
Abstract
The most predominant opportunistic fungal pathogens Candida sp, Aspergillus and Cryptococcus sp are emerging as a global problem of great concern as it is associated with increased morbidity and mortality. Although there are different classes of approved antifungal agents like polyenes, pyrimidines, azoles, and echinocandins, the morbidity and mortality rate due to systemic infection remains high, due to its toxicity, narrow spectrum of activity and tolerability. The continuous usages of these drugs are associated with resistance development and thus this is another area of emerging global problem. In addition, biofilm formation by fungal pathogens are proven to be an important virulence factor and are resistant to host defense mechanism and antifungal agents. Combination therapy has shown to be a promising choice to overcome the emergence of drug resistance, minimizing the toxic effects by reducing the drug dosage, and to increase the spectrum of killing. In this chapter, we have reviewed the last 5 years’ developments made in combinational antifungal therapy with special emphasize on biofilm related infections and have presented a reappraisal of the current practices associated with immunotherapy to treat fungal infections and future challenges.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
6.1 Introduction
The improvement in medical advances incidentally has led to an increase in the incidence of opportunistic fungal infections. The most predominant opportunistic fungal pathogens are Candida sp., Aspergillus sp., and Cryptococcus sp. (Paramythiotou et al. 2014). During the last decade, there has been an emergence of other pathogens like Zygomycetes, Fusarium, etc., (Pagano et al. 2011). These pathogens take the chance to cause disease when the immune system of an individual is altered due to a plethora of reasons like organ transplantation, cancer therapies, diabetes mellitus, immature birth, dietary factors, and nutrient deficiency (Riddell and Shuman 2012). The other contributing factor for the increase of fungal infections includes surgical problems with prolonged antibiotic treatment (Miceli et al. 2011).
Although there are different classes of approved antifungal agents like polyenes, pyrimidines, azoles, and echinocandins, the morbidity and mortality rate due to systemic infection remains high, due to its toxicity, narrow spectrum of activity, and tolerability (Bal 2010). The continuous treatment with similar kind of drugs evolves different drug resistance mechanisms in fungi, and the emergence of drug-resistant strains is a growing global problem (Allen 2010). The common drug resistance mechanisms established in fungi are (I) transport alteration (ATP-binding cassette transporters or major facilitator superfamily transporter), (II) target alteration (by mutation or structural change in target protein), (III) gene upregulation (overexpression of target protein) and utilization of compensatory and catabolic pathway, (IV) biofilm formation, (V) efflux of antifungal drug, and (VI) decreased uptake of antifungal drug (Kontoyiannis and Lewis 2002; Sanglard et al. 2009).
Combination therapy has shown to be a promising choice to overcome the emergence of drug resistance, minimizing the toxic effects by reducing the drug dosage, and to increase the spectrum of killing. The drug combinations are always selected based on the two different modes of actions of the antifungals. These are inhibitors of cell wall synthesis and inhibitors of protein/DNA synthesis or inhibitors of the cell membrane. The drug interactions in the combinational therapy are classified as synergistic, antagonistic, and additive (Chen et al. 2014). Some proposed mechanisms for synergistic drug interactions are inhibition of biochemical pathway at different stages, for example, ergosterol biosynthesis and weakening of the fungal cell membrane by combinational drug therapy using terbinafine and azoles (Barchiesi et al. 1998; Johnson et al. 2004); increased penetration of combined drugs by impairing the cell wall or cell membrane activity, for example, combination of fluconazole or amphotericin B with quinolones and rifampin allows these antibacterial agents to penetrate fungal cell to target DNA synthesis (Medoff 1983); transport interaction of antifungal drug with the help of another antifungal drug, for example, the cell membrane degradation by amphotericin B via oxidative decay allows flucytosine into the cell (Beggs and Sarosi 1982); and instantaneous inhibition of multiple fungal cell targets (Bartizal et al. 1997). Antagonism among antifungal agents might occur in one of several ways. Administration of antifungal drug at the same site and time will decrease the efficacy of drug, for example, amphotericin B and azole: amphotericin B binds with ergosterol in the cell membrane and usually blocks the synthesis of ergosterol (Cosgrove et al. 1978). Adsorption of one antifungal agent to the cell surface inhibits binding of another antifungal agent to target site of activity, for example, itraconazole and ketoconazole are lipophilic azole that bind with the cell surface and inhibit binding of amphotericin B (Johnson et al. 2004). Modification of a drug target upon previous exposure to an antifungal agent reduces the susceptibility, for example, exposure of pathogen to an azole compound results in alteration of membrane ergosterol with methylated sterol derivative, which will reduce the activity of amphotericin B (Currie et al. 1995). The selection of drugs in combination should accomplish these criteria for better treatment option.
Considering these facts, the therapy should also incorporate the appropriate dosage for eradication of fungal biofilms. Biofilm formation is an important contributing factor for virulence and an emerging global problem of great concern. The biofilms are resistant to host defense mechanism and antifungal agents (Martinez and Fries 2010). The three main drug resistance mechanisms proposed in biofilms are (I) incomplete or slow penetration of antimicrobials due to exopolysaccharide polymers; (II) resistant phenotype, known as persister cells characterized by a slower growth rate and greater resistance to antimicrobials; and (III) an altered environment that may affect the antibiotic action such that fungal biofilm persists even when appropriate antifungal treatment is given (Chatzimoschou et al. 2011). For instance, fluconazole and amphotericin combination therapy was unsuccessful in the treatment of Cryptococcus neoformans-infected hip joints due to poor penetration of drugs through the biofilm (Johannsson and Callaghan 2009). Catheter-related fungal infections are serious infectious complications that worsen the patient outcomes. The guidelines of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID) have suggested removing the catheter, but a recent report, by analyzing 842 patients from two randomized clinical trials, reveals that the removal of the catheter does not improve the patient outcome (Nucci et al. 2010). Hence, development of new strategies for fungal infection management is required. One such approach is identifying an antiquorum sensing molecule which comprises targets of biofilm, virulence factors, and hyphal formation. The most studied quorum sensing molecule in fungi is farnesol, which has a control over growth, morphogenesis, and biofilm formation. The fungus quorum sensing research is still in infancy, and further research could lead to the development of new antifungal agents (Albuquerque and Casadevall 2012). Immunotherapy is based on the premise that most opportunistic fungal infections are a result of compromised immune system. And thus, any means of modulating the immune system using immune or immunogenic components could prove to be useful in achieving immune activation in the patient that will enable pathogen clearance. Such strategy minimizes the toxic effects of conventional antifungal drugs (and in some cases even synergizes with the drug or potentiates the activity of the drug), thereby opening up a new way of combating multidrug-resistant fungal pathogens.
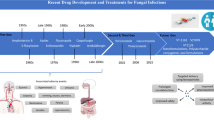
In this chapter, we have reviewed the last 5 years’ developments made in combinational antifungal therapy with special emphasis on biofilm-related infections and have presented a reappraisal of the current practices associated with immunotherapy and future challenges.
6.2 Cryptococcus Meningitis
Cryptococcus meningitis (CM) is an opportunistic neurological infection in immunocompromised individuals. It has emerged as the leading cause of morbidity and mortality in HIV-positive and HIV-negative patients (Muzoora et al. 2012). Recently, the Center for Disease Control estimated approximately one million new cases of cryptococcal meningitis each year, resulting in the death of 625,000 people worldwide. The standard therapy for CM consists of three phases: induction, consolidation, and maintenance (Vaidhya et al. 2015). The current standard initial therapy consists of amphotericin B and flucytosine (5-FC), but the availability of 5-FC is an issue in the countries of Asia and Africa, where CM is common (Perfect et al. 2010). Hence the probability of effective antifungal combination therapies is being investigated by the researchers.
The primary objectives of the therapies are to reduce the high infectious dose in the lungs and brain tissues which in turn increase the intracranial pressure and tend to have a worse prognosis. The induction therapy with amphotericin B deoxycholate (0.7–1.0 mg/-kg) combined with flucytosine (100 mg/kg/day for 2 weeks), followed by further consolidation therapy with fluconazole, is considered a standard management regime in the developed world (Denning and Hope 2010). An alternate but promising treatment strategy includes administering amphotericin B and fluconazole (both at 800 mg/14 days), followed by administration of fluconazole alone (800 mg/56 days). However, validation of this strategy is required through randomized phase III clinical trial. Fluconazole resistance is not common if it is used as a part of amphotericin B induction therapy. But continuous use of fluconazole has been shown to result in other resistant yeast infections, for example, fluconazole resistant (Govender et al. 2013). Toward the end of these antibiotic therapies, protein abnormalities may persist for years, which would cause adverse effects including nephrotoxicity (Fine and Atta 2007). Hence the combination therapy of fluconazole and flucytosine was recommended and has greater antifungal activity compared with fluconazole monotherapy, even though fluconazole has high penetration in CSF (Yao et al. 2014). However, the effect of this combination in comparison with amphotericin B is unknown (Denning and Hope 2010). Although the lipid-based formulations of amphotericin B have excellent efficiency by reducing its nephrotoxicity, the major drawback is its cost. To execute significant measures to overcome drug resistance, few studies have attempted to search for an alternative antifungal drug (Nikapitiya 2012). In search of new antifungal agents, in vivo efficacy of miltefosine, alkylphosphocholine, was evaluated for its usage as monotherapy and in combination with fluconazole and amphotericin. The monotherapy does not significantly improve survival of mice. Combination with miltefosine revealed neither synergy nor antagonism in both models, suggesting caution with the utility of miltefosine for anticryptococcal therapy (Wiederhold et al. 2013).
Apart from cost, the lipid formulation of amphotericin B is unable to pass through the blood-brain barrier. In CNS fungal infections, the permeability of lipid amphotericin B is enhanced by the inflammatory process. However, the inflammation is associated with the risk of intracranial pressure. Hence drugs directly administered into the CNS could be a suitable therapy for CN infections, especially in those cases where conventional therapy fails or a relapse of the infection occurs. Gazzoni et al. (2012) compared the in vivo efficacy of intrathecal administration of L-AmB alone and in combination with voriconazole in a murine model of cryptococcal meningitis. The animals were treated with liposomal amphotericin B administered intrathecally at 0.006 mg/kg weekly or intravenously at 10 mg/kg daily or with voriconazole administered orally at 30 mg/kg per dose twice daily or with combinations of both drugs, i.e., L-AmB i.t.c. + voriconazole or L-AmB i.v. + voriconazole at the same doses as used in the monotherapies. All treatments significantly increased the survival and reduced fungal burden in comparison with the control group, with voriconazole being less effective in comparison with all other treatments. The combination of L-AmB i.t.c. + voriconazole showed a synergistic effect in the reduction of fungal load that was significantly superior to any tested therapy (Gazzoni et al. 2012).
The increase of fungal burden in the CSF is a major issue to be focused to increase the survival. Studies were conducted to determine the therapeutic value of amphotericin B and voriconazole (alone and in combination) that would support reduction of fungal burden. Preclinical studies were conducted using BALB/c-SCID mice infected with fluconazole-resistant strains of Cryptococcus neoformans var. grubii. The survival curves and yeast quantifications indicate that amphotericin B was effective in increasing the survival of the animal without reducing the fungal burden in the lung and brain tissue (Silva et al. 2011). The rate of clearance of C. neoformans from cerebrospinal fluid is facilitated by amphotericin B and flucytosine as first-line therapy. But flucytosine is not available in many resource-limited countries (Bisson et al. 2013). Thus, combinations with different modes of action and different doses were tried with AmB + fluconazole (800 mg/day), AmB + fluconazole (600 mg twice per day), and AmB + voriconazole (300 mg twice per day) that provides a means to compare the antifungal activity of alternate regimens in small phase II studies. The combination therapy comprising rifampin and amphotericin B for treating fungal diseases is of past interest to eradicate the toxic effect of conventional AMB. In vivo studies were not conducted due to the lack of laboratory testing standards, of Cryptococcus sp. (Loyse et al. 2012).
Gamaletsou et al. (2012) experimented salvage therapy for two patients with HIV-related, refractory Cryptococcus meningitis as there are no potential comparative studies for salvage therapy. The salvage regimen provides a successful treatment, consisting of the combination of liposomal amphotericin B (3 mg/kg), intravenous voriconazole, and subcutaneous recombinant interferon γ-1b (200 μg thrice weekly). And to overcome interactions with co-administered ritonavir, voriconazole was given at an increased dose (5 mg/kg, twice daily). Oral fluconazole was given as maintenance therapy. The salvage therapy eliminates the fungal burden in the CSF after 10 weeks of therapy, also no adverse side effects, and no recurrence of infections has been observed even after 4 years of follow-up. Further validation is needed to use this strategy as initial therapy for HIV-related CM.
