
Abstract
Agroforestry systems (AFS) are recommended for Europe through the European Rural Development Council regulation 1698/2005, in recognition of their role in reducing carbon (C) emissions and promoting C sequestration which would help to fulfil the Kyoto Protocol requirements. These systems have been found to be a good tool to reduce fire risk and C release in southern European countries. The implementation of AFS could also reduce C release to atmosphere because of the value given to non-timber products, thereby reducing chances for clear cutting of trees. Furthermore, the tree component in AFS will add C into the soil through litterfall and root decomposition, which takes place at deeper soil layers than under agronomic crops or pasture. Tree management practices such as regulating tree density and planting arrangement will influence the C sequestered in the system. Compared with the tree components, the understory components of AFS have less impact on the total C sequestration. The higher inputs of residues generated by the trees in AFS than in tree-less systems may cause high soil C sequestration potential, but soil C increase depends on the incorporation and mineralization of C in the soil, which are affected by understory crop management practices.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
During the last century, the climate in Europe has changed more than in other areas of the world (IPCC 2007). Compared to the pre-industrial era, when the mean annual temperature increased by 0.8°C globally, it increased by 1.2°C in Europe. Based on theoretical models, a further increase of 1.0–5.5°C is expected by the end of the twenty-first century (Christensen et al. 2007). The increase in temperature has been most apparent in hilly areas such as the Alps, which tend to have high biodiversity and where temperature increased by 2°C during the twentieth century (EEA 2009a). This is twice the average temperature increase for the northern hemisphere. In addition, the quantity and distribution of precipitation have also changed in Europe during the twentieth century. Although there has been a 20% decrease in rainfall in southern Europe, there has been a 10–40% increase in rainfall in northern Europe. Furthermore, an increase in the frequency of extreme weather events is predicted across the European continent (EEA 2008).
Climate change may lead to an increase in the incidence of wildfire outbreaks, a decrease in biodiversity, and an increase in carbon dioxide (CO2) emissions. Wildfires are a serious threat to forest ecosystems in Europe (Rigueiro-Rodríguez et al. 2009a), and represent a major source of CO2 emissions. Any increase in temperature will aggravate the danger of forest fires by increasing the incidence of fire events, the area burnt, and the duration of fire seasons, especially in southern and central Europe (EEA 2008). Moreover, climate change in Europe may modify biodiversity through habitat loss and cause changes in dispersal capacity, phenological characteristics, life cycles, and food sources of native species. Climate change may also provoke the decoupling of predator-prey relationships, new invasions, or the spread of already established invasive alien species (EEA 2009b). It would also lead to a decline in soil organic carbon (C) stocks and an increase in CO2 emission from soils. Soils may become more susceptible to erosion, especially in the Mediterranean areas where annual soil losses may reach 200 Mg ha−1 (Correal et al. 2009). Soil degradation is already intense in parts of the Mediterranean and central Eastern Europe and may contribute to desertification (EEA 2008). Agroforestry systems (AFS) offer solutions to some of these climate change related ecosystem management problems. For example, AFS have proved to be an excellent fire prevention technique in many parts of southern Europe such as France (Etienne 1996; Etienne et al. 1996; Rigolot and Etienne 1996), Greece (Papanastasis et al. 2009), and Spain (Robles et al. 2009; Rigueiro-Rodríguez et al. 2009b). Agroforestry practices are considered good land management tools to enhance biodiversity (Rois-Díaz et al. 2006; Rigueiro-Rodríguez et al. 2011b) and augment C sequestration, compared with tree-less systems worldwide (Nair et al. 2008, 2009).
The Kyoto Protocol establishes that land use, land use change and forestry (LULUCF) activities such as afforestation, reforestation, and deforestation (Article 3.3), and forest land management, cropland management, grazing land management, and revegetation (Article 3.4) can be used to meet the greenhouse gas (GHG) emission reduction goal (UN 1998). Burley et al. (2007) indicated that forest offset projects can be based on two approaches, namely, (a) the absorption of GHG by new vegetation (i.e., sink creation and sink enhancement), and (b) displaced emissions by existing vegetation (i.e., fire risk reduction and avoided deforestation). Emission from timber harvesting, which also negatively affects soil organic matter (SOM), could be reduced by the adoption of agroforestry systems that provide benefits other than timber from forest areas. Austria, Belgium, Denmark, Finland, Ireland, Italy, Luxembourg, the Netherlands, Portugal, and Spain plan to fulfil their assigned C emissions by using the Kyoto mechanisms described in Articles 3.3 and 3.4. However, Spain and Italy are among the EU countries with the greatest focus on increasing atmospheric CO2 removals by enhancing C sink activities. Therefore, the implementation of AFS in these two countries, aimed at reducing CO2 in the atmosphere through the two LULUCF activities described by the Kyoto Protocol, should be greater than in other European countries (EEA 2009c).
Considerable efforts in land use change for the reduction of GHG emissions have been carried out in Europe. More than one million hectares of forests were planted between 1994 and 1999 in Europe (Rois-Díaz et al. 2006). According to current targets, it is expected that more than 650,000 ha of agricultural land and about 240,000 ha of non-agricultural land will be afforested in Europe during the period 2007–2013 (EU 2009). This process will involve more than 12,000 landowners. The most recent European Rural Development Report estimates that AFS will cover 60,000 ha of agricultural lands representing 3,000 landowners during the period 2007–2013 (EU 2009) as a result of the council regulation 1698/2005 (EU 2005).
