
Abstract
This chapter reviews the present knowledge and previous developments concerning the pollen transport in the atmosphere. Numerous studies are classified according to the spatial scales of the applications, key processes considered, and the methodology involved. Space-wise, local, regional and long-range scales are distinguished. An attempt of systematization is made towards the key processes responsible for the observed patterns: initial dispersion of pollen grains in the nearest vicinity of the sources at micro-scale, transport with the wind, mixing inside the atmospheric boundary layer and dry and wet removal at the regional scale, and the long-range dispersion with synoptic-scale wind, exchange between the boundary layer and free troposphere, roles of dry and wet removal, interactions with chemicals and solar radiation at the large scales.
Atmospheric dispersion modelling can pursue two goals: estimation of concentrations from known source (forward problem), and the source apportionment (inverse problem). Historically, the inverse applications were made first, mainly using the simple trajectory models. The sophisticated integrated systems capable of simulating all main processes of pollen lifecycle have been emerging only during last decade using experience of the atmospheric chemical composition modelling.
Several studies suggest the allergen existence in the atmosphere separately from the pollen grains – as observed in different parts of the world. However, there is no general understanding of the underlying processes, and the phenomenon itself is still debated. Another new area with strongly insufficient knowledge is the interactions of airborne allergens and chemical pollutants.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
5.1 Introduction
The atmospheric pathway is the fastest and the simplest way for biological agents to spread over terrestrial ecosystems. Many organisms can significantly increase the efficiency of their movements by taking advantage of air currents (Isard et al. 2005). Biota that is present in the atmosphere ranges in size from very small (viruses, bacteria, pollen and spores) to quite large (seeds, aphids, butterflies and moths, songbirds, and waterfowl) (Gage et al. 1999; Westbrook and Isard 1999). The link between these biological systems and the atmosphere is the key to understanding the population dynamics of and diseases spread by, aerobiota. Within this chapter, the emphasis is placed on identifying biologically and medically relevant temporal and spatial scales of atmospheric motions and meteorological parameters, which help control the abundance and distribution of airborne biota, such as pollen and other aeroallergens.
Biologically-relevant dispersion of bioaerosols affects the structure of ecosystems, since pollen is responsible for gene flow (Ellstrand 1992; Ennos 1994; Burczyck et al. 2004; Belmonte et al. 2008), and it contributes in determining the spatial distribution of plant species (Ellstrand 1992; Smouse et al. 2001; Sharma and Khanduri 2007; Schmidt-Lebuhn et al. 2007; Belmonte et al. 2008). Therefore, understanding the pollen gene dispersal is instrumental for the interpretation of the biogeographic range of plants and plant conservation issues. A review by Di-Giovanni and Kevan (1991) on the factors affecting the pollen dispersion in natural habitats can be recommended for the corresponding processes.
Apart from gene flow, the transport of bioaerosols causes concern because of its potential to distribute pathogens and allergens, which can affect human health, agriculture, and farming (Belmonte et al. 2000; Griffin et al. 2001a, b; Taylor 2002; Brown and Hovmoller 2002; Shinn et al. 2003; Garrison et al. 2003; Wu et al. 2004; Kellogg et al. 2004; Griffin 2007; Paz and Broza 2007; Polymenakou et al. 2008).
The pollen records from aerobiological monitoring sites have traditionally been interpreted as if the grains always originate from the local environment. Consequently, pollen forecasting tools have been designed by taking into account only local meteorological variables and phenological observations in the neighbourhood. This view is currently changing to acknowledge much broader bioaerosol movements, based on increasing evidence of pollen and spore transport at much greater distances, including continental (Belmonte et al. 2000,2008; Sofiev et al. 2006a; Siljamo et al. 2007, 2008b, c;Skjøth et al. 2009) and intercontinental scales (Prospero et al. 2005; Kellogg and Griffin 2006; Rousseau et al. 2008).
5.1.1 Basic Terms
5.1.1.1 Spatial and Temporal Scales
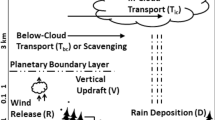
Pollen-related processes in the atmosphere take place at a wide range of scales. For the needs of the current chapter the following terms will be used following the classical definitions after Seinfeld and Pandis (2006) – Fig. 5.1:
-
Processes at the micro-scale are connected with pollen release and take place within a few metres from the plants
-
Local-scale processes include initial dispersion of grains that happen within the nearest kilometre from the source
-
Regional and meso-scales are considered as synonyms and cover the processes responsible for the dispersion and removal of the bulk of pollen grains – at distances of up to a hundred kilometres.
-
The hierarchy of the scales related to long-range transport consists of synoptic, continental, and global scales, which include processes of up to 1,000–2,000 km, up to 5,000 km, and over 5,000 km, respectively.
Connections between the above spatial and corresponding temporal scales are shown in Fig. 5.1.

Spatial and temporal scales of variability of the atmospheric constituents (Modified from Seinfeld and Pandis 2006)
Distinction between the scales is not always unequivocal and specific borders can vary depending on the specific application and criteria used. For the purposes of this review, we will also consider the cases as “local” or “regional” if the conditions at both source and the receptor points can be (roughly) described by a single or few observation stations. In the long-range transport case, such a description always requires many stations distributed over the area and atmospheric modelling as a way to evaluate the transport conditions.
5.1.1.2 Pollen Life Time in the Atmosphere
Atmospheric lifetime is the key parameter for each tracer, which has a direct connection to its spatial scale of distribution and temporal scale of its variations (Seinfeld and Pandis 2006). Several example species are marked in Fig. 5.1 following their characteristic lifetimes. With pollen and allergens, the situation is more complicated. Indeed, the pollen atmospheric lifetime of a few days (due to substantial gravitational sedimentation) defines it as a local-scale pollutant with some minor connection to regional scales (Sofiev et al. 2006a). However, as discussed below, in many cases the released amount is so large that the medical impact can be substantial at much larger distances – up to continental scales. This ambiguity is reflected in Fig. 5.1 where the pollen-related processes are delineated by a separate rectangular.
5.1.1.3 Aerobiological Phases of Pollen and Other Biogenic Aerosols in the Atmosphere
A schematic model for passively transported biological substances includes presentation of the biological material, its release in the atmosphere, dispersion, transformation, deposition, and impact (Fig. 5.2, extended from Isard and Gage 2001). Sometimes, after deposition, the grains can be subjected to refloating processes, thus being resuspended back into the air. In this chapter we will consider only release, transport, transformation, and deposition phases as the most important and studied so far.

Phases of aerobiological processes related to transport of chemically inactive biogenic aerosol (Extended from Isard and Gage 2001)
5.1.1.4 Types of Biogenic Aerosols
The most-studied biogenic aerosol is evidently the pollen grain. It transfers the male gametophyte to the female reproductive organs, which is termed as pollination process. Once pollen grains are deposited on the reproductive female organs the recognition process starts via protein exchange to allow the germination and liberation of the male gametes for fecundation. Several proteins inside the grain are considered to be allergens since, in some occasions, the human immune system can react on their presence by triggering allergenic reactions. Direct studies of these proteins in connection to atmospheric transport are very limited, therefore the main attention in this chapter will be given to pollen grains. The allergen-related studies will be included wherever possible. Other types of biogenic particulates, such as spores, seeds, etc., are beyond the scope of the review.
5.1.1.5 Main Features of Pollen and Allergen as Atmospheric Tracers
A pollen grain, from the point of view of the atmospheric transport, is a very large but comparatively light aerosol. Aerodynamic features of most of the pollen grains (except for the largest ones with diameter over 100 μm) allow the classical model considerations when the particle is considered to be embedded into all atmospheric flows including the small-scale turbulent eddies (Sofiev et al. 2006a). The life span of the particles in the atmosphere strongly depends on their deposition intensity, i.e. the sum of dry and wet deposition. For coarse particles, the gravitational settling is the most important deposition pathway (Seinfeld and Pandis 2006), which makes the sedimentation velocity the primary parameter deciding the atmospheric life time of pollen. For birch, it is about 1.2 cm sec−1 (Sofiev et al. 2006a), which corresponds to the lifetime ranging from a few tens of hours up to a couple of days depending on the vertical transport and mixing.
Pollen is not soluble in water but can easily be scavenged via impaction (in case of sub-cloud scavenging). Processes occurring inside the clouds are very poorly studied but one can expect that the grains can be embedded into the forming droplets and scavenged together with them.
A pollen allergen is usually a sub-micron aerosol with high water solubility (Vrtala et al. 1993; Taylor et al. 2004). It can therefore be scavenged with precipitation but has no substantial dry deposition velocity (see Chap. 19 of Seinfeld and Pandis 2006). As a result, one can expect the allergen to stay in air much longer than pollen – days and, possibly, weeks in the case of no precipitation.
Among the transformation processes occurring during the atmospheric transport, the most frequent one is a loss and gain of water depending on the air humidity, an ability of the pollen grain called harmomegathy (Wodehouse 1935). Sometimes this situation provokes the pollen grain rupture and allergen release. Secondly, it is a loss of viability due to temperature and UV radiation. Thirdly, a chemical damage of the grains by strong oxidants can take place in polluted environments. Finally, the release of the pollen content due to the grain rupture or (pseudo-) germination has been confirmed by several studies (Pacini et al. 2006; Motta et al. 2006; Taylor et al. 2002, 2004, 2006).
5.2 Meteorological Drivers of Pollen Dispersion in the Atmosphere
The pollen lifecycle in the atmosphere starts from the release from the anthers – arguably the smallest-scale relevant atmospheric process. In many anemophilous trees, the anther’s burst is a result of dehydration due to high temperature, solar exposure, low humidity and moderate wind (see Helbig et al. 2004; Linskens and Cresti 2000 and references therein). In other plants, floral parts, such as the filaments in grasses actively push the anthers into an exposed position, and anther’s dehiscence may be a result of passive desiccation or active reabsorption. Reabsorption enables the anther opening at any time of day, whereas evaporation may occur only during dry hours. The Asteraceae, which includes the genera of Artemisia and Ambrosia, is characterized by the secondary pollen presentation, which in this case means that the pollen mass is pushed to the top of the flower where it is exposed to atmospheric stress (von Wahl and Puls 1989; Kazlauskas et al. 2006). In this case, the pollen detachment from the capitulum can take place later than the anther opening. In particular, it could take place at any time during the day when the wind and turbulence are strong enough to pick up the pollen from the flower. In the Urticaceae, pollen emission is explosive and is caused by the spring-like release of the anthers caused by dehydration. A more detailed account of pollen release can be found in the Chap. 3 of this book and dedicated reviews, such as (Pacini and Hesse 2004).
Once the pollen grains are released into the atmosphere, the mechanical force of air flow, either induced by mean wind or by turbulent eddies, becomes the only process that keeps the grains in the air. The initial mixing and uplift driven by the turbulent motions largely determines the fraction of the released grains that will come to larger-scale dispersion (e.g. Gregory 1961). At the meso-scale, the mean wind becomes the main transport force while the turbulent mixing keeps the grains aloft and further redistributes them along the vertical – inside and beyond the atmospheric boundary layer. At regional and large scales, both horizontal and vertical mean-wind components are responsible for the transport on-par with turbulent mixing. At all scales, the processes of dry deposition, by impact or gravitational sedimentation, as well as wet deposition by scavenging with precipitation, are responsible for the removal of grains from the atmosphere. However, their importance varies being the highest at large scales.
5.3 Micro- and Local Scales
Micro- and local-scale transport of pollen grains includes the initial dispersion of pollen grains from the anthers and the transport over the first kilometre(s) from the source. An outlook of the main factors affecting the dispersion can be found in (Di-Giovanni and Kevan 1991). The knowledge about micro- to local scale pollen transport is based on field experiments with pollen release from a well-defined source. The dispersion of the pollen plume at these scales can be modelled with various types of dispersion models (Lagrangian, Gaussian plume model, large eddy simulation, or quasi-mechanistic models – e.g. Aylor et al. 2006; Arritt et al. 2007; Chamecki et al. 2009; Jarosz et al. 2003, 2004; Klein et al. 2003; Kuparinen 2006; Kuparinen et al. 2007; Schueler and Schlünzen 2006).
Experiments studying the dispersion and deposition of pollen were already made in the 1960s and 1970s by Raynor, Ogden and Hayes. For ragweed pollen, Raynor et al. (1970) used a set of point and area sources surrounded by four to five rings of samplers located concentrically from 1 to 69 m from the centre of the plot. The samples were taken at four heights from 0.5 to 4.6 m. It was found that about half of the ragweed pollen grains reaching the edge of the source are still airborne some 55–65 m further away. Extrapolation towards greater distances indicates that about 1% of the pollen grains remain airborne at 1 km. Raynor et al. (1970) state that since pollen clearly disperses similarly to other (small) particles, calculation of pollen dispersion by use of existing diffusion models should be practical. Further studies for Timothy grass and maize pollen were published by Raynor et al. (1972a,b).
Particular interest to the pollen transport was caused by the gene flow problem at local scales. Gleaves (1973) has developed a local-scale empirical gene-flow model and performed a series of sensitivity studies of the exchange efficiency of the genetic material depending on the mutual position of the plants, transport conditions, etc. A motivation for the study was that, despite “pollen can be blown hundreds of miles, the bulk of cross-pollination occurs over very short distances”. Therefore, the analysis has been performed for the transport range of a few tens of metres. Govindaraju (1988) has statistically demonstrated that the pollination mechanism directly influences the gene flow and showed that the dependence is the strongest for the wind-pollinating plants, being much weaker for animal- and self-pollinating species. Quantitative estimates for the tree species can be found in Govindaraju (1989).
