
Abstract
Climatic changes not only produce latitudinal shifts in different ecological belts, equally significant is the diverse pattern of minute but extremely important changes observed in the mountains of the arid land regions of the world. These areas of greater elevation enjoy a higher local rainfall even today and during periods of wetter climate, rainfall values increase appreciably creating large islands of, relatively, rich vegetation and attractive landscapes (Butzer and Twidale, 1966). The preceding pages of Chapters 1 and 2 of this book confirm this statements. The variation in climatic conditions prevailing on the Afro-Asian Mediterranean-Red Sea coastal lands recognizabl affect their plant life, the species of which exhibit special adaptation in structure, ecology, and associations (Kassas, 1996). Here are some concluding remarks on the relationships between climate and plant life of these coastal lands, discussing it under the two themes of climatic features and vegetation forms.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
3.1 Introduction
Climatic changes not only produce latitudinal shifts in different ecological belts, equally significant is the diverse pattern of minute but extremely important changes observed in the mountains of the arid land regions of the world. These areas of greater elevation enjoy a higher local rainfall even today and during periods of wetter climate, rainfall values increase appreciably creating large islands of, relatively, rich vegetation and attractive landscapes (Butzer and Twidale, 1966). The preceding pages of Chapters 1 and 2 of this book confirm this statements. The variation in climatic conditions prevailing on the Afro-Asian Mediterranean-Red Sea coastal lands recognizably affect their plant life, the species of which exhibit special adaptation in structure, ecology, and associations (Kassas, 1996). Here are some concluding remarks on the relationships between climate and plant life of these coastal lands, discussing it under the two themes of climatic features and vegetation forms.
3.2 Climatic Features
Climate is the master of all environmental factors, it controls not only the growth and development of the plants but also their geographical distribution. This is obvious in that the major vegetation forms of the earth are related to the different climatic types. However, other environmental factors (land-forms, habitat types, soil characteristics, etc.) are also locally effective. The climatic features associated with the Afro-Asian Mediterranean and Red Sea coastal lands are described below.
3.2.1 Afro-Asian Mediterranean Coastal Lands
Branigan and Jarrett (1975) state that the lands around the Mediterranean Sea, including the North African and SW Asian coastal lands, represent most clearly defined climatic units in the world. In summer, following the northward march of the sun, the Mediterranean region lies in the belt of the north-east trade winds and of high pressure and it is, therefore, dry. In winter, with the retreat of the sun south of the equator, the region falls under the general influence of the westerlies and the depression associated with them, and is wet. Many of the depressions travel the whole length of the Mediterranean from west to east, and most of the winter rain typical of the region owes its origin to them. It may be said, in general, that the Mediterranean region has a temperateFootnote 1 and changeable climate for the winter half of the year and a more uniform climate of the “hot dry” type during the summer.
The main characteristics of the Mediterranean climate may be summarized under four headings: winter rain, summer drought, mild winter temperature (mean ≥ 6.1°C) and hot summer (mean ≥ 21.1°C). It is also noteworthy that long periods of sun-shine and cloudless skies are experienced at all seasons.
Ayyad and Ghabbour (1993), following Meig’s climatic system, stated that the climate of the North African Med. coastal lands lies in the area of “warm coastal desert”: summer’s warmest month has a mean temperature less than 30°C and winter coldest month has a mean temperature above 10°C. Though occasional short rain-storms occur in winter, most of the days are sunny and mild. Winds are generally gentle, but violent sand-storms and dust pillars are not rare. Dry hot dust laden winds from the south known as khamasin Footnote 2 in Egypt, sirocco in Libya and Tunisia blow occasionally for about 50 days during spring and early summer. These winds develop as a result of the passing of lower pressure over the Mediterranean.
The mean annual rainfall in the North African Med. coastal lands varies from about 140 mm in the arid climate of Egypt and Libya to almost 600 mm at the transition between upper semiarid and subhumid climates in northern Tunisia and the highlands of Cyrenaica. The seasonal distribution of rainfall can be distinguished into two patterns: one in which most of the rain (60% or more) occurs during winter (November–February) and the summer is virtually dry and one in which rainfall is more or less evenly distributed between autumn, winter and spring with some rainy days during summer. The first pattern of rainfall distribution characterizes the arid and semiarid coastal ecosystems of Egypt and Libya while the second pattern occurs in Tunisia.
In the SW Asian Med coastal lands, represented by Palestine-Israel-Syria section, the Mediterranean climate is marked by a mild, rainy winter and prolonged dry and hot summer. Geographical latitude and altitude, the blocking effect of the mountain ranges and distance from the sea are among the factors which modify the climate (Zohary, 1962). The effect of latitude manifests itself in the abrupt north to south decrease in annual rainfall so that within an average of 4 latitudinal degrees (33–38°N) the rainfall drops from about 1,000 to 25 mm/year. Temperature, increases from north to south, with the mean annual temperature rising from below 16°C in the north to approximately 23°C in the extreme south of the Palestine-Israel coast. In a west to east direction, from the sea shore landwards annual rainfall and mean temperature undergo similar but less regular changes, this is because of the interference of the Israeli and Jordanian mountain ranges. As a result of their interception of rains, part of the Jordan valley is turned into a rain-shadow desert. The tempering influence of the Mediterranean Sea is greatly limited by the mountain ranges, leaving most of the Palestine-Israel area open to a widen range of seasonal and diurnal temperatures. Climatically Zohary (1962) classified the SW Asian Med. coastal land, represented by the Palestine-Israel area, into three zones:
-
1.
an arid zone with 200–25 mm/year rainfall,
-
2.
a semi-arid zone where annual rainfall ranges, between 200 and 400 mm,
-
3.
a sub-humid zone with an annual rainfall of 400–1,000 mm.
3.2.2 Afro-Asian Red Sea Coastal Lands
The Red Sea Basin occupies an area of about 810,000 km2 including essentially the Red Sea coastal plain (210,000 km) and the highlands (600,000 km2) bordering them on both sides in Africa and Asia. The entire area of the Red Sea and its coastal lands are generally hot deserts with high temperature and low precipitation. Two climatic types are recognized: the monsoonFootnote 3 type in the south with summer rainfall and the Mediterranean type in the north with winter rainfall. Rainfall regions are almost comparable on both African and Asian sides having a bimodal distribution pattern with spring and autumn peaks that mimics the Mediterranean pattern. The annual precipitation is variable, from virtually less than 5 mm in the tropical coastal lowlands to more than 200 mm in some locations where the shores of the Red Sea are near the mountain ranges. On the highlands, the annual precipitation varies from 300 mm or slightly less to over 1,000 mm. Such variation of rainfall has its effects on the diversity, density and distribution pattern of the natural vegetation (Le Houerou, 2001). Temperatures in the low lands are among the highest on the planet reaching an annual mean of about 30°C, with maxima exceeding 60°C. The potential evapotranspiration is also among the highest on the earth, between 1,800 and 3,000 mm/year. The decrease in mean annual temperature and potential evapotranspiration with altitude is about 0.55°C and 55 mm/100 m, respectively. Rainfall increases by 10% for each increase of 100 m, i.e. doubling for each 1,000 m up to 2,000–3,000 m, but then decreasing. In this context, Dallman (1998) stated that the elevation is important in relation to temperature and rainfall. In the montane countries, temperature decreases by roughly 5°C/1,000 m. Rainfall is typically greater with increase in elevation. The flora and vegetation are likewise similar and occasionally identical for comparable degrees of elevation, temperature and aridity. The two areas, north-east and south-west of the Red Sea as a whole including their highlands, share 60% of the total number of the genera and some 30% of their approximately 5,000 species (Le Houerou, 2001).
Over the whole Red Sea Basin, winter may extends from mid-October to mid-April with summer occupying the rest of the year (Anonymous, 1993). The January mean daily temperature varies from about 20°C in the far north to about 29°C in the far south. The corresponding July figures are 35 and 40°C, respectively. Rainfall over the Red Sea coastal lands as a whole is sparse, and sporadic and often very localized (Zahran and Al-Kaf, 1996). A particular location may receive no rain for years, and then can experiences a brief heavy rainfall which may not be repeated for a similar length of time. For example in Hodayda in Yemen (Asian Red Sea coast), the total annual rainfall in 1989 was 207.3 mm occurring in January (31.4 mm), February (30.5 mm), March (56.4 mm) and April (89.0 mm). In 1988, the total annual rainfall was 127 mm occurring in April (39 mm), August (61.4 mm) and September (27.5 mm). These 2 years were exceptionally wet. The mean annual rainfall on the Hodayda coast usually ranges between 35 and 60 mm, mainly between October and April.
The African Red Sea coastal lands in the Sudan receives the main part of rainfall during the winter season: October–January, i.e. Mediterranean affinity (Kassas, 1957). As the prevalent continental northerlies pass over the warm water of the Red Sea, they absorb a considerable amount of moisture, causing convectional rainfall. This is especially affected by winter cyclonic activity over the Red Sea (Hefny, 1953). Though the coast is shut off from summer monsoon influence by mountain barriers, local penetration due to low topography can bring summer cloudbursts in July–August. The convectional rain of winter decreases landward (westward). For example, Sallom (19° 22′N, 37° 06′E, Alt. 170 m), which lies on the coastal plain at a distance of about 22 km from the shore, receives less winter rain (56 mm/year) than Suakin (19° 07′N, 37° 20′E, Alt. 5 m) (122 mm/year) to its south east and Port Sudan (19° 37′N, 37° 13′, Alt. 5 m) (92 mm/year) to its northeast. In contrast, Gebeit (18° 57′N, 35° 5′E, Alt. 795 m), within the Red Sea hills 60 km inland, receives a negligible amount of winter rainfall (12 mm, in October–February) because it lies at about the northern boundary of the tropical continental climate region where the southerlies bring summer (May–September) rainfall (111 mm/year).
Variability of the annual rainfall, a characteristic feature of the arid lands, is obvious on the Sudanese Red Sea coast (Kassas, 1957). The Suakin rainfall, for example, ranges from 617 mm/year in 1896 to 33 m/year in 1953 and in Port Sudan, rainfall ranges from 422 mm/year in 1925 to 19 mm/year in 1910 (a variability of about 56%). The moisture-laden northerlies cause convectional rain at the coast which decreases inland. But as they cross the coastal plain, the northerlies encounter the Red Sea hills causing orographic rain and other forms of condensation as dew. The seaward (east) slopes of the hills receives a greater share of this rain. Again the higher hills farther inland receiver, by virture of their altitude, more of this moisture than the low hills and buttes.Footnote 4 This will have a noticeable effect on the plant cover and its altitudinal zonation pattern.