Agroforestry and Carbon Sequestration
The C sequestration potential of AFS is based on live components growing up within the system including the soil, but should also include activities such as forest fire prevention and other multifunctional outputs from the system (Rigueiro-Rodríguez et al. 2009b). The potential of C sequestration in AFS is dependent on the tree component (Nair et al. 2009). Tree presence would increase C sequestration per unit of land due to the C sequestered by the tree itself, the inputs of residues (leaves and branches) it makes on the soil, and the incorporation of roots into the soil. Trees use a greater volume of soil to build up SOM than herbaceous crops, as they are able to explore soils farther from the tree trunk and to a greater depth, assuming small tree density is used (Moreno et al. 2005). The greater soil volume explored by tree roots would enhance belowground organic matter depositions (Howlett et al. 2011). However, understory species may also be positively or negatively affected by the tree presence. The symbiotic or competitive relationship of these components (i.e., tree and understory) depends on specific edapho-climatic conditions (Rigueiro-Rodríguez et al. 2009a; Mosquera-Losada et al. 2010a). Conditions such as adequate water regime, optimal temperatures, and soil nutrient availability would promote tree growth (López-Díaz et al. 2010), but in areas with strong water deficits, usually development of pasture (or other understory species used in the AFS) is reduced due to the presence of trees.
Agroforestry as a land use option has great potential for C sequestration in Europe, as it allows for the sequestration of more C per unit of land, compared with tree-less agronomic systems (Matos et al. 2010a). Agroforestry also results in higher annual economic returns per unit of land through the whole life cycle than in exclusive forestry systems where the revenue is generally only realised at final harvest. These returns could be further increased if appropriate land management practices mainly regulating tree density and distribution are adopted (Sibbald 1996; Fernández-Núñez et al. 2007). The role of AFS in the reduction of C emissions derives from the prevention of forest fires in Mediterranean Europe, as silvopasture agroforestry practices reduce the understory woody biomass (Etienne et al. 1996; Rigueiro-Rodríguez et al. 2009b, 2010). Most AFS have also been shown to reduce soil erosion, and improve nutrient cycling, water availability for crops, soil faunal activities, and soil fertility, while at the same time sustaining high levels of crop production (Grünewald et al. 2007; Quinkenstein et al. 2009; Rigueiro-Rodríguez et al. 2009a).
Some of the most important options to increase C sequestration are those dealing with LULUCF measures. Currently, most European forests are relatively young and they act as a C sink. Growing forests sequester C, but when they reach maturity, the C annually sequestered is reduced. For this reason, forested land conservation that avoids total clear felling should be better at reducing C emissions in the future, not only because of C exported in the harvested trees, but also due to C soil emissions, once the trees are harvested (Nair et al. 2009). However, Dresner et al. (2007) highlighted that if cut timber is worth more than trees still standing in the forest, there is no incentive for farmers to protect the forest. As such, deforestation is likely to occur, regardless of the wider impacts of this such as C emissions. Nonetheless, if the agronomic component of an AFS is valuable for farmers, this would be an additional reason to prevent deforestation and thus reduce CO2 emissions (Dresner et al. 2007).
Several types of agroforestry practices are currently implemented in Europe. Silvoarable and silvopasture agroforestry practices are the most prevalent in Europe in terms of the area under those practices compared with other agroforestry practices (Eichhorn et al. 2006; Mosquera-Losada et al. 2009). They are mostly carried out in Spain and Portugal, but also in Germany, France, Italy and the UK (Dupraz et al. 2005; Grünewald et al. 2007; Mosquera-Losada et al. 2010a; Quinkenstein et al. 2009).
The tree component of an AFS may be more efficient at CO2 utilisation from the atmosphere and may have higher C returns to the soil through their litter than herbaceous crops (Gordon et al. 2006). One year after the implementation of an agroforestry system (Böhm et al. (2010), the content of organic C in soil under tree hedgerows was significantly higher as compared to field alleys (Fig. 1) in Germany, due to the higher root development in the hedgerows compared with the field alleys.

Hot water extractable organic carbon in the surface (0–30 cm) soil, 1 year (HWC 2008) and 2 years (HWC 2009) after establishing an alley cropping system, in a mining reclamation landscape, Lower Lusatia, north-eastern Germany (Source: Adapted from Böhm et al. 2010)
In agroforestry systems, C is located in five main pools, namely, aboveground plant biomass (tree and understory), plant roots (tree and understory), litter, microbial, and soil C. These pools interact with each other via different pathways of transformation and translocation, e.g., plants absorbing CO2 from the atmosphere during photosynthesis. Some CO2 is released back to the atmosphere in the process of plant respiration. Litter falling from plants and dead roots from plant material are decomposed into soil C. Some of the soil C is taken up by microbes and stored, and some becomes mineralised. Soil stores C, but as a result of the mineralisation and root respiration, part of that C is released back to the atmosphere. Furthermore, biodiversity enhancement by AFS facilitate a better nutrient use and therefore increases C sequestration compared with tree-less agronomic systems (Howlett et al. 2011; Rigueiro-Rodríguez et al. 2011b).
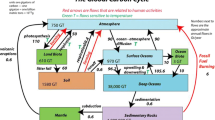
Measurement of C sequestration following land use changes from tree-less agriculture to forestland requires the evaluation of the baseline C stocks as well as the nature of the tree component and the modifications the tree causes to the understory and in the soil compartment. If silvopasture agroforestry is carried out, then the animal component and the emission of methane and nitrous oxide gases should also be taken into account (IPCC 2007). The main components and their GHG balance in a silvopasture agroforestry system including grazing animals are presented in Fig. 2.

Carbon pools in a silvopasture system including GHG emissions: (a) A schematic diagram showing the different compartments. (b) An example of the estimated quantities in each compartments in a 11 years-old Pinus radiata D. Don stand in Galicia, NW Spain (Source: Adapted from Fernández-Núñez et al. 2010)
Tree Component
Land use change through afforestation or reforestation should increase C sequestration per unit of land and the rate of C sequestered by trees within a system will depend on tree species, age, and density (Quinkenstein et al. 2009), besides the edapho-climatic conditions, management, fertilization, and land clearing, among others. Carbon sequestration by an individual tree can be estimated by allometric equations based on the tree diameter that have been recently developed in Spain (Montero et al. 2005) and Europe (Zianis et al. 2005). In their studies, 13 and 24 conifers and 15 and 31 broadleaf trees species were used to estimate the C sequestered in aboveground biomass and in roots, respectively. This was carried out for species from the Mediterranean, mountainous, and Atlantic biogeographic regions of Europe. However, most of the trees used to develop the equations were in mature, dense stands, and therefore, more research is needed to understand how C is sequestered in younger stands (Knopka et al. 2010) and growing at lower densities such as in AFS.