Several recent field experiments have been designed to estimate the three-dimensional plume shape of airborne pollen grains, as well as the amount deposited. The outcome was also used for evaluation of the dispersion models. With numerous samplers placed around the source, instrument towers, and occasionally even remote-piloted planes and aircrafts, pollen concentrations up to several tens of meters above the ground were measured. The experiments for maize were made by Jarosz et al. (2003, 2004), Aylor et al. (2003, 2006), Boehm et al. (2008), Klein et al. (2003), Arritt et al. (2007); for ragweed pollen by Chamecki et al. (2009); and for oilseed rape pollen by McCartney and Lacey (1991).
The introduction of genetically modified organisms (GMO) in agriculture favoured the research of pollen transport processes on local scales for establishing a basis for assessment of the gene flow from these plants to other crops or to natural vegetation. An overview of these efforts is given by the European Environment Agency (2002). This report considers the significance of pollen-mediated gene flow from six major crops: oilseed rape, sugar beet, potatoes, maize, wheat and barley.
The EEA (2002) report confirmed that the majority of airborne pollen is deposited at very short distance from the pollen source, although out-crossing of maize pollen has been recorded at up to 800 m from the source and, in extreme cases, there is the evidence of wind transfer of oilseed rape pollen up to at least 1.5 km (Timmons et al. 1995). Studies of maize pollen dispersal from small plots (e.g. Raynor et al. 1972b; Jarosz et al. 2003) have suggested that there is relatively little impact of pollen beyond a few hundred meters from the source. However, the impact of pollen may extend to much greater distances because the dispersal distribution has a long extending tail (Aylor et al. 2003). The tail is expected to be much more evident for large source areas (Aylor et al. 2006). Brunet et al. (2004) used measurements taken by aircraft and found the viable pollen throughout the entire depth of the atmospheric boundary layer. Their results imply that updrafts due to turbulent eddies in the boundary layer overcome the terminal fall velocity of maize pollen grains and transport pollen to considerable heights, so that it could travel long distances before settling (Arritt et al. 2007).
One of the key parameters originating from the local-scale studies is the estimation of the pollen source strength, as seen at various scales, and its dependence on the plant type and atmospheric conditions.
A detailed micro-scale experiment by Laursen et al. (2007) included 15-min measurements of the key meteorological parameters accompanied by the pollen concentration monitoring in the nearest vicinity of the plants. An empirical multi-linear regression fit was obtained for concentrations of Artemisia grains as a function of temperature, humidity, and wind speed. As was shown, wind and temperature are synergetic, i.e. higher wind and temperature both promote pollen release. Interestingly, relative humidity was not included into the equation (although measured during the experiment).
Aylor et al. (2006) estimated the maize pollen release rates 10–700 grains per square meter per second depending on weather factors and the day-to-day and diurnal variation of pollen production. In many cases pollen is not released steadily but reacts on wind gusts that “shake” the anthers. Grasses are known to disperse pollen at a certain time of the day, which varies between the species and environmental conditions. Some grass species exhibit a bimodal diurnal pattern (Davidson 1941; Subba Reddi et al. 1988).
Ambrosia pollen is initially shed from the staminate flowers in large pollen clumps containing hundreds of grains. Turbulent wind stress breaks these pollen clumps quickly into smaller ones, so that at regional scales Ambrosia pollen is released as a largely homogeneous plume. Modelling with large eddy simulation by Chamecki et al. (2009) provided an estimate of the total pollen emission of a ragweed field and its deposition partition inside the field and downwind. About 60% of pollen is deposited inside the field, 14% was deposited downwind inside the simulation domain, which is about 1,000 m long, while 26% of the total pollen remained airborne and left the domain.
Bricchi et al. (2000) found that the highest Platanus pollen deposition is recorded close to the plants and decreases quite quickly at a distance greater than 400 m. More than 88% of the pollen is deposited within a range of 2,750 m from the source.
A measuring experiment of local Betula pollen dispersion was presented by Michel et al. (2010), aiming at 3-D representation of the plume originating from an isolated birch stand.
A study by Spieksma et al. (2000) about the influence of nearby stands of Artemisia on pollen concentrations at street-level versus roof-top-level shows that the street-level concentrations were in average 4.8 times higher than the ones at 25 m on the roof.
5.4 Regional Scale
Regional-scale pollen dispersion over distances of about a hundred kilometres (or slightly more) poses different challenges in comparison with local scales. First of all, only a fraction of emitted pollen reaches these scales – so-called “regional component” (Faegri et al. 1989) or “escape fraction” (Gregory 1961) of the emission. Secondly, the observations at the receptor location are not fully representative for the pollen release area. They can, however, provide rough estimates for the transport conditions between the source and receptor points.
This connection of regional-scale meteorology and climatology with the pollen dispersion has been known for decades and is used in practically all studies. For example, the relation between measurements of Castanea sativa pollen concentration and the meteorological variables registered in different locations of Po Valley (Italy) were studied by Tampieri et al. (1977). Peeters and Zoller (1988) and Gehrig and Peeters (2000) connected the Betula and Castanea pollen distribution in Switzerland with such factors as altitude, prevailing meteorological patterns during spring time, etc.
The situations of strong regional pollen dispersion and the conditions favouring the regional-scale transport of Juniperus ashei pollen were described by Van de Water et al. (2003) and Van de Water and Levetin (2001). The authors qualitatively estimated the conditions favouring or precluding the released pollen to be transported regionally. A trajectory model HYSPLIT and five observation stations were used to obtain a complex pattern of the regional pollen distribution and to evaluate the allergenic threat in the region of Tulsa city. Conditions found to be favouring the regional-scale transport were: sunshine level of greater than 65%, temperature over 5 °C, relative humidity less than 50% and wind speed exceeding 1.8 m/s. The considered region was about 1,000 by 1,000 km with a characteristic transport distance being about 300–400 km in case of favourable conditions.
Many studies rely on single-point observations at a receptor site, thus concentrating on evaluation of the origin of the observed pollen. Strong indication of the regional-scale transport in such cases is high concentrations of pollen during night time when the local release is low.
Examples of these studies are: the multi-species study of Damialis et al. (2005) in Thessaloniki, the study of the regional impact of Copenhagen birch sources (Skjøth et al. 2008b), impact assessment of the southern England birch population onto London (Skjøth et al. 2009), evaluation of the role of regional and long-range transport on the Lithuanian pollen seasons by Veriankaité et al. (2010); the airborne monitoring during a cruise across the East Mediterranean Sea (Waisel et al. 2008). Most of the above studies provided qualitative connections between the regional-scale meteorological patterns and pollen distribution.
Some information helping to detect the pollen transport can be obtained using the local phenological observations together with the pollen counts. For example, the differences between flowering dates recorded by phenology network and pollen counts of Betula, Poaceae and Artemisia observed in Germany were correlated with the regional and large-scale transport by Estrella et al. (2006). Similar comparisons were used by Veriankaité et al. (2010), Siljamo et al. (2008b) and others. However, such comparison has to be performed with care because of very large uncertainties of the phenological observations (Siljamo et al. 2008a).
In contrast with the above studies stressing the episodic features of the regional transport, several assessments have concentrated on its long-term characteristics and impact. The comparison of multi-annual data sets of airborne and deposited pollen in northern Finland by Ranta et al. (2008) were used for qualitative description of the main transport directions in the region. The other long-term studies included the redistribution of genetically modified creeping bentgrass pollen (Van de Water et al. 2007); and paleopalynological studies (Romero et al. 2003; Hooghiemstra et al. 2006).
5.5 Long-Range Transport (LRT)
The large-scale dispersion of atmospheric constituents is controlled by synoptic-, continental-, or hemispheric- scale meteorological phenomena. In particular, it means that even combined observations at both source and receptor points cannot describe the transport conditions. In fact, in many cases the mere connection between the sources and receptors is difficult to establish due to complicated large-scale dispersion patterns. Under such constraints, the indication of the LRT can be either foreign pollen grains, which cannot be produced locally, or “wrong” time of appearance of the grains, which is significantly outside the local flowering period. Application of dispersion models in either forward or inverse mode is practically inevitable for analysis of such cases.
Studies of exotic grains observed in various parts of the globe started at least half a century ago, mainly in application to polar altitudes where the diversity of plants is comparatively low and exotic pollen is easier to find (Nichols 1967; Ritchie and Lichti-Federovich 1967; Janssen 1973; Ritchie 1974). Probably, these were the first unequivocal proofs of existence of such long-range dispersion of pollen material. Similar studies continued and extended towards temperate zones in later years (Hicks 1985; Porsbjerg et al. 2003; Hicks et al. 2001; Campbell et al. 1999; Rousseau et al. 2003, 2004, 2006). In some cases, specific meteorological conditions could be identified as a driving force for the exotic pollen appearance (Campbell et al. 1999). However, usually the number of the exotic grains found is small and the practical importance of the trans-continental transport for the human allergy and gene flow has proven to be low.
More significant amounts of pollen are episodically transported from Northern Africa, which has been identified as a source area in a number of studies. For instance, Van Campo and Quet (1982) identified several pollen types that had been transported from North Africa to Montpellier (France) together with mineral desert dust. Further south, Cannabis sativa (marihuana) pollen originating in Morocco was detected in Malaga, Southern Spain (Cabezudo et al. 1997) and Cannabis, Cupressus, Pinus, Platanus and Sambucus pollen were observed in Cordoba (South Spain) exclusively during African-dust events (Cariñanos et al. 2004). In addition, the source areas of several LRT episodes in Tenerife (Canary Islands) that originated from Mediterranean region were traced to the Saharan sector and the Sahel (Izquierdo et al. 2011).
A strong association of biological particles with desert dust was also suggested by Kellogg and Griffin (2006). Dust clouds generated by storm activity over arid land can result in mineral particles combined with viruses, bacteria (Hua et al. 2007; Hervàs et al. 2009), fungal spores (Griffin 2004, 2007; Griffin and Kellogg 2004; Griffin et al. 2001a, 2003, 2007, 2006; Kellog and Griffin 2006; Wu et al. 2004; Garrison et al. 2006; Schlesinger et al. 2006; Lee et al. 2007) and pollen being raised to altitudes over 2 km and then transported for thousands of kilometres, i.e. at a planetary scale. For example, viable microorganisms and fungal spores from Africa were sampled at Barbados after being transported by African dust plumes (Prospero et al. 2005). Intercontinental mineral dust transport has been the subject of much attention over decades (Guerzoni and Chester 1996; Goudie and Middleton 2001; Prospero et al. 1996; Zhang et al. 1997), but further research is needed on the biological component associated to the dust.
Finally, the study of aerobiological long-range transport has stimulated a new research line about the viability of pollen (Bohrerova et al. 2009), bacteria (Hervàs et al. 2009) and fungal spores (Gorbushina et al. 2007) after the long distance dispersion.
In Europe, numerous long range transport episodes have been identified in Fennoscandia (Ranta et al. 2006; Oikonen et al. 2005). Franzen and Hjelmroos (1988) observed pollen transport from Germany, Holland and England to the Swedish coast, and Franzen (1989) and Franzen et al. (1994) documented the arrival of pollen grains to Fennoscandia from the Mediterranean. A strong episode of the ragweed pollen transport from southern Europe to Finland was recorded in 2005 and traced back to the source areas in Hungary. A particularly specific spring season in Europe took place in 2006, when a strong plume of birch pollen was transported from Russia over the whole Europe and reached Iceland. The pollen cloud was mixed with the dense smoke from wild-land fires, which allowed its easy identification and follow-up by chemical observations and air quality models (e.g. Saarikoski et al. 2007).
As examples of transport episodes from East to West Europe, there are episodes of Betula pollen coming from Poland and Germany to Denmark (Skjøth et al. 2007) and from Russia to Finland (Siljamo et al. 2008b). In the opposite direction, Betula pollen arrived in Lithuania from Latvia, Sweden, Denmark, Belarus, Ukraine, Moldava, Germany and Poland (Veriankaité et al. 2010). Also, episodes of the pollen of the tree species Fagus sylvatica (beech) reaching Catalonia were traced back to central Europe (Belmonte et al. 2008). Complicated large-scale patterns have been found for Moscow by Siljamo et al. (2008b), who also reported strong bi-directional exchange of birch pollen between Russia and Finland.
The LRT episodes are not specifics of the tree pollen only. Thus, Smith et al. (2005) has registered Poaceae pollen in the UK originating from the continental Europe. The pollen spectrum of Lithuania was also affected by long range transport of several pollen types coming from other European regions (Sauliene et al. 2007).
As an outcome of a series of studies, Ambrosia distribution pathways over Europe were found to be highly irregular and episodic. Thus, the pollen registered in Poland, the Balkans and Italy was shown to originate near the Pannonian Plain (Cecchi et al. 2006, 2007; Stach et al. 2007; Kasprzyk 2008; Smith et al. 2008; Šikoparija et al. 2009) and Ukraine (Kasprzyk et al. 2010). In contrast, other studies recorded the transport in the opposite direction from France, Italy and Croatia to Hungary (Makra and Palfi 2007), as well as from France to Switzerland (Clot et al. 2002). There is also evidence of Ambrosia pollen transport to Catalonia (northeastern Spain) from France (Belmonte et al. 2000). Furthermore, an increasing risk of LRT Ambrosia episodes was detected in Scandinavia due to the rapid spread of ragweed in North-Northeast Europe (Dahl et al. 1999).