3.3 Vegetation Forms
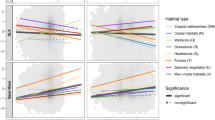
Eleven major forms of vegetation are recognized along the Afro-Asian Mediterranean and Red Sea coastal land’s (Table 3.1): mangrove, reed swamp, salt marsh, sand dune, rocky ridge, desert, Mediterranean steppe grassland, broad-leaved evergreen forest, stunted woodland (matorral), coriferous forest and scrubland vegetation. The fact that the distribution of these vegetation forms along the coastal lands is affected mainly by the prevailing climatic condition is of ecological interest. Four vegetation forms: (reed swamp, salt marsh, sand dune and desert) occur in all sections of the Afro-Asian Mediterranean and Red Sea coastal lands, while the others have different distribution patterns. Mangrove vegetation is a tropical formation, and hence, its presence is restricted to the coastal belts of the Afro-Asian Red Sea but absent from those of the Mediterranean Sea. Rocky ridge vegetation occurs in the Egyptian and Libyan sections of the North African Mediterranean coast whereas Mediterranean steppe grasslands are localized in the coastal lands of the three western countries of the North African Mediterranean coast (Morocco, Algeria and Tunisia). The same distribution pattern is seen in broad-leaved evergreen forest and stunted woodland (matorral) but both also occur in the escarpment of the Palestine-Israel coast of the SW Asian Mediterranean. Coniferous forests are ecologically related to cold climate territories and, therefore, occur at high altitudes of the North African and, SW Asian Mediterranean, and the southern Sinai and Asian Red Sea coastal lands. Scrubland vegetation is abundant in the Afro-Asian Red Sea coastal lands, including Southern Sinai, poorly represented in the Palestine-Israel section of the SW Asian Med. and almost absent from North African coastal lands. Details of each of these eleven vegetation forms may explain the relationship between them and their floristic composition with climatic conditions particularly rainfall and temperature.
3.3.1 Mangrove Vegetation
Mangrove vegetation is one of the characteristic ecological features of both African and Asian Red Sea coastal belts with almost the same latitudinal distribution, but are totally absent from the coastal belts of the North African and South-East Asian Mediterranean. It is, actually, a climatic phenomenon, mangrove forests are tropical formations that only grow in the swamps of warm and hot coastal belts between the two tropics: the Tropic of Cancer (23° 30′N) and Tropic of Capricorn (23° 30′S). Low temperature is fatal to the young seedlings of these plants, a cogent explanation for the absence of this vegetation form from the coastal belts of the cool and cold areas of the world (Chapman, 1977).
Three mangrove species are recorded along the Afro-Asian Red Sea coastal belts: Avicennia marina, Rhizophora mucronata and Bruguiera gymnorrhiza. A. marina is well developed along both coasts of the Red Sea starting from latitude 27° 14′N southwards in Egypt, the Sudan, Eritera and Djibouti (African Red Sea coast), and in Saudi Arabia and Yemen (Asian Red Sea coast). North of this latitude, A. marina forest is practically absent except for a few isolated patches with stunted bushes in the swamps of Ras Muhammed in Sinai (27° 40′N) and on the Nabq coast at the southern part of the western coast of the Gulf of Aqaba (28° 10′N). A comparable distribution pattern is recognized along the Asian Red Sea coast. The most extreme northern presence of A. marina is recorded in the coast at Duba (27° 22′N) (Zahran et al., 1983; Zahran, 2007). These findings confirm Chapman’s statement (1977): “Mangal reaches its optimal development in the tropics but it extends to the subtropics described as warm temperate where the air temperature seems cooler, particularly at night, than the range of tolerance of the seedlings of the mangrove plants”.
R. mucronata and B. gymnorrhiza mangroves have a different distribution pattern: both are absent from the northern sections of the Afro-Asian Red Sea coastal belts (north of 23°N). In Egypt R. mucronata starts to appear in the mangrove swamps of the Shalatein coast (23°N) and continues southwards to Mersa Halaib (22°N) on the Sudano-Egyptian border. R. mucronata has been also recorded further south in Suaken (Sudan) and in Djibouti (19° 15′–19°N), Zahran et al. (2009). Along the Asian Red Sea coastal belt, R. mucronata, occurs on Jizan coast, Saudi Arabia (16° 20′N) and southwards at Hodayda in Yemen (15°N). B. gymnorrhiza was recorded in Suaken (Sudan), Hodayda (Yemen) and Djbouti coasts. Nowadays, it is absent from these coasts due to the influence of man (Andrews, 1948, 1950–1956; Draz, 1956; Zahran et al., 1983, 2009).
3.3.2 Reed Swamp Vegetation
Reed swamp vegetation inhabits the lakes, lagoons and water creeks of the Afro-Asian Mediterranean and Red Sea coastal lands. The most widespread and characteristic species (helophytes) are: Phragmites australis and Typha domingensis, are recorded in almost all stands of this vegetation form. Other common swampy species include: Cyperus articulatus, C. dives, Eleocharis palustris, Lemna spp, Samolus valerandi, Scripus mucronatus, Spirodela polyrrhiza and Wolffia hyalina. There are reed-swamps species (helophytes) with restricted geographical distribution; many of them occur only in the swampy areas of the Afro-Asian Mediterranean coastal lands while others occur only in Afro-Asian Red Sea coastal lands. For example, Typha elephantina has been recorded in a swampy area in the downstream part of Wadi Gizan along the Asian Red Sea coast of Saudi Arabia (Zahran, 1982b), but has never been recorded in the swamps of the Afro-Asian Med. Coast. Andrews (1950–1956) recorded some helophytes growing in the swampy habitat of the Red Sea coast of the Sudan: (Cyperus amauropus, C. schimprianus, C. cristatus var. nigricans, C. rotundus var. nubicus, C. microbolbos etc.), and Cyperus bulbosus was recorded from the Red Sea coast of the Sudan and Egypt but absent from the Med. coast. The are also the helophytes recorded from the Afro-Asian Mediterranean coastal land but absent from those of the Red Sea coast including: Cyperus papyrus, C. compressus (= C. conglomerates), C. rotundus var. rotundus, C. capitatus, C. difformis, C. alopecuroides, Juncus subulatus, Carex divis, C. distans, Eleocharis palustris, Fimbristylis bisumbellata, Fuirena pubescens (= Scirpus pubescens), Scirpus maritimus, S. littoralis, S. holoschoenus, Phalaris canariensis, P. aquatica. etc.
3.3.3 Salt Marsh Vegetation
Littoral salt marsh vegetation occurs in coastal lands subjected to maritime influences: periodic flooding with sea water, sea water spray, sea water seepage etc. They can be found, in general, if any of the following physiographic conditions is fulfilled: the presence of estuary, the shelter of spits, off-shore barrier islands, large or small protected bays with shallow water etc. (Chapman, 1974).
Littoral salt marshes of the arid and semi-arid coastal lands are really fringes of inland deserts and their landward area being defined by their desertic qualities. Climate and terrain can be used to mark off their inland boundary. The halophytes, the plants of this vegetation, may be considered as high specialized plants most of which have great tolerance to salt. The salt tolerance of the halophytes increases both during their growth and development from generation to the next (Keith, 1958).
Zonation pattern is a universal feature of the littoral salt marsh vegetation (Chapman, 1974; Kassas, 1957) controlled mainly by tidal movement, land relief, soil salinity and underground seepage of sea waer. The species of the lower zones, near to the sea, may be very different from those of the higher zones and “in many instances, only a few inches increase in level results in a profound change” (Chapman, 1974). The zonation sequence of the littoral salt-marsh vegetation is complete only where the shore rises gently and gradually into the land.
Littoral salt marsh vegetation is well developed along the Afro-Asian Med. and Red Sea coastal lands but the geographical distribution of its floristic elements varies. Some of these species have wide ecological amplitude and grow under the different climatic conditions of these coastal lands, whereas others have a narrow range and grow either in the Mediterranean or the Red Sea coastal lands. Representative characteristic species of these three groups of halophytes are:
-
(a)
Halophytes occurring in both the Afro-Asian Med. and Red Sea coastal lands include: Arthrocnemum macrostachyum, Aeluropus lagopoides, Cyperus laevigatus, Cressa cretica, Halopeplis perfoliata, Halocnemum strobilaceum, Imperata cylindrica, Juncus rigidus, Limonium pruinosum, Nitraria retusa, Salicornia fruticosa (= Sarcocornia fruticosa), Sporobolus spicatus, Suaeda vermiculata, Tamarix aphylla, T. nilotica, T. tetragyna etc.
-
(b)
Halophytes occur only in the Afro-Asian Med. coastal lands include: Atriplex halimus, A. portulacoides, A. leucoclada var. inamoena, Frankenia revoluta, Inula crithmoides, Juncus acutus, Limoniastrum monopetalum, Salsola tetrandra, Schoenus nigricans, Sporobolus pungens, Suaeda paleastina and S. pruinosa.
-
(c)
Halophytes restricted to the littoral salt marshes of the Afro-Asian Red Sea coastal lands include: Aeluropus littoralis, Limonium axillare, Salsola spinescens, Sevada schimperi, Suaeda monoica, Tamarix amplexicaulis and T. macrocarpa.
The geographical distribution of six of these halophytes deserves extra consideration, these are: Halocnemum strolbilaceum, Nitraria retusa, Suaeda monoica, Halopeplis perfoliata, Tamarix macrocarpa and Salicornia fruticosa.
-
1.
Halocnemum strobilaceum dominates a community common in the littoral salt marshes of both areas but its presence in the Afro-Asian Red Sea coasts is restricted to the northernmost 400 km but rare or even absent in the southern sections (south of latitude 27°N).
-
2.
Nitraria retusa similarly dominates a community common in the littoral salt marshes of the Afro-Asian Med. and Red Sea coastal land but its presence is restricted to the northern 700 km stretch, i.e. north of Mersa Alam in Egypt and Umlog in Saudi Arabia, southwards, it is absent along the whole Red Sea coasts.
-
3.
Suaeda monoica is a succulent halophyte comparable in habit and habitat to Nitraria retusa. The two species have an ecological range that extends beyond the limits of the salt marsh to the fringes of the coastal desert plain. Both may form and protect mounds and hillocks of sand, though S. monoica hills can be larger (Zahran and Willis, 2008). However, these two species seem to have different geographical areas. Along the African Red Sea coast, N. retusa occurs in the northern 700 km stretch south of Suez but is absent in the south. The reverse in true for S. monoica gradually replacing N. retusa over the 300–700 km stretch south of Suez. Further south, S. monoica is a salient feature of the southern section of the Red Sea coast in Egypt as well as in the Sudan and Eritrea (Kassas, 1957; Hemming, 1961). The same geographical distribution of N. retusa and S. monoica has been noticed on the Asian Red Sea coast (Zahran, 1982b). S. monoica has also been recorded by Zohary (1962) in the inland desert of Palestine-Israel area.
-
4.