Recently, Yao et al. (2014) conducted a meta-analysis for prospective cohort studies to compare the survival benefits between fluconazole and 5-flucytosine given in combination with AmB as the initial therapy for CM. The survival rate was similar in case of AmB in combination with high-dose fluconazole, and therefore this combination is alternative for 5-flucytosine for the management of HIV-associated CM therapy, which is not available in resource-limited countries (Yao et al. 2014). Clinical data disclose that high-dose fluconazole shows no adverse liver function abnormalities (Vaidhya et al. 2015). However, high-dose fluconazole might lead to the emergence of drug-resistant strains. For patients without CNS involvement with limited pulmonary cryptococcosis and non-pulmonary cryptococcal infections in immunocompetent patients, initial therapy with fluconazole (400 mg per day) could be considered. The standard therapy of amphotericin B and flucytosine is not recommended for disseminated cryptococcosis (Probst et al. 2010).
Cryptococcus neoformans biofilms are becoming common due to the increased use of brain valves and other medical devices such as ventriculoatrial shunt catheters, peritoneal dialysis fistula, cardiac valves, and prosthetic joints. The knowledge on the mechanism of biofilm formation by Cryptococcus is still in infancy; hence, there are very limited reports on the evaluation of antifungals on C. neoformans biofilms. An in vitro study reveals that amphotericin B and caspofungin are effective against the mature biofilms of C. neoformans; however, the efficacy profile is not similar for melanized Cryptococcus biofilms. Recently, antibiofilm activity of antifolate drugs (sulfamethoxazole-trimethoprim and sulfadiazine-pyrimethamine) alone and in combinations with amphotericin B was studied (Liao et al. 2015). However clinical study and combination therapy for cryptococcal shunt infections are lacking.
6.3 Candidiasis
Candida albicans is the most prevalent infectious species among the 20 different species of Candida (Centre for Disease Control 2014). It is the normal commensal of the alimentary tract, vagina, and oral cavity. The emergence of Candida sp. as the important nosocomial pathogen is owing to the increased use of medical devices such as central venous catheters and various implanted devices (Kim et al. 2011). Although C. albicans is the most prevalent, non-albican species, C. glabrata, C. tropicalis, C. parapsilosis, C. guilliermondii, C. krusei, C. lusitaniae, and C. kefyr are also increasing recently, which is evident in a recent 5-year prospective study and population-based surveys of candidemia (Alexander et al. 2013). For candidemia management, the most frequently administered antifungal agent is fluconazole and echinocandins. The effectiveness of the antifungal agent varies with the species of Candida isolated from different clinical samples (Pfaller et al. 2012; Sardi et al. 2013).
Andes et al. (2012) conducted an individual patient-level quantitative review of randomized trials consisting of 1,915 patients for treatment of invasive candidiasis. They identified that the use of an echinocandin was associated with reduced mortality when compared to the use of a drug from either the triazole or polyene classes. And their study also supports the greater toxic effects associated with amphotericin B than triazole or echinocandin therapy. These findings lend support to recent treatment guidelines that recommend echinocandin as a first-line choice for invasive candidiasis particularly for patients who are critically ill and those with previous exposure to triazole (Andes et al. 2012).
The treatment for Candida infections varies with patient’s underlying conditions. For instance, the delayed treatment of candidemia in septic shock is shown to be an important factor of patient outcome. Septic shock due to invasive candidiasis is a near-fatal condition. Retrospective cohort studies of septic shock patients with Candida infections reveal the risk of death is exceptionally high among patients with septic shock associated with Candida infection. The antifungal therapy regime is needed to improve the clinical outcomes (Kollef et al. 2012). Invasive candidiasis is also a leading cause of mortality and morbidity in immune-compromised neonates. A comparative outcome of antifungal treatment of candidiasis in infants was provided by a large multicenter cohort study. Infants treated with L-AmB had a higher mortality when compared to infants treated with either amphotericin B deoxycholate or fluconazole. This finding may be related to inadequate penetration of L-AmB into the kidneys. There are no recommendations for combination therapy to treat life-threatening infection and deep tissue infection as the comparative outcome data are limited (Tragiannidis et al. 2015). In case of candidiasis in hematologic malignancies, echinocandins are recommended as initial therapy, in non-neutropenic patients with candidial infections, initial therapy with the echinocandin followed by fluconazole, is recommended. There are no reports on combination therapy for invasive candidiasis in leukemic patients (Walsh and Gamaletsou 2013). The summary of recent treatment recommendations for different types of candidiasis is given in Table 6.1.
The important factor for drug resistance in Candida sp. is their ability to grow and adhere as biofilms on various surfaces including the medical devices. Candida biofilms are the leading cause for intravascular catheter-related infections. The presence of biofilms on intravascular devices makes the treatment of CR-BSI difficult and resists the action of clinically important drugs including amphotericin B and azole (Chatzimoschou et al. 2011). Also a recent randomized clinical trial reveals the higher toxicity effects of amphotericin B with fluconazole (Sanati et al. 1997). Thus, many researchers have turned their focus to evaluate the antifungal therapy comprising different combinations including antimicrobial peptides, natural products, and non-antifungal agents. And many studies are being carried out to demonstrate the antimicrobial lock therapy to treat catheter-related blood infections. Antimicrobial lock therapy involves the instillation of a small volume of the highly concentrated antibiotic solution, which is allowed to stay in the lumen of the catheter for 12–72 h, to eradicate biofilm formed on the inner surface of the device. Ku et al. (2010) determined the efficacy of antibiofilm and persistence of the activity for the lock solutions of caspofungin, micafungin, and posaconazole in in vitro model of Candida biofilm on silicone catheter. Micafungin and caspofungin had significant inhibitory activity against mature C. albicans and C. glabrata biofilms, but caspofungin displayed less persistence in C. glabrata biofilms. Even though posaconazole was less effective against C. albicans biofilms, its activity was maintained. They had suggested that echinocandin lock therapy may be useful to control candidiasis in catheterized patients (Cateau et al. 2011). Until recently, the guidelines of the Infectious Diseases Society of America (IDSA) had suggested the use of antibiotic lock therapy only under special circumstances as a salvage treatment. For the management of catheter-related bloodstream infections, tigecycline, an antibacterial agent, was evaluated alone and in combination with fluconazole, amphotericin B, or caspofungin against Candida biofilms. It has been found that high-dose tigecycline is highly active against in vitro Candida biofilms; however, the combination with antifungals is not significant. Further investigation is required to utilize the antimicrobial lock therapy containing tigecycline for the management of CR-BSI (Ku et al. 2010). The antibiofilm activity of amphotericin B was found to be significantly enhanced when used in combination with polyamine biosynthesis inhibitors, 1,4-diamino-2-butanone (DAB) and difluoromethylornithine (DFMO). Further studies have shown that DAB and DFMO also enhanced the antibiofilm activities of several other antifungal agents by enhancing the formation of reactive oxygen species in biofilm cells.
In vitro, the combination of caspofungin or anidulafungin with antimicrobial peptides is very effective in killing Candida sp.; however, the efficacy was not improved in a murine model of systemic candidiasis, wherein the combination of caspofungin and antimicrobial peptide, ranalexin, was used. The enhanced efficacy was observed in the same animal model when the combination therapy of amphotericin B and lactoferrin was used. There is no literature support for the synergism between antimicrobial peptides and echinocandins. Rossignol et al. (2011) investigated the tryptophan-rich, ApoEdpL-W peptide, derived from human ApoE apolipoprotein shown to have less antifungal activity against mature biofilms and partially inhibited biofilms on medical devices.
Synergism of natural products with antibiotics is the currently developing area of phytopharmaceuticals. Many natural products including essential oils, peptides, and polyphenols are active against different shapes of Candida biofilms. Recently the effectiveness of the antifungal activity of the essential oils of Ocimum sanctum was evaluated for the fluconazole-resistant Candida sp. Besides its selective cytotoxicity, the synergism with the fluconazole and ketoconazole showed a better antifungal activity in in vitro fluconazole-resistant Candida strains (Amber et al. 2010). Similar synergistic interactions were observed in farnesol with micafungin, fluconazole, and amphotericin B against in vitro C. albicans biofilms (Katragkou et al. 2015). The in vitro effect of essential oil components of eugenol and methyleugenol alone and in combination was checked against fluconazole-sensitive and fluconazole-resistant strains of clinical Candida sp. The synergistic interactions of eugenol and methyleugenol with fluconazole are very effective killing fluconazole-resistant strains (Ahmad et al. 2010a). Mandal et al. (2011) demonstrated the activity of Tn-AFP1 from Trapa natans against biofilm inhibition of C. tropicalis in vitro. Similarly, Delattina et al. (2014) identified OSIP108, a nontoxic decapeptide from the model plant Arabidopsis thaliana, that inhibits C. albicans biofilm. These recent reports support the synergism of natural products with antifungal agents in controlling the Candida biofilms. However, the major drawback of natural products’ utility is the unestablished toxicity profiles. One such well-established compound, fulvic acid, was proven to be nontoxic in an epithelial cell line and rat model alone (Nett 2014). The information from several studies indicating synergistic interactions of non-antifungal drugs in combination with fluconazole are summarized in Table 6.2
Due to the high cost and side effects of combination therapy, the focus of research recently turned to examine the combinations with non-antifungal agents, including tetracyclines (Loreto et al. 2011), quinolones (Wang et al. 2012), calcineurin inhibitors (Chen et al. 2011), heat shock protein 90 inhibitors, calcium homeostasis regulators, and traditional medicines (Muñoz et al. 2012).
One such combination is azole with calcineurin inhibitor. The calcineurin signaling pathway is a key mediator of stress responses in C. albicans as a potential drug target for systemic candidiasis and candidal keratitis. Calcineurin inhibitors when given as a monotherapy were not suitable for other candidial infections of oropharyngeal, pulmonary, and vaginal tract. The virulence pathway inhibitor in combination with azole would increase the efficiency of the antifungal drug. The establishment of virulence pathway varies with the species. Hence, identifying suitable drug targets and finding its synergism with the antibiotics would be potential therapeutic options (Reedy et al. 2010). However, drugs targeting the inhibition of virulence expression would be suitable targets to enhance the current antifungal therapy and to reduce the evolution of drug-resistant strains. The synergism shown by calcineurin inhibitors with the antifungal agents amphotericin B and caspofungin is potentially good as the combination lock therapy in the rat catheter infection model; however, its utility is limited as the drugs have immunosuppressive effects that prevents the systemic administration.
Diclofenac, aspirin in combination with caspofungin and amphotericin B, was evaluated to treat infections of Candida albicans and C. parapsilosis. Bink et al. (2012) screened 1,600 compounds of a drug-repositioning library that enhance antibiofilm activity against C. albicans. They have found out that hexachlorophene, pyrvinium pamoate, and artesunate can act synergistically with miconazole in eradicating C. albicans biofilms. They suggested miconazole and artesunate as the potential combination therapy to treat oral and vulvovaginal candidiasis. In vivo toxicity studies are required to execute the potentiality of the miconazole and artesunate (De Cremera et al. 2015). Finasteride, a 5-α-reductase inhibitor, commonly used for the treatment of benign prostatic hyperplasia was tested alone and in combination with amphotericin B and fluconazole against C. albicans urinary biofilms. It exhibits synergistic activity in combination with fluconazole. However, further investigation is required to explore the clinical utility of finasteride in the prevention of candidiasis (Chavez-Dozal et al. 2014). Synergistic interactions were observed during in vitro combination studies of allicin and flucytosine against C. albicans, C. glabrata, and C. tropicalis (Alexander et al. 2013). L-AmB in combination with heat shock protein, HSP 90 inhibitor (Mycograb), also provides promising therapy; however, the antibody-based inhibitor is not available for clinical trials (Pachl et al. 2006). The synergistic interactions of fluconazole with non-antifungal agents are summarized in Table 6.2.
The present review recommends the necessity for standard guidelines for the management of fungal biofilm infection.
