
Abstract
Early metabolic studies of C. burnetii showed minimal metabolic activity of axenic (host cell-free) organisms in buffers adjusted to neutral pH. However, our understanding of the organism’s physiology was greatly improved upon the discovery that C. burnetii requires an acidic pH for metabolic activation. Indeed, information gained from acid activation studies coupled with contemporary analyses using transcription microarrays, metabolic pathway reconstruction and metabolite typing, led to an axenic culture system that supports robust growth of C. burnetii. While axenic culture of C. burnetii can present some technical challenges, the technique is currently facilitating new lines of investigation and development of genetic tools. Axenic cultivation of C. burnetii should also prove useful in clinical settings.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
11.1 Introduction
The discovery of C. burnetii as the causative agent of human Q fever in the late 1930s was followed almost immediately by unsuccessful attempts to cultivate the pathogen in axenic (host cell-free) medium (Cox 1938). Microscopic evaluation established an intracellular vacuolar niche for the organism and contributed to C. burnetii’s designation as an obligate intracellular bacterium. Subsequent analyses of the organism’s metabolic abilities in buffers adjusted to physiological pH showed substantial activity of central metabolic pathways (e.g., glycolysis and RNA synthesis) in bacterial extracts, but not intact cells (Baca and Paretsky 1983). The demonstration that the C. burnetii parasitophorous vacuole (PV) exhibits phagolysosomal characteristics (Burton et al. 1971, 1978) led to the discovery that a moderately acidic pH mimicking that of a phagolysosome (∼pH 4.5) is required for metabolic activation of C. burnetii (Hackstadt and Williams 1981a). Indeed, C. burnetii transport, catabolism and incorporation of both glucose and glutamate are dependent on acidic pH (Hackstadt 1983). Moreover, pH dependent transport of carbon and energy sources is required for de novo protein synthesis (Zuerner and Thompson 1983), stability of the ATP pool (Hackstadt and Williams 1981b), and membrane energization (Hackstadt 1983). Acid activation of C. burnetii metabolism was presented as a “biochemical stratagem” (Hackstadt and Williams 1981a) manifested as metabolic quiescence and stability at the neutral pH found in the extracellular environment, and metabolic activation and bacterial replication at the acidic pH found in the organism’s PV (Akporiaye et al. 1983). With improved knowledge of C. burnetii metabolic requirements, researchers revisited the possibility of C. burnetii axenic cultivation using more complex buffer systems. While de novo nucleic acid and protein biosynthesis were observed for several hours, net biomass increases indicated C. burnetii was not replicating (Chen et al. 1990).
11.2 Genomic Insight into C. burnetii Metabolic Capacity
The sequencing of multiple C. burnetii strains allowed a comprehensive look at C. burnetii ’s metabolic potential (Beare et al. 2009b; Seshadri et al. 2003). Overall, the ∼2 Mb genome of C. burnetii is roughly twice the size of most genomes of obligate intracellular rickettsiae (Ellison et al. 2008; McLeod et al. 2004) and chlamydiae (Stephens et al. 1998). Moreover, analysis of coding ORFs shows that C. burnetii central metabolic pathways, such as glycolysis, the tricarboxylic acid cycle (TCA), and pathways for nucleic acid biosynthesis, are largely intact. As discussed below, C. burnetii does have predicted metabolic peculiarities that may reflect adaptation to intracellular parasitism, including some that may impact overall metabolism.
11.2.1 Glycolysis, TCA, and Pentose Phosphate Pathways
Biochemical studies show that C. burnetii can catabolize glucose (Hackstadt and Williams 1981a, b). However, the organism does not encode the initial enzyme of glycolysis (glucokinase or hexokinase) needed to convert glucose to glucose-6-phosphate. Because C. burnetii lysates can phosphorylate glucose in the presence of carbamoyl phosphate (Paretsky et al. 1962), glucose may be phosphorylated by an alternative carbamoyl phosphate-dependent mechanism that subsequently allows oxidation of glucose via glycolysis. C. burnetii encodes a complete TCA cycle and readily metabolizes glutamate, succinate, and pyruvate (Hackstadt and Williams 1981b). However, the organism lacks the glyoxylate bypass enzymes isocitrate lyase and malate synthase, and consequently cannot utilize fatty acids as a sole source of carbon. This is in contrast to the intracellular bacterium Mycobacterium tuberculosis that relies on fatty acids as a source of carbon during intracellular growth (Schnappinger et al. 2003). The non-oxidative branch of the pentose phosphate pathway is complete in C. burnetii, allowing generation of pentose sugars for nucleic acid biosynthsis. However, the oxidative branch of the pathway is defective due to the lack of glucose-6-phosphate dehydrogenase and 6-phosphogluconate dehydrogenase. Thus, C. burnetii cannot rely on this pathway for synthesis of NADPH, a major source of reducing equivalents for biosynthetic reactions and antioxidant mechanisms. Overall, biochemical and genomic data suggest oxidation of glycolytic and TCA cycle intermediates, and substrates that funnel directly into these pathways, as critical for C. burnetii energy production.