In North America, Raynor and Hayes (1983) applied the trajectory model to cases of pollen wet deposition in Albany (New York) after long-distance transport from the source areas located at South-Southwest. The arrival of Juniperus ashei pollen released in southern Oklahoma and Texas to Tulsa has been reported by Rogers and Levetin (1998) and Van de Water et al. (2003).
In South America, extra-regional pollen of Celtis coming from the northeast and of Nothofagus from the southwest has been found to contribute to Mar de Plata City (Argentina) pollen records (Gassmann and Pérez 2006).
In the north-western India, bioaerosols collected during dust storms sporadically contained pollen from Himalayan species (Yadav et al. 2007). The presence of pollen grains from trees forming forests at much lower latitudes has been evidenced in the Arctic environment (Bourgeois 2000; Savelieva et al. 2002; Rousseau et al. 2003, 2004, 2005, 2006, 2008). Extra-regional pollen transport has also been found in Antarctic (Wynn-Williams 1991), Arctic (Campbell et al. 1999), Australia (Salas 1983; Hart et al. 2007), and New Zealand (Moar 1969).
5.6 Release of Pollen Allergens from Grains
Probably the most important process that takes place during the pollen transport is the release of allergen from the grains. In a dry atmosphere, pollen is very stable and can keep its content over years (Stanley and Linskens 1974). However, under specific conditions the allergen release can take place within minutes: (i) a high relative air humidity; (ii) thunderstorms and heavy rain; (iii) high concentrations of air pollutants. According to Behrendt et al. (1997) under humid conditions, allergens are released from pollen grains in the process that resembles pollination. The existence of free allergen in air and its importance for the pollination season were confirmed by a series of field observations, such as Busse et al. (1972) for Ambrosia, Stewart and Holt (1985) for grass, and Rantio-Lehtimaki and Matikainen (2002), Rantio-Lehtimaki (2002), Matikainen and Rantio-Lehtimaki (1999), and Rantio-Lehtimaki et al. (1994) for birch.
Pollen allergens are generally glycoproteins. The majority of them are found in a limited number of protein families. Their biological functions are presumably related to the recognition, attachment, growth and development of the pollen tube on and within the pistil, i.e. to hydrolysis of proteins, polysaccharides, and lipids, binding of metal ions and lipids, and to the cytoskeleton (Radauer et al. 2008). Allergen activity can be detected, depending on the specific role of the protein in question, both before and after germination (Alché et al. 2002; Buters et al. 2010). These proteins were found in organelles, such as mitochondria, polysaccharide particles, starch granules, and endoplasmatic reticulum (Behrendt and Becker 2001; Rodríguez-García et al. 1995). Allergens are sometimes stored in the ectexine of the outer pollen wall. Rapid elution and water solubility is considered an important prerequisite for a protein to behave as a major allergen (Grote et al. 2001; Gupta et al. 1995).
The allergen release from pollen is responsible for bio-availability of the allergen (Behrendt and Becker 2001) but it also releases into the atmosphere the new particles with entirely different transport features. Indeed, using the formulations of Sofiev et al. (2006a) and applying them to the allergen size distribution observed by Taylor et al. (2004) or Miguel et al. (2006), one obtains the sedimentation velocity for allergen to be 0.01–0.1 mm sec−1, compared to 12 mm sec−1 for pollen. Allergens can stay in air much longer than pollens and thus be transported over much longer distances. In contrast to pollen grains, which due to their size do not penetrate into the lower human airways, the allergen aerosols are respirable and can provoke stronger immune response than pollen itself (Motta et al. 2006).
The allergen liberation is well reproduced in laboratories and can occur within a few tens of minutes in favourable conditions (Behrendt et al. 1997; Behrendt and Becker 2001; Taylor et al. 2002, 2004, 2007). The release can take place through the pollen walls or through the pollen tube wall. The liberation is pH- and temperature dependent (Behrendt et al. 1997). It is yet unclear how frequent this phenomenon is present in the real atmosphere because the field measurement of allergen are extremely complicated and often give semi-qualitative results based on the indirect indicators (Matikainen and Rantio-Lehtimaki 1999; Buters et al. 2010).
Schäppi et al. (1997) found that atmospheric concentrations of birch pollen allergen are correlated with birch pollen counts, but also that concentration of fine particles associated to Bet v 1 dramatically increase with light rainfall. They suggested that deposited pollen was stimulated to germinate, and that these particles were liberated as pollen tubes dried out when the rain was over. In Derby, UK, the pollen levels affected the number of emergency visits for asthma during days with light rainfall, but not during dry days (Lewis et al. 2000).
During thunderstorms, outbreaks of allergenic reactions have been registered and a high fraction of the patients were sensitive to both grass pollen and fungal spores (Pulimood et al. 2007). It was suggested that pollen grains, spores and other bioaerosols are carried into a cloud base, where they burst. Cold outflows transport the debris downwards where it can cause the observed asthma exacerbations (Marks et al. 2001).
In contrast to results from other studies, Buters et al. (2010) in Munich did not find any Bet v 1 in the fraction 0.12–2.5 μm. A hypothesis was suggested that either no allergen was present in the particles of this size range or the allergen was absorbed to diesel soot particles and became invisible for the technique used. Diesel soot particles bound to the grass allergen Lol p 1 under in vitro conditions were previously found by Knox et al. (1997).
An indirect way to detect the allergen in the air is to monitor the long-range transport plumes. Due to sharply different features of the particles, pollen grains are deposited faster than allergen and, after some travel time, the only aerosols present in air will be the allergen. The other option is to use size-segregated observations, thus separating pollens and allergens in different filters. That way, however, is difficult because of: (i) non-ideal filtering of coarse particles during sampling, which results in contamination of the fine-particle filter with pollen grains, (ii) very low allergen concentrations in ambient air – a few nanograms m−3 which are difficult to measure, (iii) addiction of allergen to the black carbon aerosols originating from diesel exhaust, which can make the detection of the allergens complicated or impossible.
5.7 Inter-Action Between Pollen and Chemical Pollutants
At all spatial and temporal scales, pollen, spores and, if present in a free form in the air, allergens are subjected to chemical and physical interactions with other atmospheric constituents. Chemical pollution can stress both the pollinating plants and the pollen grain, which cause qualitative and quantitative changes of the pollen content. The impact starts already during the pollen formation. For example, Aina et al. (2010) found an increased amount of the allergenic proteins in the grass Poa annua if the plant is grown in soil contaminated with cadmium. Similarly, the pollen vitality of Parietaria judaica was found maximal in soils enriched with heavy metals (Fotiou et al. 2010).
Processes taking place in the air during pollen transport are very poorly studied and one can only guess the type and intensity of the involved reactions. In an attempt to classify these processes, one can consider the physical transformations (rupture of grains, phase transformations of allergen particles, coagulation with other aerosols, etc.); chemical transformations (oxidation and nitration), and biological transformations of the particles (loss of viability, germination). Some of these processes can lead to substantial changes in the atmospheric features of the particles affecting their life time and dispersion features, and some are of importance from medical and biological points of view. In this sub chapter, we will consider only changes that are induced by chemical and physical transformations and result in alterations of the atmospheric features of the particles. The other aspects of the interactions are considered in other chapters of this book.
During transport, the water content of the grains is adapted to ambient humidity via water exchange through the pollen walls (Traidl-Hoffmann et al. 2003). In a very humid environment, a process resembling germination (so-called pseudogermination) can be triggered resulting in an abrupt release of the pollen content into the air (Motta et al. 2006; Traidl-Hoffmann et al. 2003; Grote et al. 2000, 2001, 2003). Together with water, pollen also absorbs dissolved heavy metals, nitrate and sulphur (WHO 2003).
The other process revealed in laboratory conditions refers to damage of pollen by aggressive chemicals, first of all ozone and nitrogen oxides. The related chemical processes are oxidation and nitration. According to Motta et al. (2006), treatment of pollen grains by ozone leads to a substantial increase of the fraction of damaged pollen already for the ozone concentrations of 100 ppb, which can easily be observed in the real atmosphere. For NO2, the levels of a few ppm needed for the non-negligible impact were much too high to represent any real-life case, which questions the role of nitrogen oxides in the pollen rupture. Apart from NO2 and O3, oxidation agents, such as hydroxyl radicals, can damage pollen or oxidise allergen but, to our knowledge, no experiments were performed with such species.
Chemical pollution can cause morphological changes of the pollen grains. Among the effects are the collapse and thinning of the exine (Shahali et al. 2009), which increases a bioavailability of the content. Stronger leaching was one of the explanations of observed decreased allergen detection in grass pollen exposed to car exhausts (Peltre et al. 1991), as well as to O3, SO2 and NO2 (Rogerieux et al. 2007).
The reaction between the pollen allergens and air pollutants is nitration (Franze et al. 2005), which may augment the allergenic potential of the allergen.
Interactions of pollen with fine aerosols of anthropogenic origin are confirmed by observations (Behrendt and Becker 2001), who showed that small carbon particles stick to the surface of pollen grains. This process provokes pseudo-germination and pollen rupture. However, comparatively fast depositing pollens can serve as a scavenging agent for small particles. The latter process, however, can be non-negligible only for the episodes with extremely high pollen concentrations in the air.
Interaction of allergen and diesel exhaust probably rests on the coagulation mechanism. Knox et al. (1997) showed that grass allergen molecules tend to bind to micrometre-size aggregates formed by black carbon particles. Since the resulting aggregate is in the respirable range and is much less soluble than the allergen itself, these particles tend to be transported even further than allergen and are capable of penetrating deep into human airways.
Apart from allergens, pollen grains also contain an array of non-allergenic but pro-inflammatory pollen-associated lipid mediators (PALMs) and enzymes that are suggested to be involved in the pathogenesis of allergic diseases (Gunawan et al. 2008; Gilles et al. 2010). Pollen collected in streets with a heavy traffic released significantly more PALMs than pollen collected in rural surroundings (Risse et al. 2000). A similar effect was found by Behrendt et al. (1999) for grains treated with volatile organic compounds.
5.8 Modelling the Pollen Dispersion
Modelling studies of pollen dispersion in the atmosphere are not numerous and concentrate on just two main directions. The most widely used model-based approach is an inverse-modelling analysis of the observational results and determination of the source regions affecting the specific monitoring site. This direction is dominated by simple backwards trajectory analysis.
The second application area – forward pollen dispersion – aims at simulation of the actual pollen life cycle: production, release into the air, transport, deposition, and sometimes, also the health impact. The outcome of such studies is a set of concentration and deposition maps of the particular pollen type. This task normally requires comprehensive systems, detailed input information of various kinds, and substantial computational resources. Compromising approaches can be applied in this area too but they are quite rare.
5.8.1 Inverse Studies and Analysis of Observational Results
The inverse modelling studies usually pursue one of the following objectives: (i) to find out the reasons of the observed peculiar behaviour of pollen concentrations, (ii) to outline the source areas most frequently and significantly affecting certain region. The first task normally covers a short period of time, up to a few days that immediately precede the considered episode, while the second task requires the analysis of a period of a few years to accumulate sufficient information for the statistical analysis.
The most common and simple method for back-tracing the origin of the observed pollen is the backward trajectories computed using a Lagrangian trajectory model (the Lagrangian trajectory approach). Within this method, a few trajectories directed backwards in time are started from the location of the pollen monitor and evaluated a few hours or days back in time. The resulting trajectories roughly show the direction from where the air masses arrived at the observation place during the observation time period. Since the method is qualitative, extra measures can be taken to increase the value of the outcome: clusters or ensembles of trajectories can be started from different points (e.g. Smith et al. 2005; Stach et al. 2007), trajectories can also be started at different heights, a random shift of start time can be applied, etc.
The backward trajectory methodology has been used for studying the origin of the observed pollen by Gassmann and Pérez (2006) for Celtis and Nothofagus pollen in Argentina, by Stach et al. (2007) and Smith et al. (2008) for Ambrosia in Poland, by Skjøth et al. (2007) for birch in Poland, by Šaulienė and Veriankaitė (2006) and Veriankaité et al. (2010) for birch in Lithuania, by Cecchi et al. (2007) for Ambrosia in Central Italy, by Mahura et al. (2007) and Skjøth et al. (2007, 2008b) for birch in Denmark, by Skjøth et al. (2009) for birch in London, etc. The bulk of the works were based on inverse trajectories computed by the NOAA HYSPLIT model (Draxler and Hess 2010), but also by own systems, such as THOR (Skjøth et al. 2002) in Denmark. A manual construction of trajectories via summing-up the wind vectors was used by Giner et al. (1999) for evaluation of the Artemisia source areas affecting the station at the eastern coast of Spain.
Apart from the above regional-scale assessments, trajectories were also used for back-tracking the transoceanic transport. In particular, many studies quoted in Sect. 5.5 (Long-Range Transport) have used trajectories to understand the origin of the exotic pollen (e.g. Rousseau et al. 2003, 2004, 2006).
