
Abstract
Biological clocks are considered to increase an organism’s fitness in that they allow the synchronization of reproduction, physiology, and behavior with environmental conditions. Indirect evidence for the biological fitness value comes from the existence of local timing adaptations within a species. Some of the most clear cut examples of adaptations in circalunar and circadian clocks are found in the intertidal midge Clunio marinus. In adaptation to the local tidal regime, populations of this insect differ in the phase of the circadian clock, as well as in the phase, period, and zeitgeber sensitivity of the circalunar clock. These differences allow comparative genetic and molecular studies that may both shed light on the evolutionary forces shaping biological clocks and unravel the first known molecular components of a circalunar clock.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction: Selected Evolutionary Aspects of Circatidal and Circalunar Clocks
1.1 The Adaptive Value of Circatidal and Circalunar Clocks
Biological clocks are at the core of some of the most fascinating phenomena in biology, ranging from the time-compensated sun-compass orientation of the migratory monarch butterfly to the swarming of the palolo worm during a few predictable nights a year. With astonishing precision, circatidal, circadian, circa(semi)lunar, and circannual clocks capture the environmental cycles that are driven by the earth’s rotation around its axis, the earth’s revolution around the sun, and the revolution of the moon around the earth. It is reasonable to assume that biological clocks have evolved because they enhance an organism’s fitness, that is, its longevity and reproductive success, by allowing the organism to coordinate its physiology or behavior with environmental cycles. However, experimental validation of this hypothesis has turned out to be challenging. Even for the adaptive value (i.e., the ‘fitness advantage’) of circadian clocks, convincing evidence has been presented in only a few tractable systems (O’Donnell et al. 2011; Saunders 1972; Ouyang et al. 1998; Dodd et al. 2005) (for a review of other evidence, see Yerushalmi and Green 2009).
Circatidal and circalunar clocks are primarily found in marine organisms, and two major hypotheses have been discussed with respect to their adaptive value (Christy 1978; Pearse 1990; Naylor 2001).
On the one hand, circatidal and circalunar clocks may time an organism’s activity or reproduction to the most suitable times of the tidal cycle. This capacity is most important in intertidal zones, which experience strong fluctuations in physical conditions. Circalunar clocks come into play here because the tidal cycle is modulated across the lunar cycle (see Chap. 1 by Neumann, this volume, Sect. 1.2.1). This hypothesis may hold true for many crustaceans (e.g., Eurydice pulchra: Reid and Naylor 1985; Zhang et al. 2013; Excirolana chiltoni: Klapow 1972; Enright 1972; Sesarma haematocheir: Saigusa 1986; Saigusa 1980), and the marine midge Clunio marinus (see Chap. 1 by Neumann, this volume, and this chapter).
On the other hand, circalunar clocks in particular may synchronize reproduction within a population, which may be especially important for organisms with external fertilization, such as corals (Harrison et al. 1984) and marine annelids (Eunice viridis: Hauenschild et al. 1968; Platynereis dumerilii: Hauenschild 1960; Zantke et al. 2013; Typosyllis prolifera: Franke 1985). Lunar cues may also serve to synchronize reproduction in tropical regions where distinct seasonal cues are not available. A striking example may be the lunar-synchronized mass emergence of midges from tropical lakes (Corbet 1958; Hartland-Rowe 1955). There are many other ways in which lunar cycles can influence an organism’s fitness (Kronfeld-Schor et al. 2013), but in most cases evidence for the involvement of an endogenous circalunar clock is lacking.
There has been very limited experimental testing of the adaptive value of circalunar and circatidal clocks. For the marine midge Pontomyia oceana, which times its reproduction to distinct tidal situations by combining circalunar and circadian clocks (similar to C. marinus; see Chap. 1 by Neumann, this volume, and the following sections of this chapter), it was tested whether there is a fitness difference between the midges emerging in the full moon peak versus the new moon peak of a semilunar rhythm. The authors found no difference and concluded that emergence during both new moon and full moon spring tides is a bet-hedging strategy, which allows the organism to cope better with stochastic variation in the tides (Soong and Leu 2005). The same insect served to test the two hypotheses on the adaptive value of tide-related clocks in terms of the synchronization of reproduction versus timing to the tides. As already mentioned, P. oceana does not have a circatidal clock but instead relies on a circadian clock in combination with a circasemilunar clock to time emergence to the tides. The precision of circadian reproductive timing within the population was found to be higher than the accuracy of coordination with the tides (which also depends on the circalunar clock). This observation suggests that in P. oceana the adaptive value is rather synchronization of reproduction within the population (Soong et al. 2006).
1.2 Local Adaptation
Indirect evidence for the adaptive value of circatidal and circalunar clocks comes from the existence of local adaptations in these clocks, that is, from clock-controlled differences in tidal and lunar rhythms within one species that correlate with differences in the environmental cycles which the species experiences across its geographic range. Generally, adaptations are defined in terms of fitness, that is, as characteristics that enhance an organism’s survival or reproduction, relative to alternative character states. They are caused by natural selection and therefore must have a genetic basis (Futuyma 2009). Phenotypic plasticity, an environmentally induced change of a character during an organism’s lifetime, also enhances an organism’s performance in a specific environment (cf. Naylor 1989 with respect to tidal and lunar clocks). These changes are not embraced by the term adaptation in its evolutionary sense.
1.3 Temporal Isolation
Local adaptations in timing are particularly interesting if they affect reproduction. In these cases they may entail temporal isolation between populations, representing a prezygotic reproductive barrier (Dobzhansky 1937; Mayr 1942, 1970). Most studies on temporal isolation are concerned with seasonal differences in reproductive timing, be it in terrestrial (Alexander and Bigelow 1960; Tauber and Tauber 1977; Smith 1988; Filchak et al. 2000; Abbot and Withgott 2004; Levin 1978), riverine (Palstra et al. 2004), or marine habitats (Knowlton 1993; Palumbi 1994). There are some studies on diel differences in reproductive timing (Lloyd 1966), including in Drosophila, where the isolating effect of different circadian timing of activity and reproduction has been shown in the laboratory and could in one case be attributed to a single gene (Tauber et al. 2003). Studies on temporal isolation in tidal or lunar rhythms are rare: differing lunar and diel rhythms of reproduction are reported in closely related damselfish species (Foster 1987), and differing lunar rhythms of spawning were observed in sea urchins (Lessios 1984; Muthiga 2003). Organisms from the intertidal zone have hardly been studied with respect to local timing adaptations and temporal isolation, which is surprising because the fluctuation of environmental conditions is strong in the intertidal zone and tidal regimes differ tremendously between different geographic locations (see Sect. 1.2.1 by Neumann, this volume). Hence, these organisms can be expected to have pronounced and site-specific tidal and lunar rhythms.