11.2.2 Amino Acid Acquisition
Based on reconstruction of amino acid biosynthetic pathways, C. burnetii appears auxotrophic for 11 amino acids (Beare et al. 2009b; Seshadri et al. 2003). Thus, during intracellular growth, C. burnetii must acquire these amino acids via transport of free amino acids or peptides from the PV lumen. Autophagosomes, carrying defunct organelles and protein aggregates, and fluid phase endosomes, fuse with the C. burnetii PV (Beron et al. 2002; Heinzen et al. 1996). This trafficking likely provides key protein precursors required for C. burnetii growth. Indeed, the C. burnetii PV lumen is clearly a proteolytic environment as evidenced by the rapid breakdown of bovine serum albumin and the presence of active cathepsins (Howe et al. 2010). This activity is predicted to convert proteins to short peptides and free amino acids that can be transported by C. burnetii using several amino acid permeases, di-, tri- and oligo-peptide transporters, and 13 major facilitator superfamily transporters (Beare et al. 2009b). C. burnetii’s numerous amino acid auxotrophies suggest the PV has a steady supply of protein precursors, at least during the organism’s replicative phase.
11.2.3 Energy Production
The mechanism(s) of C. burnetii acid activation is poorly defined. Data from early acid activation studies suggest extracellular acidic pH has multiple beneficial effects on C. burnetii physiology including membrane energization (Hackstadt 1983) and activation of metabolite transport (Hackstadt and Williams 1983). With respect to membrane energization, a proton gradient imposed by incubation of host cell-free organisms at acidic pH (i.e., pH 3–5) results in a transient increase of the C. burnetii ATP pool (Hackstadt and Williams 1981b). This effect is consistent with proton gradient driven synthesis of ATP via the organism’s F1F0 ATP synthase. However, maintenance of the ATP pool is dependent on an oxidizable substrate, such as glutamate. Acidic pH clearly assists C. burnetii metabolite transport as demonstrated by enhanced transport of glutamate at pH 4.5 (Hackstadt and Williams 1983). Thus, a working model of C. burnetii acid activation of metabolism involves stimulation of proton symporters that acquire critical carbon and energy sources.
C. burnetii pH-dependent transport of energy sources and their subsequent oxidation generates reducing equivalents (e.g., NADH) that drives respiratory chain extrusion of protons. This process results in a proton motive force that promotes ATP synthesis. C. burnetii encodes a branched respiratory chain with distinct terminal ubiquinol cytochrome o (encoded by cyoABCDE) or cytochrome bd (encoded by cydAB) oxidases (Fig. 11.1). Cytochrome bd, with its increased affinity for oxygen, is used by bacteria under microaerobic conditions. Thus, C. burnetii may adapt to microaerobic levels of oxygen during intracellular growth. In fact, the intracellular oxygen tension of cultured eukaryotic cells and their compartments can be much lower than the extracellular oxygen tension (Hu et al. 1992). Furthermore, the level of oxygen in the spleen, a tissue that can contain C. burnetii infected macrophages, has been measured at approximately 3% (Braun et al. 2001).

Schematic overview of the C. burnetii aerobic respiratory chain. C. burnetii’s branched respiratory chain has distinct terminal ubiquinol oxidases incorporating either cytochrome o or cytochrome bd that are typically used under aerobic and microaerobic conditions, respectively. It is unclear how C. burnetii acquires ubiquinone, the predicted major quinone cofactor in electron transport activities, as key biosynthetic enzymes are missing from the pathogen’s genome
11.2.4 pH Homeostasis
C. burnetii cytoplasmic pH drops when the organism is incubated in an acidic buffer without an oxidizable energy source. Associated with this pH drop is depletion of the ATP pool and loss of viability (Hackstadt and Williams 1981b). Thus, like other more extreme acidophiles, maintenance of cytoplasmic neutrality by C. burnetii is key for survival and the organism likely deploys several mechanisms to maintain pH homeostasis. As described above, proton extrusion via the electron transport chain could contribute to the regulation of cytoplasmic pH. Moreover, energy requiring expulsion of protons could occur by the F0F1-ATPase reverse reaction, and by K+/H+ and Na+/H+ antiporters. Proton consumption via decarboxylation of amino acids can also be utilized as a mechanism to prevent cytoplasmic acidification (Kashiwagi et al. 1992). Ammonia generation during conversion of glutamine to glutamate (Harth et al. 1994) is another example of how C. burnetii might couple amino acid metabolism to cytoplasmic pH homeostasis.