A next step in the complexity of the methodology is consideration of a large number of Lagrangian particles – from a few thousands and up to a few millions. Each particle is transported with wind, thus drawing a trajectory, and is randomly relocated at every model time step, thus reflecting the impact of atmospheric diffusion. This approach is called as the “Lagrangian particle random-walk” method. In comparison with few trajectories, the large number of particles allows reproducing the actual shape of the clouds and makes it possible to include linear transformation processes, dry, and wet deposition. Most of the presently used Lagrangian models follow that paradigm: FLEXPART (Stohl et al. 2005), SILAM-L (Sofiev et al. 2006b), DERMA (Sorensen 1998), SNAP (Saltbones et al. 2001).
The statistical analysis of a great number of back trajectories from receptor sites has turned out to be an efficient tool to identify sources and sinks of atmospheric trace substances or to reconstruct their average spatial distribution (Ashbaugh 1983; Seibert et al. 1994). A trajectory statistical method was successfully applied to identify source areas for beech pollen recorded in Catalonia (Belmonte et al. 2008) and for pollen of various species recorded on Tenerife (Izquierdo et al. 2011).
Lagrangian particle models, being more realistic than Lagrangian trajectory ones, still inherit some weaknesses of the Lagrangian approach – first of all, limited spatial representativeness of a single Lagrangian particle. As a result, even for regional studies an astronomical number of particles have to be evaluated (Veriankaité et al. 2010; Siljamo, et al. 2008b; Sofiev et al. 2006a). It was also shown that the trajectory models show difficulties in mountain regions, where airflow patterns are known to be complex (Pérez-Landa et al. 2007a, b) and hard to explore with few trajectories. It has therefore been suggested that more sophisticated (Eulerian) approaches could be applied for such environments (Smith et al. 2008; Šikoparija et al. 2009).
Compared with the back-trajectories approaches, adjoint dispersion modelling offers a more rigorous instrument to identify the sources (Marchuk 1982). The idea of the method is to explicitly compute the sensitivity of the observed values to emission fluxes, chemical transformations and meteorological processes, which can affect the particular observation. Adjoint methods (both explicitly identified in the studies and implicitly included into the numerical systems) are extensively used in observational analysis at a local scale (Kuparinen 2006; Kuparinen et al. 2007; Rannik et al. 2003), in the source apportionment of greenhouse gas emissions (Bergamaschi et al. 2005), analysis of regional air quality (e.g. Saarikoski et al. 2007; Prank et al. 2008, 2010), and for the regional-scale source apportionment of the observed pollen (Veriankaité et al. 2010; Sofiev et al. 2006a, b; Siljamo et al. 2008b).
An example of comparison of the three methodologies is a study of Veriankaité et al. (2010), where the Lagrangian trajectory model HYSPLIT was used in combination with the SILAM model, which was run in two setups: as a back-tracking Lagrangian particle model and as an Eulerian model in adjoint mode. Despite qualitatively similar conclusions of all three analyses, a general recommendation was favouring the most comprehensive and least subjective approach of Eulerian adjoint modelling. It was noted that the results of trajectory analysis suffered from limited representativeness of a single trajectory and depended on subjective selection of the trajectory setup. The problems can become acute in meteorologically complicated situations, such as passage of a frontal line, strong cyclonic activity, etc.
As a post-processing of the model output for quantitative source identification, a number of analytical inversion techniques have been developed and applied in air pollution research (Wotawa et al. 2003; Seibert and Frank 2004; Stohl et al. 2009). Such methods have not yet been applied to the problem of identifying pollen sources. The same is true for variational methods of source apportionment (Sofiev and Atlaskin 2007).
5.8.2 Forward Simulations and Forecasting of the Pollen Seasons
Forward model simulations aim at predicting the pollen emission from plants, dispersion in the atmosphere and computing on its basis the concentrations of the grains at any distance from the sources. Similar to the inverse modelling, both Lagrangian and Eulerian approaches have been developed for these tasks.
A series of works addressed the local-scale dispersion of pollen from genetically modified plants: see review of Kuparinen (2006), applications for maize by Arritt et al. (2007), Jarosz et al. (2004), and Aylor et al. (2006), theoretical considerations of Novotny and Perdang (2003) and Fraile et al. (2006), meso-scale Lagrangian model of Kuparinen et al. (2007), etc.
In the 1980s, the significance of atmospheric conditions for the regional pollen dispersion was considered and the trajectory model applied for prediction of the transport direction (Davis and Main 1984, 1986). Another work was connected with the transport of marihuana pollen from Northern Africa to Europe (Cabezudo et al. 1997). The transport of Junipeerus Ashei pollen towards the city of Tulsa, USA, was studied by Van de Water et al. (2003) and Van de Water and Levetin (2001), who used a separately estimated release of pollen in combination with trajectory and weather pattern analysis. The result was a qualitative forecast of the threat to the downwind regions, covering a territory of about 1,000 km2. The outcome was encouraging: during the two seasons of 1998–1999, only a single occurrence of “high” or “very high” pollen concentrations in the city of Tulsa was not directly linked to “moderate” or “severe” forecast threat predictions. Other applications include the study of Pasken and Pietrowicz (2005) for oak, and the study by Kawashima and Takahashi (1999) for Japanese cedar (both emission and dispersion were predicted).
The results of trajectory model applications in 1990s showed a principal possibility to predict the pollen distribution by means of dispersion modelling. However, actual forecasting for large areas, such as Europe, could be approached only by integrated modelling systems, which incorporate descriptions of all the parts of the pollen cycle: production, release, transport, transformation, and deposition (Helbig et al. 2004; Sofiev et al. 2006a, 2009; Siljamo et al. 2004a, b, 2006, 2007, 2008a, b, c; Vogel et al. 2008). The corresponding biological, mathematical and physical sub-models are then driven only by historical, actual and forecasted meteorological information.
The input information for the pollen dispersion models consists of: (i) map of distribution of the source plants (see maps in Sofiev et al. 2006a; Skjøth et al. 2008a, 2010), (ii) information for the phenological model covering the whole region of computations and predicting both start and end of the flowering season (Helbig et al. 2004; Linkosalo et al. 2010; Siljamo et al. 2007, 2008c; Efstathiou et al. 2011), (iii) information on the pollen season severity obtained from observations or predicted from historical meteorological and pollen data (Ranta and Satri 2007), (iv) meteorological data produced by numerical weather prediction models. One has to warn against the climatology-based approaches, when the model is driven by long-term averaged parameters, such as the flowering date expressed as Julian date of the year: such approaches are bound to fail in every specific situation (Sofiev et al. 2006a).
Observational information on pollen distribution, to our knowledge, is not used in any of the presently existing systems. The same is true for the remote-sensing observations, despite showing that they provide valuable information about the phenological stages of some plants (Høgda et al. 2002).
The pollen emission and dispersion computations are usually split into two stages: prior to the season and during the season. Before the season, the only important process is the accumulation of the heat sum and other relevant parameters, which control the pollen development. Upon fulfilling some condition (e.g., reaching the prescribed heat sum threshold), the actual pollination starts. During that stage, the model follows the propagation of the flowering season but also responds to the short-term meteorological stress, such as cold weather, diurnal cycle of temperature and humidity, precipitation, etc. These variations can be parameterised as a function of daytime (e.g. Mahura et al. 2009) or predicted dynamically from the basic meteorological parameters (Linkosalo et al. 2010; Helbig et al. 2004; Siljamo et al. 2008b).
5.9 Discussion
In this section we will briefly touch on a few aspects of the pollen and allergen dispersion in the atmosphere, which bring together the above sections and illustrate interconnections between them. The topics are by no means exhaustive and the reader is referred to the cited literature sources for a more comprehensive analysis.
5.9.1 Quantification of Pollen Loads at Different Scales
As shown above, the pollen dispersion in the atmosphere is relevant at a very wide range of scales (Fig. 5.1) – from micro-scale up to continental. There are very few pollutants, which distribution can be relevant for so large variety of spatial and temporal scales. The main reason for this is a large variety of processes where the pollen impact is important – especially, biological and medical.
For successful fertilization, the density of the pollen cloud must be thousands of grains per cubic metre, though this level varies greatly for different species. At lower concentrations, probability of fertilization becomes too small to be biologically important (Gleaves 1973). Such concentrations usually can be obtained only in the source vicinity, i.e. biologically relevant distances hardly exceed tens or hundreds of metres for herbaceous species and, possibly, a few kilometres for wind-pollinating trees.
With regard to human allergy, concentrations much lower than those near the source are important (see the chapter on the pollen allergy threshold). In some cases, a few tens of grains per cubic metre can be sufficient to trigger the first symptoms. As demonstrated by the above-quoted studies, such concentrations can easily be observed at hundreds and, possibly, thousands of kilometres distance from the sources.
Finally, the inter-continental transported pollen, though existing (see the LRT section), can hardly play any practical role due to extremely low concentrations at the receptor point. One can, however, use exotic pollen as a natural tracer, which can reveal the pathway and frequency of the inter-continental exchange. Pollen, therefore, can serve as a marker of distribution of other species, whose observations may be not possible or very complicated.
A very wide range of relevant scales poses serious challenges to pollen monitoring and modelling. To date, the most widely used methodology of pollen monitoring is the Hirst (1952) trap. This is a low-volume device (about 0.01 m3 min−1), which is prone to significant uncertainties. The problem exacerbates further due to sharp gradients of pollen concentrations occurring in the vicinity of the sources: moving the trap higher or lower or repositioning it may lead to strong changes.
From the modelling point of view, the major challenge is also the near-source distribution and a fraction of pollen that reaches the regional scale of transport. All regional models currently ignore the complexity of the micro-scale processes replacing them with some variations of the “escape fraction” of Gregory (1961). The micro-scale models, to the contrary, concentrate of reproducing the patterns in the nearest vicinity of the source while the regional tails of the distributions are considered unimportant for the applications.
These deficiencies in both current monitoring and modelling techniques are to be addressed in future research in order to end up with comprehensive descriptions of pollen dispersion at all relevant spatial and temporal scales.
5.9.2 Variety of Modelling Approaches: Any Chance for Harmonization?
A variety of model types used for the pollen-related simulations is largely caused by historical reasons and conditions when the availability and simplicity of the systems were the primary selection criterion. However, the capabilities of the Lagrangian trajectory models are still sufficient for some tasks, first of all, for analysis of some specific measurements obtained at a single station. As long as a simple direction of transport is enough, these models are the reasonable choice. For more sophisticated observation analysis one has to rely on Lagrangian particle or Eulerian systems. Concerning the prediction of the pollen concentrations (the forward modelling tasks), the deficiencies of Lagrangian approaches (first of all, limited representativeness of a single particle) become impossible to overcome, so for these tasks the comprehensive Eulerian models are inevitable.
A specific problem of Lagrangian trajectory systems is selection of the type of trajectories. For about half a century it has been known that, for example, isobaric trajectories should not be used because they can produce entirely wrong results (Danielsen 1961). The isentropic trajectories are somewhat better but can still deviate from the true pathways, which can be drawn only by fully 3D trajectories. In that sense, the Lagrangian particle models, usually based on 3D trajectories, are preferable.
Considering the comprehensive systems for the pollen forecasts, one has to pay special attention to the phenological models as a part of the emission sub-system. There have been numerous phenological models developed for many species (see the Chap. 3, “Pollen season”, and Sect. 4.3.2 “Process based phenological models”, this issue). However, practically all of these models have been developed and evaluated within a limited territory. An attempt to use these models outside their native region leads to unpredictable results. The experience of SILAM system (presently, the only European-scale pollen forecasting system) shows that none of the existing phenological models can satisfactorily describe the birch phenology over the whole continent. The thermal-sum model used in SILAM had to be reparameterised for each sub-region independently (Sect. 4.3.3.1 “Application to large areas”). Understanding the differences between the regions and the reasons why the same taxa require different accumulated heat in different regions is a challenge for the future.
References
Aina, R., Asero, R., Ghiani, A., Marconi, G., Albertini, E., & Citterio, S. (2010). Exposure to cadmium-contaminated soils increases allergenicity of Poa annua L. pollen. Allergy, 65, 1313–1321. ALL2364 [pii], 10.1111/j.1398-9995.2010.02364.x [doi].
Alché, J. D., Castro, A. J., & Rodriguez-García, M. I. (2002). Localization of transcripts corresponding to the major allergen from olive pollen (Ole e I) by electron microscopic non-radioactive in situ RT-PCR. Micron, 33, 33–37.
Arritt, R. W., Clark, C. A., Goggi, S., Lopez Sanchez, H., Westgate, M. E., & Riese, J. M. (2007). Lagrangian numerical simulations of canopy air flow effects on maize pollen dispersal. Field Crops Research, 102, 151–162.
Ashbaugh, L. L. (1983). A statistical trajectory technique for determining air pollution source regions. Journal of the Air Pollution Control Association, 33, 1096–1098.
Aylor, D. E., Schultes, N. P., & Shields, E. J. (2003). An aerobiological framework for assessing cross-pollination in maize. Agricultural and Forest Meteorology, 119, 111–129.
Aylor, D. E., Boehm, M. T., & Shields, E. J. (2006). Quantifying aerial concentrations of maize pollen in the atmospheric surface layer using remote-piloted airplanes and Lagrangian stochastic modelling. Journal of Applied Meteorology and Climatology, 45, 1003–1015.
Behrendt, H., & Becker, W.-F. (2001). Localization, release and bioavailability of pollen allergens: The influence of environmental factors. Current Opinion in Immunology, 13, 709–715.
Behrendt, H., Becker, W. M., Fritzsche, C., Sliwa-Tomczok, W., Tomczok, J., Friedrichs, K. H., & Ring, J. (1997). Air pollution and allergy: Experimental studies on modulation of allergen release from pollen by air pollutants. International Archives of Allergy and Immunology, 113, 69–74.
