
Abstract
This chapter revealed the efficacy of three predominant dragonfly species found in a natural population where the survey of mosquito population was conducted. Nymphs of dragonflies belonging to family Libellulidae, Neurothemis fluctuans, Orthetrum sabina, and Orthetrum chrysis, were used as predators on the IV instar of mosquito larvae, Aedes albopictus, Aedes aegypti, and Culex quinquefasciatus. The daily feeding rates varied among predators and mosquito species. The mean numbers of mosquito larvae consumed by the predators were different between the mosquito species. Aedes aegypti was the most preferred prey for Orthetrum sabina and Neurothemis fluctuans. However, Orthetrum chrysis consumed more of Culex quinquefasciatus in contrast to other prey species. Feeding activities peaked during light-on in contrast to light-off. The results of variation factors that influenced the predation activities were significant and further discussed in this chapter. The factors that were assessed in the experiments included the water volume, predator species, predator density, and prey density and species. This chapter lends support to the potential use of Odonata species as an eco-friendly method of mosquito population eradication.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Mosquitoes play a significant role from the standpoint of human welfare because the females are bloodsucking in which many species bite people and at the same time serve as vectors in transmission of several fatal human diseases (Triplehorn and Johnson 2005). A number of Aedes (Stegomyia) spp. may act as a vector in these situations, depending on the geographic area, including A. aegypti, A. albopictus, A. polynesiensis, and other members of the A. scutellaris group. The serious transmission cycle from a public health standpoint is the urban endemic/epidemic cycle in large urban centers of the tropics. Humans are infected with dengue viruses when bitten by an infective mosquito. A. aegypti, the principal vector of dengue, is small in size with black-white body striations and a highly domesticated tropical mosquito that prefers to lay eggs in artificial containers commonly found in and around homes, for example, flower vases, old automobile tires, buckets, and trash that collect rainwater in general (Gubler 1998). In summary, demographic and societal changes, decreasing resources for vector-borne infectious disease prevention and control, and changes in public health policy have all contributed to increased epidemic dengue activity (Gubler 1998).
Normal chemical controls that were used in the eradication of adult mosquitoes were fogging with DDT and larvicide, for the larvae stages (Jatanasen 1997). Larviciding, the application of chemicals to kill mosquito larvae or pupae in the water, is generally more effective and target specific than applying chemicals to kill adult mosquitoes (adulticiding) but seemed less permanent. Adulticiding is usually the least efficient mosquito control technique. However, it is the only way to kill adult mosquitoes and is the last line of defense in reducing mosquito populations. Adulticides are typically applied as an ultralow-volume (ULV) spray where small amounts of insecticides dispersed by either truck-mounted equipment or aircraft. Adulticides labeled for mosquito control included the organophosphates malathion and naled, some natural pyrethrins, and synthetic pyrethroids (permethrin, resmethrin, and sumithrin). Insecticide selection and time of application should be based on the distribution and behavior of the target mosquito species (Dykstra 2008). Many synthetic insecticides are widely used for controlling adult and larval mosquito populations. However, the harmful effects of chemicals on nontarget populations and the development of resistance to these chemicals in mosquitoes along with the recent resurgence of different mosquito-borne diseases as reported by Milam et al. (2000) have prompted many researches to explore alternative, simple, sustainable methods for the potential control of mosquitoes.
Utilizing biological organisms to control mosquitoes had been proven, not only to be eco-friendly but as well constitute means by which more effective and sustainable control can be achieved. Control of mosquito larvae with biological agents that are their competitors and predators is more convenient and alleviates the need for frequent chemical applications (Kumar and Hwang 2006). There are various organisms that can act as biocontrol agents to control mosquito populations, thus avoiding the use of chemicals that are harmful to the environment. It is desirable to use biocontrol agents that can adapt to the mosquito breeding habitats, which are found naturally, and pose no danger to humans living in the area (Rishikesh et al. 1988, Spielman et al. 1993). Many biocontrol agents are able to disperse by themselves enabling them to spread and build up viable populations (Caltagirone 1981, Bellows 2001, Headrick and Goeden 2001).
