
Abstract
Endometriosis is an oestrogen-dependent inflammatory disease. Endometriosis is often associated with pain symptoms such as dysmenorrhoea, dyspareunia, dyschezia, dysuria and low back pain. Although increased immune cells in peritoneal fluid, adhesions, retrograde menstruation and prostaglandins are considered to be causes of pain symptoms in endometriosis, the underlying mechanisms by which pain is generated still remain unknown. Recently numerous studies have focused on nerve fibres and neurotrophins in eutopic endometrium and endometriotic lesions from women with endometriosis as well as in animal models as a source of pain generation. Nerve fibres in eutopic and ectopic endometrium may be activated and/or sensitised by many inflammatory mediators to cause pain and tenderness. Neurotrophins are known to regulate the survival, development and function of nerve fibres. However, many other molecules may act as a neurotrophic factor in endometriosis. Increased numbers of nerve fibres, increased amount of neurotrophins and different types of nerve fibres in endometriosis may explain why women with endometriosis experience pain.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Different Types of Nerve Fibres
Nerve fibres in the body arise from cell bodies in dorsal root ganglia (DRG) and nerve fibres in the head arise from cell bodies in trigeminal ganglion. Nerve fibres are categorised into six groups, namely Aα, Aβ, Aγ, Aδ, B and C fibres. Cell bodies with the largest diameters give rise to myelinated Aα, Aβ and Aγ fibres, and cell bodies with small and medium diameter give rise to thinly myelinated Aδ and B and unmyelinated C fibres. Aα, Aβ and Aγ fibres are large (5–15 μm in diameter) with myelin sheaths around them and they conduct at a rapid rate (20–100 m/s). Aα, Aβ and Aγ fibres carry the sensation of light pressure to deep muscle and soft touch to skin and do not produce pain. Pain impulses originate at nociceptors, which are sensory neurons that respond to potentially damaging stimuli. When receptors receive pain stimuli, the stimuli are transmitted to the central nervous system (CNS) by either Aδ fibres or C fibres via DRG [1]. Aδ fibres are smaller fibres with myelin sheaths and they conduct at a slower rate (10–25 m/s) than Aα or Aβ fibres due to their size (1–5 μm in diameter). C fibres are small unmyelinated fibres (0.3–1 μm in diameter) and they conduct at a rate of 0.5–2 m/s. Aδ fibres mediate rapid, acute and sharp pain (first pain), and C fibres mediate dull and diffuse pain (second pain) to the CNS [2].
2 Nociceptors and Pain Transmission
There are two types of nociceptors, namely high threshold mechanoreceptors and polymodal nociceptors. High threshold mechanoreceptors respond to mechanical damage such as cutting, crushing or pinching, and polymodal nociceptors respond to all kinds of damaging stimuli, such as irritating chemicals released from injured tissue. Information from high threshold mechanoreceptors are rapidly transmitted to the brain mainly by Aδ fibres, and the location of pain can be recognised. However, information from polymodal nociceptors is transmitted to the brain slowly and the location of pain is usually unrecognised.
Nociceptors can be further divided into peptidergic which contains peptides, such as substance P (SP) and calcitonin gene-related peptide (CGRP), and non-peptidergic which binds isolectin B4 (IB4) [3]. Aδ and C fibres express many of the molecules that have been implicated in the pain activation and sensitisation. These molecules include numerous ion channels and receptors for chemical mediators in their sensory endings. Most nociceptors express only part of the receptors, but prolonged inflammation can lead to an up-regulation of receptors for excitatory compounds. It is likely that the expression of some receptors for chemical mediators may be increased in endometriosis.
The pain from the periphery enters the spinal cord via DRG and it is passed to the nociceptive second-order neurons in the dorsal horn of the grey matter. Second-order neurons are divided into two groups, namely nociceptive-specific (NS) neurons and wide dynamic range (WDR) neurons [4]. Neurotransmitters such as glutamic acid and SP are released from the nerve terminals and they can activate DL-α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptor (AMPA) and N-methyl-d-aspartic acid (NMDA) receptors on the second-order neurons. The pain goes to the thalamus and then enters the cerebral cortex or cerebral limbic system. Not all neurotransmitters activate AMPA or NMDA, and some neurotransmitters such as gamma-aminobutyric acid (GABA) and serotonin suppress pain sensation. Neurotransmitters can actually control pain transmission in the brain and spinal cord.
3 Pain-Producing Substances
The sensitisation and activation of nociceptors after inflammation result from the release of a variety of chemicals by damaged cells and tissues in the vicinity of the injury. These substances include bradykinin, histamine, prostaglandins (PGs), serotonin and nerve growth factor (NGF). They sensitise (lower the threshold) or activate the terminals of the nociceptor by interacting with cell-surface receptors expressed by these neurons. Primary sensory neurons have been shown to contain bioactive peptides that can cause local inflammation. These peptides include SP, CGRP, neuropeptide Y and vasoactive intestinal polypeptide (VIP). Inflammatory mediators activate the non-selective cation channel transient receptor potential vanilloid 1 (TRPV1) which leads to an influx of calcium. Tryptase released from degranulated mast cells cleaves PAR-2 at the plasma membrane of sensory nerve endings. Increased calcium causes depolarisation of the nerve and activated PAR-2 stimulates the release of bioactive peptides including SP and CGRP from sensory nerve endings.
