
Abstract
This chapter reexamines our research on secondary students’ understanding of genetics in terms of gene conceptions and reasoning when they learned genetics with multiple external representations (MERs). In our Australian study, teachers in three Perth schools included interactive BioLogica activities which feature manipulable MERs to varying degrees in their teaching. In our Hong Kong study, students used BioLogica in an after-school program for which the teacher and researcher provided bilingual support and both group and individual feedback. The results of our studies—from a cross-case analysis of students’ gene conceptions and genetics reasoning based on interviews, online two-tier tests, computer log files, and other data sources—indicated that most students improved their understanding of genetics to varying extent in terms of sophistication of their gene conceptions and the six types of genetics reasoning. The findings suggest that MERs supported understanding of genetics but not for all students. We compare the Australian and Hong Kong studies in terms of students’ genetics reasoning, explore how students learned complex content in biology using MERs within different learning contexts, and discuss the potential of visual-graphical and bilingual representations for scaffolding the learning of English language learner (ELL) students.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Introduction
In this chapter, we first discuss the theoretical aspects of learning genetics and learning with multiple external representations (MERs) by reviewing the literature relevant to our studies. Next, we reexamine our Australian study focusing on students’ understanding in terms of gene conceptions and genetics reasoning from a cross-case analysis of data from three senior secondary schools in Perth (Tsui & Treagust, 2007, 2010). We also report on our recent Hong Kong study (Tsui, 2009), compare its results with those of the Australian study, and explore how students learned complex content in biology using MERs within different learning contexts, including the role of language in learning. Pseudonyms are used throughout this chapter to maintain anonymity of all participants in our studies.
Genetics is Conceptually and Linguistically Difficult
Over the past decades, research has shown that genetics not only is a conceptually difficult topic in school biology because that knowledge is organized at multiple levels but also is a linguistically difficult content area because of its large content-specific vocabulary (e.g., Bahar, Johnstone, & Hansell, 1999; Hackling & Treagust, 1984; Horwitz & Christie, 2000; Pearson & Hughes, 1988; Stewart, 1982; Venville & Treagust, 1998; Wood, 1996). Learning genetics requires multilevel thinking—phenotypes of an organism are at the macroscopic level, whereas cells and chromosomes are at the microscopic level, DNA is at submicroscopic level, and genotypes are at the symbolic level (e.g., Johnstone, 1991). Student understanding of genetics also depends on dealing with these concepts and processes simultaneously at several levels of organization, on connecting them as an interrelated whole (Marbach-Ad & Stavy, 2000), and on reasoning with concepts and processes across ontologically distinct levels (e.g., genes or DNA molecules are informational but the traits they control are physical) (Duncun & Reiser, 2007).
Science educators have recently called for improving the ways to teach the complexity of the gene concept and for using better approaches to address both the complex content of genetics and the inadequate current instructional methods and materials in schools (e.g., Duncan, Rogat, & Hmelo-Silver, 2009; Venville & Donovan, 2005). Genetics literacy—“being able to comprehend, use or respond to information about genetic phenomena and technologies” (Duncan et al., 2009, p. 657)—is needed for all citizens in order to better understand the emerging contemporary issues such as genetic modifications, genomics, or cloning and to make informed judgments and decisions.
Learning Genetics as Understanding: Gene Conceptions and Genetics Reasoning
In our studies, we considered student learning of genetics as conceptual understanding in terms of their gene conceptions and genetics reasoning. Theoretically, we drew on a multidimensional conceptual change framework (Tyson, Venville, Harrison, & Treagust, 1997) to address the acknowledged limitations of the traditional, largely epistemological conceptual change model of Posner, Strike, Hewson, and Gertzog (1982). In reexamining the results, we consider Vygotskian perspectives that emphasize the role of social and cultural contexts and that of language in learning (Vygotsky, 1968, 1978), as well as some perspectives about learning from psycholinguistic research (e.g., Kroll & Hermans, 2011; Lin, 2006).
The first focus in our Australian study was on student understanding as developing ontological conceptual change in conceptualizing the gene from being a particle to a sequence of instructions as in Venville and Treagust’s (1998) study in which the grade 10 students developed their conceptions through a pathway indicating their progressively more sophisticated mental models of the gene (inactive particle gene → active particle gene → sequence of instructions gene → productive sequence of instructions gene). The second focus was on students’ understanding in terms of reasoning that can be diagnosed by a two-tier instrument (Treagust, 1988) which we developed and used in three Perth schools for pre- and post-instructional evaluation of students’ genetics reasoning (Tsui & Treagust, 2010). The two-tier diagnostic instrument was subsequently modified and used in our Hong Kong study. The two-tier test items evaluate students’ genetics reasoning using Hickey and Kindfield’s (1999) matrix of reasoning (Tsui & Treagust, 2003, 2010) (see Table 15.1).