Medlock and Snow (2008) identified five categories of possible mosquito habitats: permanent freshwater, temporary woodland pools or flooded habitats, brackish water salt marshes, artificial container habitats, and phytotelmatas. Each of these natural or artificial places would have their specific food webs including the mosquitoes populating these habitats that are preyed upon by varied predator types. The tree holes and bamboo nodes which are in the phytotelmata category are specialized habitats known to be exploited by Aedes mosquitoes (Yates 1979), not easily reached by normal predators, but there had been reports by Corbet (1999) and Juliano and Gravel (2002) that dragonflies were able to disperse to these specialized niche areas for oviposition behavior.
Dragonflies (Order: Odonata) are known not only for their dispersal capability but also for their predaceous behavior as adults as well as immature stages. The females will seek water bodies to lay their eggs, and once hatched the Odonata larvae are voracious and known to be important predators of mosquito larvae living in sympatric. Water bodies that are utilized by females to lay eggs are not limited to large or flowing waters but instead could be confined in stagnant aquatic habitats, man-made or natural, that is simultaneously exploited by container-dwelling mosquitoes. The dragonflies and damselflies are true enemies of mosquitoes as the odonate larvae are able to prey on the mosquito larvae (Breene et al. 1990) and the adults are efficient predators of airborne adult mosquitoes. One example of a dragonfly that had been investigated by Singh et al. (2010) as biocontrol agent was the larvae of Brachythemis contaminata (Libellulidae) that efficiently preyed upon the larvae of Anopheles stephensi, Culex quinquefasciatus, and Aedes aegypti. The frequently cited success story of dragonflies as biocontrol agents was the work of Sebastian et al. (1990) that was conducted in a country, formerly known as Myanmar (now Burma). They released dragonfly larvae into water held containers in homes where there were known to accommodate Ae. aegypti mosquitoes. The resulting effect was the rapid disappearance of mosquito larvae with the introduction of the dragonfly predators in which led to a strategic management program of mosquito control by a systematic release of dragonfly larvae during the monsoon season (the time when dengue fever was being transmitted by the mosquito vectors). There was an impressive drop in mosquito infestations that would not be achieved even with the traditional use of chemical insecticidal treatments.
Although odonate larvae had not been investigated as much as the utilization of guppies or other predaceous aquatic insects as mosquito biocontrol, but their long life cycle, high predation capacity and sharing of habitats with mosquito immatures as well as adults made them highly appropriate for consideration as biological control agents.
2 Methodology Followed
The species of dragonfly nymphs that were used in these experiments came from the family group of Libellulidae, known to thrive in stagnant waters: Orthetrum chrysis, Orthetrum sabina, and Neurothemis fluctuans which were aptly the dominant species found in the natural study areas where mosquito monitoring studies were conducted, the work reported elsewhere (Saleeza et al. 2011). All the individuals used in the experiments were measured for their body lengths and widths using a digital caliper prior to experiments to ensure constant or standard selection for species group’s body size. The mosquito larvae and their predator dragonfly nymphs were maintained and kept in laboratory aquariums separately.