11.3 Establishment of a C. burnetii Axenic Culture System
Early work establishing conditions that support transient acid activation of C. burnetii metabolism provided a foundation on which to further define nutritional and physiochemical conditions that support prolonged axenic metabolic activity. Using contemporary tools, such as biochemical pathway reconstruction, transcriptomics, and metabolite typing, conditions were defined that not only support prolonged C. burnetii metabolic activity, but also axenic replication.
11.3.1 First Generation Medium Supporting Sustained Metabolic Activity
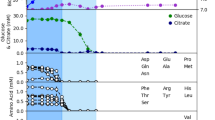
The first stage in designing an axenic culture system for C. burnetii was to establish a medium that supported metabolic activity, in the form of protein and ATP synthesis, for several hours (Omsland et al. 2008). The known requirement of a moderately acidic environment (∼pH 4.5) for C. burnetii metabolic activation (Hackstadt and Williams 1981a) prompted screening of several buffers to identify one with both good buffering capacity at acidic pH and the ability to support C. burnetii metabolic activity. Buffers based on acetate and succinate were deleterious to C. burnetii de novo protein synthesis. However, a citrate buffer supported high metabolic activity and provided excellent buffering capacity.
Sodium (Na+), potassium (K+) and chloride (Cl−) make up the principal ion gradients used by microbes to transport nutrients and also help establish physiological osmotic pressure in living systems. The eukaryotic cell cytoplasm contains high concentrations of K+ (∼140 mM) relative to Na+ (∼15 mM) and Cl− (∼15 mM). Conversely, these levels are roughly reversed (K+, ∼5 mM; Na+, ∼145 mM; and Cl−, ∼110 mM) in interstitial fluid. Surprisingly, although the C. burnetii PV is located in the host cell cytoplasm, pathogen metabolic activity was enhanced in a citrate salts buffer (CSB) containing ion levels resembling interstitial fluid (Omsland et al. 2008). This metabolic effect may reflect PV interactions with fluid phase endosomes that traffic material from the extracellular space to the PV lumen (Heinzen et al. 1996).
CSB was then supplemented with three complex nutrient sources to compensate for C. burnetii’s auxotrophies and to exploit the organism’s otherwise sophisticated metabolic machinery: (1) neopeptone, a combination of peptides, nucleotides and vitamins, (2) fetal bovine serum (FBS), and (3) RPMI cell culture medium. Supplementation of CSB with individual nutrient sources only marginally improved C. burnetii metabolic activity. However, a considerable increase in metabolic activity was observed when the nutrient sources were combined, creating a first generation medium called Complex Coxiella Medium (CCM) (Omsland et al. 2008). C. burnetii metabolic activity was optimal within a narrow range of nutrient concentrations. In fact, addition of extra nutrients, such as succinate or pyruvate, actually caused a decline in C. burnetii ATP pools and de novo protein synthesis. Incomplete oxidation of excess substrate resulting in generation of inhibitory intermediates may explain C. burnetii sensitivity to certain carbon sources (Teixeira de Mattos and Neijssel 1997). The negative effects of succinate and pyruvate on C. burnetii metabolism may also reflect suboptimal biochemical routing of the oxidized substrate (Tesh et al. 1983).
11.3.2 Second Generation Medium Supporting Axenic Replication
Although CCM supported prolonged C. burnetii metabolic activity, increases in genome equivalents (GE) indicating cell division were not observed. Therefore, transcription microarray technology was used to assess the overall metabolic state of C. burnetii in CCM with the goal of defining potential medium nutritional deficiencies (Omsland et al. 2009). When compared to the transcriptome of C. burnetii replicating in Vero host cells, ribosomal gene expression of C. burnetii in CCM was dramatically downregulated, suggesting suboptimal protein synthesis could be blocking replication. Thus, an additional source of amino acids (casamino acids) was added to CCM in an attempt to boost pathogen de novo protein synthesis (Omsland et al. 2009). A high concentration of L-cysteine was also added as this supplement is necessary for axenic growth of Legionella pneumophila (Ewann and Hoffman 2006), a close phylogenetic relative of C. burnetii. The new medium called Acidified Citrate Cysteine Medium (ACCM), supported a considerable increase in C. burnetii de novo protein synthesis. However, cell division was still not observed.
As discussed, the presence of a branched respiratory chain in C. burnetii suggests the organism might adjust to alterations in oxygen concentration during intracellular growth. Previous testing of media had been conducted under ambient (∼20%) oxygen conditions. Therefore, the effect of oxygen availability on C. burnetii metabolic activity was probed using Phenotype Microarrays incubated in 20, 5 or 2.5% oxygen environments. The number of oxidized substrates increased as oxygen level decreased, suggesting C. burnetii is a microaerophile. Subsequent incubation of C. burnetii in ACCM in a 2.5% oxygen environment resulted in robust growth with approximately a 3 log10 increase in C. burnetii GE observed over a 6 day incubation (Omsland et al. 2009). Importantly, the infectivity of axenically propagated bacteria for Vero cells was similar to host cell-propagated bacteria. C. burnetii colony growth was also achieved using semi-solid ACCM-agarose plates.