As indicated by Table 15.1, genetics reasoning in our studies required students to use both logical reasoning (domain-general dimension) and information in their subject content (domain-specific dimension) for understanding. Novice reasoners often use mental representations of only one antecedent condition in reasoning tasks to arrive at the conclusion, whereas expert reasoners use two or more antecedent conditions in such reasoning processes and become more reflective and active in seeking alternatives and making inferences to draw conclusions (Lawson, 1992).
Therefore, we can explain how students reason in completing the tasks of genetics reasoning Types I–IV (see Table 15.1). For example, to solve pedigree problems that require Types III and IV reasoning, students need to reason by mapping given phenotypes to unknown genotypes of the parents (effect-to-cause), respectively, within and between generations. Mapping in Types III and IV is more difficult compared to that in Types I and II (cause-to-effect) because the former is not a one-to-one mapping, that is, more than one genotype may correspond to the same given phenotype. In solving human pedigree problems, Hackling and Lawrence (1988) also pointed out that the expert problem solvers are able to identify critical cues in the problems, test hypotheses with genotypes assigned to phenotypes, and use given evidence to support or falsify an alternative hypothesis before arriving at the answer. It was based on these six types of reasoning that we designed the interview reasoning tasks (Tsui & Treagust, 2003) and the two-tier diagnostic instrument in the Australian study (Tsui & Treagust, 2010).
Multiple Representations and BioLogica
Biology teachers have long been using different external representations (ERs) in classroom teaching to communicate ideas to students by voice, writing, drawings, diagrams, images, gestures, and so on. Students’ conceptions can be regarded as their internal representations or mental models of an object or event (Duit & Glynn, 1996) constructed from the ERs of these entities. Models of scientific objects or processes can be considered as ERs for modeling in model-based learning which plays a central role in science education (Justi & Gilbert, 2002).
Visualizations, as ERs, have been important in learning since the advent of computer technology and are now being widely used for learning science and in the media to convey scientific information (Gobert, 2005). In our studies, we explored the pedagogical functions of using more than one form of external representations or multiple external representations (MERs) in learning (cf. van Someren, Reimann, Boshuizen, & de Jong, 1998). In particular, we utilized Ainsworth’s (1999) functional taxonomy of MERs to argue that MERs can support learning in three ways: (1) by providing/supporting complementary information and/or cognitive processes, (2) by constraining interpretations or misinterpretations of phenomena, and (3) by promoting the construction of a deeper understanding of concepts through abstraction, such as detecting and extracting a subset of relevant elements from a representation; extension or extending knowledge learned in one representation to new situations with other representations; and relations, such as translating between two or more unfamiliar representations. However, learning with MERs may not always be useful because of the new costs and challenges (Ainsworth, Bibby, & Wood, 1997).
In this chapter, we argue that MERs appear to be a promising construct for improving learning of complex concepts in biology because biological knowledge is hierarchically organized (Marbach-Ad & Stavy, 2000) at ontologically distinct levels (Duncun & Reiser, 2007). The computer-based activities of BioLogica (Concord Consortium, 2002)—a hypermodel (Horwitz, 1995; Horwitz & Tinker, 2001) or an interactive, exploratory environment for learning genetics—were used in our studies. BioLogica features dynamically linked MERs of genetics that allow users to manipulate objects of genetics represented at different levels of biological organization—DNA, genes, chromosomes, gametes, cells, organisms, pedigrees, and populations—and observe the changes in their behavior as a result of manipulation in ways constrained by models based on transmission genetics and molecular/cellular mechanisms (Buckley et al., 2004; Gobert et al., 2011) (see Fig. 15.1).

A screenshot of BioLogica activity Meiosis showing organism level, cell level, and chromosome level of dragons (an imaginary species)
BioLogica guides learners’ interaction with the activities through a sequence of challenges, monitors their progress, and provides learners with feedback and helpful hints as they work through progressively more challenging activities. The interactions in these activities are controlled and implemented by a software component called activity scripts (Horwitz & Tinker, 2001) having different pedagogical functions—such as narratives, tasks and puzzles, representational assistance, reasoning models, explanations and feedback on actions and responses, embedded assessment questions, and reflective questions—that mediated the students’ conceptual learning and reasoning (Buckley et al., 2004). The learning goals of eight BioLogica activities completed by most of the students in our studies are shown in Table 15.2.
The data about the students’ interactions (e.g., which screen a student used in answering a question or what graphic objects the student accessed and in what order) can be logged by the BioLogica program in the form of log files automatically generated and saved on an individual computer, a school network, or a remote server of a research center so that teachers and researchers can analyze the logged data. As Horwitz and Tinker (2001) explained, the log files that track student interactions in BioLogica are “a promising research tool that allows us to obtain at a distance detailed information about student thinking, knowledge, and problem-solving strategies” (p. 13).