Three species of dragonfly nymphs were exposed with all three species of mosquito larvae Ae. albopictus, Ae. aegypti, and Cx. quinquefasciatus within their individual isolated aquaria. Nine aquaria were used which contained pond water and were oxygenated using air pumps. During the experiment, three species of dragonfly nymphs Orthetrum chrysis, Orthetrum sabina, and Neurothemis fluctuans were allowed to feed on 100 IV instar mosquito larvae of Ae. albopictus, Ae. aegypti, and Cx. quinquefasciatus. During the duration of experiments, the water temperatures ranged from 24 to 29 °C, pH at levels of 6.1–6.3, and dissolved oxygen maintained at 5.2–6.3 mg/l. The number of mosquito larvae consumed by each of the dragonfly nymphs was counted at every 3 h interval for the total period of 24 h, in three replicates. The duration of time taken for first attack by each dragonfly nymph to approach or consume the prey was recorded. The numbers of mosquito larvae ingested by the dragonfly nymphs were obtained by pouring the aquaria water through a fine mesh sieve to collect all of the mosquito larvae that were not consumed and immediately transferred to a white pan for counting. After each 3 h interval, the aquaria were replenished with the number of larvae that were eaten, along with the same volume of water, to maintain the same prey density. This experiment was conducted three times on three separate days (n = 3) with the same number of nymphs for accuracy. After 24 h, all remaining mosquito larvae and dragonfly predators were removed from the aquaria. These mosquito larvae and dragonfly nymphs were not used in subsequent experiments. The active period of dragonfly feeding on mosquitoes was determined by exposure to light and dark hours, that is, 12 h in the daylight and 12 h in the nighttime. The experiments conducted revealed the prey-predation relationships, feeding rate, predators active foraging times, as well as the exposures to all three species of the mosquitoes: Ae. albopictus, Ae. aegypti, and Cx. quinquefasciatus would enable preferred prey choice to be determined.
In a separate experiment to assess the predation efficiency when exposed to various conditions, a total of 36 aquaria were used, according to the protocols as listed below. This experiment was conducted in three replicates: Aquaria A, Orthetrum chrysis (1 × 1 × 100), single-dragonfly nymph with 1 L of water volume and 100 IV instar of mosquito larvae; Aquaria B, Orthetrum chrysis (1 × 2 × 100), single-dragonfly nymph with 2 L of water volume and 100 IV instar of mosquito larvae; Aquaria C, Orthetrum chrysis (2 × 1 × 100), two-dragonfly nymph with 2 L of water volume and 100 IV instar of mosquito larvae; Aquaria D, Orthetrum chrysis (1 × 1 × 200), single-dragonfly nymph with 2 L of water volume and 200 IV instar of mosquito larvae; Aquaria E, Orthetrum sabina (1 × 1 × 100), single-dragonfly nymph with 1 L of water volume and 100 IV instar of mosquito larvae; Aquaria F, Orthetrum sabina (1 × 2 × 100), single-dragonfly nymph with 2 L of water volume and 100 IV instar of mosquito larvae; Aquaria G, Orthetrum sabina (2 × 1 × 100), two-dragonfly nymph with 1 L of water volume and 100 IV instar of mosquito larvae; Aquaria H, Orthetrum sabina (1 × 1 × 200), single-dragonfly nymph with 1 L of water volume and 200 IV instar of mosquito larvae; Aquaria I, Neurothemis fluctuans (1 × 1 × 100), single-dragonfly nymph with 1 L of water volume and 100 IV instar of mosquito larvae; Aquaria J, Neurothemis fluctuans (1 × 2 × 100), single-dragonfly nymph with 2 L of water volume and 100 IV instar of mosquito larvae; Aquaria K, Neurothemis fluctuans (2 × 1 × 100), two-dragonfly nymph with 1 L of water volume and 100 IV instar of mosquito larvae; and Aquaria L, Neurothemis fluctuans (1 × 1 × 200), single-dragonfly nymph with 1 L of water volume and 200 IV instar of mosquito larvae.
3 Results
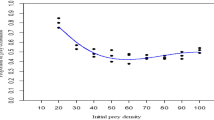
Figure 13.1 showed the number of mosquito larvae species consumed by three species of Odonata Neurothemis fluctuans, Orthetrum sabina, and Orthetrum chrysis. Overall, Orthetrum sabina consumed the highest number of mosquitoes. Both Neurothemis fluctuans and Orthetrum sabina preferred Ae. aegypti in contrast to Orthetrum chrysis which was skewed toward feeding on Cx. quinquefasciatus larvae. Overall, the dragonfly species showed daylight active feeding in contrast to dark hours (t test: T(7,9) = 2.80, p = 0.01) and this contrast was most apparent in Aedes aegypti (t test: T(1,3) = 27.02, p = 0.01) when compared to dragonflies consumption on other prey species.