The growth rate of tree species is a significant factor in promoting C sequestration. Annual estimates of C sequestered by tree biomass of Eucalyptus globulus Labill, Pinus pinaster Ait., Pinus radiata D. Don and Castanea sativa Mill. in Spain were 5.14, 1.58, 1.11, and 0.52 Mg C ha−1, respectively (Pardos 2010). Differences in growth rates explain why, after 10 years, P. radiata, a species with a high growth rate, sequestered eight times more C per tree than Betula alba L. at densities of 833 and 2,500 trees ha−1 in Spain (Fig. 3). Similarly, species like poplar or eucalyptus were able to sequester C faster than species such as P. radiata, B. alba, P. pinaster. However, these three species grew and sequestered C faster than the other common silvopastoral tree species such as Pinus sylvestris L., Quercus petraea L., Quercus robur L., or Fagus sylvatica L. (Pardos 2010). Gordon et al. (2006) highlighted the importance of using fast growing tree species in silvopastoral systems to reduce C emissions in Canada. They estimated that net C sequestration of a poplar-based silvopastoral system was almost three times more than that reached by a monoculture pasture system. However, if trees grow quickly, C sequestered for a given period of time is reduced as trees will be harvested earlier (Fernández-Núñez et al. 2010). The time required for C sequestration to occur is longer for slow growing species than for fast growing species. Therefore, once harvested, the fast growing species emit C into the atmosphere earlier than slow growing species, mainly from SOM mineralization. Moreover, sawn timber production is usually associated with slow growing species, which are retained for longer time than pulp and paper from fast growing species such as Eucalyptus spp.

Carbon stock in the tree stand and individual tree of Pinus radiata D. Don and Betula alba L. Eleven years after establishment at two densities (833 and 2,500 trees/ha−1) in Galicia, NW Spain
The production of Robinia pseudoacacia L. in an alley cropping system has received considerable interest in Germany as an alternative to agricultural crops as well as an additional wood source, while simultaneously acting as a potential C sink to counterbalance greenhouse gases emissions. Average aboveground biomass production of R. pseudoacacia ranged from 0.04 to 9.5 Mg ha−1 year−1 for 1–14 years of growth, respectively on reclaimed sites in north-eastern Germany (Quinkenstein et al. 2011).
The C sequestration of afforested or reforested lands also depends on land management and soil type. Fertilization carried out to enhance crop production in AFS indirectly increases tree growth in some edapho-climatic conditions (Dupraz et al. 2005). In acidic soils of Galicia, Spain (water pH = 4.5), the C sequestered by P. radiata (1,667 trees ha−1) 11 years after afforestation was 4.09 Mg C ha−1 when no fertilizer was applied. The amount of C sequestered by the tree component significantly increased to 7 Mg C ha−1 when sewage sludge was used as fertilizer in the same soil. However, these values were lower than those reported for agricultural lands (initial soil water pH = 6.9), which were afforested at high density (2,500 trees ha−1) (Fig. 3; Fernández-Núñez et al. 2010). Soil fertility improvements usually increase growth rates and symbiosis. However, facilitation between the tree and the understory should be promoted in the early tree ages in order to enhance resource use and increase C sequestration (Mosquera-Losada et al. 2006, 2011b). The use of legumes such as clover (Trifolium spp.) in the sown mixture, increased tree growth and was found to promote symbiosis between P. radiata and understory (López-Díaz et al. 2010). However, the increase in ryegrass (Lolium perenne L.) density during the year of plantation establishment reduced P. radiata growth due to competition between the tree and ryegrass for soil resources (Mosquera-Losada et al. 2011b).
Tree density is another factor that affects C sequestration. Fernández-Núñez et al. (2010) reported from Galicia, Spain, that land that had previously been under agriculture when afforested with P. radiata at 833 or 2,500 trees ha−1 was able to sequester 40.8 and 102.4 Mg C ha−1 11 years after plantation in tree roots and aboveground biomass, respectively, despite the fact that C sequestered per tree was higher at a low density (48 and 40 kg C tree−1, respectively: Fig. 3). Similar results were also found for B. alba planted at these densities in the same area.
In the Atlantic biogeographic region of Europe, tree stands were established at higher tree densities than in the Mediterranean dehesa area to promote timber production (Serrada et al. 2008). Due to the intraspecific competition in the high density stands, tree roots may not spread far away from the tree trunks compared with low density stands. There have been few published studies where the differences in root system profiles with respect to the distance from the tree for low versus high density stands have been measured. In the Mediterranean environments, Moreno et al. (2005) reported that most fine roots of Quercus ilex L. trees were below 80 cm depth, while herbaceous plant roots were mainly located in the top 30 cm soil layer. Drought conditions could have a great effect on tree root distribution within the soil profiles of Mediterranean systems. The same effect could be simulated by competition within the herbaceous layer in more northern European countries if AFS with low tree densities were implemented. If tree roots are located below the herbaceous understory rhizosphere, then competition for soil resources between trees and herbaceous plants is reduced. Implementation of agroforestry could increase the volume of soil explored by roots (the upper part of the soil explored by the herbaceous component and the lower part by the tree component) in low density stands compared with tree-less pastures. The amount of fine roots that are considered to be the main source of organic matter within a soil C pool (Dresner et al. 2007) would also increase.
