
Abstract
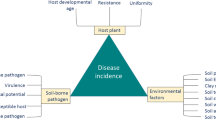
Soilborne pathogens are difficult to manage, especially since the use of methyl bromide has been phased out in most countries. Resistance against many soilborne pathogens is hardly available and fungicides are effective only to a limited extent. In organic agriculture, many problems related to soilborne pathogens are avoided by applying wide rotations, but still some polyphagous soilborne pathogens can be highly problematic, especially since most chemical crop protectants are not allowed. In addition, wide rotations are often economically unprofitable. Therefore, alternative practices to manage soilborne pathogens are needed. In this review, the occurrence of soilborne pathogens in three types of cropping systems are evaluated: (i) continuous cultivation of single crops in monoculture, (ii) crop rotation, and (iii) mixed cropping, i.e., cultivation of multiple crops in the same field at the same time. Both continuous cropping and crop rotation have been investigated extensively. Therefore, in this chapter we focus on mixed-cropping systems in relation to soilborne pathogens, their potential to suppress soilborne diseases, and the mechanisms underlying disease suppression. In general, mixed cropping is practiced to optimize nutrient uptake, control soil erosion, suppress the epidemic spread of airborne pathogens, and improve crop yields per unit of area. While mixed cropping has received attention for its effects on airborne pests and pathogens, the effects on soilborne pathogens are poorly known. In 30 out of 36 publications, mixed cropping showed a significant reduction in soilborne disease and in six, no or a positive effect on disease incidence or severity was found. Diseases caused by splash-dispersed pathogens were less severe in mixed-cropping systems in ten out of 15 studies. The magnitude of disease reduction in mixed compared to single crops varied, from a 63% reduction to a 100% increase in disease. Host dilution appeared to be the most important mechanism of disease suppression for both soilborne and splash-dispersed pathogens (12 and five cases, respectively). Although the use of mixed cropping for soilborne disease suppression is still in its infancy, the wide range of biological effects and interactions observed holds promise for further optimization and management of soilborne diseases, for example, by selecting plant species and cultivars that provide an optimal combination of root architectures.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Mixed cropping
- intercropping
- soilborne pathogens
- crop rotation
- microclimate
- monocropping
- multiple cropping
- disease management
- allelopathy
- ISR (induced systemic resistance)
- SAR (systemic acquired resistance)
- microbial antagonism
5.1 Introduction
During the past decades, intensified mechanization and the use of synthetic fertilizers and crop protectants have substantially increased agricultural yields. However, these practices also resulted in an array of adverse environmental side effects, including soil erosion, water pollution, eutrophication, and reduced innate soil fertility (Gliessman 2001). Acquisition of capital-intensive and crop-specific machinery further narrowed rotations. Although these negative side effects of intensive agriculture counteract the initial increase in food production per unit of area (Matson et al. 1997), ultimately they may lead to a decline in total food production because of land becoming unproductive due to soil erosion and pollution. On the other hand, increasing demands for agricultural products can be met only when high yields per unit of area are achieved, especially when productive land is falling short (Hill 2007). Therefore, it is necessary to find more sustainable ways of cultivating crops without sacrificing on the yield.
Narrow rotations of cash crops have resulted in a high incidence of soilborne diseases (Garrett and Cox 2006). Although genetic resistance and effective pesticides are insufficiently available, many soilborne pathogens, such as Gaeumannomyces graminis var. graminis, can be managed by wide crop rotations (Werker and Gilligan 1990) and other cultural measures (Cook 2001). However, wide crop rotations are, from an economic point of view, undesirable in areas where arable land is limited. Soil fumigants can be highly effective, especially for the control of nematodes, but they have a strong negative impact on non-target organisms and therefore their use is discouraged or prohibited (Martin 2003; Schneider et al. 2003). Methyl bromide, the most common soil fumigant for decades, was added to the list adopted by the Montreal protocol in 1997 and will be banned completely in 2015 (Gullino et al. 2003, Liu et al. 2007). Most soil fumigants are costly and generally too expensive for low-value crops like cereals or for use by subsistence farmers in the developing countries. The application of methods specifically designed to control soilborne pathogens, such as biological soil disinfestation, soil solarization, and flooding, is also often too costly, so they are applicable only to capital-intensive crops (Blok et al. 2000).
While mixed cropping has received attention for its effects on airborne pests (Björkman et al. 2008; Bukovinszky et al. 2004; Risch et al. 1983) and pathogens (Mundt 2002a; Wolfe 1985), the effects on soilborne pathogens barely have attracted attention. In this review, we evaluate how cropping systems and in particular mixed cropping can affect soilborne pathogens. We first define the different types of cropping systems and specifically continuous single-crop cultivation (monoculture), crop rotation (i.e., change of crop diversity in time), and mixed cropping (i.e., any type of growing multiple crops in the same field at the same time). Then we will in short assess and discuss how these cropping systems can affect the dynamics of soilborne diseases. The effects of mixed cropping on soilborne and splash-dispersed fungal and bacterial pathogens will be discussed as well as the mechanisms underlying disease suppression by mixed cropping. We end this review with recommendations and options for the use of mixed cropping that may contribute to improving the sustainability of agricultural production.