SP and CGRP can contribute to the inflammatory response by causing vasodilation, plasma extravasation (leakage of proteins and fluid from postcapillary venules) and cellular infiltration by interacting with endothelial cells, arterioles, mast cells, neutrophils and immune cells [5]. SP acts on mast cells in the vicinity of sensory nerve endings to evoke degranulation and the release of histamine, which further induces a release of SP and NGF. SP also acts on platelets to release serotonin, providing a positive feedback [6]. CGRP inhibits SP degradation by a specific endopeptidase (SPE) [7] and enhances SP release, amplifying the effects [6]. VIP induces vasodilation and histamine release from mast cells. Histamine and serotonin levels rise in the extracellular space, secondarily sensitising nearby nociceptors. This leads to a gradual spread of hyperalgesia and/or tenderness.
4 Research on Nerve Fibres in the Uterus
Research on nerve fibres in the human uterus began about 1680 [8]. Kilian [9] was one of the first researchers to investigate nerve fibres in the human endometrium [9]. Frankenhauser [10] reported unmyelinated nerve fibres to the smooth muscle of the myometrium with branches to the stroma and the lining epithelium of the human endometrium [10]. Patenko [11], Kostlin [12] and Clivio [13] also demonstrated a fine plexus of unmyelinated nerve fibres in the submucosa from which fine fibrils extended to the epithelium of the human endometrium. Von Gawronsky [14] showed large nerve bundles extending parallel to the endometrial-myometrial junction, with fine branches into the human endometrial stroma [14]. Labhardt [15] and Mabuchi [16] were unable to demonstrate nerve fibres in the human endometrium; however, Dahl [17] described numerous fine nonmodulated nerve fibres in the human endometrium, ending in small treelike branches in the stroma and as straight fibres between the cells of the epithelium. Stöhr [18] reported nerve fibres in the mucosa of the human uterus, even fine fibres to the epithelium. Davis [19] did not find nerve fibres beneath the mucosa of the body of the human uterus. Brown and Hirsch [20] reported nerve fibres in the basal layer of the endometrium in the infantile uterus but were unable to demonstrate the mode of termination in those tissues. These investigators used the silver reduction methods to stain nerve fibres in the human uterus. There is a discrepancy between these studies and it is considered that the silver staining method did not clearly differentiate the nerves and reveal the nerve fibres in the uterus compared with immunohistochemistry.
State and Hirsch [21] used Goldner’s modification of Masson’s trichrome stain to demonstrate nerve fibres in human uteri with no obvious pathologic changes. Some nerve fibres were present in the lower third of the basal layer of the endometrium that have branches terminating in the stroma, in the basolar arterioles and at the origin of the spiral arterioles. No nerve fibres were detected beyond the basal layer of the endometrium. They also demonstrated some nerve fibres in the myometrium and at the endometrial-myometrial junction. In this study, the nerve fibres in the endometrium were found to be unmyelinated. Koppen [22], Stöhr [23], Krantz [24] and Witt [25] reported that branches of nerve fibres accompanying arteries had been clearly revealed only in the basal layer of the endometrium and there were no nerve fibres in the outer two-thirds of the endometrium by Goldner’s modification of Masson’s trichrome stain.
Lerner et al. [26] reported abnormal innervation in the myometrium from women with chronic pelvic pain and dysmenorrhoea. There was a marked proliferation of unmyelinated nerve bundles in the myometrium, but nerve fibres in the endometrium were not mentioned in this study. Quinn and Kirk [27] investigated uterine innervation in normal and some clinical conditions such as adenomyosis and chronic pelvic pain. They performed immunohistostaining using an antibody against protein gene product (PGP9.5). They demonstrated nerve bundles at the endometrial-myometrial interface and throughout the myometrium in nulliparous and parous subjects with no histologic abnormality. There was nerve fibre proliferation throughout the myometrium with small-diameter nerve fibres eccentric courses throughout the myometrial stoma in some subjects with chronic pelvic pain.
Some researchers have identified types of nerve fibres in the human uterus. SP- and CGRP-immunoreactive nerve fibres were present in the human myometrium [28]. Heinrich et al. [29] demonstrated neuropeptide (NPY), SP, neurotensin (NT) and VIP-immunoreactive nerve fibres in the basal layer of the endometrium and myometrium in nonpregnant women. Lynch et al. [30] and Helm et al. [31] also detected VIP-immunoreactive nerve fibres in the endometrium and myometrium in women with no pathologic abnormality.
Nerve fibres in the endometrium in animals have also been investigated by some researchers. A number of tyrosine hydroxylase (TH), dopamine β-hydroxylase (DβH), NPY and VIP-immunoreactive nerve bundles and fibres were present in equine endometrium [32], and CGRP-immunoreactive nerve fibres were in equine and rat endometrium [32, 33], secretoneurin (SN)-immunoreactive nerve fibres were in rat endometrium [34], VIP-immunoreactive nerve fibres were in rat and porcine endometrium [35, 36], adrenergic and acetylcholine (ACh)-immunoreactive nerve fibres were in sheep endometrium [37], and AChE- and NPY-immunoreactive nerve fibres were in rat endometrium [38].