A large-scale study on model-based learning using BioLogica in schools across the United States indicated that the experimental groups outperformed the control groups in understanding genetics (Buckley et al., 2004). Further analyses showed that those students with better understanding of models as multiple representations learned significantly better about the content of genetics in BioLogica activities than did those with less understanding of models as such (Gobert et al., 2011).
Research Questions
In this chapter we attempt to focus on two research questions about the understanding of genetics in terms of their gene conceptions and genetics reasoning by discussing and comparing the results of our Australian and Hong Kong studies: (1) What are the students’ pre-instructional and post-instructional gene conceptions? (2) In what ways and how do the MERs of BioLogica promote students’ genetics reasoning?
Method
Research Approach
In our Australian study, we adopted an interpretive research approach (Erickson, 1998) involving largely qualitative case study methods (Merriam, 1998). In particular, we drew on Merriam’s three major features that characterize a case study—particularistic, descriptive, and heuristic—and the research strategies (e.g., prolonged engagement, persistent observation, member checks, and triangulation) for increasing the research rigor of a qualitative case study (cf. Denzin & Lincoln, 1994; Erickson, 1998).
School Context
The three case schools in our Australian study were School F (a state co-ed school), School O (an independent girls’ school), and School U (a state co-ed school) in Perth, Western Australia. The study was first conducted in a 10th-grade class in School F when genetics was taught and then in Schools O and U in the following year with similar methods. The four participating biology teachers had teaching experiences ranging from 9 to 27 years, and their participating students (72 girls and 17 boys), aged from 14 to 18 years, in three 10th-grade classes (Schools F and O) and two 12th-grade classes (School U), were mostly Australian-born and native speakers of English. Research ethics (e.g., voluntary participation, informed consent, use of pseudonyms for participants) was strictly followed (Tsui & Treagust, 2007, 2010).
The second case study involved action research in a government-subsidized girls’ secondary school in Hong Kong, a special administrative region of China since 1997 (Tsui, 2009). This study was a learning project—a collaboration between the first author and a biology teacher with support from the school—for improving students’ scientific reasoning and writing biology in English (Tsui, 2009). The 20 participating students who volunteered to take part were 10th-grade girls of average age of 15.6 years; they were all Hong Kong Chinese with English as their second language and Chinese as their first language (their native dialect is Cantonese and written language is Modern Standard Chinese). Before the study, the 10th graders in this Chinese Medium of Instruction (CMI) school had completed their first 3 years (grades 7–9) of secondary education in CMI. Since the beginning of the first semester of their 10th-grade year, these English-as-a-second-language (ESL) or English language learner (ELL) students had used English as the medium of instruction (EMI) for learning some subjects, including biology. This change in the medium of instruction is common in many CMI schools in Hong Kong. They had not learned genetics before this study because genetics was part of their 11th-grade biology curriculum.
Over 8 weeks in the second semester, these 10th graders learned genetics in weekly after-school computer sessions using BioLogica activities. Their biology teacher, Ms Chan, who had 15 years of teaching experience, collaborated with the first author to provide scaffolding and support in all the weekly computer learning sessions. Both the teacher and the first author are bilingual speakers of English and Cantonese.
Data Collection and Analysis
Australian Study
Although the participating teachers in the three schools all included BioLogica activities for their student learning, they also used other teaching aids and learning resources. The data from multiple sources—before, during, and after teaching with BioLogica activities—were collected: transcripts of semi-structured student interviews, online results of the two-tier pretests/posttests and open-ended questionnaires (delivered by WebCT, Curtin University’s then e-learning system), computer log files on students’ usage of BioLogica, classroom observation field notes and audio recordings transcripts, the first author’s reflective journals, and teachers’ handouts and other school documents.
To evaluate Australian students’ understanding in terms of gene conceptions, we analyzed their open-ended questionnaire responses, interview and lesson transcripts, and other qualitative data. We interviewed 26 target students in the three Perth schools, selected from each class on the basis of their scores in the online pretests on genetics reasoning to include students from high and low groups. The interview protocols used were the same in the three schools except that for School U no reasoning tasks were included (Tsui & Treagust, 2007, 2010). We used the two-tier posttest to diagnose students’ understanding of genetics in terms of reasoning and analysis of some target students’ log files. Both the two-tier tests and interview reasoning tasks were designed to evaluate students’ six types of genetics reasoning.