Number of three types of mosquito larvae species consumed by three species of dragonfly predators
Several interesting outcomes were apparent in the experimental setup where three species of Odonata were exposed and tested with multiple variables in terms of differences in mosquito species, water volume, number of predators, and mosquito densities. Overall, the predation activities that had significant result outcomes were for the differences in prey densities and when increased number of predators were present, resulting in obvious competition between them [multiple correlation coefficient (R): Y = 72.44–25.92X1 + 14.22X2 = 0.72, where Y = predation, X1 = mosquito densities, and X2 = number of predators]. Table 13.1 shows the regression equation for the experiment with different species of Odonata. Here, when the predators were Orthetrum sabina (Y = 66.22–27.66 X1 + 23.77 X2; R = 0.82) and Orthetrum chrysis (Y = 61.00–19.88 X1 + 16.33 X2; R = 0.65), the significant factors that influenced predation performance were water volume and number of predators. However, for Neurothemis fluctuans (Y = 94.370–31.07 X3; R = 0.77), it was the prey densities that significantly affected predation activities, where there is higher predator feeding rate with increased prey densities (R = 0.77).
Tables 13.1, 13.2, and 13.3 reveal the mean number of predation activities ± SE by three different Odonata species on the three different types of prey (different mosquito species) with variations in water volume, number of predators, and mosquito densities. For the predators, Orthetrum sabina and Orthetrum chrysis, they showed increased predatory performances with high water volume and in contrast lowered their performances when in the presence of competitors (two individuals present). Interestingly, Neurothemis fluctuans predatory behaviors were affected by the increment of prey densities where they showed frenzy feeding acts with higher prey densities, from 100 to 200 mosquito larvae, and the larvae consumption increased until satiation levels were achieved.
4 Discussions
Investigations on aspects of biocontrol against mosquito larvae had been well documented, and the interests were not only confined to the tropical areas. A number of different predators were studied for their potentials as biological control agents, for example, Rhantus sikkimensis and larvae of Toxorhynchites splendens (Aditya et al. 2006; Aditya et al. 2007), Diplonychus sp. and Anisops sp. (Shaalan et al. 2007), odonate nymphs (Chandra et al. 2006; Mandal et al. 2008), Acilius sulcatus (Coleoptera: Dytiscidae) (Chandra et al. 2008), Mesocyclops (Copepoda: Cyclopoida) (Marten 1990), planaria (Dugesia bengalensis) (Kar and Aditya 2003), diving beetles (Ohba and Takagi 2010), and guppies (Poecilia reticulata) (Seng et al. 2008).
Some of the investigations were useful and practical as they were conducted in both the laboratory and the field. One such work was done by Marti et al. (2006) using three species of dragonfly nymphs, Neurothemis fluctuans, Orthetrum sabina, and Orthetrum chrysis, as predators against the three species of mosquitoes Ae. albopictus, Ae. aegypti, and Cx. quinquefasciatus. The results revealed preferential prey choices by the predators where Orthetrum sabina and Neurothemis fluctuans consumed larger quantities of Ae. aegypti larvae in contrast to other mosquito prey species, while Orthetrum chrysis preferred Cx. quinquefasciatus. They concluded that the predation rates from highest to lowest performance were Orthetrum sabina > Orthetrum chrysis > Neurothemis fluctuans. These trends were in support for the current work as reported here. Mandal et al. (2008) investigated the prey-predation abilities of a number of Odonata species nymphs against the mosquito prey, Cx. quinquefasciatus, where they found that the daily feeding rates of these predator species had varied capacities. The outcomes in descending values of larvae quantities per day were I. forcipata (64), A. flavifrons (57), R. ignipennis (45), S. durum (25), and C. kashmirum (14).