Tree C sequestration also depends on the species. Evergreen trees retain C in the leaves for longer period of time than deciduous tree species, which cause regular inputs of organic matter into the soil, apart from the roots. Evergreen tree litterfall is usually low until canopy closure. Afterwards, the relatively low understory light levels may cause an accumulation of litter on the forest floor. Density affects the dynamics of the tree litter inputs into the soil. A dense P. radiata canopy caused an accumulation of a thick litter layer of several centimetres above the soil a few years after canopy closure, which prevented herbaceous plant establishment and reduced biodiversity and soil C sequestration potential. Litter biomass also depends on tree density, which was higher in high density forests (6.25 Mg ha−1 at 2,500 trees ha−1) than in low density stands (4.26 Mg ha−1 at 833 trees ha−1) in a P. radiata-afforested land 11 years after planting (Fernández-Núñez et al. 2010). The thick litter layer could emit large quantities of C once the forest stand is harvested. However, no accumulation of litterfall on the soil was observed in a silvopastoral system established with B. alba at 2,500 trees ha−1 or 833 trees ha−1 due to the low growth rate of birch as compared with radiata pine. Higher soil temperatures in birch stand, comparable with P. radiata, increased birch litter decomposition, promoting soil C sequestration (Howlett et al. 2011).
Understory Component
Compared to the tree and the soil C pools, the amount of C sequestered in the understory component of the AFS is relatively small (Fernández-Núñez et al. 2010). In European AFS, the understory component may be a crop (e.g., a cereal or leguminous crop) in the silvoarable systems or herbaceous or woody plants in the silvopastoral systems. Arable systems have lower C sequestration potential than herbaceous pasture or understory woody plants and involve annual crops that are usually harvested within a year of sowing, and the biomass is exported from the system. Crop management practices such as plowing, liming, and fertilization may cause either soil C increase or losses. Improvement of soil fertility increases the growth of AFS components and therefore soil inputs of C. However, management activities may also result in better aeration, increased pH, and enhanced soil fertility, promoting microbial activity and organic matter mineralisation, in turn, leading to lower SOM levels (Reijneveld et al. 2010).
Perennial grasslands and shrublands may store C within their tissues for a longer period of time than arable crops. The large area of the European Union allocated to grasslands in different biogeographic regions (33% and 25% of the Atlantic and continental biogeographic regions of Europe; EEA 2006) offers a high potential for C sequestration. This potential, however, is dependent on the edapho-climatic conditions and land management practices adopted (Follet et al. 2001; Schanabel et al. 2001). The input of organic matter to grassland soils is very important (Sanderson and Wätzold 2010) and would increase the SOM content. Mature pasturelands, however, show no net annual C uptake when all sources and sinks are considered (Suyker and Verma 2001; Gianelle et al. 2004). Follet et al. (2001) concluded that improved grassland management could enable C sequestration to continue for 25–50 years until a new equilibrium of soil C content is reached. After that, the improved grasslands would no longer serve as C sinks. Even though grasslands may sequester C, grazing by livestock animals may result in CH4 or N2O emissions (IPCC 2007). When the animal stocking rate is adjusted to the production of grasslands, the C losses with GHGs are offset by the C sequestered (Fernández-Núñez et al. 2010).
Understory shrubs sequester more C than herbaceous plants. However, the risk of C emissions caused by fires associated with forestlands is increased by these shrubs, making the presence of woody vegetation understory very hazardous in the Mediterranean countries of Europe and in the southern Atlantic biogeographic region of Europe, where summers are too dry and fire risk is high (Rigueiro-Rodríguez et al. 2009b). Prevention of forest fires mitigates C emissions (Burley et al. 2007). Agroforestry practices could be successfully implemented to reduce the emissions of C caused by fires. For instance, shrub grazing by goats in silvopastoral systems reduces the amount of combustible vegetation in the understory and encourages a less flammable herbaceous layer (Rigueiro-Rodríguez et al. 2011a). Understory vegetation transformation from shrubs to a grass is thus promoted by grazing of shrubs by animals as well as by soil nutrient cycling through animal faeces and urine deposition (Rigueiro-Rodríguez et al. 2009b).
Soil Component
The soil represents the most important pool of C storage in terrestrial ecosystems, accounting for about 75% of total stored C (Lal 2005; Dresner et al. 2007). Soil C sequestration depends on edapho-climatic conditions, which may increase or reduce the organic matter inputs (i.e., the quantity of plant residues), incorporation of organic matter into the soil, and organic matter mineralisation (Nieder et al. 2003). Soil properties such as clay content determine the extent of C enrichment in humus. Organic matter inputs usually create a C gradient from the surface to the lower layers of the soil worldwide (Fig. 4; Howlett et al. 2011).

Total soil carbon concentration by depth in an afforested and non-fertilized soil in Galicia, NW Spain
Temperature and humidity are the main drivers of SOM production, incorporation, and mineralisation (Theng et al. 1989). If temperature and humidity are optimal for aboveground biomass production as in the Atlantic climate, the inputs of organic matter into the soil are greater than that in less favourable climatic conditions such as in the Mediterranean climate. For this reason, the higher potential productivity of crops in the Spanish Atlantic region is an important indicator of higher SOM as compared to the Spanish Mediterranean region (i.e., 3 and 0.4 Mg ha−1 year−1 for Atlantic and Mediterranean regions, respectively: Pardos 2010). In the dehesas located in the Mediterranean area, the presence of mature trees and, therefore, the rate of incorporation of their residues are associated with higher SOM levels below the tree than away from the tree in AFS established under trees without canopy closure (Moreno and Obrador 2007). In some cases, the degree of incorporation of plant residues into the soil may be restricted by high humidity and low temperature. In P. radiata stands, the closure of tree canopies caused an annual accumulation of about 7 Mg ha−1 of litterfall in Galicia (Fernández-Núñez et al. 2010).