Behrendt, H., Tomczok, J., Sliwa-Tomczok, W., Kasche, A., Ebner von Eschenbach, C., Becker, W. M., & Ring, J. (1999). Timothy grass (Phleum pratense L.) pollen as allergen carriers and initiators of an allergic response. International Archives of Allergy and Immunology, 118, 414–418.
Belmonte, J., Vendrell, M., Roure, J. M., Vidal, J., Botey, J., & Cadahía, A. (2000). Levels of Ambrosia pollen in the atmospheric spectra of Catalan aerobiological stations. Aerobiologia, 16, 93–99.
Belmonte, J., Alarcón, M., Avila, A., Scialabba, E., & Pino, D. (2008). Long-range transport of beech (Fagus sylvatica L.) pollen to Catalonia (north-eastern Spain). International Journal of Biometeorology, 52, 675–687. doi:10.1007/s00484-008-0160-9.
Bergamaschi, P., Krol, M., Dentener, F., Vermeulen, A., Meinhardt, F., Graul, R., Ramonet, M., Peters, W., & Dlugokencky, E. J. (2005). Inverse modelling of national and European CH4 emissions using the atmospheric zoom model TM5. Atmospheric Chemistry and Physics, 5, 2431–2460.
Boehm, M. T., Aylor, D. E., & Shields, E. J. (2008). Maize pollen dispersal under convective conditions. Journal of Applied Meteorology and Climatology, 47, 291–307.
Bohrerova, Z., Bohrer, G., Cho, K. D., Bolch, M. A., & Linden, K. G. (2009). Determining the viability response of pine pollen to atmospheric conditions during long-distance dispersal. Ecological Applications, 19(3), 656–667.
Bourgeois, J. C. (2000). Seasonal and interannual pollen variability in snow layers of arctic ice caps. Review of Palaeobotany and Palynology, 108, 17–36.
Bricchi, E., Frenguelli, G., & Mincigrucci, G. (2000). Experimental results about Platanus pollen deposition. Aerobiologia, 16, 347–352.
Brown, J. K. M., & Hovmoller, M. S. (2002). Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science, 297, 537–541.
Brunet, Y., Foueillassar, X., Audran, A., Garrigou, D., & Dayau, S. (2004). Evidence for long-range transport of viable maize pollen. Reprints, 16th Conference on Biometeorology and Aerobiology. Vancouver, Canada. American Meteorological Society, CD-ROM, P4A.2.
Burczyk, J., DiFazio, S. P., & Adams, W. T. (2004). Gene flow in forest trees: How far do genes really travel? Forest Genetics, 11, 1–14.
Busse, W. W., Charles, E. R., & Hoehne, J. H. (1972). Where is the allergic reaction in ragweed asthma? II. Demonstration of ragweed antigen in airborne particles smaller than pollen. The Journal of Allergy and Clinical Immunology, 50, 289–293.
Buters, J. T., Weichenmeier, I., Ochs, S., Pusch, G., Kreyling, W., Boere, A. J., Schober, W., & Behrendt, H. (2010). The allergen Bet v 1 in fractions of ambient air deviates from birch pollen counts. Allergy, 65(7), 850–858. doi:ALL2286 [pii], 10.1111/j.1398-9995.2009.02286.x.
Cabezudo, B., Recio, M., Sanchez-Laulhe, J. M., Trigo, M. M., Toro, F. J., & Polvorinos, F. (1997). Atmospheric transportation of marihuana pollen from North Africa to the southwest of Europe. Atmospheric Environment, 31, 3323–3328.
Campbell, I. D., McDonald, K., Flannigan, M. D., & Kringayark, J. (1999). Long-distance transport of pollen into the Arctic. Nature, 399, 29–30.
Cariñanos, P., Galán, C., Alcázar, P., & Domínguez, E. (2004). Analysis of the particles transported with dust-clouds reaching Córdoba, southwestern Spain. Archives of Environmental Contamination and Toxicology, 46, 141–146.
Cecchi, L., Morabito, M., Paola Domeneghetti, M., Crisci, A., Onorari, M., & Orlandini, S. (2006). Long distance transport of ragweed pollen as a potential cause of allergy in central Italy. Annales of Allergy Asthma and Immunology, 96, 86–91.
Cecchi, L., Torrigiani Malaspina, T., Albertini, R., Zanca, M., Ridolo, E., Usberti, I., Morabito, M., Dall’ Aglio, P., & Orlandini, S. (2007). The contribution of long-distance transport to the presence of Ambrosia pollen in central northern Italy. Aerobiologia, 23, 145–151.
Chamecki, M., Meneveau, C., & Parlange, M. B. (2009). Large eddy simulation of pollen transport in the atmospheric boundary layer. Journal of Aerosol Science, 40, 241–255.
Clot, B., Schneiter, D., Tercier, Ph, Gehrig, R., Peeters, A., Thibaudon, M., & Clot, B. (2002). Ambrosia pollen in Switzerland: Local production or transport? Allergie et Immunologie, 34, 126–128.
Dahl, A., Strandhede, S.-V., & Wihl, J.-A. (1999). Ragweed: An allergy risk in Sweden? Aerobiologia, 15, 293–297.
Damialis, A., Gioulekas, D., Lazopoulou, C., Balafoutis, C., & Vokou, D. (2005). Transport of airborne pollen into the city of Thessaloniki: The effects of wind direction, speed and persistence. International Journal of Biometeorology, 49, 139–145.
Danielsen, E. F. (1961). Trajectories: Isobaric, isentropic and actual. Journal of Meteorology, 18, 479–486.
Davis, J. M., Main, C. E. (1984). A regional analysis of the meteorological aspects of the spread and development of blue mold on tobacco. Boundary-Layer Meteorology, 28, 271–304.
Davis, J. M., Main, C. E. (1986). Applying atmospheric trajectory analysis to problems in epidemiology. Plant Disease, 70, 490–497.
Davidson, A. (1941). A note on anthesis in some common grasses near Johannesburg, and the relation of anthesis to collection of pollen for medical purposes. Journal of South African Botany, 7, 145–152.
Di-Giovanni, F., & Kevan, P. G. (1991). Factors affecting pollen dynamics and its importance to pollen contamination: A review. Canadian Journal of Forest Research, 21, 1155–1170.
Draxler, R. R., Hess, G. D. (2010). Description of the HYSPLIT_4 modelling system. NOAA Technical Memorandum ERL ARL-224NOAA Air Resources Laboratory; 27 pp. http://www.arl.noaa.gov/ready/hysplit4.html, http://www.arl.noaa.gov/documents/reports/arl-224.pdf. Last access 25 July 2012.
Efstathiou, C., Isukapalli, S., & Georgopoulos, P. (2011). A mechanistic modelling system for estimating large-scale emissions and transport of pollen and co-allergens. Atmospheric Environment 45(13), 2260–2276. [online] Available from: http://dx.crossref.org/10.1016%2Fj.atmosenv.2010.12.008 (last access 25.07.2012).
Ellstrand, N. C. (1992). Gene flow by pollen: Implications for plant conservation genetics. Oikos, 63, 77–86.
Ennos, R. A. (1994). Estimating the relative rates of pollen and seed migration among plant-populations. Heredity, 72, 250–259.
Estrella, N., Menzel, A., Krämer, U., & Behrendt, H. (2006). Integration of flowering dates in phenology and pollen counts in aerobiology: Analysis of their spatial and temporal coherence in Germany (1992-1999). International Journal of Biometeorology, 54, 49–59.
European Environment Agency. (2002). Genetically modified organisms (GMOs): The significance of gene flow through pollen transfer. Environmental Issue Report No 28, ISBN: 92-9167-411-7, Copenhagen, 75 pp. http://www.eea.europa.eu/publications/environmental_issue_report_2002_28 (last access 25.07.2012).
Faegri, K., Iversen, J., & Krzywinski, K. (1989). Textbook of pollen analysis. Toronto: Wiley. 328 pp.
Fotiou, C., Damialis, A., Krigas, N., Halley, J. M., & Vokou, D. (2010). Parietaria judaica flowering phenology, pollen production, viability and atmospheric circulation, and expansive ability in the urban environment: Impacts of environmental factors. International Journal of Biometeorology, 55, 35–50. doi:10.1007/s00484-010-0307-3.
Fraile, R., Calvo, A. I., Castro, A., Fernandez-Gonzalez, D., & Garcıa-Ortega, E. (2006). The behavior of the atmosphere in long-range transport. Aerobiologia, 22, 35–45.
Franze, T., Weller, M. G., Niessner, R., & Poschl, U. (2005). Protein nitration by polluted air. Environmental Science and Technology, 39, 1673–1678.
Franzen, L. (1989). A dustfall episode on the Swedish west-coast, October 1987. Geografiska Annaler Series A, Physical Geography, 71, 263–267.
Franzen, L., & Hjelmroos, M. (1988). A coloured snow episode on the Swedish west coast, January 1987. A quantitative study of air borne particles. Geografiska Annaler Series A, Physical Geography, 70, 235–243.
Franzen, L., Hjelmroos, M., Kallberg, P., Brorstromlunden, E., Juntto, S., & Savolainen, A. L. (1994). The yellow-snow episode of northern Fennoscandia, March-1991: A case-study of long-distance transport of soil, pollen and stable organic-compounds. Atmospheric Environment, 28, 3587–3604.
Gage, S., Isard, S. A., & Colunga, G. M. (1999). Ecological scaling of aerobiological dispersal processes. Agricultural and Forest Meteorology, 97, 249–261.
Garrison, V. H., Shinn, E. A., Foreman, W. T., Griffin, D. W., Holmes, C. W., Kellogg, C. A., Majewski, M. S., Richardson, L. L., Ritchie, K. B., & Smith, G. W. (2003). African and Asian dust: From desert soils to coral reefs. Bioscience, 53, 469–480.
Garrison, V. H., Foreman, W. T., Genualdi, S., Griffin, D. W., Kellogg, C. A., Majewski, M. S., Mohammed, A., Ramsubhag, A., Shinn, E. A., Simonich, S. L., & Smith, G. W. (2006). Saharan dust – a carrier of persistent organic pollutants, metals and microbes to the Caribbean? Revista Biologia Tropical (Int. J. Trop. Biol. ISSN-0034-7744) 54(3), 9–21.
Gassmann, M. I., & Pérez, C. F. (2006). Trajectories associated to regional and extra-regional pollen transport in the southeast of Buenos Aires province, Mar Del Plata (Argentina). International Journal of Biometeorology, 50, 280–291. doi:10.1007/s00484-005-0021-8.
Gehrig, R., & Peeters, A. G. (2000). Pollen distribution at elevations above 1000 m in Switzerland. Aerobiologia, 16, 69–74.
Gilles, S., Jacoby, D., Blume, C., Mueller, M. J., Jakob, T., Behrendt, H., Schaekel, K., & Traidl-Hoffmann, C. (2010). Pollen-derived low-molecular weight factors inhibit 6-sulfo LacNAc + dendritic cells’ capacity to induce T-helper type 1 responses. Clinical and Experimental Allergy, 40, 269–278. doi:CEA3369 [pii], 10.1111/j.1365-2222.2009.03369.x.
Giner, M. M., Garcia, C. J. S., & Selles, G. J. (1999). Aerobiology of Artemisia airborne pollen in Murcia (SE Spain) and its relationship with weather variables: Annual and intradiurnal variations for three different species. Wind vectors as a tool in determining pollen origin. International Journal of Biometeorology, 43, 51–63.
Gleaves, J. T. (1973). Gene flow mediated by wind-borne pollen. Heredity, 31, 355–366.
Gorbushina, A. A., Kort, R., Schuite, A., Lazarus, D., Schnetger, B., Brumsack, H., Broughton, W. J., & Favet, J. (2007). Life in Darwin’s dust: Intercontinental transport and survival of microbes in the nineteenth century. Environmental Microbiology, 9(12), 2911–2922. doi:10.1111/j.1462-2920.20.2007.01461.x.
Goudie, A. S., & Middleton, N. J. (2001). Saharan dust storms: Nature and consequences. Earth-Science Reviews, 56, 179–204.
Govindaraju, D. R. (1988). Relationship between dispersal ability and levels of gene flow in plants. Oikos, 52, 31–35.
Govindaraju, D. R. (1989). Estimates of gene flow in forest trees. Biological Journal of the Linnean Society, 37, 345–357.
Gregory, P. H. (1961). The microbiology of the atmosphere. New York: Interscience. 251 pp.
Griffin, D. W. (2004). Terrestrial microorganism at an altitude of 20,000 m in Earth’s atmosphere. Aerobiologia, 20, 135–140.
Griffin, D. W. (2007). Atmospheric movement of microorganisms in clouds of desert dust and implications for human health. Clinical Microbiology Reviews, 20, 459–477.
Griffin, D. W., & Kellogg, C. A. (2004). Dust storms and their impact on ocean and human health: Dust in Earth’s atmosphere. EcoHealth, 1, 284–295.
Griffin, D. W., Kellogg, C. A., & Shinn, E. A. (2001a). Dust in the wind: Long range transport of dust in the atmosphere and its implications for global public and ecosystems health. Global Change and Human Health, 2, 20–33.
Griffin, D. W., Garrison, V. H., Herman, J. R., & Shinn, E. (2001b). African desert dust in the Caribbean atmosphere: Microbiology and public health. Aerobiologia, 17, 203–213.