5.2 Design of Cropping Systems to Manage Soilborne Diseases
In modern agriculture, cultivation of single crops in a rotation is the most common cropping system for a vast range of crop species worldwide. If properly designed, crop rotation is the most efficient (cultural) practice to reduce the incidence and severity of soilborne diseases (Cook and Veseth 1991). However, crop rotation is not always practiced. In highly mechanized productions, continuous cultivation of the same single crop is regularly practiced, whereas in areas where mechanization, artificial fertilizers, and crop protectants are too costly, diverse forms of mixed cropping are encountered regularly. Disease suppression related to crop rotation and continuous single-crop production has been extensively investigated (Mazzola 2002; Schneider 1982; Weller et al. 2002). However, the effects of mixed cropping on soilborne pathogens have received considerably less attention. Where in literature effects of mixed cropping on soilborne pathogens are reported, they often appear just as a co-observation in studies on crop productivity. The main reasons why the effects on soilborne pathogens have received little attention are the inconspicuous nature of soilborne diseases (Cook 2001), the aspecific disease symptoms, and the inherent difficulty of designing experiments with mixed-cropping systems. A typical example of a disease with aspecific symptoms is Potato Early Dying (Rowe et al. 1987), caused by Verticillium dahliae, which is often erroneously held for drought stress. Furthermore, disease can go unnoticed for some time as is the case for spinach wilt caused by Verticillium dahliae, which induces symptoms only after bolting so that disease is not observed in fresh produce (duToit et al. 2005).
5.2.1 Successive Cultivation of a Single Crop
Continuous cultivation of the same single crop in the same field is practiced in areas where the number of crops that can be grown is agronomically and economically limited (Cook 2001). Under these conditions, mechanization makes cultivation more economically feasible but at the same time hinders the adoption of a more diversified crop rotation. In continuous crop cultivation, inoculum densities of soilborne pathogens increase without exception and a certain degree of damage has to be accepted (Shipton 1975). Some cultural measures including reduced tillage can enhance the survival of certain pathogens (Meynard et al. 2003; Pankhurst et al. 2002). Regular tillage can lead to burial of inoculum of Pseudocercosporella herpotrichoides and limit disease progress in the following season (Colbach and Meynard 1995). On the other hand, reduced tillage and direct drilling resulted in suppression of Gaeumannomyces graminis var. graminis (Pankhurst et al. 2002) because of increased soil organic carbon concentrations and consequently higher microbial activity compared to conventional tillage. Also stimulation of microbial activity through organic amendments can reduce pathogen inoculum or activity (Hoitink and Boehm 1999).
For certain pathosystems, natural disease suppression is known to be induced during continuous cultivation (Schneider 1982; Weller et al. 2002), e.g., Gaeumannomyces graminis in wheat and barley (Gerlagh 1968; Raaijmakers and Weller 1998; Weller et al. 2002), Rhizoctonia solani in sugar beet (Hyakumachi and Ui in Sturz and Christie 2003), Streptomyces scabies in potato (Menzies 1959), and Fusarium oxysporum f. sp. melonis in melon (Alabouvette 1999). Induction of disease suppression can take multiple years and generally it is lost after growing other crops (Shipton 1975). The mechanisms involved have been studied extensively and are linked to the microbial community in soil or the rhizosphere. The best-known mechanisms include antibiotic production (e.g., by strains of Pseudomonas fluorescens), competition by closely related non-pathogenic strains (e.g., competition for carbon by nonpathogenic Fusarium oxysporum), and parasitism (e.g., by Trichoderma spp.) (Weller et al. 2002). For these types of disease suppression to develop and to sustain, both the pathogen and a susceptible host plant need to be present and a certain level of damage has to be accepted. Overall, adequate disease suppression in continuous monocropping systems can be induced in several pathogen–crop combinations. However, other pathogens on the same crop can become problematic. Moreover, the unpredictable time span needed for induction of specific disease suppression and the inflexibility of the cropping system, result in limited applicability of this system for soilborne disease management.
5.2.2 Crop Rotation
Crop rotation is the practice of growing crops on the same field sequentially in time. Crop rotation is commonly practiced to avoid the buildup of soilborne pathogens (Cook and Veseth 1991), to maintain a balanced soil fertility, and to avoid intensive soil tillage before planting root crops (Termorshuizen 2001). The beneficial effect of crop rotation against many soilborne pathogens is due to their limited host range (Krupinsky et al. 2002). The host-dependent reproduction of most pathogens (Garrett and Cox 2006) limits inoculum buildup and viability of the inoculum present diminishes in time when nonhosts are grown (Cook 2001). Alternations of dicotyledonous with monocotyledonous crops are effective in limiting the inoculum levels of the majority of soilborne plant pathogens (Agrios 1997). Alternation with hosts that do not support inoculum production can be a measure to reduce the amount of pathogen inoculum. For example, sugar beet is a host to Verticillium dahliae, but hardly contributes to inoculum buildup, as microsclerotia have not yet been produced at the time when roots are harvested (A.J. Termorshuizen, personal observation).