Roots are also an important part of the C balance in terrestrial ecosystems because they transfer large amounts of C into the soil. More than half of the C assimilated by the plant is transported belowground via root growth and turnover, root exudates (of organic substances) and litter deposition, and roots may contribute up to 33% to C sequestered in ecosystems (Fernández-Núñez et al. 2010). The dynamics of growth, decay, and root turnover are some of the least understood aspects of belowground interactions in agroforestry (Nair et al. 1999). There is much information on C sequestration in the topsoil layer of 0–20 cm. However, information on deeper soil layers, where most of the tree roots occur, is lacking in most environments, but some studies have been carried out in the Spanish dehesa agrosilvopastoral system (Moreno and Obrador 2007). Roots of trees and grass or crops have different root length and depth profiles. Tree roots are longer and deeper in soil than grass or crop roots, and in soils under trees, a considerable amount of C is stored below the plow layer (50 cm). This C is also better protected from disturbance, which leads to longer residence time in the soil. Most of the root biomass of annual crops and grasses consists of fine roots (diameter < 2 mm). Fine roots of both trees and crops have a relatively fast turnover (measured in days to weeks), but lignified coarse roots of trees decompose much more slowly once trees are harvested and may contribute substantially to belowground C pools (Vanlauwe et al. 1996).
Carbon inputs to the soil are also affected by litterfall. Higher biomass production per tree and per hectare obtained in previously agricultural lands afforested with P. radiata and B. alba at high stem density increased soil C more than low tree density 5 years after the establishment of both trees (Fernández-Núñez et al. 2010). However, differences in SOM between density treatments or species disappeared 10 years after afforestation, probably due to the lack of litterfall incorporation under high density stands. This can be explained by the low temperatures and high humidity experiences differentially by both systems. Incorporation of residues into the soil is the first step to increasing SOM.
Alley cropping systems have also come into focus in the reclamation of post-mining areas where the initial content of SOM is generally close to zero and soil fertility is very low (Nii-Annang et al. 2009). The increase in SOM in reclaimed areas depends on the amount of biomass production and return to soil as well as mechanisms for C protection and retention. Due to its high potential for litterfall production and nitrogen fixation, R. pseudoacacia improves soil physical, chemical, and biological properties by increasing SOM, thereby converting mine spoils into productive and sustainable soils (Grünewald et al. 2007).
High amounts of litterfall increase fire risk in European Mediterranean areas (Delabraze 1986) and, therefore, the risk of C emissions to the atmosphere. Implementation of silvopasture has been shown to reduce fire risk through the enhancement of litter incorporation into the soil as nitrogen is added with the urine of the animals and C/N relationship is reduced (Etienne et al. 1996; Rigolot and Etienne 1996).
It is well known that soil management activities such as plowing or fertilization may reduce or increase SOM content. Matos et al. (2010a, b) investigated the effect of conversion from silvopasture to arable land and reported lower contents of total organic carbon (TOC) and total nitrogen (TN) in arable soils than silvopasture. The composition and distribution of SOM also differed between these two systems. The light fraction C content declined with depth in silvopasture system, while there were no such depth-related differences in arable system. This can be attributed to tillage in arable systems, which leads to the disturbance of upper soil layers causing an increase in mineralization rates, CO2 emissions from soils, and the reduction of soil C. Soil management through fertilization also affects soil C storage. Mosquera-Losada et al. (2010b) reported that the addition of sewage sludge (pH around 7) in acidic soils (water pH = 4.5) increased SOM content through the input of organic matter as well as calcium via the sewage sludge (Fig. 5). The SOM content was not modified when mineral nitrogen was added, as incorporation of organic residues through the improvement of soil pH was not promoted. In a Populus canadensis Moench silvopastoral system developed on a Galician (Spain) acid soils with pH around 5.5, the SOM content in winter was related to pasture production in the preceding autumn (r2 = 0.93; % SOM = 0.48 autumn production [Mg ha−1] +8.87 p < 0.05), which suggested that organic matter was incorporated but not mineralized (Mosquera-Losada et al. 2011a). Therefore, SOM seems to have increased when herbaceous autumn production was high, causing an increase in organic matter inputs into the soil.

Soil organic matter (SOM) and pH (water) under different fertilization treatments of Pinus radiata D. Don in Galicia. NF: no fertilization; N160, N320 and N480 refer to160, 320, and 480 kg N ha−1, respectively; MIN: inputs of a mineral compound fertilizer 500 kg of 8N: 24P2O5: 16 K2O. The letters on graph indicate significance of differences between treatments at p = 0.05 according to Duncan´s multiple range test (Source: López-Díaz et al. 2007; Rigueiro-Rodríguez et al. 2011a)
It has been proposed that C stored in the soil could be linked to different soil-size fractions (Lal 2005). However, there have been only very few studies evaluating C storage in different soil-size fractions in treeless versus AFS. Carbon associated with macroaggregates (250–2,000 μm), microaggregates (53–250 μm) and silt clay (<53 μm) can have mean residence time of 1–10, 1–25, and 100–1,000 years, respectively (Parton et al. 1987; Schimel et al. 1994). One study carried out in Galicia, Spain, showed that the broadleaf B. alba sequestered more C in the 250–2,000 μm size class as compared to soils under the conifer P. radiata. However, pastures had more C than pine silvopasture in soils with finer particle sizes fractions of less than 250 μm (Howlett et al. 2011).
Conclusion
Agroforestry systems have great potential to enhance C sequestration compared with tree-less agronomic systems, and therefore their implementation should be considered as a land use option in Europe. The limited number of studies undertaken so far at various locations and systems in Europe have shown that the factors that contribute to higher C sequestration under AFS include greater above-and below-ground spatial heterogeneity in the vegetation (trees and crops), production of higher amounts of plant biomass, more extensive root exploration of rhizosphere and increased litterfall inputs to the soil. Further studies are needed on all these as well as other aspects of the soil and associated vegetation to evaluate different components of agroforestry systems, including trees, the understory, animals and their interactions, under specific edapho-climatic conditions. The implementation of AFS contributes to an overall sustainable land management based on the increase of soil fertility by C enrichment in humus and the potential of C sequestration in the soil–plant system.