Griffin, D. W., Kellogg, C. A., Garrison, V. H., Lisle, J. T., Borden, T. C., & Shinn, E. A. (2003). Atmospheric microbiology in the northern Caribbean during African dust events. Aerobiologia, 19, 143–157.
Griffin, D. W., Westplhal, D. L., & Gray, M. A. (2006). Airborne microorganisms in the African desert dust corridor over the mid-Atlantic ridge, Ocean Drilling Program, Leg 209. Aerobiologia, 22, 211–226. doi:10.1007/s10453-006-9033-z.
Griffin, D. W., Kubilay, N., Koçak, M., Gray, M. A., Borden, T. C., & Shinn, E. A. (2007). Airborne desert dust and aeromicrobiology over the Turkish Mediterranean coastline. Atmospheric Environment, 41, 4050–4062.
Grote, M., Vrtala, S., Niederberger, V., Valenta, R., & Reichelt, R. (2000). Expulsion of allergen-containing materials from hydrated rye grass (Lolium perenne) pollen revealed by using immunogold field emission scanning and transmission electron microscopy. The Journal of Allergy and Clinical Immunology, 105, 1140–1145.
Grote, M., Vrtala, S., Niederberger, V., Wierman, R., Valenta, R., & Reichelt, R. (2001). Release of allergen-bearing cytoplasm from hydrated pollen: A mechanism common to variety of grass (Poaceae) species revealed by electron microscopy. The Journal of Allergy and Clinical Immunology, 108, 109–115.
Grote, M., Valenta, R., & Reichelt, R. (2003). Abortive pollen germination: A mechanism of allergen release in birch, alder and hazel revealed by immunogold electron microscopy. The Journal of Allergy and Clinical Immunology, 111, 1017–1023.
Guerzoni, S., & Chester, R. (1996). The impact of desert dust across the Mediterranean. Dordrecht: Kluwer Academic Publishers. 389 pp.
Gunawan, H., Takai, T., Kamijo, S., Wang, X. L., Ikeda, S., Okumura, K., & Ogawa, H. (2008). Characterization of proteases, proteins, and eicosanoid-like substances in soluble extracts from allergenic pollen grains. International Archives of Allergy and Immunology, 147, 276–288. doi:000144035 [pii] 10.1159/000144035 [doi].
Gupta, N., Sriramarao, P., Kori, R., & Rao, P. V. (1995). Immunochemical characterization of rapid and slowly released allergens from the pollen of Parthenium hysterophorus. International Archives of Allergy and Immunology, 107, 557–565.
Hart, M. A., de Dear, R., & Beggs, P. J. (2007). A synoptic climatology of pollen concentrations during the six warmest months in Sydney, Australia. International Journal of Biometeorology, 51, 209–220.
Helbig, N., Vogel, B., Vogel, H., & Fiedler, F. (2004). Numerical modelling of pollen dispersion on the regional scale. Aerobiologia, 20(1), 3–19.
Hervàs, A., Camarero, L., Reche, I., & Casamayor, E. O. (2009). Viability and potential for immigration of airborne bacteria from Africa that reach high mountain lakes in Europe. Environmental Microbiology, 11(6), 1612–1623.
Hicks, S. (1985). Modern pollen deposition records from Kuusamo, Finland. Grana, 24, 167–184.
Hicks, S. H., Tinsley, A., Huusko, C., Jensen, M., Hättestrand, M., Gerasimedes, A., & Kvavadze, E. (2001). Some comments on spatial variation in arboreal pollen deposition: First records from the Pollen Monitoring Programme (PMP). Review of Palaeobotany and Palynology, 117, 183–194.
Hirst, J. M. (1952). An automatic volumetric spore trap. Annals of Applied Biology, 39(2), 257–265.
Høgda, K. A., Karlsen, S. R., Solheim, I., Tommervik, H., & Ramfjord, H. (2002). The start dates of birch pollen seasons in Fennoscandia studied by NOAA AVHRR NDVI data. Proceeding of IGARSS. June 24–28, 2002, Toronto. ISBN 0-7803-7536-X.
Hooghiemstra, H., Lezine, A. M., Leroy, S. A. G., Dupont, L., & Marret, F. (2006). Late quaternary palynology in marine sediments: A synthesis of the understanding of pollen distribution patterns in the NW African setting. Quaternary International, 148, 29–44.
Hua, N., Kobayashi, F., Iwasaka, Y., Shi, G., & Naganuma, T. (2007). Detailed identification of desert-originated bacteria carried by Asian dust storms to Japan. Aerobiologia, 23, 291–298.
Isard, S. A., & Gage, S. H. (2001). Flow of life in the atmosphere: An airscape approach to understanding invasive organisms. East Lansing: Michigan State University Press. 240 pp.
Isard, S. A., Gage, S. H., Comtois, P., & Russo, J. M. (2005). Principles of the atmospheric pathway for invasive species applied to soybean rust. Bioscience, 55(10), 851–861.
Izquierdo, R., Belmonte, J., Avila, A., Alarcón, M., Cuevas, E., & Alonso-Pérez, S. (2011). Source areas and long-range transport of pollen from continental land to Tenerife (Canary Islands). International Journal of Biometeorology, 55(1), 67–85. doi: 10.1007/s00484-010-0309-1.
Janssen, C. R. (1973). Local and regional pollen deposition. In J. H. B. Birks & R. G. West (Eds.), Quaternary plant ecology (pp. 30–43). Oxford: Blackwell Scientific.
Jarosz, N., Loubet, B., Durand, B., McCartney, A., Foueillassar, X., & Huber, L. (2003). Field measurements of airborne concentration and deposition rate of maize pollen. Agricultural and Forest Meteorology, 119, 37–51.
Jarosz, N., Loubet, B., & Huber, L. (2004). Modelling airborne concentrations and deposition rate of maize pollen. Atmospheric Environment, 38, 5555–5566.
Kasprzyk, I. (2008). Non-native Ambrosia pollen in the atmosphere of Rzeszow (SE Poland); Evaluation of the effect of weather conditions on daily concentrations and starting dates of the pollen season. International Journal of Biometeorology, 52, 341–351. doi:10.1007/s00484-007-0129.
Kasprzyk, I., Myszkowska, D., Grewling, Ł., Stach, A., Šikoparija, B., Skjøth, C. A., & Smith, M. (2010). The occurrence of Ambrosia pollen in Rzeszów, Kraków and Poznań, Poland: Investigation of trends and possible transport of Ambrosia pollen from Ukraine. International Journal of Biometeorology. doi:10.1007/s00484-010-0376-3.
Kawashima, S., & Takahashi, Y. (1999). An improved simulation of mesoscale dispersion of airborne cedar pollen using a flowering-time map. Grana, 38, 316–324.
Kazlauskas, M., Sauliene, I., & Lankauskas, A. (2006). Airborne Artemisia pollen in Siauliai (Lithuania) atmosphere with reference to meteorological factors during 2003-2005. Acta Biologica Universitatis, 6, 1–2.
Kellogg, C. A., & Griffin, D. W. (2006). Aerobiology and the global transport of desert dust. Trends in Ecology & Evolution, 21, 638–644.
Kellogg, C. A., Griffin, D. W., Garrison, V. H., Peak, K. K., Royall, N., Smith, R. R., & Shinn, E. A. (2004). Characterization of aerosolized bacteria and fungi from desert dust events in Mali, West Africa. Aerobiologia, 20, 99–110.
Klein, E. K., Lavigne, C., Foueilassar, X., Gouyon, P. H., & Laredo, C. (2003). Corn pollen dispersal: Quasi-mechanistic models and field experiments. Ecological Monographs, 73, 131–150.
Knox, R. B., Suphioglu, C., Taylor, P., Desai, R., Watson, H. C., Peng, J. L., & Bursill, L. A. (1997). Major grass pollen allergen Lol p 1 binds to diesel exhaust particles: Implications for asthma and air pollution. Clinical and Experimental Allergy, 27, 246–251.
Kuparinen, A. (2006). Mechanistic models for wind dispersal. Trends in Plant Science, 11, 298–301.
Kuparinen, A., Markkanen, T., Riikonen, H., & Vesala, T. (2007). Modelling air-mediated dispersal of spores, pollen and seeds in forested areas. Ecological Modelling, 208, 177–188.
Laursen, S. C., Reiners, W. A., Kelly, R. D., & Gerow, K. G. (2007). Pollen dispersal by Artemisia tridentata (Asteraceae). International Journal of Biometeorology, 51, 465–481.
Lee, A. K. Y., Lau, A. P. S., Cheng, J. Y. W., Fang, M., & Chan, C. K. (2007). Source identification analysis for the airborne bacteria and fungi using a biomarker approach. Atmospheric Environment, 41, 2831–2843.
Lewis, S. A., Corden, J. M., Forster, G. E., & Newlands, M. (2000). Combined effects of aerobiological pollutants, chemical pollutants and meteorological conditions on asthma admissions and A & E attendances in Derbyshire UK, 1993-96. Clinical and Experimental Allergy, 30, 1724–1732.
Linkosalo, T., Ranta, H., Oksanen, A., Siljamo, P., Luomajoki, A., Kukkonen, J., & Sofiev, M. (2010). A double-threshold temperature sum model for predicting the flowering duration and relative intensity of Betula pendula and B. pubescens. Agricultural and Forest Meteorology, 150(12), 1579–1584.
Linskens, H. F., & Cresti, M. (2000). Pollen allergy as an ecological phenomenon: A review. Plant Biosystems, 134(3), 341–352.
Mahura, A., Korsholm, S. U., Baklanov, A. A., & Rasmussen, A. (2007). Elevated birch pollen episodes in Denmark: Contributions from remote sources. Aerobiologia, 23, 171–179.
Mahura, A., Baklanov, A., & Korsholm, U. (2009). Parameterization of the birch pollen diurnal cycle. Aerobiologia, 25, 203–208.
Makra, L., & Palfi, S. (2007). Intra-regional and long-range ragweed pollen transport over southern Hungary. Acta Climatologica Et Chorologica, 40–41, 69–77.
Marchuk, G. I. (1982). Mathematical modelling in the environmental problems. Moscow: “Nauka” publisher. 320 pp, (in Russian).
Marks, G. B., Colquhoun, J. R., Girgis, S. T., Koski, M. H., Treloar, A. B., Hansen, P., Downs, S. H., & Car, N. G. (2001). Thunderstorm outflows preceding epidemics of asthma during spring and summer. Thorax, 56, 468–471.
Matikainen, E., & Rantio-Lehtimaki, A. (1999). Semiquantitative and qualitative analysis of pre seasonal airborne birch pollen allergens in different particle sizes. Grana, 37, 293–297.
McCartney, H. A., & Lacey, M. E. (1991). Wind dispersal of pollen from crops of oilseed rape (Brassica napus L.). Journal of Aerosol Science, 22, 467–477.
Michel, D., Rotach, M. W., Gehrig, R., & Vogt, R. (2010). Experimental investigation of micrometeorological influences on birch pollen emission. Arbeitsberichte der MeteoSchweiz, 230, 37 pp.
Miguel, A. G., Taylor, P. E., House, J., Glovsky, M. M., & Flagan, R. C. (2006). Meteorological influences on respirable fragment release from Chinese Elm pollen. Aerosol Science and Technology, 40(9), 690–696.
Moar, N. T. (1969). Possible long-distance transport of pollen to New Zealand. New Zealand Journal of Botany, 7, 424–426.
Motta, A. C., Marliere, M., Peltre, G., Sterenberg, P. A., & Lacroix, G. (2006). Traffic-related air pollutants induce the release of allergen-containing cytoplasmic granules from grass pollen. International Archives of Allergy and Immunology, 139, 294–298.
Nichols, H. (1967). Pollen diagrams from Sub-Arctic Central Canada. Science, 155, 1665–1668.
Novotny, E., & Perdang, J. (2003). Simulations of pollen transport by wind. SGR (Scientist for Global Responsibility), 3 pp.
Oikonen, M., Hicks, S., Heino, S., & Rantio-Lehtimaki, A. (2005). The start of the birch pollen season in Finnish Lapland: Separating non-local from local birch pollen and the implication for allergy sufferers. Grana, 44, 181–186.
Pacini, E., & Hesse, M. (2004). Cytophysiology of pollen presentation and dispersal. Flora, 199, 273–285.
Pacini, E., Guarnieri, M., & Nepi, M. (2006). Pollen carbohydrates and water content during development, presentation and dispersal: A short review. Protoplasma, 228, 73–77.
Pasken, R., & Pietrowicz, J. A. (2005). Using dispersion and mesoscale meteorological models to forecast pollen concentrations. Atmospheric Environment, 39, 7689–7701.
Paz, S., & Broza, M. (2007). Wind direction and its linkage with Vibrio cholerae dissemination. Environmental Health Perspectives, 115(2), 195–200.
Peeters, A. G., & Zoller, H. (1988). Long range transport of Castanea sativa pollen. Grana, 27, 203–207.
Peltre, G., Derouet, L., & Cerceau-Larrival, M. T. (1991). Model treatments simulating environmental action on allergenic Dactylis glomerata pollen. Grana, 30, 59–61.
Pérez-Landa, G., Ciais, P., Gangoiti, G., Palau, J. L., Carrara, A., Gioli, B., Miglietta, F., Schumacher, M., Millán, M. M., & Sanz, M. J. (2007a). Mesoscale circulations over complex terrain in the Valencia coastal region, Spain – Part 2: Modeling CO2 transport using idealized surface fluxes. Atmospheric Chemistry and Physics, 7, 1851–1868.