1.4 Examples of Local Adaptation in Circatidal and Circalunar Cocks
Local adaptation in a circatidal clock is reported for the sand beach isopod Euridyce pulchra. This crustacean displays a tidal rhythm of swimming activity that is modulated across the lunar cycle (Reid and Naylor 1986). Entrainment of the tidal rhythm by cycles of mechanical disturbance was found to be most effective when the 2-h mechanical stimulus was presented during the daytime of high tide on spring tide days. For isopods from two different geographic locations, the time of day when most effective mechanical entrainment occurred, was found to differ according to the tidal regimes at their place of origin (Fig. 7.1).

Mean total activity (±SD) over five tidal cycles of the isopod Eurydice pulchra after exposure to a regime of 2-h agitation every 12 h for 4 days. The x-axis gives the time of day of the entraining 2-h agitation pulse. Each point on the x-axis is an independent experiment with 4–10 groups of 20 specimens. The last two points are repetitions of the first two for ease of comparison. The y-axis gives mean total activity, which is an approximation for the quality of entrainment of the tidal rhythm by the 2-h agitation pulse, plotted for populations from Swansea (black) and Newborough (grey). Spring tide high tides occur around dusk and dawn in Swansea and around noon and midnight in Newborough. For both locations, entrainment is most effective around the time of local spring high tides. (Adapted from Reid and Naylor 1986)
Local adaptations in a circalunar clock are found in the marine midge Clunio marinus, for which the components of the circalunar and circadian timing system have been progressively characterized throughout the 1960–1990s (see Chap. 1 by Neumann, this volume). In the past years this species has become accessible to genetic and molecular studies. The remainder of this chapter focuses on the timing adaptations of this re-emerging model system, which may inspire both chronobiologists and evolutionary biologists.
2 Local Timing Adaptations in Clunio marinus
2.1 Introduction to C. marinus
An overview on the life cycle of C. marinus, its endogenous clocks, and their experimental corroboration is given in Chap. 1 by Neumann (for more details see Neumann 1966, 1986, 1995). Briefly, C. marinus (Diptera, Chironomidae) is a marine midge that is found in the rocky intertidal zone along the European Atlantic Coast (Fig. 7.2). The larvae build their tubes in pads of small algae and sand, which are found in the lower midlittoral on protected rocks that are almost permanently submerged. However, the adults need these larval substrates to be dry for oviposition, as they literally “glue” the egg masses onto suitable sites. To overcome the problem, Clunio has an extremely reduced adult lifespan of 1–2 h, and adult emergence is timed to coincide with the spring tide low tides, that is, the low tides on days around new moon and full moon, when the water levels are generally lowest. Upon emergence, the midges mate and oviposit immediately, then die in the rising tide. A circalunar clock controls development to ensure that pharate pupae, that is, pupae ready to emerge, are only present during spring tide days. A circadian clock controls the daily timing of adult emergence. Notably, in C. marinus there is no evidence for a circatidal clock. The midge achieves synchronization of its life cycle with the tides only by the combination of the circadian clock with the circalunar clock. Both clocks have been shown to be free running and temperature compensated, and the entraining cues (‘zeitgebers’) are known: the circadian clock is entrained by the light–dark (LD) cycle, whereas the circalunar clock responds to moonlight, tidal turbulence, or tidal temperature fluctuations. Along the European coastline, Clunio populations differ in at least four aspects of these timing systems: (1) phase of the diel emergence rhythm with respect to the zeitgeber cycle, (2) phase of the lunar emergence rhythm with respect to the zeitgeber cycle, (3) zeitgeber sensitivity of the circalunar clock, and (4) period of the circalunar clock. These aspects are discussed in detail below.

Geographic range and locally adapted diel emergence times of selected laboratory strains of Clunio marinus. (a) Diel emergence peaks of males (open circles) and females (filled circles) are plotted against time in the artificial LD cycle in the laboratory. Diamonds with whiskers are mean emergence times ± SD. Arrows with whiskers are time of low tide on spring tide days ± SD at the place of origin of the respective laboratory strains. (b) The places of origin of the five laboratory strains as indicated by black circles correspond to the five regions analyzed in the population genetic study reviewed in Sect. 7.4. Grey shaded area is the known distribution of Clunio marinus. Black lines along the coast indicate predominantly rocky coast; grey lines indicate sandy coast, which is generally not inhabited by Clunio marinus. Numbers indicate sampled subpopulations: 1 Punta Helena; 2 St. Jean-de-Luz; 3 Erromardie; 4 Guéthary; 5 Grandcamp; 6 Port-en-Bessin; 7 Westwatt, 8 Vorhafen; 9 Düne; 10 Kviturdvikpollen. Other samples (11 Greystones/Ireland; 12 Reykjavík/Iceland) are only used in the haplotype network (Fig. 7.6). (Adapted from Kaiser et al. 2010)
2.2 Phase of the Diel Emergence Rhythm
Differences in diel emergence time were the first variable clock properties to be observed in C. marinus (Neumann 1966). As delineated in Sect. 1.2.1 by Neumann (this volume), the daytime of low tide on spring tide days is close to constant for a given geographic location, but it varies tremendously between geographic locations. Laboratory strains of Clunio from different geographic locations emerge at different times relative to the artificial LD cycle, and these differences correlate well with the time of low tide on spring tide days at their place of origin (Fig. 7.2). There is no evidence for involvement of a circatidal clock, given there is no daily delay of emergence according to the tides, and in each strain adults emerge only during one of the two daily low tides. The adaptive value of these diel timing differences has not been tested experimentally. However, it is intuitively clear that, given the short lifespan of Clunio adults, individuals emerging only a few hours early or late will find neither mating partners nor suitable substrates for oviposition, which must severely impair their fitness. The genetic basis of these adaptations in the circadian clock is discussed in Sect. 7.3.