Halopeplis perfoliata is a succulent halophyte recorded on the littoral salt marshes of both the Afro-Asian Med. and Red Sea coastal lands. Its presence and distribution along the Red Sea coastal belts is of ecological interest. Along the African Red Sea coast within the 950 km stretch from Suez to Mersa Kilies, H. perfoliate is recorded in one locality (55 km south of Ras Gharib, 295 km south of Suez) but is otherwise very rare or absent. South of Mersa Kilies, H. perfoliata and its community become common features of the littoral salt marsh vegetation. It is also very common further south in the Sudan and Eriterea (Kassas, 1957; Hemming, 1961). Almost, the same geographical distribution of H. perfoliata has been recorded by Zahran (1982b) on the Asian Red Sea coast.
-
5.
Tamarix macrocarpa (= T. passerinoides) is absent from the Afro-Asian Med. coastal lands and is rare along the Egyptian Red Sea coast. Its presence is limited to a narrow area: El-MallahaFootnote 5 20–40 km south of Ras Gharib (260–280 km south of Suez) where there is an inland depression separated from the shore by an elevated raised beach and fed with seawater through underground passage. T. macrocarpa has not been recorded from the Yemeni Red Sea coast (Wood, 1997) but has been further north in Saudi Arabia by Migahid and Hammouda (1978).
-
6.
Salicornia fruticasa (= Sarcocornia fruticosa) is a widespread succulent halophyte along Afro-Asian Med. coastal land but is absent from the Asian Red Sea coast. On the African Red Sea coast, S. fruticosa is recorded from only one site, the El-Mallaha saline depression in Egypt, where it dominates pure stands with 50–100% cover. Andrews (1950–1956) recoded S. fruiticosa from the Sudanese Red Sea littoral salt marshes.
3.3.4 Sand Dune Vegetation
Along the Afro-Asian Mediterranean and Red Sea coastal belts lie chains of sand dunes which, due to their proximity to the seas, are more humid and exposed to maritime influences such as sea-water spray. As rainfall is considerably higher along the Afro-Asian Med. coasts than those of the Red Sea, the fresh water of rain is usually stored in the dunes and is frequently obtained by digging carefully to a 3–4 m depth. Such fresh (rain) water, having a lower specific gravity than the saline water below, can form a layer above it. There may be a hard pan of limestone rock underlying the sand which prevents percolation of rain water, resulting in the sand dunes acting as reservoirs of fresh water.
Plants growing in the sand dune habitats (psammophytes) are highly specialized and can have the ability to elongate vertically on burial with sand (Girgis, 1973). They are also subjected to partial exposure of their underground organs often without being seriously affected. Many psammophytes develop extensive superifical roots that make use of dew.
Plant diversity of the sand dune vegetation of the Afro-Asian Mediterranean coastal lands is rather richer than that of the Afro-Asian Red Sea. This is, presumably, due to the relatively high amount of rainfall in the Mediterranean relative to the levels in the Red Sea coastal lands. The geographical distributions of species recorded from these coastal lands are mainly affected by rainfall and temperature. The psammophytes restricted to the North-African and SW Asian Med. coastal lands include: Ammophila arenaria, Lygeum spartum, Elymus farctus, Lotus polyphyllus, Thymelaea hirsuta, Lycium europaeum, Silene succulanta, Asparagus stripularis, Allium roseum, Convolvulus althaeoides, Pancratium maritimum, Phlomis floccosa, Salvia lanigera, Zygophyllum aegyptium (an endemic), Echium angustifolium subsp. sericeum, E. glomeratum Artemisia monosperma, Moltkiopsis ciliata, Scrophularia hypericifolia, Salsola kali, Cakile maritima, Ipomoea litteralis, Lotus creticus, Medicago marina, Stipagrostis scoparia (= Aristida scoparia), Lolium gaudini, Astragalus fruticosum (= A. tomentosum), A. spinosum, Euphorbia paralias, Prosopis farcta (= Lagonychium farctum), and Ziziphus lotus. All of these are absent from the Red Sea coastal lands. Psammophytes inhabiting both the Afro-Asian Med. and Red Sea coastal lands include: Cyperus conglomeratus, Retama raetam, Cutandia memphitica, Panicum turgidum, Pennisetum divisum, Eremobiun aegyptiacum, Echinops spinosissimus, Halplophyllum tuberculatum, Neurada procumbens, Deverra tortusa (= Pituranthos tortuosus), Cynodon dactylon, Stipagrostis plumosa, Alhagi graecorum, Tamarix aphylla and Zygophyllum album. Only two species are restricted in their distribution to the coastal sand dunes of the Afro-Asian Red Sea coastal lands, namely: Atriplex farinosa, and Halopyrum mucronatum. A. farinosa is widespread along the whole length of the African and Asian Red Sea coastal lands inhabiting the low sand dunes close to the sea. However, H. mucronatum has been recorded only from very limited areas of both Red Sea coasts: from Mersa Abu Ramad (33 km) north of Mersa Halaib at the Sudano-Egyptian border, from the Eritrean coast (African Red Sea coast) and from the Saudi Arabian coast at Jizan.
3.3.5 Rocky Ridge Vegetation
The rocky ridge is a unique geomorphological landscape of the western section of the North African Mediterranean coastal land in Libya and in Egypt. Embabi (2004) stated that, although the surface of the Mediterranean coastal plain in Egypt slopes gently towards the sea, the plain is characterized by a sequence of low carbonate ridges (bars) which are roughly parallel to the present coastline. These ridges are separated from each others by sabkha-lagoonal depressions interrupted in parts by secondary transverse bars. The number of these coastal ridges ranges between 4 and 8 reaching altitude of 10–110 m. All extend for long distances along the coastal plain parallel to the shoreline. The first (nearest the sea), and the second bars are almost continuous and prominent (El-Shazly and Shata, 1969).
Submerged ridges are also recognized in the offshore areas. These are composed of oolitic limestone, marl with shell fragments and calcareous sand, lying 0.71–15.79 m below sea level. Butzer (1960) recognized eleven ridges lying at up to 11 m below sea level and two others at 40 m below sea level some 6 km offshore.
The coastal ridges of the North African Med. Coastal lands are veneered with a hard brown calcareous duricrusts (calceretes) which are either exposed or covered by a thin layer of recent aeolian sand. The tops of these ridges are composed of hard oolitic limestone with horizons of palaeo-soil (Rashed, 1998). This may indicate that they have been deposited during a generally arid period with two relatively short periods of wet climate (El-Asmar and Wood, 2000).
El-Morsy (2008) and Zahran and Willis (2008) found that the rocky ridge vegetation in Libya and Egypt is formed mainly of elements restricted geographically to the Mediterranean region and absent from the Red Sea coastal lands, there are few species recorded from both coastal areas. The characteristic Mediterranean elements include: Thymus capitatus, Glaucium grandiflorum, Ononis natrix, Panicum repens, Asparagas aphyllus, Bellevalia fruticosum, Echium sericeum, Helianthemum peruviana, Lygeum spartum, Lycium europaeum, Thymelaea hirsuta, Salvia lanegera, Chenolea arabica (= Bassia arabica), Globularia alypum, Scilla peruviana, Reaumuria hirtella, Iris sisyrinchum, Stipa parviflora, Haloxylon scoparium, Noaea mucronata, Plantago albicans, Ephedra alata, Carduus getulus, Bupleurum nodiflorum, Arisarum vulgare, Aegilops kotschyi, Lotus glaber, and L. creticus. Apart from these, the flora of rocky ridge vegetation comprises species of wide geographical distribution and may grow in the other habitats of the Red Sea costal lands e.g. Deverra tortusa, Gymnocarpos decander, Aeluropus lagopoides, Echinops spinosus, Helianthemum lippii, Globularia arabica, Plantago albicans, Stipa capensis etc.
3.3.6 Desert Vegetation
The coastal desert vegetation is the most prominent and widespread type in the Afro-Asian Mediterranean and Red Sea Coastal lands. It predominates in desert areas extending between the inland borders of the littoral salt marshes and/or coastal sand dunes and the coastal plateaus of the Med. coasts or the ranges of hills and mountains of the Red Sea coasts on the inland side. Due to its intermediate position, the ecological features of the desert vegetation, especially on its fringes, show transitional characters.
The coastal desert vegetation is far away from maritime influences and has non-saline habitat but climate and soil aridity are the main environmental factors. It presents a complicated pattern owing to the difference in topography, characters of the surface deposits and the relationship with mountain groups. The habitat is essentially a gravel-covered plain traversed by the mid- and downstream extremities of the main wadis. The downstream parts of these wadis, particularly along the Afro-Asian Red Sea coastal lands and south Sinai can form deltaic basins.
Within these coastal desert ecosystem soil transporting agencies, mainly climatic (wind and rain), are actively operating. The alluvial deposits carried by rainfall range in particle sizes from silt to coarse gravels and boulders and often build compact terraces on the sides of the water courses (wadis). The building and destruction of these terraces are mainly physical processes independent of the vegetation. Aeolian deposits carried by wind, on the other hand, are sandy superimposed as sheets, mounds or hills of various heights and extent. These sandy bodies are usually built around plants (phytogenic) and their sizes seems to depend upon the plant speices.
Unlike the zonation of littoral salt marsh vegetation, coastal desert vegetation shows a mosaic pattern and distinct seasonal aspects due to the preponderant growth of therophytes (ephemeral, annuals, and biennials) after rain. These short-lived plants usually become dry and disappear vegetatively during the dry seasons but their seeds are dormant in the soil until the next rains when they germinate to give new seedlings. This aspect of seasonal phenology is essentially a climatic feature, characteristic of desert vegetation but not seen in the salt marsh plants. High soil salinity in the latter habitat prohibits the growth of most therophytes.
Desert vegetation contains the largest number of species, and the floristic composition of its communities is usually much more elaborate than the simple composition of the salt-marsh vegetation. Its framework is formed of perennial xerophytes associated with therophytes during the rainy seasons. Thus, the desert vegetation can be classified into two main types: ephemeral and perennial vegetation types.
-
(A)
Ephemeral vegetation type
This type include all short lived plants (therophytes) that appear in the deserts after rain. It indicates soil conditions that do not allow water to be stored over the year: soil westness is maintained during only part of the year. This may be due either to the scantiness of rainfall or also to surface deposits being too shallow. Green patches of ephemeral vegetation are a recurring phenomenon seen in the wadis and plains of the coastal deserts after the rains usually forming a mosaic of patches each of which may be dominated by one or more species. One type of ephemeral growth may recur in the same patch for several years, possibly due to the availability of seeds where parent plants have existed.