References
Böhm C, Quinkenstein A, Freese D, Hüttl RF (2010) Energieholzproduktion in Agroforstsystemen als ein Beitrag zur Verbesserung des Bodenschutzes. In: Tagungsband zu 6. Marktredwitzer Bodenschutztage “Bodenschutz in Europa – Ziele und Umsetzung”, S 78–84
Burley J, Ebeling J, Costa PM (2007) C sequestration as a forestry opportunity in a changing climate. In: Freer-Smith PH, Boradmeadow MSJ, Lynch JM (eds.) Forestry and climate change. CAB International, Wallingford, UK, pp 31–37
Christensen JH, Hewitson B, Busuioc A, Chen A, Gao X, Held I, Jones R, Kolli RK, Kwon WT, Laprise R, Magaña Rueda V, Mearns L, Menéndez CJ, Räisänen J, Rinke A, Sarr A, Whetton P (2007) Regional climate projections. In: Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M, Miller HL (eds.) Climate change 2007: the physical science basis contribution of working group I to the fourth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK, pp 849–940
Correal E, Erena M, Ríos S, Robledo A, Vicente M (2009) Agroforestry systems in southeastern Spain. In: Rigueiro-Rodríguez A, McAdam J, Mosquera-Losada MR (eds.) Agroforestry in Europe: current status and future prospects. Springer, Dordrecht, pp 183–210
Delabraze P (1986) Sylviculture méditeranéenne. In: Lanier L (ed.) Précis de Sylviculture. ENGREF, Nancy, pp 362–376
Dresner S, Ekins P, McGeevor K, Tomei J (2007) Forest and climate change global understandings and possible responses. In: Freer-Smith PH, Boradmeadow MSJ, Lynch JM (eds.) Forestry and climate change. CAB International, Wallingford, UK, pp 38–48
Dupraz CC, Burgess PJ, Gavaland A, Graves AR, Herzog F, Incoll LD, Jacson N, Keesman K, Lawson G, Lecomte I, Mantzanas K, Mayus M, Palma J, Papanastasis V, Paris P, Pilbeam DJ, Reisner Y, van Noordwijk M, Vincent G,van der Werf W (2005) SAFE (Silvoarable Agroforestry for Europe ) synthesis report. SAFE project (August 2001–January 2005). Available at http://www1.montpellier.inra.fr/safe/english/results/final-report/SAFE%20Final%20Synthesis%20Report.pdf. Accessed 9 Dec 2010
EEA (2006) Land accounts for Europe 1990–2000. Towards integrated land and ecosystem accounting. Available at http://www.eea.europa.eu/publications/eea_report_2006_11. Accessed 9 Dec 2010
EEA (2008) Impacts of Europe´s changing climate. EEA briefing 2008–03. Available at http://www.eea.europa.eu/publications/briefing_2008_3. Accessed 12 Jan 2011
EEA (2009a) Regional climate change and adaptation. The Alps facing the challenge of changing water resources. Available at http://www.eea.europa.eu/publications/alps-climate-change-and-adaptation-2009. Accessed 12 Jan 2011
EEA (2009b) Progress towards the European 2010 biodiversity target. EEA report no 4/2009. Available at http://www.eea.europa.eu/publications/progress-towards-the-european-2010-biodiversity-target. Accessed 12 Jan 2011.
EEA (2009c) Greenhouse gas emission trends and projections in Europe 2009. Tracking progress towards Kyoto targets. Available at http://www.eea.europa.eu/publications/eea_report_2009_9. Accessed 12 Jan 2011
Eichhorn MP, Paris P, Herzog F, Incoll LD, Liagre F, Mantzanas K, Mayus M, Moreno G, Papanastasis VP, Pilbeam DJ, Pinsanelli A, Dupraz C (2006) Silvoarable systems in Europe - past, present and future prospects. Agroforest Syst 67:29–50
Etienne M (1996) Western silvopastoral systems. INRA, Paris, 276 p
Etienne M, Drezko M, Rigolot E (1996) Browse impact in silvopastoral systems participating in fire prevention in the French Mediterranean region. In: Étienne M (ed.) Western European silvopastoral systems. INRA, Paris, pp 93–102
EU (2005) Council Regulation (EC) No 1698/2005 of 20 September 2005 on support for rural development by the European Agricultural Fund for Rural Development (EAFRD). Available at http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2005:277:0001:0040:EN:PDF. Accessed 12 Jan 2011
EU (2009) Report on implementation on forestry measures under the rural development regulation 1698/2005 for the period 2007–2013. Directorate-general for agriculture and rural development. Available at http://ec.europa.eu/agriculture/fore/publi/report_exsum_en.pdf. Accessed 12 Jan 2011
Fernández-Núñez E, Mosquera-Losada MR, Rigueiro-Rodríguez A (2007) Economic evaluation of different land use alternatives: forest, grassland and silvopastoral systems. Grassland Sci Eur 12:508–511
Fernández-Núñez E, Rigueiro-Rodríguez A, Mosquera-Losada MR (2010) C allocation dynamics one decade after afforestation with Pinus radiata D. Don and Betula alba L. under two stand densities in NW Spain. Ecol Eng 36:876–890
Follet RF, Kimble JM, Lal R (2001) The potential of US grazing lands to sequester carbon and mitigate the greenhouse effect. CRC Lewis Publishers, Boca Raton, 442 p
Gianelle D, Zorer R, Marcolla B, Cescattia A, Vescovo L (2004) Carbon flux measurements in a mountain grassland in the Italian Alps. Grassland Sci Eur 9:157–159
Gordon AM, Thevathasan NV (2006) How much carbon can be stored in Canadian agroecosystems using a silvopastoral approach? In: Mosquera-Losada MR, McAdam J, Rigueiro-Rodríguez A (eds.) Silvopastoralism and sustainable land management. CABI Publishing, Wallingford, UK, pp 210–219
