
Abstract
We assess the status and structure of coral reef fish assemblages in the large remote and unfished Chagos Archipelago, comparing fish biomass and structure among atolls and across the wider western Indian Ocean. We then assess the longer term trends in reef shark abundances and the stability of fish assemblages through the 1998 climate induced habitat disturbance. Diego Garcia atoll had the lowest standing reef fish biomass estimates, while Peros Banhos and the Great Chagos Bank had the highest. Further, the biomass of larger bodied, higher trophic level, and most fishery target families was higher in the northern atolls, indicating a potential impact of the recreational fishery operating around Diego Garcia. Fish biomass in the northern atolls of Chagos was up to six times greater than estimates from even the more successful small marine protected areas elsewhere in the western Indian Ocean region. Interestingly, the biomass recorded around Diego Garcia was comparable to the highest values reported elsewhere in the region, suggesting that although the recreational fishery has had some impact at that atoll, it is far from overfished. Reef shark abundance has declined substantially since the 1970s, largely due to illegal fishing activities, however the numbers recorded in 2010 and 2012 were higher than those recorded in 1996 and 2006. The 1998 mass coral bleaching event did not have a major impact on reef fish assemblages in Chagos, with specialised coral feeding fishes being numerically dominant over less specialised coral feeders by 2012. The Chagos Archipelago provides an example of the biomass and community structure of reef fishes that many reefs likely had in the past, and also demonstrates the considerable resilience that coral reefs can exhibit when they are not otherwise stressed by chronic human pressures.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Introduction
Coral reef fishes are conspicuous components of coral reef ecosystems, representing the most diverse vertebrate communities on Earth (Jones et al. 2002). Reef fishes have attracted a great deal of research and management attention because of their vulnerability to fishing, and because many fishes have important functional roles within coral reef ecosystems. These functions include herbivory which mediates the competition between corals and benthic algal communities (Bellwood et al. 2004), predation on invertebrates that modify habitat (McClanahan 2000), or piscivory that mediates competitive interactions among fish (Hixon 1991). Fishes also form some of the strongest ecosystem service links between coral reefs and human societies, providing benefits such as food and income through fisheries and tourism (Polunin and Roberts 1996, Williams and Polunin 2000).
The sensitivity of fishes to depletion through fishing activities has been documented in a range of studies with rapid depletion of fish biomass even with relatively light fishing pressure. The most vulnerable groups are often species with higher trophic levels, such as groupers and snappers (Russ and Alcala 1989; McClanahan 1994; Jennings and Polunin 1997; McClanahan et al. 2008). Additionally, there are significant changes in reef fish size resulting in ecosystems being dominated by small-bodied individuals and species (Dulvy et al. 2004; Graham et al. 2005; McClanahan and Omukoto 2011). Even recreational fishing can have substantial impacts on fish populations (Coleman et al. 2004). Given the known sensitivity of reef fish assemblages to fishing, much interest has developed around management techniques to redress declines and restore pristine ecosystems (Pitcher 2001; McClanahan 2011).
A great deal of management and research attempting to restore coral reef ecosystems has focused on no-take marine protected areas (or marine reserves), with most studies documenting increases in the abundance, biomass and sometimes the diversity of reef fish assemblages within well-enforced marine reserves (Russ et al. 2005; McClanahan et al. 2007a; Stockwell et al. 2009). However, considerable debate has questioned whether marine reserves, which are generally <10 km2 (McClanahan et al. 2009), are capable of promoting biomass and trophic structures that foster ecological processes in a ‘pristine’ environment (Graham et al. 2011a). Indeed, recent studies in remote, uninhabited atolls of the NW Hawaiian Islands and northern Line Islands in the Pacific have documented reef fish biomass, dominated by top predators, that far exceeds the biomass values quantified for even relatively large and old marine reserves (Friedlander and DeMartini 2002; Sandin et al. 2008; DeMartini et al. 2008; Williams et al. 2011). The Chagos Archipelago is likely to be a similar reference site in the Indian Ocean, as it covers a very large area (with ∼60,000 km2 of potential reef area) and the northern atolls have been uninhabited since the early 1970s with very little fishing activity even before the current no-take areas status was declared in April 2010 (Sheppard et al. 2012). Extensive fish surveys have been conducted across much of the western Indian Ocean and these provide a useful context for evaluating the status of the Chagos (Graham et al. 2008; McClanahan et al. 2011).
Studies in remote Pacific locations have highlighted the high abundance and biomass of reef sharks (Sandin et al. 2008; Nadon et al. 2012). In Chagos, however, sharks are one of the few groups that have been substantially depleted through illegal fishing activities. The relative abundance of reef sharks in the northern atolls of Chagos are estimated to have declined by ∼90% from the 1970s to 2006 (Anderson et al. 1998; Spalding 2003; Graham et al. 2010). Consequently, the trophic structure of reef fish communities at Chagos may differ from other remote locations. These results emphasize the importance of monitoring reef shark populations into the future, as efforts to enforce the recently declared Chagos no-take marine protected area develop and take effect.
Reef fishes are also sensitive to disturbances to their coral reef habitat (Jones and Syms 1998). If reef corals die and the physical structure of the reef subsequently erodes, reef fish abundance and diversity can decline (reviewed by Wilson et al. 2006; Pratchett et al. 2008). Fish species specialized on corals for food (Pratchett et al. 2006; Graham 2007), habitat (Munday 2004, Wilson et al. 2008a, Pratchett et al. 2012) or settlement (Jones et al. 2004) typically show the largest declines in abundance. Body size also appears to be a key determinant of fish species vulnerability to habitat degradation, with smaller body size classes showing the greatest declines (Graham et al. 2006, 2007; Wilson et al. 2008b, 2010). Chagos, like much of the northern Indian Ocean, was badly impacted by the 1998 thermal anomaly event, with up to 80% of the corals dying on many reefs (Sheppard 1999). However, the reefs have shown a strong recovery and by 2010 coral cover had returned to pre-1998 levels (Ateweberhan et al. 2011; Sheppard et al. 2012). It is useful to understand how reef fish have responded to these substantial benthic changes in remote locations such as Chagos, where local anthropogenic impacts are minimal and to allow comparisons with other locations with substantial local human impacts.
In this chapter we assess the reef fish biomass and trophic structure at Chagos, comparing data within and among atolls (Salomon, Peros Banhos, Great Chagos Bank and Diego Garcia), with a particular focus on the effects of recreational fishing around Diego Garcia. These data are then put into the context of the wider western Indian Ocean, including smaller marine reserves embedded in fished coastlines. Trends in reef shark abundances at Chagos are updated from Graham et al. (2010) to include data from 1975 to 2012. Finally, we assess the impacts of habitat change in Chagos on reef fish assemblages, compared to other locations in the western Indian Ocean, with a particular focus on the importance of specialization in corallivorous fishes.
Reef Fish Biomass Among Atolls of Chagos
It is estimated that there are at least 784 species of fishes in Chagos (e.g. Winterbottom and Anderson 1999). Spalding (1999) compared the density and diversity of fishes at Chagos to the relatively nearby Seychelles in 1996, reporting slightly higher species richness per site in Chagos, but slightly lower species over smaller replicate count areas (Spalding 1999). Following the 1998 mass bleaching substantial declines in species richness and abundances of certain guilds (e.g. coral feeding fishes) were reported across the western Indian Ocean where coral cover declined, but the fish assemblages at Chagos displayed relative stability (Graham et al. 2008; Sheppard et al. 2012; see ‘Habitat degradation effects on reef fishes in Chagos’ below). All of this work indicated that reef fish assemblages in Chagos tolerated the 1998 disturbance well, but the biomass of these assemblages had not been quantified.