Depending on the growth form of the dominant (or most abundant) species, three types of ephemeral vegetation are recognized in Afro-Asian Red Sea and Med. coastal deserts (Zahran and Willis, 2008; Zohary, 1962; El-Morsy, 2008). One is dominated by succulent plants, a second by grasses and a third by herbaceous species. Zygophyllum simplex, Aizoon canariense and Spergula fallax are widespread succulent ephemerals in the Afro-Asian Med. and Red Sea coastal deserts. Trianthema crystallina and Tribulus pentandrus represent succulent ephemerals restricted in their distribution to Red Sea coastal deserts, whereas Mesembryanthemum spp. (M. nodiflorum, M. crystallinum) represent succulent ephemerals restricted to Med. coastal deserts. The tissues of these succulent plants can store water for use later in the growing season. They have shallow roots and survive throughout a season longer than the other ephemeral.
The vegetation of the ephemeral grasses is of special importance for the nomadic herdsmen for whom it is valuable pasture. Bromus fasciculatus, Eragrostis cilianensis, Schismus barbatus, Setaria verticilata, Stipa capensis and Stipagrostis hirtigluma, are among the widespread ephemeral grasses growing in both Afro-Asian Med. and Red Sea coastal deserts. Ephemeral grasses restricted to the Med. coastal deserts but absent from the Red Sea coasts include: Bromus diandrus, Dactyloctenium aegyptium, Eleusine indica subsp. indica, Eragrostis aegyptiaca, Hordeum spontaneum, Lolium rigidum, and Triticum aestivum whereas those recorded only from Afro-Asian Red Sea coastal deserts are represented by: Aristida mutabilis, Brachiaria deflexa, Cenchrus pennisetiformis, Digitaria ciliaris, Eragrostis ciliaris and Trichoneura mollis (= Diplachne arenaria).
The herbaceous ephemeral type of vegetation may be dominated by one of a great variety of species or can be mixed species with no obvious dominant. These plants, like the grasses, can provide valuable grazing sites. Within Red Sea coastal deserts, patches of ephemerals dominanted by one of the following species are recorded: Arnebia hispidissima, Astragalus vogelii subsp. vogelii, A. eremophilus, Ifloga spicata, Cleome brachycarpa, Plantago parviflora, Senecio flavus, and Tribulus pentandrus. The herbaceous ephemerals of the Afro-Asian Med. coastal deserts also contain very many species, such as: Arnebia decumbens, A. linearifolia, Asphodelus viscidulus, Astraglus boeticus, A. tribuloides var. tribuloides, A. mareoticus, A. annularis, Filago contracta, Malva sylvestris, M. aegyptia, Plantago squarrosa, Raptianus raphanistrum, Senecio vulgaris, Linum decumbens, Lythrum hyssopifolia, Erucaria crassifolia, Sinapis alba, S. arvensis subsp. arvensis, Brassica rapa, Lobularia arabica, Centaurium spicatum, Notobasis syriaca. Species recorded in both Afro-Asian Med. and Red Sea coastal deserts include: Asphodelus tenuifolius, Filago desertorum, Malva parviflora, Neurada procumbens, Shouwia thebaica, Senecio glaucus subsp. coronopifolius, and Tribulus terrestris.
-
(B)
Perennial vegetation type
Xerophytic perennial vegetation of the Afro-Asian Med. and Red Sea coastal deserts can be classified into two main subtypes: suffrutescentFootnote 6 and frutescent.Footnote 7 The vegetation of the suffrutescent subtype consists of two layers: an upper layer (30–120 cm) that includes the dominant species and a ground layer (< 30 cm) that includes associated therophytes and cushion-forming perennials such as Cleome droserifolia, Cucumis prophetarum subsp. prophetarum, Fagonia indica, F. mollis, Blepharis edulis etc. (Red Sea coastal desert), Astragalus alexandrinm, Fagonia bruguieri, and F. cretica, F. glutinosa (Med. coastal desert), and Citrullus colocynthis, and Polycarpaea repens (from both Med. and Red Sea coastal deserts). Frutescent perennial vegetation comprises three layers-the two of the suffrutescent subtype and a higher (< 120 cm) one including the dominant. This vegetation includes some trees more than 5 m such as Acacia raddiana and Balanites aegyptiaca.
-
I.
Suffrutescent perennial vegetation
Three units of different species are recognized in the suffrutescent perennial vegetation: succulent half-shrubs, grasslands and woody plants.
-
(a)
The succulent half-shrubs are represented by the following species: Anabasis articulata and Hammada elegans (= Haloxylon salicornicum) in both Med. and Red Sea costal deserts; Haloxylon scoparium, H. negevensis and H. persicum are restricted to the Mediterranean, whereas Zygophyllum coccineum and Z. decumbens only occur on the Red Sea coastal deserts.
-
(b)
Grassland perennial vegetation is represented by:
-
(i)
Perennial grasses recorded in both Med. and Red Sea coastal deserts: Panicum turgidum, Pennisetum setaceum, Saccharum spontaneum subsp. aegyptiaca, Lasiurus scindicus, Centropodia forsskaolii, Cenchrus ciliaris, Hyparrhenea hirta and Polypogon viridis.
-
(ii)
Perennial grasses recorded in the Med. but absent from the Red Sea coastal deserts include: Panicum repens, Saccharum spontaneum subsps. spontaneum, Sorghum halepense, Stipagrostis scoparia, S. lanata, Agropyron cistatum, A. obtusa, Dichanthium annulatum, Lolium perenne, Leersia hexanda, and Oryzopsis miliacea.
-
(iii)
Perennial grasses recorded from the Red Sea but absent from the Med. coastal deserts include: Pennisetum divisum, Stipagrostis acutifolia, S. raddiana, Dicanthium foveolatum, Dactyloctenium scindicum, Cenchrus setigerus, and Enneopogon desvauxii.
-
(i)
-
(c)
Woody vegetation is represented by shrublets with various distribution patterns. Some species are widespread and grow in both coastal desert areas such as: Zilla spinosa, Cornulaca monacantha, Calligonum comosum, Artemisia herba-alba, A. judaica, Achillea fragrantissima, Ephedra alata, Gymnocarpos decander, Iphiona mucronata, and Pulicarea crispa. Those restricted to Med. coastal deserts include: Achillea santolina, Peganum harmala, Ranunculus asiaticus, R. millefolius, Noaea mucronata, Echium angustifolium, E. glomeratum, and Ephedra aphylla, while those restricted to the Red Sea coastal deserts include: Launaea spinosa, Abutilon pannosum, A. fruticosum, Hibiscus micranthus, Cocculus pendulus, Aerva javanica, A. lanata, Periploca aphylla, Cadaba rotundifolia, C. farinosa, Solenostemma arghal, Commicarpus helenae, Fagonia indica, F. thebaica, and F. boulosii (an endemic, Boulos, 1995) etc.
-
II
Frutescent perennial vegetation
Details of this vegetation type are dealt with in Section 3.3.8.4 (scrubland vegetation) of this book.
-
II
-
(a)
-
I.
3.3.7 Mediterranean Steppe Grasslands
There are extensive areas of purely Mediterranean grasses in Morocco, Algeria and Tunisia, i.e. restricted only to the western section of this coastal land. This grassland is formed mainly of the tough tussocky grasse Stipa tenacissima, called: esparto by the Spaniards and alfa by the Arabs (Branigan and Jarrett, 1975). S. tenacissima is indigenous to the south of Spain and the western section of North Africa but absent elsewhere. It grows well in dry, sunny situations on the Algerian and Tunisian coasts. The main shoots can grow to a heights of 1 m and the leaves to 15–90 cm in length. When young, S. tenacissima serves as a feed for cattle and sheep, but after a few years’ growth it acquires great toughness of texture. On account of the tenacity of their fiber and their flexibility, the leaves have for centuries been employed in the making of such useful articles as baskets, sandals, ropes and mats. Today, they are used as raw material for paper making.
The vast alfa steppes of North Africa are associated with an abundant growth of undershrubs such as Artemisia spp. and Rhanterium spp. in addition to scattered trees of Pistacia atlantica. Lygeum spartum which is very common grass in the sand dune and rocky ridge habitats of the eastern section of the North African Med. coastal land is also a common associate of S. tenacissima. When aridity is combined with a high water table, creating saline flats, the salt tolerant bushes of Atriplex, Salsola and Suaeda species constitute an extensive formation.
3.3.8 Forests of the Afro-Asian Mediterranean and Red Sea Coastal Lands
In the Mediterranean basin, including the North African and SW Asian coastal lands, the term “forest” should not necessarily bring to mind an image of high, dense stands of trees with closed canopies. Mediterranean forests are highly diverse in their architecture, appearance and woody plant species composition (Blondel and Aronson, 1999). Mediterranean forests are highly varied in growth form, morphology, physiology and phenology of the dominant trees in each region. Four leaf types occur in varying combination. Firstly leaves may be sclerophyllous and evergreen leathery in texture and often spiny or prickly. A second group has laurel-like leaves that are somewhat softer and shiny but still evergreen, like the foliage in many tropical forest trees. Thirdly, they may be semi-deciduous and remain on their stems over winter, with reduced or terminated growth and photosynthesis. Leaves are not shed until spring when they are replaced by a new crop of leaves. Examples of such species are Quercus faginea and Q. infectoria. The fourth group has typically deciduous leaves such as predominate in northern temperate forest trees e.g. Carpinus, Corylus, Ostrya and Zekova species.
Mediterranean forests also differ in the structure or physiognomy they assume under human management. Example include forest where all conifers have been removed but not the oaks. The largest and most diverse evergreen sclerophyllous forest in the Mediterranean area today are the “lauriphyllous” forests, relicts of a now virtually extinct Tertiary flora previously widespread in southern Europe and Northern Africa. These forest include species of the tropical family Lauraceae (e.g. Apollonia barbujana, Laurus azorica, Persea indica and Octea foetens) as well as several endemic broad-leaved evergreen trees such as Arbutus canariensis, Myrica faya and Visnea mocanera. All of these trees share their broad-leaved schlerophyllous leaf shape with the laurel (Laurus nobilis) which still occurs widely throughout the Mediterranean Basin. At high altitudes, open formations of coniferous forest containing species of Abies, Cedrus, Cupressus, Juniperus and Pinus are found with an understorey of spiny shrubs such as species of Astragalus and Genista.
In the Red Sea basin, including the southern mountainous section of Sinai, the term forest may be applied to the open scrubland vegetation that characterizes the montane countries where the amount of the orographic rain is relatively higher than the annual rainfall of the surrounding coastal deserts. This amount of rain associated with suitable temperature enable woody xerophytic trees and shrubs of, for example Acacia spp., or Balanites aegyptiaca etc. to grow and dominate forming a vegetation form in the wadis and low altitudes of the slopes of the mountains. At still higher altitudes, where temperature is low, coniferous forest of Juniperus and Pinus may also occur.
The vegetation forms that may represent the forests of the Afro-Asian Mediterranean and Red Sea coastal lands are: broad-leaved evergreen forests, stunted woodland (matorral), coniferous forest and scrubland vegetation (Table 3.1). The geographical distribution and floristic composition of these forms may reflect the relationship between vegetation and climatic factors prevailing in these coastal lands.