11.4 Practical Aspects of C. burnetii Axenic Culture
Axenic cultivation of C. burnetii is enabling new lines of investigation that are difficult or impossible to conduct with bacteria propagated in host cells. However, axenic growth can be challenging and attention to detail is important for success. In this section, technical issues that are important to successful axenic culture of C. burnetii are discussed.
11.4.1 Preparation of Acidified Citrate Cysteine Medium (ACCM)
ACCM supports substantial growth of C. burnetii. However, the organism is highly sensitive to medium variations so care must be taken during medium preparation. For example, C. burnetii tolerates a moderate range of nutrient concentrations with elevated concentrations of certain nutrients, including some carbon sources, readily inhibiting C. burnetii metabolic activity and growth (Omsland et al. 2008). Moreover, ion conditions, especially the concentration of chloride, have considerable effects on metabolic activity (Omsland et al. 2008). The ability of C. burnetii to grow in ACCM is even affected by the water used to prepare the medium. Reproducible culture can be achieved using regular deionized water. The sensitivity of C. burnetii to ion and nutrient concentrations also means that a humidified incubator should be used for cultivation of C. burnetii in ACCM to prevent medium evaporation. Finally, C. burnetii acidophilic metabolism is highly sensitive to extracellular pH (Hackstadt and Williams 1981a). Therefore, care should be taken to adjust ACCM to pH 4.75 as moderate (e.g., 0.5 pH units) deviations from this value may compromise growth. The acidic pH of ACCM may contribute to medium instability as culture is inconsistent if the medium is more than 1 week old.
11.4.2 Modifications of ACCM
ACCM is a complex nutrient medium containing chemically undefined components, namely FBS, neopeptone, and casamino acids. We recently found that 1% FBS can be replaced with 1 mg/ml of synthetic methyl-β-cyclodextrin (Mβ-CD) (Omsland et al. 2011). Mβ-CD is used in other bacteriologic media where it may sequester toxic metabolites (Marchini et al. 1995; Imaizumi et al. 1983). The modified medium termed ACCM-2 supports improved growth of C. burnetii in liquid medium and as colonies in solid medium.
11.4.3 Growth of C. burnetii in ACCM
C. burnetii cultures can be established in ACCM with or without shaking. Gentle shaking (75 RPM) improves C. burnetii growth in ACCM while vigorous shaking can cause protein precipitation. Robust growth occurs in shaken 125 ml Erlenmeyer flasks containing 20 ml of medium (Fig. 11.2a) or 250 ml Erlenmeyer flasks containing 40 ml of medium. C. burnetii appears highly sensitive to several detergents (A. Omsland and R. Heinzen, unpublished observations). Thus, disposable polyethylene terepthalate glycol (PETG) Erlenmeyer flasks with 0.2 μm filtered screw top caps are preferred. Without shaking, culture vessels should provide a large air-liquid interface to promote gas exchange. T-25 and T-75 tissue culture flasks containing 7 and 20 ml of medium, respectively, work well in this manner. Although liquid cultures can be started using fewer than 10 organisms, an inoculum of 1.0 × 106 organisms/ml will develop to stationary phase (∼109 organisms/ml) in approximately 6 days (Omsland et al. 2009), similar to the organism’s growth kinetics in cultured cells (Coleman et al. 2004). A critical parameter that affects C. burnetii axenic growth is oxygen concentration. C. burnetii replicates in ACCM when oxygen levels are between 1% and 5%. Declines in growth are observed in 7.5% oxygen while no growth is observed in 10% oxygen. An incubator with automatic monitoring and gas adjustments ensures a reliable oxygen environment.

Axenic culture of C. burnetii. (a) Mock and C. burnetii Nine Mile (phase II)-inoculated ACCM-2 following 6 days of incubation with shaking in a 2.5% oxygen environment. (b) Representative colony formation by C. burnetii Nine Mile (phase II) in ACCM-agarose following 6 days of incubation in a 2.5% oxygen environment
11.4.4 Storage of ACCM Cultured C. burnetii
ACCM-cultivated C. burnetii are sensitive to repeated freeze-thaw cycles, but excellent viability is observed when stocks are stored in ACCM containing a cryoprotectant, such as 10% dimethyl sulfoxide. Nonetheless, for optimal viability, C. burnetii stocks should be stored as single-use aliquots.
11.4.5 C. burnetii Colony Growth in ACCM-Agarose
An important advance enabled by axenic growth is the ability to obtain C. burnetii clones by colony formation in ACCM-agarose. Minute colonies form when organisms are embedded in semi-solid medium using a soft agarose overlay method (Fig. 11.2b). C. burnetii will not grow directly on the surface of solid ACCM. This behavior may reflect a requirement by C. burnetii to be submersed in an acid bath for proper membrane energization and transporter function. The use of a thin overlay medium promotes outgrowth of C. burnetii colonies in one focal plane, thus facilitating identification and picking of isolated colonies.