Green manure or cover crops cultivated in wintertime can be part of the crop rotation. The main reason to grow a green manure crop is to protect soil from erosion and to prevent leaching of mineralized nitrogen. In narrow rotations with a high pressure of soilborne pathogens, the choice of the optimal green manure crop can be a challenge. For example, to reduce nitrate leaching in sandy soils in wintertime in the Netherlands, it is now obligatory to grow a green manure crop following maize cultivation. Due to the late harvest of maize, the choice of green manure crops is usually limited to a grass or winter cereal, which to a great extent resembles maize with respect to its host status for nematodes. The single option farmers have is to harvest their maize earlier, so that they can still sow mustard. Several green manures are known for their capacity to reduce diseases caused by soilborne pathogens. Incorporation of several Brassica species has been shown to reduce disease incidence caused by Rhizoctonia solani, Phytophthora erythroseptica, Pythium ultimum, Sclerotinia sclerotiorum, or Fusarium sambucinum in potato (Larkin and Griffin 2007). The underlying mechanism involves the production of toxic volatiles during decomposition of the cruciferous organic matter. Marigold (Tagetes spp.) is grown as a green manure to specifically suppress Pratylenchus penetrans (Kimpinski et al. 2000), which is likely due to toxic plant exudates.
The effective length of crop rotation as a method to manage specific soilborne pathogens depends on the survival of the pathogen. For example, the resting spores of Spongospora subterranea, the causal agent of powdery scab of potato, can survive for many years in the absence of a host (Jeger et al. 1996), while the survival of Gaeumannomyces graminis is limited to only a few years at most (Gerlagh 1968). Crop rotation is therefore not suitable to manage powdery scab, but it can be a valuable measure to manage take-all disease caused by G. graminis (Cook 2001). For various other soilborne pathogens, e.g., Verticillium dahliae, Rhizoctionia solani, root knot nematodes (Meloidogyne spp.) and root lesion nematodes (Pratylenchus spp.), the design of a proper rotation can be difficult because these pathogens are capable of infecting and/or surviving on multiple hosts.
Crop rotation is a flexible disease management system that is capable of reducing disease losses caused by many soilborne pathogens. However, the need for rotating high-value crops with lower-value crops and the relatively high risk of losing a complete crop make this system often less attractive to farmers.
5.2.3 Mixed-Cropping Systems
Mixed cropping is defined as the cultivation of a mixture of two (or more) crops together in the same field (Trenbath 1976; Willey 1979). There are various types of mixed cropping (Geno and Geno 2001; Vandermeer 1990), each of which may affect soilborne pathogens differently (Table 5.1, Fig. 5.1). Mixed-cropping systems can be characterized according to the degree to which roots of different crop species interact, which is determined not only by the mixed-cropping system but also by the root architecture of each of the crops in the mixture (de Kroon 2007; Weaver 1926).

Mixed-cropping systems
We define here mixed cropping sensu stricto as the practice of growing multiple crops simultaneously without a specific spatial structure. This way of cropping is used frequently in slash-and-burn fallow agriculture or ley farming with multilines or species mixtures (e.g., broadcast-sown grass-clover mixes). In a mixed setting, distances between hosts are generally greater than when grown as single crops and disease will spread more slowly (host dilution). Also allelopathy (Natarajan et al. 1985), microclimate change (Luthra and Vasudeva 1940), root camouflage (Gilbert et al. 1994), and microbial antagonism have been proposed as potential mechanisms underlying the disease suppression induced by mixed cropping (Abadie et al. 1998; Soleimani et al. 1996).
Strip mixed cropping is the “strip-wise simultaneous cultivation of multiple crops in rows, wide enough to permit independent cultivation but still sufficiently narrow to interact agronomically” (quoted from: Vandermeer 1990) (Fig. 5.2). Typically, the width of the strips is adapted to the size of the machinery to be used. Since the crops co-occur on a narrow strip, belowground interactions between the different crop species occur relatively infrequently and therefore the effects on soilborne pathogens are considered to be minor.

Strip mix crop (Photo courtesy of Tim McCabe 1999, USDA-NRCS)
Relay mixed cropping is the simultaneous cultivation of multiple crops during only part of their field period. The second crop is planted at the time when the first crop reaches its reproductive stage but has not yet been harvested. When root systems of both crops overlap sufficiently, disease-suppressive effects due to allelopathy, microbial antagonism, or physical separation between pathogen and host may occur. Because of the time gap between sowing of both crops (strip), tillage between rows of the standing crop can affect pathogen establishment and spread by burial of inoculum (Colbach and Meynard 1995; Meynard et al. 2003).
Row mixed cropping is defined as the production of multiple crops alternately planted in rows. It can be done in an additive design, where both crops are sown at their single densities (Fig. 5.3) or in a replacement design, where one crop is replaced by the other (Fig. 5.4). Irrespective of plant density, disease can spread within rows like in single-culture cropping systems, but between rows the alternate crop(s) can act as a barrier (Michel et al. 1997). Here, host dilution (replacement design), allelopathy, root camouflage, and microbial antagonism may play a role in disease suppression.

Mixed crop, Brussels sprouts–barley, additive design (Photo: G.A. Hiddink)

Mixed crop, triticale–clover, replacement design (Photo: G.A. Hiddink)
Multistorey mixed cropping (Fig. 5.5) is the cultivation of tall perennials combined with shorter biannual or annual crops and is practiced in orchards, tree nurseries, and agroforestry. The area between the rows is used to grow a cover crop to suppress weeds, fix nitrogen, reduce nutrient leaching, and increase the productive surface area. Allelopathy is a possible mechanism of disease suppression, but also roots can act as a physical barrier for pathogen spread, root camouflage, and microbial antagonism.

