
Abstract
A lipopolysaccharide (LPS) is considered to be one of the major determinants of virulence expression and infection of virulent Coxiella burnetii. The LPSs from virulent phase I (LPS I) and from avirulent phase II (LPS II) bacteria were investigated for their chemical composition, structure and biological properties. LPS II is of rough (R) type in contrast to LPS I, which is phenotypically smooth (S) and contains a noticeable amount of two sugars virenose (Vir) and dihydrohydroxystreptose (Strep), which have not been found in other LPSs and can be considered as unique biomarkers of the bacterium. Both sugars were suggested to be located mostly in terminal positions of the O-specific chain of LPS I (O-PS I) and to be involved in the immunobiology of Q fever. There is a need to establish a more detailed chemical structure of LPS I in connection with prospective, deeper studies on mechanisms of pathogenesis and immunity of Q fever, its early and reliable diagnosis, and effective prophylaxis against the disease. This will also help to better understanding of host-pathogen interactions and contribute to improved modulation of pathological reactions which in turn are prerequisite for research and development of vaccines of new type. A fundamental understanding of C. burnetii LPS biosynthesis is still lacking. The intracellular nature of the bacterium, lack of genetic tools and its status as a selected agent have made elucidating basic physiological mechanisms challenging. The GDP-β-D-Vir biosynthetic pathway proposed most recently is an important initial step in this endeavour. The current advanced technologies providing the genetic tools necessary to screen C. burnetii mutants and propagate isogenic mutants might speed the discovery process.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
4.1 Introduction
Upon serial passage in an immunoincompetent host, virulent C. burnetii undergoes a modification of its LPS what is traditionally referred to as phase variation (Stoker and Fiset 1956; Ftacek et al. 2000). C. burnetii phase variation includes the non-reversible switch from the virulent phase I (S) LPS I, which has a full length O-PS I chain (Mayer et al. 1988; Skultety et al. 1998) to the avirulent phase II (R) LPS II, which is missing the O-PS I chain and the saccharides located in the LPS I outer core (Toman and Skultety 1996; Toman et al. 2009). Several studies have indicated differential protein expression between the small cell variant (SCV) and large cell variant (LCV) cells of the bacterium (McCaul et al. 1991; Heinzen and Hackstadt 1996; Seshadri et al. 1999; Coleman et al. 2004, 2007) but the structural studies of the LPSs isolated from these variants have not been performed thus far. Although the structure of lipid A proximal core region of LPS I, represented by LPS II, was well established (Toman and Skultety 1996), both composition and structural features of the LPS I outer core and O-PS I chain have not been fully defined thus far (Toman et al. 2009).
LPS I is of particular biological, immunological and medical significance. It plays an essential role in interaction between the microbe and host including pathogenicity and immunogenicity of the agent. It is capable of inducing antibody response and is considered to be a protective immunogen (Williams and Waag 1991; Gajdosova et al. 1994; Hussein et al. 2001). Thus, it has been reported recently that O-PS I and its epitopes play a critical role in the antibody response to C. burnetii (Slaba et al. 2003; Vadovic et al. 2005), are involved in both signal transduction pathways (Dellacasagrande et al. 2000a, b) and LPS trafficking (Pretat et al. 2009), and may help to provide protection when included in a subunit vaccine (Zhang et al. 2007). Furthermore, research that advances the objective of rapid and specific detection of the infectious pathogen and differentiation of the individual isolates is centered on the development of monoclonal antibodies (mAbs) directed against the unique sugars present in the O-PS I chain (Palkovicova et al. 2009). However, both LPS I and LPS II were shown to be poor endotoxins. Investigations of their endotoxin activities showed that they were up to 1,000-fold less endotoxic than LPSs of Escherichia coli and Salmonella typhimurium (Amano et al. 1987). In this chapter, the structure, genetics, biosynthesis, and biological properties of the C. burnetii LPS will be reviewed.
4.2 Lipopolysaccharide Overview
LPSs are complex macromolecules found in the Gram-negative bacteria cell walls that may form up to approximately 75% of the outer leaflet of their outer membranes. They are essential for the physical organization and function of these membranes, and thus, for bacterial growth and multiplication. They are targets for bacteriophages, possess binding sites for antibodies and non-immunoglobulin serum factors, and in this way, they are involved in specific recognition and elimination of bacteria by the host defense system (Alexander and Rietschel 2001). LPSs may prevent the activation of complement and uptake of bacteria by phagocytes, and thus, they play an important role in the bacterial virulence. It has been presented in numerous studies (Alexander and Rietschel 2001; Raetz and Whitfield 2002) that LPSs display a broad spectrum of biological activities such as lethal toxicity and pyrogenicity, up- and down- regulation of several biological processes and in this manner they can contribute to the pathogenic potential of many Gram-negative bacteria. Moreover, LPSs activate strongly B-cells, granulocytes and mononuclear cells being considered as potent immunostimulators. In addition, there has been ever increasing evidence on an extensive involvement of LPSs in certain signal transduction pathways (Takeda and Akira 2005; Lu et al. 2008).
LPSs of Gram-negative bacteria share the common structural features and consist usually of three structural domains: (1) lipid A, which is attached to the outer bacterial membrane of the cell wall via acyl tails and is responsible for the endotoxic properties of LPS, (2) a short non-repeating inner and outer core, which is attached to lipid A and extends outwardly, and (3) O-PS chain, which is usually composed of repeating saccharide units attached to the lipid A – core and is responsible for serological heterogeneity among bacterial isolates (Alexander and Rietschel 2001). Minimally for survival, Gram-negative bacteria require lipid A and one 3-deoxy-D-manno-oct-2-ulosonic acid (Kdo). Bacteria with only these two subunits exhibit a deep (R) LPS phenotype (Raetz and Whitfield 2002) while bacteria with lipid A – Kdo plus additional saccharide units but not O-PS are generally described as (R) LPS bacteria. In contrast, (S) LPS bacteria are characterized by a lipid A – core attached to a fully developed O-PS chain. In addition to numerous functions of (S) LPS mentioned above, it may serve also as a permeability barrier against external agents such as hydrophobic antibiotics and to help to maintain the structural integrity of the Gram-negative cell wall (Raetz and Whitfield 2002). Moreover, (S) LPS has been a defined virulence factor in many clinically relevant pathogens. For example, (R) LPSs of Vibrio cholerae mutants were impaired in the intestinal epithelia colonization in the mouse model, which reflected their increased sensitivity to complement and cationic peptides (Nesper et al. 2001). Studies with uropathogenic E. coli O75:K5 showed that the chemistry and length of wild type O-PS degraded the complement system by inhibiting the assembly of the membrane attack complex (Joiner 1988; Burns and Hull 1998). The increased O-PS length also decreased the sensitivity of E. coli to neutrophil bactericidal/permeability-increasing protein (Weiss et al. 1986). The (S) LPS of Shigella flexneri influenced the polar localization of its invasion protein and was essential for invasiveness and subsequent inter- and intracellular spread (Sandlin et al. 1995, 1996; Van den Bosch et al. 1997). Finally, experiments with the burned mouse model and corneal mouse model showed that Pseudomonas aeruginosa (S) LPS is a critical virulence factor required for colonization (Rocchetta et al. 1999).
The importance of LPS as a virulence factor for the intracellular pathogens is less clear. It was shown in Legionella spp. that a virulence-associated LPS epitope caused most community acquired legionellosis (Helbig et al. 1995) and a phase-variable expression of LPS contributed to virulence by mediating serum complement resistance (Luneberg et al. 1998). However, the O-acetyl groups that modify the Legionella O-PS, hypothesized to be critical for virulence, were shown to have no impact on serum resistance (Luck et al. 2001). Francisella tularensis blue strain (S) LPS was more virulent than gray strain (R) LPS, but the mechanism of action is still unknown (Gunn and Ernst 2007). Brucella can perturb bactericidal activity in macrophages by influencing the host cell response to its advantage through its LPS (Jimenez de Bagues et al. 2005). The C. burnetii phase II attenuation in the immunocompetent hosts may be explained by several observations. The phase II organisms are more susceptible to complement mediated lysis relative to the phase I organisms (Vishwanath and Hackstadt 1988). It has been shown that phase I C. burnetii can infect and propagate in human dendritic cells (DC) while inhibiting their ability to induce maturation or inflammatory cytokine production (Shannon et al. 2005). In the same study, phase II bacteria significantly induced DC maturation and a robust inflammatory response.
4.2.1 Lipopolysaccharide Phase Variation
The generally accepted definition of phase variation is the reversible on-off switching mechanism of cell surface structures under various environmental conditions (Van der Woude and Baumler 2004). Although most O-PS operons are expressed constitutively (Raetz and Whitfield 2002), mechanisms for the LPS phase variation have been described for several bacteria. For example, the O-PS of Yersinia enterocolitica O3 is regulated by a repressor gene located outside of the O-PS gene cluster which is expressed at 37°C but not at 25°C (Al-Hendy et al. 1991; Skurnik and Toivanen 1993). The Legionella pneumophila O-PS is modulated by an unstable 30 kb genetic element that when integrated into the chromosome produces full length O-PS, but when incorporated in its native plasmid express an (R) LPS phenotype (Luneberg et al. 2001). Although Haemophilus influenzae lacks O-PS, its LPS structure is a critical virulence factor and is modulated by slipped-stranded mispairing (Weiser et al. 1989). Additionally, growth phase dependent expression of RfaH, a transcriptional anti-termination factor, was shown to modulate O-PS in S. enterica serovar Typhi (Rojas et al. 2001; Nagy et al. 2008).