2.3 Phase of the Lunar Emergence Rhythm
It was noticed early that Clunio laboratory strains of different geographic origin emerge at different times of the artificial moonlight cycle in the laboratory (Neumann 1966). This observation remained enigmatic for a long time, as Clunio populations in the field emerge around the spring tide days, that is, at the same time of the lunar cycle, irrespective of geographic location. Only recently were the different lunar emergence times shown to represent local adaptations to the tidal regime (Kaiser et al. 2011).
The conceptual background to understanding these lunar timing adaptations was established in two experiments, which are described in detail in Sect. 1.5.4 by Neumann (this volume; see Figs. 1.6 and 1.7). Briefly, one experiment showed that because of the circadian gating of Clunio moonlight sensitivity, moonlight is only an effective zeitgeber around midnight. The other experiment brought forth the idea that the tides may modulate the intensity of moonlight that reaches the submerged larval substrates of Clunio. In the light of these findings, it seems reasonable to assume that the time point of moonlight perception in the field depends on three conditions (Fig. 7.3). (1) Clunio must be sensitive to moonlight, which is only the case around midnight. (2) The moon must be in the sky around midnight, which is true during the moonlit quarters of the lunar cycle from waxing half moon to waning half moon. (3) The larval habitat in the lower midlittoral must be effectively exposed to moonlight by a low tide around midnight. The latter condition dramatically narrows the window for moonlight perception to a few days and is fulfilled during specific days in the lunar cycle for each geographic location, depending on the local tidal regime (compare site 1 and site 2 in Fig. 7.3a). Thus, the days of moonlight perception in the field are site specific, whereas the days of Clunio emergence during the spring tides are the same for all populations; this may explain the strain-specific phase relationship between the artificial moonlight stimulus and the emergence peak in the laboratory.

Local adaptation in lunar emergence times of Clunio marinus. (a) Schematic relationship of the moon phases, tides, Clunio’s moonlight sensitivity, and the locally adapted diel and lunar emergence times for two localities with differing tidal regimes (site 1, site 2). Time of day is plotted against days of the lunar cycle. Black area represents dark phase; grey shading indicates when the moon is in the sky. Red box marks the circadian period of sensitivity of Clunio to moonlight. Fig. 7.3 (continued) Hypothetically, moonlight is best perceived when the time of low tide (blue dotted lines) falls to midnight (yellow box). As tidal regimes differ for other places along the coast (compare blue dotted lines in site 1 and site 2), the time when moonlight is best perceived differs. Nevertheless, all known Clunio populations emerge during the spring tides (around new moon and/or full moon). Thus, they must respond to the moonlight stimulus with a different delay of their emergence peak (yellow bars below the graph). Note that the required strain-specific phase of circalunar emergence time (yellow bars) is correlated with the required strain-specific phase of circadian emergence time relative to LD (green bars) because both are adaptations to the tidal regime. (b) Lunar emergence times of Clunio laboratory strains relative to the artificial moonlight stimulus, plotted against the time between the days with low tide at midnight and spring tide days in the field (according to the model in a). Error bars are standard deviations. Squares mark emergence peaks that fall to full moon, circles mark emergence peaks that fall to new moon in the field. Correlation coefficient and p value of the correlation are given in the graph (Pearson’s product–moment correlation). Strain identities: 1 Vigo; 2 Santander; 3 St. Jean-de-Luz; 4 Port-en-Bessin; 5 Lulworth; 6 Studland; 7 Bembridge; 8 Roscoff; 9 Concarneau; 10 Camaret-sur-Mer; 11 St. Briac-sur-Mer. (Adapted from Kaiser et al. 2011)
The testable prediction resulting from this hypothesis is that, for each Clunio population, the number of days between the artificial moonlight stimulus and the adult emergence peak in the laboratory should correspond to the number of days between the days with low tides around midnight and the spring tide days in the field, as has been tested for 11 laboratory strains; the results depict a perfect correlation (Fig. 7.3b). This finding has two general implications for the study of moonlight perception in circalunar clocks. First, it argues for caution when interpreting the artificial moonlight stimulus given in laboratory experiments; it may not necessarily represent full moon. Second, the modulation of light intensity by the tides makes moonlight basically a ‘tidal stimulus.’ Its perception depends on a circadian modulation of receptor sensitivity, and the very same mechanism has been discussed for the perception of other tidal stimuli, such as tidal turbulence cycles or tidal temperature cycles (see Sect. 1.5.5 by Neumann, this volume).
The genetic basis of the strain-specific phase relationship between moonlight zeitgeber and lunar emergence time is described in Sect. 7.3.
2.4 Zeitgeber Sensitivity of the Circalunar Clock
Three different stimuli have been shown to act as zeitgebers for entrainment of the circalunar clock in C. marinus: moonlight, tidal cycles of water turbulence, and tidal cycles of water temperature (for details of the mechanisms, see Sect. 1.5.5). The geographic distribution of the sensitivity to these zeitgebers suggests a certain degree of local adaptation (Table 7.1). Whereas all laboratory strains from southern regions respond to moonlight, the Helgoland strain from the North Sea does not; similar to other strains from northern regions, it responds strongly to tidal cycles of water turbulence or temperature. The suggested adaptive value of the lack of moonlight sensitivity in the Helgoland strain is that moonlight may not be a reliable time cue at northern latitudes, where the moon is low above the horizon and nights are short or even bright in summer may thus not penetrate the water surface effectively. The Bergen strain, which comes from a sheltered fjord location, responds much more strongly to temperature cycles than to turbulence cycles, which may correlate to the general lack of surf or waves that characterizes its habitat.
Notably, most populations responded to all tested zeitgebers (compare Table 7.1), suggesting that Clunio generally relies on the combination of several time cues to assure entrainment of the circalunar clock. In this sense, local adaptation may not involve the acquisition of habitat-specific zeitgeber sensitivities, but rather the loss of sensitivity to zeitgebers that are unreliable or absent or not required in a particular habitat.