Multistorey mix crop (Photo courtesy of: Gary Kramer 2001, USDA-NRCS)
Natural vegetation consists mostly of multiple species and can be considered to be closely related to (zero-tillage) mixed cropping. The disease-suppressive mechanisms that operate in natural ecosystems are probably comparable to the mixed cropping or multistorey mixed-cropping system.
As may be clear from the definitions of the different types of mixed cropping, mixed cropping can have many appearances and characteristics. These characteristics often determine if soilborne diseases can be suppressed and what mechanisms for suppression can be held responsible for this disease suppression.
5.3 Disease Reduction in Mixed-Cropping Systems
In 30 out of the 36 studies where the fate of soilborne pathogens was investigated in mixed-cropping systems, soilborne disease was significantly reduced in the mixtures. In the remaining six studies, there was no or a negative effect of mixed cropping on disease suppression (Table 5.2). In ten cases, a positive effect was reported for splash-dispersed pathogens against five with no or negative effects (Table 5.2). The most investigated crop appeared to be wheat, where in nine out of 15 cases (wheat as main crop) disease was reduced in the mixture. Clover was most important as secondary crop in six mixtures with a disease reduction in five of those mixtures. In the following sections, we will discuss the most important proposed disease-suppressive mechanisms and try to explain how they could be operational in mixed-cropping systems.
5.3.1 Host Dilution
In most studies that report a reduction in soilborne diseases or pathogens in mixed-cropping systems, host dilution is assumed to play a crucial role (Table 5.3). The magnitude of disease reduction is variable but can be as much as 50% (Table 5.2). Host dilution is also regarded as the dominant disease-reducing mechanism for airborne pathogens in mixed-cropping systems (Mundt 2002a). The effect of host dilution will likely be a reduction in disease incidence rather than disease severity on infected plants (Burdon and Chilvers 1982). Host dilution might have direct (an effect on the pathogen itself) as well as indirect effects (influencing other factors than the pathogen) on disease suppression in mixed crops. An increased inter-host distance reduces the spread of pathogens. In Pythium garden cress experiments, a distance of 6 cm or more prevented disease spread (Burdon and Chilvers 1975). Similarly, spread of Rhizoctonia damping-off in radish–mustard mixtures decreased with increasing densities of the nonhost mustard plants and spread halted at host densities below a threshold density (Otten et al. 2005). When the distance between host plants becomes shorter than the threshold distance, pathogen expansion can become invasive. The threshold distance is affected by the availability of nutrient resources and interactions with competing microbial communities. These thresholds can be determined based on the percolation theory developed in physics (Bailey et al. 2000). Based on this theory Bailey et al. (2000) calculated the probability of invasive spread of Rhizoctonia solani in microcosms with hosts at varying distances. This, however, is only applicable for pathogens that are able to bridge the gaps between hosts from a nutrient base.
At increasing densities of susceptible roots, disease spread may accelerate if secondary root infections occur as can be the case for G. graminis (Bailey and Gilligan 2000) and R. solani (Otten et al. 2005). Such secondary infections likely occur at a lower rate because of larger inter-root distances in mixed-crop systems. For pathogens with a wide host range such as R. solani, slightly or moderately susceptible plants may also serve as nutrient source without expressing striking disease symptoms (Otten et al. 2005), thus reducing the host dilution effect. The intensity of root intermingling in mixed cropping may be an important determinant for the interference processes (Kroon 2007) and the level of disease suppression may therefore be determined by the crops or cultivars grown and their root architectures. In contrast to pathogens capable of bridging the gaps between host plants by transporting nutrients from a substrate base, host dilution has hardly an effect on pathogens without this capacity, such as powdery scab (Spongospora subterranea), Verticillium wilt, and clubroot (Plasmodiophora brassicae).
For splash-dispersed pathogens in mixed cropping, the host dilution effect is comparable to that of airborne pathogens, influencing disease incidence more than disease severity. The non-host crop simply acts as a physical barrier, thus reducing disease spread as has been shown for Pseudocercosporella herpotrichoides, the causal agent of eyespot in cereals (Villich-Meller 1992). The barrier function can reduce the impact of raindrops thus reducing dispersal, and it can intercept splashing spores that would reach a host plant under conditions of monoculture (Ntahimpera et al. 1998; Soleimani et al. 1996).
5.3.2 Allelopathy
Allelopathy is defined as any biochemical interaction among plants, including those mediated by microorganisms, resulting in either detrimental or beneficial effects on the interacting plants (Wu et al. 2001). In four studies, allelopathy was suggested to play a role in disease suppression in mixed cropping (Table 5.2). When watermelon was intercropped with rice, allelopathic substances from rice roots reduced production and germination of conidia of Fusarium oxysporum f. sp. melonis, leading to a 67% reduction in wilt (Ren et al. 2007). The allelopathic exudates only reduced Fusarium conidial density in the rhizosphere and not in bulk soil indicating a limited diffusion. Delayed germination of spores of F. udum, causing wilt in pigeon pea, has been attributed to allelopathic substances exuded from sorghum roots (Natarajan et al. 1985). To be effective in inhibiting rhizosphere-inhabiting pathogens, allelopathic substances should be present at sufficiently high concentrations in the micro sites where the pathogen is located, and roots of mixed crops should be in close proximity.