The current work reported on specific prey preferences shown by odonate predators where dragonfly nymphs of Orthetrum sabina and Neurothemis fluctuans captured more of the Ae. aegypti larvae in contrast to the other two mosquito species, whereas Orthetrum chrysis consumed more of Cx. quinquefasciatus larvae. Two contributing factors that influenced the selectivity of preys by the predators would be the prey’s escape strategies and the predatory abilities of predators. Observations conducted in this work, showed that Orthetrum sabina individuals were very active and aggressive when compared to the other two species, in which they consumed the most of the Aedes in contrast to the Culex mosquito larvae. The contributing factor to this high capture rates on Aedes compared to Culex could be deduced from work done by Yee et al. (2004) where they found different strategies in the mosquito larvae feeding behavior; the former prey species spent more of their activity time trashing below the water surfaces, and Culex spent more time at the surfaces. This evidential stratification in foraging areas made Aedes to be the targeted prey for dragonfly predators since dragonflies spent most of the time stalking for preys at bottom levels making Culex tendency to escape predation.
Additionally, the prey posture could be the contributing factor to the high success rate of capturing Aedes. Kar and Aditya (2003) worked on planaria as predators for mosquitoes where they found that planarians preferred Anopheles larvae rather than Culex species. This was explained to be due to the behavior of Anopheles larvae, in which they frequently were at rest and had resting postures in parallel to the water surfaces; this position made them easy targets for attack by the planarians. The Culex larvae were moving actively and thus impossible for captures by predators. Their work further reported exposure to different developmental stages of prey, where planarians selected larvae more than eggs or pupa regardless of mosquito species, whether Anopheles and Culex. Interestingly, planarians avoided the active 1st instars of mosquito species and foraged more on the less active 2nd and 3rd instars. Selection of prey based on profitability was apparent in studies done by Aditya et al. (2007), who found that the Toxorhynchites splendens consumed more of the prey Armigeres subalbatus compared to Culex quinquefasciatus larvae which were bigger in biomass. Thus, more predation efforts were directed toward Armigeres subalbatus as this would be the cost-effective foraging strategy. The rate of predation, however, dropped when prey turned to pupa. Similarly, work conducted by Ghosh et al. (2005) found that the predators used in their experiments ate greater quantities of mosquito prey larvae compared to pupa.
Predatory foraging decisions were also affected by dilution factors as displayed by Orthetrum sabina and Orthetrum chrysis where their attack behaviors decreased when water volume was increased. The tendencies for preys able to escape were enhanced with increased water volume and predators were less successful in their attacks in which led to waning of feeding rates. Such findings had been earlier reported by Mandal et al. (2008) for their experiments on dragonfly larvae predated on Cx. quinquefasciatus. Although increasing the water volume seemed to be of positive impact for mosquito preys in being provided with increased escape areas or routes, such factor could also be translated as increased foraging area for the predators. Such perspective was adopted by Shaalan et al. (2007), who worked on Hemiptera (Diplonychus sp. and Anisops sp.) as predators on mosquito larvae and found that the adults of Diplonychus sp. regardless of increased water volume foraged efficiently with quick attack movements taking advantage of the increased foraging areas.
5 Conclusion
Biocontrol has gained serious attention evident by the extensive investigations that had been supported by various biocontrol agents expressed by several authors, for example, the use of fishes were recommended by Medlock and Snow (2008) being advantageous as able to be maintained in either natural or ornamental water bodies; Nyamah et al. (2011) promoted the use of Toxorhynchites spp. claimed to be efficient predators to the mosquito larvae as well as being environmental friendly; Chatterjee et al. (2007) vouched the use of dragonfly larvae, Brachytron pratense, that was proven in his studies to be more efficient than the well-known larvivorous fishes like the Gambusia affinis in suppressing the mosquito larvae populations. One ideal method proposed by Kumar and Hwang (2006), in using dragonflies as biocontrol agents, would be to select bodies of water known to be breeding grounds for mosquitoes and create miniature biotopes adjacent to these water bodies in which native dragonfly nymphs could be artificially introduced and once hatched would predate on the airborne adult mosquitoes, while the female dragonflies would lay eggs in the mosquito habitats to hatch out into voracious larvae exploiting the mosquito larvae.