The genetic basis of zeitgeber sensitivity has so far not been investigated.
2.5 Period of the Circalunar Clock and Emergence During New Moon Versus Full Moon
With respect to the period of the circalunar clock, two classes have been described in C. marinus. There are strains with a semilunar rhythm that emerge during both full moon and new moon spring tides in the field. Two of these strains were tested in the laboratory and showed a free-running period of the circasemilunar clock of 11 days (He strain from Helgoland, Germany) or 13–14 days (Por strain, from Port-en-Bessin, Normandie/France), corresponding closely to half a lunar cycle of 14.77 days (Neumann 1966). There are also strains that have a lunar rhythm. Most of these strains emerge during new moon spring tides only (e.g., Jean strain, from St. Jean-de-Luz, Basque Coast/France), but one of them was found to emerge during full moon spring tides only (Ros strain, from Roscoff, Bretagne/France; see Neumann 1989). The free-running period of the circalunar clock in the Jean strain was measured to be 26–27 days (Neumann 1966).
It is unclear whether these differences represent local adaptations. The level of new moon versus full moon low tides can differ by 1–2 m along the European Atlantic Coast. However, magnitude and direction of the effect change gradually and are not consistent throughout the years. In some years full moon low tides are lower during Clunio emergence season in summer, and in other years new moon low tides are lower. In this respect, emergence in a semilunar rhythm may represent a bet-hedging strategy, as is reported for Pontomyia oceana (Soong and Leu 2005). Most strains with a lunar rhythm emerge during dark phase, and an explanation for emergence during new moon versus full moon in strains with a lunar rhythm might be found in factors associated with the increased nightly illumination during full moon nights, for example, an increased risk of predation.
There are hints to a genetic basis for the differences in the period of the lunar rhythm (see Sect. 7.3).
2.6 Adaptive Loss of the Lunar Rhythm
In two geographic areas, C. marinus is reported to have lost the lunar rhythm. The arctic population of Tromsø in northern Norway has completely abandoned both the circalunar and the circadian rhythm of adult emergence and instead relies on a tidal hourglass mechanism (see Sect. 1.4.4 by Neumann, this volume). These changes can be considered an adaptation to arctic summer with more than 2 months of permanent daylight, which precludes entrainment of the circadian clock and thereby also zeitgeber perception for the circalunar clock.
Clunio midges from the Baltic Sea were first recorded as C. marinus, but were later described as an independent species, C. balticus (Heimbach 1978). It is likely that C. balticus was derived from C. marinus within the past 10,000 years, as subfossil Clunio head capsules can be found in Baltic Sea sediments only within the past 8,000–10,000 years (Hofmann and Winn 2000). The two species can still be interbred in the laboratory (Heimbach 1978) and cannot be distinguished by standard phylogenetic markers (T. Kaiser, unpublished data). Supposedly, in adaptation to the virtual absence of tides in the Baltic Sea, C. balticus has lost the lunar rhythm of emergence. In contrast to C. marinus, the adults of C. balticus do not rely on exposure of the larval habitat but instead swarm and lay their egg masses on the open water surface, sometimes far offshore. Unlike the egg masses of C. marinus, egg masses of C. balticus penetrate the water surface and sink to the larval substrates (Endraß 1976), which are red algae at a depth of 2–20 m. Presumably to ensure sufficient aggregation of mating partners in the open sea, the diel emergence window of C. balticus is extremely narrow (30–60 min); it is timed to and modulated by dusk. While these adaptations make C. balticus independent from the tides, they do not impede the species from settling in places with tides, beyond the Baltic Sea. Indeed, sympatric populations of C. marinus and C. balticus are described from Bergen in Norway, where heterospecific matings are prevented by their differing circadian emergence times (Heimbach 1978). The stable co-occurrence of both species in this site as a consequence of temporal isolation was the basis for giving species status to C. balticus. This interesting evolutionary scenario may represent a case of ongoing ecological speciation and certainly deserves further investigation.
In summary, local adaptations in circalunar and circadian timing occur at a multitude of levels in C. marinus. Two of them—namely, phase of the lunar rhythm and phase of the diel rhythm relative to the respective zeitgeber cycles—have been studied at the genetic and molecular level. These experiments are described in the following sections.
3 The Genetic Basis of Circalunar and Circadian Emergence Times of Clunio marinus
3.1 Crossing Experiments
As early as 1967 Dietrich Neumann reported genetic control of the strain-specific diel emergence times in C. marinus (Neumann 1967). Recently, these results were confirmed, and beyond that, genetic control of the strain-specific lunar emergence times was discovered (Kaiser et al. 2011). Both studies relied on crossing experiments between two specific laboratory strains, which differ considerably in lunar and diel emergence time (Fig. 7.4a, b; panels 1 and 2). The two strains originate from the shores of St. Jean-de-Luz (Basque Coast, France) and Port-en-Bessin (Normandie, France) and are therefore referred to as Jean and Por strains (see Fig. 7.2). F1 hybrids of these two strains emerge at intermediate times between the parents with respect to both the lunar and the diel emergence time; the backcross progeny are intermediate between the backcross parents (Fig. 7.4a, b; panels 3 and 4), which documents a genetic basis to both strain-specific diel and lunar emergence times. The lack of 1:1 segregation in the backcross progeny indicates that each trait is controlled by two or more independent genetic factors (Kaiser et al. 2011).