An interesting question is whether allelopathy causes death of the pathogen propagules (Ren et al. 2007) or only delays germination (Natarajan et al. 1985). In the latter case, the effect would resemble fungistasis, which is the general phenomenon of restriction of germination and growth of fungal propagules in soil (Lockwood 1977). A high level of soil fungistasis is often assumed to be accompanied by a high level of general disease suppression (Hornby 1983; Janvier et al. 2007; Lockwood 1977). Fungistasis can however also be regarded as a mechanism of delayed activity if conditions are unfavorable for the pathogen, which is also the case if non-lethal allelopathic substances are formed temporarily. The effect can be detrimental, but beneficial to the pathogen as germination in absence of a host plant is, generally, not a desirable trait for pathogens. Roots of non-hosts can sometimes stimulate the germination of the survival propagules of the pathogen (Mol and van Riessen 1995) leading to a decline in the inoculum density. In relay mixed crops, this premature germination might have a disease-suppressive effect, especially in combination with inoculum burial and enhanced microbial antagonism.
Biofumigation has been proposed as a mechanism to suppress soilborne pathogens when Brassica species are used in mixed-cropping systems (Hauggaard-Nielsen and Jensen 2005; Kirkegaard and Sarwar 1998). However, with the exception of the work by Zewde et al. (2007), convincing field data are not yet available. This is in contrast with studies on the biofumigation potential of Brassica crop residues (Kirkegaard and Sarwar 1998, Smolinska et al. 2003), which showed disease suppression for various soilborne pathogens especially in controlled greenhouse experiments.
5.3.3 Microbial Antagonists
In five of the cropping systems listed in Table 5.2, enhanced antagonistic populations were proposed as a main mechanism for disease reduction in mixed-cropping systems. In three cases, pseudomonads and probably antibiotics were involved. For example, wheat root infection by G. graminis var. tritici was reduced by 25% in wheat-trefoil (Medicago lupulina) mixes (Lennartsson 1988). Maximum reduction (73%) in fusarium wilt was reached when bottle gourd was mixed with Chinese chive because of stimulation of Pseudomonas gladioli populations on the Chinese chive roots (Arie et al. 1987). Also, increased occupation of available niches by non-pathogenic Fusaria was held responsible for increased disease suppression in oil-palm–legume mixed cropping (Abadie et al. 1998). The build up of populations of antagonistic microorganisms has been studied mostly in single-crop systems. It seems that the natural build up of antagonists to levels where they are effective takes place mostly as a result of selection or coevolution, i.e., continuous cultivation of the same single crop in the presence of the pathogen (Schneider 1982; Weller et al. 2002). Nevertheless, also in these agro-ecosystems the fate of the same, but introduced antagonistic microorganisms is often inconsistent (Whipps 2001). Rhizosphere microbial communities, including pathogens, antagonists, and plant-growth-promoting bacteria are crop- and cultivar-specific (Germida and Siciliano 2001; Smith et al. 1999) and it might be worthwhile to investigate if these communities can be manipulated by the choice of cultivars in a mixed-crop setting. Crop- or cultivar-specific resistance against races of pathogens is widely known and often applied in mixed crops (Mundt 2002a). Mazzola and Gu (2002) used wheat to stimulate the natural antagonistic populations of fluorescent pseudomonads, which led to control of apple replant disease. The rhizospheres of old wheat cultivars were less aggressively colonized by fluorescent pseudomonads than those of modern ones (Germida and Siciliano 2001). Among tomato lines, genetic differences correlated with Pythium suppression by Bacillus cereus and growth of this biocontrol agent on seeds (Smith et al. 1999). Also legumes may stimulate and support antagonistic Rhizobium bacteria in the rhizosphere (Dakora 2003; Simpfendorfer et al. 1999), which might result in increased pathogen suppression in mixed crops. When growing white clover together with triticale, take-all disease was reduced (Hiddink et al. 2004; Hiddink 2008), although the exact disease-suppressive mechanism remains elusive.
In mixed crops, increased plant diversity leads to more diverse root exudates and consequently to a more diverse rhizosphere-inhabiting microbial community (Kowalchuk et al. 2002; Westover et al. 1997). Rhizospheres of mixed crops support different bacterial and fungal microbial communities compared to the corresponding single-crop rhizospheres (Hiddink et al. 2004; Song et al. 2007). On the other hand, the effect of mixed cropping on the bulk soil microbial community has not been shown (Hiddink et al. 2005a; Kowalchuk et al. 2002). In a more biodiverse setting, the likelihood to encounter microorganisms with antagonistic properties is higher, but at the same time their densities are expected to be lower under these conditions. However, if a higher biodiversity would mean a higher diversity in functions, a higher rate of consumption of root exudates could be expected, which relates to the root camouflage concept proposed by Gilbert et al. (1994). Although increased microbiological diversity is often referred to as an important indicator for soil health (Doran and Zeiss 2000; Mäder et al. 2002; Van Elsas et al. 2002), with respect to disease suppression, its effects can be both positive (more consumption of root exudates, more antagonists) and negative (potentially effective antagonists suffer more from competition and fail to establish and be active).