Hong Kong Study
In this study, only five sources of data used in the Australian study were collected—interviews of students, open-ended questionnaire (gene conception) and two-tier posttest (genetics reasoning), BioLogica log files (tracking student interactions with MERs), and teacher’s handouts and other documents. We also analyzed students’ written answers to the parallel open-ended questionnaire in the paper-and-pencil pretest and posttest What do you know about a gene? for identifying their gene conceptions using the framework of Venville and Treagust (1998). We interviewed four target students, from the high- and low-ability group based on their school examination results, before and after instruction. Unlike the Australian study, we used the two-tier posttest only to diagnose students’ understanding of genetics in terms of reasoning to respect the biology teacher’s suggestion. We also conducted analyses of the log files and correlation analyses to explore the relations between students’ genetics reasoning and other variables.
Results
Identifying Common Gene Conceptions
In a cross-case analysis of the Australian students’ gene conceptions before and after instruction—based on their responses to an open-ended questionnaire What do you know about a gene? in the online pretest and posttest—we identified five common gene conceptions of the 10th graders in a way similar to the findings of Venville and Treagust (1998). A student could hold more than one gene conception. As shown in Table 15.3, the most common gene conception was: “A gene is from parents/grandparents.”
In our Hong Kong study, the analysis of students’ written answers to the same parallel open-ended questionnaire in the paper-and-pencil pretest and posttest (What do you know about a gene?) indicated that their gene conceptions could be categorized into four gene conceptions along a pathway of progressively more sophisticated conceptions of the gene as reported by Venville and Treagust (1998) (see Table 15.4). These results suggest that the Hong Kong students improved their understanding of the gene in terms of developing progressively sophisticated conceptions of the gene after their learning with eight BioLogica activities they had done weekly over 2 months.
Learning to Write Genetics with Confidence: Some Examples
Although only some Hong Kong students could fully express their understanding in writing about genetics, most of them improved their confidence in writing English despite their grammatical and other errors. Bilingual support (e.g., bilingual glossary of genetic terms in English and Modern Standard Chinese), on-site scaffolding, weekly feedback of the first author to the students by returning to them their log files with feedback comments, and collaborative classroom discourse with mixed-code (English and Cantonese) discussions—all appeared to be conducive to their learning (Tsui, 2009). For example, some students like Mei-ling (see Figs. 15.2 and 15.3) who used Chinese and diagrams to represent their gene conceptions at the pretest became more confident at the posttest to write in English; some used mixed-code in their representations.

Pretest answers of Mei-ling (16 years old) who used Chinese to describe her gene conception. Her pre-instructional gene conception was categorized as C2 (see Table 15.4)

Posttest answers of Mei-ling (16 years old) who wrote in English but used Chinese “遺傳” for inherit and “分裂” for divide to complete this mixed-code sentence to represent her post-instructional gene conception which remained unchanged as C2 (see Table 15.4)
The pretest and posttest open-ended questionnaires were both What do you know about a gene? and at the pretest students were allowed to write in either Chinese or English and use diagrams to illustrate their answers, but at the posttest they were asked to write in English (for some examples of students’ answers, see Figs. 15.2, 15.3, 15.4, and 15.5).

Pretest answers of Lai-ming (16 years old) whose answers were in Chinese. Her pre-instructional gene conception was categorized as C1 (see Table 15.4)

Posttest answers of Lai-ming (16 years old) whose gene conception had progressed from C1 to C3 after instruction (see Table 15.4)
Comparing Genetics Reasoning of Students from Hong Kong and Perth
In our Australian study, for the students in all three schools, a paired t test indicated that their genetics reasoning posttest scores were significantly higher than their pretest scores at p < 0.01 (see Table 15.5).
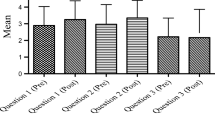
One limitation in our Hong Kong study is that there was no genetics reasoning pretest for a pre-post comparison; however, an analysis of descriptive statistics of the posttest scores (M = 31.25, SD = 16.42) of the Hong Kong students (N = 20) showed that their posttest reasoning patterns by types were as predicted according to the difficulty level of the six types of reasoning (Tsui & Treagust, 2003, 2010). Analyses also showed that they had achieved the similar patterns at the posttest comparable to those of their Australian counterparts as indicated in Figs. 15.6 and 15.7; nevertheless their mean scores were much lower because they had not previously studied genetics in school.

Genetics reasoning by types in two-tier posttest of Hong Kong 10th-grade students (N = 20)

Genetics reasoning by types in two-tier posttest of Australian 10th graders (n = 56; n = 33 for Types II and V items) and 12th graders (n = 14)
Analysis of other results suggest that student performance in genetics reasoning in our Hong Kong study depended on their prior knowledge of biology and English language proficiency as indicated by Pearson correlation analyses using the students’ school examination scores before the study—prior knowledge of biology (r = .512; p = .021, two-tailed; N = 20) and English language proficiency (r = .57; p = .008, two-tailed; N = 20) were significantly correlated with the genetics reasoning scores in the posttest of the study (p < .05).