To prepare ACCM-agarose plates, combine 10 ml 2× ACCM (pH 4.75, 37°C) with 10 ml 0.5% UltraPure Agarose preheated to 50°C, then pour the mixture into 100 mm Petri dish creating a 0.25% ACCM-agarose base. After the medium base has solidified, prepare the 0.1% agarose overlay medium by combining 1.25 ml 2× ACCM (37°C), 0.75 ml water (37°C), and 0.5 ml 0.5% UltraPure Agarose (50°C). Add the C. burnetii inoculum (maximum 0.1 ml) to the overlay medium, mix, then pour over the ACCM-agarose base. Incubate plates at 4°C for 30 min to promote solidification of the low percentage overlay medium. Remove excess liquid in ACCM plates by placing them in a biosafety cabinet, lids cracked, for 20–30 min or until condensation is gone.
Colonies develop in ACCM-agarose in 4–7 days when incubated in a 2.5% oxygen environment. Colonies may be as small as 0.05 mm in diameter (Omsland et al. 2009). The reason for small colony size is unclear, but electron microscopic analysis reveals that 6 day old colonies are primarily comprised of C. burnetii small cell variants (SCV) which are non-replicative, stationary phase cells (Coleman et al. 2004; Omsland et al. 2011). Nutrient depletion in the colony microenvironment may drive C. burnetii into stationary phase and halt further colony expansion.
11.4.6 Selection and Recovery of C. burnetii Genetic Transformants Using ACCM-Agarose
Axenic culture in both ACCM and ACCM-agarose greatly facilitates isolation of C. burnetii genetic transformants. Compared to cell culture-based methods, where clonal isolation of C. burnetii transformants can take 8–12 weeks (Beare et al. 2009a), transformants can be isolated in as little as 16 days using axenic techniques (Beare et al. 2010; Omsland et al. 2011). Picking of C. burnetii colonies is facilitated by first locating isolated colonies using a dissecting microscope. Colonies can then be extracted from solid ACCM-agarose with a micro-pipette tip with an enlarged opening (approximately 0.5 mm). The clonal populations of C. burnetii can then be expanded by culture in ACCM following release of embedded organisms by repeated pipetting.
11.4.7 Determination of C. burnetii Viability Using ACCM-Agarose
Enumeration of C. burnetii in stock preparations is commonly conducted using an infectious focus-forming unit (FFU) assay or by determining the number of GE by quantitative PCR (Q-PCR) (Coleman et al. 2004). The FFU assay relies on immunofluorescent detection of PVs in cultured host cells. This method is complicated by the fact that phase I and phase II C. burnetii have dramatically different infectivity for cultured host cells. For example, the infection efficiency of phase II C. burnetii for Vero cells is approximately 5 GE per FFU while several hundred phase I C. burnetii are required to produce a single FFU (Cockrell et al. 2008; R. A. Heinzen, unpublished data). While the GE assay is sensitive and accurate, it does not distinguish between live and dead bacteria. The plating efficiency of the C. burnetii Nine Mile phase II strain in ACCM-agarose is nearly 1 GE per colony (Beare et al. 2010). Thus, quantification of C. burnetii by a conventional colony forming unit assay using ACCM-agarose plates is an alternative method to accurately enumerate the number of live organisms in a given preparation.
11.5 Summary
Axenic culture has removed the severe experimental obstacles associated with C. burnetii’s obligate reliance on a eukaryotic host cell for growth. Perhaps most importantly, ACCM culture techniques are aiding development of new genetic tools for C. burnetii (Chen et al. 2010; Voth et al. 2011; Beare et al. 2010). Moreover, biochemical and physiological studies can now be conducted without the metabolic background of the host cell.
C. burnetii axenic growth requirements have in vivo correlates. Oxygen gradients that occur within biological systems, including across biological membranes, can be associated with oxygen concentrations that are considerably lower than atmospheric oxygen levels (i.e., ∼20%) (Hu et al. 1992; Khan et al. 2003). C. burnetii microaerophilic metabolism was hypothesized based on the organism’s branched respiratory chain containing terminal oxidases typically associated with bacterial responses to normoxic and microaerobic levels of oxygen. The physiological basis of enhanced C. burnetii metabolism in a microaerobic environment is unclear, but may include sensitivity to oxidative stress. Technical difficulties in measuring ion levels in acidic vacuoles have prevented direct measurement of PV Na+, K+, and Cl− concentrations. However, C. burnetii’s requirement for specific levels of these ions in ACCM likely correlates with their in vivo roles in physiological processes such as metabolite transport and pH homeostasis. Finally, a pH of ∼4.75 is firmly established for the C. burnetii PV (Akporiaye et al. 1983; Maurin et al. 1992), a value that correlates with optimal pathogen growth in ACCM.