For bulk soil, an increased bacteria diversity is sometimes related to increased disease suppression. Hiddink et al. (2005a) reported that higher diversity indices for bulk soil bacteria were correlated with a lower disease severity. Suppression of corky root of tomato, caused by Pyrenochaeta lycopersici, was related to a more diverse actinomycete community in bulk soil (Workneh and van Bruggen 1994). Although mixed cropping could increase rhizosphere microbial diversity at intensive intermingling of different roots, the effect on bulk soil biodiversity seems limited (Hiddink et al. 2005a).
Discussing the effect of microbial diversity on disease suppression is complicated since proper methods to quantify diversity are still under development. Cultivation-based approaches do not take into account the non-culturable species, whereas cultivation-independent approaches such as analysis by Denaturing Gradient Gel Electrophoresis (DGGE) underestimate the microbial diversity in soil as only the most abundant species (approximately 0.1–1% of the microorganisms present) are detected (Muyzer et al. 1993). One may assume, however, that the abundant species will also harbor species that contribute to competition for nutrients and space. Another challenge is linking microbial diversity to ecological function (Hiddink et al. 2005a; Nannipieri et al. 2003). The degree of functional redundancy (with respect to disease suppression) could perhaps be regarded as a reliable measure for disease suppression, but how this redundancy could be measured is as yet unclear (Giller et al. 1997; Nannipieri et al. 2003). This could explain why a high biodiversity can be considered a desirable trait, but until indicators quantifying functional redundancy have developed this topic will remain largely speculative.
There clearly is a contradiction between desiring a high functional diversity on the one hand and a high establishment of a given antagonist on the other hand. In soils with a high microbial diversity, a low conduciveness for establishment and growth of an introduced antagonist or pathogen is to be expected. If disease suppression would be controlled by a single antagonist, a high microbial diversity would then be an undesirable trait of soils. This is in line with the observation that establishment of pseudomonads in organic soils (which showed a higher microbial diversity) is more limited than in conventional soils (Hiddink et al. 2005b).
5.3.4 Microclimate
Mixed cropping generally changes the microclimate. Higher soil coverage leads to lower soil temperatures which have been associated with lower disease incidence of Macrophomina phaseolina and Rhizoctonia solani in cotton–sorghum mixtures (Luthra and Vasudeva 1940). The lower level of disease severity of the splash-dispersed Pseudocercosporella herpotrichoides in wheat–clover systems was attributed to a higher decomposition rate of organic material that serves as a base for survival of the pathogen spores (Soleimani et al. 1996). However, increased moisture content in the mixed crop could have increased soilborne pathogens such as Pythium spp., which can survive and disperse more easily in moist soils. Likewise, airborne diseases such as halo blight caused by Pseudomonas syringae pv. phaseolicola could be more severe in mixed bean/maize than in a single bean crop (Mabagala and Saettler 1992).
5.3.5 Induced Systemic Resistance (ISR) and Systemic Acquired Resistance (SAR)
Mixed cropping can bring about ISR (induced by non-pathogenic microorganisms) or SAR (stress inducers like water stress, salinity, allelopathic substances, or pathogens) if one crop creates the right condition for ISR/SAR inducers for which the alternate crop is sensitive (Hamerschmidt et al. 2001). Both ISR and SAR can be interpreted as a form of increased generalized resistance in response to an external stress (Agrios 1997). The response starts from a localized point and can spread throughout the whole plant as a result of signal transduction. Induced resistance could be due to direct effects of stress-inducing root exudates or indirect effects via root-exudate-affected microbial populations (Kloepper et al. 1992). ISR has been mentioned as a mechanism for reduction of several airborne pathogens such as powdery mildew in barley cultivar mixtures (Chin and Wolfe 1984). However, neither ISR nor SAR have been suggested to play a role in suppression of soilborne pathogens in mixed crops (Table 5.2), probably because of difficulties to prove this experimentally.
5.3.6 Nutrients and Disease Development
Nutrients can affect disease development above and belowground (Walters and Bingham 2007). In mixed crops, uptake of nitrogen from undersown clover reduced take-all disease severity in barley (Garrett and Mann 1948). Not only the amount but also the form of nitrogen is important. Exudation of ammonium from clover roots (Paynel and Cliquet 2003) may lead to a reduction in the rhizosphere pH in cereal roots, thereby influencing the antagonistic microbial population and decreasing infection by G. graminis (Sarniquet et al. 1992; Smiley 1978). Also, availability of several other elements such as potassium, phosphorus, sulfur, and silicon will influence disease development directly or indirectly (e.g., Walters and Bingham 2007) in mixed crops but are not further discussed in this review.