Analyses of Students’ Log Files
The computer log files that tracked students’ interaction with BioLogica were subsequently analyzed to explore how students learned during the computer activities. Log file specifications (e.g., time in screen, interaction time, inputs to model, answers, typing time) and their analysis can be useful for understanding how students interact with the MERs in terms of their model-based learning and their modeling skills (Buckley et al., 2004; Gobert, 2005).
There was also a limitation in our Australian study. We did not have a complete set of log files so that we could only analyze some case studies of students’ log files to examine how they interacted with the MERs of BioLogica and with each other (from classroom audio recordings) during the activities. For example, a dyad of 12th graders, Helena and May of School U in Perth, had the following episode in which they had dialogic interactions while working on a task of the Monohybrid activity that could be interpreted by juxtaposing Helena’s log file with a reconstructed screenshot of the BioLogica program she was using (see Table 15.6 and Fig. 15.8).

A screenshot of the BioLogica activity Monohybrid reconstructed based on the information of the corresponding log file of Helena (from 16:13:45 to 16:14:10)
Verbatim transcription of the audio recordings of the two students’ dialogic interactions in Table 15.6 indicates that Helena was being encouraged (e.g., lines 4–6 in Table 15.6) and scaffolded (lines 9–12; 14–15) by May, her more confident peer, during this same predict-observe-explain (POE) task on which they were working at their own computers next to each other. After May had read out the question on the screen (lines 18–22 in Table 15.6), Helena did not answer the question to predict what would happen as indicated in the log file (no text was logged between markup tags “</action>” and “<action>” before the time 16:14:10), but she went on with the POE task to breed 30 baby dragons to observe what happened next. The last part of the log file in Table 15.6 summarizes the results of Helena’s action. Then, in the next part the computer would ask the users to explain (not shown in Table 15.6).
In the Hong Kong study, we had collected a complete set of the log files of all students and we analyzed in detail the log files of four target students and the students’ errors in the activities. The following episode illustrates Mei-yee’s (one of the four target students) interactions with the computer during the Monohybrid activity. She was working on a task of Type II reasoning (see Table 15.1) as illustrated by a snippet of the log file tracking her interactions with BioLogica that corresponded to the reconstructed screenshot at 16:31:48 (see Figs. 15.9 and 15.10).

A snippet of Mei-yee’s log file corresponding to the computer-user interactions that followed the screenshot in Fig. 15.10

A reconstructed screenshot of Mei-yee’s interactions with BioLogica Monohybrid activity as tracked by the log file with the snippet shown in Fig. 15.9
As the log file in Fig. 15.9 indicates, at 16:28:10, Mei-yee had just successfully completed her first task to use the Punnett square to work out the possible combinations of alleles in a monohybrid cross between two dragon parents (a fancy-tailed dad and plain-tailed mom, i.e., Tt × tt). Then, she was asked to select all the zygotes in the Punnett square that would develop into plain-tailed baby dragons in order to work out the proportion of plain-tailed baby dragons in the offspring.
The log file in Fig. 15.9 continues to indicate that after Mei-yee’s first attempt to select the right zygotes failed, she repeatedly viewed the chromosomes of the parents’ to check out their genotypes (by clicking on the dragon icons). However, she made another wrong attempt before she finally selected the right zygotes at 16:31:48. That is, the two zygotes with genotype tt or two of the four possible cases in the cross (tt × Tt) that would develop into plain-tailed baby dragons. In so doing, she had achieved the two learning goals of the Monohybrid activity for using a Punnett square in solving problems in Mendelian genetics (see Table 15.2).
We next analyzed the log files of three selected activities— Meiosis, Monohybrid, and Mutations—by counting the number of students’ errors in using the BioLogica activities, including their wrong answers to questions and unsuccessful attempts to solve problems (e.g., Mei-yee made two errors during part of the Monohybrid activity as indicated by the log file in Fig. 15.9). We wanted to find out the relation between these students’ errors and their genetics reasoning as indicated by the two-tier posttest results. The results of an SPSS correlation analysis indicated that students’ genetics reasoning skills (r = −.428; p = 0.034, one-tailed; N = 19) had significant negative correlation (p < .05, one-tailed) with their errors in using these three BioLogica activities. These results suggest that the tasks, puzzles, and embedded assessment questions of the BioLogica activities can be used to evaluate students’ understanding of genetics in terms of the six types of reasoning as they work through the progressively challenging activities.