TH-immunoreactive nerve fibres are considered to regulate uterine contractility, uterine blood flow and endometrial secretion as those nerve fibres are often associated with blood vessels, endometrial glands and myometrial smooth muscle. SP and CGRP are co-expressed in a subpopulation of nerve fibres (sensory Aδ and C). SP induces contraction in the human uterus [28] and controls blood flow [39]. In contrast, CGRP is a potent vasodilator [40] and inhibitor of spontaneous contractile activity in the human uterus [28]. Also CGRP inhibits uterine contractility caused by SP in rat uterus [41], regulates sensory transmission and glandular secretion [42, 43] and has a proliferative effect on human endothelial cells [43]. NPY is known to co-exist with noradrenaline and considered to regulate vascular tone and exert inhibitory effect on myometrial contractility [28]. VIP is a potent vasodilator of the uterine artery and involves in smooth muscle relaxation, blood flow increase and secretion [36]. ACh is associated with myometrial and vascular smooth muscle [44] and evokes contraction of the myometrium [45]. VIP co-exists with ACh [46] and promotes relaxation of the myometrium through inhibition of the excitatory action of ACh [47]. CGRP also inhibits ACh-stimulated uterine contraction and this is dose dependent [33]. SN is contained in sensory C fibres and parasympathetic neurons [34] and co-exists with SP and CGRP [48]. SN may regulate vascular control and smooth muscle contraction and play a role in the process of neurogenic inflammation that involves the activation of sensory C fibres, which release SP, neurokinin A, CGRP and nitric oxide from their peripheral terminals to increase vascular permeability, protein extravasation, tissue oedema, vasodilation and activation and recruitment of inflammatory immune cells. TH and DβH are present in adrenergic nerve fibres and they are associated with vascular and nonvascular smooth muscle and participate in the regulation of myometrial contractions and blood flow [49]. It would be interesting to see how these nerve fibres are correlated with variation in pain symptoms and response to hormone treatment.
5 Nerve Fibres and Neurotrophins in the Uterus from Women with Endometriosis
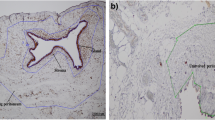
Quinn and Kirk [27] reported that there was widespread nerve fibre proliferation (small-diameter nerve fibres) in the uterine isthmus suggesting nerve fibre damage in the uterine isthmus in women complaining of chronic pelvic pain. Few of these women had endometriosis, but they did report widespread nerve fibre proliferation in the uterus and cervix in one woman with advanced endometriosis and with painful periods and intercourse. These nerve fibres were prominently seen around arteries and veins throughout the uterine isthmus. Quinn and Armstrong [50] reported widespread nerve fibre proliferation with extensive perivascular nerve fibre proliferation around both arteries and veins in the myometrium in an endometriosis patient with dysmenorrhoea and dyspareunia. In another study, increased numbers of nerve fibres were seen in the myometrium from women with advanced endometriosis compared with women without endometriosis [51]. These studies demonstrated the proliferation of nerve fibres; however, types of nerve fibres in the endometrium and myometrium from women with endometriosis were not investigated in these studies. It has been demonstrated that no nerve fibres were detected in the functional layer of the endometrium from women without endometriosis; however, there were several small nerve fibres present in the functional layer of the endometrium from women with endometriosis (the mean density of nerve fibres: 13.4 mm2) [52]. There were more nerve fibres in the basal layer of the endometrium and myometrium from women with endometriosis than women without endometriosis. Several thick nerve trunks were seen in the basal layer of the endometrium or at the endometrial-myometrial interface in women with endometriosis, but these nerve fibres were not seen in women without endometriosis. Many small nerve fibres were also present throughout the basal layer of the endometrium, but only a few nerve fibres were seen in women without endometriosis. Many more nerve fibres and nerve trunks were seen in the myometrium in women with endometriosis than in women without endometriosis. In women with endometriosis, nerve fibres in the functional layer of the endometrium were sensory C fibres; sensory C, sensory Aδ and adrenergic fibres in the basal layer; and sensory C, sensory Aδ, adrenergic and cholinergic fibres in the myometrium [53]. These results indicate that abnormal innervation in the uterus may be associated with pain generation in women with endometriosis who suffer from chronic pelvic pain.
Progestogens and combined oral contraceptives are often used to treat women with endometriosis-associated pain. Progestogens and combined oral contraceptives significantly reduced the nerve fibre density in the functional and the basal layers of the endometrium and myometrium from women with hormonally treated endometriosis compared with that from women with untreated endometriosis [54]. Hormonal treatments may reduce pain symptoms by decreasing nerve fibres in the endometrium and myometrium in women with endometriosis.
6 Conscious Pain Mapping
Several groups have studied conscious pain mapping since it has been assumed that endometriotic lesions are the source of localised pain and tenderness, and conscious pain mapping may help to identify potential areas causing pelvic pain. Palter and Olive [55] first described conscious pain mapping to identify a focal source of pain and generalised visceral hypersensitivity in a majority of women with chronic pelvic pain (CPP). They found that deep infiltrating sclerotic endometriosis of the rectum, active peritoneal endometriosis and an adhesion of small bowel were apparent sources of cause of pain. Demco [56] found that most women localised or mapped their pain to their endometriotic lesions and the surrounding peritoneum. In this study, pain extended beyond the lesions to normal-looking peritoneum. Almeida and Val-Gallas [57] found 48 positive findings in 50 women with conscious pain mapping and Howard et al. [58] also reported that conscious pain mapping successfully identified tender lesion in 70 % of women. It seems that most endometriotic lesions themselves appear to be capable of generating pain stimuli, and conscious pain mapping can be useful in detecting a source of pain. However, not all endometriotic lesions produce pain and it is likely that there may be other sources of pain generation in women with endometriosis.