5.4 Similarities and Differences Between Disease-Suppressive Mechanisms in the Different Cropping Systems
All three cropping systems, continuous monocropping, crop rotation, and mixed cropping, can contribute to the management of certain soilborne pathogens. Crop rotation is the most commonly applied method to manage soilborne pathogens. However, while rotation schemes can reduce specific soilborne pathogens, for several other, more generalist pathogens, crop rotation is not necessarily a proper solution. Also, wide crop rotations can be undesirable from an economic point of view. Continuous cultivation of the same crop can result in a persistent decline of a pathogen, as is the case for take-all disease of cereal crops. Continuous cultivation of the same crop has not been “invented” as a management tool for soilborne pathogens perse, but induction of disease suppression is a complementary benefit in situations where no options other than continuous cultivation of single crops are available. This specific suppression usually is only active against a single pathogen leaving opportunities for other soilborne pathogens to develop and cause disease. Mixed cropping has been practiced for ages in all sorts of combinations, although not specifically designed for suppression of soilborne pathogens, but rather as an insurance against crop failures and soil erosion.
In all three types of cropping systems, multiple disease-reducing mechanisms are active, but mixed cropping offers the most diverse form of disease suppression because root systems of different crop species interact. In mixed cropping systems, the most important disease-reducing mechanism appears to be host dilution. The magnitude of this effect depends on the planting density, the type of mixed cropping, and root architecture of the crops grown. Competition will affect the distribution of roots in mixed crops (de Kroon 2007; reviewed by Hauggaard-Nielsen and Jensen 2005). Allelopathic effects, nutrient concentrations, and water flow will determine how the roots interact and the diversity of (microbial) interactions in the rhizosphere (Bowen and Rovira 1976). Furthermore, as long as host species are mix-cropped with non-hosts in lower densities, host dilution will inevitably lead to a reduction in the number of diseased plants per area.
Other factors that result in disease suppression, such as allelopathy and antagonism induced by the non-host crop, depend on characteristics of all crops present in the mix. Biofumigation using Brassica species in mixed cultivation has received attention recently, but its effectiveness is still limited (Hiddink et al. 2005a). Breeding for Brassica species exhibiting higher glucosinolate contents is an option to increase their effectiveness (Matthiessen and Kirkegaard 2006). More effective suppression can be expected from legumes, which can excrete allelopathic root exudates and support potentially antagonistic microorganisms, besides fixing nitrogen (Dakora 2003). Also the use of specific crops and cultivars that support antagonistic microorganisms (Mazzola and Gu 2002; Smith et al. 1999) can be a valuable tool to create mixtures that actively suppress soilborne pathogens.
5.5 Practical Feasibility of Mixed Cropping
Although it is clear that mixed cropping can reduce soilborne diseases, it also has an inherent weakness: the presence of multiple crop species may bring about a greater variety of soilborne pathogens albeit likely at lower densities for each of the crops. An important question is whether and how mixed crops should be rotated and what the choice of rotation crops in time should be. When rotated, mixtures of wheat or barley containing oats resulted in lower disease levels in the crops the following year than mixtures of barley and wheat (Vilich 1993). An additional question that should be addressed is: Does mixed cropping of two crops continuously for two (or more) years lead to less disease than growing those same two crops in rotation? It is surprising that, to the best of our knowledge, no answer to this question is available in the literature. The answer to this question can be complex, as was shown by Hiddink (2008). In this study, take-all disease was lower during three consecutive years in a triticale–white clover field compared to single-cropping triticale. However, in the fourth year, Fusarium infected white clover and reduced its stand, which in turn caused an increase in take-all in triticale in the mixture to a disease level above that obtained in the single-cropped triticale. Soilborne pathogens with broad host ranges or long-term survival structures are likely to be less suppressed in mixed crops grown repeatedly. If pathogens like Fusarium in clover (Hiddink 2008) are not actively suppressed by the co-occurring crop, inoculum will continue to build up and rotating the crops in the mixture would have been a better tool to suppress the pathogens. To manage mixed crops for the suppression of soilborne diseases requires advanced skills of the farmer and knowledge of the pathogens that might cause diseases in both mixed-crop components. It can be more labor-intensive and not suitable for mechanized production of all crops. Certain crops are not suitable to grow in mixed crops because of their weak competiveness. The degree of intercrop competition is decisive whether a certain combination can be grown. Thus, although club root, caused by Plasmodiophora brassicae, was reduced in a barley–Brussels sprouts mixed crop, yield of Brussels sprouts was reduced by nearly 50% because of competition by barley (Hiddink 2008). However, often an overall yield increase is observed in mixed crops. This effect is generally expressed as the Land Equivalent Ratio (LER) (Vandermeer 1990). The LER is the sum of the yields of both components per unit of land area combined divided by the area of land needed to obtain the same yields when both components are grown as single crops (Vandermeer 1990). Mixed crops have been grown for ages, because of their yield stability and mixed cropping is still practiced for this reason in tropical regions (Vandermeer 1990). Co-occurring crops compensate for failure of one of the crops due to soil and airborne pathogens, weeds, temperature-, and water stress (Vandermeer 1990). This kind of growth compensation is an important reason for mixed cropping.
Overall, we conclude that it is interesting to consider mixed cropping where land-use efficiency and yield assurance are important reasons for practicing mixed cropping. However, application of mixed crops as tools for soilborne pathogen management is still in its infancy and not yet reliable enough.