Emergence patterns of parental strains, hybrids, and backcrosses of two populations of Clunio marinus under light-dark cycles (LD 14:10) and artificial moonlight entrainment. (a) Diel rhythm plotted as the fraction of individuals that emerged during 30-min intervals, for the parental strains of Port-en-Bessin (Por) and St. Jean-de-Luz (Jean), the F 1 generation, and the backcross generation (BC). Daytime is given in hours after the middle of dark phase (“hour 0”), which necessarily makes the middle of the light phase “hour 12.” Hatched area marks dark phase. (b) Lunar rhythm plotted as the fraction of individuals that emerged during each day of the artificial moonlight cycle. Arrows mark days with artificial moonlight. Additive data of two lunar cycles are shown. (From Kaiser et al. 2011)
3.2 Linkage Mapping and QTL Analysis
One family of the backcross progeny from the aforementioned crosses served to construct a linkage map of the C. marinus genome (Kaiser and Heckel 2012). The map has three linkage groups (Fig. 7.5), corresponding to the three chromosomes known from light microscopy (Michailova 1980). It has a genetic length of 167–193 cM (centimorgans) and the genome has a physical size of 95–140 million base pairs (Mb), as estimated by flow cytometry. The map also contains the locations of most circadian clock genes known from insects, as well as a number of light receptor genes, which are considered candidates for being involved in the timing differences between the strains.

Linkage map of the Clunio marinus genome including the quantitative trait loci (QTLs) for diel and lunar emergence times. There are three linkage groups corresponding to the three chromosomes. Map length in centimorgans (cM) is given to the left, marker names to the right. Locations of a number of light receptor and clock genes are given. For period, vrille, casein kinase 1a, cryptochrome 1, lark, and cycle only approximate positions are known. The QTLs for the differences in diel emergence time are light grey; QTLs for differences in lunar emergence time are dark grey. They overlap with the loci of cOpsin2, timeout, and cryptochrome 1 respectively. (Adapted from Kaiser and Heckel 2012)
Quantitative trait locus (QTL) analysis, a statistical method testing the association of the patterns of inheritance of genetic markers with the pattern of inheritance of a given trait of interest, identified two QTLs controlling lunar emergence time, one on linkage group 1 and one on linkage group 2 (Fig. 7.5). For diel emergence time, two QTLs were identified, both on linkage group 1. The lunar QTL on linkage group 1 overlaps with one of the circadian QTLs. The estimated additive effects of these loci account for 85 % of the difference in diel emergence time and the full difference in lunar emergence time between the strains, indicating that these loci are the only major-effect loci for the two traits. The QTLs overlap with the location of the clock gene timeout/timeless 2 as well as the locations of the light receptors (insect type) cryptochrome 1 and ciliary opsin 2. These genes remain candidates for being involved in the timing differences. All other mapped candidate genes, including the well-studied circadian clock genes period, cycle/bmal, clock, timeless, and (mammalian type) cryptochrome 2, are not in the QTLs. For these genes, a major effect on strain-specific timing can be excluded.
3.3 Genetic Nonindependence of Diel and Lunar Emergence Times
In the backcross progeny of the crossing experiment, diel and lunar emergence times were correlated. Individuals emerging early in the artificial moonlight cycle also emerged early in the LD cycle, and individuals emerging late in the artificial moonlight cycle also emerged late in the LD cycle, suggesting a genetic nonindependence of both traits (Kaiser et al. 2011). This finding was underscored by QTL mapping, which showed that the QTLs with the largest effect on diel or lunar emergence time, respectively, are overlapping (Fig. 7.5). The resolution of the genetic map does not allow one to conclude whether the correlation is the result of independent but close-by circadian and circalunar genes (genetic linkage) or is caused by a single gene affecting both timing traits (pleiotropy). In the latter case we may further distinguish an effect of the circadian clock as a whole on the lunar rhythm (modular pleiotropy; e.g., the known involvement of the circadian clock in moonlight perception in C. marinus), from the effect of a single gene being involved in both circalunar and circadian clocks (gene pleiotropy). Deciding between these possible explanations will likely require the identification of the underlying genes.
Irrespective of the mechanism, the correlation of lunar and diel emergence times can itself be considered adaptive in the intertidal zone, as it stabilizes well-adapted combinations of diel and lunar emergence times. This correlation is beneficial, as each geographic location requires a specific combination of adaptations in lunar and diel emergence time to ensure emergence during the low tide of spring tide days. As both diel and lunar adaptations are adaptations to the tidal regime, they change in a correlated manner along the coastline (compare Fig. 7.3a). If lunar and diel timing are found to be controlled by independent but linked genes, it is conceivable that selection may have shaped genome architecture toward genetic linkage of those genes.
3.4 A Genetic Basis of the Period of a Lunar Rhythm?
The crossing experiment is in principle also informative on the nature of a lunar rhythm (Jean strain) versus a semilunar rhythm (Por strain). However, it is not clear whether the minor second peaks in the lunar rhythm of the F1 and the backcross progeny (see Fig. 7.4) represent a genetic influence of the Por strain or an artefact of artificial moonlight entrainment (which sometimes produces small second peaks in the Jean strain as well). All individuals in the minor peak in the mapping family of the backcross progeny inherited a region of linkage group 2, ranging from marker groups 2-M16 to 2-M22, from the Por grandparent. If genetic control of the minor peak were to be confirmed, the genetic factors controlling the trait are expected to be in this genomic region.
3.5 Future Prospects
The reference genome sequence of Clunio marinus is close to completion, and population samples of the Por and Jean strains have been resequenced to identify which genes within the QTLs show strain-specific genetic variation (Kaiser, von Haeseler, Tessmar-Raible, unpublished). This step will reduce the number of candidate genes to a manageable level for analysis. The required molecular tools are currently being established for C. marinus.
The availability of a reference genome will not only facilitate molecular studies of Clunio’s complex timing systems but also boost studies investigating the evolutionary causes and consequences of these timing adaptations. These topics are the subject of the following section.
4 The Evolutionary Background of Timing Adaptations in Clunio marinus
C. marinus has been be found at basically all rocky sites along the European mainland coast, on the British Isles, and in Iceland. At each geographic location, the populations are adapted to the local tidal regime in various aspects of the circadian and circalunar clocks (see Sect. 7.2 and Fig. 7.2). In the light of these findings, several questions must be raised with respect to the evolutionary history and population genetics of these populations: (1) When were the European coasts colonized? Namely, in which timeframe did the timing differences evolve? (2) Were the European coasts colonized from a single source or from several sources? Colonization from a single source would imply that the timing adaptations evolved after colonization. (3) Are these populations genetically isolated today? (4) If so, is genetic isolation the result of temporal isolation? And do the local timing adaptations drive genetic divergence between the populations? Answering these questions may also allow indirect conclusions to be drawn on the evolutionary mechanisms that shaped the timing adaptations. These issues were addressed in a recent study (Kaiser et al. 2010) that included a hierarchical sampling of ten populations from five regions along the European mainland coast (see Fig. 7.2b), at two very different scales of geographic separation. Regions were separated by 650–1,150 km. Within regions, populations were separated by 2–20 km only. From each population, 18 males were analyzed with nuclear and mitochondrial genetic markers: microsatellites, amplified fragment length polymorphisms (AFLPs), and mitochondrial DNA sequences (cytochrome oxidase I). The following conclusions could be drawn with respect to the afore-mentioned questions:
-
1.