6.4 Aspergillosis
Aspergillus exposure to human is quite common due to their ubiquitous nature. In immune-compromised host, it causes aspergillosis. Invasive aspergillosis (IA) has become increasingly common in recent days resulting in a mortality ranging from 40 % to 80 % (Ascher et al. 2012; Aguado et al. 2015). The predominant causative agent is A. fumigatus which also finds concurrence in a recent literature study reported from 1985 to 2013. The study comprised of 116 liver transplant recipients with invasive aspergillosis. The most common infecting species are Aspergillus fumigatus (73 %), A. flavus (14 %), and A. terreus (8 %). Voriconazole improves the survival among the commonly used AMB and itraconazole (Barchiesi et al. 2015; Ogawa et al. 2015). However, an increase in drug resistance restricts the usage of azole for treatment and necessitates the combinational therapy approach.
A randomized, double-blind, placebo-controlled, multicenter trial comprising 454 patients with hematologic malignancies and hematopoietic stem cell transplantation suspected with invasive aspergillosis were treated with voriconazole and anidulafungin. The study showed the improvement of survival when compared with monotherapy (Marr et al. 2015). In another retrospective study that was conducted between 1993 and 2008, the combination of L-AmB and echinochandins for the management of IA in hematological malignancies was analyzed, and the results suggested that the combination therapy offered no improvement in terms of response and mortality (Mihu et al. 2010).
Though sinus aspergillosis was believed to be a rare disease for many years, recently there has been an increase in the number of cases. Chronic invasive aspergillosis of the sinus occurs in mildly immunodeficient patients. Two of the most important treatment regimens for invasive aspergillosis of the sinus are surgical debridement and antifungal therapy. The pharmacodynamic interactions of amphotericin B and voriconazole were assessed using conventional in vitro tests that mimic human serum concentrations. The combination was more effective than monotherapy regimens (Siopi et al. 2015).
Various formulations of L-AmB differ in terms of toxicity and efficacy which concurs with a recent finding by Olson et al. (2015) while comparing, AmBisome and Lambin, in Aspergillus fumigatus mice model. Lambin was more toxic than the other, emphasizing the adequacy requirement to compare liposomal formulations. The in vitro combination studies of nystatin-intralipid with caspofungin demonstrated effective synergistic interaction against A. terreus when compared with voriconazole and 5-fluorocytosine (Semis et al. 2015; Shadomy et al. 1975). In a murine aspergillosis model, the combinations of L-AmB, micafungin, and deferasirox improved the fungal clearance and patient survival. The in vitro efficacy of PTX3, a glycoprotein produced by monocytes in combination with voriconazole, improved patient survival and clearance of fungal burden (Giudice et al. 2012). Erythropoietin in combination with AMB improved the outcome of in vivo disseminated aspergillosis.
The synergistic interactions vary with the species and resistance mechanism. In a similar kind of study, the synergistic interactions of posaconazole and caspofungin varied with 20 different wild and resistant types of A. fumigatus, with ten different resistant mechanisms. The interactions are insignificant when compared with wild isolates (Mavridou et al. 2015). Similarly, while investigating the efficacy of the combination of voriconazole and anidulafungin in an in vitro model of invasive pulmonary mice infected with triazole-resistant Aspergillus, the addition of anidulafungin does not significantly improve the voriconazole activity, whereas another study suggests the combination of triazole and echinocandin as the treatment strategy for triazole-resistant isolates (Lepak et al. 2013). The efficacy varied with the type of strain and the drug resistance pattern evolved. A study by Jeans et al. (2012) has proven the efficacy of voriconazole and caspofungin in an in vivo model of pulmonary aspergillosis infected with different species of Aspergillus. The synergism between A. flavus and A. niger was found to be significant while not in the case of A. fumigatus (Zhang et al. 2014). In a neutropenic A. fumigatus murine model, the synergistic interactions of voriconazole and anidulafungin were observed in voriconazole-susceptible strain, but additive in resistant type, which creates concern for the management of infections by voriconazole-resistant strains.
The inappropriate use of antibiotics is another major factor for the emergence of drug-resistant strains. A recent cross-sectional cohort study states that systemic antifungal therapy was administered irrespective of the presence of invasive fungus infection (Ascher et al. 2012; Olson et al. 2015). Based on this, a recent survey study conducted revealed the weakness in the knowledge on current guidelines of invasive candidiasis and invasive aspergillosis among the European physicians (Valerio et al. 2015).
A. fumigatus biofilms are difficult to manage with conventional antifungal agents; hence, even after antimycotic therapy, these infections are not eradicated and are emerging as life-threatening infections (Mowat et al. 2007). Melanin produced by A. fumigatus during infection is required for lung tissue invasion, conferring less susceptibility to drugs (Bowyer et al. 2011). Biofilms of A. fumigatus are resistant to itraconazole and, some extent, to caspofungin due to its extracellular polymeric substances (EPS) (Singh et al. 2011). An in vitro finding reveals A. fumigatus are not responding to amphotericin B, when they have a prior exposure to voriconazole, suggesting the improper usage of azole will be a hindrance for treatment with amphotericin B. Synergistic interactions were observed in combination with caspofungin and amphotericin B or voriconazole against Aspergillus biofilms (Liu et al. 2012). Another in vitro study with combinations of amphotericin B or Lamb with alginate lyase, the degrading enzyme of EPS (Bugli et al. 2013; Rajendran et al. 2015), suggesting the EPS as a suitable target for antibiofilm activity. There are very limited clinical studies that are focused on the management of biofilm-associated A. fumigatus infections.
In a recent literature review, data addresses the significance of combination antifungal therapy in the treatment of invasive aspergillosis. The information suggests that evidence is not convincing enough to support the use of combination antifungal therapy (Garbati et al. 2012). Another systematic review comprising animal and clinical studies addressed the efficacy of combination therapy of triazole and echinocandin in treatment of invasive aspergillosis, suggesting a development toward improved overall survival (Day et al. 2013; Marr et al. 2015).
In summary, combination therapies appear to be conflicting in some studies, and the limited clinical data are not convincing to support combination antifungal therapy over monotherapy. In addition, well-sketched clinical trials focusing on management of biofilm-related fungal infections are immediately needed to overcome the emergence of drug resistance in these pathogens.
6.5 Immunotherapy Against Fungal Infections
Opportunistic fungal infections become a major challenge to health care since a majority of them have affected immune-compromised individuals such as HIV, hypersensitive, autoimmune, or transplant patients. Most of these infections in the case of normal persons are easily combated by the immune system, which is not the case with patients whose immune system is suppressed. Added to this are the problems associated with hospital multiple drug-resistant pathogens. Opportunistic fungal infections such as candidiasis, aspergillosis, and cryptococcosis are three of the leading causes of significant mortality in both hospital-derived infections and in those of immune-compromised patients (Truan et al. 1994). Studies also clearly suggest that due to modified lifestyles and exposure, these fungal infections are set to increase and dominate the concerns of medical industry in the future (Kauffman 2001). This creates the need for either combinatorial therapies or rapid discovery of novel antibiotics. However, most such strategies are replete with adverse side effects and may not be favorable, though the benefits far outweigh the risks. One way forward to combat such opportunistic fungal infections, with nil or minimal side effects, is immunotherapy (Ahmad et al. 2010b).
Majority of the immunotherapy strategies rely on immunomodulation of the patient’s immune system to prime it against the fungal pathogen. This indirect method has many advantages, including lack of side effects and resistance development (Ahmad et al. 2010b; Owais et al. 2010), and more importantly such treatment to some extent can be catered to suit an individual patient’s needs. However, it’s important to recognize that host-fungi interaction (Ahmad et al. 2010b), immune responses of the host to fungi, and, importantly, immune escape mechanisms of fungi are quite complicated and the underlying mechanisms of fungal infections are beginning to be unraveled. With better understanding of these components, there is a better hope of developing much more effective immune modulators. In the next few sections, we will primarily focus on the use of immunotherapy as the latest and futuristic treatment for three of the major opportunistic pathogenic fungi: Candida, Aspergillus, and Cryptococcus.
6.5.1 Antimicrobial Proteins
These form one of the first lines of innate immune defense against a variety of pathogens and are widely distributed in animal kingdom. These are usually cationic and amphipathic peptides (Owais et al. 2010) and expressed in cells of the innate immunity such as macrophages, mast cells, and neutrophils, as well as in nonimmune cells such as keratinocytes and epithelial cells. The antimicrobial proteins can either directly defend the pathogens (Braff et al. 2005) or stimulate cells such as neutrophils (Sorensen et al. 1997) to degranulate resulting in inflammatory responses (Hancock and Diamond 2000). These antimicrobial peptides comprise cathelicidins (Sorensen et al. 1997), β-defensins, substance P, RANTES (Braff et al. 2005), LL-37 (Larrick et al. 1995; Nagaoka et al. 2001), etc. Apart from immune cell activation, these peptides can also activate epithelial cells upon microbial invasion leading to proinflammatory cytokine signaling (Van Watering et al. 1997) and toll-like receptor (TLR) activation (Akira et al. 2006). This could end up activating and recruiting immune cells to the site of infection and thus boosting the host immune defense. Thus, components of the innate immunity can be tweaked that could prove to be effective not only in controlling the spread of infection but also end up activating the adaptive immune components. However, the effectiveness of the use of these antimicrobial peptides as a therapy against opportunistic fungal pathogens needs more attention.
6.5.2 Cytokines
Both pro- and anti-inflammatory cytokines are regulators of innate and adaptive immune responses (Iwasaki and Medzhitov 2015), including cell recruitment, immune cell activation, and immune cell differentiation. Nowhere is this more pronounced than in the interplay of cytokines during development of T helper subsets (Romani 2000). Studies have shown the importance of cytokines in not only modulating immune responses against fungal pathogens (Mencacci et al. 1998) but have also demonstrated mechanisms involved in hijacking of the host cytokine network for immune escape by the fungal pathogens (Netea et al. 2006). Cytokines such as proinflammatory TNF-α and IFN-γ are unregulated (Rodriguez-Adrian et al. 1998) upon infection by Candida and Aspergillus, resulting in the activation of phagocytic cells. Cytokines can modulate immune cells very effectively if properly applied and are one of the potential immunotherapeutic strategies against opportunistic fungal infections. TNF is a proinflammatory cytokine and has been found to be very effective against A. fumigatus (Steinbach and Stevens 2003). TNF has pleiotrophic effect on phagocytic cells, including cell activation and phagolysosomal killing of fungal pathogens (Cisalpino et al. 1996). The study by Steinbach and Stevens (2003) has shown the importance of TNF-α in suppressing immunodeficiency induced by dexamethasone, and the mechanism consists of upregulation of TLR and IFN-γ and preventing leukocyte apoptosis (van der Meer et al. 2005). Although both Th1 and Th2 are essential for anticryptococcal immunity, Th1 are very effective (Hoag et al. 1997). Thus, higher levels of Th1 cytokines such as TNF-α and IFN-γ are associated with increased fungal clearance (Milam et al. 2007; Wormley et al. 2007). More importantly, TNF-α is necessary for IFN-γ production (Bekker et al. 2001) which results in macrophage activation. So, it is clear that TNF therapy could potentially stimulate IFN activity leading to better antifungal responses. Apart from this, pathogen-associated molecular patterns (PAMPs) are potent antigens recognized by innate pattern recognition receptors (PRRs) such as TLRs. Both TNF-α and IFN-γ synergize to increase the cell surface expression of TLRs (Bosisio et al. 2002) resulting in better recognition and clearance of Candida, Aspergillus (Tsiodras et al. 2008), and Cryptococcus (Pappas et al. 2004). Thus, TNF-α immunotherapy could be a potential therapeutic strategy against fungal pathogens.