Pérez-Landa, G., Ciais, P., Sanz, M. J., Gioli, B., Miglietta, F., Palau, J. L., Gangoiti, G., & Millán, M. M. (2007b). Mesoscale circulations over complex terrain in the Valencia coastal region, Spain – Part 1: Simulation of diurnal circulation regimes. Atmospheric Chemistry and Physics, 7, 1835–1849.
Polymenakou, P. N., Mandalakis, M., Stephanou, E. G., & Tselepides, A. (2008). Particle size distribution of airborne microorganisms and pathogens during an intense African dust event in the Eastern Mediterranean. Environmental Health Perspectives, 116, 292–296.
Porsbjerg, C., Rasmussen, A., & Backer, A. (2003). Airborne pollen in Nuuk, Greenland, and the importance of meteorological parameters. Aerobiologia, 19, 29–37.
Prank, P., Sofiev, M., Kaasik, M., Ruuskanen, T., Kukkonen, J., & Kulmala, M. (2008). The origin and formation mechanics of aerosol during a measurement campaign in Finnish Lapland, evaluated using the regional dispersion model SILAM. In C. Borrego & A. I. Miranda (Eds.), Air pollution modeling and its application XIX (NATO science for peace and security series-C: Environmental security, pp. 530–538). Berlin: Springer.
Prank, M., Sofiev, M., Denier van der Gon, H. A. C., Kaasik, M., Ruuskanen, T., & Kukkonen, J. (2010). A refinement of the emission data for Kola Peninsula based on inverse dispersion modelling. Atmospheric Chemistry and Physics Discussions, 10, 15963–16006.
Prospero, J. M., Barett, K., Churcha, T., Dentener, F., Duce, R. A., Galloway, J. N., Levy, H., II, Moody, J., & Quinn, P. (1996). Atmospheric deposition of nutrients to the North Atlantic Basin. Biogeochemistry, 35, 27–73.
Prospero, J. M., Blades, E., Mathison, G., & Naidu, R. (2005). Interhemispheric transport of viable fungi and bacteria from Africa to the Caribbean with soil dust. Aerobiologia, 21, 1–19.
Pulimood, T. B., Corden, J. M., Bryden, C., Sharples, L., & Nasser, S. M. (2007). Epidemic asthma and the role of the fungal mold Alternaria alternata. The Journal of Allergy and Clinical Immunology, 120, 610–617. doi:S0091-6749(07) 00970-0 [pii], 10.1016/j.jaci.2007.04.045 [doi].
Radauer, C., Bublin, M., Wagner, S., Mari, A., & Breiteneder, H. (2008). Allergens are distributed into few protein families and possess a restricted number of biochemical functions. The Journal of Allergy and Clinical Immunology, 121, 847.e7–852.e7. DOI: S0091-6749(08)00163-2.
Rannik, U., Markkanen, T., Raittila, J., Hari, P., & Vesala, T. (2003). Turbulence statistics inside and over forest: Influence on footprint prediction. Boundary-Layer Meteorology, 109, 163–189.
Ranta, H., & Satri, P. (2007). Synchronised inter-annual fluctuations of flowering intensity affects the exposure to allergenic tree pollen in north Europe. Grana, 46(4), 274–284.
Ranta, H., Kubin, E., Siljamo, P., Sofiev, M., Linkosalo, T., Oksanen, A., & Bondestam, K. (2006). Long distance pollen transport cause problems for determining the timing of birch pollen season in Fennoscandia by using phenological observations. Grana, 45(4), 297–304.
Ranta, H., Sokol, C., Hicks, S., Heino, S., & Kubin, E. (2008). How do airborne and deposition pollen samplers reflect the atmospheric dispersal of different pollen types? An example from northern Finland. Grana, 47, 285–296.
Rantio-Lehtimaaki, A., Viander, M., & Koivikko, A. (1994). Airborne birch pollen antigens in different particle sizes. Clinical and Experimental Allergy, 24, 23–28.
Rantio-Lehtimaki, A. (2002). Siitepolyallergeenit sisalyss. Allergia & Asthma, 2, 25–27 (in Finnish).
Rantio-Lehtimaki, A., & Matikainen, E. (2002). Pollen allergen reports help to understand preseason symptoms. Aerobiologia, 18, 135–140.
Raynor, G. S., & Hayes, J. V. (1983). Testing of the air resources laboratories trajectory model on cases of pollen wet deposition after long-distance transport from known source regions. Atmospheric Environment, 17(2), 213–220.
Raynor, G. S., Ogden, E. C., & Hayes, J. V. (1970). Dispersion and deposition of ragweed pollen from experimental sources. Journal of Applied Meteorology, 9, 885–895.
Raynor, G. S., Ogden, E. C., & Hayes, J. V. (1972a). Dispersion and deposition of timothy pollen from experimental sources. Agricultural Meteorology, 9, 347–366.
Raynor, G. S., Ogden, E. C., & Hayes, J. V. (1972b). Dispersion and deposition of corn pollen from experimental sources. Agronomy Journal, 64, 420–427.
Risse, U., Tomczok, J., Huss-Marp, J., Darsow, U., & Behrendt, H. (2000). Health-relevant interaction between airborne particulate matter and aeroallergens (pollen). Journal of Aerosol Science, 31, 27–28.
Ritchie, J. C. (1974). Modern pollen assemblages near arctic tree Line, Mackenzie Delta Region, Northwest-Territories. Canadian Journal of Botany, 52, 381–396.
Ritchie, J. C., & Lichti-Federovich, S. (1967). Pollen dispersal phenomena in Arctic-Subarctic Canada. Review of Palaeobotany and Palynology, 3, 255–266.
Rodríguez-García, M. I., Fernández, M. C., Alché, J. D., & Olmedilla, A. (1995). Endoplasmic reticulum as a storage site for allergenic proteins in pollen grains of several Oleaceae. Protoplasma, 187, 111–116. doi:10.1007/bf01280238.
Rogerieux, F., Godfrin, D., Senechal, H., Motta, A. C., Marliere, M., Peltre, G., & Lacroix, G. (2007). Modifications of Phleum pratense grass pollen allergens following artificial exposure to gaseous air pollutants (O3, NO2, SO2). International Archives of Allergy and Immunology, 143, 127–134. doi:000099079 [pii] 10.1159/000099079 [doi].
Rogers, C. A., & Levetin, E. (1998). Evidence of long-distance transport of mountain cedar pollen into Tulsa, Oklahoma. International Journal of Biometeorology, 42, 65–72.
Romero, O. E., Dupont, L., Wyputta, U., Jahns, S., & Wefer, G. (2003). Temporal variability of fluxes of eolian-transported freshwater diatoms, phytoliths, and pollen grains off Cape Blanc as reflection of land-atmosphere-ocean interactions in northwest Africa. Journal of Geophysical Research-Oceans, 108(C5), 3153–3164. doi:10.1029/2000JC000375/2003.
Rousseau, D. D., Duzer, D., Cambon, G. V., Jolly, D., Poulsen, U., Ferrier, J., Schevin, P., & Gros, R. (2003). Long distance transport of pollen to Greenland. Geophysical Research Letters, 30, 1765. doi:10.1029/2003GL017539.
Rousseau, D. D., Duzer, D., Etienne, J.-L., Cambon, G., Jolly, D., Ferrier, J., & Schevin, P. (2004). Pollen record of rapidly changing air trajectories to the North Pole. Journal of Geophysical Research, 109, D06116. doi:10.1029/2003JD003985.
Rousseau, D. D., Schevin, P., Duzer, D., Cambon, G., Ferrier, J., Jolly, D., & Poulsen, U. (2005). Pollen transport to southern Greenland: New evidences of a late spring long distance transport. Biogeosciences Discussions, 2(4), 709–715.
Rousseau, D. D., Schevin, P., Duzer, D., Cambon, G. V., Ferrier, J., Jolly, D., & Poulsen, U. (2006). New evidence of long distance pollen transport to southern Greenland in late spring. Review of Palaeobotany and Palynology, 141, 277–286. doi:10.1016/j.revpalbo.2006.05.001.
Rousseau, D. D., Schevin, P., Ferrier, J., Jolly, D., Andreasen, T., Ascanius, S. E., Hendriksen, S. E., & Poulsen, U. (2008). Long-distance pollen transport from North America to Greenland in spring. Journal of Geophysical Research-Biogeosciences, 113. doi:10.1029/2007JG000456.
Saarikoski, S., Sillanpää, M., Sofiev, M., Timonen, H., Saarnio, K., Teinilä, K., Karppinen, A., Kukkonen, J., & Hillamo, R. (2007). Chemical composition of aerosols during a major biomass burning episode over northern Europe in spring 2006: Experimental and modelling assessments. Atmospheric Environment, 41, 3577–3589.
Salas, M. R. (1983). Long-distance pollen transport over the southern Tasman Sea: Evidence from Macquarie Island. New Zealand Journal of Botany, 21, 285–292.
Saltbones, J., Bartnicki, J., & Foss, A. (2001). Handling of fallout processes from nuclear explosions in a severe nuclear accident program (SNAP). met.no report no. 157/2003, http://ebookbrowse.com/saltbones-pdf-d25441461 (last access 25.07.2012), 13 pp. (shortened English version).
Šaulienė, I., & Veriankaitė, L. (2006). Application of backward air mass trajectory analysis in evaluating airborne pollen dispersion. Journal of Environmental Engineering and Landscape Management, 14(3), 113–120.
Sauliene, I., Veriankaite, L., & Lankauskas, A. (2007). The analysis of the impact of long distance air mass to airborne pollen concentration. Cross-border cooperation in researches of biological diversity. Acta Biologica Universitatis Daugapiliensis Supplement, 1, 61–74.
Savelieva, L. A., Dorozhkina, M. V., & Pavlova, E. Y. (2002). Modern annual deposition and aerial pollen transport in the Lena Delta. Polarforschung, 70, 115–122.
Schäppi, G. F., Suphioglu, C., Taylor, P. E., & Knox, R. B. (1997). Concentrations of the major birch tree allergen Bet v 1 in pollen and respirable fine particles in the atmosphere. The Journal of Allergy and Clinical Immunology, 100, 656–661. doi:S0091-6749(97)70170-2.
Schlesinger, P., Mamane, Y., & Grishkan, I. (2006). Transport of microorganisms to Israel during Saharan dust events. Aerobiologia, 22, 259–273. doi:10.1007/s10453-006-9038-7.
Schmidt-Lebuhn, A. N., Seltmann, P., & Kessler, M. (2007). Consequences of the pollination system on genetic structure and patterns of species distribution in the Andean genus Polylepis (Rosaceae): A Comparative study. Plant Systematics and Evolution, 266, 91–103. doi:10.1007/s00606-007-0543-0.
Schueler, S., & Schlünzen, K. H. (2006). Modelling of oak pollen dispersal on the landscape level with a mesoscale atmospheric model. Environmental Modelling and Assessment, 11, 179–194.
Seibert, P., Kromp-Kolb, H., Balterpensger, U., Jost, D.T., Schwikowski, M., Kasper A., Puxbaum, H. (1994). Trajectory analysis of aerosol measurements at high alpine sites. In P. M. Borrel, P. Borrell, T. Cvitas, W. Seiler (Eds.), Transport and transformation of pollutants in the troposphere (pp. 689–693). The Hague: Academic.
Seibert, P., & Frank, A. (2004). Source-receptor matrix calculation with a Lagrangian particle dispersion model in backward mode. Atmospheric Chemistry and Physics, 4, 51–63. http://www.atmos-chem-phys.net/4/51/2004/ (last access 25.07.2012).
Seinfeld, J. H., & Pandis, S. N. (2006). Atmospheric chemistry and physics: From air pollution to climate change (2nd ed.). New York: Wiley Interscience, Wiley. 1203 pp.
Shahali, Y., Pourpak, Z., Moin, M., Zare, A., & Majd, A. (2009). Impacts of air pollution exposure on the allergenic properties of Arizona cypress pollens. Journal of Physics Conference Series, 151, 012027. doi: 10.1088/1742-6596/151/1/012027.
Sharma, C. M., & Khanduri, V. P. (2007). Pollen-mediated gene flow in Himalayan long needle pine (Pinus roxburghii Sargent). Aerobiologia, 23, 153–158. doi:10.1007/s10453-007-9056-0.
Shinn, E. A., Griffin, D. W., & Seba, D. B. (2003). Atmospheric transport of mold spores in clouds of desert dust. Archives of Environmental Health, 58, 498–504.
Šikoparija, B., Smith, M., Skjøth, C. A., Radišić, P., Milkovska, S., Šimić, S., & Brandt, J. (2009). The Pannonian plain as a source of Ambrosia pollen in the Balkans. International Journal of Biometeorology, 53, 263–272.
Siljamo, P., Sofiev, M., & Ranta, H. (2004a). An approach to simulation of long-range atmospheric transport of natural allergens: An example of birch pollen. In C. Borrego & A. -L. Norman, (Eds.), Air pollution modelling and its applications XVII (pp. 331–340). New York: Springer (2007). ISBN-10: 0-387-28255-6.
