity. J Mammal Evol. doi:10.1007/s10914-012-9220-3
Sánchez-Villagra MR, Kay RF (1997) A skull of Proargyrolagus, the oldest argyrolagid (Late Oligocene Salla Beds, Bolivia), with brief comments concerning its paleobiology. J Vert Paleontol 17:717–724
Sánchez-Villagra MR, Kay RF, Anaya-Daza F (2000) Cranial anatomy and palaeobiology of the miocene marsupial Hondalagus altiplanensis and a phylogeny of argyrolagids. Palaeontol 43(2):287–301
Sbarra N (1963) La Plata tuvo una Universidad al aire libre. In: Universidad nueva y ámbitos culturales platenses. Ed. UNLP, La Plata
Seba A (1734) Locupletissimi rerum naturalium thesauri accurata descriptio, et iconibus artificiosissimis expressio, per universam physices historiam. Opus, cui, in hoc rerum genere, nullum par exstitit. Ex toto terrarum orbe collegit, digessit, et depingendum curavit, vol I. Wetstenium. Smith & Janssonio-Waesbergios, Amsterdam
Sigé B (1971) Les Didelphoidea de Laguna Umayo (Formation Vilquechico, Crétacé supérieur, Pérou), et le peuplement marsupial d’Amérique du Sud. Compt Rendus Acad Sci 273:2479–2481
Sigé B (1972) La faunule de mammifères du Crétacé supérieur de Laguna Umayo (Andes péruviennes). Bull Mus Nat Hist Nat 99 Sc Terre 19:375–409
Sigé B, Sempere T, Butler RF, Marshall LG, Crochet J-Y (2004) Age and stratigraphic reassessment of the fossil-bearing Laguna Umayo red mudstone unit, SE Peru, from regional stratigraphy, fossil record, and paleomagnetism. Géobios 37:771–794
Simpson GG (1932a) Some new or little known mammals from the Colpodon beds of Patagonia. Amer Mus Novit 575:1–12
Simpson GG (1932b) The supposed occurrences of Mesozoic mammals in South America. Amer Mus Novit 530:1–9
Simpson GG (1935a) Descriptions of the oldest known South American mammals, from the Río Chico formation. Amer Mus Novit 793:1–25
Simpson GG (1935b) Note of the classification of Recent and fossil opossums. J Mammal 16:134–137
Simpson GG (1938) A new marsupial from the Eocene of Patagonia. Amer Mus Novit 989:1–5
Simpson GG (1948) The beginning of the age of mammals in South America, Part 1. Bull Amer Mus Nat Hist 91:1–232
Simpson GG (1950) History of the fauna of Latin America. Am Sci 38:361–389
Simpson GG (1967) The beginning of the age of mammals in South America, Part 2. Bull Amer Mus Nat Hist 137:1–259
Simpson GG (1972) Didelphidae from the Chapadmalal Formation in the Museo Municipal de Ciencias Naturales of Mar del Plata. Publ Mus Cs Nat Trad Mar del Plata 2:1–40
Simpson GG (1974) Notes on the Didelphidae (Mammalia, Marsupialia) from the Huayquerian (Pliocene) of Argentian. Amer Mus Novit 2259:1–15
Simpson GG (1980) Splendid isolation: the curious history of South American mammals. Yale Univesity Press, New Haven
Sinclair E (1906) Marsupialia of the Santa Cruz beds. In: Reports of the Princeton University Expeditions to Patagonia, 1896–1899, 4 (Paleontology). Princeton
Szalay FS (1982) A new appraisal of marsupial phylogeny and classification. In: Archer M (ed) Carnivorous marsupials, vol 2. R Zool Soc New South Wales, Sydney
Szalay FS (1994) Evolutionary history of the marsupials and an analysis of osteological characters. Cambridge University Press, New York
Tejedor MF, Goin FJ, Gelfo JN, López G, Bond M, Carlini AA, Scillato-Yané GJ, Woodburne MO, Chornogubsky L, Aragón E, Reguero M, Czaplewski N, Vincon S, Martin GM, Ciancio M (2009) New Early Eocene mammalian fauna from Western Patagonia, Argentina. Amer Mus Novit 3638:1–42
Tomassini RL, Montalvo CI, Deschamps CM, Manera T (2013) biochronology of the Monte Hermoso formation (early Pliocene) at its type locality, Buenos Aires Province, Argentina. J South Ameri Earth Sci 48:31–42
Tonni EP (2009) Los mamíferos del Cuaternario de la región pampeana de Buenos Aires, Argentina. In: Ribeiro AM, Girardi Bauermann C, Saldanha Scherer C (eds) Quaternario do Rio Grande do Sul. Integrando Conhecimentos. Monogr Soc Brasileira Paleontol, Porto Alegre