Although it is difficult to be definitive in the possible prospect of natural predators as effective in mosquito control, but what is obviously essential is care must be taken not to reduce their numbers by environmental manipulation or intensive use of agricultural pesticides. Direct strategies to enhance their numbers by making habitats more suitable for their continued survival and existence in the ecosystems should be the essential actions taken (Medlock and Snow 2008). The current concerns are also pertaining to what had been proposed by Kumar and Hwang (2006); in order to achieve an acceptable range of control, a sound knowledge of various attributes of interactions between a pest population and the predator to be introduced is desirable.
6 Future Focus
Further work is necessary, to determine the proper methodology for the mass rearing and augmentative release of the biocontrol agents that would make this biocontrol procedure feasible for widespread application. It is also important for understanding under what set of environmental conditions a predator will be effective in reducing mosquito populations.
References
Aditya G, Ash A, Saha GK (2006) Predatory activity of Rhantus sikkimensis and larvae of Toxorhynchites splendens on mosquito larvae in Darjeeling, India. J Vector Borne Dis 43:66–72
Aditya G, Bhattacharyya S, Kundu N, Kar PK, Saha GK (2007) Predatory efficiency of the sewage drain inhabiting larvae of toxorhynchites Splendens Wiedemann on culex quinquefasciatus say and Armigeres subalbatus (Coquillett) larvae. Southeast Asian J Trop Med Public Health 38(5):799–807
Bellows TS (2001) Restoring population balance through natural enemy introductions. Biol Control 21:199–205
Breene RG, Sweet MH, Olson JK (1990) Analysis of the gut contents of naiads of Enallagma civile (Odonata: Coenagrionidae) from a Texas pond. J Am Mosq Control 6:547–548
Caltagirone LE (1981) Landmark examples in classical biological control. Annu Rev Entomol 26:213–232
Chatterjee S, Ghosh NM, Chandra G (2007) Eco-friendly control of mosquito larvae by Brachytron pratense nymph. J Environ Health 69(8):44–48
Chandra G, Chatterjee SN, Ghosh A (2006) Role of Dragonfly (Brachytron pratense) Nymphs as a Biocontrol Agent or Larval Mosquitoes. Bull Panel Kesehat 34(4):147–151
Chandra G, Bhattacharjee I, Chatterjee SN, Ghosh A (2008) Mosquito control by larvivorous fish. Indian J Med Res 127:13–27
Corbet PS (1999) Dragonflies: behavior and ecology of odonata. Cornell University Press, Ithaca, 829 pp
Dykstra L (2008) Guidance for surveillance, prevention and control of mosquito borne disease. Washington State Department of Health, Washington, 137pp
Ghosh A, Mandal S, Bhattacharjee I, Chandra G (2005) Biological control of vector mosquitoes by some common exotic fish predators. Turk J Biol 29:167–171
Gubler DJ (1998) Dengue and dengue hemorrhagic fever. Clin Microbiol Rev 11(3):480–496
Headrick DH, Goeden RD (2001) Biological control as a tool for ecosystem management. Biol Control 21:249–257
Jatanasen S (1997) Environment manipulation and health education in Aedes aegypti control in Thailand. Bull WHO 36:363–368
Juliano SA, Gravel ME (2002) Predation and the evolution of prey behavior: an experiment with tree hole mosquitoes. Behav Ecol 13(3):301–311
Kar S, Aditya AK (2003) Biological control of mosquitoes by aquatic planaria. TISCIA 34:15–18
Kumar R, Hwang JS (2006) Larvicidal efficiency of aquatic predators: a perspective for mosquito biocontrol. Zoolog Stud 45(4):447–466
Mandal SK, Ghosh A, Bhattacharjee I, Chandra G (2008) Biocontrol efficiency of odonate nymphs against larvae of the mosquito, Culex quinquefasciatus Say, 1823. Acta Trop 106(2):109–114