IFN-γ synergizes with TNF-α in a variety of immune functions including Th1 differentiation (Milam et al. 2007), TLR expression (Tsiodras et al. 2008), and phagocyte activation (Bekker et al. 2001). In rodents, the use of IFN-γ has shown good results against systemic cryptococcosis (Lutz et al. 2000), and the same cytokine was also successfully tested in one human patient (Netea et al. 2004). Studies have shown the effectiveness of IFN-γ against a variety of fungal pathogens including Candida, Aspergillus, and Cryptococcus (Rodriguez-Adrian et al. 1998; Shahid et al. 2010). IFN-γ has been shown to stimulate phagocytic activity and intracellular killing of A. fumigatus (Steinbach and Stevens 2003; Shahid et al. 2010), through stimulating generation of free radical production (Steinbach and Stevens 2003). In murine models, immunotherapy using TNF-α and IFN-γ was shown to produce significant reduction in mortality due to Aspergillus infection (Shahid et al. 2010). However, one issue with IFN-γ usage is that it reduces anticryptococcal activity of human macrophage and thus its use in humans is debatable (Reardon et al. 1996). Nevertheless, in 2010, the Infectious Diseases Society of America has approved recombinant IFN-γ against persistent cryptococcosis (Antachopoulos and Walsh 2012). Gallin et al. (1995) have shown that the use of IFN-γ under clinical settings is very effective in the case of patients suffering from chronic granulomatous disease (CGD) and who are especially prone to Aspergillus (Antachopoulos and Roilides 2005). Unlike anticryptococcal action, macrophage anti-Candida activity is enhanced by IFN-γ treatment (Redmond et al. 1993; Baltch et al. 2005). But other studies in rodents have shown lack of an effect of IFN-γ (Brummer and Stevens 1987; Marcil et al. 2002) in case of pulmonary macrophages, but peritoneal or peripheral blood PMNs (from both human and murine) showed good anti-Candida activity after IFN-γ treatment (Djeu et al. 1986; Kullberg et al. 1993). In spite of the lack of strong evidence for IFN-γ immunotherapy, Dignani et al. (2005) showed in the case of three patients with candidiasis that IFN-γ therapy was actually beneficial. Similarly, IFN-γ immunotherapy of an HIV-infected patient proved to be beneficial (Riddell et al. 2001; Bodasing et al. 2002). Nonetheless, large-scale clinical trials are the need of the hour to better understand the efficacy of this cytokine for immunotherapy.
IL-12, like TNF-α and IFN-γ, is a Th1 cytokine (Milam et al. 2007; Wormley et al. 2007), and Th1 activation and responses are vital for strong antifungal responses against Candida, Aspergillus, and Cryptococcus. IL-12 has been shown to inhibit Th2 responses, thereby increasing anti-candidiasis response of the host (Morrison et al. 1986; Deepe 1997). Other studies have shown the potential of IL-12 in enhancing the antifungal efficacy of fluconazole against Candida (Afonso et al. 1994), murine cryptococcosis (Muchmore et al. 1968), and aspergillosis (Allendoerfer et al. 1996). From these studies, it is recognized that one way by which IL-12 could exert its antifungal effect is through enhancement in IFN-γ levels (Antachopoulos and Roilides 2005; Kawakami et al. 1996; Jacobson et al. 2000), synergization with IL-18 in the induction of anticryptococcal activity (Qureshi et al. 1999), and suppression of IL-4 activity (Qureshi et al. 1999). However, IL-12 usage has been shown to exacerbate mucosal disease in mouse (Morein et al. 1984) and produce adverse reactions on the immune system (Murphy et al. 1993). Thus, more studies are warranted to check the efficacy of this cytokine as an immunotherapy agent.
Colony-stimulating factors (CSFs) are essential for phagocyte development (especially in the case of neutropenia) and function and have been approved for humans (Rodriguez-Adrian et al. 1998). Being able to facilitate antifungal activity of phagocytes, CSFs are one of the promising immunotherapy agents. Approved and accepted since 1991 for patient use (Welte et al. 1996), granulocyte colony-stimulating factor (G-CSF) is effective in prolonging phagocyte and respiratory burst activity against Aspergillus infection (Polak-Wyss 1991; Liles et al. 1997) and Candida hyphae (Gaviria et al. 1999; Kullberg et al. 2004). However, when compared to other CSFs, G-CSF also produces anti-inflammatory cytokines and could be useful against endotoxins in immune-compromised patients (Gomez et al. 1995). G-CSF also favors a Th2 response and so can be beneficial against Candida (Condon et al. 1996) and Aspergillus (Cisalpino et al. 1996). However, anticryptococcal immunity requires a robust Th1 activation, and thus G-CSF may not prove to be successful here. However, neutrophils from G-CSF-treated HIV patients have been shown to be effective against Cryptococcus and Candida (Han and Cutler 1995), and this was shown to be due to enhanced superoxide anion generation. Granulocyte-macrophage colony-stimulating factor (GM-CSF) is very important for the development and activation of neutrophils (Rodriguez-Adrian et al. 1998) and macrophage (Steinbach and Stevens 2003). In murine models, Brummer and Stevens (1987) demonstrated the usefulness of GM-CSF in rescuing dexamethasone-induced suppression of macrophage and synergistically working with TNF-α (in potentiating anti-Aspergillus function of macrophage (Bodey 1994)). Rowe et al. (1995) have shown that GM-CSF was effective against aspergillosis in clinical trials, but issues remain (Kong and Levine 1967; Bodey et al. 1993; Groll et al. 1996). However, when compared to other CSFs, studies have shown that GM-CSF is more effective (Kirkland and Fierer 1985; Kirkland et al. 1991) and thus is more promising. In combination with antifungal therapy, GM-CSF administration was shown to improve clinical outcome of Candida infection (Vazquez et al. 2000), but controlled trials are the need of the hour as far as GM-CSF is concerned.
Macrophage colony-stimulating factor (M-CSF), important for mononuclear phagocyte functions (Shahid et al. 2010), has been demonstrated to be very effective against Candida infections (Mencacci et al. 1996). A study employing rabbits (Gonzalez et al. 2001) has shown that M-CSF is very effective against pulmonary aspergillosis and resulted in higher numbers of activated alveolar macrophages. But its usefulness, when compared to G-CSF or GM-CSF, is less clear (Steinbach and Stevens 2003). Currently, human recombinant G-CSF and GM-CSF are available for clinical use but not so in the case of M-CSF (Hubel et al. 2002). Thus, extensive studies are needed to confirm the effectiveness of these three CSFs as potent immunotherapy agents.
6.5.3 Antibodies
Antibody represents one of the best effector arms of the humoral immune response and is important for establishing a robust secondary immune response. Being highly specific in their action, antibodies are receiving a lot of interest as adjuncts for antifungal therapy. Antibody-mediated immunity (AMI) thus is a potential immunotherapeutic strategy and can be activated by direct mechanism or by indirect mechanism (Casadevall and Pirofski 2012). In the case of direct action, antibodies can interfere with the fungal pathogen and inhibit Cryptococcus virulence production (Martinez et al. 2004) and resistance development (Martinez and Casadevall 2005), Candida replication (Brena et al. 2007), gene expression in Cryptococcus (McClelland et al. 2010), and metabolism in Candida (Brena et al. 2011). By their indirect action, antibodies have been shown to enhance antifungal responses against Cryptococcus by inducing phagocytic activity (Schlagetter and Kozel 1990), complement activation (Shapiro et al. 2002), NK cell-mediated killing (Nabavi and Murphy 1986), and proinflammatory responses (Feldmesser et al. 2002). Thus, for both Candida and Cryptococcus, a variety of monoclonal antibodies have been developed (Ullmer et al. 1993; Beno et al. 1995) but only very few against Aspergillus (Torosantucci et al. 2009). In most of these studies, it is clear that administration of mAbs produces a variety of antifungal effects including opsonization (Rachini et al. 2007), inhibition of cell growth (Rodrigues et al. 2000), reduction of capsule/cell wall thickness (Rachini et al. 2007), regulating cytokine production and interplay (Beenhouwer et al. 2001), and in general triggering inflammation (Rivera et al. 2005). But, patient response to mAb administration remains inconsistent since their function is affected by dose and immune parameters of the patient (Casadevall and Pirofski 2006). In addition, the actual antigen load can also influence antibody activity (Casadevall and Pirofski 2003). Under these circumstances, studies have started to show the importance of idiotypic antibodies, especially against Candida (Van De Veerdonk et al. 2010), primarily because these produce highly specific immunotherapy and affect the so-called “natural antibody” network.
6.5.4 Transfusion of Leukocytes
Opportunistic infection by fungal pathogens such as Candida, Aspergillus, and Cryptococcus is high in the case of immune-compromised patients, including those with neutropenia. The use of blood transfusion, especially using leukocytes, represents an effective strategy for immunotherapy. In spite of early failures, leukocyte transfusion is gaining importance now due to advances in our understanding of leukocyte development and function. Thus, for example, use of CSFs along with leukocyte transfusion increases the effectiveness of immunotherapy (Casadevall 1995; Deepe 1997). Indeed, clinical trials have shown that the use of G-CSF-pretreated leukocytes for transfusion is very effective against Candida (Condon et al. 1996) and Aspergillus (Cisalpino et al. 1996) infections. In fact phase 1/2 trial using G-CSF primer leukocytes has been reported to be effective (Cox et al. 1988). Bozza et al. (2004) have reported that transfer of antigen-primed dendritic cells results in enhanced adaptive immune response against Candida. In vitro priming of leukocytes, by appropriate cytokine or fungal antigen, and subsequent transfer of the patient represent a simple strategy for antifungal immunotherapy (Bacci et al. 2002). It is recognized that neutrophils are effective against disseminated candidiasis, whereas cell-mediated immunity is triggered against mucosal candidiasis (Deepe 1997). In the case of Aspergillus infection of mononuclear cells (Deepe 1994) and cryptococcosis, macrophages (Rohatgi and Pirofski 2015) appear to be important. Tramsen et al. (2007) have shown the effectiveness of anti-Candida T cells after transfusion, especially against the hyphal forms and synergize with neutrophils to restrict Candida infection (Tramsen et al. 2007). In the case of cryptococcosis, Th1 responses are vital (Hoag et al. 1997) and higher levels of Th1 cytokines such as TNF and IFN are associated with increased fungal clearance (Milam et al. 2007; Wormley et al. 2007). In addition to this, a recent work by Muller et al. (2007) has demonstrated the importance of IL-17 and the associated Th-17 response in immune modulation of C. neoformans-infected mice. Moreover, such adoptive transfer of leukocyte also has the added advantage of increasing the activity of specific components of either innate or adaptive immune system, resulting in a robust response. Thus, it is clear that transfusion of a specific type of leukocyte after priming, based on the nature of fungal infection, could be an important immunotherapy strategy in the case of immune-compromised patients.
6.5.5 Vaccines
Antigens from fungus have been successfully isolated and have been used as vaccine candidates. An excellent review by Casadevall and Pirofski (2012) shows that some of these antigens include β-glucan (Torosantucci et al. 2009) and cell wall glycoprotein (Chaturvedi et al. 2005) from A. fumigatus, Hsp 90 (Matthews et al. 1991), aspartyl proteinase (Sandini et al. 2011), phosphoglycerate kinase (Calcedo et al. 2011) from C. albicans and melanin (Rosas et al. 2001), glucosylceramide (Rodrigues et al. 2000), β-glucan (Torosantucci et al. 2009), and glucuronoxylomannan (Beenhouwer et al. 2007) from C. neoformans. These studies have shown that most of these antigens elicit excellent antibody-mediated immunity in the host and thus are quite successful (for more information, please refer to specific reviews on antifungal vaccines). However, a successful vaccination requires intact cellular components and thus might be an issue in the case of immune-compromised individuals. However, vaccination strategy has worked well in murine models of both systemic and mucosal candidiasis (Torosantucci et al. 2005), using diphtheria toxoid CRM197 and laminarin conjugate. Similarly, vaccine conjugates containing mannan too have been successful against disseminated candidiasis and vaginal infection by Candida in mice (Han et al. 1999). The use of inert antigens from the fungal pathogens appears to be preferred since it is safer when compared to attenuated or live fungus. But, such a strategy may not be very effective in terms of the antigenicity or purity of the antigen preparation. Under these circumstances recombinant technology provides an excellent opportunity to precisely select and express the desired epitope to be used as an effective vaccine. Nevertheless, the use of adjuvants, such as cytokines, has opened up huge possibilities in antifungal vaccine development (Deepe 1997), which allows us to modulate the cellular response postvaccination. Another way forward is DNA vaccines, as has been shown for virulent bacterial infections such as Mycobacterium (Tascon et al. 1996). But antifungal DNA vaccines are yet to be reported for major fungal pathogens. In addition, extensive studies are needed to analyze antifungal vaccines in humans to better understand the mechanisms involved and the therapeutic effectiveness reached. This will enable us to develop an effective antifungal vaccination for use in the general population.