Timeframe of colonization: The haplotype network of COI sequences showed one major haplotype for each region, with a star-like expansion of minor haplotypes derived from the major haplotypes (Fig. 7.6). This is a signal of recent colonization, likely after the last Ice Age, within the last 20,000 years. The nuclear AFLP markers support this view, in that more than 80 % of the markers lack polymorphism all across Europe.
Fig. 7.6 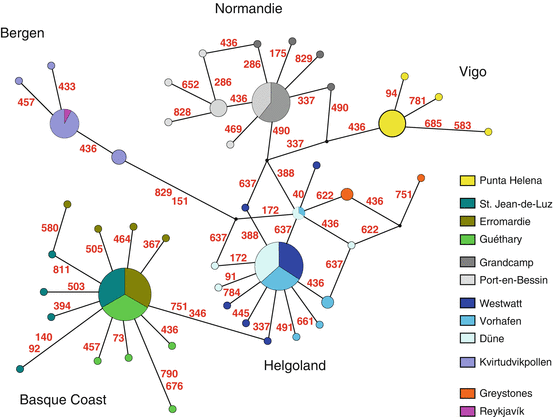
Haplotype network for COI sequences. Each circle represents a haplotype; size of the circle equals haplotype frequency in the sample. Lines and red numbers indicate the mutations separating the haplotypes and the position of the mutation along the sequence. Geographic origins of the haplotypes are color coded (see insert). Small black dots are hypothetical haplotypes. Each region has its own set of haplotypes; only Iceland shares its haplotype with Norway. (From Kaiser et al. 2010)
-
2.
Colonization sources: The colonization source does not seem to be contained within the sampled regions, as all of them seem to be of recent origin. With respect to the number of colonization sources, the data are contradictory. Mitochondrial markers suggest colonization from five different sources (Fig. 7.6), but polymorphism in nuclear markers—if present—is shared and suggests a common source of all populations. As the identity and number of the colonization sources is unresolved, it remains unclear which of the timing adaptations were already present before colonization.
-
3.
Genetic isolation: Within regions at a scale of 2–20 km, genetic differentiation is very low, indicating that these populations admix freely (Table 7.3). Between regions, on a scale of 650–1,150 km, mitochondrial haplotypes are perfectly private (Fig. 7.6). Genetic polymorphism in nuclear markers is shared, but allele frequencies differ tremendously, supporting strong genetic isolation (Table 7.2). Most likely these regions have not had much genetic exchange since they were first colonized. Long stretches of sandy coast (which are not habitable for Clunio) seem to represent particularly strong barriers to gene flow: Vigo and Basque Coast, which are the only regions situated within the same stretch of continuous rocky coast, are less differentiated than the other regions (Table 7.2).
Table 7.2 Genetic differentiation in nuclear markers as measured by pairwise F ST between regions Table 7.3 Genetic differentiation in nuclear markers as measured by pairwise F ST between populations within regions -
4.
Temporal isolation: A test for temporal isolation was done at the scale of regions. If temporal isolation matters, genetic differentiation between the regions should be correlated to the timing difference between the regions. At the time of the study, local adaptation in the lunar rhythm was not yet fully understood, so that only the overlap of the diel emergence peaks (compare Fig. 7.2a) served as an approximation of timing differences. The diel timing differences show no correlation to the genetic differentiation, indicating that at the scale of regions temporal isolation does not matter. Geographic isolation seems to be the major isolating force at this scale.

In the light of the foregoing, the evolutionary background of the timing adaptations seems to differ depending on the geographic scales. At the scale of regions, the most likely scenario is that discrete stretches of rocky coast were colonized in rare and separate events. Strong geographic isolation between the regions likely facilitated the subsequent evolution of timing adaptations.
At the scale of populations within the same stretch of continuous rocky coast, geographic isolation is not very effective. If these populations have local adaptations in their circadian and circalunar clocks, these must have evolved or are being maintained, despite gene flow, only as a result of strong local selection. This idea represents a very different evolutionary mechanism compared to the one found at the scale of regions and deserves further investigation.
5 Concluding Remarks
The local timing adaptations of C. marinus in its circalunar and circadian clocks offer the unique opportunity to access and investigate the yet unknown circalunar clock and its interaction with the circadian clock by means of experimental and genetic dissection. At the same time, C. marinus provides a natural setup to study a multitude of evolutionary processes associated with biological clocks, ranging from the selective forces that act on biological clocks, to the temporal isolation that these clocks may impose on a species. In a very fruitful way, functional and evolutionary studies can shed light on each other in C. marinus. Evolutionary genomic scans for patterns of selection or differentiation between the locally adapted Clunio strains may help to identify genes that are involved in determining the local timing adaptations, possibly including the first genetic components of circalunar clocks known in any species. Conversely, knowing which genes are involved in circadian and circalunar clocks and determining their molecular interactions may foster our understanding of the evolutionary history of Clunio populations and of the evolutionary forces that shape local timing adaptations and temporal isolation.
Beyond this, the genus Clunio comprises a few dozens of species worldwide, and there are many more timing adaptations to be studied that may allow further experimental analysis of circadian and circalunar clocks, and perhaps circatidal and circannual clocks as well. Notably, the species Clunio tsushimensis from the seas of Japan was reported to display a tidal modulation in its diel emergence time (Saigusa and Akiyama 1995; Oka and Hashimoto 1959). As yet, it has not been tested whether this is a direct response to the tides, or whether there is an underlying circatidal clock. Furthermore, C. tsushimensis emerges year round and the tidal regime in its habitat is characterized by a distinct inequality between the levels of the two daily low tides. In summer, the morning low tides are lower, whereas in winter the evening low tides are lower. Consequently, C. tsushimensis was found to emerge during the morning low tides in summer and during the evening low tides in winter (Saigusa and Akiyama 1995; Oka and Hashimoto 1959). A preliminary laboratory study suggested that the seasonal switch in diel emergence times is directly controlled by photoperiod (Neumann 1983).