Larger bodied fish contribute disproportionately to standing biomass estimates of fish assemblages. However large fish are much more vulnerable to fishing (Dulvy et al. 2004; Olden et al. 2007; Graham et al. 2011b), and substantial declines in reef fish biomass have been detected on coral reefs associated with increasing fishing pressure or efficiency (McClanahan 1994; Jennings and Polunin 1996; Russ and Alcala 1989; Newman et al. 2006; Cinner et al. 2009, 2013). Fish biomass is thought to be important to the functioning of coral reefs, maintaining a range of ecosystem processes (McClanahan et al. 2011; Mora et al. 2011). It is therefore critical to have reliable estimates of reef fish assemblage biomass in the absence of fishing pressure. Small marine reserves can substantially enhance fish biomass compared to adjacent fished areas (e.g. Russ et al. 2005; McClanahan et al. 2007a), but large, remote and unfished locations in the Pacific far outstrip the biomass values recorded in even the most successful, well managed, small marine reserves (Friedlander and DeMartini 2002; Stevenson et al. 2007; Sandin et al. 2008). In Chagos, a small seasonal (June – August) Mauritian fishery operated in the archipelago from the early 1970s until 2010, and was under license since 1991 (Mees 1996). This fishery was relatively small, targeting grouper and snappers on banks and in deeper water and there was no uptake of licences in some years (Mees 1996, C. Mees, personal communication). It is therefore unlikely that this fishery affected the reef fish populations at the depth and habitats that we have quantified them here.
At Diego Garcia there is also a small recreational fishery associated with the US Navy base. Affects from this recreational fishery on local fish communities may be significant and needs to be assessed.
Reef fish biomass at Chagos was first quantified in the 2010 Chagos scientific expedition. During this trip 18 sites were surveyed using underwater visual census techniques on the outside of Salomon and Peron Banhos atolls and around the Three Brothers and Eagle Island on the Great Chagos Bank (Fig. 19.1). A further 6 sites were surveyed on the outside of Diego Garcia during the 2012 expedition using the same techniques (Fig. 19.1). At each site fish were surveyed within four 50 by 5 m belt transects, by the same observer (N. Graham), along the 7–9 m depth contour. All diurnally active, non-cryptic, reef associated fishes larger than 8 cm were identified to species, counted and length estimated to the nearest centimeter. Larger more mobile species were surveyed on a first pass of the transect, while smaller fish, principally damselfishes, were surveyed within a 2 m belt on the return pass. Biomass was estimated for each counted individual using the length estimate and published length-weight relationships for that species or genus (Froese and Pauly 2012). Benthic cover, was quantified every 50 cm along each 50 m transect and structural complexity was estimated on a 6 point visual scale following Wilson et al. (2007).

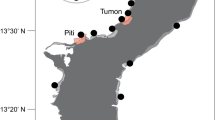
Map of sites surveyed: (a) Diego Garcia, (b) Chagos archipelago and (c) The western Indian Ocean
Diego Garcia had similar coral cover to the Great Chagos Bank and Peros Banhos atoll sites, but lower cover than Salomon atoll (Table 19.1 and Fig. 19.2a). Sites at Salomon atoll had the highest coral cover, but cover did not differ significantly to Peros Banhos. Diego Garcia also had among the lowest structural complexity values, and was similar only to the Great Chagos Bank, while sites at Peros Banhos had the highest values (Table 19.1 and Fig. 19.2b).

Variation in (a) Hard coral cover (%) and (b) Structural complexity among atolls within the Chagos archipelago. Error bars represent one standard error of the mean. Horizontal lines show homogeneous subsets from post hoc comparisons using the Tukey test
In terms of total fish biomass, Diego Garcia had the lowest while Peros Banhos and the Great Chagos Bank had the highest values (Table 19.1 and Fig. 19.3a). Pairwise comparisons indicate that total fish biomass between Diego Garcia and Salomon atoll were similar, however the mean values at Diego Garcia are less than half those of Salomon. Shark biomass was not statistically different among atolls (Table 19.1 and Fig. 19.3b), although this may be due to high variation within atolls likely due to the relatively small sampling units for these large mobile organisms. Despite this variation, the Great Chagos Bank appears to have considerably higher shark biomass than the other three atolls. If the total fish biomass is broken down into families typically targeted in fisheries (e.g. groupers, snappers, and emperors) versus non-targeted fish families (e.g. damselfish, butterflyfish, and angelfish), significant variation among atolls is apparent for targeted biomass (Table 19.1 and Fig. 19.3c), but not non-targeted biomass (Fig. 19.3d). Targeted fish biomass was highest at Peros Banhos, intermediate on the Great Chagos Bank and lowest at Salomon followed by Diego Garcia (Fig. 19.3c).

Variation in (a) Total fish biomass, (b) Shark biomass, (c) Targeted fish biomass and (d) Untargeted fish biomass (kg/ha) among atolls in the Chagos archipelago. Error bars represent one standard error around the mean. Horizontal lines show homogeneous subsets from post hoc comparisons using the Tukey test
Higher levels of reef structural complexity can have a positive influence on reef fish biomass, and is generally more important than coral cover for fish biomass (Cinner et al. 2009). If any effect of reef structural complexity on targeted fish biomass is controlled for with an analysis of covariance, the difference in biomass among atolls still comes out as highly significant, explaining 29% of the total variance in the data (structural complexity, F = 8.23, p < 0.01; atoll, F = 21.45, p < 0.0001). Structural complexity is therefore influencing reef fish biomass at Chagos, but the difference among atolls occurs independent of structural complexity, which can be noted from the patterns in structural complexity versus biomass among atolls (Figs. 19.2 and 19.3).
Breaking down the fish biomass by trophic level and body size classes produces some illuminating patterns (based on species level information obtained from FishBase; Froese and Pauly 2012). The fish communities around the northern atolls appear to be represented by higher trophic level and larger sized fish than those around Diego Garcia (Fig. 19.4). Only ∼35% of the community around Diego Garcia is composed of fish with a trophic level greater than 3.5, while this portion of the community makes up ∼60–70% of the community in the three northern atolls (Fig. 19.5a). Similarly, fish with a body size greater than 50 cm only make up ∼ 25% of the community biomass around Diego Garcia, whereas they make up between 40 and 60% in the northern atolls (Fig. 19.5b). Large bodied fish are typically the most vulnerable to exploitation (Olden et al. 2007), and can decline substantially in response to fairly light fishing pressure (Dulvy et al. 2004; Wilson et al. 2010). Similarly, higher trophic level organisms are typically vulnerable to exploitation and are conspicuously missing in many food webs (Estes et al. 2011). These patterns, along with the biomass analyses (Fig. 19.3), indicate that the recreational fishery around Diego Garcia is reducing fish biomass and size compared to the relatively unfished northern atolls.

High fish biomass at Chagos. (a) A large coral trout, Plectropomus laevis, (b) A school of paddletail snapper, Lutjanus gibbus, (c) A school of captain parrotfish, Chlorurus enneacanthus, (d) A mix of snapper species along the reef bottom, including red snapper, Lutjanus bohar, paddeltail snapper, Lutjanus gibbus, and midnight snapper, Macolor niger (Photo’s: N Graham)

Relative contributions of fish (a) Trophic levels and (b) Body size classes (cm) to the total fish biomass among atolls within the Chagos archipelago
Community composition of fish varied considerably among sites and atolls (multidimensional scaling plot, MDS, Fig. 19.6). All the sites around Diego Garcia fall to the far left hand side of the ordination, while the other atolls, in particular Peros Banhos, spread out towards the right hand side of the ordination (Fig. 19.6a). The 10 most important fish families driving these patterns are shown as vectors (Fig. 19.6b), with almost all families, including all those that are potentially targets of fishing, increasing to the right side of the ordination. This indicates that in almost all cases the biomass of these important families is higher in the northern atolls than around Diego Garcia, potentially highlighting the influence of recreational fishing on the fish community biomass.