In contrast, C. burnetii phase variation is historically defined as a non-reversible switch from virulent phase I cells containing LPS I to avirulent phase II cells with largely modified LPS II (Stoker and Fiset 1956; Williams and Waag 1991). It has been proposed that the observed LPS modification proceeds through gradual reduction of O-PS I during the phase variation yielding the LPS II which is the only LPS present in the C. burnetii phase II cells (Diaz and Lukacova 1998). In contrast, a completely different insight into this complex phenomenon was brought by Ftacek et al. (2000) based on their study of both composition and structure of LPSs isolated during serial passage of C. burnetii in embryonated hen eggs. It is a well-known fact that LPS molecules isolated from most (S) strains/isolates of Gram-negative bacteria are heterogeneous in size. They may contain some (R) LPSs, and in several cases variously truncated LPS molecules (Keenleyside and Whitfield 1999). Therefore, it can be assumed that strains/isolates of Gram-negative bacteria contain cell populations that may express multiple LPS structures. As the similar observation was also made by Ftacek et al. (2000), it has been suggested that a redistribution of the existing LPS populations takes place during the C. burnetii phase variation due to an increasing prevalence of those cells in the whole cell population that express LPS molecules with truncated O-PS I and those being of (R) type. It appears that in an attempt to optimize the balance between the expenditure of energy for the synthesis of complete LPS I molecules and the need to resist the microbicidal activities of the phagolysosome of the host immune system, C. burnetii has evolved genetic mechanisms of the transition from an energy-demanding (phase I) to a lesser energy-demanding (phase II) states. Thus, the mutants or strains/isolates that prefer biosynthesis of the incomplete LPS I subclasses might have a growth advantage.
Investigations of the O-PS biosynthesis region in the phase II Nine Mile strain (NM II, RSA 439) revealed that a large group of the genes was deleted (Hoover et al. 2002; Denison et al. 2007). Nevertheless, some phase II isolates contained no apparent deletions (Denison et al. 2007). To our best knowledge, the phase variation mechanisms have not been elucidated by bioinformatics tools in C. burnetii thus far and similarly, no R → S transition has ever been observed in the C. burnetii phase II strains/isolates. Thus, molecular mechanisms influencing the LPS modifications during the C. burnetii phase variation remain unclear at present.
4.3 Structural Studies of a Lipopolysaccharide from C. burnetii
4.3.1 Composition and Structure of LPS II
The first, more detailed chemical study of LPS II reported that it had similar sugar composition to LPS I although with quantitative differences (Baca et al. 1980). The following studies showed that LPS II and LPS I differed in the composition of constituent sugars (Schramek and Mayer 1982; Amano and Williams 1984; Mayer et al. 1988). It has been suggested, however, that LPS II contains a “Kdo-like substance” in the lipid A proximal region that is different from the enteric Kdo typical for most bacterial LPSs. Moreover, the sugar sequence in the inner core region of LPS II was not fully characterized. In their initial study, Toman et al. (1993) confirmed the presence of enteric Kdo in LPS II. Later, methylation-linkage analysis of LPS II demonstrated the presence of three Kdo residues with a structural arrangement similar to that of many enterobacterial LPSs (Toman and Skultety 1994). From other sugar residues, two terminal D-mannoses (Man), 2- and 3,4-linked D-glycero-D-manno-heptoses (D,D-Hep) were established (Toman and Skultety 1996). The presence of the latter sugar was quite surprising as most bacterial LPSs contain L-glycero-D-manno-heptose as the constituent sugar of the inner core region. Fast atom bombardment (FAB)- and electrospray ionization (ESI)-mass spectrometries (MS) of the lipid A deprived LPS II established the sequence of its sugar residues (Toman and Skultety 1996). The chemical structure of lipid A proximal region of LPS II is shown in Fig. 4.1. The subsequent matrix assisted laser desorption ionization (MALDI) – MS measurements confirmed the previous results and established the molecular mass of LPS II to 2841.58 (Toman et al. 2003b).

Chemical structure of lipid A proximal region of LPS II from Coxiella burnetii in avirulent phase II. Hep D-glycero-D-manno-heptose, Kdo 3-deoxy-D-manno-oct-2-ulosonic acid, Man D-mannose
In the last decade, many (R) LPSs have been found in pathogenic wild type bacteria including those colonizing especially the mucosal surfaces of the respiratory and urogenital tracts, such as Neisseria meningitidis, N. gonorrhoeae, H. influenzae, Bordetella pertussis or Chlamydiaceae (Holst 1999; Caroff et al. 2000). At present, it is not known with certainty whether these bacteria are not able to biosynthetize (S) LPSs or whether the genes encoding synthesis of O-specific chains are suppressed or deleted by regulatory mechanisms induced by environmental cues (Burns and Hull 1998; Heinrichs et al. 1999; Holst 1999). However, occurrence of such (R) type LPSs in the pathogenic wild-type bacteria indicates that the O-chain cannot be considered as the only major factor of pathogenicity in the respective bacterium and that it is dispensable without deleterious effects for it.
4.3.2 Composition and Structure of LPS I
As mentioned earlier, LPS I plays an important role in the interaction of C. burnetii with host, its pathogenicity and immunogenicity and is considered to be a protective immunogen (Amano and Williams 1984; Williams and Waag 1991; Gajdosova et al. 1994; Hussein et al. 2001). Several structural studies of this polymer were reported in the past but the data obtained brought only limited information on its structural features (Schramek et al. 1985; Amano et al. 1987; Toman and Kazar 1991; Skultety et al. 1998). LPS I contains, in addition to the sugar residues found in LPS II and some frequently occuring sugars, two unusual sugar units in its O-PS I, namely virenose (Vir, 6-deoxy-3-C-methylgulose) and dihydrohydroxystreptose [Strep, 3-C-(hydroxymethyl)lyxose]. Both sugars have not been found in other bacterial LPSs and can be considered unique biomarkers of C. burnetii. The enantiomeric forms and ring conformations of both saccharides were established from the optical rotation and nuclear magnetic resonance (NMR) data (Toman et al. 1998). Vir was found to be the D-gulo enantiomer with the 4C1 ring conformation and Strep was shown to be the L-lyxo enantiomer also with the 4C1 conformation. However, a structural analysis of LPS I showed that Strep was present in the parent LPS I in a furanose form (Toman 1991). Therefore, it was suggested that a furanose to pyranose tautomerization took place in the course of the isolation procedure (Toman et al. 1998). Recently, candidate genes involved in the synthesis of Vir and Strep have been suggested (Hoover et al. 2002; Thompson et al. 2003) and their protein products NDP-hexose 3-C-methyltransferase TylCIII (CBU0691) and methyltransferase FkbM family (CBU0683) were identified by the proteomic analysis (Skultety et al. 2005). However, the presence of galactosaminuronyl-α-(1 → 6)-glucosamine structural motif reported to occur in O-PS I (Amano et al. 1987) could not be confirmed in our laboratory very recently. Methylation analyses of two polysaccharide fractions of LPS I revealed the presence of terminal Vir, Strep, and Man, 4-substituted Vir, 4-substituted Man, and 2,3- and 3,4-disubstituted D,D-Hep (Vadovic et al. 2005). From amino sugars, 4-substituted D-glucosamine (GlcN) was detected. The methylation data demonstrated the pyranose form of Vir, Man, D,D-Hep and GlcN, and the furanose form of Strep. It could be anticipated from previous investigations (Toman and Skultety 1996) that 2,3- and 3,4-disubstituted D,D-Hep, and two terminal Man were from the core region of LPS I and thus, other terminal and substituted sugars should be located in the O-PS I chain. It was suggested in earlier studies (Schramek et al. 1985; Amano et al. 1987; Toman 1991; Skultety et al. 1998) that Vir and Strep were located almost exclusively in terminal positions. However, the recent findings have indicated that this might be true only for Strep as Vir is also (1 → 4)-linked (Vadovic et al. 2005). Similarly, Man is present in terminal positions but about 23% of it could also be involved in 1 → 4 linkages. A faster progress in a more detailed structural characterization of O-PS I is hampered by the presence of several O-PS I populations differing one from another in size, shape, and chemical composition. Therefore, only a tentative structural arrangement of sugar residues in LPS I can be given at present as shown in Fig. 4.2.

Preliminary structural arrangement of the sugar residues of LPS I from Coxiella burnetii in virulent phase I. GalNAc N-acetyl-D-galactosamine, GlcNAc N-acetyl-D-glucosamine, Glc D-glucose, Hep D-glycero-D-manno-heptose, Kdo 3-deoxy-D-manno-oct-2-ulosonic acid, Man D-mannose, Strep dihydrohydroxystreptose [3-C-(hydroxymethyl)-L-lyxose], Vir virenose (6-deoxy-3-C-methyl-D-gulose). The sugars in bold are prevailing in the O-specific chain
In the past, the C. burnetii clonal derivative RSA 514 named “Crazy” (Cr) was isolated from the placental tissue of guinea pig infected with the phase I Nine Mile (NM I, RSA 493) strain for 343 days (Hackstadt et al. 1985). It reacted with phase I antiserum, but to a much lesser extent than the NM I strain, and displayed no reactivity with phase II antiserum. The isolated LPS Cr gave one band at about 14 kDa on SDS-PAGE in contrast to LPS I and LPS II (Toman et al. 2009). The compositional analysis revealed the presence of Man, Glc, D,D-Hep, Strep, and GlcN in a molar ratio 3.1:0.1:1.0:1.5:1.2, respectively. No Vir was found. Both composition and structure of LPS II were shown to be a result of large chromosomal DNA deletions in the phase II cells of NM I strain (Hoover et al. 2002). However, deletions in the variant RSA 514 were larger extending on both ends beyond the phase II deletion junctions. Surprisingly, structural complexity of the LPS Cr appeared to be intermediate to LPS I and LPS II. The reasons for this discrepancy are still unknown.