References
Afonso LCC, Scharton TM, Viera LQ, Wysocka M, Trinchieri G, Scott P (1994) The adjuvant effect of interleukin-12 in a vaccine against Leishmania major. Science 263:235–237
Aguado JM, Vázquez L, Fernández-Ruiz M, Villaescusa T, Ruiz-Camps I, Barba P, Silva JT, Batlle M, Solano C, Gallardo D, Heras I, Polo M, Varela R, Vallejo C, Olave T, López-Jiménez J, Rovira M, Parody R, Cuenca-Estrella M (2015) Serum galactomannan versus a combination of Galactomannan and Polymerase chain reaction – based Aspergillus DNA detection for early therapy of invasive aspergillosis in high-risk hematological patients: a randomized controlled trial. Clin Infect Dis 60(3):405–414
Ahmad A, Khan A, Khan LA, Manzoor N (2010a) In vitro synergy of eugenol and methyleugenol with fluconazole against clinical Candida isolates. J Med Microbiol 59(10):1178–1184
Ahmad I, Khan MSA, Zahin M, Owais M, Shahid M, Mehmood Z, Pant AB et al (2010b) Combinational antifungal therapy and recent trends in drug discovery. In: Ahmad I (ed) Combating fungal infections. Springer-Verlag, Berlin. doi:10.1007/978-3-642-12173-9_10
Akira S, Uematsu S, Takeuchi O (2006) Pathogen recognition and innate immunity. Cell 124:783–801
Albuquerque P, Casadevall A (2012) Quorum sensing in fungi – a review. Med Mycol 50(4):337–345
Alexander BD, Johnson MD, Pfeiffer CD, Jiménez-Ortigosa C, Catania J, Booker R, Castanheira M, Messer SA, Perlin DS, Pfaller MA (2013) Increasing echinocandin resistance in Candida glabrata: clinical failure correlates with presence of FKS mutations and elevated minimum inhibitory concentrations. Clin Infect Dis 56(12):1724–1732
Allen UD (2010) Antifungal agents for the treatment of systemic fungal infections in children. Paediatr Child Health 15(9):603
Allendoerfer R, Maresca B, Deepe GS (1996) Cellular immune responses to recombinant heat shock protein 70 from Histoplasma capsulatum. Infect Immun 64:4123–4128
Amber K, Aijaz A, Immaculata X, Luqman KA, Nikhat M (2010) Anticandidal effect of Ocimum sanctum essential oil and its synergy with fluconazole and ketoconazole. Phytomedicine 17(12):921–925
Andes DR, Safdar N, Baddley JW, Playford G, Reboli AC, Rex JH, Sobel JD, Pappas PG, Kullberg BJ (2012) Impact of treatment strategy on outcomes in patients with candidemia and other forms of invasive candidiasis: a patient-level quantitative review of randomized trials. Clin Infect Dis 54:1110–1122, cis021
Angela Bacci*, Claudia Montagnoli*, Katia Perruccio†
Antachopoulos C, Roilides E (2005) Cytokine and fungal infections. Br J Haematol 129:583–596
Antachopoulos C, Walsh TJ (2012) Immunotherapy of Cryptococcus infections. Clin Microbiol Infect 18:126–133
Ascher SB, Smith B, Watt K, Benjamin DK, Cohen-Wolkowiez M, Clark RH, Benjamin DK, Moran C (2012) Antifungal therapy and outcomes in infants with invasive Candida infections. Pediatr Infect Dis J 31(5):439
Bacci A, Montagnoli C, Perruccio K, Bozza S, Gaziano R, Pitzurra L, Velardi A, D’Ostiani CF, Cutler JE, Romani L (2002) Dendritic cells pulsed with fungal RNA induce protective immunity to Candida albicans in hematopoetic transplantation. J Immunol 168:2904–2913
Bal AM (2010) The echinocandins: three useful choices or three too many? Int J Antimicrob Agents 35(1):13–18
Baltch AL, Bopp LH, Smith RP, Ritz WJ, Carlyn CJ, Michelsen PB (2005) Effects of voriconazole, granulocyte-macrophage colony-stimulating factor, and interferon gamma on intracellular fluconazole-resistant Candida glabrata and Candida krusei in human monocyte-derived macrophages. Diagn Microbiol Infect Dis 52:299–304
Barchiesi F, Di Francesco LF, Compagnucci P, Arzeni D, Giacometti A, Scalise G (1998) In-vitro interaction of terbinafine with amphotericin B, fluconazole and itraconazole against clinical isolates of Candida albicans. J Antimicrob Chemother 41(1):59–65
Barchiesi F, Mazzocato S, Mazzanti S, Gesuita R, Skrami E, Fiorentini A, Singh N (2015) Invasive aspergillosis in liver transplant recipients: epidemiology, clinical characteristics, treatment, and outcomes in 116 cases. Liver Transpl 21(2):204–212
Bartizal K, Gill CJ, Abruzzo GK, Flattery AM, Kong L, Scott PM, Smith JG, Leighton CE, Bouffard A, Dropinski JF, Balkovec J (1997) In vitro preclinical evaluation studies with the echinocandin antifungal MK-0991 (L-743,872). Antimicrob Agents Chemother 41:2326–2332
Beenhouwer DO, Shapiro S, Feldmesser M, Casadevall A, Scharff MD (2001) Both Th1 and Th2 cytokines affect the ability of monoclonal antibodies to protect mice against Cryptococcus neoformans. Infect Immun 69:6445–6455
Beenhouwer DO, Yoo EM, Lai CW, Rocha MA, Morrison SL (2007) Human immunoglobulin G2 (IgG2) and IgG4, but not IgG1 or IgG3, protect mice against Cryptococcus neoformans infection. Infect Immun 75:1424–1435
Beggs WH, Sarosi GA (1982) Further evidence for sequential action of amphotericin B and 5-fluorocytosine against Candida albicans. Chemotherapy 28:341–344
Bekker LG, Freeman S, Murray PJ, Ryffel B, Kaplan G (2001) TNF-α controls intracellular mycobacterial growth by both inducible nitric oxide synthase-dependent and inducible nitric oxide synthase-independent pathways. J Immunol 166:6728–6734
Beno DW, Stover AG, Mathews HL (1995) Growth inhibition of Candida albicans hyphae by CD81 lymphocytes. J Immunol 154:5273–5281
Bink A, Kucharíková S, Neirinck B, Vleugels J, Van Dijck P, Cammue BP, Thevissen K (2012) The nonsteroidal antiinflammatory drug diclofenac potentiates the in vivo activity of caspofungin against Candida albicans biofilms. J Infect Dis 206(11):1790–1797
Bisson GP, Molefi M, Bellamy S, Thakur R, Steenhoff A, Tamuhla N, Rantleru T, Tsimako I, Gluckman S, Ravimohan S, Weissman D, Tebas P (2013) Early versus delayed antiretroviral therapy and cerebrospinal fluid fungal clearance in adults with HIV and cryptococcal meningitis. Clin Infect Dis 56(8):1165–1173
Bodasing N, Seaton RA, Shankland GS, Pithie A (2002) Gamma-interferon treatment for resistant oropharyngeal candidiasis in an HIV-positive patient. J Antimicrob Chemother 50:765–766
Bodey GP (1994) The potential role of granulocyte-macrophage colony stimulating factor in therapy of fungal infections: a commentary. Eur J Clin Microbiol Infect Dis 13:363–366
Bodey GP, Anaissie E, Gutterman J, Vadhan-Raj S (1993) Role of granulocyte macrophage colony-stimulating factor as adjuvant therapy for fungal infections in patients with cancer. Clin Infect Dis 17:705–707
Bosisio D, Polentarutti N, Sironi M, Bernasconi S, Miyake K, Webb GR, Martin MU, Mantovani A, Muzio M (2002) Stimulation of toll-like receptor 4 expression in human mononuclear phagocytes by interferon-γ: a molecular basis for priming and synergism with bacterial lipopolysaccharide. Blood 99:3427–3431
Bowyer P, Moore CB, Rautemaa R, Denning DW, Richardson MD (2011) Azole antifungal resistance today: focus on Aspergillus. Curr Infect Dis Rep 13(6):485–491
Bozza S, Montagnoli C, Gaziano R, Rossi G, Nkwanyuo G, Bellocchio S, Romani L (2004) Dendritic cell-based vaccination against opportunistic fungi. Vaccine 22:857–864
Braff MH, Bardan A, Nizet V, Gallo RL (2005) Cutaneous defense mechanisms by antimicrobial peptides. J Invest Dermatol 125:9–13
Brena S, Omaetxebarria MJ, Elguezabal N, Cabezas J, Moragues MD, Ponton J (2007) Fungicidal monoclonal antibody C7 binds to Candida albicans Als3. Infect Immun 75:3680–3682
Brena S, Cabezas-Olcoz J, Moragues MD, de Fernandez LI, Dominguez A, Quindos G, Ponton J (2011) Fungicidal monoclonal antibody C7 interferes with iron acquisition in Candida albicans. Antimicrob Agents Chemother 55:3156–3163
Brummer E, Stevens DA (1987) Activation of pulmonary macrophages for fungicidal activity by gamma-interferon or lymphokines. Clin Exp Immunol 70:520–528
Bugli F, Posteraro B, Papi M, Torelli R, Maiorana A, Sterbini FP, Posterarod P, Sanguinettia M, De Spirito M (2013) In vitro interaction between alginate lyase and amphotericin B against Aspergillus fumigatus biofilm determined by different methods. Antimicrob Agents Chemother 57(3):1275–1282
Calcedo R, Ramirez-Garcia A, Abad A, Rementeria A, Pontón J, Hernando FL (2011) Phosphoglycerate kinase and fructose bisphosphate aldolase of Candida albicans as new antigens recognized by human salivary IgA. Rev Iberoam Micol 29:172–174
Casadevall A (1995) Antibody immunity and invasive fungal infections. Infect Immun 63:4211–4218
Casadevall A, Pirofski LA (2003) Antibody-mediated regulation of cellular immunity and the inflammatory response. Trends Immunol 24:474–478
Casadevall A, Pirofski LA (2006) A reappraisal of humoral immunity based on mechanisms of antibody-mediated protection against intracellular pathogens. Adv Immunol 91:1–44
Casadevall A, Pirofski LA (2012) Immunoglobulin in defense, pathogenesis and therapy of fungal diseases. Cell Host Microbe 11:447–456
Cateau E, Berjeaud JM, Imbert C (2011) Possible role of azole and echinocandin lock solutions in the control of Candida biofilms associated with silicone. Int J Antimicrob Agents 37(4):380–384
Centre for Disease Control (CDC) (2014) http://www.cdc.gov/fungal/diseases/candidiasis/genital/index.html
Chaturvedi AK, Kavishwar A, Shiva Keshava GB, Shukla PK (2005) Monoclonal immunoglobulin G1 directed against Aspergillus fumigatus cell wall glycoprotein protects against experimental murine aspergillosis. Clin Diagn Lab Immunol 12:1063–1068
Chatzimoschou A, Katragkou Α, Simitsopoulou M, Antachopoulos C, Georgiadou E, Walsh TJ, Roilides E (2011) Activities of triazole-echinocandin combinations against Candida species in biofilms and as planktonic cells. Antimicrob Agents Chemother 55(5):1968–1974
Chavez-Dozal AA, Lown L, Jahng M, Walraven CJ, Lee SA (2014) In vitro analysis of Finasteride activity against Candida albicans urinary biofilm formation and filamentation. Antimicrob Agents Chemother 58(10):5855–5862
Chen SC, Slavin MA, Sorrell TC (2011) Echinocandin antifungal drugs in fungal infections: a comparison. Drugs 71(1):11–41
Chen X, Ren B, Chen M, Liu MX, Ren W, Wang QX, Zhang LX, Yan GY (2014) ASDCD: antifungal synergistic drug combination database. PLoS One 9(1):e86499
Cisalpino CS, Puccia R, Yamauchi LM, Cano MI, da Silva JF, Travassos LR (1996) Cloning, characterization, and epitope expression of the major diagnostic antigen of Paracoccidioides brasiliensis. J Biol Chem 271:4553–4560
Condon C, Watkins SC, Celluzzi CM, Thompson K, Falo LD (1996) DNA-based immunization by in vivo transfection of dendritic cells. Nat Med 2:1122–1127
Cornely OA, Pappas PG, Young JAH, Maddison P, Ullmann AJ (2011) Accumulated safety data of micafungin in therapy and prophylaxis in fungal diseases. Expert Opin Drug Saf 10(2):171–183
Cosgrove RF, Beezer AE, Miles RJ (1978) In vitro studies of amphotericin B in combination with the imidazole antifungal compounds clotrimazole and miconazole. J Infect Dis 138:681–685