Multidimensional scaling analysis of fish family biomass. (a) Plot of the spatial variation in fish family level biomass among sites in Chagos. Each site is coloured according to its corresponding atoll. (b) The relative contribution and direction of influence of the top ten fish families to the observed variation among sites. Only fish families with a correlation > 0.3 with either axis MDS1 or MDS2 were selected for representation in the vector plot
Focusing specifically on Diego Garcia provides some interesting patterns. Among the six sites around the atoll there were significant differences in hard coral cover (Table 19.2 and Fig. 19.7a) and structural complexity (Table 19.2 and Fig. 19.7b). There was a gradient from a coral cover of 8% and a relatively low structural complexity value of 1.5 at Cannon Point site 1 to a high coral cover of 59% at Barton Point west and a high structural complexity of 3.4 in Barton Point east. Fish biomass showed considerable variability among sites, but there was no significant difference when assessing total fish biomass (Fig. 19.7c). However, if only fish families known to be targeted in fisheries of the region (based on body size, and susceptibility to fishing gears) were assessed, biomass at Cannon Point site 2 was lower than at Middle island (Table 19.2 and Fig. 19.7d). Interestingly, the sites at Cannon Point are adjacent to the Navy base, and as such may experience the most recreational fishing pressure. Although both coral cover and structural complexity were generally lower at the Cannon Point sites, regression analysis showed no significant relationship between benthic reef structure and total, targeted or untargeted fish biomass at Diego Garcia.

Variation in (a) Hard coral cover (%), (b) Structural complexity, (c) Total fish biomass (kg/ha) and (d) Targeted fish biomass (kg/ha) among sites at Diego Garcia. Error bars represent one standard error of the mean. Horizontal lines at a same level show homogeneous subsets from post hoc comparisons using the Tukey test
Comparison of Fish Biomass Across the Wider Indian Ocean
Overfishing on coral reefs is a pervasive problem, which is thought to have started centuries ago in some locations (Jackson et al. 2001; McClanahan and Omukoto 2011). Research indicates that the first effects of fishing are often the most severe in terms of biomass reduction, as slow-growing and large bodied fishes are vulnerable to fishing, and also contribute significantly to standing biomass (Reynolds et al. 2001). Given widespread depletion of fish assemblages in many countries, small marine reserves have become a dominant management tool attempting to reverse depletion trends and return fish biomass to high levels (Graham et al. 2011a). Putting the Chagos fish biomass in a broader geographic context produces an insightful view of the status of some of these smaller marine reserves that are embedded in fished landscapes. Broad spatial comparisons also gives biomass estimates in the northern atolls context and provides a clearer picture of just how heavily impacted the fish assemblages are around Diego Garcia. Fish biomass and coral cover has been collected in fished areas and marine reserves using comparable survey methods across 8 other western Indian Ocean countries (Fig. 19.1c). These data, collected principally by T McClanahan, N Graham and S Wilson, cover over 100 sites across the region and provide a basis for comparisons with fish and coral communities in Chagos. Some of the marine reserves in the region have reached asymptotes in fish biomass recovery (McClanahan et al. 2009), and prior to these surveys of Chagos, the relatively lightly fished Maldives was the site with the greatest recorded reef fish biomass (McClanahan 2011). Nevertheless, both historical records of fish catch from before the thirteenth century (McClanahan and Omukoto 2011) and evaluations of life history characteristics indicated that these reserves were not in pristine condition (McClanahan and Humphries 2012).
Hard coral cover varies substantially across the region (Table 19.3 and Fig. 19.8). The Seychelles have the lowest coral cover with an average of 14 ± 3%, Diego Garcia falls within the middle group with average hard coral cover of 38 ± 4%, while the northern atolls in Chagos have among the highest coral cover values in the region, with an average of 44 ± 4% (Fig. 19.8). The western Indian Ocean was one of the regions most affected by the 1998 thermal anomaly; losing ∼45% total coral cover (Ateweberhan et al. 2011). However, the impacts varied greatly around the region with extensive coral decline at the low latitude island nations (Seychelles, Chagos and Maldives) whilst impacts at higher latitude nations to the south, such as Mauritius, Tanzania, Mozambique and Madagascar, were minimal (McClanahan et al. 2007b; Graham et al. 2008; Ateweberhan et al. 2011).

Variation in hard coral cover (%) across nine countries in the western Indian Ocean. Diego Garcia and Chagos other atolls (Salomon, Peros Banhos and Great Chagos Bank) are presented separately. Error bars represent one standard error around the mean
Many of the patterns in coral cover reported here are a consequence of the 1998-bleaching event (Fig. 19.8). For example, the inner Seychelles lost >90% of its coral cover in 1998, and recovery of cover has been slow and variable among sites (Wilson et al. 2012). Similarly, both the Maldives and Kenya still have reduced coral cover at many sites in the aftermath of the 1998 bleaching event (McClanahan 2008; Ateweberhan et al. 2011). Given the substantial declines in coral cover in Chagos following 1998 (Sheppard 1999), the high cover reported here reflects substantial recovery not seen in other locations similarly impacted across the region. Indeed, the rapid recovery of hard coral cover in Chagos has been well documented (Sheppard et al. 2008; 2012) and contrasts markedly to many other locations in the region (Ateweberhan et al. 2011).
We do not have consistent structural complexity data across all of these locations but it is expected that structural complexity and coral cover will be positively correlated at this scale (Graham et al. 2008). Importantly, reef habitats in Chagos support much higher fish biomass than equivalent sites in the western Indian Ocean with comparable coral cover. This suggests human population density, distance to markets and community level economic development (which relate to fishing pressure and efficiency) are likely to be the key determinants of variation in reef fish biomass across the region (Cinner et al. 2009).
Total fish biomass in the three northern atolls of Chagos dwarf the biomass seen at all other reef sites across the wider region (Table 19.3 and Fig. 19.9a). Indeed, biomass values in the northern atolls are six times greater than those recorded from even the most successful small marine reserves in the region (McClanahan et al. 2009, 2011). A great deal of this difference is due to greater abundance of higher trophic level fishes and those with a larger overall body size in the northern atolls (Graham and McClanahan in press). The performance of the marine reserves across the region, in terms of building up fish biomass compared to fished areas, varies substantially (Fig. 19.9a). Some locations, such as Kenya, have relatively large (up to ∼ 30 km2) and old (up to ∼ 40 years) marine reserves that are well enforced and compliance is high. These reserves recover from the effects of fishing in ∼20 years, when estimates of fish biomass asymptote (McClanahan and Graham 2005), indicating they have reached their full biomass potential. Nevertheless, they continue to change in terms of the composition of the fish fauna towards slow-maturing and growing species (McClanahan and Humphries 2012).

Variation in reef fish biomass across nine countries in the western Indian Ocean including sites in fished and unfished areas. (a) Total fish biomass (kg/ha), (b) Total fish biomass (kg/ha) broken down by targeted and untargeted fish families. The open portion of the bars in panel (b) represent families that could not be easily assigned as targeted or untargeted. The y axis on panel (b) Have been cut off at 1,600 kg/ha to facilitate comparison between Diego Garcia and the other eight countries. Error bars represent 1 standard error around the mean
Biomass in the northern atolls of Chagos is, therefore, much higher than even the most effective smaller reserves. Biomass differences are attributable to greater abundance of larger, higher trophic level fishes in the northern atolls. Many of these fish, such as the grey reef shark, have limited reef fidelity and large home ranges (Heupel et al. 2010). Hence, it is unlikely that small reserves embedded in fished landscapes are adequately protecting the top end of the foodweb, with larger fish exposed to fishing pressure when they move outside park boundaries (Graham and McClanahan in press). The archaeological record of fish bones also support this contention of reduced trophic level and other life history characteristics in these marine reserves (McClanahan and Omukoto 2011).
Fish biomass at Diego Garcia is much more comparable to some of the other locations across the region (Fig. 19.9a, b). It should be noted however, that Diego Garcia is similar to the highest values reported across the region, for example the Maldives, Mayotte and Kenyan marine reserves (McClanahan et al. 2011). When the fish community is broken down by species targeted by fisheries, versus those that are not, the differences become more apparent. While the northern atolls have an exceptionally high targeted fish biomass (mean 4,930 kg/ha), Diego Garcia had values similar to the best performing marine reserves in the region (Fig. 19.9b). This indicates that although the recreational fishery around Diego Garcia does seem to be having an impact of the fish biomass compared to the northern atolls, the biomass in Diego Garcia is still very high, and is far from an overfished status.