Discussion and Conclusions
The findings of our studies suggest that the MERs of BioLogica provided students with complementary information and processes about genetics across the dynamically linked levels of organization. These manipulable MERs, particularly the visual-graphical representations of the genetic phenomena, co-deployed simultaneously with scripts or texts—including narratives, tasks and puzzles, representational assistance, reasoning models, and explanations and feedback—are pedagogically useful in mediating the students’ learning (Buckley et al., 2004). From a conceptual change perspective, the MERs increase the intelligibility of the gene concept so that students can continue to engage in their learning toward developing more sophisticated gene conceptions. The progressively challenging BioLogica activities are useful in developing students’ reasoning skills. The MERs in the activities allow students to initially start to think about the genetic phenomena at the macro level (organisms, pedigrees, and populations) before moving on to understandings at the micro level (cells and chromosomes), at the submicro level (DNA), and at the symbolic level (genetic code and genotypes). In other words, students’ interpretation of a less familiar or more abstract representation of a genetic phenomenon is being constrained by the more familiar dragons in BioLogica in ways compatible to Ainsworth’s (1999) functional taxonomy of MERs.
The participating teachers in the three Australian case schools played an important role in determining what and how students benefited from their learning with MERs by providing various classroom contexts for learning. They took different approaches in using BioLogica activities in their teaching to suit their beliefs and their students’ learning styles, thus providing different learning opportunities for students during the genetics course.
The results of our Hong Kong case study largely corroborated what we had found in our Australian study in the terms of the range of student gene conceptions and their reasoning skills. It is interesting that the reasoning pattern of the Hong Kong students was similar to that of the Australian counterparts in Perth notwithstanding the linguistically and culturally different learning contexts across the schools in the two cities. Visualization can play an important role in scaffolding knowledge construction and conceptual understanding for the Hong Kong students who are English language learners as shown by some studies (e.g., Dixon, 1995).
Most students in our Hong Kong study appeared to have learned some reasoning skills and improved in their confidence to write biology in English. Although the Hong Kong students had not learned genetics before the study, their prior knowledge of genetics in Chinese acquired from the media and the Internet appeared to have helped their understanding of genetics in English. Code-mixing in classroom discourse helped these ELL students to access and capitalize on their L1 (Cantonese) linguistic resources for learning concepts in science in their L2 (English) (e.g., Lin, 2006). This interpretation is compatible with the sociocultural perspectives of learning, especially in terms of the constructs of zone of proximal development (Vygotsky, 1978) and verbal thought (Vygotsky, 1968).
Psycholinguistic research has indicated that bilinguals are able to learn concepts by way of the developmental shift from lexical mediation between their L1 and L2 to direct word-concept access or conceptual links in their L1 and then L2 (Kroll & Hermans, 2011) as shown in Fig. 15.11.

A proposed model adapted from Kroll and Hermans (2011, p. 18) to illustrate how Hong Kong ELL students might possibly learn the concept of the gene in a bilingual way. The links indicated by dotted arrows will become solid when learners have acquired better L2 skills
Accordingly, such a shift depends on the bilingual learners’ increasing ability to directly process the concepts in L2 without L1 mediation. For example, using mixed-coding and code-switching, a Hong Kong science teacher can embed key terms in L2 and concepts in a rich L1 semantic context and then illustrate L2 abstract scientific concepts with concrete L1 everyday life experiences and examples and so on (Lin, 2006). In terms of the second pedagogical function of MERs, the interpretation of biological concepts in a less familiar L2 representation can be constrained by its more familiar L1 representation for better understanding.


Unlike the Australian study, where the teachers taught genetics and used BioLogica activities to a lesser or greater extent in class to support student learning, the Hong Kong students learned genetics largely from BioLogica activities. The Hong Kong students completed all eight BioLogica activities and were seldom absent from the after-school program. Therefore, we have reason to believe that the causal relation between the usage of the interactive activities and student understanding should be stronger in the Hong Kong study. Just as in our Australian study, we found that mere engagement in the BioLogica activities interacting with the MERs may not be useful for developing deep understandings. Apart from the difference in the individual and classroom factors, interactions with the MERs in BioLogica need to be mindful and intrinsically motivated for students to benefit from such interactions in developing their understanding (Tsui & Treagust, 2004).
We summarize here the implications of our research discussed in this chapter. First, the MERs of BioLogica within different classroom contexts—Australian teachers’ stories and games, web-based activities, and other approaches, as well as Hong Kong’s bilingual and mixed-code classroom discourse—appeared to provide different learning opportunities for students to undergo conceptual change toward developing more sophisticated gene conceptions. Second, complementary and constraining functions of the MERs appeared to promote students’ construction of deep understanding of the genetic phenomena. Thus, they were able to move on to coherently relate the hierarchically arranged objects and events of genetics, abstract the genetic phenomena (phenotypes and inheritance patterns) to symbols (genotypes and DNA code) for reasoning and problem solving, and extend such understandings (e.g., sex-linkage) to real-life human examples. Third, MERs of BioLogica appeared to provide scaffolding for ELL students with limited English language skills for developing scientific reasoning by way of visual-graphical representations dynamically linked to texts. Furthermore, bilingual representations and discourse also might have scaffolded these ELL students to develop better understanding.