4.3.3 Composition and Structure of Lipid A
The lipid A portion of LPS is linked to the core oligosaccharide mostly via Kdo and serves as the hydrophobic anchor of LPS in the bacterial outer membrane. Lipid A is established as the endotoxic principle of bacterial LPSs and represents in terms of both chemical composition and structure the most conserved region in them (Alexander and Rietschel 2001; Raetz and Whitfield 2002). Lipid A plays a major role in the pathogenesis of bacterial infections and is important contributor to massive inflammation, sepsis, and septic shock leading to fatalities in Gram-negative bacteria infections. It also promotes the activation of the innate immune system via induction of inflammatory cytokines released by human cells (Alexander and Rietschel 2001). Structurally, lipid A is typically composed of a β-D-GlcN-(1 → 6)-α-D-GlcN disaccharide backbone carrying two phosphate groups at positions O-1 and O-4′. Both phosphates can be further substituted with groups such as ethanolamine, ethanolamine phosphate, ethanolamine diphosphate, GlcN, 4-amino-4-deoxy-L-arabinopyranose, and D-arabinofuranose (Rietschel et al. 1987; Vadovic et al. 2007). There are attached up to four acyl chains by ester or amide linkages to the GlcN disaccharide. These chains can then in turn be substituted by further fatty acids to provide the lipid A moieties with up to seven acyl substituents, which vary quite considerably among species in the nature, number, length, order, and saturation (Alexander and Rietschel 2001).
It appears that the major contributing factors to endotoxicity are the number and lengths of acyl chains present and the phosphorylation state of the disaccharide backbone. For example, the structures with only one phosphate at either O-1 or O-4′ appear in most assays to be ∼1,000-fold less active than the highly endotoxic lipid A of E. coli as it was found in the naturally occurring monophosphorylated Bacteroides fragilis lipid A (Rietschel et al. 1987; Alexander and Rietschel 2001). However, phosphates by themselves do not appear to be essential as the substitution with the phosphono-oxyethyl group does not alter activity of the compound, suggesting that only correctly placed negative charges can restore activity (Ulmer et al. 1992; Erridge et al. 2002). The GlcN monosaccharide preparations phosphorylated and acylated in various positions lack activity in general, suggesting that the disaccharide backbone is also required for the optimum recognition by the humoral/cellular receptors (Aschauer et al. 1990). Nevertheless, lipids A with 2,3-diamino-2,3-dideoxy-D-glucose replacing GlcN in the backbone have similar activity than those having GlcN as seen, e.g. in Campylobacter jejuni (Moran et al. 1991; Alexander and Rietschel 2001).
Naturally, much work was concentrated on the role of the nature, number and lengths of acyl chains attached to the lipid A. For example, structures similar to that of the E. coli lipid A with two phosphates, but with seven or five fatty acids were less active by a factor of approximately 100 (Rietschel et al. 1993; Erridge et al. 2002). Thus, it appears that the hexaacylated lipid A species consisting of two 3-hydroxyacyl and two 3-acyloxyacyl residues limited in length of individual acid chains to 12 or 14 carbons of either E. coli or N. meningitidis like – types are optimally recognized by the mammalian receptors to express the full spectrum of endotoxic activities (Alexander and Rietschel 2001; Erridge et al. 2002).
Wollenweber et al. (1985) analyzed the fatty acid composition in the C. burnetii lipids A from LPS I and LPS II. The authors identified more than 50 different 3-acyloxyacyl residues to be involved in the amide linkage, and together with other fatty acids found, suggested an enormous heterogeneity of both lipid A components. In contrast, Toman et al. (2003a) could not find any 3-acyloxyacyl residues in the investigated lipids A from the LPSs I of Henzerling and S strains, and in addition, their fatty acid compositions were considerably less complex than those published by Wollenweber et al. (1985). The authors showed that both lipids A were tetraacylated with two amide-linked 3-hydroxy and two ester-linked nonhydroxylated fatty acids attached to the GlcN dissaccharide. Differences were found in the small structural details only. Further, in the LPS I of Priscilla strain, two tetraacylated lipid A species were found as the major components despite a noticeable microheterogeneity of the analyzed sample (Toman et al. 2004). They possessed the classical backbone of diphosphorylated GlcN disaccharide, in which both reducing-end GlcN I and terminal GlcN II carried amide-linked iso-branched or normal (n) 3-hydroxyhexadecanoic fatty acids. One of the species had ester-linked n-hexadecanoic acids at both GlcNs while the other had ester-linked anteiso-branched pentadecanoic instead of n-hexadecanoic acid at GlcN II. The lipid A moiety had a cubic inverted aggregate structure, and the inclination angle of the GlcN disaccharide backbone plane of the lipid A part with respect to the membrane normal was around 40°. The lipid A readily intercalated into phospholipid liposomes mediated by the LPS-binding protein. The lipid A-induced tumor necrosis factor (TNF) production in the human mononuclear cells was one order of magnitude lower than that found for lipid A of S. minnesota. In another study (Zamboni et al. 2004), an asymmetrical structure of the C. burnetii lipid A was suggested where a 3-acyloxyacyl chain, alternatively composed of 3-hydroxytetra- to hexadecanoic acids, was attached to the amide-linked 3-hydroxylated fatty acid at the N-2′ of the GlcN II unit. However, the proposed structure needs further verification.
It was reported (Alexander and Rietschel 2001) that variation of the lipid A domain of LPS serves as one strategy utilised by Gram-negative bacteria to promote survival by providing resistance to components of the innate immune system and helping to evade recognition by Toll-like receptor 4 (TLR 4). Thus, it was of interest to see if the long-term survival of C. burnetii in host led to modifications in its lipid A in comparison with the known structures for lipids A from the strains Henzerling, S and Priscilla (Toman et al. 2003a, 2004). The recent study of lipid A from the clonal derivative RSA 514 has revealed no substantial modifications in the basic chemical composition and structure proposed for lipid A of C. burnetii (Vadovic et al. 2009). It appears that lipids A from various strains/isolates of C. burnetii represent tetraacylated molecular species with the conserved basic structural features. Some differences are found in nature, length and possibly also in distribution of fatty acids along the GlcN disaccharide. These fatty acid variations contribute to a remarkable diversity of the C. burnetii lipid A although its biochemical synthesis is a highly conserved process like in other Gram-negative bacteria. Nevertheless, further studies are needed to confirm symmetrical (Toman et al. 2003a, 2004; Vadovic et al. 2009) or asymmetrical (Zamboni et al. 2004) structural model of the lipid A. The proposed structural features of C. burnetii lipid A are shown in Fig. 4.3.

Basic chemical structure of lipid A of Coxiella burnetii. Models with symmetrical R1 = H3C–(CH2)n–CH2–CH2–CO or H3C–(CH2)n–CH(OH)–CH2–CO, R2 = H (Toman et al. 2003a; Toman et al. 2004; Vadovic et al. 2009) and asymmetrical R1 = H, R2 = H3C–(CH2)n–CH(OH)–CH2–CO (Zamboni et al. 2004) structures have been proposed; n = 12–14, X = C14–16
Distinct structural features of the C. burnetii lipid A might be the reason for its lower endotoxic activity as compared to the classical forms of enterobacterial lipids A, found, e.g. in E. coli, N. meningitidis or Salmonella spp. Nevertheless, C. burnetii and its LPS were shown to exhibit various immunomodulatory activities despite their low endotoxic activity (see below). This behavior could be elucidated by applying the previously presented conformational concept of endotoxicity (Seydel et al. 1999), a conical shape of the lipid A of C. burnetii and a sufficiently high inclination of the sugar backbone plane with respect to the membrane (Toman et al. 2004).
4.4 Biosynthesis of Lipopolysaccharides
4.4.1 Nomenclature
The complex biochemistry of LPS biosynthesis and expression causes confusion when naming and studying LPS related genes. Contributing factors that add to the confusion include different names for duplicated genes located in different loci of a bacterium, different names for the same gene in different bacteria or the same name for genes that are not related in different bacteria. Additionally, some contemporary LPS biosynthesis investigators still use nomenclature that reflects the historical assignment of a gene by the Demerec System (Demerec et al. 1966). To help to resolve the limitations inherent to the Demerec System, a new system was proposed by Reeves et al. (1996) known as the Bacterial Polysaccharide Synthesis and Gene Nomenclature (BPGN). This system expanded the number of gene possibilities from 26 (number of letters in the alphabet) to 17,576 possibilities and is organized by function. Since both Demerec and BPGN nomenclature systems are currently in use, the usage of gene names in this chapter will reflect those which are most frequently found in literature.
4.4.2 O-Polysaccharide Genetics
The structural heterogeneity observed in various O-PS chains correlates to the genetic variation found within their encoding loci (Raetz and Whitfield 2002). Despite this variation the clusters themselves tend to be highly organized (Stroeher et al. 1998; Trefzer et al. 1999; Luneberg et al. 2000; Raetz and Whitfield 2002; King et al. 2009). Present just upstream of many polysaccharide gene clusters is a 39-bp JUMPstart/Operon Polarity Suppressor site that is involved in transcriptional anti-termination shown to require RfaH (Hobbs and Reeves 1994; Wang et al. 1998). This finding is consistent with a hypothesis that the O-PS clusters are transcribed as a single locus, which may contain 7 to over 17 genes (Raetz and Whitfield 2002). In E. coli and S. enterica, the O-PS locus is usually flanked by his and gnd (Raetz and Whitfield 2002) while in Pseudomonas spp. is flanked by ihfB and wbpM (Rocchetta et al. 1999). It has been suggested that these flanking genes are target sites for the lateral transfer of O-PS clusters (Bisercic et al. 1991; Nelson and Selander 1994). Further supporting this hypothesis is the observation that the O-PS gene clusters typically have a GC content lower than the genome average (Samuel and Reeves 2003).
The currently accepted organizational paradigm for genes located within the O-PS locus, referred to as wb (BPGN) or rfb (Demeric), is by function: (1) pathway genes, (2) processing genes and (3) transferase genes. Pathway genes include those genes involved in the activated carbohydrate biosynthesis. These genes share a high degree of identity among a wide range of species (Trefzer et al. 1999; Raetz and Whitfield 2002; Samuel and Reeves 2003). Processing genes encode proteins that mediate O-PS transport from the cytosol to the periplasm and are usually adjacent within the wb locus (Raetz and Whitfield 2002). Transferase genes catalyze the sequential attachment of sugar units to the growing O-PS chain. The glycosyltransferases are often dispersed throughout the wb locus and have low levels of similarity with one notable exception, wecA (rfe, Demeric) (Raetz and Whitfield 2002). The wecA gene encodes a glycosyltransferase responsible for the transfer of the first saccharide called the primer saccharide to the hydrophobic lipid carrier molecule, undecaprenol phosphate (und-P), in E. coli. The initial transfer of the primer saccharide to und-P is required for the O-PS elongation in the cytosol (Raetz and Whitfield 2002). Unlike other transferases, wecA is located outside of the wb locus. Bioinformatic analysis of various WecA glycosyltransferases showed a high degree of similarity and it was once thought they might have wide substrate specificity when compared to other glycosyltransferases (Raetz and Whitfield 2002). However, recent evidence has shown that WecA is specific for only GlcNAc transfer to und-P (Rush et al. 2010).