Various studies have shown that the first effects of fishing are often the most substantial in terms of reducing fish biomass, mainly because the slow-growing and large bodied fish that constitute most of biomass are the first to be caught and removed (e.g. Jennings and Polunin 1996, McClanahan and Omukoto 2011), and even recreational fishing can have substantial impacts on fish biomass (Coleman et al. 2004). The recreational fishery around Diego Garcia should be very carefully controlled and monitored to prevent any further declines in fish biomass in the future, and ideally management put in place to try to rebuild these fish stocks toward the status of the northern atolls. Diego Garcia is currently the only atoll in Chagos not included in the no-take marine protected area, however this does not mean more restrictive and targeted management, such as protecting a large portion of the atoll, or putting caps on fishing effort or restrictions of specific species, could not be achieved. It should be noted that some of the marine reserves in other countries across the region are not enhancing fish biomass compared to fished areas in the same countries. This is likely due to a combination of some of the areas being small and not very old (McClanahan et al. 2009) and in some locations weak compliance by fishers and heavy fishing around the borders of the reserves (Pollnac et al. 2010; Daw et al. 2011).
It is clear that the biomass of reef fish in the northern atolls of Chagos is remarkably high, especially in the context of the wider western Indian Ocean region. The most likely explanation for this is the lack of fishing at a large scale in Chagos compared to the other countries. This may be thought of as an exploitation gap; the effect fishing has on reef fish assemblages compared to semi-pristine unfished locations. The key ecosystem processes fish provide (Bellwood et al. 2004; McClanahan et al. 2011; Mora et al. 2011) suggests that the high, relatively unexploited, biomass of fish in Chagos is key to the stability and health of the coral reef ecosystem in Chagos as a whole. Indeed, high abundance/biomass of the reef fish assemblage is a likely reason recovery of reefs in Chagos from the 1998 thermal anomaly was rapid compared to other reefs in the region. Locations such as Chagos are increasingly rare in the world’s oceans and provide unique opportunities to understand what coral reef ecosystems should look like and how they function with minimal human impacts. They also represent some of the few areas likely to persist in a coral dominated and relatively intact state in the face of climate change.
Aside from Chagos, exceptional reef fish biomass and coral reef ecosystem condition has been documented from several locations in the Pacific Ocean. The uninhabited, and now protected, northwest Hawaiian Islands is one such example, where fish biomass, particularly the biomass of apex predators, is far greater than the biomass recorded around the populated main Hawaiian Islands (Friedlander and DeMaritini 2002). Similarly, the remote uninhabited northern Line Islands, in particular Palmyra and Kingman atolls, have exceptional reef fish biomass compared to the populated and fished southern Line Islands (Stevenson et al. 2007; DeMartini et al. 2008; Sandin et al. 2008; Williams et al. 2011). The biomass on these remote unfished Pacific locations is comparable to the biomass reported here for Chagos (for example biomass at Kingman atoll was reported as 5,500 kg/ha; Sandin et al. 2008). However, many of the locations in the Pacific report top predators, such as reef sharks, dominating the biomass of the assemblage (Friedlander and DeMartini 2002; Sandin et al. 2008). The contribution of fish trophic groups to the composition of fish biomass in Chagos is much more balanced (Graham and McClanahan in press). It must however be noted that reef sharks are one of the few groups of fish that have experienced some declines in Chagos, largely due to illegal fishing. It is therefore important to understand longer term trends in reef shark abundance and composition in Chagos.
Trends in Reef Shark Relative Abundances 1975–2012
Reef sharks are some of the most important apex predators in coral reef ecosystems, with evidence of their presence influencing trophic structure (DeMartini et al. 2008), prey fish demographics (Ruttenberg et al. 2011) and prey fish behaviour (Madin et al. 2010). Many species of sharks are vulnerable to over-exploitation due to their ‘slow’ life history traits, including late age at maturity, large body size, and low fecundity with a long gestation period (Reynolds et al. 2001). As such, reported declines in shark abundances has increased over the past two decades, in conjunction with increases in the trade for shark fin (Fong and Anderson 2002; Clarke et al. 2007; Dulvy et al. 2008). For example, spatial comparisons between remote coral reef locations or no-go preservation areas and more heavily used reef areas indicate substantial differences in shark abundances (Robbins et al. 2006, Stevenson et al. 2007; DeMartini et al. 2008; Sandin et al. 2008; Hisano et al. 2011).
The Chagos archipelago is not immune to the effects of shark fishing. Indeed, reef sharks are one of the only reef associated fish groups that have been heavily targeted (Sheppard et al. 2012). Aside from the small Mauritian reef fishery pre-2010, there was a licensed blue-water fishery in the territory largely for tuna, which stopped with the creation of the no-take marine protected area in 2010. Between 2006 and 2010 all licensed fishing vessels had to declare the quantities of shark fin bycatch and other products on board upon inspection by the Chagos fishery patrol vessel and in log books. Furthermore, shark finning was banned in 2006 and use of wire trace on longlines has been banned in the consolidated fisheries ordinance since 1999, with an associated penalty of £100,000 (Anon 2007). The majority of the fishing for reef associated sharks is thought to be by illegal vessels principally from Sri Lanka (Anderson et al. 1998). The number of Sri Lankan fishing vessels detected in the archipelago between 2002 and 2009 ranged from 3 to 26 per year, with number of arrests ranging from 1 to 8 (Graham et al. 2010). Vessels arrested usually have a hold full of sharks, retaining both the carcass and the fins.
The impacts of illegal fishing on the reef shark populations in Chagos has been well documented, with a 90% decline in the number of sharks observed per scientific dive between the 1970s and 1996 (Anderson et al. 1998), a small increase by 2001 (Spalding 2003) and a number similar to 1996 observed in 2006 (Graham et al. 2010). Grey reef sharks consistently made up the majority of the sharks observed across the entire sampling period (Fig. 19.10). Between 1996 and 2006 there was a small decline in the relative number of tawny nurse sharks and an increase in the proportion of silvertip sharks seen (Graham et al. 2010).

A grey reef shark, Carcharhinus amblyrhynchos, which numerically dominate the reef shark assemblages of the Chagos Archipelago as observed by divers (Photo: N Graham)
We update earlier assessments of relative shark abundance by including data collected in the 2010 and 2012 Chagos expeditions. The earlier data (1975–1996) collected by Charles Anderson, Ron Crosby, Charles Sheppard and Mark Spalding was collated and compiled by Charles Anderson to investigate changes in shark abundance in the archipelago through time (Anderson et al. 1998). This was updated with data from 2001 collected by Mark Spalding (Spalding 2003), and data collected by Nick Graham in 2006 (Graham et al. 2010). Data presented here continue this time series and were collected by Nick Graham on the 2010 and 2012 expeditions. All observations were made on the outer reef slopes of Salomon, Peros Banhos and the Eagle and Brothers group of the Great Chagos Bank. Dives were typically ∼1 h in duration and between 5 m and 25 m depth. From 1996 onwards the species of shark was also recorded. Although density cannot be estimated with these observational data, they are thought to provide reliable data on the relative abundance of sharks among years (Anderson et al. 1998).
The number of sharks seen per dive declined from 4.2 in the 1970s to 0.7 by 1996, increased in 2001 at 1.4 sharks per dive and declined again to 0.4 by 2006 (Anderson et al. 1998; Spalding 2003; Graham et al. 2010). Updating these data to include observations from 2010 and 2012 provides cause for some cautious optimism. The number of sharks seen per dive in 2010 rose to 1.5 and in 2012 an average of 1.0 shark was seen per dive (Fig. 19.11). These numbers are still fluctuating, and are a long way from the values recorded in the 1970s, however it is positive to see the very low numbers in 1996 and 2006 have not been repeated. It should be noted that the 95% confidence intervals around the 2010 and 2012 estimates are relatively large. Observational data of this kind are inherently variable, and in 2010 and 2012 up to 4 and 5 sharks were seen on individual dives respectively, while other dives recorded zero sharks. Continuing this time series will be important to see if these higher abundances are maintained or improved upon in the future.

Grey reef sharks continue to dominate the assemblage on the outer reef slopes in terms of proportional abundance (Fig. 19.12). The substantial contribution of silvertip sharks seen in 2006 (Graham et al. 2010), did not continue into 2010 and 2012, although the proportion did remain higher than in 1996 and 2001 (Fig. 19.12). Tawny nurse sharks were recorded in similar proportion to that seen in 1996 and 2001. Interestingly, blacktip reef sharks and, in particular, whitetip reef sharks that were absent from counts in 2006 and 2010, were recorded in 2012. It is positive to see all five species of shark being represented in the most recent surveys, again offering cautious optimism.