To conclude, these two case studies have provided some detailed evidence and thick descriptions (Merriam, 1998) for the claims that learning with multiple representations can be pedagogically useful (cf. Ainsworth, 1999) within different learning contexts for students’ conceptual understanding and reasoning in biology, particularly for students from diverse backgrounds. We believe this is important at a time when the latest trend of science education is directed toward globalization (cf. Chiu & Duit, 2011).
References
Ainsworth, S. (1999). The functions of multiple representations. Computers in Education, 33(2/3), 131–152.
Ainsworth, S., Bibby, P. A., & Wood, D. J. (1997). Information technology and multiple representations: New opportunities – new problems. Journal of Information Technology for Teacher Education, 6(1), 93–104.
Bahar, M., Johnstone, A. H., & Hansell, M. H. (1999). Revisiting learning difficulties in biology. Journal of Biological Education, 33(2), 84–86.
Buckley, B. C., Gobert, J. D., Kindfield, A. C. H., Horwitz, P., Tinker, R., Gerlits, B., et al. (2004). Model-based teaching and learning with BioLogicatm: What do they learn? How do they learn? How do we know? Journal of Science Education and Technology, 13(1), 23–41.
Chiu, M.-H., & Duit, R. (2011). Globalization: Science education from an international perspective. Journal of Research in Science Teaching, 48(6), 553–566.
Concord Consortium. (2002). BioLogica teachers’ guide. Retrieved June, 2002, from http://mac.concord.org/downloads/
Denzin, N., & Lincoln, Y. S. (Eds.). (1994). Handbook of qualitative research. Thousand Oaks, CA: Sage.
Dixon, J. K. (1995). Limited English proficiency and spatial visualization in middle school students’ construction of the concepts of reflection and rotation. Bilingual Research Journal, 19(2), 221–247.
Duit, R., & Glynn, S. (1996). Mental modelling. In G. Welford, J. Osborne, & P. Scott (Eds.), Research in science education in Europe: Current issues and themes (pp. 166–176). London: The Falmer Press.
Duncun, R. G., & Reiser, B. J. (2007). Reasoning across ontologically distinct levels: Students’ understanding of molecular genetics. Journal of Research in Science Teaching, 44(7), 938–959.
Duncan, R. G., Rogat, A. D., & Hmelo-Silver, C. E. (2009). A learning progression for deepening students’ understandings of modern genetics across the 5th-10th grades. Journal of Research in Science Teaching, 46(6), 655–674.
Erickson, F. (1998). Qualitative research methods for science education. In B. J. Fraser & K. G. Tobin (Eds.), International handbook of science education (pp. 1115–1173). Dordrecht, The Netherlands: Kluwer.
Gobert, J. D. (2005). A model of molecular visualization. In J. K. Gilbert (Ed.), Visualization in science education (Vol. 1, pp. 61–72). Dordrecht, The Netherlands: Springer.
Gobert, J. D., O’Dwyerb, L., Horwitz, P., Buckley, B. C., Levy, S. T., & Wilensky, U. (2011). Examining the relationship between students’ understanding of the nature of models and conceptual learning in biology, physics, and chemistry. International Journal of Science Education, 33(5), 653–684.
Hackling, M. W., & Lawrence, A. J. (1988). Expert and novice solutions of genetic pedigree problems. Journal of Research in Science Teaching, 25(7), 531–546.
Hackling, M. W., & Treagust, D. F. (1984). Research data necessary for meaningful review of grade ten high school genetics curricula. Journal of Research in Science Teaching, 21(1), 197–209.
Hickey, D. T., & Kindfield, A. C. H. (1999, April). Assessment-oriented scaffolding of student and teacher performance in a technology-supported genetics environment. Paper presented at the annual meeting of the American Educational Research Association, Montreal, Canada.
Horwitz, P. (1995). Linking model to data: Hypermodels for science education. The High School Journal, 79(2), 148–156.
Horwitz, P., & Christie, M. A. (2000). Computer-based manipulatives for teaching scientific reasoning: An example. In M. J. Jacobson & R. B. Kozma (Eds.), Innovations in science and mathematics education: Advanced design for technologies of learning (pp. 163–191). Hillsdale, NJ: Lawrence Erlbaum Associates.
Horwitz, P., & Tinker, R. (2001). Pedagogica to the rescue: A short history of hypermodels. Concord Newsletter, 5(1), 1–4.