7 Nerve Fibres in Endometriotic Lesions
Tamburro et al. [59] demonstrated the expression of transforming growth factor β1 (TGFβ1) in nerve fibres in endometriotic lesions from women with dysmenorrhoea associated with endometriosis. There was a significant correlation between the dysmenorrhoea and maximal intensity of staining of TGFβ1 and a relationship between the colour of the lesions and maximal intensity of staining of TGFβ1. Greater maximal intensity of staining of TGFβ1 was seen in red, deep lesions than in black and normal peritoneum. TGFβ1 is increased in the peritoneal fluid of women with endometriosis [60] and can increase cyclooxygenase-2 (COX-2) activity to produce more PGs [59]. Since there were no morphologic differences in the nerve fibres between endometriotic peritoneal lesions and normal peritoneum, substances that are present in the nerve fibres appear to be associated with causing pelvic pain in women with endometriosis. Tulandi et al. [61] demonstrated nerve fibres in endometriotic lesions in women with endometriosis by using an antibody against neurofilament (NF). They reported that the distance between endometrial glands and nerve fibres in endometriotic lesions from women with pain was closer than in women with no pain. Quinn and Kirk [62] reported widespread nerve fibre proliferation (small-diameter nerve fibres) in endometriotic lesions from a woman with pelvic pain. Berkley et al. [63] demonstrated nerve fibres in endometriotic lesions in a rat model and these nerve fibres were protein gene product 9.5 (PGP9.5), CGRP, SP and vesicular monoamine transporter (VMAT) immunoreactive. SP and CGRP are present in myelinated and unmyelinated sensory nerve fibres (Aδ fibres and C fibres), and VMAT is present in sympathetic fibres.
In peritoneal endometriotic lesion from endometriosis patients who presented pain symptoms, nerve fibres were innervated by sensory C, sensory Aδ, cholinergic and adrenergic fibres, and the mean density of nerve fibres was 16 mm2. Progestogens and combined oral contraceptives significantly reduced nerve fibre density in peritoneal endometriotic lesions from hormone-treated women with endometriosis (the mean density of nerve fibres: 10.6 mm2) compared with peritoneal endometriotic lesions from untreated women with endometriosis (the mean density of nerve fibres: 16.3 mm2) [65]. Nerve fibres expressing growth-associated protein 43, which is a marker of neuronal development and sprouting in peritoneal endometriotic lesion as well as SP, have also been reported by another group [66]. The same group has shown the relationship between the severity of pain symptoms and the density of nerve fibres in peritoneal endometriotic lesions [67]. There were more nerve fibres stained with PGP 9.5 and NF in endometriosis patients whose pain score was at least 3 or more than those whose pain score was 2 or less. This study has shown that there is a correlation between the density of nerve fibres and pain severity in peritoneal endometriotic lesions in women with endometriosis. Alvarez et al. have developed peritoneal endometriotic lesions in a rat model by implanting with autologous uterus in the gastrocnemius muscle [68]. Those lesions showed peptidergic nerve fibres (CGRP-positive), non-peptidergic nerve fibres (IB4-positive) as well as GAP43-positive nerve fibres. Since no studies have demonstrated IB4-positive nerve fibres in endometriotic lesions from women with endometriosis, it would be interesting to investigate the presence of IB4-positive nerve fibres and to find out the roles of these nerve fibres in endometriosis.
Deep infiltrating endometriosis (DIE) is often associated with severe pain and is defined as endometriotic lesions penetrating into the retroperitoneal space or the wall of the pelvic organs for a distance of 5 mm or more [69]. Patients with deep pelvic infiltrating endometriosis usually have much stronger pain than those with other types of endometriosis [69–71]. Anaf et al. [70] have demonstrated that endometriosis patients with DIE had significantly higher preoperative pain scores than patients with peritoneal or ovarian endometriosis. In their study, mast cells located <25 μm from nerve fibres were significantly more abundant in DIE than in peritoneal and ovarian endometriosis. Wang et al. have reported that there were more nerve fibres in DIE (the mean density of nerve fibres: 68 mm2) than in peritoneal endometriosis (the mean density of nerve fibres: 16 mm2) [72]. Those nerve fibres were sensory C, sensory Aδ, cholinergic and adrenergic fibres. When compared with the locations of DIE, more nerve fibres were detected in DIE in the sigmoid colon, appendix and rectum (the mean density of nerve fibres: 172 mm2) than in the uterosacral ligament, cul-de-sac or peritoneal sidewall (the mean density of nerve fibres: 68 mm2).
There are many substances secreted from endometriotic lesions including PGs [73], tumour necrosis factor-α (TNF-α) [74] and NGF [75]. These substances could sensitise and/or activate sensory and sympathetic nerve fibres that are present in endometriotic lesions. TRPV1, a key molecule in nociception, has been identified in the human cervix uteri in the nonpregnant state [76]. The levels of TRPV1 can be increased in painful inflammatory diseases, and TRPV1 can trigger the release of SP and CGRP from peripheral nerve terminals during inflammation [77]. It would be interesting to see whether TRPVI may be present in ectopic endometrium in women with endometriosis.
Most women with endometriosis present some kind of pain symptoms such as dysmenorrhoea or dyspareunia. However, a few endometriosis patients do not show any pain symptoms. When peritoneal endometriotic lesions from women with endometriosis who did not have any pain symptoms were stained with PGP 9.5, some nerve fibres were still detected. Therefore, not only the presence of nerve fibres but also types of nerve fibres or molecules in endometriotic lesions may be associated with pain generation in women with endometriosis. There may be some molecules which can desensitise nerve fibres in those who do not experience pain symptoms. Further studies will be required as to nerve fibres and substances secreted from endometriotic lesions from endometriosis patients who do not have any pain symptoms.
8 Neurotrophins (NGF, BDNF, NT-3, NT-4/5) in Eutopic and Ectopic Endometrium
NGF belongs to the neurotrophin family, together with brain-derived neurotrophic factor (BDNF) [78], neurotrophins-3 (NT-3) [79] and neurotrophins-4/5 (NT-4/5) [80]. NGF exerts its effects by binding to a tyrosine kinase A receptor (TrkA), and BDNF and NT-4/5 bind TrkB, and NT-3 binds TrkC instead [81]. p75, a low-affinity glycoprotein, also binds all neurotrophins [82].