Several advances in C. burnetii genetic transformation have been made using ACCM (Chen et al. 2010; Voth et al. 2011; Beare et al. 2010). For example, secretion into the host cell cytoplasm of several potential C. burnetii effector molecules was recently demonstrated with a shuttle vector developed using axenic procedures (Chen et al. 2010; Voth et al. 2011). Continued development of genetic methods should soon allow fulfillment of molecular Koch’s postulate for putative C. burnetii virulence genes. ACCM culture will also aid investigations of mechanisms used by C. burnetii to persist in the harsh conditions of its PV, and the biological bases of developmental form transitions. For instance, C. burnetii is highly sensitive to certain short chain fatty acids, such as propionate, during growth in ACCM (unpublished data). In L. pneumophila, nutritional cues including amino acid availability and exposure to short chain fatty acids can trigger a stringent response that drives transition of replicative to transmissive forms that are biologically reminiscent of C. burnetii large cell variant (LCV) and SCV developmental forms (Sauer et al. 2005; Edwards et al. 2009; Dalebroux et al. 2009). Thus, these molecules may also trigger stringent response genes that regulate C. burnetii morphological transitions, a hypothesis that is testable with ACCM culture techniques.
C. burnetii can be grown to high numbers in embryonated eggs (Williams et al. 1981). However, bacteria purified from eggs or mammalian host cells (Cockrell et al. 2008) contain eucaryotic host cell contamination, making these preparations suboptimal for biochemical analyses or vaccine production. Additionally, the homogenization and centrifugation steps involved in purification can generate infectious aerosols, making the procedure potentially hazardous. Purification of C. burnetii from ACCM is simple, safe and results in organisms free of host cell contamination.
ACCM cultures of the avirulent C. burnetii Nine Mile phase II isolate can be started from tissue samples of infected SCID mice that contain as few as 10 organisms (Omsland et al. 2011). Virulent C. burnetii isolates also grow well in ACCM; thus, it should be possible to isolate these organisms from infected immunocompetent animals and human patients using ACCM culture techniques. Recent Q fever outbreaks (Gilsdorf et al. 2008; Schimmer et al. 2009) highlight the need for improved clinical and epidemiological methods. Axenic isolation of C. burnetii should prove valuable in these settings.
11.6 Disclosure Statement
A Patent Cooperation Treaty patent application (No. PCT/US2010/024706) has been filed on ACCM.
References
Akporiaye ET, Rowatt JD, Aragon AA, Baca OG (1983) Lysosomal response of a murine macrophage-like cell line persistently infected with Coxiella burnetii. Infect Immun 40:1155–1162
Baca OG, Paretsky D (1983) Q fever and Coxiella burnetii: a model for host-parasite interactions. Microbiol Rev 47:127–149
Beare PA, Howe D, Cockrell DC, Omsland A, Hansen B, Heinzen RA (2009a) Characterization of a Coxiella burnetii ftsZ mutant generated by Himar1 transposon mutagenesis. J Bacteriol 191:1369–1381
Beare PA, Unsworth N, Andoh M, Voth DE, Omsland A, Gilk SD, Williams KP, Sobral BW, Kupko JJ 3rd, Porcella SF, Samuel JE, Heinzen RA (2009b) Comparative genomics reveal extensive transposon-mediated genomic plasticity and diversity among potential effector proteins within the genus Coxiella. Infect Immun 77:642–656
Beare PA, Omsland A, Cockrell DC, Howe D, Heinzen RA (2010) Expansion of the Coxiella burnetii genetics tool box using axenic culture. Abstracts of the 24th Meeting of the American Society for Rickettsiolgy, abstracts 94, Stevenson, WA
Beron W, Gutierrez MG, Rabinovitch M, Colombo MI (2002) Coxiella burnetii localizes in a Rab7-labeled compartment with autophagic characteristics. Infect Immun 70:5816–5821
Braun RD, Lanzen JL, Snyder SA, Dewhirst MW (2001) Comparison of tumor and normal tissue oxygen tension measurements using OxyLite or microelectrodes in rodents. Am J Physiol Heart Circ Physiol 280:H2533–H2544
Burton PR, Kordova N, Paretsky D (1971) Electron microscopic studies of the rickettsia Coxiella burnetii: entry, lysosomal response, and fate of rickettsial DNA in L-cells. Can J Microbiol 17:143–150