Relative species-level contribution to shark assemblages seen per scientific dive in the Chagos Archipelago, 1996–2012. Triaenodon obesus: whitetip reef shark; Carcharhinus melanopterus: blacktip reef shark; Carcharhinus albimarginatus: silvertip shark; Carcharhinus amblyrhynchos: grey reef shark; Nebrius ferrugineus: tawny nurse shark (Updated from Spalding 2003 and Graham et al. 2010)
Illegal vessels with sharks on board are still being detected in Chagos (N Graham pers obs), and fishes with slow life histories such as sharks are typically depleted far more rapidly than they recover. It is therefore important that enforcement continues to develop and employ new technologies that will improve the protection of shark populations in the no-take area, and that this is augmented with other multi-faceted conservation measures (Graham et al. 2010). If the no-take area can be successfully protected, there is every reason to believe that given sufficient time shark populations in Chagos can recover to densities recorded in the 1970s. Given the exceedingly high fish biomass results given above, it will be interesting to see what added contribution to these values recovering shark populations make, or if they reduce the biomass of lower trophic levels through predation.
Habitat Degradation Effects on Reef Fishes in Chagos
One of the greatest threats to reef fish assemblages, other than fishing, is habitat degradation and loss. Substantial changes in coral cover have been documented through time in all coral reef regions of the world, with considerable ongoing declines in the Caribbean (e.g., Gardner et al. 2003) and Indo-west Pacific (e.g., Bruno and Selig 2007). Much of this change is associated with pulse disturbances, such as coral bleaching, crown-of-thorns starfish outbreaks, and tropical storms (Bruno and Selig 2007). Similarly, there was marked decline in coral cover across the Indian Ocean due to the 1998 thermal anomaly followed by some recovery (Ateweberhan et al. 2011). There is now substantial evidence that such habitat degradation has substantial negative effects on the abundance of many trophic groups of fish, on reef fish diversity, and can lead to local extinctions of highly specialized species (Wilson et al. 2006; Pratchett et al. 2008, 2011). Interestingly, the suite of species highly vulnerable to population declines following habitat degradation is different to the suite of species vulnerable to fishing (Wilson et al. 2008b, 2010; Graham et al. 2011b). While large bodied, often piscivorous species are typically most vulnerable to fishing, small bodied species that use live coral or the structure it provides for shelter, settlement or food are most vulnerable to habitat loss (Graham et al. 2011b). This is a concern, because both fishing and habitat degradation occur together on many of the world’s coral reefs, effectively resulting in complementary effects that greatly extend the range of fishes facing significant population declines. We have ascertained above that fishing is having a negligible impact on reef fish communities in the northern atolls of Chagos. However, it is imperative to also assess if the reef fish assemblages are under threat from habitat change.
The greatest impact to coral cover and associated reef habitat in Chagos since scientific study of the reefs began was the 1998 thermal anomaly (Sheppard 1999). This thermal anomaly had severe, but variable, impacts on reefs across the western Indian Ocean. In order to assess the effects of this large scale habitat disturbance to reef fish assemblages, a large-scale study identified all the surveys of reef condition and associated fish assemblage structure from the Indian Ocean region conducted in the mid-1990s before the thermal anomaly and repeated these surveys using identical methods in 2005 and 2006 after the thermal anomaly. Analyses of these data showed that at locations where coral and associated structure had been lost, fish species richness, abundance of fish that feed on coral or plankton, and fish with a maximum body size less than 20 cm declined (Graham et al. 2008; MacNeil and Graham 2010). Chagos was interesting, because although coral mortality in 1998 had been up to 80% at many sites (Sheppard 1999), recovery of coral cover by 2006 was already substantial (Graham et al. 2008; Sheppard et al. 2008). Fish species richness showed very little change after the thermal anomaly in Chagos (Fig. 19.13a), although abundance of coral feeding fish (corallivores) declined in proportion with the amount of coral that was lost across the region (Graham et al. 2008). The extent of corallivore decline was however less severe than many other locations in the western Indian Ocean, largely because coral cover had recovered substantially in Chagos by the 2006 repeat surveys (Graham et al. 2009; Fig. 19.13b).

Change in (a) Reef fish species richness and (b) The abundance of corallivores, across seven countries in the Indian Ocean before and after the 1998 coral mortality event. Chagos sites represented by red circles, and all other data as blue squares (Adapted from Graham et al. (2008))
The corallivores are one of the most vulnerable fish groups to coral loss worldwide (Pratchett et al. 2008). Of particular interest is the degree of specialization among different species of butterflyfish. The family can be crudely broken into non-coral feeders, facultative coral feeders (i.e. those species that feed on coral, but will also feed on benthic algae and mobile invertebrates), and obligate corallivores (i.e. those species that feed exclusively on corals) (e.g. Pratchett 2005; Fig. 19.14). The extent to which abundances decline following coral mortality varies greatly among these classifications, with obligate specialists showing the greatest population losses (Pratchett et al. 2006; Graham 2007). If the measure of specialization is improved to include the number of coral species fed upon by different species of butterflyfishes, the importance of dietary specialization becomes even more evident. For example, at Lizard Island on the Great Barrier Reef, Australia, a gradient in specialization exists among 10 obligate coral feeding butterflyfish from a species that will only feed on a subset of less than 10 species of corals, to a more generalist feeder that will consume tissue from over 50 species of coral. Following a crown-of-thorns starfish outbreak that caused extensive loss of live coral cover at Lizard Island, the most specialized butterflyfish declined by ∼80%, while the more generalist feeder only declined by ∼20% (Pratchett 2001; Pratchett et al. 2008).

Butterflyfish from the Chagos Archipelago. (a) Chaetodon meyeri are obligate coral feeders, but are known to feed on at least 24 coral genera in Chagos (Pratchett et al. in press). (b) Two Chaetodon auriga and a Chaetodon kleinii from the Chagos Archipelago. Both species are facultative coral feeders, taking between 10% and 30% of their bites from corals in Chagos (Pratchett et al. in press) (Photo’s: M Pratchett)
Interestingly, the extent of decline in corallivores in Chagos after the 1998 thermal anomaly was not related to degree of specialization (Graham 2007; Graham et al. 2009). This was perhaps due to the rapid recovery of coral on reefs in Chagos masking any effects. However, from a range of studies worldwide, it is clear that the degree of dietary specialization confers a significant disadvantage to coral feeding fishes in the face of coral loss and increasing habitat disturbances (Lawton et al. 2011). The obvious question, therefore, is what is the advantage of such dietary specialization. Noting that coral cover had recovered to pre-1998 levels by the 2010 Chagos expedition (Sheppard et al. 2012), M. Pratchett quantified the degree of feeding specialization of butterflyfish at the archipelago to compare to their patterns of numerical dominance in the assemblage. Interestingly, there was a very strong positive relationship between feeding specialization and abundance, with the most specialized being most abundant (Pratchett et al. in press). Such information from a unique and relatively undisturbed location such as Chagos provides a starting point to understanding why some species are so specialized. Specialization may have evolved to provide a competitive advantage, perhaps associated with the quality of the tissue of different corals (Graham 2007), during long geological periods of relatively stable conditions on coral reefs (Pandolfi et al. 2011). However, specialized feeding on corals is likely to confer a significant disadvantage as coral cover declines due to the effects of climate change and human impacts.
Dire predictions for the future of coral dominated ecosystems under global climate change suggest that if carbon emissions continue under a business as usual scenario, reefs may be functionally lost by the middle of the century (Sheppard 2003; Hoegh-Guldberg et al. 2007; Veron et al. 2009). However, it is also emerging that different species of reef building corals and fishes are differentially vulnerable to the effects of climate change and other disturbances (McClanahan et al. 2007b; Graham et al. 2011b; Pandolfi et al. 2011), and composition of coral reef communities may change substantially rather than be lost altogether. Coral reefs will also have a much better chance of bouncing back from disturbances and coping with a certain degree of climatic warming if local human derived stresses can be greatly reduced (Hughes et al. 2010). Chagos is an excellent example, being one of the few locations in the Indian Ocean that is largely free of human impact, and one of the few that has recovered rapidly from the 1998 thermal anomaly (Sheppard et al. 2012). It is highly likely that large remote locations, such as Chagos, will be some of the longest lasting and best hopes for the future of coral dominated ecosystems as our climate continues to change.