Cox RA, Kennell W, Boncyk L, Murphy JW (1988) Induction and expression of cell-mediated immune responses in inbred mice infected with Coccidioides immitis. Infect Immun 56:13–17
Currie B, Sanati H, Ibrahim AS, Edwards JE, Casadevall A, Ghannoum MA (1995) Sterol compositions and susceptibilities to amphotericin B of environmental Cryptococcus neoformans isolates are changed by murine passage. Antimicrob Agents Chemother 39:1934–1937
Day JN, Chau TTH, Wolbers M, Mai PP, Dung NT, Mai NH, PhuNghia HD, Phong ND, Thai CQ, Thai LH, Chuong LV, Sinh DX, Duong VA, Hoang TN, Campbell JI, Sieu TPM, Baker SG, Chau NVV, Hien TT, Lalloo DG, Farrar JJ (2013) Combination antifungal therapy for cryptococcal meningitis. N Engl J Med 368(14):1291–1302
De Cremera K, Lanckackerc E, Coolsa TL, Baxa M, Bruckera KD, Cosc P, Cammuea BPA, Thevissen K (2015) Artemisinins, new miconazole potentiators resulting in increased activity against Candida albicans biofilms. Antimicrob Agents Chemother 59(1):421–426
Deepe GS (1994) Role of CD81 T cells in host resistance to systemic infection with Histoplasma capsulatum in mice. J Immunol 152:3491–3500
Deepe GS (1997) Prospects for the development of fungal vaccines. Clin Microbiol Rev 10:585–596
Delattina N, De Bruckera K, Craikb DJ, Chenevalb O, Fröhlichc M, Veberc M, Girandonc L, Davisd TR, Weeksd AE, Kumamotod CA, Cose P, Coenyef T, Conincka BD, Cammuea BPA, Thevissen K (2014) Plant-derived decapeptide OSIP108 interferes with Candida albicans biofilm formation without affecting cell viability. Antimicrob Agents Chemother 58(5):2647–2656
Denning DW, Hope WW (2010) Therapy for fungal diseases: opportunities and priorities. Trends Microbiol 18(5):195–204
Dignani MC, Rex JH, Chan KW, Dow G, deMagalhaes-Silverman M, Maddox AM, Walsh T, Anaissie E (2005) Immunomodulation with interferon-gamma and colony-stimulating factors for refractory fungal infections in patients with leukemia. Cancer 104:199–204
Djeu JY, Blanchard DK, Halkias D, Friedman H (1986) Growth inhibition of Candida albicans by human polymorphonuclear neutrophils: activation by interferon-g and tumor necrosis factor. J Immunol 137:2980–2984
Feldmesser M, Mednick A, Casadevall A (2002) Antibody-mediated protection in murine Cryptococcus neoformans infection is associated with subtle pleotrophic effects on the cytokine and leukocyte response. Infect Immun 70:1571–1580
Fine DM, Atta MG (2007) Review: kidney disease in the HIV-infected patient. AIDS Patient Care STDs 21(11):813–824
Gallin JI, Farber JM, Holland SM, Nutman TB (1995) Interferon-gamma in the management of infectious diseases. Ann Intern Med 123:216–222
Gamaletsou MN, Sipsas NV, Kontoyiannis DP, Tsiakalos A, Kontos AN, Stefanou I, Kordossis T (2012) Successful salvage therapy of refractory HIV-related cryptococcal meningitis with the combination of liposomal amphotericin B, voriconazole, and recombinant interferon-γ. Diagn Microbiol Infect Dis 74(4):409–411
Garbati MA, Alasmari FA, Al-Tannir MA, Tleyjeh IM (2012) The role of combination antifungal therapy in the treatment of invasive aspergillosis: a systematic review. Int J Infect Dis 16(2):e76–e81
Gaviria JM, Van Burik JA, Dale DC, Root RK, Liles WC (1999) Modulation of neutrophil-mediated activity against the pseudohyphal form of Candida albicans by granulocyte colony-stimulating factor (G-CSF) administered in vivo. J Infect Dis 179:1301–1304
Gazzoni AF, Capilla J, Mayayo E, Guarro J (2012) Efficacy of intrathecal administration of liposomal amphotericin B combined with voriconazole in a murine model of cryptococcal meningitis. Int J Antimicrob Agents 39(3):223–227
Giudice PL, Campo S, De Santis R, Salvatori G (2012) Effect of PTX3 and voriconazole combination in a rat model of invasive pulmonary aspergillosis. Antimicrob Agents Chemother 56(12):6400–6402
Gomez FJ, Allendoerfer R, Deepe GS (1995) Vaccination with recombinant heat shock protein 60 from Histoplasma capsulatum protects mice against pulmonary histoplasmosis. Infect Immun 63:2587–2595
Gonzalez CE, Lyman CA, Lee S, del Guercio C, Roilides E, Bacher J, Gehrt A, Feuerstein E, Tsokos M, Walsh TJ (2001) Recombinant human macrophage colony-stimulating factor augments pulmonary host defences against Aspergillus fumigatus. Cytokine 15:87–95
Govender NP, Meintjes G, Bicanic T, Dawood H, Harrison TS, Jarvis JN, Karstaedt AS, Maartens G, McCarthy KM, Rabie H, Variava E, Venter WDF, Boulware DR, Chiller T, Meya DB, Scriven J (2013) Guideline for the prevention, diagnosis and management of cryptococcal meningitis among HIV-infected persons: 2013 update. S Afr J HIV Med 14(2):76–86
Groll AH, Shah PM, Mentzel C, Schneider M, Just-Nuebling G, Huebner K (1996) Trends in the postmortem epidemiology of invasive fungal infections at a university hospital. J Infect 33:23–32
Han Y, Cutler JE (1995) Antibody response that protects against disseminated candidiasis. Infect Immun 63:2714–2719
Han Y, Ulrich MA, Cutler JE (1999) Candida albicans mannan extract-protein conjugates induce a protective immune response against experimental candidiasis. J Infect Dis 179:1477–1484
Hancock RE, Diamond G (2000) The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol 8(9):402–410
Hoag KA, Lipscomb MF, Izzo AA, Street NE (1997) IL-12 and IFN-gamma are required for initiating the protective Th1 response to pulmonary cryptococcosis in resistant C.B-17 mice. Am J Respir Cell Mol Biol 17:733–739
http://www.cdc.gov/fungal/diseases/candidiasis/thrush/index.html
Hubel K, Dale DC, Liles WC (2002) Therapeutic use of cytokines to modulate phagocyte function for the treatment of infectious diseases: current status of granulocyte colony-stimulating factor, granulocyte-macrophage colony-stimulating factor, macrophage colony-stimulating factor, and interferon-gamma. J Infect Dis 185:1490–1501
Iwasaki A, Medzhitov R (2015) Control of adaptive immunity by the innate immune system. Nat Immunol 16:343–353
Jacobson MA, Hardy D, Connick E, Watson J, DeBruin M (2000) Phase 1 trial of a single dose of recombinant human interleukin—12 in human immunodeficiency virus—infected patients with 100–500 CD4 Cells/μL. J Infect Dis 182(4):1070–1076
Jeans AR, Howard SJ, Al-Nakeeb Z, Goodwin J, Gregson L, Warn PA, Hope WW (2012) Combination of voriconazole and anidulafungin for treatment of triazole-resistant Aspergillus fumigatus in an in vitro model of invasive pulmonary aspergillosis. Antimicrob Agents Chemother 56(10):5180–5185
Jensen SB, Peterson DE (2014) Oral mucosal injury caused by cancer therapies: current management and new frontiers in research. J Oral Pathol Med 43(2):81–90
Johannsson B, Callaghan JJ (2009) Prosthetic hip infection due to Cryptococcus neoformans: case report. Diagn Microbiol Infect Dis 64(1):76–79
Johnson MD, MacDougall C, Ostrosky-Zeichner L, Perfect JR, Rex JH (2004) Combination antifungal therapy. Antimicrob Agents Chemother 48(3):693–715
Katragkou A, McCarthy M, Alexander EL, Antachopoulos C, Meletiadis J, Jabra-Rizk MA, Petraitis V, Roilides E, Walsh TJ (2015) In vitro interactions between farnesol and fluconazole, amphotericin B or micafungin against Candida albicans biofilms. J Antimicrob Chemother 70(2):470–478
Kauffman CA (2001) Fungal infections in older adults. Clin Infect Dis 33:550–555
Kawakami K, Tohyama M, Xie Q, Saito A (1996) IL-12 protects mice against pulmonary and disseminated infection caused by Cryptococcus neoformans. Clin Exp Immunol 104:208–214
Kim JS, Holtom P, Vigen C (2011) Reduction of catheter-related bloodstream infections through the use of a central venous line bundle: epidemiologic and economic consequences. Am J Infect Control 39(8):640–646
Kirkland TN, Fierer J (1985) Genetic control of resistance to Coccidioides immitis: a single gene that is expressed in spleen cells determines resistance. J Immunol 135:548–552
Kirkland TN, Zhu S, Kruse D, Hsu L, Seshan KR, Cole GT (1991) Coccidioides immitis fractions which are antigenic for immune T lymphocytes. Infect Immun 59:3952–3961
Kobayashi C, Hanadate T, Niwa T, Hirano Y, Yoshiyasu T, So M, Matsui K (2015) Safety and efficacy of micafungin for prophylaxis against invasive fungal infections in Japanese patients undergoing hematopoietic stem cell transplantation: results of a post-marketing surveillance study. J Infect Chemother 21:438–443
Kollef M, Micek S, Hampton N, Doherty JA, Kumar A (2012) Septic shock attributed to Candida infection: importance of empiric therapy and source control. Clin Infect Dis 54(12):1739–1746
Kong Y, Levine HB (1967) Experimentally induced immunity in the mycoses. Bacteriol Rev 31:35–53
Kontoyiannis DP, Lewis RE (2002) Antifungal drug resistance of pathogenic fungi. Lancet 359(9312):1135–1144
Ku TSN, Palanisamy SK, Lee SA (2010) Susceptibility of Candida albicans biofilms to azithromycin, tigecycline and vancomycin and the interaction between tigecycline and antifungals. Int J Antimicrob Agents 36(5):441–446
Kullberg BJ, Van’t Wout JW, Hoogstraten C, Hoogstraten C, van Furth R (1993) Recombinant interferon-g enhances resistance to acute disseminated Candida albicans infection in mice. J Infect Dis 168:436–443
Kullberg BJ, Oude Lashof AM, Netea MG (2004) Design of efficacy trials of cytokines in combination with antifungal drugs. Clin Infect Dis 39:S218–S223
Larrick JW, Hirata M, Balint RF, Lee J, Zhong J, Wright SC (1995) Human CAP18: a novel antimicrobial lipopolysaccharide-binding protein. Infect Immun 63:1291
Lars Tramsen1, Olaf Beck1, Friedhelm R. Schuster3,4
Laua AF, Kabira M, Chena SCA, Playford EG, Marriottd DJ, Jonese M, Lipmanf J, McBrydeg E, Gottliebh T, Cheungi W, Seppeltj I, Iredella J, Sorrell TC (2015) Candida colonization as a risk marker for invasive candidiasis in mixed medical-surgical intensive care units: development and evaluation of a simple, standard protocol. J Clin Microbiol 53(4):1324–1330
Lepak AJ, Marchillo K, VanHecker J, Andes DR (2013) Impact of in vivo triazole and echinocandin combination therapy for invasive pulmonary aspergillosis: enhanced efficacy against Cyp51 mutant isolates. Antimicrob Agents Chemother 57(11):5438–5447
Liao Z, ZhangGuan X, Zhu Z, Yao X, Yang Y, Jiang Y, Cao Y (2015) Enhancement of the antibiofilm activity of amphotericin B by polyamine biosynthesis inhibitors. Int J Antimicrob Agents (in press)
Liles WC, Huang JE, van Burik JA, Bowden RA, Dale DC (1997) Granulocyte colony stimulating factor administered in vivo augments neutrophil-mediated activity against opportunistic fungal pathogens. J Infect Dis 175:1012–1015
Liu W, Li L, Sun Y, Chen W, Wan Z, Li R, Liu W (2012) The interaction of echinocandin caspofungin with amphotericin B or voriconazole against the Aspergillus biofilms in vitro. Antimicrob Agents Chemother 56:6414–6416, AAC-00687