3.3.8.1 Broad-Leaved Evergreen Forests
This type of forest lies entirely within the limits of olive tree cultivation and may be regarded as the most characteristic type of the Mediterranean vegetation. The chief trees are the evergreen oaks of various species, the most numerous being holm oak (Quercus ilex), the cork oak (Q. suber) and kermes oak (Q. coccifera). The last species (Q. coccifera) is the most widespread in the three western North African countries (Morocco, Algeria and Tunisia) and also occur in the Levant on the Palestine-Israel coast. Q. coccifera has more stunted appearance than other evergreen oaks and its timber is of little economic value. Unfortunately, most of the regional broad-leaved evergreen forests have been cut down and replaced by cultivated trees or by the land left derelict so that there are only a few representative vegetation areas with too little rain for agriculture; exceptions may occur where oaks are of economic value as in the cork oaks (Q. suber).
The broad-leaved evergreen forests of the North African Mediterranean coastal land are formed of widely spaced oak trees with dense mats of xerophytic woody shrubs, undershrubs and stunted trees in between such as brooms (Sarothamnus scoparius, Cytisus spp., Genista spp., Spartum spp., Leguminosae), rosemary (Rosmarinus officinalis, Labiatae), myrtle (Myrtus communis, myrtaceae), gorse (Ulex europeaus, Leguminosae), privet (Ligustrum vulgare, Oleocae), rockrose (species of Helianthemum, Tuberaria, Citrus etc.), and Laurel (Laurus mobilis, Apollonia barbujana, Laurus azorica, Persea indica and Octea foetens, Lauraceae) forming a ground vegetation and above them rise the taller figs (Ficus carica, F. alba), wild olive (Olea europaea) terebinth (Pistacia terebinthus) and the strawberry trees arbutus (Arbutus unido, A. canariensis, Murica faya and Visnea mocanera), while bulbous plants such as narcissi (Narcissus poeticus), tulips (Tulipa stylosa) and lilies (Lilium spp.) are scattered throughout the forest.
In certain coastal areas where summer drought is too intense or the soil too thin for major tree growth, oaks show stunted growth or they may disappear completely to be replaced by other woody trees such as wild olive (Olea europaea), locus tree (carob, Ceratonia siliqua) and mastic tree (Pistacia lentiscus).
3.3.8.2 Stunted Woodlands (Matorrals)
The stunted woodlands of the Mediterranean Basin is the undergrowth of broad-leaved evergreen forest which remains after the uncontrolled destruction of trees and/or depredations of goats causing degeneration (Branigan and Jarrett, 1975). This vegetation forms an extensive and virtually impenetrable tangle of woody, thorny shrubs and dwarfed twisted trees rising to a height of not more than 3 m. It covers vast areas of Morocco, Algeria, Tunisia and Libya (North African Med. Coast) as well as the Palestine-Israel area of the SW Asian Med. Coast. “Woodland is, generally, found in areas that are suitable for agriculture use, so that much of it has been converted into pasture or cleared for growing grains” (Dallman, 1998).
This stunted woodland has diverse names, with almost every region or country with its own name or names to designate the diverse local names: garrigue or gariga and maguis or macchia in France and Italy, xerovuni in Greece, matorral and tomillares in Spain, choresh or maquis in the Palestine-Israel region. In many countries, the term maquis (or mecchia) refers to the first major stage in forest degradation, followed by garrigue, phrygana or batha which are all of still lower stature and complexity than maquis. The distinction between garrigue and maquis is considered by many geographers and phytoecologists to depend on the substrate. Garrigues are said to occur primarily on limestone and include the full range of species associated with holm-oak (Quercus ilex), while maquis is reserved for those formations occurring on acid, siliceous soils. In addition to the cohort of species found on nearby garrigues, maquis includes such calciphobeFootnote 8 marker (indicator) species such as strawberry tree (Arbutus unido, A. canariensis) and heath family trees (species of Calluna, Erica etc.), as well as certain rock-roses e.g. Cistus ladanifer, lavenders (e.g. Lavandula vera), and other shrubs.
The most characteristic feature of the matorrals is that they include a fine-grained mosaic of almost all the growth forms recognized by plant ecologists. Their under storey is rich with the full range of plant life-forms of Raunkiaer (1934): therophytes (ephemerals, annuals and biennials), cryptophytes (perennial plants with buds below the soil), geophytes (plants with perennating organs buried in the ground e.g. bulbs, rhizomes, corms, etc.), chamaephytes (plants with resting buds slightly above the ground but less than 25 cm), hemicryptophytes (with their buds at the level of the ground) and pahnerophytes (trees, shrubs or vines whose perennating buds are more than 25 cm above the level of the soil).
Matorrals are dominated by shrubs with evergreen, broad and small, stiff and thick (sclerophyllous) leaves, an overstory of small trees may present and with or without an understorey of annuals and herbaceous perennials (Di Castri, 1981). Matorral has also been defined by Zohary (1962) as any sclerophyllous evergreen dense vegetation type that may attain 4–6 m in height. The prominent dominant species include various species of oaks, carobs and lentisk in addition to different species of Arbutus. Daphne, Laurus, Phillyrea, Myrtus, Rhamnus and Viburnum, all of which are sclerophyllous. The understorey of the matorrals of the North-African and SW Asian Med. Coastal lands where human pressure is higher, tends to have fewer shrubs and more hemicryptophytes (Blondel and Aronson, 1999).
Though matorrals appear predominantly evergreen, yet some of their woody plants are in fact winter-deciduous. Examples are the maples (species of Acer), and species of Quercus, Pistacia, and Rhus. The brooms are notably common in Morocco, and have the distinction of bearing evergreen stems that are photosynthetically active all year round. Most of these species in the so-called retamoidFootnote 9 group have small deciduous leaves that fall readily during drought.
Maquis, is the most widespread form of vegetation in the Mediterranean Basin with a Med. climate. It is highly varied with respect to dominant species and height of vegetation. Dallman (1998) classified maquis into two types: high maquis and low maquis. High maquis includes tall shrubs and small trees of varying height and favours shady slopes and sheltered locations. Dominant plants such as kermes oak (Quercus coccifera) and holm oak (Q. ilex) have dense branches and small, dark green, leathery, evergreen leaves. Strawberry tree (Arbutus unedo) and heather tree (Erica arborea) are also common. Interspersed among these are shrubs such as myrtle (Myrtus communis) and Spanish broom (Genista hispanica). Low maquis, consists mainly of evergreen shrubs including rock rose and oleander. Rock rose in the genus Cistus (Cistaceae) is a dominant aromatic shrub common in Morocco and Iberia. Cistus species bear large numbers of five-petalled, rose like flowers over a period of several weeks in the spring. Flower colours range from white to pink through violet and in most species last for no more than 1 day. Sage-leaved cistus (Cistus salviifolius) has white flowers and narrow leaves while C. crispus has pink flowers. Associated with Cistus is the genus Cytinus, a parasitic species that grows on the roots of Cistus. These low-growing plants have yellow, white or pale pink flowers with conspicuous orange or red-coloured bracts. Oleander (Nerium oleander Footnote 10) is a poisonous plant (Apocynaceae), with a milky sap., avoided by herbivorous animals. N. oleander is abundant in many wadis often shaded by plane trees (Platanus orientalis) in the eastern part of the Mediterranean. N. oleander is one of the dominant species growing on stony and gravelly banks or beds of permanent and ephemeral runnels in the mountains and plains of Palestine-Israel area, SW Asian Med. coastal lands (Zohary, 1962). Baumann (1993) found that oleander leaves are stuffed into mouse holes, the mice die after they nibble the leaves.
Zohary (1962) reported that maquis vegetation in Palestine-Israel Med. coast is represented by the following species: Genista shacelata, Olea europaea, Calycotome villosa, Pistacia lentiscus, Ceratonia siliqua, Quercus calliprinos, Q. ithaburensis, Crataegus azarolus, Rhamnus palaestina and Amygdalus communis.
Garrigne consists of low shrubs dotted overhills with intervening bare, stony or shady patches. The term garrigue is a widely used French term for the vegetation type, also called Batha in the Palestine-Israel Med. coast where it accounts for more than 40% of all hilly, upland terrain. Much of the garrigue has been heavily grazed for thousands of years (Shmida and Barbour, 1982). This may explain why many of the most prominent species of the present status of this vegetation type are spiny plants not attractive to grazing animals. They often form distinct cushion shapes, leaving rooms from bulbs and orchids in the intervening spaces. Among these cushion plants are: the Greek spiny spurge (Euphorbia acanthothamnos) and thorny burnet (Sarcopoterium spinosum) both native to the eastern coast of the Med. Basin. In SW Med. coast of Palestine-Israel area, the Mediterranean batha (garrigue) vegetation is represented by (Zohary, 1962): Asparagus aphyllus, Salvia triloba, Cistus villosus, Thymus capitatus, Poterium spinosum, Satureja thymbra and Teucrium polium.
Garrigue vegetation contains many plants producing two types of foliage referred to as seasonally dimorphic. Jerusalem sage (Phlomis fruticosa), for example, has relatively large and soft leaves which emerge in the winter rainy season. Later in spring and summer, a more drought-tolerant foliage is produced. Garrigue vegetation contains many aromatic shrubs that emit oils with a pungent odor particularly in the heat of day (Dallman, 1998). Plants of the mint family are common in the Mediterranean Basin e.g. rosemary (Rosmarinus officinalis) and Spanish lavender (Lavandula stoechas) both with blue to violet flowers. Jerusalem sage (Phlomis fruticosa) has very attractive bright yellow flowers. Lavandula stoechas is an aromatic plant growing wild on Palestine-Israel Med. coast but introduced elsewhere long ago as an ornamental plant. Its oil has long appealed to man, but in fact in nature discourages consumption by foraging animals.
Bulbs are most abundant in the garrigue of North Africa particularly in Morocco, including: narcissus (Narcissus bulbocodium, N. tazetta), crocus (Crosus biflorus), tulip (Tulipa doerfleri) and iris (Iris species).
The areas covered with maquis and garrigue in the North African and SW Asian Medi. coastal lands have been estimated by Dallman (1998), in million hectares, as follows: Morocco (5.2), Algeria (2.4), Tunisia (0.8), Libya (0.5), Palestine-Israel (0.1), Lebanon (0.1) and Syria (0.4).
3.3.8.3 Coniferous Forests
Coniferous means related to or belonging to the plant group Coniferae. Conifer is any gymnosperm tree or shrub of the group coniferae typically producing cones and evergreen leaves. This group includes the pines (Pinus spp.), spruce (Picea spp.), firs (Abies spp.) larches (Larix spp.), yews (Taxus spp.), juniper (Juniperus spp.) cedars (Cedrus spp.), cypressus (Cupressus spp.). All conifers belong geographically to the North Hemisphere in areas with cold climate latitudinally and altitudinally.