Siljamo, P., Sofiev, M., Ranta, H., Kalnina, L., & Ekebom, A. (2004b). Long-range atmospheric transport of birch pollen. Problem statement and feasibility studies. Proceedings of Baltic HIRLAM workshop, St. Petersburg, November 17–20, 2003. (pp. 100–103). HIRLAM publications, SMHI Norrkoping, Sweden.
Siljamo, P., Sofiev, M., Severova, E., Ranta, H., & Polevova, S. (2006). On influence of long-range transport of pollen grains onto pollinating seasons. Developments in Environmental Science, 6. DOI: 10.1016/S1474-8177(70)06074-3. C. Borrego & E. Renner (Eds.). Air pollution modelling and its applications XVIII, (pp. 708–716) Amsterdam: Elsevier.
Siljamo, P., Sofiev, M., & Ranta, H. (2007). An approach to simulation of long-range atmospheric transport of natural allergens: An example of birch pollen. In C. Borrego & A.-L. Norman (Eds.), Air pollution modeling and its applications XVII, (pp. 331–339). New York: Springer.
Siljamo, P., Sofiev, M., Ranta, H., Linkosalo, T., Kubin, E., Ahas, R., Genikhovich, E., Jatczak, K., Jato, V., Nekovar, J., Minin, A., Severova, E., & Shalaboda, V. (2008a). Representativeness of point-wise phenological Betula data observed in different parts of Europe. Global Ecology and Biogeography, 17(4), 489–502. doi:10.1111/j.1466-8238.2008.00383.x.
Siljamo, P., Sofiev, M., Severova, E., Ranta, H., Kukkonen, J., Polevova, S., Kubin, E., & Minin, A. (2008b). Sources, impact and exchange of early-spring birch pollen in the Moscow region and Finland. Aerobiologia, 24, 211–230. doi:10.1007/s10453-008-9100-8.
Siljamo, P., Sofiev, M., Linkosalo, T., Ranta, H., & Kukkonen, J. (2008c). Development and application of biogenic emission term as a basis of long-range transport of allergenic pollen. In C. Borrego & A. I. Miranda (Eds.), Air pollution modelling and its application XIX. NATO Science for Peace and Security Series C: Environmental Security (pp. 154–162). Dordrecht: Springer.
Skjøth, C. A., Hertel, O., & Ellermann, T. (2002). Use of the ACDEP trajectory model in the Danish nation-wide background monitoring programme. Physics and Chemistry of the Earth, 27, 1469–1477.
Skjøth, C. A., Sommer, J., Stach, A., Smith, M., & Brandt, J. (2007). The long-range transport of birch (Betula) pollen from Poland and Germany causes significant pre-season concentrations in Denmark. Clinical and Experimental Allergy, 37, 1204–1212.
Skjøth, C. A., Geels, C., Hvidberg, M., Hertel, O., Brandt, J., Frohn, L. M., Hansen, K. M., Hedegard, G. B., Christensen, J. H., & Moseholm, L. (2008a). An inventory of tree species in European essential data input for air pollution modelling. Ecological Modelling, 217, 292–304.
Skjøth, C. A., Sommer, J., Brandt, J., Hvidberg, M., Geels, C., Hansen, K. M., Hertel, O., Frohn, L. M., & Christensen, J. H. (2008b). Copenhagen – a significant source to birch (Betula) pollen? International Journal of Biometeorology, 52, 453–462.
Skjøth, C. A., Smith, M., Brandt, J., & Emberlin, J. (2009). Are the birch trees in Southern England a source of pollen in North London? International Journal of Biometeorology, 53, 75–86.
Skjøth, C. A., Smith, M., Sikoparija, B., Stach, A., Myszkowska, D., Kasprzyk, I., Radisic, P., Stjepanovic, B., Hrga, I., Apatini, D., Magyar, D., Páldy, A., & Ianovici, N. (2010). A method for producing airborne pollen source inventories: An example of Ambrosia (ragweed) on the Pannonian Plain. Agricultural and Forest Meteorology, 150, 1203–1210.
Smith, M., Emberlin, J., & Kress, A. (2005). Examining high magnitude grass pollen episodes at Worcester, United Kingdom, using back-trajectory analysis. Aerobiologia, 21(2), 85–94.
Smith, M., Skjøth, C. A., Myszkowska, D. A. U., Puc, M., Stach, A., Balwierz, Z., Chlopek, K., Piotrowska, K., Kasprzyk, I., & Brandt, J. (2008). Long-range transport of Ambrosia pollen to Poland. Agricultural and Forest Meteorology, 148, 1402–1411.
Smouse, P., Dyer, R. J., Westfall, R. D., & Sork, V. L. (2001). Two-generation analysis of pollen flow across a landscape. I. Male gamete heterogeneity among females. Evolution, 55, 260–271. doi:10.1111/j.0014-3820.2001.tb01291.
Sofiev, M., & Atlaskin, E. (2007). An example of application of data assimilation technique and adjoint modelling to an inverse dispersion problem based on the ETEX experiment. In C. Borrego & A. Norman (Eds.), Air pollution modelling and its application XVII (pp. 438–448). New York: Springer.
Sofiev, M., Siljamo, P., Ranta, H., & Rantio-Lehtimaki, A. (2006a). Towards numerical forecasting of long-range air transport of birch pollen: Theoretical considerations and a feasibility study. International Journal of Biometeorology, 50, 392–402.
Sofiev, M., Siljamo, P., Valkama, I., Ilvonen, M., & Kukkonen, J. (2006b). A dispersion modelling system SILAM and its evaluation against ETEX data. Atmospheric Environment, 40, 674–685. doi:10.1016/j.atmosenv.2005.09.069.
Sofiev, M., Bousquet, J., Linkosalo, T., Ranta, H., Rantio-Lehtimaki, A., Siljamo, P., Valovirta, E., & Damialis, A. (2009). Pollen, allergies and adaptation. Chapter 5. In K. Ebi, G. McGregor & I. Burton (Eds.), Biometeorology and adaptation to climate variability and change (pp. 75–107). ISBN 978-4020-8920-6, Dordrecht: Springer.
Sorensen, J. H. (1998). Sensitivity of the DERMA long-range Gaussian dispersion model to meteorological input and diffusion parameters. Atmospheric Environment, 32, 4195–4206.
Spieksma, F. T. M., van Noort, P., & Nikkels, H. (2000). Influence of nearby stands of Artemisia on street-level versus roof-top-level ratio’s of airborne pollen quantities. Aerobiologia, 16, 21–24.
Stach, A., Smith, M., Skjøth, C. A., & Brandt, J. (2007). Examining Ambrosia pollen episodes at Poznan (Poland) using back-trajectory analysis. International Journal of Biometeorology, 51, 275–286.
Stanley, R. G., & Linskens, H. F. (Eds.). (1974). Pollen: Biology – biochemistry – management. Berlin: Springer.
Stewart, G. A., & Holt, P. G. (1985). Submicronic airborne allergens. The Medical Journal of Australia, 143(9), 426–427.
Stohl, A., Forster, C., Frank, A., Seibert, P., & Wotawa, G. (2005). Technical note: The Lagrangian particle dispersion model FLEXPART version 6.2. Atmospheric Chemistry and Physics, 5, 2461–2474.
Stohl, A., Seibert, P., Arduini, J., Eckhardt, S., Fraser, P., Greally, B. R., Lunder, C., Maione, M., Mühle, J., O’Doherty, S., Prinn, R. G., Reimann, S., Saito, T., Schmidbauer, N., Simmonds, P. G., Vollmer, M. K., Weiss, R. F., & Yokouchi, Y. (2009). An analytical inversion method for determining, regional and global emissions of greenhouse gases: Sensitivity studies and application to halocarbons. Atmospheric Chemistry and Physics, 9, 1597–1620. www.atmos-chem-phys.net/9/1597/2009/.
Subba Reddi, C., Reddi, N. S., & Atluri Janaki, B. (1988). Circadian patterns of pollen release in some species of Poaceae. Review of Palaeobotany and Palynology, 54, 11–42.
Tampieri, F., Mandrioli, P., & Puppi, G. L. (1977). Medium range transport of airborne pollen. Agricultural Meteorology, 18, 9–20.
Taylor, D. A. (2002). Dust in the wind. Environmental Health Perspectives, 110, A80–A87.
Taylor, P. E., Flagan, R. C., Valenta, R., & Glovsky, M. M. (2002). Release of allergens as respirable aerosols: A link between grass pollen and asthma. The Journal of Allergy and Clinical Immunology, 109, 51–56.
Taylor, P. E., Flagan, R. C., Miguel, A. G., Valenta, R., & Glovsky, M. M. (2004). Birch pollen rupture and the release of aerosols of respirable allergens. Clinical and Experimental Allergy, 34, 1591–1596.
Taylor, P. E., Card, G., House, J., Dickinson, M. H., & Flagan, R. C. (2006). High-speed pollen release in the white mulberry tree, Morus alba L. sexual plant. Reproduction, 19, 19–24.
Taylor, P. E., Jacobson, K. W., House, J. M., & Glovsky, M. M. (2007). Links between pollen, atopy and the asthma epidemic. International Archives of Allergy and Immunology, 144, 162–170. doi:000103230 [pii] 10.1159/000103230 [doi].
Timmons, A., O’Brien, E., Charters, Y., Dubbels, S., & Wilkinson, M. (1995). Assessing the risks of wind pollination from fields of genetically modified Brassica napus ssp. oleifera. Euphytica, 85, 417–423.
Traidl-Hoffmann, C., Kasche, A., Menzel, A., Jakob, T., Thiel, M., Ring, J., & Behrendt, H. (2003). Impact of pollen on human health: More than allergen carriers? International Archives of Allergy and Immunology, 131, 1–13. doi:10.1159/000070428.
Van Campo, M., & Quet, L. (1982). Pollen and red dust transport from South to North of the Mediterranean area. Comptes Rendus des Seances de l’Academie des Sciences Serie III Sciences de la Vie, 295, 61–64.
Van de Water, P. K., & Levetin, E. (2001). The contribution of upwind pollen sources to the characterization of Juniperus ashei phenology. Grana, 40, 133–141.
Van de Water, P. K., Keever, T., Main, C. E., & Levetin, E. (2003). An assessment of predictive forecasting of Juniperus ashei pollen movement in the Southern Great Plains, USA. International Journal of Biometeorology, 48, 74–82.
Van de Water, P., Watrud, L. S., Lee, E. H., Burdick, C., & King, G. A. (2007). Long-distance GM pollen movement of creeping bentgrass using modeled wind trajectory analysis. Ecological Applications, 17(4), 1244–1256.
Veriankaité, L., Siljamo, P., Sofiev, M., Sauliené, I., & Kukkonen, J. (2010). Modelling analysis of source regions of long range transported birch pollen that influences allergenic seasons in Lithuania. Aerobiologia, 26, 47–62.
Vogel, H., Pauling, A., & Vogel, B. (2008). Numerical simulations of birch pollen dispersion with an operational weather forecast system. International Journal of Biometeorology, 52, 805–814.
von Wahl, P.-G., & Puls, K. E. (1989). The emission of mugwort polen (Artemisia vulgaris L.) and its flight in the air. Aerobiologia, 5, 55–63.
Vrtala, S., Grote, M., Duchene, M., van Ree, R., Kraft, D., Scheiner, O., & Valenta, R. (1993). Properties of tree and grass pollen allergens: Reinvestigation of the linkage between solubility and allergenicity. International Archives of Allergy and Immunology, 102, 160–169.
Waisel, Y., Ganor, E., Epshtein, V., Stupp, A., & Eshel, A. (2008). Airborne pollen, spores, and dust across the East Mediterranean sea. Aerobiologia, 24, 125–131.
Westbrook, J. K., & Isard, S. A. (1999). Atmospheric scales of biotic dispersal. Agricultural and Forest Meteorology, 97, 263–274.
WHO. (2003). Phenology and human health: Allergic disorders. Copenhagen: WHO Regional Office for Europe. 55 pp.
Wodehouse, R. P. (1935). Pollen grains. Their structure, identification and significance in science and medicine. New York: MacGraw-Hill.
Wotawa, G., De Geer, L.-E., Denier, P., Kalinowski, M., Toivonen, H., D’Amours, R., Desiato, F., Issartel, J.-P., Langer, M., Seibert, P., Frank, A., Sloan, C., & Yamazawa, H. (2003). Atmospheric transport modelling in support of CTBT verification – overview and basic concepts. Atmospheric Environment, 37, 1565–1573.
Wu, P. C., Tsai, J. C., Li, F. C., Lung, S. C., & Su, H. J. (2004). Increased levels of ambient fungal spores in Taiwan are associated with dust events from China. Atmospheric Environment, 38, 4879–4886.
Wynn-Williams, D. D. (1991). Aerobiology and colonization in Antarctica: The BIOTAS programme. Grana, 30, 380–393.
Yadav, S., Chauhan, M. S., & Sharma, A. (2007). Characterisation of bio-aerosols during dust storm period in N-NW India. Atmospheric Environment, 41, 6063–6073.
Zhang, W. Y., Arimoto, R., & An, Z. S. (1997). Dust emission from Chinese desert sources linked to variations in atmospheric circulation. Journal of Geophysical Research, 102(23), 28041–28147.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Sofiev, M. et al. (2013). Airborne Pollen Transport. In: Sofiev, M., Bergmann, KC. (eds) Allergenic Pollen. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-4881-1_5
Download citation
DOI: https://doi.org/10.1007/978-94-007-4881-1_5
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-4880-4
Online ISBN: 978-94-007-4881-1
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)