Liu S, Hou Y, Chen X, Gao Y, Li H, Sun S (2014) Combination of fluconazole with non-antifungal agents: a promising approach to cope with resistant Candida albicans infections and insight into new antifungal agent discovery. Int J Antimicrob Agents 43(5):395–402
Loreto ÉS, Mario DAN, Denardi LB, Alves SH, Santurio JM (2011) In vitro susceptibility of Pythium insidiosum to macrolides and tetracycline antibiotics. Antimicrob Agents Chemother 55(7):3588–3590
Loyse A, Wilson D, Meintjes G, Jarvis JN, Bicanic T, Bishop L, Rebe K, Williams A, Jaffar S, Bekker LG, Wood R, Harrison TS (2012) Comparison of the early fungicidal activity of high-dose fluconazole, voriconazole, and flucytosine as second-line drugs given in combination with amphotericin B for the treatment of HIV-associated cryptococcal meningitis. Clin Infect Dis 54(1):121–128
Lutz JE, Clemons KV, Stevens DA (2000) Enhancement of antifungal chemotherapy by interferon-gamma in experimental systemic cryptococcosis. J Antimicrob Chemother 46:437–442
Mandal SM, Migliolo L, Franco OL, Ghosh AK (2011) Identification of an antifungal peptide from Trapa natans fruits with inhibitory effects on Candida tropicalis biofilm formation. Peptides 32(8):1741–1747
Marcil A, Harcus D, Thomas DY, Whiteway M (2002) Candida albicans killing by RAW 264.7 mouse macrophage cells: effects of Candida genotype, infection ratios, and gamma interferon treatment. Infect Immun 70:6319–6329
Marr KA, Schlamm HT, Herbrecht R, Rottinghaus ST, Bow EJ, Cornely OA, Heinz WJ, Jagannatha S, Koh LP, Kontoyiannis DP, Lee DG, Nucci M, Pappas PG, Slavin MA, Queiroz-Telles F, Selleslag D, Walsh TJ, Wingard JR, Maertens JA (2015) Combination antifungal therapy for invasive aspergillosis: a randomized trial. Ann Intern Med 162(2):81–89
Martinez LR, Casadevall A (2005) Specific antibody can prevent fungal biofilm formation and this effect correlates with protective efficacy. Infect Immun 73:6350–6362
Martinez LR, Fries BC (2010) Fungal biofilms: relevance in the setting of human disease. Curr Fungal Infect Reports 4(4):266–275
Martinez LR, Moussai D, Casadevall A (2004) Antibody to Cryptococcus neoformans glucuronoxylomannan inhibits the release of capsular antigen. Infect Immun 72:3674–3679
Matthews RC, Burnie JP, Howat D, Rowland T, Walton F (1991) Autoantibody to heat shock protein 90 can mediate protection against systemic candidosis. Immunology 74:20–24
Mavridou E, Meletiadis J, Rijs A, Mouton JW, Verweij PE (2015) The strength of synergistic interaction between Posaconazole and Caspofungin depends on the underlying azole-resistance mechanism of Aspergillus fumigatus. Antimicrob Agents Chemother 59:1738–1744, AAC-04469
McClelland EE, Nicola AM, Prados-Rosales R, Casadevall A (2010) Ab binding alters gene expression in Cryptococcus neoformans and directly modulates fungal metabolism. J Clin Invest 120:1355–1361
Medoff G (1983) Antifungal action of rifampin. Rev Infect Dis J 5(suppl. 3):S614–S619
Mencacci A, Spaccapelo R, Del Sero G, Enssle KH, Cassone A, Bistoni F, Romani L (1996) CD41 T-helper-cell responses in mice with low-level Candida albicans infection. Infect Immun 64:4907–4917
Mencacci A, del Sero G, Cenci E, d’Ostiani CF, Bacci A, Montagnoli C, Kopf M, Romani L (1998) Endogenous IL-4 is required for development of protective CD4+ T helper type 1 cell responses to Candida albicans. J Exp Med 187:307–317
Mendling W (2015) Guideline: vulvovaginal candidosis (AWMF 015/072), S2k (excluding chronic mucocutaneous candidosis). Mycoses 58(1):1–15
Miceli MH, Díaz JA, Lee SA (2011) Emerging opportunistic yeast infections. Lancet Infect Dis 11(2):142–151
Mihu CN, Kassis C, Ramos ER, Jiang Y, Hachem RY, Raad II (2010) Does combination of lipid formulation of amphotericin B and echinocandins improve outcome of invasive aspergillosis in hematological malignancy patients? Cancer 116(22):5290–5296
Milam JE, Herring-Palmer AC, Pandrangi R, McDonald RA, Huffnagle GB, Toews GB (2007) Modulation of the pulmonary type 2 T-cell response to Cryptococcus neoformans by intratracheal delivery of a tumor necrosis factor alpha-expressing adenoviral vector. Infect Immun 75:4951–4958
Morein B, Sundquist B, Hoglund S, Dalsgaard K, Osterhaus A (1984) Iscom, a novel structure for antigenic presentation of membrane proteins from enveloped viruses. Nature 308:457–460
Morrison LA, Lukacher AE, Braciale VL, Fan DP, Braciale TJ (1986) Differences in antigen presentation to MHC class I- and class II-restricted influenza virus-specific cytolytic T lymphocyte clones. J Exp Med 163:903–921
Mowat E, Butcher J, Lang S, Williams C, Ramage G (2007) Development of a simple model for studying the effects of antifungal agents on multicellular communities of Aspergillus fumigatus. J Med Microbiol 56(9):1205–1212
Muchmore HG, Felton FG, Salvin SB, Rhoades ER (1968) Delayed hypersensitivity to cryptococcin in man. Sabouraudia 6:285–288
Muller U, Stenzel W, Kohler G, Werner C, Polte T, Hansen G (2007) IL-13 induces disease-promoting type 2 cytokines, alternatively activated macrophages and allergic inflammation during pulmonary infection of mice with Cryptococcus neoformans. J Immunol 179:5367–5377
Muñoz A, Marcos JF, Read ND (2012) Concentration‐dependent mechanisms of cell penetration and killing by the de novo designed antifungal hexapeptide PAF26. Mol Microbiol 85(1):89–106
Murphy JW, Hidore MR, Wong SC (1993) Direct interactions of human lymphocytes with the yeast-like organism, Cryptococcus neoformans. J Clin Invest 91:1553–1566
Muzoora CK, Kabanda T, Ortu G, Ssentamu J, Hearn P, Mwesigye J, Longleya N, Jarvisb JN, Jaffard S, Harrisonb TS (2012) Short course amphotericin B with high dose fluconazole for HIV-associated cryptococcal meningitis. J Infect 64(1):76–81
Nabavi N, Murphy JW (1986) Antibody-dependent natural killer cell-mediated growth inhibition of Cryptococcus neoformans. Infect Immun 51:556–562
Nagaoka I, Hirota S, Niyonsaba F, Hirata M, Adachi Y, Tamura H, Heumann D (2001) Cathelicidin family of antibacterial peptides CAP18 and CAP11 inhibit the expression of TNF-α by blocking the binding of LPS to CD14+ cells. J Immunol 167:3329
Netea MG, Brouwer AE, Hoogendoorn EH, Van der Meer JW, Koolen M, Verweij PE, Kullberg BJ (2004) Two patients with cryptococcal meningitis and idiopathic CD4 lymphopenia: defective cytokine production and reversal by recombinant interferon- gamma therapy. Clin Infect Dis 39:e83–e87
Netea MG, Van der Meer JW, Kullberg BJ (2006) Role of the dual interaction of fungal pathogens with pattern recognition receptors in the activation and modulation of host defence. Clin Microbiol Infect 12:404–409
Nett JE (2014) Future directions for anti-biofilm therapeutics targeting Candida. Expert Rev Anti-Infect Ther 12(3):375–382
Nikapitiya C (2012) Bioactive secondary metabolites from marine microbes for drug discovery. Adv Food Nutr Res 65(8):363–387
Nucci M, Anaissie E, Betts RF, Dupont BF, Wu C, Buell DN, Kovanda L, Lortholary O (2010) Early removal of central venous catheter in patients with candidemia does not improve outcome: analysis of 842 patients from 2 randomized clinical trials. Clin Infect Dis 51(3):295–303
Ogawa T, Matsumoto K, Tsujimoto K, Hishiya N, Yamada Y, Uno K, Kasaharaa K, Maedaa K, Nariob K, Mikasaa K, Morita K (2015) Chronic invasive sinus and intracerebral aspergillosis controlled by combination therapy with micafungin and a daily dose of 400 mg itraconazole oral solution. J Infect Chemother 21(2):134–137
Olson JA, Schwartz JA, Hahka D, Nguyen N, Bunch T, Jensen GM, Adler-Moore JP (2015) Toxicity and efficacy differences between liposomal amphotericin B formulations in uninfected and Aspergillus fumigatus infected mice. Med Mycol 53(2):107–118
Owais M, Ansari MA, Ahmad I, Zia Q, Pierard G, Chauhan A et al (2010) Innate immunity in pathogenesis and treatment of dermatomycosis. In: Ahmad I (ed) Combating fungal infections. Springer-Verlag, Berlin. doi:10.1007/978-3-642-12173-9_10
Pachl J, Svoboda P, Jacobs F, Vandewoude K, Hoven BV, Spronk P, Masterson G, Malbrain M, Aoun M, Garbino J, Takala J, Drgona L, Burnie J, Matthews R (2006) A randomized, blinded, multicenter trial of lipid-associated amphotericin B alone versus in combination with an antibody-based inhibitor of heat shock protein 90 in patients with invasive candidiasis. Clin Infect Dis 42(10):1404–1413
Pagano L, Akova M, Dimopoulos G, Herbrecht R, Drgona L, Blijlevens N (2011) Risk assessment and prognostic factors for mould-related diseases in immunocompromised patients. J Antimicrob Chemother 66:i5–i14
Pappas PG, Rex JH, Sobel JD, Filler SG, Dismukes WE, Walsh TJ, Edwards JE (2004) Guidelines for treatment of candidiasis. Clin Infect Dis 38:161–189
Paramythiotou E, Frantzeskaki F, Flevari A, Armaganidis A, Dimopoulos G (2014) Invasive fungal infections in the ICU: how to approach, how to treat. Molecules 19(1):1085–1119
Perfect JR, William ED, Francoise D, David LG, John RG, Richard JH, Thomas SH, RA Larsen RA, Lortholary O, Nguyen MH, Pappas PG, Powderly WG, Singh N, Sobel JD, Sorrell TC (2010) Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the Infectious Diseases Society of America. Clin Infect Dis 50(3):291–322
Pfaller M, Neofytos D, Diekema D, Azie N, Meier-Kriesche HU, Quan SP, Horn D (2012) Epidemiology and outcomes of candidemia in 3648 patients: data from the Prospective Antifungal Therapy (PATH Alliance®) registry, 2004–2008. Diagn Microbiol Infect Dis 74(4):323–331
Pieralisi N, Godoy J, Yamada S, Santana R, Svidzinski T (2014) Oral lesions and colonization by yeasts in hemodialysis patients. J Oral Pathol Med 44(8): 585–590
Polak-Wyss A (1991) Protective effect of human granulocyte colony-stimulating factor (hG-CSF) on Cryptococcus and Aspergillus infections in normal and immunosuppressed mice. Mycoses 34:205–215
Probst C, Pongratz G, Capellino S, Szeimies RM, Scholmerich J, Fleck M, Salzberger B, Ehrenstein B (2010) Cryptococcosis mimicking cutaneous cellulitis in a patient suffering from rheumatoid arthritis: a case report. BMC Infect Dis 10(1):239
Qureshi MH, Zhang T, Koguchi Y, Nakashima K, Okamura H, Kurimoto M, Kawakami K (1999) Combined effects of IL-12 and IL-18 on the clinical course and local cytokine production in murine pulmonary infection with Cryptococcus neoformans. Eur J Immunol 29:643–649
Rachini A, Pietrella D, Lupo P, Torosantucci A, Chiani P, Bromuro C, Proietti C, Bistoni F, Cassone A, Vecchiarelli A (2007) An anti-beta-glucan monoclonal antibody inhibits growth and capsule formation of Cryptococcus neoformans in vitro and exerts therapeutic, anticryptococcal activity in vivo. Infect Immun 75:5085–5094
Rajendran R, Mowat E, Jones B, Williams C, Ramage G (2015) Prior in vitro exposure to voriconazole confers resistance to amphotericin B in Aspergillus fumigatus biofilms. Int J Antimicrob Agents 46:342–345