NGF was strongly expressed near endometriotic glands in peritoneal and ovarian endometriosis [64, 75], and neurotrophin-3 (NT-3) expression was also observed in peritoneal endometriosis [66]. In ovarian endometriosis, NGF, BDNF, NT-3, NT-4/5 and NTRK2 mRNA expressions were detected by quantitative reverse transcriptase-polymerase chain reaction (RT-PCR) [83]. Browne et al. have demonstrated that NT-4/5 and BDNF mRNAs concentrations in eutopic endometrium detected on the antibody microarrays and reverse transcriptase-polymerase chain reaction were much higher in endometriosis patients than in women without endometriosis [84]. However, another study has reported that there was no difference in amounts of NGF, BDNF, NT-3, TrkA and NGFRp75 in eutopic endometrium between women with and without endometriosis using immunofluorescence staining, Western blot and a neuronal growth assay [85]. The discrepancy may be due to different kinds of pain symptoms, severity of pain or the density of nerve fibres among those patients. Much more detailed studies will be needed to determine the expression of these neurotrophins.
NGF has several effects through a variety of mechanisms. NGF stimulates the development, growth and survival of neurons, particularly of sensory neurons [86]. NGF increases synthesis and release of neuropeptides such as SP and CGRP from sensory nerve endings [87] and stimulates degranulation of mast cells. Recent evidence has implicated NGF as a key mediator of inflammation and pain. NGF is up-regulated in nerve fibres associated with the inflamed area during inflammation. NGF can increase the sensitivity and excitability of peripheral neurons to cause pain by directly or indirectly sensitising sensory nerve fibres [88]. NGF in turn activates mast cells and neutrophils which can release additional inflammatory mediators such as histamine, serotonin and bradykinin to cause hypersensitivity [89]. NGF has also been reported to promote angiogenesis [90]. NGF induces proliferation of endothelial cells [91] and of vascular smooth muscle cells [92] and stimulates the production of vascular endothelial growth factor (VEGF) [93]. NGF is expressed in vascular endothelial cells [94] and a variety of cell types such as T and B lymphocytes [95], mast cells [96], eosinophils [97], monocytes/macrophages [98], neutrophils and basophils [99]. Increased angiogenesis in eutopic endometrium in women with endometriosis demonstrated by several studies [100, 101] and increased numbers of immune cells may lead to further production of NGF, promoting further nerve fibre outgrowth and pain sensation.
9 Effects of Oestrogen on Nerve Fibres
It is well established that oestrogen has multiple effects on the female reproductive system and peripheral nervous system including pain sensitivity and neural regulation of vascular function. Endometriosis is oestrogen dependent for continued growth and proliferation, and it usually becomes less active with menopause as the oestrogen level decreases. However, oestrogen replacement therapy can reactivate the disease. Oophorectomy significantly decreased NGF and BDNF levels while oestrogen treatment increased these levels [102, 103], and oestrogen also up-regulated NGF and BDNF expressions in the endometrium [104–106]. Several studies have shown that neuroprotective effects of oestrogen were blocked by ER antagonists [107–109]. Ovariectomy caused a significant decrease in NGF protein content in the uterus, and short-term treatment of ovariectomised mice with oestrogen and/or progesterone increased uterine NGF mRNA and restored NGF protein to concentrations similar to intact control mice [110].
In animals, stronger immunostaining of NGF and TrkA was observed in luminal epithelial cells and glandular cells in the estrous period and early pregnancy as compared to the non-breeding period in the uterus of the wild ground squirrels [111]. p75 was immunolocalised only in luminal epithelial and glandular cells during the estrous period, early pregnancy and non-breeding period. The mean mRNA levels of NGF and TrkA and p75 were significantly higher in the estrous period and early pregnancy as compared to the non-breeding period.
Strong immunostaining of NGF and its receptors NTRK1 and TNFRSF1B was observed in uteri of golden hamsters on the day of proestrus as compared to the other stages of the estrous cycle [112]. There was a positive correlation between uterine NGF expression and plasma concentrations of estradiol-17beta, and estradiol-17beta stimulated expression of NGF and its two receptors in the uterus.
Neutral endopeptidase (NEP) is the enzyme responsible for degradation of SP and oestrogen treatment of ovariectomised rats resulted in a four-fold decrease in uterine NEP relative to control ovariectomised rats, resulting in increased SP levels [113]. Angiotensin-converting enzyme (ACE) is an enzyme responsible for degradation of bradykinin and treatment of postmenopausal women with oestrogen caused an increase in bradykinin and a decrease in ACE [114]. Steroidal hormones may induce further production of neuropeptides and neurotransmitters by altering enzyme expressions to sensitise sensory nerve terminals to cause prolonged pain sensation.
Oestrogen receptor-alpha (ER-alpha) and ER-beta are expressed in sensory neurons of the dorsal root ganglia (DRG) [115], so oestrogen could directly act on sensory nerve fibres to enhance axon outgrowth. Receptors for other steroidal hormones such as progesterone may be expressed in nerve fibres and they may also have direct effects on nerve fibres.
10 Prostaglandins and Neurotrophins
The correlation between prostaglandins and endometriosis-associated pain is well established. PGE2 is present in sensory and sympathetic nerve fibres and can sensitise and activate sensory nerve fibre terminals to induce pain [116]. Prostaglandins may act neuroprotectively to induce the production and release of neurotrophins to promote the survival and outgrowth of nerve fibres. Toyomoto et al. [117] reported that PGE2 induced and stimulated the secretion and synthesis of NGF and BDNF, and Kanda et al. [118] also demonstrated that PGE2 enhanced the production of neurotrophin-4 (NT-4) via EP3 receptor.