Ugalde MF (2006) Difusión en el período de Desarrollo Regional: algunos aspectos de la iconografía Tumaco-Tolita. Bull Instit Français d’Études Andines 35(3):397–407
Villarroel C, Marshall LG (1982) Geology of the Deseadan (early Oligocene) age Estratos Salla in the Salla-Luribay basin, Bolivia, with description of new Marsupialia. Geobios Mem Spec 6:201–211
Villarroel C, Marshall LG (1983) Two new late Tertiary marsupials (Hathlyacyninae and Sparassocyninae) from the Bolivian Altiplano. J Paleontol 57:1061–1066
Voss RS, Jansa SA (2009) Phylogenetic relationships and classification of didelphid marsupials, an extant radiation of New World metatherian mammals. Bull Amer Mus Nat Hist 322:1–177
Williamson TE, Brusatte SL, Carr TD, Weil A, Standhardt BR (2012) The phylogeny and evolution of Cretaceous-Palaeogene metatherians: cladistic analysis and description of new early Palaeocene specimens from the Nacimiento Formation, New Mexico. J Syst Palaeontol 10(4):625–651
Williamson TE, Brusatte SL, Wilson GP (2014) The origin and early evolution of metatherian mammals: the Cretaceous record. ZooKeys 465:1–76. doi:10.3897/zookeys.465.8178
Wilson DE, Mittermeier RA (eds) (2015) Handbook of the Mammals of the World, vol 5, Monotremes and Marsupials. Lynx Edicions, Barcelona, p 799
Winge A (1893) Jordfundne og nulevende Pungdyr (Marsupialia) fue Lagoa Santa, Minas Gerais, Brasilien. Med, Udsigt over Pungdyrenes Slaegtskab. E Mus Lund 11(2):1–149
Woodburne MO, Zinsmeister WJ (1982) Fossil land mammal from Antarctica. Science 218:284–286
Woodburne MO, Zinsmeister WJ (1984) The first land mammal from Antarctica and its biogeographic implications. J Paleontol 58:913–948
Woodburne MO, Goin FJ, Bond M, Carlini AA, Gelfo JN, López GM, Iglesias A, Zimicz AN (2013) Paleogene Land Mammal Faunas of South America; a response to global climatic changes and indigenous floral diversity. J Mamm Evol. doi:10.1007/s10914-012-9222-1
Woodburne MO, Goin FJ, Bond M, Carlini AA, Gelfo JN, López GM, Iglesias A, Zimicz AN (2014a) Paleogene Land Mammal Faunas of South America; a response to global climatic changes and indigenous floral diversity. J Mamm Evol. doi:10.1007/s10914-012-9222-1
Woodburne MO, Goin FJ, Raigemborn MS, Heizler M, Gelfo JN, Oliveira EV (2014b) Revised timing of the South American early Paleogene land mammal ages. J S Am Earth Sci 54:109–119
Wyss AR, Norell MA, Flynn JJ, Novacek MJ, Charrier R, Salinas P, McKenna M, Meng J (1990) A new Early Tertiary mammal fauna from central Chile: implications for Andean tectonics. J Vert Paleontol 10:518–522
Wyss AR, Flynn JJ, Norell MA, Swisher CCIII, Novacek MJ, McKenna MC, Charrier R (1994) Paleogene mammals from the Andes of Central Chile: a preliminary taxonomic, biostratigraphic, and geochronologic assessment. Amer Mus Novit 3098:1–31
Zetti A (1967) Sobre la presencia del género Marmosa (Didelphidae, Marsupialia) en sedimentos de la Formación Epecuén (Plioceno Medio). Ameghiniana 5(5):169–173
Zimicz AN (2014) Avoiding competition: the ecological history of metatherian carnivores. J Mamm Evol 21(4):383–393
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Copyright information
© 2016 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Goin, F.J., Woodburne, M.O., Zimicz, A.N., Martin, G.M., Chornogubsky, L. (2016). Introduction. In: A Brief History of South American Metatherians. Springer Earth System Sciences. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-7420-8_1
Download citation
DOI: https://doi.org/10.1007/978-94-017-7420-8_1
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-017-7418-5
Online ISBN: 978-94-017-7420-8
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)