Clearly, there are many more enigmatic clocks to be unraveled in the unique model system Clunio.
References
Abbot P, Withgott JH (2004) Phylogenetic and molecular evidence for allochronic speciation in gall-forming aphids (Pemphigus). Evolution 58(3):539–553
Alexander RD, Bigelow RS (1960) Allochronic speciation in field crickets, and a new species, Acheta veletis. Evolution 14(3):334–346
Christy JH (1978) Adaptive significance of reproductive cycles in the fiddler crab Uca pugilator: a hypothesis. Science 199(4327):453–455
Corbet PS (1958) Lunar periodicity of aquatic insects in Lake Victoria. Nature (Lond) 182(4631):330–331
Dobzhansky T (1937) Genetics and the origin of species. Columbia University Press, New York
Dodd AN, Salathia N, Hall A, Kevei E, Toth R, Nagy F, Hibberd JM, Millar AJ, Webb AAR (2005) Plant circadian clocks increase photosynthesis, growth, survival, and competitive advantage. Science 309(5734):630–633
Endraß U (1976) Physiological adaptations of a marine insect. 2. Characteristics of swimming and sinking egg-masses. Mar Biol 36(1):47–60
Enright JT (1972) Virtuoso isopod: circa-lunar rhythms and their tidal fine-structure. J Comp Physiol 77(2):141–162
Filchak KE, Roethele JB, Feder JL (2000) Natural selection and sympatric divergence in the apple maggot Rhagoletis pomonella. Nature (Lond) 407(6805):739–742
Foster SA (1987) Diel and lunar patterns of reproduction in the Caribbean and Pacific sergeant major damselfishes Abudefduf saxatilis and Abudefduf troschelii. Mar Biol 95(3):333–343
Franke H-D (1985) On a clocklike mechanism timing lunar-rhythmic reproduction in Typosyllis prolifera (Polychaeta). J Comp Physiol Sens Neural Behav Physiol 156(4):553–561
Futuyma DJ (2009) Evolution. Sinauer Associates, Sunderland
Harrison PL, Babcock RC, Bull GD, Oliver JK, Wallace CC, Willis BL (1984) Mass spawning in tropical reef corals. Science 223(4641):1186–1189
Hartland-Rowe R (1955) Lunar rhythm in the emergence of an ephemeropteran. Nature (Lond) 176:657
Hauenschild C (1960) Lunar periodicity. Cold Spring Harbor Symp Quant Biol 25:491–497
Hauenschild C, Fischer A, Hofmann DK (1968) Untersuchungen am pazifischen Palolowurm Eunice viridis (Polychaeta) in Samoa. Helgol Wiss Meeres 18(3):254–295
Heimbach F (1976) Semilunare und diurnale Schlüpfrhythmen südenglischer und norwegischer Clunio-Populationen (Diptera, Chironomidae). Ph.D. thesis, Universität Köln, Köln
Heimbach F (1978) Sympatric species, Clunio marinus Hal. and Cl. balticus n. sp. (Dipt., Chironomidae), isolated by differences in diel emergence time. Oecologia (Berl) 32(2):195–202
Hofmann W, Winn K (2000) The littorina transgression in the Western Baltic Sea as indicated by subfossil Chironomidae (Diptera) and Cladocera (Crustacea). Int Rev Hydrobiol 85(2-3):267–291
Kaiser TS, Heckel DG (2012) Genetic architecture of local adaptation in lunar and diurnal emergence times of the marine midge Clunio marinus (Chironomidae, Diptera). PLoS One 7(2):e32092
Kaiser TS, Neumann D, Heckel DG, Berendonk TU (2010) Strong genetic differentiation and postglacial origin of populations in the marine midge Clunio marinus (Chironomidae, Diptera). Mol Ecol 19(14):2845–2857
Kaiser TS, Neumann D, Heckel DG (2011) Timing the tides: genetic control of diurnal and lunar emergence times is correlated in the marine midge Clunio marinus. BMC Genet 12:49
Klapow LA (1972) Fortnightly molting and reproductive cycles in sand-beach isopod, Excirolana chiltoni. Biol Bull 143(3):568–591
Knowlton N (1993) Sibling species in the sea. Annu Rev Ecol Syst 24:189–216
Kronfeld-Schor N, Dominoni D, de la Iglesia H, Levy O, Herzog ED, Dayan T, Helfrich-Forster C (2013) Chronobiology by moonlight. Proc R Soc B Biol Sci 280(1765)
Lessios HA (1984) Possible prezygotic reproductive isolation in sea-urchins separated by the Isthmus of Panama. Evolution 38(5):1144–1148
Levin DA (1978) The origin of isolating mechanisms in flowering plants. In: Hecht MK, Steere WC, Wallace B (eds) Evolutionary biology, vol 11. Plenum Press, New York, pp 185–317
Lloyd JE (1966) Studies on the flash communication system in Photinus fireflies. Misc Publ Mus Zool Univ Mich 130:1–95
Mayr E (1942) Systematics and the origin of species from the viewpoint of a zoologist. Columbia biological series, vol 13. Columbia University Press, New York
Mayr E (1970) Populations, species, and evolution: an abridgement of animal species and evolution. Belknap Press, Cambridge
Michailova P (1980) A review of the European species of genus Clunio Haliday, 1855 (Diptera, Chronomidae). Zool Anz 205(5-6):417–432
Muthiga NA (2003) Coexistence and reproductive isolation of the sympatric echinoids Diadema savignyi Michelin and Diadema setosum (Leske) on Kenyan coral reefs. Mar Biol 143(4):669–677
Naylor E (1989) Temporal aspects of adaptation in the behavioural physiology of marine animals. In: 21st European marine biology symposium. Polish Academy of Sciences, Gdansk, pp 123–135
Naylor E (2001) Marine animal behaviour in relation to lunar phase. Earth Moon Planets 85-86:291–302