4.4.3 O-Polysaccharide Genetic Analysis of C. burnetii
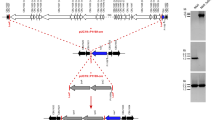
As mentioned earlier, the clonal derivative Cr had a large chromosomal deletion (25,997 bp) that overlapped the NM II deletion. The deletion should correspond to the loss of Vir in this and also the NM II strain and should explain the nonreversible phase variation observed with the NM strain (Vodkin and Williams 1986; Hoover et al. 2002). However, it does not explain the presence of LPS II in the NM II strain that lacks also Strep and the outer core saccharides. Furthermore, a recent study has shown that several phase II C. burnetii strains/isolates have a functional C. burnetii wb locus but still exhibit the (R) LPS phenotype (Denison et al. 2007). This finding might suggest that genes outside of the deleted region are required for both O-PS I and the outer core biosynthesis.
Further comparative genomic analysis revealed three regions that might contain genes necessary to express O-PS I in LPS I. Region 1 (605781–653889) includes the deleted region of the strains NM I (RSA 493) and Cr (RSA 514) defined by Hoover et al. (2002) and may contain the genes needed for GDP-β-D-Vir biosynthesis. Region 2 (775635–813578) may contain those genes needed for the expression of sugars constituting the O-PS I backbone, and region 3 (1760021–1768371) may contain the genes needed for the expression of Strep (Seshadri et al. 2003). Within the region 1, the genes that are normally associated with a wb operon, like genes for an activated saccharide biosynthesis, the glycosyl transferase genes and processing genes, have been identified (Hoover et al. 2002). Just outside of the NM II (RSA 439) and Cr (RSA 514) deleted regions, there are two adjacent genes that code for an ABC transporter system, O-antigen export system permease protein (CBU0703) and LPS/O-antigen export permease (CBU0704). The ABC transporter dependent pathways typically transport the linear O-PS structures versus branched structures characteristic of the Wzy dependent pathway (Raetz and Whitfield 2002). This evidence might support a hypothesis that the O-PS I in LPS I is essentially a linear polymer. However, this hypothesis could not be confirmed by the glycomic studies mentioned above. An exhaustive bioinformatic search has failed to identify any of the genes necessary for the Wzy dependent pathway thus far.
Although the GC content of the deleted region is slightly lower than the chromosomal GC content (43% in RSA 493 and 39% in the deleted region of RSA 439), there are no apparent target sites where the lateral transfer might occur. The bioinformatic analysis indicates no JUMPstart/ops sequence and several genes, some divergently transcribed, that may not play a role in the O-PS I biosynthesis.
4.4.4 O-Polysaccharide Biosynthesis and Export Mechanisms of C. burnetii
Three O-PS biosynthesis pathways have been described: (1) ABC-transporter dependent, (2) Wzy dependent and (3) synthase dependent. Each pathway initiates the O-PS biosynthesis by attaching a sugar-1-phosphate catalyzed by a membrane bound glycosyltransferase to und-P. The model glycosyltransferases are WecA in E. coli and WbaP in Salmonella spp., which transfer GlcNAc and D-galactose to und-P, respectively (Raetz and Whitfield 2002). Nevertheless, other initial glycosyltransferases have been shown to initiate the O-PS biosynthesis. For example, the Pseudomonas aeruginosa WbpL catalyzes the transfer of N-acetyl-D-fucosamine (D-FucNAc) and N-acetyl-D-quinovosamine (D-QuiNAc) to und-P (King et al. 2009). It is likely that other initial glycosyltransferases exist but have not been characterized thus far. For example, the unique primer saccharide legionaminic acid could be transferred to und-P by an undefined enzyme in L. pneumophila (Knirel et al. 1994).
Structural studies indicated that GlcNAc was present in the O-PS I of LPS I from the NM I strain (Toman and Kazar 1991), and GlcNAc and GalNAc were both identified in the O-PS I of LPS I from the C. burnetii strain Priscilla in phase I (Hussein et al. 2001). Additionally, C. burnetii undecaprenyl-phosphate α-N-acetylglucosamine phosphotransferase (CBU0533) has a strong sequence homology to the characterized WecA in the model organisms. Taken together, we hypothesized that GlcNAc or GalNAc could be the initial saccharides in the O-PS I chain and CBU0533 might catalyze their attachment to und-P. To prove this hypothesis, we constructed an E. coli O157:H7 ZAP198 ΔwecA mutant by a modified protocol described by Emmerson et al. (2006). The mutant was verified genotypically by polymerase chain reaction and phenotypically by silver staining and immunoblot. The wild type E. coli O157:H7 LPS was restored to the ZAP198 ΔwecA mutant when the native wecA was expressed in trans. However, despite the bioinformatic similarity between CBU0533 to characterized WecAs, it failed to complement the ZAP198 ΔwecA mutant in trans.
Several possibilities may explain this observation: (1) CBU0533 does not catalyze the transfer of GlcNAc or GalNAc to und-P, (2) it is not active in E. coli O157:H7, or (3) the E. coli O157:H7 lipid carrier molecule is not a CBU0533 substrate. It is well known that the lipid carrier molecules are used in both eukaryotic and prokaryotic cells to transport glycans across the lipid bilayers. The most characterized prokaryotic lipid carrier molecules have 11 isoprene units, 3 in trans and 8 in cis positions for a total of 55 carbons (C55). Thus, the name “undecaprenyl” phosphate derives from this finding (Raetz and Whitfield 2002). Many laboratories have shown that both length of polyisoprenol carrier molecule and nature of α-isoprene unit drive the specificity and efficiency of the initial glycosyltransferase reaction in eukaryotes (Szkopinska et al. 1992; Rush et al. 1993; D’Souza-Schorey et al. 1994; Kean et al. 1994; McLachlan and Krag 1994; Dotson et al. 1995) and in prokaryotes (Rush et al. 1997; Chen et al. 2007). An in vitro evidence from the Rocky Mountain Laboratory (RML) in Hamilton, MT, has shown that CBU0533 does not transfer GlcNAc to und-P (unpublished data). In light of the bioinformatic analysis and experimental work performed in our laboratory and at RML, we have concluded that the C. burnetii RSA 493 CBU0533 encodes a wecA-like gene. Future studies on the O-PS I biosynthesis in LPS I should focus on the attachment of the first monosaccharide to the lipid carrier molecule.
After attachment of the primer saccharide to und-P, the various O-PS pathways differ in their polymerization and transport mechanisms. In the ABC transporter pathway, rather than adding O-PS units to a growing O-PS chain in the periplasm, the O-PS is completely formed at the inner leaflet of the inner membrane by a series of glycosyltransferase reactions. This pathway is found in those bacteria that generate the O-PS homopolymers while heteropolymers are usually synthesized via the Wzy pathway (Raetz and Whitfield 2002). The completed O-PS subsequently transits to periplasm via the ABC transporter system (encoded by wzm and wzt) where it is ligated to the lipid A – core by WaaL (Raetz and Whitfield 2002). How modal distribution is achieved in the ABC transporter dependent pathway is not fully understood. However, non-reducing terminal modifications have been shown to regulate the O-PS length in E. coli and M. smegmatis and may signal for its transport through its ABC transporter (Weisman and Ballou 1984; Clarke et al. 2004).
It should be mentioned here that the ABC transporters are a large class of importers and exporters observed in both eukaryotic and prokaryotic cells. Because these transporters are important in the nutrient uptake and drug resistance, they are the subject of ongoing research (Hollenstein et al. 2007). The ABC transporters that move O-PS from cytosol to periplasm are not as well characterized. However, the fundamental transport mechanism appears to be conserved. Wzm is a membrane bound protein that facilitates the O-PS transport through the inner membrane (Raetz and Whitfield 2002). The C. burnetii O-antigen export system permease protein (CBU0703) is adjacent to the putative O-PS I biosynthesis genes and is annotated as wzm. The hydropathy plots using the Kyte-Doolittle scale (Bowen 2008 ) of CBU0703 detected hydrophobic regions typical of the characterized Wzm proteins. The C. burnetii LPS/O-antigen export permease (CBU0704) is annotated as wzt. Bioinformatic analysis has suggested that the C. burnetii Wzt has a classical ATP-binding cassette motif for which the transport system is named. This enzyme is responsible for binding the und-P linked O-PS I, and providing the machinery required for transport at the expense of ATP hydrolysis.
4.4.5 Tentative Biosynthesis Pathway of D-Virenose in C. burnetii
A fundamental understanding of O-PS I biosynthesis in LPS I is still lacking. The intracellular nature of C. burnetii, lack of genetic tools and its status as a potential biowarfare agent have made elucidating of its basic physiological mechanisms challenging. Most recently, an attempt has been made to elucidate the enzymatic steps responsible for the formation of guanosine-5′-diphosphate (GDP)-β-D-Man in the bacterium, which were bioinformatically predicted as the initial steps of GDP-β-D-Vir biosynthesis (Narasaki et al. 2011). Based on both structural determination of Vir (Toman et al. 1998) and analysis of the genes deleted in the RSA 439 variant (Hoover et al. 2002), it has been hypothesized that GDP-β-D-mannose is synthesized from fructose-6-phosphate (F6P) in three successive reactions that include: (i) isomerization to D-mannose-6-phosphate (M6P) catalyzed by a mannose-6-phosphate isomerase (PMI), (ii) conversion to mannose-1-phosphate (M1P) mediated by a phosphomannomutase (PMM) and (iii) addition of GDP by a GDP-mannose pyrophosphorylase (GMP) (Fig. 4.4). GDP-β-D-mannose is then likely converted to GDP-6-deoxy-β-D-lyxo-hex-4-ulopyranose, a Vir intermediate, by a GDP-D-mannose dehydratase (GMD). To prove the suggested pathway in C. burnetii, three ORF (CBU0671, CBU0294 and CBU0689) annotated as bifunctional type II PMI, as PMM or GMD were functionally characterized by complementation of corresponding E. coli mutant strains and in enzymatic assays. However, CBU0671 failed to complement an E.coli manA (PMM) mutant strain but complementation of manC (GMP) mutant strain restored the capsular PS biosynthesis. CBU0294 complemented a P. aeruginosa algC (GMP) mutant strain and showed phosphoglucomutase activity in a pgm E. coli mutant strain. Despite the inability to complement a manA mutant, recombinant C. burnetii PMI protein showed PMM enzymatic activity in biochemical assays. CBU0689 showed dehydratase activity and determined kinetic parameters were in agreement with the data reported from other organisms.