Conifers make their appearance in the region of the true Mediterranean climate where mountains are high enough and/or air temperature is low enough to enable conifer trees and shrubs to grow and reproduce. The lower limit of conifers varies in altitude with climatic conditions. On the west Mediterranean coast there is usually a zone of mixed deciduous coniferous trees from about 450 m upwards but in cooler areas e.g. the Balkan Peninsula, conifers may be found in lower altitudes of the mountainous slopes. One species of pines, the pinaster (Pinus pinaster), frequently occurs near sea level (Branigan and Jarrett, 1975). Coniferous forest are actually adapted to a low-temperature climate, the opposite to mangrove which never appears except in hot coastal swamps of tropical seas and oceans (see Section 3.3.1).
Three coniferous forests dominated by pines are recognized by Dallman (1998) in the Mediterranean Basin: Stone (or Umbrella) pine, Maritime pine and Aleppo pine, all commonly growing close to the sea.
-
(a)
Stone or umbrella pines (Pinus pinea) develop into an umbrella shape and most consistently grow up to 25 m high. It has been propagated since ancient times for its edible seeds called pignole, pignon or pine nuts. The seeds are bone on large, shiny, red-brown cones that take 3 years to mature, ripening in early spring to release seeds on very hot summer days or after a fire. Nuts are harvested by pulling the closed cones off the trees with a long hooked pole. One cone may contain 100 nuts. Dallman (1998) referred to P. pinea growing in the Iberian Peninsula, Italy, Greece and Turkey and Greco (1966) recorded it among the eight pine species of Algeria in the western section of the North African Med. Coast.
-
(b)
Maritime pine (Pinus pinaster) grows primarily in the western Med. coast from Morocco in the south to Spain, France and Italy in the north. It is more frost sensitive than the Stone pine and since the sixteenth century it has been Europe’s primary source of turpentine. P. pinaster trees have branches extending from the top third of a reddish trunk that reaches a height of 30 m. It has dark foliage and can form dense woods with an understorey of evergreen plants of the maquis such as tree heather (Erica arborea), strawberry tree (Arbutus unedo) and rock rose (Cistus monspeliensis). Plantings of P. pinasters have been used to stabilize sand dunes.
-
(c)
Aleppo pine (Pinus halepensis) is found along the hotter parts of the Mediterranean coastal lands mainly in North Africa (Morocco, Algeria and Tunisia) and SW Asia (in Palestine-Israel) coastal high lands (mountains). P. halepensis is the most drought tolerant, and is the most susceptible to fire of these three pines. It has a round or pyramidal crown, most common and most widely distributed pine near coasts. It is usually found in groves of scattered trees mixed with maquis or garrigue vegetation of lavender (Lavandula stoechas), rosmary (Rosmarinus officinalis), thyme (Thymus spp.) and rockrose (Cistus monspeliensis) growing in the intervening open places. Aleppo pine is rich in resin used to flavour wines.
Apart from the trees of Pinus species, the coniferous forests of the North African and SW Asian Med. Coastal lands includes trees of cupressus (Cupressus species), cedars (Cedrus species), juniper (Juniperus species) and fir (Abies species). For example, Greco (1996) recorded in the flora of the Algeria coastal mountains eight species of Pinus: P. burta, P. canariensis, P. coulteri, P. halepensis, P. insignis, P. pinea and P. radiate, six species of Cupressus: C. arizonica, C. atlantica, C. demakhar, C. dupreziana, C. glabra, and C. sempervirens, and one species of each of Juniperus (J. phoenicea) and Cedrus (C. atlantica). The coniferous forests of the Palestine-Israel and Jordan coastal mountains contain: Juniperus phoenica, J. oxycedrus, Cupressus sempervirens and Pinus halepensis (Zohary, 1962; Al-Eisawi, 1996).
Though most of the Afro-Asian Red Sea coastal lands, including the southern part of Sinai, are located within the latitudes of the tropical hot climate (29° 50′N to 12° 35′N, 43° 3′E), yet their vegetation forms comprise coniferous forest essentially in the higher altitudes of the coastal mountains where temperature is very low and suitable for the growth. For example, in the southern mountains of Sinai the highest peak “St Katherine” is up to 2,641 m above sea level and the air temperature in winter is usually below zero. In Gebels Halal and Yalaq of the southern Sinai coniferous forest occurs in Juniperus phoenicea trees associated with: Origanum isthmicus (endemic in Gebel Halal) Astoma seselifolium, Ephedra aphylla, E. campylopoda, Rubia tenuifolia and Cupressus sternbergia. Cupressus sempervirens has been recorded by Migahid et al. (1959) in Gebel Halal, but neither Täckholm (1974) nor Boulos (1995, 1999) recorded it in the flora of Egypt. However, it is well known that few Cupressus sempervirens trees have been cultivated for centuries in the gardens of the Monastry in St Katherine in south Sinai. Thus, the Cupressus shrubs recorded by Migahid et al. (1959) might not be natural, but might also have been cultivated around the natural spring known as AinFootnote 11 El-Goweirat of Gebel Hala. Today these plants are absent.
In the African Red Sea coastal mountains, the coniferous forests are not well developed. A few Juniperus procera trees have been recorded by Andrews (1950–1956) on the Red Sea coastal mountains of the Sudan. J. procera and other coniferous trees are absent from the Egyptian Red Sea coastal mountains (Täckholm, 1956; Boulos, 1995; Zahran and Willis, 2008). However the situation is different in the Asian Red Sea coastal lands. Dense coniferous forests characterize the south western mountains of Saudi Arabia (the Asir). Allered (1968) estimated about 25 million acres of coniferous forest at the tops of these mountains. The dominant coniferous trees are Juniperus procera and J. polycarpos with abundant growth of Cupressus sempervirens (Migahid and Hammouda, 1978). The highest altitude (about 3,000 m) on Al-Sudda summit, 25 km south of Abha city, represents the water shed zone of these mountains. In this zone, Juniperus trees establish a closed canopy with little understorey. Trees up to 17–20 m high and 70–85 years old have been recorded (Zahran, 1982b). At the lower altitudes (< 2,000 m), Juniperus trees still predominate associated with trees and shrubs of other species such as Olea chrysophylla, Pistacia palestina, Dodonaea viscosa, Commiphora spp., and Dracaena ombet., woody herbs, such as Psiada arabica, Eryops arabica, Lavandula dentata, and Euphorbia retusa, grasses, such as Themeda triandra, and Hyparrhenia hirta and succulents such as Euphorbia thi and Caralluma spp. (C. retrospeciens, C. pedicillata and C. sinaica). In zones lower than 1,000 m, where there are noticeable change in climatic conditions, mainly an increase in air temperature, Juniperus trees disappear and Acacia spp. (A. etbaica, A. nubica etc.) dominate. The associates are: Commiphora spp. (C. africana, C. opobalsamum, C. quadricincta), Grewia tenax, Olea chrysophylla, Ficus pseudosycomorus, and Adenium arabicus. Acacia asak, A. mellifera and A. etbaica scrubland are abundant in these lower zones associated with Themeda triandra, Artemisia judaica, Pulicaria crispa, Euphorbia cuneata, Scorzonera intricta, Aloe vera, and Euphorbia nubica.
In the coastal mountains, of Yemen, Wood (1997) stated that Juniperus procera is the only native conifer. It grows on the altitude ranging between 1,800 and 3,300 m. Although Juniper woodland is probably the natural climax there, its thick vegetation, which is such a characteristic feature of the Asir Mountains of Saudi Arabia, is not recognized in Yemen. This presumably is due to cutting and overgrazing through many centuries. In fact, coniferous forest throughout the Afro-Asian Med. and the Red Sea coastal lands have been and still are exposed to uncontrolled exploitation for many purposes e.g. building constructions, ship building, furniture, railway sleepers, harbour works and pulp and paper production. Many areas have been unfortunately, deforested with subsequent soil erosion. Projects aiming at conservation, rehabilitation and afforestation of these precious coniferous forest are badly needed.
3.3.8.4 Scrubland Vegetation
Scrubland is an arid land vegetation type formed mainly of woody trees, shrubs and undershrubs associated with some succulents, grasses and herbs, all are xerophytes. It is a characteristic ecological feature of the wadis and mountains of the Red Sea coastal deserts and the southern part of Sinai. Zohary (1962) recorded 60 tropical species of this vegetation from the Palestine-Israel coast. Boulos (1983) referred to a few medicinal woody trees and shrubs belonging to this vegetation form from North African countries.
The floras of the scrubland vegetation of the Afro-Asian Red Sea coastal lands comprise elements belonging to various tropical floras, most important among them are the Sudanian elements, such as species of Acacia, Phoenix, Balanites, Dracaena, Moringa, Salvadora, Hyphaene, and Caralluma.
The geographical distribution of scrubland vegetation form along the Afro-Asian Mediterranean and Red Sea coastal deserts and its altitudinal zonation on the coastal mountains has been discussed by many authors e.g. Tothill (1948), Kassas (1956, 1957, 1960), Ozenda (1958), Keay (1959), Hemming (1961), Zohary (1962), Greco (1966), Kassas and Zahran (1971), Pottier-Alapetite (1981), Boulos (1983, 1995, 1999, 2000, 2002, 2005), Konig (1986), Wood (1997), and Zahran and Willis (2008). These and other studies have clarified the relationship between climatic conditions and the latitudinal distribution of scrubland vegetation and its floristic elements. The altitudinal zonation pattern of this flora on the slopes of the mountains of the Red Sea also has been discussed. These are outlined below.
-
A.
Geographical distribution
-
a.
Acacia spp.
More than 10 species of Acacia are recorded in the Afro-Asian Mediterranean and Red Sea coastal lands, each has its own distribution pattern.
-
1.
Acacia raddiana (= A. tortilis subsp. raddiana) is a woody tree (> 12 m high) that seems to have wide ecological amplitude. It grows everywhere in the Afro-Asian Med. and Red Sea coastal lands but with varying abundance.
-
2.
Acacia pachyceras var. najdensis (= A. negevensis) is a tree 3–8 m high, restricted to the SW Med. coastal desert and south Sinai but absent elsewhere.
-
3.
Acacia tortilis (= A. tortilis subsp. tortilis) is a tree up to 5 m high, dominant in the wadis of Afro-Asian Red Sea coastal deserts, common in Sinai and Palestine-Israel desert but absent from the North African Med. Coast.
-
4.
Acacia laeta is a shrub or tree up to 5 m high, abundant in the Afro-Asian Red Sea coastal desert, common in the SW Asian Med. coast but absent from the North African Med. coast.
-
5.
Acacia ehrenbergiana is a shrub 1.5–4 m high, common in the Afro-Asian Red Sea coastal deserts including south Sinai but completely absent from Afro-Asian Med. Coast.