COX-2 is an enzyme which catalyses the synthesis of prostaglandins from arachidonic acid and COX-2 may have a key function with respect to inflammation and pain. Previous studies have shown that COX-2 is expressed in neurons and COX-2 expression in eutopic endometrium in women with endometriosis was higher than in those without endometriosis [119]. Also, increased COX-2 expression in endometriotic lesions of different anatomical sites (ovarian, peritoneal and deep infiltrating endometriosis) has been reported [120–122], resulting in further modification of the production of prostaglandins in different sites.
11 Molecules That May Be Associated with Nerve Fibre Growth in Endometriosis
Several molecules are present in eutopic endometrium and endometriotic lesions in women with endometriosis and some of them may have effects on nerve fibre outgrowth. Insulin-like growth factor I (IGF-I) was present in eutopic endometrium and endometriotic lesions in women with endometriosis [123]. IGF-1 induced sensory nerve fibre growth and potentiated the NGF-induced neuritogenesis [124, 125]. Oestrogen also regulated the expression and synthesis of IGF-I and its receptor [126, 127].
Bcl-2 was present in eutopic endometrium and endometriotic lesions in women with endometriosis [128]. Bcl-2 is expressed in neurons and promoted axonal growth rates in sensory nerve fibres [129, 130]. Oestrogen up-regulated the expression of Bcl-2 [131], and oestrogen treatment increased the number of Bcl-2 immunoreactive nerve fibres [132]. Both oestrogen and progesterone can increase the expression of Bcl-2 [133, 134] and Bcl-2 has been shown to inhibit neuronal death [133]. Therefore oestrogen and progesterone may directly affect cell survival or prevent neuronal cell death in neurons by Bcl-2-induced inhibition of cell death and axonal growth, resulting in increased nerve fibre densities.
Hepatocyte growth factor (HGF) was up-regulated in eutopic endometrium in women with endometriosis [135]. HGF alone had no outgrowth-promoting activity, but it co-operated with NGF in enhancing axonal growth of sensory nerves and also enhanced the neurotrophic activities of NGF [136]. Oestrogen also increased the production of HGF by peritoneal macrophages in women with endometriosis [137].
Significantly increased expression of heat shock protein 27 (HSP 27) was reported in eutopic endometrium in women with endometriosis [138]. HSP27 was expressed by sensory nerve fibres [139, 140] and significantly increased survival for rat sensory and sympathetic nerve fibres after axotomy or NGF withdrawal [141]. HSP expression was stimulated by oestrogen in the endometrium [142].
Fibroblast growth factor 9 (FGF-9) was present in endometriotic lesions in women with endometriosis [143]. FGF-9 enhanced survival of cholinergic nerve fibres [144] and administration of 17β-estradiol induced FGF-9 expression in endometriotic lesions [143].
These molecules may be associated with increased nerve fibre densities, specially sensory and autonomic neurons in women with endometriosis to induce pain, and steroidal hormones may promote the synthesis of these molecules in women with endometriosis
12 Angiogenic Molecules
Novella-Maestre et al. [145] have demonstrated that antiangiogenic treatment with cabergoline significantly reduced the number of immature blood vessels, nerve fibres, mast cells and macrophages in endometriotic lesions in a mouse model of endometriosis. Vascular endothelial growth factor (VEGF) is a secreted vascular mitogen that is specific for endothelial cells and plays an important regulatory role in vascular growth during development [146]. VEGF is increased in DIE [147] and peritoneal fluid [148] in women with endometriosis compared with women without. It is accumulating evidence that VEGF acts as a neurotrophic and neuroprotective factor [149]. VEGF application to cultured peripheral adult ganglia caused significant neuritic outgrowth [150] and peripheral nerve regeneration in vivo has been shown to be enhanced by VEGF application [151]. There are some proangiogenic factors, such as IL-8 [152], hepatocyte growth factor (HGF) [153], erythropoietin [154], angiogenin [155], macrophage migration inhibitory factor [156], neutrophil-activating factor [157] and TNF-α [158], and these proangiogenic factors may also have neurotrophic effects and be involved in nerve fibre growth in endometriosis.
13 Conclusions
Increased numbers of nerve fibres in eutopic endometrium (both the functional layer and the basal layer), myometrium and endometriotic lesions may contribute in some way to the mediation of pain in women with endometriosis. It was always been assured that the pain of endometriosis is generated in the endometriotic lesions, but the findings in some studies allow the possibility that some of the pain may actually arise within the endometrium.
Endometriosis is an inflammatory condition and many chemical and inflammatory mediators such as bradykinin, prostaglandins as well as neurotrophins can be released from both eutopic endometrium and endometriotic lesions. These substances can directly activate and/or sensitise sensory nerve endings by interacting with cell-surface receptors and trigger the release of pain mediators from other cells and afferent nerve fibres. Those pain mediators sensitise nerve endings, resulting in an increased response to painful stimuli. Prostaglandins and other arachidonic acid derivatives can increase the sensitivity of nerve endings to bradykinin or other pain-producing substances, leading to a secondary sensitisation of nearby sensory nerve fibres.
There may be a correlation between pain severity and nerve fibre density; however, it seems that some nerve fibres in eutopic and ectopic endometrium are not associated with pain generation. Further research will be needed to better understand the roles of nerve fibres and molecules secreted from eutopic and ectopic endometrium from women with endometriosis.
References
Adrienne ED, Ardem P. Nociceptors: the sensors of the pain pathway. J Clin Invest. 2010;120:3760–72.