Grünewald H, Brandt BKV, Schneider BU, Bens O, Kendzia G, Hüttl RF (2007) Agroforestry systems for the production of woody biomass for energy transformation purposes. Ecol Eng 29:319–328
Howlett DS, Mosquera-Losada MR, Nair PKR, Nair VD, Rigueiro-Rodríguez A (2011) Soil C storage in silvopastoral systems and a treeless pasture in northwestern Spain. J Environ Qual 40:1–8
IPCC (2007) Climate change 2007 — synthesis report. Contribution of working groups I, II and III to the fourth assessment report of the intergovernmental panel on climate change. Available at: http://www.ipcc.ch/publications_and_data/publications_ipcc_fourth_assessment_report_synthesis_report.htm. Accessed Jan 2011
Knopka B, Patik J, Moravcik M, Lukac M (2010) Biomass partitioning and growth efficiency in four naturally regenerated forest tree species. Basic Appl Ecol 11(3):234–243
Lal R (2005) Forest soils and C sequestration. For Ecol Manag 220:242–258
López-Díaz ML, Mosquera-Losada MR, Rigueiro-Rodríguez A (2007) Lime, sewage sludge and mineral fertilization in a silvopastoral system developed in very acid soils. Agroforest Syst 70:91–101
López-Díaz ML, Rigueiro-Rodríguez A, Mosquera-Losada MR (2010) Influence of pasture botanical composition and fertilization treatments on tree growth. For Ecol Manag 257:1363–1372
Matos ES, Freese D, Mendonca ES, Slazak A, Huttl RF (2010a) C, nitrogen and organic C fractions in topsoil affected by conversion from silvopastoral to different land use systems. Agroforest Syst. doi:10.1007/s10457-010-9314-y
Matos ES, Freese D, Ślązak A, Bachmann U, Veste M, Hüttl RF (2010b) Organic-C and nitrogen stocks and organic-C fractions in soil under mixed pine and oak forest stands of different ages in NE Germany. J Plant Nutr Soil Sci 173:654–661
Montero G, Ruiz-Peinado R, Muñoz M (2005) Producción de biomasa y fijación de CO2 por los bosques españoles, vol 13, INIA: Serie forestal. INIA, Madrid, 270 p
Moreno G, Obrador JJ (2007) Effects of trees and understory management on soil fertility and nutritional status of holm oaks in Spanish dehesas. Nutr Cycle Agroecosyst 78:253–264
Moreno G, Obrador JJ, Cubera E, Dupraz C (2005) Fine root distribution in dehesas of central-western Spain. Plant Soil 277:153–162
Mosquera-Losada MR, Fernández-Núñez E, Rigueiro-Rodríguez A (2006) Pasture, tree and soil evolution in silvopastoral systems of Atlantic Europe. For Ecol Manag 232:135–145
Mosquera-Losada MR, McAdam J, Romero-Franco R, Santiago-Freijanes JJ, Rigueiro-Rodríguez A (2009) Definitions and components of agroforestry practices in Europe. In: Rigueiro-Rodríguez A, McAdam J, Mosquera-Losada MR (eds.) Agroforestry in Europe. Current status and future prospects. Springer, Dordrecht, pp 3–20
Mosquera-Losada MR, Ferreiro-Domínguez N, Rigueiro-Rodríguez A (2010a) Fertilization in pastoral and Pinus radiata D. Don silvopastoral systems developed in forest and agronomic soils of Northwest Spain. Agric Ecosyst Environ 139(4):618–628
Mosquera-Losada MR, Muñoz-Ferreiro N, Rigueiro-Rodríguez A (2010b) Agronomic characterisation of different types of sewage sludge: policy implications. Waste Manage 30(3):492–503
Mosquera-Losada MR, Morán-Zuloaga D, Rigueiro-Rodríguez A (2011a) Effects of lime and sewage sludge on soil, pasture production and tree growth in a six year old Populus canadensis Moench silvopastoral system. J Plant Nutr Soil Sci 174:145–153
Mosquera-Losada MR, Cuiña-Cotarelo R, Rigueiro-Rodríguez A (2011b) Effect of understory vegetation management through liming and sewage sludge fertilization on soil fertility and Pinus radiata D. Don growth after reforestation. Eur J For Res. doi:10.1007/s10342-011-0489-1
Nair PKR, Buresh RJ, Mugendi DN, Latt CR (1999) Nutrient cycling in tropical agroforestry systems: myths and science. In: Buck LE, Lassoie JP, Fernandes ECM (eds.) Agroforestry in sustainable agricultural systems. CRC Press, Boca Raton, pp 1–31
Nair PKR, Gordon A, Mosquera-Losada MR (2008) Agroforestry. Encyclopaedia Ecol 1:101–110
Nair PKR, Kumar BM, Nair VD (2009) Agroforestry as a strategy for C sequestration. J Plant Nutr Soil Sci 172:10–23
Nieder R, Benbi DK, Isermann K (2003) Soil organic matter dynamics. In: Benbi DK, Nieder R (eds.) Handbook of processes and modelling in the soil-plant system. Haworth Press, Binghamton, pp 346–408
Nii-Annang S, Grünewald H, Freese D, Hüttl R, Dilly O (2009) Microbial activity, organic C accumulation and 13C abundance in soils under alley cropping systems after 9 years of recultivation of quaternary deposits. Biol Fert Soils 45(5):531–538
Papanastasis VP, Mantzanas K, Dini-Papanastasi O, Ispikoudis I (2009) Traditional agroforestry systems and their evolution in Greece. In: Rigueiro-Rodríguez A, McAdam J, Mosquera-Losada MR (eds.) Agroforestry in Europe. Current status and future prospects. Springer, Dordrecht, pp 89–110
Pardos JA (2010) Los ecosistemas forestales y el secuestro de C ante el calentamiento global. INIA, Madrid, 253 p
Parton WJ, Schimel DS, Cole CV, Ojima DS (1987) Analysis of factors controlling soil organic matter levels in Great Plains grasslands. Soil Sci Soc Am J 51:1173–1179
Quinkenstein A, Wöllecke J, Böhm C, Grünewald H, Freese D, Schneider BU, Hüttl RF (2009) Ecological benefits of the alley cropping agroforestry system in sensitive regions of Europe. Environ Sci Policy 12(8):1112–1121