Burton PR, Stueckemann J, Welsh RM, Paretsky D (1978) Some ultrastructural effects of persistent infections by the rickettsia Coxiella burnetii in mouse L cells and green monkey kidney (Vero) cells. Infect Immun 21:556–566
Chen SY, Vodkin M, Thompson HA, Williams JC (1990) Isolated Coxiella burnetii synthesizes DNA during acid activation in the absence of host cells. J Gen Microbiol 136:89–96
Chen C, Banga S, Mertens K, Weber MM, Gorbaslieva I, Tan Y, Luo ZQ, Samuel JE (2010) Large-scale identification and translocation of type IV secretion substrates by Coxiella burnetii. Proc Natl Acad Sci U S A 107:21755–21760
Cockrell DC, Beare PA, Fischer ER, Howe D, Heinzen RA (2008) A method for purifying obligate intracellular Coxiella burnetii that employs digitonin lysis of host cells. J Microbiol Methods 72:321–325
Coleman SA, Fischer ER, Howe D, Mead DJ, Heinzen RA (2004) Temporal analysis of Coxiella burnetii morphological differentiation. J Bacteriol 186:7344–7352
Cox HR (1938) A filter-passing infectious agent isolated from ticks. III. Description of organism and cultivation experiments. Public Health Report 53:2270–2276
Dalebroux ZD, Edwards RL, Swanson MS (2009) SpoT governs Legionella pneumophila differentiation in host macrophages. Mol Microbiol 71:640–658
Edwards RL, Dalebroux ZD, Swanson MS (2009) Legionella pneumophila couples fatty acid flux to microbial differentiation and virulence. Mol Microbiol 71:1190–1204
Ellison DW, Clark TR, Sturdevant DE, Virtaneva K, Porcella SF, Hackstadt T (2008) Genomic comparison of virulent Rickettsia rickettsii Sheila Smith and avirulent Rickettsia rickettsii Iowa. Infect Immun 76:542–550
Ewann F, Hoffman PS (2006) Cysteine metabolism in Legionella pneumophila: characterization of an L-cystine-utilizing mutant. Appl Environ Microbiol 72:3993–4000
Gilsdorf A, Kroh C, Grimm S, Jensen E, Wagner-Wiening C, Alpers K (2008) Large Q fever outbreak due to sheep farming near residential areas, Germany, 2005. Epidemiol Infect 136:1084–1087
Hackstadt T (1983) Estimation of the cytoplasmic pH of Coxiella burnetii and effect of substrate oxidation on proton motive force. J Bacteriol 154:591–597
Hackstadt T, Williams JC (1981a) Biochemical stratagem for obligate parasitism of eukaryotic cells by Coxiella burnetii. Proc Natl Acad Sci U S A 78:3240–3244
Hackstadt T, Williams JC (1981b) Stability of the adenosine 5′-triphosphate pool in Coxiella burnetii: influence of pH and substrate. J Bacteriol 148:419–425
Hackstadt T, Williams JC (1983) pH dependence of the Coxiella burnetii glutamate transport system. J Bacteriol 154:598–603
Harth G, Clemens DL, Horwitz MA (1994) Glutamine synthetase of Mycobacterium tuberculosis: extracellular release and characterization of its enzymatic activity. Proc Natl Acad Sci U S A 91:9342–9346
Heinzen RA, Scidmore MA, Rockey DD, Hackstadt T (1996) Differential interaction with endocytic and exocytic pathways distinguish parasitophorous vacuoles of Coxiella burnetii and Chlamydia trachomatis. Infect Immun 64:796–809
Howe D, Shannon JG, Winfree S, Dorward DW, Heinzen RA (2010) Coxiella burnetii phase I and II variants replicate with similar kinetics in degradative phagolysosome-like compartments of human macrophages. Infect Immun 78:3465–3474
Hu H, Sosnovsky G, Swartz HM (1992) Simultaneous measurements of the intra- and extra-cellular oxygen concentration in viable cells. Biochim Biophys Acta 1112:161–166
Imaizumi A, Suzuki Y, Ono S, Sato H, Sato Y (1983) Heptakis(2,6-O-dimethyl)beta-cyclodextrin: a novel growth stimulant for Bordetella pertussis phase I. J Clin Microbiol 17:781–786
Kashiwagi K, Miyamoto S, Suzuki F, Kobayashi H, Igarashi K (1992) Excretion of putrescine by the putrescine-ornithine antiporter encoded by the potE gene of Escherichia coli. Proc Natl Acad Sci U S A 89:4529–4533
Khan N, Shen J, Chang TY, Chang CC, Fung PC, Grinberg O, Demidenko E, Swartz H (2003) Plasma membrane cholesterol: a possible barrier to intracellular oxygen in normal and mutant CHO cells defective in cholesterol metabolism. Biochemistry 42:23–29
Marchini A, D’apolito M, Massari P, Atzeni M, Copass M, Olivieri R (1995) Cyclodextrins for growth of Helicobacter pylori and production of vacuolating cytotoxin. Arch Microbiol 164:290–293
Maurin M, Benoliel AM, Bongrand P, Raoult D (1992) Phagolysosomes of Coxiella burnetii-infected cell lines maintain an acidic pH during persistent infection. Infect Immun 60:5013–5016