Marten GG (1990) Elimination of Aedes albopictus from Tire Piles by introducing Macrocyclops albidus (Copepoda, Cyclopidae). J Am Mosq Control Assoc 6:689–693
Marti GA, Azpelicueta ML, Tranchida MC, Pelizza SA, Garcia JJ (2006) Predation efficiency of indigenous larvivorous fish species on Culex pipens L. larvae (Diptera: Cuculidae) in drainage ditch in Argentica. J Vector Ecol 31(1):102–106
Medlock JM, Snow KR (2008) Natural predators and parasites of British mosquitoes – a review. Eu Mos Bull 25:1–11
Milam CD, Farris JL, Wilhide JD (2000) Evaluating mosquito control pesticides for effect on target and nontarget organisms. Arch Environ Contam Toxicol 39:324–328
Nyamah MA, Sulaiman S, Omar B (2011) Field observation on the efficacy of Toxorhynchites splendens (Wiedemann) as a biocontrol agent against Aedes albopictus (Skuse) larvae in a cemetery. Trop Biomed 28(2):312–319
Ohba SY, Takagi M (2010) Predatory ability of adult diving Beetles on the Japanese encephalitis vector Culex tritaeniorhynchus. J Am Mosq Control Assoc 26(1):32–36
Rishikesh N, Dubitiskij AM, Moreau CM (1988) Malaria vector control. Biol Control 118:1227–1250
Saleeza SNR, Norma-Rashid Y, Sofian-Azirun M (2011) Mosquitoes larval breeding habitat in urban and suburban areas, Peninsular Malaysia. World Acad Sci Eng Technol 58:569–573
Sebastian A, Sein MM, Thu MM, Corbet PS (1990) Suppression of Aedes aegypti (Diptera: Culicidae) using augmentative release of dragonfly larvae (Odonata: Libellulidae) with community participation in Yangon, Myanmar. Bull Entomol Res 80:223–232
Seng CM, Setha T, Nealon J, Socheat D, Chantha N, Nathan MB (2008) Community-based use of the larvivorous fish Poecilia reticulata to control the dengue vector Aedes aegypti in domestic water storage containers in rural Cambodia. J Vector Ecol 33(1):139–144
Shaalan EAS, Canyon DV, Muller R, Younes MWF, Abdel-Wahab H, Mansour AH (2007) A mosquito predator survey in Townsville, Australia, and an assessment of Diplonychus sp. and Anisops sp. predatorial capacity against Culex annulirostris mosquito immatures. J Vector Ecol 32(1):16–21
Singh RK, Dhima RC, Dua VK, Joshi BC (2010) Entomological investigations during an outbreak of dengue fever in Lai Kuan town, Nainital district of Uttarakhand, India. J Vector Borne Dis 47:189–192
Spielman A, Kitron U, Pollack RJ (1993) Time limitation and the role of research in the worldwide attempt to eradicate malaria. J Med Entomol 30:6–19
Triplehorn CA, Johnson NF (2005) Introduction to the study of insects. Borror and DeLong’s. Belmont, Thomson Brooks/Cole
Yates MG (1979) The biology of the tree-hole breeding mosquito Aedes geniculatus in southern England. Bull Entomol Res 69:611–628
Yee DA, Kasevaraju B, Juliano SA (2004) Larval feeding behavior of three co-occurring species of container mosquitoes. J Vector Ecol 29(2):315–322
Acknowledgments
We would like to thank the Institute of Medical Research for the mosquito larvae used in the experimentation, the Putrajaya Health Office and Kuala Selangor Health Office for sampling area permissions, and Mr. Mohaiyyidin who assisted in the fieldwork. This project was funded by UMRG grant RG209-13SUS and IPPP grants PS209/2009C and PV065/2011B.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer India
About this chapter
Cite this chapter
Norma-Rashid, Y., Saleeza, S.N.R. (2014). Eco-friendly Control of Three Common Mosquito Larvae Species by Odonata Nymphs. In: Sahayaraj, K. (eds) Basic and Applied Aspects of Biopesticides. Springer, New Delhi. https://doi.org/10.1007/978-81-322-1877-7_13
Download citation
DOI: https://doi.org/10.1007/978-81-322-1877-7_13
Published:
Publisher Name: Springer, New Delhi
Print ISBN: 978-81-322-1876-0
Online ISBN: 978-81-322-1877-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)