Quinkenstein A, Böhm C, Matos E, Freese D, Hütt RF (2011) Assessing the carbon sequestration in short rotation coppices of Robinia pseudoacacia L. on marginal sites in northeast Germany. In: Kumar BM, Nair PKR (eds.) Carbon sequestration potential of agroforestry systems: opportunities and challenges. Springer, Dordrecht, pp 201–216
Reijneveld JA, Kuikman PJ, Oenema O (2010) Changes in soil organic matter content of grassland and maize land in the Netherlands between 1970 and 2009. Grassland Sci Eur 15:3–32
Rigolot E, Etienne M (1996) Litter thickness on tree covered fuel-breaks maintained by grazing. In: Étienne M (ed.) Western European silvopastoral systems. INRA, Paris, pp 111–124
Rigueiro-Rodríguez A, Fernández-Núñez E, González-Hernández P, McAdam JH, Mosquera-Losada MR (2009a) Agroforestry systems in Europe: productive, ecologial and social perspectives. In: Rigueiro-Rodríguez A, McAdam J, Mosquera-Losada MR (eds.) Agroforestry in Europe. Current status and future prospects. Springer, Dordrecht, pp 43–66
Rigueiro-Rodríguez A, López-Díaz ML, Mosquera-Losada MR (2011a) Organic matter and chromium evolution in herbage and soil in a Pinus radiata silvopastoral system in North-West Spain after sewage sludge and lime application. Commn Soil Sci Plant 12:1–14
Rigueiro-Rodríguez A, Santiago-Freijanes JJ, Fernández-Núñez E, Mosquera-Losada MR (2009b) Sistemas biológicos de prevención de incendios forestales. Los sistemas agroforestales. In: Vélez R (ed.) La defensa contra incendios forestales. Fundamentos y experiencias, 2nd edn. McGraw-Hill/Intramericana, Madrid, pp 447–451
Rigueiro-Rodríguez A, Rois-Díaz M, Mosquera-Losada MR (2010) Integrating silvopastoralism and biodiversity conservation. Sustainable Agric Rev 5:359–373
Rigueiro-Rodríguez A, Mosquera-Losada MR, Fernández-Núñez E (2011b) Afforestation of agricultural land with Pinus radiata D. Don and Betula alba L. in NW Spain: effects on soil pH, understorey production and floristic diversity eleven years after establishment. Land Degrad Dev. DOI:10.1002/ldr.1072
Robles AB, Ruíz-Mirazo J, Ramos ME, González-Rebollar JL (2009) Role of livestock grazing in sustainable use, naturalness promotion in naturalization of marginal ecosystems of southeastern Spain (Andalusia). In: Rigueiro-Rodríguez A, McAdam J, Mosquera-Losada MR (eds.) Agroforestry in Europe. Current status and future prospects. Springer, Dordrecht, pp 211–232
Rois-Díaz M, Mosquera-Losada MR, Rigueiro-Rodríguez A (2006) Biodiversity indicators on silvopastoralism across Europe. Available at http://www.efi.int/files/attachments/publications/tr_21.pdf. Accessed 12 Jan 2011
Sanderson MA, Wätzold F (2010) Balancing trade-offs in ecosystem functions and services in grassland management. Grassland Sci Eur 15:639–648
Schanabel TT, Franzluebbers AJ, Stout WL, Sanderson MA, Stuedemann JA (2001) Pasture management effects on soil carbon sequestration. In: Follet RF, Kimble JM, Lal R (eds.) Potential of US grazing lands to sequester carbon. CRC Lewis Publishers, Boca Raton, pp 291–322
Schimel DS, Braswell BH, Holland EA, Mckeown R, Ojima DS, Painter TH, Parton WJ, Townsend AR (1994) Climatic, edaphic, and biotic controls over storage and turnover of C in soils. Glob Biogeochem Cy 8(3):279–293
Serrada R, Montero G, Reque JA (2008) Compendio de Aselvicultura Aplicada en España. INIA, Madrid, 1178 p
Sibbald A (1996) Silvopastoral systems on temperate sown pastures a personal perspective. In: Etienne M (ed.) Western European silvopastoral systems. INRA, Paris, pp 23–36
Suyker AE, Verma SB (2001) Year-round observations of the net ecosystem exchange of carbon dioxide in a native tallgrass prairie. Glob Change Biol 7:279–289
Theng BKG, Tate KR, Sollins P, Moris N, Madkarni N, Tate R (1989) Constituents of organic matter in temperate and tropical soils. In: Coleman DC, Oades JM, Uehara G (eds.) Dynamics of soil organic matter in tropical ecosystems. College of Tropical Agriculture and Human Resources, University of Hawaii, Honolulu, pp 5–32
UN (1998) Kyoto protocol to the United Nations framework convention on climate change. Available at http://unfccc.int/resource/docs/convkp/kpeng.pdf. Accessed 12 Jan 2011
Vanlauwe B, Nwoke OC, Sanginga N, Merckx R (1996) Impact of the residue quality on the carbon and nitrogen mineralization of leaf and root residues of three agroforestry species. Plant Soil 183:221–231
Zianis D, Muukkonen P, Mäkipää R, Mencuccini M (2005) Biomass and stem volume equations for tree species in Europe. Silva fennica monographs 4. The Finnish society of forest science. Tammer-Paino Oy, Tampere, 63 p
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Mosquera-Losada, M.R., Freese, D., Rigueiro-Rodríguez, A. (2011). Carbon Sequestration in European Agroforestry Systems. In: Kumar, B., Nair, P. (eds) Carbon Sequestration Potential of Agroforestry Systems. Advances in Agroforestry, vol 8. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-1630-8_3
Download citation
DOI: https://doi.org/10.1007/978-94-007-1630-8_3
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-1629-2
Online ISBN: 978-94-007-1630-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)