Conclusion
The rapid recovery of coral cover at Chagos following the 1998 thermal anomaly, the stability of the fish assemblages through this event and the exceptionally high reef fish biomass on the reefs all provide compelling evidence of the unique status of the coral reefs of Chagos. The archipelago provides an example of community structure that most reefs likely resembled in the past, and also demonstrates the considerable resilience of reef ecosystems that are largely devoid of chronic anthropogenic pressures. It also indicates the high sensitivity of the ecosystem to even light recreational fishing. Such a location is not only useful as a reference point to compare with other reefs and management strategies globally (Knowlton and Jackson 2008), but is also likely to act as a large reservoir of diversity as climate change and other human pressures interact and cause uncertain futures for coral reef ecosystems. Chagos is not immune to all human impacts, and the reduced fish biomass around Diego Garcia and shark abundances testament. The recent establishment of the no-take marine protected area around Chagos can facilitate a move towards improved enforcement, appropriate international policies and engagement with fishers and consumers to help reverse trends in reef shark abundance and promote recovery of their populations. Similarly, better monitoring and regulations of any continuing recreational fishery around Diego Garcia may help enhance the reef fish biomass of that atoll towards the status of the northern atolls.
References
Anderson C, Sheppard C, Spalding M, Crosby R (1998) Shortage of sharks at Chagos. Shark News 10:1–12
Anon (2007) The fisheries (Conservation and management) ordinance 2007. British Indian Ocean Territory Administration, London
Ateweberhan M, McClanahan TR, Graham NAJ, Sheppard C (2011) Episodic heterogeneous decline and recovery of coral cover in the Western Indian Ocean. Coral Reefs 30:739–752
Bellwood DR, Hughes TP, Folke C, Nyström M (2004) Confronting the coral reef crisis. Nature 429:827–833
Bruno JF, Selig ER (2007) Regional decline of coral cover in the Indo-Pacific: timing, extent, and subregional comparisons. PLoS One 2:e711
Cinner JE, McClanahan TR, Daw TM, Graham NAJ, Maina J, Wilson SK, Hughes TP (2009) Linking social and ecological systems to sustain coral reef fisheries. Curr Biol 19:206–212
Cinner JE, Graham NAJ, Huchery C, MacNeil MA (2013) Global impacts of local human population density and distance to markets on the condition of coral reef fisheries. Conserv Biol doi:10.1111/j.1523-1739.2012.01933.x
Clarke SC, Milner-Gulland EJ, Bjørndal T (2007) Perspective: social, economic and regulatory drivers of the shark fin trade. Mar Resour Econ 22:305–327
Coleman FC, Figueira WF, Ueland JS, Crowder LB (2004) The impact of United States recreational fisheries on marine fish population. Science 305:1958–1960
Daw T, Cinner JE, McClanahan TR, Graham NAJ, Wilson SK (2011) Design factors and socioeconomic variables associated with ecological responses to fishery closures in the western Indian Ocean. Coast Manage 39:412–424
DeMartini EE, Friedlander AM, Sandin SA, Sala E (2008) Differences in fish-assemblage structure between fished and unfished atolls in the northern Line Islands, central Pacific. Mar Ecol Prog Ser 365:199–215
Dulvy NK, Polunin NVC, Mill AC, Graham NAJ (2004) Size structural change in lightly exploited coral reef fish communities: evidence for week indirect effects. Can J Fish Aquat Sci 61:466–475
Dulvy NK, Baum JK, Clarke S, Compagno LJV, Cortes E, Domingo A, Fordham S, Flowler S, Francis MP, Gibson C, Martinez J, Musick JA, Soldo A, Stevens JD, Valenti S (2008) You can swim but can’t hide: the global status and conservation of oceanic pelagic sharks and rays. Aquat Conserv Mar Freshwat Ecosyst 18:459–482
Estes JA, Terborgh J, Brashares JS, Power ME, Berger J, Bond WJ, Carpenter SR, Essington TE, Holt RD, Jackson JBC (2011) Trophic downgrading of planet earth. Science 333:301
Fong QSW, Anderson JL (2002) International shark fin markets and shark management: an integrated market preference–cohort analysis of the blacktip shark (Carcharhinus limbatus). Ecol Econ 40:117–130
Friedlander AM, DeMartini EE (2002) Contrasts in density, size, and biomass of reef fishes between the northwestern and the main Hawaiian Islands: the effects of fishing down apex predators. Mar Ecol Prog Ser 230:253–264
Froese R, Pauly D (2012) FishBase (www.fishbase.org). World wide web electronic publication
Gardner TA, Côté IM, Gill JA, Grant A, Watkinson AR (2003) Long-term region-wide declines in Caribbean corals. Science 301:958–960
Graham NAJ (2007) Ecological versatility and the decline of coral feeding fishes following climate driven coral mortality. Mar Biol 153:119–127
Graham NAJ, McClanahan TR (in press) The last call for marine wilderness? BioScience
Graham NAJ, Dulvy NK, Jennings S, Polunin NVC (2005) Size-spectra as indicators of the effects of fishing on coral reef fish assemblages. Coral Reefs 24:118–124
Graham NAJ, Wilson SK, Jennings S, Polunin NVC, Bijoux J, Robinson J (2006) Dynamic fragility of oceanic coral reef ecosystems. Proc Natl Acad Sci USA 103:8425–8429
Graham NAJ, Wilson SK, Jennings S, Polunin NVC, Robinson J, Bijoux JP, Daw TM (2007) Lag effects in the impacts of mass coral bleaching on coral reef fish, fisheries, and ecosystems. Conserv Biol 21:1291–1300
Graham NAJ, McClanahan TR, MacNeil MA, Wilson SK, Polunin NVC, Jennings S, Chabanet P, Clark S, Spalding MD, Letourneur Y, Bigot L, Galzin R, Öhman MC, Garpe KC, Edwards AJ, Sheppard CRC (2008) Climate warming, marine protected areas and the ocean-scale integrity of coral reef ecosystems. PLoS One 3:e3039
Graham NAJ, Wilson S, Pratchett M, Polunin N, Spalding M (2009) Coral mortality versus structural collapse as drivers of corallivorous butterflyfish decline. Biodivers Conserv 18:3325–3336
Graham NAJ, Spalding MD, Sheppard CRC (2010) Reef shark declines in remote atolls highlight the need for multi-faceted conservation action. Aquat Conserv Mar Freshwat Ecosyst 20:543–548
Graham NAJ, Ainsworth TD, Baird AH, Ban NC, Bay LK, Cinner JE, De Freitas DM, Diaz-Pulido G, Dornelas M, Dunn SR, Fidelman PIJ, Foret S, Good TC, Kool J, Mallela J, Penin L, Pratchett MS, Williamson DH (2011a) From microbes to people: tractable benefits of no-take areas for coral reefs. Oceanogr Mar Biol Annu Rev 49:105–136
Graham NAJ, Chabanet P, Evans RD, Jennings S, Letourneur Y, MacNeil MA, McClanahan TR, Ohman MC, Polunin NVC, Wilson SK (2011b) Extinction vulnerability of coral reef fishes. Ecol Lett 14:341–348
Heupel MR, Simpfendorfer CA, Fitzpatrick R (2010) Large-scale movement and fidelity of grey reef sharks. PLoS One 5:e9650
Hisano M, Connolly SR, Robbins WD (2011) Population growth rates of reef sharks with and without fishing on the Great Barrier Reef: robust estimation with multiple models. PLoS One 6:e25028
Hixon MA (1991) Predation as a process structuring coral reef fish communities. In: Sale PF (ed) The ecology of fishes on coral reefs. Academic Press, New York, pp 475–508
Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS, Greenfield P, Gomez E, Harvell CD, Sale PF, Edwards AJ, Caldeira K, Knowlton N, Eakin CM, Iglesias-Prieto R, Muthiga N, Bradbury RH, Dubi A, Hatziolos ME (2007) Coral reefs under rapid climate change and ocean acidification. Science 318:1737–1742
Hughes TP, Graham NAJ, Jackson JBC, Mumby PJ, Steneck RS (2010) Rising to the challenge of sustaining coral reef resilience. Trends Ecol Evol 25:633–642
Jackson JBC, Kirby MX, Berger WH, Botsford LW, Bourque BJ, Bradbury RH, Cooke R, Erlandson J, Estes JA, Hughes TP, Kidwell S, Lange CB, Lenihan HS, Pandolfi JM, Peterson CH, Steneck RS, Tegner MJ, Warner RR (2001) Historical overfishing and the recent collapse of coastal ecosystems. Science 293:629–637
Jennings S, Polunin NVC (1996) Effects of fishing effort and catch rate upon the structure and biomass of Fijian reef fish communities. J Appl Ecol 33:400–412
Jennings S, Polunin NVC (1997) Impacts of predator depletion by fishing on the biomass and diversity of non-target reef fish communities. Coral Reefs 16:71–82
Jones GP, Syms C (1998) Disturbance, habitat structure and the ecology of fishes on coral reefs. Aust J Ecol 23:287–297
Jones GP, Caley MJ, Munday PL (2002) Rarity in coral reef fish communities. In: Sale PF (ed) Coral reef fishes dynamics and diversity in a complex ecosystem. Academic, San Diego, pp 81–102
Jones GP, McCormick MI, Srinivasan M, Eagle JV (2004) Coral decline threatens fish biodiversity in marine reserves. Proc Natl Acad Sci USA 101:8251–8258
Knowlton N, Jackson JBC (2008) Shifting baselines, local impacts, and global change on coral reefs. PLoS Biol 6:e54
Lawton RJ, Messmer V, Pratchett MS, Bay LK (2011) High gene flow across large geographic scales reduces extinction risk for a highly specialised coral feeding butterflyfish. Mol Ecol 20:3584–3598
MacNeil MA, Graham NAJ (2010) Enabling regional management in a changing climate through Bayesian meta-analysis of a large-scale disturbance. Glob Ecol Biogeogr 19:412–421
Madin EMP, Gaines SD, Warner RR (2010) Field evidence for pervasive indirect effects of fishing on prey foraging behavior. Ecology 91:3563–3571
McClanahan TR (1994) Kenyan coral reef lagoon fish: effects of fishing, substrate complexity, and sea urchins. Coral Reefs 13:231–241
McClanahan TR (2000) Recovery of a coral reef keystone predator, Balistapus undulatus, in East African marine parks. Biol Conserv 94:191–198
McClanahan TR (2008) Response of the coral reef benthos and herbivory to fishery closure management and the 1998 ENSO disturbance. Oecologia 155:169–177
McClanahan TR (2011) Coral reef fish communities in management systems with unregulated fishing and small fisheries closures compared with lightly fished reefs – Maldives vs Kenya. Aquat Conserv Mar Freshwat Ecosyst 21:186–198
McClanahan TR, Graham NAJ (2005) Recovery trajectories of coral reef fish assemblages within Kenyan marine protected areas. Mar Ecol Prog Ser 294:241–248
McClanahan TR, Humphries A (2012) Differential and slow life history responses of fishes to coral reef closures. Mar Ecol Prog Ser 469:121–131
McClanahan TR, Omukoto JO (2011) Implications of modern versus historical fisheries catches for marine protected area goals. Conserv Biol 25:945–955
McClanahan TR, Graham NAJ, Calnan JM, MacNeil MA (2007a) Toward pristine biomass: reef fish recovery in coral reef marine protected areas in Kenya. Ecol Appl 17:1055–1067
McClanahan TR, Ateweberhan M, Sebastian CR, Graham NAJ, Wilson SK, Guillaume MMM, Bruggemann JH (2007b) Western Indian Ocean coral communities: bleaching responses and susceptibility to extinction. Mar Ecol Prog Ser 337:1–13
McClanahan TR, Hicks CC, Darling SE (2008) Malthusian overfishing and efforts to overcome it on Kenyan coral reefs. Ecol Appl 18:1516–1529
McClanahan TR, Graham NAJ, Wilson SK, Letourneur Y, Fisher R (2009) Effects of fisheries closure size, age, and history of compliance on coral reef fish communities in the western Indian Ocean. Mar Ecol Prog Ser 396:99–109
McClanahan TR, Graham NAJ, MacNeil MA, Muthiga NA, Cinner JE, Bruggemann JH, Wilson SK (2011) Critical thresholds and tangible targets for ecosystem-based management of coral reef fisheries. Proc Natl Acad Sci USA 108:17230–17233
Mees CC (1996) The mauritian banks fishery: a review and spatial analysis. Technical report. Marine Resources Assessment Group London
Mora C, Aburto-Oropeza O, Ayala Bocos A, Ayotte PM, Banks S, Bauman AG, Beger M, Bessudo S, Booth DJ, Brokovich E, Brooks A, Chabanet P, Cinner JE, Cortés J, Cruz-Motta JJ, Cupul Magaña A, DeMartini EE, Edgar GJ, Feary DA, Ferse SCA, Friedlander AM, Gaston KJ, Gough C, Graham NAJ, Green A, Guzman H, Hardt M, Kulbicki M, Letourneur Y, López Pérez A, Loreau M, Loya Y, Martinez C, Mascareñas-Osorio I, Morove T, Nadon M-O, Nakamura Y, Paredes G, Polunin NVC, Pratchett MS, Reyes Bonilla H, Rivera F, Sala E, Sandin SA, Soler G, Stuart-Smith R, Tessier E, Tittensor DP, Tupper M, Usseglio P, Vigliola L, Wantiez L, Williams I, Wilson SK, Zapata FA (2011) Global human footprint on the linkage between biodiversity and ecosystem functioning in reef fishes. PLoS Biol 9:e1000606
Munday PL (2004) Habitat loss, resource specialization, and extinction on coral reefs. Glob Change Biol 2004:1–6
Nadon MO, Baum JK, Williams ID, McPherson JM, Zgliczynski BJ, Richards BL, Schroeder RE, Brainard RE (2012) Re-creating missing population baselines for Pacific reef sharks. Recreación de las Líneas de Base Poblacionales Faltantes para Tiburones de Arrecife en el Pacífico. Conserv Biol 26:493–503
Newman MJH, Paredes GA, Sala E, Jackson JBC (2006) Structure of Caribbean coral reef communities across large gradient of fish biomass. Ecol Lett 9:1216–1227
Olden JD, Hogan ZS, Zanden MJV (2007) Small fish, big fish, red fish, blue fish: size-biased extinction risk of the world’s freshwater and marine fishes. Glob Ecol Biogeogr 16:694–701
Pandolfi JM, Connolly SR, Marshall DJ, Cohen AL (2011) Projecting coral reef futures under global warming and ocean acidification. Science 333:418–422
Pitcher TJ (2001) Fisheries managed to rebuild ecosystems? Reconstructing the past to salvage the future. Ecol Appl 11:601–617
Pollnac R, Christie P, Cinner JE, Dalton T, Daw TM, Forrester GE, Graham NAJ, McClanahan TR (2010) Marine reserves as linked social-ecological systems. Proc Natl Acad Sci USA 107:18262–18265
Polunin NVC, Robert CM (eds) (1996) Reef fisheries. Chapman & Hall, London
Pratchett MS (2001) Dynamics of outbreak populations of crown-of-thorns starfish (Acanthaster planci L.), and their effects on coral reef ecosystems. Ph.D. thesis, James Cook University
Pratchett MS (2005) Dietary overlap among coral-feeding butterflyfishes (Chaetontidae) at Lizard Island, northern Great Barrier Reef. Mar Biol 148:373–382
Pratchett MS, Graham NAJ, Cole AJ (in press) Specialist corallivores dominate butterflyfish assemblages in the Chagos Archipelago, central Indian Ocean. J Fish Biol