Schematic presentation of putative GDP-β-D-virenose biosynthesis pathway. Ado-Met S-adenosyl methionine, F6P fructose-6-phosphate, GDP guanosine-5′-diphosphate, GFS fucose synthetase (GDP-4-keto-6-deoxymannose epimerase/reductase), GMD GDP-D-mannose dehydratase, GMP GDP-D-mannose pyrophosphorylase, GTP guanosine-5′-triphosphate, M1P D-mannose-1-phosphate, M6P D-mannose-6-phosphate, MET methyltransferase, NADP nicotinamide adenine dinucleotide phosphate, PES perosamine synthetase, PMI mannose-6-phosphate isomerase, PMM phosphomannomutase, PP i inorganic pyrophosphate
GDP-6-deoxy-β-D-lyxo-hex-4-ulopyranose formed by GMD is the metabolic intermediate of GDP-L-fucose, GDP-colitose, GDP-perosamine, GDP-D-rhamnose and GDP-6-deoxy-D-talose (Samuel and Reeves 2003). The enzymes required to generate the final steps for GDP-perosamine (perosamine synthetase, CBU0830) and GDP-L-fucose (fucose synthetase, GDP-4-keto-6-deoxymannose epimerase/reductase, CBU0688) have been identified in the C. burnetii genome (Seshadri et al. 2003). However, we have not detected any perosamine or L-fucose in the compositional analyses of LPSs I obtained from various strains/isolates of C. burnetii. Nevertheless, the suggested biosynthesis pathway for Vir has established an initial step towards a better understanding of the O-PS I formation, which is the only known virulence factor of C. burnetii.
4.5 Functional Characteristics of the C. burnetii Lipopolysaccharide
TNF is a cytokine involved in systemic inflammation and is a member of a group of cytokines that stimulate the acute phase reaction. It is a protein with a molecular mass of 17 kDa and is produced mainly by mononuclear phagocytes following induction with an LPS (Flebbe et al. 1990). The primary role of TNF is in the regulation of immune cells. It is able to induce apoptotic cell death, to induce inflammation, and to inhibit tumorigenesis and viral replication. Dysregulation of TNF production has been implicated in a variety of human infectious diseases including Q fever (Locksley et al. 2001; Capo et al. 1999). Although LPS I and LPS II are weak endotoxins their ability to induce TNF was reported (Toman et al. 2004). TNF and IL-1β production is increased in monocytes from patients with Q fever endocarditis, whereas it remains low in patients with uncomplicated acute Q fever (Capo et al. 1996; Dellacasagrande et al. 2000a). Thus, the TNF production seems to be related to the disease activity. However, the TNF production does not directly reflect the virulence of C. burnetii since avirulent phase II bacteria were also shown to induce this cytokine (Tujulin et al. 1999). In fact, avirulent bacteria were even more potent than virulent phase I organisms at stimulating the TNF production (Dellacasagrande et al. 2000b). It has already been reported in this chapter that lipids A of LPS I and LPS II have almost identical chemical structures, and therefore, other factors should be involved in this phenomenon. One could hypothesize that presence or absence of O-PS I in these LPSs might play a role. However, it appears more likely that a limited efficiency of TNF production is not a consequence of poor activity of LPS I as this has been shown to be more potent than LPS II at stimulating the TNF release (Dellacasagrande et al. 2000b). It has been suggested that virulent bacteria, which bind poorly to monocytes, likely present fewer LPS I molecules to the target cells than do avirulent ones that efficiently bind to monocytes. The effect of activation on the uptake efficiency was also supported by the finding that neutralizing anti-TNF antibodies decrease the C. burnetii internalization by monocytes from patients with Q fever endocarditis but do not affect the long term survival of bacteria (Dellacasagrande et al. 2000a). Moreover, a neutralization of TNF with specific antibodies prevented macrophage apoptosis and the eradication of C. burnetii (Dellacasagrande et al. 1999). Recently, the strain specific differences in the TNF induction have been published (Kubes et al. 2006). The highest TNF production was monitored with both cells and LPSs of the C. burnetii strains Scurry and Priscilla. Both strains were shown to be genetically very close by the multiple locus variable number tandem repeats analysis (Svraka et al. 2006). On the contrary, previous SDS-PAGE and immunoblot analyses of their LPSs I indicated that these might be structurally and antigenically distinct (Hackstadt 1986; Skultety et al. 1998).
Macrophages, DCs and neutrophils sense invading microbial pathogens by so-called pattern recognition receptors (PRRs) to activate rapidly the innate host defence system and to promote triggering of the whole array of adaptive immune responses (O’Neill 2006; Kawai and Akira 2008). Toll-like receptors (TLRs) are the principal membrane-associated innate sensors that recognize conserved pathogen associated molecular patterns (PAMPs) at the cell surface or in the intracellular endosomal compartments (Doyle and O’ Neill 2006; O’Neill 2006). TLR 4, the predominant signal transduction receptor for the Gram-negative bacterial LPSs, is critical for the host defence against Gram-negative bacterial pathogens (Alexander and Rietschel 2001; Takeda and Akira 2005). In addition to LPS, TLR 4 recognizes other structurally unrelated microbial PAMPs like chlamydial heat shock proteins 60 and 70 (Bulut et al. 2002) and pneumolysin (Malley et al. 2003). MD-2, an extracellular protein, is essential for conferring LPS sensitivity to TLR4 (Shimazu et al. 1999), and CD14 enhances LPS responses by facilitating LPS binding to MD-2, enables MyD88-independent signaling pathways by LPS. TLR 2 has been shown to recognize lipoproteins and lipopeptides from Gram-positive bacteria, mycoplasma, and mycobacteria (Lien et al. 1999) in cooperation with TLR 1 or TLR 6 (Hajjar et al. 2001; O’Neill 2006). Recent results suggest, however, that TLR 4 and TLR 2 polymorphisms are associated with susceptibility to infectious diseases (Lorenz et al. 2000; Awomoyi et al. 2007).
TLR 4 was reported to be involved in the recognition of LPS I, initial activation of macrophages, and inflammatory response associated with the C. burnetii infection (Honstettre et al. 2004). It was shown later, however, that lipid A from C. burnetii antagonized activation of TLR 4 by the highly endotoxic lipids A from E. coli and B. pertussis, and by the E. coli LPS (Zamboni et al. 2004). A similar competitive inhibition of LPS activation of the TLR 4-dependent responses was reported for other lipids A, such as those isolated from Rhodobacter sphaeroides and lipid IVA (Golenbock et al. 1991). This fact appeared to be in agreement with observations that LPSs of some bacterial species failed to activate TLR 4 and that these bacteria activated host cells through a TLR 2-dependent mechanism (Hirschfeld et al. 2001; Girard et al. 2003; Darveau et al. 2004). It has already been mentioned in this chapter that the chemical structure of C. burnetii lipid A differs considerably from those reported for the classical enterobacterial lipids A that activate predominantly TLR 4. The recent findings in the field have indicated that TLR 4 is involved in the cytoskeleton remodeling induced by C. burnetii, controls the immune response against it through granuloma formation and cytokine production, and is dispensable for the bacterial clearance in vivo (Honstettre et al. 2004). TLR 2 seems to be also involved in the TNF and IFN-γ production induced by avirulent variants of C. burnetii in macrophages from TLR 2-deficient mice (Zamboni et al. 2004). Moreover, these macrophages have been shown to be highly permissive for the intracellular growth of bacteria. Experiments in vivo have also indicated that TLR 2 is involved in the granuloma formation similar to that of TLR 4 (Meghari et al. 2005). Thus, a synergistic action of both TLRs has been proposed in some signal transduction pathways. However, a more detailed role of LPS I and LPS II in these events remains to be elucidated.
It has already been reported that phase I C. burnetii replicated within the human DCs without inducing maturation or inflammatory cytokine production (Shannon et al. 2005). In contrast, the phase II bacteria induced a dramatic maturation of the cells. A model has been proposed whereby LPS I masks TLR ligands from innate immune recognition by DCs, thereby allowing replication without significant maturation or inflammatory cytokine production. This immune evasion strategy was suggested to allow C. burnetii to persist in an immunocompetent host. However, these findings raised several points that are discussed in a more detail in the dedicated chapters of this book.
It has been well known for many years that during acute Q fever C. burnetii induces antibodies in host against its phase II (protein) antigens, while in the later stages of the disease, and especially in its persistent/chronic form, high titers of antibodies are directed against phase I (LPS I) antigen (Marrie and Raoult 1997). Initial work has already been performed towards elucidation of the interaction of phase I antibodies with the LPS I antigen (Vadovic et al. 2005). A noticeable decrease in the serological activity of the O-antigen was monitored when Vir and Strep were selectively removed from its O-PS I chain. However, it is still not known with certainty whether the immunoreactive epitopes are located only at both sugars in terminal positions or also at those Vir residues located in the O-PS I backbone.