Neumann D (1966) Die lunare und tägliche Schlüpfperiodik der Mücke Clunio: Steuerung und Abstimmung auf die Gezeitenperiodik. Z Vgl Physiol 53(1):1–61
Neumann D (1967) Genetic adaptation in emergence time of Clunio populations to different tidal conditions. Helgol Wiss Meeres 15(1-4):163–171
Neumann D (1983) Die zeitliche Programmierung von Tieren auf periodische Umweltbedingungen. Rhein-Westfael Akad Wiss Nat- Ing- Wirtschwiss 324:31–62
Neumann D (1986) Life cycle strategies of an intertidal midge between subtropic and arctic latitudes. In: Taylor F, Karban R (eds) The evolution of insect life cycles. Springer, New York, pp 3–19
Neumann D (1989) Circadian components of semilunar and lunar timing mechanisms. J Biol Rhythms 4(2):285–294
Neumann D (1995) Physiologische Uhren von Insekten: zur Ökophysiologie lunarperiodisch kontrollierter Fortpflanzungszeiten. Naturwissenschaften 82(7):310–320
Neumann D, Heimbach F (1979) Time cues for semilunar reproduction rhythms in European populations of Clunio marinus. I. The influence of tidal cycles of mechanical disturbance. In: Naylor E, Hartnoll RG (eds) Cyclic phenomena in marine plants and animals. Pergamon Press, Oxford, pp 423–433
Neumann D, Heimbach F (1984) Time cues for semilunar reproduction rhythms in European populations of Clunio marinus. 2. The influence of tidal temperature cycles. Biol Bull 166(3):509–524
O’Donnell AJ, Schneider P, McWatters HG, Reece SE (2011) Fitness costs of disrupting circadian rhythms in malaria parasites. Proc R Soc B Biol Sci 278(1717):2429–2436
Oka H, Hashimoto H (1959) Lunar periodicity in the propagation of a Pacific species of Clunio (Diptera, Chironomidae). Biol Zentralbl 78(4):545–559
Ouyang Y, Andersson CR, Kondo T, Golden SS, Johnson CH (1998) Resonating circadian clocks enhance fitness in Cyanobacteria. Proc Natl Acad Sci USA 95(15):8660–8664
Palstra AP, De Graaf M, Sibbing FA (2004) Riverine spawning and reproductive segregation in a lacustrine cyprinid species flock, facilitated by homing? Anim Biol 54(4):393–415
Palumbi SR (1994) Genetic divergence, reproductive isolation, and marine speciation. Annu Rev Ecol Syst 25:547–572
Pearse JS (1990) Lunar reproductive rhythms in marine invertebrates: maximizing fertilization? Adv Invertebr Reprod 5:311–316
Reid DG, Naylor E (1985) Free-running, endogenous semilunar rhythmicity in a marine isopod crustacean. J Mar Biol Assoc UK 65(1):85–91
Reid DG, Naylor E (1986) an entrainment model for semilunar rhythmic swimming behavior in the marine isopod Eurydice pulchra Leach. J Exp Mar Biol Ecol 100(1–3):25–35
Saigusa M (1980) Entrainment of a semilunar rhythm by a simulated moonlight cycle in the terrestrial crab, Sesarma haematocheir. Oecologia (Berl) 46(1):38–44
Saigusa M (1986) The circa-tidal rhythm of larval release in the incubating crab Sesarma. J Comp Physiol Sens Neural Behav Physiol 159(1):21–31
Saigusa M, Akiyama T (1995) The tidal rhythm of emergence, and the seasonal variation of this synchrony, in an intertidal midge. Biol Bull 188(2):166–178
Saunders DS (1972) Circadian control of larval growth rate in Sarcophaga argyrostoma. Proc Natl Acad Sci USA 69(9):2738–2740
Smith DC (1988) Heritable divergence of Rhagoletis pomonella host races by seasonal asynchrony. Nature (Lond) 336(6194):66–67
Soong K, Leu Y (2005) Adaptive mechanism of the bimodal emergence dates in the intertidal midge Pontomyia oceana. Mar Ecol Prog Ser 286:107–114
Soong KY, Chen JY, Tsao CJ (2006) Adaptation for accuracy or for precision? Diel emergence timing of the intertidal insect Pontomyia oceana (Chironomidae). Mar Biol 150(2):173–181
Tauber CA, Tauber MJ (1977) Sympatric speciation based on allelic changes at three loci: evidence from natural populations in two habitats. Science 197(4310):1298–1299
Tauber E, Roe H, Costa R, Hennessy JM, Kyriacou CP (2003) Temporal mating isolation driven by a behavioral gene in Drosophila. Curr Biol 13(2):140–145
Yerushalmi S, Green RM (2009) Evidence for the adaptive significance of circadian rhythms. Ecol Lett 12(9):970–981
Zantke J, Ishikawa-Fujiwara T, Arboleda E, Lohs C, Schipany K, Hallay N, Straw AD, Todo T, Tessmar-Raible K (2013) Circadian and circalunar clock interactions in a marine annelid. Cell Rep 5(1):99–113
Zhang L, Hastings MH, Green EW, Tauber E, Sladek M, Webster SG, Kyriacou CP, Wilcockson DC (2013) Dissociation of circadian and circatidal timekeeping in the marine crustacean Eurydice pulchra. Curr Biol 23(19):1863–1873
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Japan
About this chapter
Cite this chapter
Kaiser, T.S. (2014). Local Adaptations of Circalunar and Circadian Clocks: The Case of Clunio marinus . In: Numata, H., Helm, B. (eds) Annual, Lunar, and Tidal Clocks. Springer, Tokyo. https://doi.org/10.1007/978-4-431-55261-1_7
Download citation
DOI: https://doi.org/10.1007/978-4-431-55261-1_7
Published:
Publisher Name: Springer, Tokyo
Print ISBN: 978-4-431-55260-4
Online ISBN: 978-4-431-55261-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)