Harkins SW, Davis MD, Bush FM, Kasberger J. Suppression of first pain and slow temporal summation of second pain in relation to age. J Gerontol A Biol Sci Med Sci. 1996;51:M260–5.
Molliver DC, Wright DE, Leitner ML, Parsadanian AS, Doster K, Wen D, Yan Q, Snider WD. IB4-binding DRG neurons switch from NGF to GDNF dependence in early postnatal life. Neuron. 1997;19:849–61.
Miraucourt LS, Dallel R, Voisin DL. Glycine inhibitory dysfunction turns touch into pain through PKCgamma interneurons. PLoS One. 2007;2:e1116.
Maggi CA. Tachykinins and calcitonin gene-related peptide (CGRP) as co-transmitters released from peripheral endings of sensory nerves. Prog Neurobiol. 1995;45:1–98.
Steinhoff M, Stander S, Seeliger S, Ansel JC, Schmelz M, Luger T. Modern aspects of cutaneous neurogenic inflammation. Arch Dermatol. 2003;139:1479–88.
Le Greves P, Nyberg F, Terenius L, Hokfelt T. Calcitonin gene-related peptide is a potent inhibitor of substance P degradation. Eur J Pharmacol. 1985;115:309–11.
Wilissius T. Cerebri anatome nervorumque descriptio et usus. I Opra Omnia. Geneva; 1680. p. 1.
Kilian FM. Ztschr F Rat Med. 1851;10:41.
Frankenhauser F. Die nerven der Gebärmutter und ihre Endigungen in der glatten Muskelfascrn: Ein Beitrag zur Anatomie und Gynäkologie, Insug. Dissert, Jcna, Fr. Manke; 1867.
Patenko T. Zentralbl F Gynäk. 1880;19:442.
Kostlin R. Die Nervendigungen in den weiblichen Geschlechtsorganen. Fortschr Med. 1894;12:411–21.
Clivio I. Contributo alla conoscenza delle terminazioni nervose dell’utero, Pavia, tipog e legat.coop; 1894.
Von Gawronsky N. Arch F Gynäk. 1894;47:271.
Labhardt A. Arch F Gynäk. 1906;80:135.
Mabuchi K. Mitt. a. d. med. Fak. d. k. Univ. Tokyo; 1924. 81: 385.
Dahl W. Ztschr F Geburtsh U Gynäk. 1916;78:539.
Stöhr P. In: von Möllendorff W, editors. Handbuch der mikroscopischen Anatomie des Menschen. Berlin: Julius Springer; 1928. 4(1):393.
Davis AA. J Obst Gynaec Brit Emp. 1933;40:481.
Brown WH, Hirch EF. The intrinsic nerves of the immature human uterus. Am J Pathol. 1941;17:731–9.
State D, Hirsch E. The distribution of the nerves to the adult endometrium. Arch Pathol. 1941;32:939.
Koppen K. Results of a histologic study of uterine innervation. Arch Gynakol. 1950;177:354–91.
Stöhr PH Jr. Mikroskopische Anatomie des vegetativcn Nervensystems. In: Handbuch der mikroskopischen Anatomic des Menschen, Bd.IV/5, Nervensystem. Berlin-Göttingen-Haidelberg: Springer; 1957.
Krantz KE. Innervation of the human uterus. Ann N Y Acad Sci. 1959;75:770–84.
Witt HJ. Strunkturelementc und funktionelle Gesamtheit des Endometriuma. Lichtoptische Morphologie. I. Das normale menschliche Endometrium. Hrsg. V. H. Schmidt-Matthiesen. Stuttgart: Georg Thieme; 1963.
Lerner EJ, Jaffe M, Ree HJ, McDuff Jr HC. Proliferation of myometrial nerves in a patient with severe dysmenorrhea. R I Med J. 1985;68:265–7.
Quinn MJ, Kirk N. Differences in uterine innervation at hysterectomy. Am J Obstet Gynecol. 2002;187:1515–9. discussion 1519–20.
Samuelson UE, Dalsgaard CJ, Lundberg JM, Hokfelt T. Calcitonin gene-related peptide inhibits spontaneous contractions in human uterus and fallopian tube. Neurosci Lett. 1985;62:225–30.
Heinrich D, Reinecke M, Forssmann WG. Peptidergic innervation of the human and guinea pig uterus. Arch Gynecol. 1986;237:213–9.
Lynch EM, Wharton J, Bryant MG, Bloom SR, Polak JM, Elder MG. The differential distribution of vasoactive intestinal polypeptide in the normal human female genital tract. Histochemistry. 1980;67:169–77.
Helm G, Ottesen B, Fahrenkrug J, Larsen JJ, Owman C, Sjoberg NO, Stolberg B, Sundler F, et al. Vasoactive intestinal polypeptide (VIP) in the human female reproductive tract: distribution and motor effects. Biol Reprod. 1981;25:227–34.
Bae SE, Corcoran BM, Watson ED. Immunohistochemical study of the distribution of adrenergic and peptidergic innervation in the equine uterus and the cervix. Reproduction. 2001;122:275–82.
Shew RL, Papka RE, McNeill DL. Calcitonin gene-related peptide in the rat uterus: presence in nerves and effects on uterine contraction. Peptides. 1990;11:583–9.
Collins JJ, Wilson K, Fischer-Colbrie R, Papka RE. Distribution and origin of secretoneurin-immunoreactive nerves in the female rat uterus. Neuroscience. 2000;95:255–64.
Houdeau E, Prudhomme MJ, Rousseau JP. Regional difference in the distribution of vasoactive intestinal polypeptide-immunoreactive nerve fibres along the uterus and between myometrial muscle layers in the rat. Histochem J. 1998;30:525–9.