Johnstone, A. H. (1991). Why is science difficult to learn? Things are seldom what they seem. Journal of Computer Assisted Learning, 7, 75–83.
Justi, R. S., & Gilbert, J. K. (2002). Science teachers’ knowledge about and attitudes towards the use of models and modelling in learning science. International Journal of Science Education, 24(12), 1273–1292.
Kroll, J. F., & Hermans, D. (2011). Psycholinguistic perspectives on language processing in bilinguals. In M. S. Schmid & W. Lowie (Eds.), Modeling bilingualism: From structures to chaos (pp. 15–36). Amsterdam/Philadelphia: John Benjamins.
Lawson, A. E. (1992). What do tests of “formal” reasoning actually measure? Journal of Research in Science Teaching, 29(9), 965–983.
Lin, A. (2006). Beyond linguistic purism in language-in-education policy and practice: Exploring bilingual pedagogies in a Hong Kong science classroom. Language and Education, 20(4), 287–305.
Marbach-Ad, G., & Stavy, R. (2000). Students’ cellular and molecular explanations of genetic phenomena. Journal of Biological Education, 34(4), 200–205.
Merriam, S. B. (1998). Qualitative research and case study applications in education. San Francisco: Jossey-Bass.
Pearson, J. T., & Hughes, W. J. (1988). Problems with the use of terminology in genetics education: 1 A literature review and classification scheme. Journal of Biological Education, 22(3), 178–182.
Posner, G. J., Strike, K. A., Hewson, P. W., & Gertzog, W. A. (1982). Accommodation of a scientific conception: Toward a theory of conceptual change. Science Education, 66(2), 211–227.
Stewart, J. H. (1982). Difficulties experienced by high school students when learning basic Mendelian genetics. The American Biology Teacher, 44(2), 80–84. 89.
Treagust, D. F. (1988). Development and use of diagnostic tests to evaluate students’ misconception in science. International Journal of Science Education, 10(2), 159–169.
Tsui, C.-Y. (2009, October). Developing scientific reasoning with multiple external representations: A case study in a Hong Kong secondary school. Paper presented at the International Conference of the East-Asian Science Education (EASE), Taipei, Taiwan.
Tsui, C.-Y., & Treagust, D. F. (2003). Genetics reasoning with multiple external representations. Research in Science Education, 33(1), 111–135.
Tsui, C.-Y., & Treagust, D. F. (2004). Motivational aspects of learning genetics with interactive multimedia. The American Biology Teacher, 66(4), 277–286.
Tsui, C.-Y., & Treagust, D. F. (2007). Understanding genetics: Analysis of secondary students’ conceptual status. Journal of Research in Science Teaching, 44(2), 205–235.
Tsui, C.-Y., & Treagust, D. F. (2010). Evaluating secondary students’ scientific reasoning in genetics using a two-tier diagnostic instrument. International Journal of Science Education, 32(8), 1073–1098.
Tyson, L. M., Venville, G. J., Harrison, A. G., & Treagust, D. F. (1997). A multidimensional framework for interpreting conceptual change events in the classroom. Science Education, 81, 387–404.
van Someren, M. W., Reimann, P., Boshuizen, H. P. A., & de Jong, T. (Eds.). (1998). Learning with multiple representations. London: Pergamon.
Venville, G. J., & Donovan, J. (2005). Searching for clarity to teach the complexity of the gene concept. Teaching Science, 51(3), 20–24.
Venville, G. J., & Treagust, D. F. (1998). Exploring conceptual change in genetics using a multidimensional interpretive framework. Journal of Research in Science Teaching, 35, 1031–1055.
Vygotsky, L. S. (1968). Thought and language. Cambridge, MA: MIT Press.
Vygotsky, L. S. (1978). Mind in society: The development of higher psychological processes. Cambridge, MA: Harvard University Press.
Wood, R. (1996). From my DNA to Darwin: Learning and teaching in genetics. D.Phil thesis, University of Waikato, Hamilton, New Zealand.
Acknowledgments
Our thanks go to the participating teachers and students in Perth and Hong Kong schools. We are grateful to Paul Horwitz of the Concord Consortium of the USA for granting us permission for using BioLogica in our studies and for reviewing an earlier draft of this chapter. We also thank Barbara Buckley for her comments on the final draft of this chapter.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Tsui, CY., Treagust, D.F. (2013). Secondary Students’ Understanding of Genetics Using BioLogica: Two Case Studies. In: Treagust, D., Tsui, CY. (eds) Multiple Representations in Biological Education. Models and Modeling in Science Education, vol 7. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-4192-8_15
Download citation
DOI: https://doi.org/10.1007/978-94-007-4192-8_15
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-4191-1
Online ISBN: 978-94-007-4192-8
eBook Packages: Humanities, Social Sciences and LawEducation (R0)