The other Acacia spp., are: A. albida (= Faidherbia albida), tree up to 18 m high, A. etbaica, a shrub or small tree, 4–6 m high, A. mellifera, a shrub or small tree, 6 m high, A. oerfota (= A. nubica), a shrub or small tree, 1.5–4 high and A. asak, a shrub or small tree, 6 m high. All of threse are restricted in distribution to the Afro-Asian Red Sea coastal deserts but absent from the south Sinai as well as from the whole of the Afro-Asian Mediterranean coastal lands. Acacia cyanophylla is a tree 5 m high, recroded by Migahid and Hammouda (1978) in the Red Sea coastal mountains of Saudi Arabia but recorded neither from Egypt, nor from Yemen. A. edgeworthii is a shrub 1.5 m high recorded by Wood (1997) from the stony steppe between 200 and 400 m altitude from the Tihama mountains of the Yemen coast but recorded neither in Saudi Arabia nor from the African Red Sea coast. However, A. albida and A. cyanophylla are known to be cultivated in Yemen (Wood, 1997). Kassas (personal communication) informed the author that A. cyanophylla and A. salegna are cultivated in Egypt, both are not African species but introduced from Australia.
-
1.
-
b.
Other woody and succulent species
Apart from the woody Acacia species, the scrubland vegetation of the Afro-Asian Red Sea coastal lands contains many other woody and succulent trees, shrubs and undershrubs, all are xerophytes. These species can be classified according to their geographical distribution into three groups: species restricted to the Afro-Asian Red Sea coastal deserts, species restricted to south Sinai, and (3) species growing in both regions.
-
1.
Scrubland species restricted to the Afro-Asian Red Sea coastal deserts and absent from south Sinai are mostly woody plants, with a few succulents. The woody species include: Commiphora africana (a shrub or tree), C. opobalsamum (a strong smelling shrub or small tree), C. quadricincta (an armed shrub), Dodonaea viscosa (a shrub), Dracaena ombet (a small stout tree growing at high altitudes in the mountains), Euclea schemperi (a shrub or small tree), Grewia villosa (a shrub), G. tembensis (a shrub), Sida ovata (a shrub), Cadaba farinosa (a shrub) C. glandulosa (a shrub), C. rotundifolia (a shrub), Zilla spinosa subsp. spinosa (a shrub), Boscia senegalensis (a shrub or small tree), B. angustifolia (a shrub or small tree), Aerva lanata (a shrublet), Maerua oblongifolia (a shrub), Solanum incanum (an undershrub), S. careens (an undershrub), Abutilon bidentatum (an undershrub), A. longicuspe (an undershrub), Senna holosericea (= Cassia holosericea, an under shrub) Rhus abyssinica (a tree or shrub), and Olea europaea subsp. cuspidata (= O. europaea subsp. africana, a tree or large shrub), Jasminum floribundum (a shrub or scrambler), J. fluminense (a climbing or scrambling, a shrub), Sterculia africana (a tree), and Cordia sinensis (= C. gharaf, a tree or shrub). The succulent species of this group include: Euphorbia thi (a shrub), E. cuneata (a shrub), and two Caralluma species, namely: C. acutangula and C. retrospiciens (both Cactus like bushes).
The flora of Yemen comprises the greatest number of Caralluma species (20 species) among the other floras of the countries of the Afro-Asian Red Sea coastal lands (Wood, 1997). The species recorded in the Tihama mountains include: Caralluma acutangula (= C. retrospiciens), C. commutata, C. deflersiana, C. penicillata (abundant in Acacia and Euphorbia scrub), C. plicatiloba, C. quadrangular (endemic to SW Arabian Peninsula) and C. subulata (endemic to SW Arabian Peninsula).
-
2.
Scrubland species restricted to south Sinai include: Capparis spinosa var. aegyptiaca (a shrub), Solanum sinaicum (an undershrub), Rhamnus lycioides subsp. graeca (a shrub) and R. disperma (a shrub) and two succulent undershrubs: Caralluma sinaica, C. europaea (= C. negevensis).
-
3.
Scrubland species growing in the Red Sea coastal deserts as well as in south Sinai include the following: Ficus palmata (F. pseudosycomorus, a shrub), Maerua crassifolia (a shrub or small tree), Rhus tripartita (a shrub), Salvadora persica (a shrub), Pistacia khinjuk (a tree), Balanites aegyptiaca (a tree), Ziziphus spina-christi (a tree), Tamarix aphylla (a tree or shrub), Lycium shawii var. shawii (a shrub), Leptadenia pyrotechnica (a shrub or small tree), Maerua crassifolia (a shrub or small tree), Hyphaene thebaica (a tree), Phoenix dactylifera (a tree), Calotropis procera (a shrub), Retama raetam (a shrub), Launaea spinosa (a shrub), Moringa peregrina (a tree or shrub), Grewia tenax (a shrub), Aerva javanica var. javanica (an undershrub), Senna italica (= Cassia obovata, an undershrub), S. alexandrina (= Cassia acutifolia, an undershrub) and Cocculus pendulus (a liane or shrub).
Species belong to the scrubland vegetation, both woody and succulents, have been also recorded in the Afro-Asian Mediterranean coastal lands. The flora of Palestine-Israel coastal lands comprises about 60 tropical species many of them belong to the scrubland vegetation form (Zohary, 1962). These include: Acacia spp. (A. pachaceras var. najdensis (= A. negevensis), A. raddiana, A. tortilis and A. laeta), Calotropis procera, Ziziphus spina-christi, Balanites aegyptiaca, Sebestena gharaf,Footnote 12 Grewia villosa, Maerua crassifolia, Abutilon pannosum (= A. muticum), A. fruticosum, Solanum incanum and Cassia obovata. Pottier-Alapetite (1981) reported few tropical species from the flora of Tunisia, namely: Acacia raddiana, Calligonum comsum, Capparis spinosa, Retama raetam and Tamarix aphylla. Ozenda (1958) recorded a large number of tropical genera and species from the inland North African desert, far from the Med. Coast. Most of these are elements of the scrubland vegetation of the Red Sea coastal deserts such as: Acacia albida, A. raddiana, Balanites aegyptiaca, Boscia salicifolia, Cadaba farinosa, C. glandulosa, Calligonum comosum, Caltropis procera, Capparis spinosa, C. galeata, C. decidua, Caralluma tombuctuensis, C. venenosa, Ficus carica, F. salicifolia, F. insens, Grewia populifolia, Hyphaene thebaica, Phoenix dactylifera, Leptadenia pyrotechnica, L. heterophyllum (= L. arborea), Lycium shawii (var shawii), Maerua crassifolia, M. angolensis, Olea laperrini (endemic), Panicum turgidum, Pistacia atlantica (endemic), Ziziphus mauritianus, Rhamnus lycioides, Salvadora persica, Tamarix aphylla and Zilla spinosa.
Interesting notes on Olea europaea and Caralluma species are given by Zohary (1962) and Boulos (2000). Olea europaea subsp. europaea var. europaea is widely cultivated in all countries of the Afro-Asian Med. coastal lands, whereas O. europaea subsp. europaea var. sylvestris grows naturally on the North African Med. coast (Boulos, 2000). Zohary (1962) recorded O. europaea var. oleaster in the carob-lentisk maquis vegetation of the SW Asian Med. coast of Palestine-Israel, mentioning also that the genus Caralluma is represented by two related endemic species and one endemic variety. C. negevensis is a Saharo-Sindian species, C. aaronis is an Irano-Turanian species, while C. europaea var. judaica is Mediterranean. C. maris-mortus (= C. sinaica) is an endemic species. He added “All the species of Caralluma are without doubt relics of the Tertiary period during which time tropical vegetation prevailed over most parts of Palestine”.
-
1.
-
a.
-
B.
Altitudinal zonation
Altitudinal zonation is obvious in the scrubland vegetation of the Red Sea coastal mountains of Egypt, the Sudan and Saudi Arabia.
-
(a)
Red Sea coastal mountains of Egypt
The range of mountains extending along the Egyptian Red Sea coast can be classified under two main sections: 1. the mountains of the northern section facing the western coast of the Gulf of Suez from Suez (27° 20′N) to Hurghada (27° 14′N) and the mountains of the southern section facing the western coast of the Red Sea proper from Hurghada to Mersa Halaieb (22°N) at the Sudano-Egyptian border. The mountains of the northern section are (from north southwards): Gebel Ataqa (817 m), Gebel Kahalya (660 m), Gebel Akheider (367 m), Gebel El-Galala El-Bahariya (700 m), Gebel El-Galala El-Qiblya (1,200 m), Gebel Abu Dokhan (1,705 m), Gebel Qattar (1,963 m), Gebel Shayeb El-Banat (2,187 m) and Gebel Umm Anab (1,782 m). Gebel Shayed El-Banat is the highest of these peaks extending some 40–50 km to the west of Hurghada and representing the southern part of the Gulf of Suez and NE part of the Red Sea proper (Zahran and Willis, 2008).
The mountains of the southern section are the Gebel Nugrus group (24° 40′N to 24° 50′N) including: Gebel Migif (1,198 m), Gebel Zabara (1,360 m), Gebel Nugrus (1,504 m) and Gebel Hafafit (857 m), Gebel Samiuki group comprises three main blocks: Gebel Abu Hamamid (1,786 m), Gebel Samiuki (1,283–1,486 m), and Gebel Hamata (1,977 m) and Gebel Elba group on the Sudano-Egyptian border (22°N) and comprises Gebel Elba (1,428 m), Gebel Shindeib (1,911 m), Gebel Shindodai (1,526 m), Gebel Shillal (1,409 m), Gebel Makim (1,871 m) and Gebel Asotriba (2,217 m). Gebel Elba is particularly favoured by its position near the sea. The richness of the plant life of Gebel Elba area is so notable that it is considered to be one of the main phytogeographical regions of Egypt (Täckholm, 1974).
Plant life of the Egyptian Red Sea coastal mountains depends on a combination of factors: (i) expanses of water body traversed by wind before reaching the coast, (ii) distance from the shore line to the mountains and the topographic features of the stretch, (iii) altitudes and (iv) exposure. Plant life on mountains (scrubland vegetation) facing limited body of water of the Gulf of Suez (i.e. northern section), indicate an arid climate. By contrast, plant life on the Elba group indicate a less arid climate and present an example of a mist oasis. Altitudinal zones of vegetation are recognizable on the slopes of the higher mountains differences between the plants of seaward and of leeward slopes are obvious (Kassas and Zahran, 1971).