Reardon CC, Kim SJ, Wagner RP, Kornfeld H (1996) Interferon-gamma reduces the capacity of human alveolar macrophages to inhibit growth of Cryptococcus neoformans in vitro. Am J Respir Cell Mol Biol 15:711–715
Redmond HP, Shou J, Gallagher HJ, Kelly CJ, Daly JM (1993) Macrophage-dependent Candidacidal mechanisms in the murine system. Comparison of murine Kupffer cell and peritoneal macrophage Candidacidal mechanisms. J Immunol 150:3427–3433
Reedy JL, Filler SG, Heitman J (2010) Elucidating the Candida albicans calcineurin signaling cascade controlling stress response and virulence. Fungal Genet Biol 47(2):107–116
Riddell J, Shuman EK (2012) Epidemiology of central nervous system infection. Neuroimaging Clin N Am 22(4):543–556
Riddell LA, Pinching AJ, Hill S, Ng TT, Arbe E, Lapham GP, Ash S, Hillman R, Tchamouroff S, Denning DW, Parkin JM (2001) A phase III study of recombinant human interferon gamma to prevent opportunistic infections in advanced HIV disease. AIDS Res Hum Retroviruses 17:789–797
Rivera J, Zaragoza O, Casadevall A (2005) Antibody-mediated protection against Cryptococcus neoformans pulmonary infection is dependent on B cells. Infect Immun 73:1141–1150
Rodrigues ML, Travassos LR, Miranda KR, Franzen AJ, Rozental S, De Souza W, Alviano CS, Barreto-Bergter E (2000) Human antibodies against a purified glucosylceramide from Cryptococcus neoformans inhibit cell budding and fungal growth. Infect Immun 68:7049–7060
Rodriguez-Adrian LJ, Grazziutti ML, Rex JH, Anaissie EJ (1998) The potential role of cytokine therapy for fungal infection in patients with cancer: is recovery from neutropenia all that is needed? Clin Infect Dis 26:1270–1278
Rohatgi S, Pirofski LA (2015) Host immunity to Cryptococcus neoformans. Future Microbiol 10:565–581
Romani L (2000) Innate and adaptive immunity in Candida albicans infections and saprophytism. J Leukoc Biol 68:175–178
Rosas AL, Nosanchuk JD, Casadevall A (2001) Passive immunization with melanin-binding monoclonal antibodies prolongs survival in mice with lethal Cryptococcus neoformans infection. Infect Immun 69:3410–3412
Rossignol T, Kelly B, Dobson C, d’Enfert C (2011) Endocytosis-mediated vacuolar accumulation of the human ApoE apolipoprotein-derived ApoEdpL-W antimicrobial peptide contributes to its antifungal activity in Candida albicans. Antimicrob Agents Chemother 55(10):4670–4681
Rowe JM, Andersen JW, Mazza JJ, Bennett JM, Paietta E, Hayes FA, Oette D, Cassileth PA, Stadtmauer EA, Wiernik PH (1995) A randomized, placebo-controlled phase III study of granulocyte–macrophage colony-stimulating factor in adult patients (15 to 70 years of age) with acute myelogenous leukemia: a study of the Eastern Cooperative Oncology Group (E1490). Blood 86:457–462
Sanati H, Ramos CF, Bayer AS, Ghannoum MA (1997) Combination therapy with amphotericin B and fluconazole against invasive candidiasis in neutropenic-mouse and infective-endocarditis rabbit models. Antimicrob Agents Chemother 41(6):1345–1348
Sandini S, La VR, Deaglio S, Malavasi F, Cassone A, De BF (2011) A highly immunogenic recombinant and truncated protein of the secreted aspartic proteases family (rSap2t) of Candida albicans as a mucosal anticandidal vaccine. FEMS Immunol Med Microbiol 62:215–224
Sanglard D, Coste A, Ferrari S (2009) Antifungal drug resistance mechanisms in fungal pathogens from the perspective of transcriptional gene regulation. FEMS Yeast Res 9(7):1029–1050
Sardi JCO, Scorzoni L, Bernardi T, Fusco-Almeida AM, Giannini MM (2013) Candida species: current epidemiology, pathogenicity, biofilm formation, natural antifungal products and new therapeutic options. J Med Microbiol 62(1):10–24
Schlagetter AM, Kozel TR (1990) Opsonization of Cryptococcus neoformans by a family of isotype-switch variant antibodies specific for the capsular polysaccharide. Infect Immun 58:1914–1918
Schlamm HT, Marr KA (2015) Combination therapy for invasive aspergillosis: controversies and conclusions. Curr Fungal Infect Rep 9(2):130–134
Semis R, Nahmias M, Lev S, Frenkel M, Segal E (2015) Evaluation of antifungal combinations of nystatin-intralipid against Aspergillus terreus using checkerboard and disk diffusion methods. J Mycol Méd/J Med Mycol 25(1):63–70
Shadomy S, Wagner G, Espinel-Ingroff E, Davis BA (1975) In vitro studies with combinations of 5-fluorocytosine and amphotericin B. Antimicrob Agents Chemother 8:117–121
Shahid M, Tripathi T, Khardori N, Huq A, Ahmad I et al (2010) Cytokine therapy: possible tools in management of fungal infection. In: Ahmad I (ed) Combating fungal infections. Springer, Berlin. doi:10.1007/978-3-642-12173-9_16
Shapiro S, Beenhouwer DO, Feldmesser M, Taborda C, Carroll MC, Casadevall A, Scharff MD (2002) Immunoglobulin G monoclonal antibodies to Cryptococcus neoformans protect mice deficient in complement component C3. Infect Immun 70:2598–2604
Silva EG, Paula CR, Dias ALT, Chang MR, da Silva Ruiz L, Gambale V, Prates RA, Ribeiro MS (2011) Combination efficacy of voriconazole and amphotericin B in the experimental disease in immunodeficient mice caused by fluconazole-resistant Cryptococcus neoformans. Mycopathologia 171(4):261–266
Singh R, Shivaprakash MR, Chakrabarti A (2011) Biofilm formation by zygomycetes: quantification, structure and matrix composition. Microbiology 157(9):2611–2618
Siopi M, Siafakas N, Vourli S, Zerva L, Meletiadis J (2015) Optimization of polyene-azole combination therapy against aspergillosis using an in vitro pharmacokinetic-pharmacodynamic model. Antimicrob Agents Chemother 59:3973–3983, AAC-05035
Sorensen OE, Arnljots K, Cowland JB, Bainton DF, Borregaard N (1997) The human antibacterial cathelicidin, hCAP-18, is synthesized in myelocytes and metamyelocytes and localized to specific granules in neutrophils. Blood 90:2796
Steinbach WJ, Stevens DA (2003) Review of newer antifungal and immunomodulatory strategies for invasive aspergillosis. Clin Infect Dis 37:S157–S187
Tascon RE, Colston MJ, Ragno S, Stavropoulos E, Gregory D, Lowrie D (1996) Vaccination against tuberculosis by DNA injection. Nat Med 2:888–898
Torosantucci A, Bromuro C, Chiani P, De Bernardis F, Berti F, Galli C, Norelli F, Bellucci C, Polonelli L, Costantino P, Rappuoli R, Cassone A (2005) A novel glyco-conjugate vaccine against fungal pathogens. J Exp Med 202:597–606
Torosantucci A, Chiani P, Bromuro C, De BF, Palma AS, Liu Y, Mignogna G, Maras B, Colone M, Stringaro A, Zamboni S, Feizi T, Cassone A (2009) Protection by anti-beta-glucan antibodies is associated with restricted beta-1,3 glucan binding specificity and inhibition of fungal growth and adherence. PLoS One 4:e5392
Tragiannidis A, Tsoulas C, Groll AH (2015) Invasive candidiasis and Candidaemia in neonates and children: update on current guidelines. Mycoses 58(1):10–21
Tramsen L, Beck O, Schuster FR, Hunfeld KP, Latgé JP, Sarfati J, Röger F, Klingebiel T, Koehl U, Lehrnbeche T (2007) Generation and characterization of anti-Candida T cells as potential immunotherapy in patients with Candida infection after allogeneic hematopoietic stem-cell transplant. J Infect Dis 196:485–492
Truan G, Epinat JC, Rougeulle C, Cullin C, Pompon D (1994) Cloning and characterization of a yeast cytochrome b5 encoding gene which suppresses ketoconazole hypersensitivity in a NADPH P-450 reductase deficient strain. Gene 142:123–127
Tsiodras S, Samonis G, Boumpas DT (2008) Fungal infections complicating tumor necrosis factor α blockade therapy. Mayo Clin Proc 83:181–194
Ullmer JB, Donnelly JJ, Parker SE, Rhodes GH, Felgner PL, Dwarki VJ, Gromkowski SH, Deck RR, DeWitt CM, Friedman A, Hawe LA, Leander KR, Martinez D, Perry HC, Shiver JW, Montgomery DL, Liu MA (1993) Heterologous protection against influenza by injection of DNA encoding a viral protein. Science 259:1745–1749
Vaidhya SA, Gupta BB, Jha RK, Kumar R (2015) Combination versus Monotherapy for the treatment of HIV associated cryptococcal meningitis. J Clin Diagn Res: JCDR 9(2):14
Valerio M, Vena A, Bouza E, Reiter N, Viale P, Hochreiter M, Giannella M, Muñoz P (2015) How much European prescribing physicians know about invasive fungal infections management? BMC Infect Dis 15(1):80
Van De Veerdonk FL, Netea MG, Joosten LA, Van Der Meer LWM, Kullberg BJ (2010) Novel strategies for the prevention and treatment of Candida infections: the potential of immunotherapy. FEMS Microbiol Rev 34:1063–1075
van der Meer JW, Popa C, Netea MG (2005) Side effects of anticytokine strategies. Neth J Med 63:78–80
Van Wetering S, Mannesse-Lazeroms SP, Van Sterkenburg MA, Daha MR, Dijkman JH, Hiemstra PS (1997) Effect of defensins on interleukin-8 synthesis in airway epithelial cells. Am J Physiol 272:L888
Vazquez JA, Hidalgo JA, De Bono S (2000) Use of sargramostim (rh-GM-CSF) as adjunctive treatment of fluconazole-refractory oropharyngeal candidiasis in patients with AIDS: a pilot study. HIV Clin Trials 1:23–29
Walsh TJ, Gamaletsou MN (2013) Treatment of fungal disease in the setting of neutropenia. ASH Educ Program Book 1:423–427
Wang Y, Damu GL, Lv JS, Geng RX, Yang DC, Zhou CH (2012) Design, synthesis and evaluation of clinafloxacin triazole hybrids as a new type of antibacterial and antifungal agents. Bioorg Med Chem Lett 22(17):5363–5366
Welte K, Gabrilove J, Bronchud MH, Platzer E, Morstyn G (1996) Filgrastim (r-metHuG-CSF): the first 10 years. Blood 88:1907–1929
Wiederhold NP, Najvar LK, Bocanegra R, Kirkpatrick WR, Sorrell TC, Patterson TF (2013) Limited activity of miltefosine in murine models of cryptococcal meningoencephalitis and disseminated cryptococcosis. Antimicrob Agents Chemother 57(2):745–750
Wormley FL, Perfect JR, Steele C, Cox GM (2007) Protection against cryptococcosis by using a murine gamma interferon-producing Cryptococcus neoformans strain. Infect Immun 75(3):1453–1462
Yao ZW, Lu X, Shen C, Lin DF (2014) Comparison of flucytosine and fluconazole combined with amphotericin B for the treatment of HIV-associated cryptococcal meningitis: a systematic review and meta-analysis. Eur J Clin Microbiol Infect Dis 33(8):1339–1344
Zhang M, Su X, Sun WK, Chen F, Xu XY, Shi Y (2014) Efficacy of the combination of voriconazole and caspofungin in experimental pulmonary Aspergillosis by different Aspergillus species. Mycopathologia 177(1–2):11–18
Author information
Authors and Affiliations
Corresponding authors
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer India
About this chapter
Cite this chapter
Rathore, S.S., Ramakrishnan, J., Raman, T. (2016). Recent Advancements in Combinational Antifungal Therapy and Immunotherapy. In: Basak, A., Chakraborty, R., Mandal, S. (eds) Recent Trends in Antifungal Agents and Antifungal Therapy. Springer, New Delhi. https://doi.org/10.1007/978-81-322-2782-3_6
Download citation
DOI: https://doi.org/10.1007/978-81-322-2782-3_6
Published:
Publisher Name: Springer, New Delhi
Print ISBN: 978-81-322-2780-9
Online ISBN: 978-81-322-2782-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)