The presence of unique biomarkers Vir and Strep in LPS I and their favourable location in its O-PS I provides an excellent opportunity to generate mAbs against them that would enable rapid, sensitive and specific detection of the virulent form of the bacterium. So far, a mAb (IgG2b subclass) has been generated that was proved to be highly specific for the presence of Vir in the bacterium and LPS I as only Vir-containing C. burnetii strains/variants and their LPSs reacted with the mAb (Palkovicova et al. 2009). In addition, no cross-reaction was observed with the cells and LPSs from the selected species of the families Rickettsiaceae, Piscirickettsiaceae, Chlamydiaceae and Enterobacteriaceae.
Most recently, the intracellular trafficking of LPS I and LPS II in murine bone marrow-derived macrophages has been investigated (Pretat et al. 2009). It has been found that the intracellular trafficking of LPS I is similar to that of LPS II and that the high portions of both LPSs have not used the endosomal route. It was proposed that the LPSs transit through non-classical endocytic or uncharacterized pathways. Moreover, the results suggest that LPS I may be involved in a subverted conversion of the C. burnetii phagosome by interfering with the endosomal pathway.
More information on the biological and functional characteristics/properties of the C. burnetii LPSs can be found in the dedicated chapters of this book.
4.6 Summary
Although the C. burnetii lipid A and inner core structures of LPS I are well characterized, the composition and structure of its outer core and O-PS I chain are not entirely resolved. More detailed investigations are necessary in this direction, and in addition, there is a need to better characterize the role of individual LPS I components in pathogenesis and immunity of Q fever. Thus, it is important to investigate effects of these biological response modifiers on both humoral and cellular immune systems. For example, knowledge on the structure/function relationship of the LPS I antigens involved in the induction of immunity and pathological reactions during the infection and administration of the experimental vaccines is not sufficient. A better understanding of both composition and structure of the biologically active outer membrane components (LPS I and proteins) of C. burnetii will be of crucial importance not only in a more detailed elucidation of host-pathogen interactions but also in modulation of pathological reactions, which in turn is a prerequisite for development of vaccines of new type.
The intracellular nature of C. burnetii, the lack of genetic tools and its status as a selected agent have made elucidating its basic O-polysaccharide biosynthesis and export mechanisms together with other principal physiological mechanisms challenging. Future technologies that provide the genetic tools necessary to screen C. burnetii mutants and propagate isogenic mutants might speed the discovery process in this area.
References
Alexander C, Rietschel ET (2001) Bacterial lipopolysaccharides and innate immunity. J Endotoxin Res 7:167–202
Al-Hendy A, Toivanen P, Skurnik M (1991) Expression cloning of Yersinia enterocolitica O:3 rfb gene cluster in Escherichia coli K12. Microb Pathog 10:47–59
Amano K, Williams JC (1984) Chemical and immunological characterization of lipopolysaccharides from phase I and phase II Coxiella burnetii. J Bacteriol 160:994–1002
Amano K, Williams JC, Missler SR et al (1987) Structure and biological relationships of Coxiella burnetii lipopolysaccharides. J Biol Chem 262:4740–4747
Aschauer H, Grob A, Hildebrandt J et al (1990) Highly purified lipid X is devoid of immunostimulatory activity. Isolation and characterization of immunostimulating contaminants in a batch of synthetic lipid X. J Biol Chem 265:9159–9164
Awomoyi AA, Rallabhandi P, Pollin TI et al (2007) Association of TLR4 polymorphisms with asymptomatic respiratory syncytial virus infection in high-risk infants and young children. J Immunol 179:3171–3177
Baca OG, Martinez IL, Aragon AS et al (1980) Isolation and partial characterization of a lipopolysaccharide from phase II Coxiella burnetii. Can J Microbiol 26:819–826
Bisercic M, Feutrier JY, Reeves PR (1991) Nucleotide sequences of the gnd genes from nine natural isolates of Escherichia coli: evidence of intragenic recombination as a contributing factor in the evolution of the polymorphic gnd locus. J Bacteriol 173:3894–3900
Bowen RA (2008) Protein hydrophobicity plots. http://www.vivo.colostate.edu/molkit/hydropathy/index.html
Bulut Y, Faure E, Thomas L et al (2002) Chlamydial heat shock protein 60 activates macrophages and endothelial cells through Toll-like receptor 4 and MD2 in a MyD88-dependent pathway. J Immunol 168:1435–1440
Burns SM, Hull SI (1998) Comparison of loss of serum resistance by defined lipopolysaccharide mutants and an acapsular mutant of uropathogenic Escherichia coli O75:K5. Infect Immun 66:4244–4253
Capo C, Zugun F, Stein A et al (1996) Upregulation of tumor necrosis factor-α and interleukin-1β in Q fever endocarditis. Infect Immun 64:1638–1642
Capo C, Amirayan N, Ghigo E et al (1999) Circulating cytokine balance and activation markers of leucocytes in Q fever. Clin Exp Immunol 115:120–123
Caroff M, Brisson JR, Martin A et al (2000) Structure of the Bordetella pertussis 1414 endotoxin. FEBS Lett 477:8–14
Chen MM, Glover KJ, Imperiali B (2007) From peptide to protein: comparative analysis of the substrate specificity of N-linked glycosylation in Campylobacter jejuni. Biochemistry 46:5579–5585
Clarke BR, Cuthbertson L, Whitfield C (2004) Nonreducing terminal modifications determine the chain length of polymannose O-antigens of Escherichia coli and couple chain termination to polymer export via an ATP-binding cassette transporter. J Biol Chem 279:35709–35718
Coleman SA, Fischer ER, Howe D et al (2004) Temporal analysis of Coxiella burnetii morphological differentiation. J Bacteriol 186:7344–7352
Coleman SA, Fischer ER, Cockrell DC et al (2007) Proteome and antigen profiling of Coxiella burnetii developmental forms. Infect Immun 75:290–298
Darveau RP, Pham TT, Lemley K et al (2004) Porphyromonas gingivalis lipopolysaccharide contains multiple lipid A species that functionally interact with both toll-like receptors 2 and 4. Infect Immun 72:5041–5051
Dellacasagrande J, Capo C, Raoult D et al (1999) IFN-γ-mediated control of Coxiella burnetii survival in monocytes: the role of cell apoptosis and TNF. J Immunol 162:2259–2265
Dellacasagrande J, Ghigo E, Capo C et al (2000a) Coxiella burnetii survives in monocytes from patients with Q fever endocarditis: involvement of tumor necrosis factor. Infect Immun 68:160–164
Dellacasagrande J, Ghigo E, Machergui-El Hammami S et al (2000b) Alpha(v)beta(3) integrin and bacterial lipopolysaccharide are involved in Coxiella burnetii-stimulated production of tumor necrosis factor by human monocytes. Infect Immun 68:5673–5678
Demerec M, Adelberg EA, Clark AJ et al (1966) A proposal for a uniform nomenclature in bacterial genetics. Genetics 54:61–76
Denison AM, Massung RF, Thompson HA (2007) Analysis of the O-antigen biosynthesis regions of phase II isolates of Coxiella burnetii. FEMS Microbiol Lett 267:102–107
Diaz QM, Lukacova M (1998) Immunological consequences of Coxiella burnetii phase variation. Acta Virol 42:181–185
Dotson SB, Rush JS, Ricketts AD et al (1995) Mannosylphosphoryldolichol-mediated O-mannosylation of yeast glycoproteins: stereospecificity and recognition of the alpha-isoprene unit by a purified mannosyltransferase. Arch Biochem Biophys 316:773–779
Doyle SL, O’ Neill LA (2006) Toll-like receptors from the discovery of NF-χB to new insights into transcriptional regulations in innate imunity. Biochem Pharmacol 72:1102–1113
D’Souza-Schorey C, McLachlan KR, Krag SS et al (1994) Mammalian glycosyltransferases prefer glycosyl phosphoryl dolichols rather than glycosyl phosphoryl polyprenols as substrates for oligosaccharyl synthesis. Arch Biochem Biophys 308:497–503
Emmerson JR, Gally DL, Roe AJ (2006) Generation of gene deletions and gene replacements in Escherichia coli O157:H7 using a temperature sensitive allelic exchange system. Biol Proced Online 8:153–162
Erridge C, Bennett-Guerrero E, Poxton IR (2002) Structure and function of lipopolysaccharides. Microbes Infect 4:837–851
Flebbe LM, Chapes SK, Morrison DC (1990) Activation of C3H/HeJ macrophage tumoricidal activity and cytokine release by R-chemotype lipopolysaccharide preparations. J Immunol 145:505–514
Ftacek P, Skultety L, Toman R (2000) Phase variation of Coxiella burnetii strain Priscilla: influence of this phenomenon on biochemical features of its lipopolysaccharide. J Endotoxin Res 6:369–376
Gajdosova E, Kovacova E, Toman R et al (1994) Immunogenicity of Coxiella burnetii whole cells and their outer membrane components. Acta Virol 38:339–344
Girard R, Pedron T, Uematsu S et al (2003) Lipopolysaccharides from Legionella and Rhizobium stimulate mouse bone marrow granulocytes via toll-like receptor 2. J Cell Sci 116:293–302
Golenbock DT, Hampton RY, Qureshi N et al (1991) Lipid A-like molecules that antagonize the effects of endotoxins on human monocytes. J Biol Chem 266:19490–19498