The effect of topography on precipitation is a universal phenomenon but is more pronounced near the coast. Though located in arid regions, the Red Sea coastal mountains, particularly the Gebel Elba group, can cause ample orographic precipitation. The northern blocks of mountains receive lesser water from orographic rain. Such differences in the amounts of rainfall have their effects on the plants of these mountains. In terms of the number of species recorded from the four mountain groups it was found that: 53 species from the Gebel Shayeb El-Banat group, 92 species from the Gebel Nugrus group, 125 species from the Gebel Samiuki group and 458 species from the Gebel Elba group. Though it is not the highest, Gebel Elba is the nearest to the sea (20–25 km) facing NE bend of the shore in such a manner that it faces in northward direction an almost endless stretch of water. In contrast, Gebel Shayeb El-Banat faces only a narrow stretch of water (the mouth of the Gulf of Suez). The amount of orographic rain is rather higher on Gebel Elba than that on Gebel Shayeb El-Banat.
In Gebel Elba, plant growth on the slopes of different exposures are obvious: north and east slopes have richer plant growth than south and west slopes. The scrubland vegetation on north slopes varies in relation to altitude, but on south slopes there are few differences except perhaps between the more or less barren slopes and the vegetated runnels that dissect them. Four main altitudinal zones are distinguished on the north and east slopes of Gebel Elba (Kassas and Zahran, 1971):
-
(a)
a basal zone of Euphorbia cuneata scrub,
-
(b)
a middle zone of Euphorbia nubica scrub,
-
(c)
a high zone of Acacia etbaica scrub and,
-
(d)
a tope zone with a patchwork variety of plants.
The altitudinal limits of these zones are not fixed by absolute levels but related to a combination of altitude, degree of slope, air temperature, wind velocity and other climatic and physiographic features. The fourth top zone comprises stands of Dracaena ombet, Dodonaea viscosa, Delonix elata, Euclea schimperi, Rhus abyssinica, Ficus salicifolia and Pistacia khinjuk. Within this zone, ferns and bryohytes also abound e.g. Adiantum capillus veneris, Actiniopteris australis, Fumaria pallenscens, Riccia spp., and Mannia androgyne. This may indicate that the plants growing at the top of Elba do not belong to the arid climate regions as they comprises a great variety of species of wet areas with high water requirements. On the south and west slopes of Gebel Elba, plants growing are mostly confined to runnels with an open scrubland type dominated by Commiphora opobalsamum. At the mountain base the runnels are mostly dominated by Acacia tortilis scrublands (for details please see section “In Egypt”, II. Coastal mountains of Chapter 2).
-
(b)
Red Sea coastal mountains of the Sudan
The Red Sea coastal mountains of the Sudan can be represented by the Erkwit Plateau (19°7′N, 37°20′E, Alt. 1,080 m). Erkwit is a deserted summer resort, lies at about 45 km to the south-west of Suakin on the Red Sea coast and about 30 km to the east of Sinkat on the inland plain at the edge of a steep escarpment dropping abruptly to the Red Sea plains. At its northern boundary are Gebel Nakeet (1,176 m) and Gebel Essit (1,143 m), these two mountains drop to Khor Dahand which separates Erkwit from the barren hills on the other side of the Khor. At the eastern boundary is Gebel Sela (1,273 m) the highest evergreen mountain of the district. At the southern boundary are Gebel Tatasi (1,190 m), Gebel Lagagribab (1,209 m), and Gebel Auliai (1,191 m) separated by Khor Amat from the barren mountain further south (Gebel Erbaba, 1,923 m). At the eastern boundary are Gebel Hadast (1,147 m) and Gebel Mastiokriba (1,113 m) which also drop to Khor that separates the Erkwit Plateau from the desert plain to its east (Kassas, 1956).
Erkwit receives a rainfall (218 mm) greater than the neighbouring areas, as compared to data from two stations: Suakin (181 mm) to the NE and Sinkat (127 mm) to the west. Suakin, a Red Sea port, represents areas with winter rainfall. Sinkat, which lies on the inland plain, represent areas with summer rainfall. Erkwit lies in between and receives both the summer and winter rainfalls. Tothill (1948) stated that the climate of Erkwit may be due to the happy combination of: latitude, situation and elevation. Its latitude is close to 19°N which divides the Sudan into an arid desert region to the north and a tropical continental region to the south (Ireland, 1948). In the north prevails the dry northerlies while in the south the southerlies bring summer rainfall. Erkwit enjoys the maritime modification of the Red Sea: as the continental northerlies pass over the warm water of the sea they absorb a considerable amount of moisture. Being situated on a westerly bend of the Red Sea, Erkwit is exposed to about 650 km of open sea in the direction facing the northerlies (the N.E. Trades). The moisture-laden wind meets no dissipating obstacle before impinging on the cool hills of Erkwit and orographic precipitation consequently occur. After passing over the Erkwit Plateau the northerlies resume their dry continental characteristics. During the winter months, the Erkwit Plateau is frequently swathed in clouds for weeks. This entails considerable dew precipitation which is more marked the higher the elevation, and which supplies the vegetation with a valuable water resources (Kassas, 1956).
The vegetation of Erkwit has a trizonal pattern: an arid, a transitional and a moist zone (Andrews, 1948). However, Kassas (1956) point out the features of five vegetation zones dominated and co-dominated by: Maytenus senegalensis (zone I), Maytenus senegalensis – Euphorbia abyssinica (zone II), Euphorbia abyssinica (zone III), Dracaena ombet – Euphorbia abyssinica (zone IV) and Euphorbia thi (zone V). Such zonation is actually a climatic phenomeon related to the wetness and dryness of the zones (for details of these five altitudinal zones please see section “In the Sudan” of Chapter 2).
The relationship between the local climate and vegetation is obvious in the Erkwit Oasis, deduced from the following (Kassas, 1956):
-
1.
The total plant cover varies from 70% in particular habitats of the wettest zone I to only 5% in the driest zone (zone V),
-
2.
The distribution of ferns, mosses and liverworts is limited to zones I and II evidence for interzonal differences in water resources available to the roots, Atmospheric moisture is much greater in zone I. gradually decreasing downwards in zones III, IV and V.
-
3.
Vegetation is denser on the seaward slopes of the mountains than the leeward ones. This shows, again, that the atmospheric humidity (mist and wind-born water) is an effective water resource.
-
4.
Generally, the conditions that made the limited mountainous area of Erkwit into an oasis with thick vegetation and a wide range of plant diversity amidst arid country are: firstly, the combination of summer rainfall of the territory to its west and winter rainfall of the Red Sea to its east; secondly, it receives sea-mists and wind-born moisture which face no obstacle until they meet the edge of the Erkwit plateau.
-
5.
Local differences in physiographic factors, altitudes and distance from the edge of the escarpment cause the zonal pattern of the vegetation due to difference in the amount of available water.
-
1.
-
(c)
Red Sea coastal mountains of Saudi Arabia
The altitudinal zonation of the vegetation of the Saudi Arabian-Asian Red Sea coastal mountains is described in details by Konig (1986) in six locations: Tanuma, Gabal Sauda-ad-Darb, Slope of Hilly Tihama, Gabal Sauda, Al-Ulayya-Bisa, and draining runnels and wadis.
A short note on the zonation pattern of Al-Ulayya-Bisa region is given below.
The mountains of Al-Ulayya-Bisa region reach on altitude of 2,200 m. and the escarpment to the Red Sea is comparatively moderate so that rain shadow effects along the eastern slopes are not so marked (mean annual = 137 mm) as compared with that of Gabal Sauda, with an elevation up to 2,650 m, being 559 mm/year. Here, five zones of vegetation are recognized depending upon elevation: 2,200 m (Juniperus woodland), 2,000 m (Olea sclerophyllous scrub), 1,800 m (Acacia gerrardi xerophytic scrubland), 1,600 m (semi-desert) and 1,400 m (Acacia-Commiphora xerophytic scrubland). The Juniperus woodland of the higher altitude include: Juniperus excelsa, Euryops arabicus, Euphorbia schimperiana and Cultia richardiana. Below 2,100 m, Acacia gerradii becomes more characteristic of the vegetation, beginning with Juniperus-Acacia gerrardii woodland with Cultia richardiana, Sageretia thea ssp. thea and Dodonea viscosa. This community is replaced by Olea-Acacia gerrarrdii woodland below 2,000 m with characteristic species: Acacia gerrardii, Olea europaea spp. africana, Barbeya oleoides, Pistacia chenensis ssp. falcata, Jasminum floribundum, Euclea schemperi, Ficus salicifolia, Psiada punctulata, Carissa edulis and Hypoestes forsskaolii. With increasing distance from the watershed, only Acacia gerrardii xerophytic scrubland occurs, which is replaced below 1,850 m by semi-desert vegetation characterized by scattered dwarf shrubs such as: Teucrium yemense, Campylanthus pungens, Pulicaria crispa, P. somalensis ssp. somalensis, Heliotropuim strigosum, Morettia canescens, Crotalaria emarginella, Salvia aegyptiaca, Lavandula pubescens, Aerva persica, Farsetia longisiliqua, Fagonia bruguieri, Indigofera spinosa, Periploca aphylla, Otostegia fruticosa, and Lycuim shawii, in addition to grasses such as Eneapogon schimperianus, Tetrapogon villosus, Danthoniopsis barbata and Eneapogon brachystachyus. A particularly conspicuous species is the highly endangered Dracaena ombet found between 1,850 and 1,700 m where it grows in a number of places on the NW slopes. Below 1,600 m, Acacia – Commiphora xerophytic scrubland with A. asak, A. hamulosa, A. tortilis, Commiphora myrrha, C. opobalsamum and Euphorbia cuneata occurs.
-
(a)
Notes
- 1.
Temperate–having climate intermediate between tropical and polar; moderate or mild in temperature.
- 2.
Khamasin is the Arabic for fifty.
- 3.
Monsoon = A seasonal wind of south Asia that blows from the SW in summer bringing heavy rains and from the north east in winter. The rainy season where the SW monsoon blows is April–October.
- 4.
Buttes = isolated steep-sided flat topped hills.
- 5.
El-Mallaha is the Arabic wording for the highly saline land.
- 6.
Suffrutescent = undershrubby.
- 7.
Frutescent = shrubby.
- 8.
Calciphobe = Chalk – hating plants, plants shunning chalk or limestone.
- 9.
Retama raetam is a soon leafles desert shrub (Täckholm, 1974).
- 10.
Nerium oleander has been introduced to Egypt as an ornamental plants but never seen naturally growing in Egypt’s Med. Coastal land.
- 11.
Ain is the Arabic for spring.
- 12.
Prof. L. Boulos informed the author that Sebestena gharaf could be Cordia gharaf (C. sinensis).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2010 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Zahran, M. (2010). Climate–Vegetation Relationships: Perspectives. In: Gilbert, F. (eds) Climate - Vegetation. Plant and Vegetation, vol 4. Springer, Dordrecht. https://doi.org/10.1007/978-90-481-8595-5_3
Download citation
DOI: https://doi.org/10.1007/978-90-481-8595-5_3
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-90-481-8594-8
Online ISBN: 978-90-481-8595-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)