5.6 Conclusion
In spite of the frequently observed disease or pathogen suppression (40 out of 51 observations) in mixed cropping, this system will not be a panacea for combating soilborne plant pathogens. However, in some cases it can contribute substantially to the management of soilborne pathogens. Design of mixed-cropping systems as a tool for suppressing plant pathogens is still in its infancy compared to continuous monocropping and crop rotation. The available literature is limited and scattered. In this literature review we showed that the most frequently observed disease-suppressive mechanism is host dilution (17 times for soilborne and splash-dispersed pathogens combined). Likely, however, multiple factors affect the extent of disease suppression. We think that much can be done to optimize the disease-suppressive effects based on allelopathy and antagonism. Although we focused on effects of mixed cropping on soilborne pathogens, other benefits should also be considered when evaluating mixed cropping. Reduction in plant pests and weeds has been reported widely (Baumann et al. 2001; Bukovinszky 2004). Reduced growth of one crop results in lower competition and can increase the production of the accompanying crop and thus increase overall yield stability per unit of area. This could be especially useful when no direct control measures such as pesticides are available. Another important benefit of mixed cropping is the higher potential yield per unit of area of cultivated land. This would reduce the plant production acreage needed to produce a certain amount thus using the available production factors more efficiently and reducing nutrient leaching, water runoff, and soil erosion per unit of yield. More production per area of land also means that competing claims for land needed for the production of human food and animal feed and for the production of bio-fuels can be relieved to some extent if they can be grown on the same area of land at the same time.
References
Agrios GN (1997) Plant pathology. Academic, London, UK, pp. 635
Arie T, Namba S, Yamashita S, Doi Y, Kijima T (1987) Pseudomonas gladioli (Japanes title). Annu Rev Phytopathol Soc Jpn 53:531–539
Bailey DJ, Otten W, Gilligan CA (2000) Saprotrophic invasion by the soil-borne fungal plant pathogen Rhizoctonia solani and percolation thresholds. New Phytol 146:535–544. http://www.jstor.org/stable/2588935
Garrett KA, Cox CM (2006) Applied biodiversity science: managing emerging diseases in agriculture and linked natural systems using ecological principles. In: Ostfeld R, Keesing F, Eviner V (eds) Infectious disease ecology: the effects of ecosystems on disease and of disease on ecosystems. Princeton University Press, Princeton, NJ, pp 368–386
Geno L and Geno B (2001) Polyculture production – principles, benefits and risks of multiple cropping land management systems for Australia, pp 105. Publication No. 01/34, Rural Industries Research and Development Corporation (RIRDC), Kingston, ACT
Gu Y-H, Mazzola M (2003) Modification of fluorescent pseudomonad community and control of apple replant disease induced in a wheat cultivar-specific manner. Appl Soil Ecol 24:57–72. doi: 10.1016/S0929-1393(03)00066-0
Hiddink GA, Termorshuizen AJ, Raaijmakers JM, van Bruggen AHC (2004) Effect of mixed cropping on rhizosphere microbial communities and plant health. In: Book of abstracts international congress rhizosphere 2004, Munich, Germany, 12–17 Sept 2004
Lennartsson M (1988) Effects of organic soil amendments and mixed species cropping on take-all disease of wheat. In: Allen P, van Dusen D (eds) Global perspectives on agroecology and sustainable agricultural systems: proceedings of the sixth International scientific conference of the international federation of organic agriculture movements, Santa Cruz, August 18–20, 1986, pp 575–580
Trenbath BR (1976) Plant interactions in mixed crop communities. In: Papendick RI, Sanchez PA, Triplett GB (eds) Multiple cropping. ASA Special Publication No. 27, ASA, SSSA, CSSA, Madison, WI, pp 129–169
Vandermeer JH (1990) Intercropping. In: Carrol CR, Vandermeer JH, Rosset P (eds) Agroecology. Mcgraw Hill, New York, pp 481–516
Van Rheenen HA, Hasselbach OE, Muigai SGS (1981) The effect of growing beans together with maize on the incidence of bean diseases and pests. Neth J P1ant Pathol 87:193–199
Weaver JE (1926) Root development of field crops. McGraw-Hill, New York. http://www.soilandhealth.org. Accessed on 2010
Wu H, Pratley J, Lemerle D, Haig T (2001) Allelopathy in wheat (Triticum aestivum). Ann Appl Biol 139:1–9
Zewde T, Fininsa C, Sakhuja PK, Ahmed S (2007) Association of white rot (Sclerotium cepivorum) of garlic with environmental factors and cultural practices in the North Shewa highlands of Ethiopia. Crop Protection 26:1566–1573. doi: 10.1016/j.cropro.2007.01.007
Acknowledgments
We thank Dr. J.M. Raaijmakers for his valuable comments and suggestions after reviewing this manuscript. This work was part of the project “Enhanced Biodiversity” funded by the section Earth and Life Sciences of the Dutch Scientific Organization (NWO-ALW, project number 014.22.032).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2010 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Hiddink, G.A., Termorshuizen, A.J., van Bruggen, A.H.C. (2010). Mixed Cropping and Suppression of Soilborne Diseases. In: Lichtfouse, E. (eds) Genetic Engineering, Biofertilisation, Soil Quality and Organic Farming. Sustainable Agriculture Reviews, vol 4. Springer, Dordrecht. https://doi.org/10.1007/978-90-481-8741-6_5
Download citation
DOI: https://doi.org/10.1007/978-90-481-8741-6_5
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-90-481-8740-9
Online ISBN: 978-90-481-8741-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)