Gunn JS, Ernst RK (2007) The structure and function of Francisella lipopolysaccharide. Ann NY Acad Sci 1105:202–218
Hackstadt T (1986) Antigenic variation in the phase I lipopolysaccharide of Coxiella burnetii isolates. Infect Immun 52:337–340
Hackstadt T, Peacock MG, Hitchcock PJ et al (1985) Lipopolysaccharide variation in Coxiella burnetii: intrastrain heterogeneity in structure and antigenicity. Infect Immun 48:359–365
Hajjar AM, O’Mahony DS, Ozinsky A et al (2001) Cutting edge: functional interactions between toll-like receptor (TLR) 2 and TLR1 or TLR6 in response to phenol-soluble modulin. J Immunol 166:15–19
Heinrichs DE, Whitfield C, Valvano MA (1999) Biosynthesis and genetics of lipopolysaccharide core. In: Brade H, Opal SM, Vogel SN, Morrison DC (eds) Endotoxin in health and disease. Marcel Dekker, New York/Basel, pp 305–330
Heinzen RA, Hackstadt T (1996) A developmental stage-specific histone H1 homolog of Coxiella burnetii. J Bacteriol 178:5049–5052
Helbig JH, Luck PC, Knirel YA et al (1995) Molecular characterization of a virulence-associated epitope on the lipopolysaccharide of Legionella pneumophila serogroup 1. Epidemiol Infect 115:71–78
Hirschfeld M, Weiss JJ, Toshchakov V et al (2001) Signaling by toll-like receptor 2 and 4 agonists results in differential gene expression in murine macrophages. Infect Immun 69:1477–1482
Hobbs M, Reeves PR (1994) The JUMPstart sequence: a 39 bp element common to several polysaccharide gene clusters. Mol Microbiol 12:855–856
Hollenstein K, Dawson RJ, Locher KP (2007) Structure and mechanism of ABC transporter proteins. Curr Opin Struct Biol 17:412–418
Holst O (1999) Chemical structure of the core region of lipopolysaccharides. In: Brade H, Opal SM, Vogel SN, Morrison DC (eds) Endotoxin in health and disease. Marcel Dekker, New York/Basel, pp 115–154
Honstettre A, Ghigo E, Moynnault A et al (2004) Lipopolysaccharide from Coxiella burnetii is involved in bacterial phagocytosis, filamentous actin reorganization, and inflammatory responses through toll-like receptor 4. J Immunol 172:3695–3703
Hoover TA, Culp DW, Vodkin MH et al (2002) Chromosomal DNA deletions explain phenotypic characteristics of two antigenic variants, phase II and RSA 514 (Crazy), of the Coxiella burnetii nine mile strain. Infect Immun 70:6726–6733
Hussein A, Kovacova E, Toman R (2001) Isolation and evaluation of Coxiella burnetii O-polysaccharide antigen as immunodiagnostic reagent. Acta Virol 45:173–180
Jimenez de Bagues MP, Gross A, Terraza A et al (2005) Cellular bioterrorism: how Brucella corrupts macrophage physiology to promote invasion and proliferation. Clin Immunol 114:227–238
Joiner KA (1988) Complement evasion by bacteria and parasites. Ann Rev Microbiol 42:201–230
Kawai T, Akira S (2008) Toll-like receptor and RIG-I-like receptor signaling. Ann NY Acad Sci 1143:1–20
Kean EL, Rush JS, Waechter CJ (1994) Activation of GlcNAc-P-P-dolichol synthesis by mannosylphosphoryldolichol is stereospecific and requires a saturated alpha-isoprene unit. Biochemistry 33:10508–10512
Keenleyside WJ, Whitfield C (1999) Genetics and biosynthesis of lipopolysaccharide O-antigens. In: Brade H, Opal SM, Vogel SN, Morrison DC (eds) Endotoxin in health and disease. Marcel Dekker, New York/Basel, pp 331–358
King JD, Kocincova D, Westman EL et al (2009) Review: lipopolysaccharide biosynthesis in Pseudomonas aeruginosa. Innate Immun 15:261–312
Knirel YA, Rietschel ET, Marre R et al (1994) The structure of the O-specific chain of Legionella pneumophila serogroup 1 lipopolysaccharide. Eur J Biochem 221:239–245
Kubes M, Kuzmova Z, Gajdosova E et al (2006) Induction of tumor necrosis factor alpha in murine macrophages with various strains of Coxiella burnetii and their lipopolysaccharides. Acta Virol 50:93–99
Lien E, Sellati TJ, Yoshimura A et al (1999) Toll-like receptor 2 functions as a pattern recognition receptor for diverse bacterial products. J Biol Chem 274:33419–33425
Locksley RM, Killeen N, Lenardo MJ (2001) The TNF and TNF receptor superfamilies: integrating mammalian biology. Cell 104:487–501
Lorenz E, Mira JP, Cornish KL et al (2000) A novel polymorphism in the toll-like receptor 2 gene and its potential association with staphylococcal infection. Infect Immun 68:6398–6401
Lu YC, Yeh WC, Ohashi PS (2008) LPS/TLR4 signal transduction pathway. Cytokine 42:145–151
Luck PC, Freier T, Steudel C et al (2001) Point mutation in the active site of Legionella pneumophila O-acetyltransferase results in modified lipopolysaccharide but does not influence virulence. Int J Med Microbiol 291:345–352
Luneberg E, Zahringer U, Knirel YA et al (1998) Phase-variable expression of lipopolysaccharide contributes to the virulence of Legionella pneumophila. J Exp Med 188:49–60
Luneberg E, Zetzmann N, Alber D et al (2000) Cloning and functional characterization of a 30 kb gene locus required for lipopolysaccharide biosynthesis in Legionella pneumophila. Int J Med Microbiol 290:37–49
Luneberg E, Mayer B, Daryab N et al (2001) Chromosomal insertion and excision of a 30 kb unstable genetic element is responsible for phase variation of lipopolysaccharide and other virulence determinants in Legionella pneumophila. Mol Microbiol 39:1259–1271
Malley R, Hennecke P, Morse SC et al (2003) Recognition of pneumolysin by toll-like receptor 4 confers resistance to pneumococcal infection. Proc Natl Acad Sci USA 100:1966–1971
Marrie TJ, Raoult D (1997) Q fever – a review and issues for the next century. J Antimicrob Agents 8:145–161
Mayer H, Radziejewska-Lebrecht J, Schramek S (1988) Chemical and immunochemical studies on lipopolysaccharides of Coxiella burnetii phase I and phase II. Adv Exp Med Biol 228:577–591
McCaul TF, Banerjee-Bhatnagar N, Williams JC (1991) Antigenic differences between Coxiella burnetii cells revealed by postembedding immunoelectron microscopy and immunoblotting. Infect Immun 59:3243–3253
McLachlan KR, Krag SS (1994) Three enzymes involved in oligosaccharide-lipid assembly in Chinese hamster ovary cells differ in lipid substrate preference. J Lipid Res 35:1861–1868
Meghari S, Honstettre A, Lepidi H et al (2005) TLR2 is necessary to inflammatory response in Coxiella burnetii infection. Ann NY Acad Sci 1063:161–166
Moran AP, Zähringer U, Seydel U et al (1991) Structural analysis of the lipid A component of Campylobacter jejuni CCUG 10936 (serotype O:2) lipopolysaccharide. Description of a lipid A containing a hybrid backbone of 2-amino-2-deoxy-D-glucose and 2,3-diamino-2,3-dideoxy-D-glucose. Eur J Biochem 198:459–469
Nagy G, Palkovics T, Otto A et al (2008) “Gently rough”: the vaccine potential of a Salmonella enterica regulatory lipopolysaccharide mutant. J Infect Dis 198:1699–1706
Narasaki CT, Mertens K, Samuel JE (2011) Characterization of the GDP-D-mannose biosynthesis pathway in Coxiella burnetii: the initial steps for GDP-β-D-virenose biosynthesis. PLoS One 6(10):e25514
Nelson K, Selander RK (1994) Intergeneric transfer and recombination of the 6-phosphogluconate dehydrogenase gene (gnd) in enteric bacteria. Proc Natl Acad Sci USA 91:10227–10231
Nesper J, Lauriano CM, Klose KE et al (2001) Characterization of Vibrio cholerae O1 El tor galU and galE mutants: influence on lipopolysaccharide structure, colonization, and biofilm formation. Infect Immun 69:435–445
O’Neill LA (2006) How toll-like receptors signal: what we know and what we don’t know. Curr Opin Immunol 18:3–9
Palkovicova K, Ihnatko R, Vadovic P et al (2009) A monoclonal antibody specific for a unique biomarker virenose in a lipopolysaccharide of Coxiella burnetii. Clin Microbiol Infect 15:183–184
Pretat L, Toman R, Vadovic P et al (2009) Intracellular trafficking of the Coxiella burnetii lipopolysaccharide. Clin Microbiol Infect 15:185–187
Raetz CR, Whitfield C (2002) Lipopolysaccharide endotoxins. Ann Rev Biochem 71:635–700
Reeves PR, Hobbs M, Valvano MA et al (1996) Bacterial polysaccharide synthesis and gene nomenclature. Trends Microbiol 4:495–503
Rietschel ET, Brade H, Brade L et al (1987) Lipid A, the endotoxic centre of bacterial lipopolysaccharides: relation of chemical structure to biological activity. Prog Clin Biol Res 231:25–53
Rietschel ET, Kirikae T, Schade U et al (1993) The chemical structure of bacterial endotoxin in relation to bioactivity. Immunobiology 187:169–190
Rocchetta HL, Burrows LL, Lam JS (1999) Genetics of O-antigen biosynthesis in Pseudomonas aeruginosa. Microbiol Mol Biol Rev 63:523–553
Rojas G, Saldias S, Bittner M et al (2001) The rfaH gene, which affects lipopolysaccharide synthesis in Salmonella enterica serovar Typhi, is differentially expressed during the bacterial growth phase. FEMS Microbiol Lett 204:123–128
Rush JS, Shelling JG, Zingg NS et al (1993) Mannosylphosphoryldolichol-mediated reactions in oligosaccharide-P-P-dolichol biosynthesis. Recognition of the saturated alpha-isoprene unit of the mannosyl donor by pig brain mannosyltransferases. J Biol Chem 268:13110–13117
Rush JS, Rick PD, Waechter CJ (1997) Polyisoprenyl phosphate specificity of UDP-GlcNAc: undecaprenyl phosphate N-acetylglucosaminyl 1-P transferase from E.coli. Glycobiology 7:315–322
Rush JS, Alaimo C, Robbiani R et al (2010) A novel epimerase that converts GlcNAc-P-P-undecaprenol to GalNAc-P-P-undecaprenol in Escherichia coli O157. J Biol Chem 285:1671–1680