Mcleod MP, Qin X, Karpathy SE, Gioia J, Highlander SK, Fox GE, Mcneill TZ, Jiang H, Muzny D, Jacob LS, Hawes AC, Sodergren E, Gill R, Hume J, Morgan M, Fan G, Amin AG, Gibbs RA, Hong C, Yu XJ, Walker DH, Weinstock GM (2004) Complete genome sequence of Rickettsia typhi and comparison with sequences of other rickettsiae. J Bacteriol 186:5842–5855
Omsland A, Cockrell DC, Fischer ER, Heinzen RA (2008) Sustained axenic metabolic activity by the obligate intracellular bacterium Coxiella burnetii. J Bacteriol 190:3203–3212
Omsland A, Cockrell DC, Howe D, Fischer ER, Virtaneva K, Sturdevant DE, Porcella SF, Heinzen RA (2009) Host cell-free growth of the Q fever bacterium Coxiella burnetii. Proc Natl Acad Sci U S A 106:4430–4434
Omsland A, Beare PA, Hill J, Cockrell DC, Howe D, Hansen B, Samuel JE, Heinzen RA (2011) Isolation from animal tissue and genetic transformation of Coxiella burnetii are facilitated by an improved axenic growth medium. Appl Environ Microbiol 77:3720–3725
Paretsky D, Consigli RA, Downs CM (1962) Studies on the physiology of rickettsiae. III. Glucose phosphorylation and hexokinase activity in Coxiella burnetii. J Bacteriol 83:538–543
Sauer JD, Bachman MA, Swanson MS (2005) The phagosomal transporter A couples threonine acquisition to differentiation and replication of Legionella pneumophila in macrophages. Proc Natl Acad Sci U S A 102:9924–9929
Schimmer B, Dijkstra F, Vellema P, Schneeberger PM, Hackert V, Ter Schegget R, Wijkmans C, Van Duynhoven Y, Van Der Hoek W (2009) Sustained intensive transmission of Q fever in the south of the Netherlands, 2009. Euro Surveill 14:19210
Schnappinger D, Ehrt S, Voskuil MI, Liu Y, Mangan JA, Monahan IM, Dolganov G, Efron B, Butcher PD, Nathan C, Schoolnik GK (2003) Transcriptional adaptation of Mycobacterium tuberculosis within macrophages: insights into the phagosomal environment. J Exp Med 198:693–704
Seshadri R, Paulsen IT, Eisen JA, Read TD, Nelson KE, Nelson WC, Ward NL, Tettelin H, Davidsen TM, Beanan MJ, Deboy RT, Daugherty SC, Brinkac LM, Madupu R, Dodson RJ, Khouri HM, Lee KH, Carty HA, Scanlan D, Heinzen RA, Thompson HA, Samuel JE, Fraser CM, Heidelberg JF (2003) Complete genome sequence of the Q-fever pathogen Coxiella burnetii. Proc Natl Acad Sci U S A 100:5455–5460
Stephens RS, Kalman S, Lammel C, Fan J, Marathe R, Aravind L, Mitchell W, Olinger L, Tatusov RL, Zhao Q, Koonin EV, Davis RW (1998) Genome sequence of an obligate intracellular pathogen of humans: Chlamydia trachomatis. Science 282:754–759
Teixeira De Mattos MJ, Neijssel OM (1997) Bioenergetic consequences of microbial adaptation to low-nutrient environments. J Biotechnol 59:117–126
Tesh MJ, Morse SA, Miller RD (1983) Intermediary metabolism in Legionella pneumophila: utilization of amino acids and other compounds as energy sources. J Bacteriol 154:1104–1109
Voth DE, Beare PA, Howe D, Sharma UM, Samoilis G, Cockrell DC, Omsland A, Heinzen RA (2011) The Coxiella burnetii cryptic plasmid is enriched in genes encoding type IV secretion system substrates. J Bacteriol 193:1493–1503
Williams JC, Peacock MG, Mccaul TF (1981) Immunological and biological characterization of Coxiella burnetii, phases I and II, separated from host components. Infect Immun 32:840–851
Zuerner RL, Thompson HA (1983) Protein synthesis by intact Coxiella burnetii cells. J Bacteriol 156:186–191
Acknowledgements
I would like to thank Robert A. Heinzen for critical review of this manuscript, and Anita Mora and Austin Athman for graphic illustrations. This work was funded by the Intramural Research Program of the National Institutes of Health, National Institute of Allergy and Infectious Diseases
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Omsland, A. (2012). Axenic Growth of Coxiella burnetii . In: Toman, R., Heinzen, R., Samuel, J., Mege, JL. (eds) Coxiella burnetii: Recent Advances and New Perspectives in Research of the Q Fever Bacterium. Advances in Experimental Medicine and Biology, vol 984. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-4315-1_11
Download citation
DOI: https://doi.org/10.1007/978-94-007-4315-1_11
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-4314-4
Online ISBN: 978-94-007-4315-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)