Pratchett MS, Wilson SK, Baird AH (2006) Declines in the abundance of Chaetodon butterflyfishes following extensive coral depletion. J Fish Biol 69:1–12
Pratchett MS, Munday PL, Wilson SK, Graham NAJ, Cinner JE, Bellwood DR, Jones GP, Polunin NVC, McClanahan TR (2008) Effects of climate-induced coral bleaching on coral-reef fishes: ecological and economic consequences. Oceanogr Mar Biol Annu Rev 46:251–296
Pratchett MS, Hoey AS, Wilson SK, Messmer V, Graham NAJ (2011) Changes in biodiversity and functioning of reef fish assemblages following coral bleaching and coral loss. Diversity 3:424–452
Pratchett MS, Coker DJ, Jones GP, Munday PL (2012) Specialization in habitat use by coral reef damselfishes and their susceptibility to habitat loss. Ecol Evol 2:2168–2180
Reynolds JD, Jennings S, Dulvy NK (2001) Life histories of fishes and population responses to exploitation. In: Reynolds JD, Mace GM, Redford KH, Robinson JG (eds) Conservation of exploited species. Cambridge University Press, Cambridge, pp 47–168
Robbins WD, Hisano M, Connolly SR, Choat JH (2006) Ongoing collapse of coral-reef shark population. Curr Biol 16:2314–2319
Russ GR, Alcala AC (1989) Effects of intense fishing pressure on an assemblage of coral reef fishes. Mar Ecol Prog Ser 56:13–27
Russ GR, Stockwell B, Alcala AC (2005) Inferring versus measuring rates of recovery in no-take marine reserves. Mar Ecol Prog Ser 292:1–12
Ruttenberg BI, Hamilton SL, Walsh SM, Donovan MK, Friedlander A, DeMartini E, Sala E, Sandin SA (2011) Predator-induced demographic shifts in coral reef fish assemblages. PLoS One 6:e21062
Sandin SA, Smith JE, DeMartini EE, Dinsdale EA, Donner SD, Friedlander AM, Konotchick T, Malay M, Maragos JE, Obura D, Pantos O, Paulay G, Richie M, Rohwer F, Schroeder RE, Walsh S, Jackson JBC, Knowlton N, Sala E (2008) Baselines and degradation of coral reefs in the Northern Line Islands. PLoS One 3:e1548
Sheppard CRC (1999) Coral decline and weather patterns over 20 years in the Chagos Archipelago, Central Indian Ocean. Ambio 28:472–478
Sheppard CRC (2003) Predicted recurrences of mass coral mortality in the Indian Ocean. Nature 425:294–297
Sheppard CRC, Harris A, Sheppard ALS (2008) Archipelago-wide coral recovery patterns since 1998 in the Chagos Archipelago, central Indian Ocean. Mar Ecol Prog Ser 362:109–117
Sheppard CRC, Ateweberhan M, Bowen BW, Carr P, Chen CA, Clubbe C, Craig MT, Ebinghaus R, Eble J, Fitzsimmons N, Gaither MR, Gan CH, Gollock M, Guzman N, Graham NAJ, Harris A, Jones R, Keshavmurthy S, Koldewey H, Lundin CG, Mortimer JA, Obura D, Pfeiffer M, Price ARG, Purkis S, Raines P, Readman JW, Riegl B, Rogers A, Schleyer M, Seaward MRD, Sheppard ALS, Tamelander J, Turner JR, Visram S, Vogler C, Vogt S, Wolschke H, Yang JMC, Yang SY, Yesson C (2012) Reefs and islands of the Chagos Archipelago, Indian Ocean: why it is the world’s largest no-take marine protected area. Aquat Conserv Mar Freshwat Ecosyst 22:232–261
Spalding MD (1999) Biodiversity patterns in Chagos reef fishes. In: Sheppard CRC, Seaward MRD (eds) The ecology of the Chagos Archipelago. Westbury Publishing, Great Britain, pp 119–136
Spalding MD (2003) Partial recovery of sharks in the Chagos Achipelago. Shark News 15:12–13
Stevenson C, Katz LS, Micheli F, Block B, Heiman KW, Perle C, Weng K, Dunbar R, Witting J (2007) High apex predator biomass on remote Pacific Island. Coral Reefs 26:47–51
Stockwell B, Jadloc CRL, Abesamis RA, Alcala AC, Russ GR (2009) Trophic and benthic responses to no-take marine reserve protection in the Philippines. Mar Ecol Prog Ser 389:1–15
Veron JEN, Hoegh-Guldberg O, Lenton TM, Lough JM, Obura DO, Pearce-Kelly P, Sheppard CRC, Spalding M, Stafford-Smith MG, Rogers AD (2009) The coral reef crisis: the critical importance of 350 ppm CO2. Mar Pollut Bull 58:1428–1436
Williams ID, Polunin NVC (2000) Differences between protected and unprotected reefs of the western Caribbean in attributes preferred by dive tourists. Environ Conserv 27:382–391
Williams ID, Richards BL, Sandin SA, Baum JK, Schroeder RE, Nadon MO, Zgliczynski B, Craig P, McIlwain JL, Brainard RE (2011) Differences in reef fish assemblages between populated and remote reefs spanning multiple Archipelagos across the Central and Western Pacific. J Mar Biol 2011:1–14, Article ID 826234
Wilson SK, Graham NAJ, Pratchett MS, Jones GP, Polunin NVC (2006) Multiple disturbances and the global degradation of coral reefs: are reef fishes at risk or resilient? Glob Change Biol 12:2220–2234
Wilson SK, Graham NAJ, Polunin NVC (2007) Appraisal of visual assessments of habitat complexity and benthic composition on coral reefs. Mar Biol 151:1069–1076
Wilson SK, Burgess SC, Cheal AJ, Emslie M, Fisher R, Miller I, Polunin NVC, Sweatman HPA (2008a) Habitat utilization by coral reef fish: implications for specialists vs. generalists in a changing environment. J Anim Ecol 77:220–228
Wilson SK, Fisher R, Pratchett MS, Graham NAJ, Dulvy NK, Turner RA, Cakacaka A, Polunin NVC, Rushton SP (2008b) Exploitation and habitat degradation as agents of change within coral reef fish communities. Glob Change Biol 14:2796–2809
Wilson SK, Fisher R, Pratchet MS, Graham NAJ, Dulvy NK, Turner RA, Cakacaka A, Polunin NVC (2010) Habitat degradation and fishing effects on the size structure of coral reef fish communities. Ecol Appl 20:442–451
Wilson SK, Graham NAJ, Fisher R, Robinson JI, Nash K, Chong-Seng K, Polunin NVC, Aumeeruddy R, Quatre R (2012) The effect of macroalgal expansion and marine protected areas on coral recovery following a climatic disturbance. Conserv Biol 26:995–1004
Winterbottom R, Anderson RC (1999) Fishes of the Chagos Archipelago. In: Sheppard CRC, Anderson RC (eds) Ecology of the Chagos Archipelago. Westbury Publishing, Great Britain, pp 101–117
Acknowledgements
Funding for this work was provided by the British Foreign and Commonwealth Office, the Leverhulme Trust, Fisheries Society of the British Isles, Western Indian Ocean Marine Science Association and research fellowships from the Australian Research Council and Queensland Smart Futures Fund. The research could not have happened without the organization of Chagos expeditions by Charles Sheppard and Heather Koldewey. Many thanks to Bob Long for help in the field around Diego Garcia and to Cindy Huchery for help with figures and analyses. The Wildlife Conservation Society through grants from the Western Indian Ocean Marine Science Association supported much of the larger western Indian Ocean data collection.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Graham, N.A.J., Pratchett, M.S., McClanahan, T.R., Wilson, S.K. (2013). The Status of Coral Reef Fish Assemblages in the Chagos Archipelago, with Implications for Protected Area Management and Climate Change. In: Sheppard, C. (eds) Coral Reefs of the United Kingdom Overseas Territories. Coral Reefs of the World, vol 4. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-5965-7_19
Download citation
DOI: https://doi.org/10.1007/978-94-007-5965-7_19
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-5964-0
Online ISBN: 978-94-007-5965-7
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)