Samuel G, Reeves P (2003) Biosynthesis of O-antigens: genes and pathways involved in nucleotide sugar precursor synthesis and O-antigen assembly. Carbohydr Res 338:2503–2519
Sandlin RC, Lampel KA, Keasler SP et al (1995) Avirulence of rough mutants of Shigella flexneri: requirement of O antigen for correct unipolar localization of IcsA in the bacterial outer membrane. Infect Immun 63:229–237
Sandlin RC, Goldberg MB, Maurelli AT (1996) Effect of O side-chain length and composition on the virulence of Shigella flexneri 2a. Mol Microbiol 22:63–73
Schramek S, Mayer H (1982) Different sugar composition of lipopolysaccharides isolated from phase I and pure phase II cells of Coxiella burnetii. Infect Immun 38:53–57
Schramek S, Radziejewska-Lebrecht J, Mayer H (1985) 3-C-branched aldoses in lipopolysaccharide of phase I Coxiella burnetii and their role as immunodominant factors. Eur J Biochem 148:445–461
Seshadri R, Hendrix LR, Samuel JE (1999) Differential expression of translational elements by life cycle variants of Coxiella burnetii. Infect Immun 67:6026–6033
Seshadri R, Paulsen IT, Eisen JA et al (2003) Complete genome sequence of the Q-fever pathogen Coxiella burnetii. Proc Natl Acad Sci USA 100:5455–5460
Seydel U, Wiese A, Schromm AB et al (1999) A biophysical view on the function and activity of endotoxins. In: Brade H, Opal SM, Vogel SN, Morrison DC (eds) Endotoxin in health and disease. Marcel Dekker, New York/Basel, pp 195–219
Shannon JG, Howe D, Heinzen RA (2005) Virulent Coxiella burnetii does not activate human dendritic cells: role of lipopolysaccharide as a shielding molecule. Proc Natl Acad Sci USA 102:8722–8727
Shimazu R, Akashi S, Ogata H et al (1999) MD-2, a molecule that confers lipopolysaccharide responsiveness on toll-like receptor 4. J Exp Med 189:1777–1782
Skultety L, Toman R, Patoprsty V (1998) A comparative study of lipopolysaccharides from two Coxiella burnetii strains considered to be associated with acute and chronic Q fever. Carbohydr Polymers 35:189–194
Skultety L, Hernychova L, Toman R et al (2005) Coxiella burnetii whole cell lysate protein identification by mass spectrometry and tandem mass spectrometry. Ann NY Acad Sci 1063:115–122
Skurnik M, Toivanen P (1993) Yersinia enterocolitica lipopolysaccharide: genetics and virulence. Trends Microbiol 1:148–152
Slaba K, Hussein A, Palkovic P et al (2003) Studies on immunological role of virenose and dihydrohydroxystreptose present in the Coxiella burnetii phase I lipopolysaccharide. Ann NY Acad Sci 990:505–509
Stoker MG, Fiset P (1956) Phase variation of the Nine Mile and other strains of Rickettsia burneti. Can J Microbiol 2:310–321
Stroeher UH, Jedani KE, Manning PA (1998) Genetic organization of the regions associated with surface polysaccharide synthesis in Vibrio cholerae O1, O139 and Vibrio anguillarum O1 and O2: a review. Gene 223:269–282
Svraka S, Toman R, Skultety L et al (2006) Establishment of a genotyping scheme for Coxiella burnetii. FEMS Microbiol Lett 254:268–274
Szkopinska A, Swiezewska E, Chojnacki T (1992) On the specificity of dolichol kinase and DolPMan synthase towards isoprenoid alcohols of different chain length in rat liver micro-somal membrane. Int J Biochem 24:1151–1157
Takeda K, Akira S (2005) Toll-like receptors in innate immunity. Int Immunol 17:1–14
Thompson HA, Hoover TA, Vodkin MH et al (2003) Do chromosomal deletions in the lipopolysaccharide biosynthetic regions explain all cases of phase variation in Coxiella burnetii strains? An update. Ann NY Acad Sci 990:664–670
Toman R (1991) Basic structural features of a lipopolysaccharide from the Coxiella burnetii strain Nine Mile in the virulent phase I. Acta Virol 35:224
Toman R, Kazar J (1991) Evidence for the structural heterogeneity of the polysaccharide component of Coxiella burnetii strain Nine Mile lipopolysaccharide. Acta Virol 35:531–537
Toman R, Skultety L (1994) Analysis of the 3-deoxy-D-manno-2-octulosonic acid region in a lipopolysaccharide isolated from Coxiella burnetii strain Nine Mile in phase II. Acta Virol 38:241–243
Toman R, Skultety L (1996) Structural study on a lipopolysaccharide from Coxiella burnetii strain Nine Mile in avirulent phase II. Carbohydr Res 283:175–185
Toman R, Skultety L, Kazar J (1993) On the determination of “Kdo-like substance” in the lipopolysaccharide from Coxiella burnetii strain Nine Mile in phase II. Acta Virol 37:196–198
Toman R, Skultety L, Ftacek P et al (1998) NMR study of virenose and dihydrohydroxystreptose isolated from Coxiella burnetii phase I lipopolysaccharide. Carbohydr Res 306:291–296
Toman R, Hussein A, Palkovic P et al (2003a) Structural properties of lipopolysaccharides from Coxiella burnetii strains Henzerling and S. Ann NY Acad Sci 990:563–567
Toman R, Hussein A, Slaba K et al (2003b) Further structural characteristics of the lipopolysaccharide from Coxiella burnetii strain Nine Mile in low virulent phase II. Acta Virol 47:129–130
Toman R, Garidel P, Andra J et al (2004) Physicochemical characterization of the endotoxins from Coxiella burnetii strain Priscilla in relation to their bioactivities. BMC Biochem 5:1 http://www.biomedcentral.com/1471-2091/5/1
Toman R, Skultety L, Ihnatko R (2009) Coxiella burnetii glycomics and proteomics – tools for linking structure to function. Ann NY Acad Sci 1166:67–78
Trefzer A, Salas JA, Bechthold A (1999) Genes and enzymes involved in deoxysugar biosynthesis in bacteria. Nat Prod Rep 16:283–299
Tujulin E, Lilliehook B, Macellaro A et al (1999) Early cytokine induction in mouse P388D1 macrophages infected by Coxiella burnetii. Vet Immunol Immunopathol 68:159–168
Ulmer AJ, Heine H, Feist W et al (1992) Biological activity of synthetic phosphono-oxyethyl analogues of lipid A and lipid A partial structures. Infect Immun 60:3309–3314
Vadovic P, Slaba K, Fodorova M et al (2005) Structural and functional characterization of the glycan antigens involved in immunobiology of Q fever. Ann NY Acad Sci 1063:149–153
Vadovic P, Fodorova M, Toman R (2007) Structural studies of lipid A of Piscirickettisa salmonis, the etiological agent of the salmonid rickettsial septicemia. Acta Virol 51:249–259
Vadovic P, Fuleova A, Ihnatko R et al (2009) Structural studies of lipid A from a lipopolysaccharide of the Coxiella burnetii isolate RSA 514 (Crazy). Clin Microbiol Infect 15:198–199
Van den Bosch L, Manning PA, Morona R (1997) Regulation of O-antigen chain length is required for Shigella flexneri virulence. Mol Microbiol 23:765–775
Van der Woude MW, Baumler AJ (2004) Phase and antigenic variation in bacteria. Clin Microbiol Rev 17:581–611
Vishwanath S, Hackstadt T (1988) Lipopolysaccharide phase variation determines the complement-mediated serum susceptibility of Coxiella burnetii. Infect Immun 56:40–44
Vodkin MH, Williams JC (1986) Overlapping deletion in two spontaneous phase variants of Coxiella burnetii. J Gen Microbiol 132:2587–2594
Wang L, Jensen S, Hallman R et al (1998) Expression of the O antigen gene cluster is regulated by RfaH through the JUMPstart sequence. FEMS Microbiol Lett 165:201–206
Weiser JN, Love JM, Moxon ER (1989) The molecular mechanism of phase variation of Haemophilus influenzae lipopolysaccharide. Cell 59:657–665
Weisman LS, Ballou CE (1984) Biosynthesis of the mycobacterial methylmannose polysaccharide. Identification of a 3-O-methyltransferase. J Biol Chem 259:3464–3469
Weiss J, Hutzler M, Kao L (1986) Environmental modulation of lipopolysaccharide chain length alters the sensitivity of Escherichia coli to the neutrophil bactericidal/permeability-increasing protein. Infect Immun 51:594–599
Williams JC, Waag DM (1991) Antigens, virulence factors, and biological response modifiers of Coxiella burnetii: strategies for vaccine development. In: Williams JC, Thompson HA (eds) Q fever: the biology of Coxiella burnetii. CRC Press, Boca Raton, pp 175–222
Wollenweber HW, Schramek S, Moll H et al (1985) Nature and linkage of fatty acids present in lipopolysaccharides of phase I and phase II Coxiella burnetii. Arch Microbiol 142:6–11
Zamboni DS, Campos MA, Torrecilhas ACT et al (2004) Stimulation of toll-like receptor 2 by Coxiella burnetii is required for macrophage production of pro-inflammatory cytokines and resistance to infection. J Biol Chem 279:54405–54415
Zhang G, Russell-Lodrigue KE, Andoh M et al (2007) Mechanisms of vaccine-induced protective immunity against Coxiella burnetii infection in BALB/c mice. J Immunol 179:8372–8380
Acknowledgement
This research was supported in part by the grant 2/0026/12 from the Scientific Grant Agency of Ministry of Education of Slovak Republic and the Slovak Academy of Sciences.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Narasaki, C.T., Toman, R. (2012). Lipopolysaccharide of Coxiella burnetii . In: Toman, R., Heinzen, R., Samuel, J., Mege, JL. (eds) Coxiella burnetii: Recent Advances and New Perspectives in Research of the Q Fever Bacterium. Advances in Experimental Medicine and Biology, vol 984. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-4315-1_4
Download citation
DOI: https://doi.org/10.1007/978-94-007-4315-1_4
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-4314-4
Online ISBN: 978-94-007-4315-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)