Rodriguez R, Pozuelo JM, Martin R, Arriazu R, Santamaria L. Stereological quantification of nerve fibers immunoreactive to PGP 9.5, NPY, and VIP in rat prostate during postnatal development. J Androl. 2005;26:197–204.
Renegar RH, Rexroad Jr CE. Uterine adrenergic and cholinesterase-positive nerves and myometrial catecholamine concentrations during pregnancy in sheep. Acta Anat (Basel). 1990;137:373–81.
Papka RE, Cotton JP, Traurig HH. Comparative distribution of neuropeptide tyrosine-, vasoactive intestinal polypeptide-, substance P-immunoreactive, acetylcholinesterase-positive and noradrenergic nerves in the reproductive tract of the female rat. Cell Tissue Res. 1985;242:475–90.
Otsuka M, Konishi S, Yanagisawa M, Tsunoo A, Akagi H. Role of substance P as a sensory transmitter in spinal cord and sympathetic ganglia. Ciba Found Symp. 1982;91:13–34.
Sato S, Hayashi RH, Garfield RE. Mechanical responses of the rat uterus, cervix, and bladder to stimulation of hypogastric and pelvic nerves in vivo. Biol Reprod. 1989;40:209–19.
Shew RL, Papka RE, McNeill DL, Yee JA. NADPH-diaphorase-positive nerves and the role of nitric oxide in CGRP relaxation of uterine contraction. Peptides. 1993;14:637–41.
Gibson SJ, Polak JM, Bloom SR, Sabate IM, Mulderry PM, Ghatei MA, McGregor GP, Morrison JF, et al. Calcitonin gene-related peptide immunoreactivity in the spinal cord of man and of eight other species. J Neurosci. 1984;4:3101–11.
Onuoha GN, Alpar EK. Levels of vasodilators (SP, CGRP) and vasoconstrictor (NPY) peptides in early human burns. Eur J Clin Invest. 2001;31:253–7.
Adham N, Schenk EA. Autonomic innervation of the rat vagina, cervix, and uterus and its cyclic cariation. Am J Obstet Gynecol. 1964;104:508–16.
Traurig HH, Papka RE. Autonomic efferent and visceral sensory innervation of the female reproductive system: special reference to the functional roles of nerves in reproductive organs. In: Maggi CA, editor. Nervous control of the urogenital system. Chur: Harwood Academic; 1993. p. 103–41.
Houdeau E, Boyer PA, Rousseau A, Rousseau JP. Coexpression of neuropeptide Y and vasoactive intestinal polypeptide in pelvic plexus neurones innervating the uterus and cervix in the rat. Cell Tissue Res. 1997;288:285–92.
Stjernquist M, Owman C. Interaction of noradrenaline, NPY and VIP with the neurogenic cholinergic response of the rat uterine cervix in vitro. Acta Physiol Scand. 1987;131:553–62.
Kirchmair R, Marksteiner J, Troger J, Mahata SK, Mahata M, Donnerer J, Amann R, Fischer-Colbrie R, Winkler H, Saria A. Human and rat primary C-fibre afferents store and release secretoneurin, a novel neuropeptide. Eur J Neurosci. 1994;6:861–8.
Vera PL, HaaseE B, Schramm LP. Origins of the sympathetic innervation of the cervical end of the uterus in the rat. Brain Res. 1997;747:140–3.
Quinn M, Armstrong G. Uterine nerve fibre proliferation in advanced endometriosis. J Obstet Gynaecol. 2004;24:932–3.
Atwal G, du Plessis D, Armstrong G, Slade R, Quinn M. Uterine innervation after hysterectomy for chronic pelvic pain with, and without, endometriosis. Am J Obstet Gynecol. 2005;193:1650–5.
Tokushige N, Markham R, Russell P, Fraser IS. High density of small nerve fibres in the functional layer of the endometrium in women with endometriosis. Hum Reprod. 2006;21:782–7.
Tokushige N, Markham R, Russell P, Fraser IS. Different types of small nerve fibers in eutopic endometrium and myometrium in women with endometriosis. Fertil Steril. 2007;88:795–803.
Tokushige N, Markham R, Russell P, Fraser IS. Effects of hormonal treatment on nerve fibers in endometrium and myometrium in women with endometriosis. Fertil Steril. 2008;90:1589–98.
Palter SF, Olive DL. Office microlaparoscopy under local anesthesia for chronic pelvic pain. J Am Assoc Gynecol Laparosc. 1996;3:359–64.
Demco L. Mapping the source and character of pain due to endometriosis by patient-assisted laparoscopy. J Am Assoc Gynecol Laparosc. 1998;5:241–5.
Almeida Jr OD, Val-Gallas JM. Conscious pain mapping. J Am Assoc Gynecol Laparosc. 1997;4:587–90.
Howard FM, El-Minawi AM, Sanchez RA. Conscious pain mapping by laparoscopy in women with chronic pelvic pain. Obstet Gynecol. 2000;96:934–9.
Tamburro S, Canis M, Albuisson E, Dechelotte P, Darcha C, Mage G. Expression of transforming growth factor beta1 in nerve fibers is related to dysmenorrhea and laparoscopic appearance of endometriotic implants. Fertil Steril. 2003;80:1131–6.
Pizzo A, Salmeri FM, Ardita FV, Sofo V, Tripepi M, Marsico S. Behaviour of cytokine levels in serum and peritoneal fluid of women with endometriosis. Gynecol Obstet Invest. 2002;54:82–7.
Tulandi T, Felemban A, Chen MF. Nerve fibers and histopathology of endometriosis-harboring peritoneum. J Am Assoc Gynecol Laparosc. 2001;8:95–8.
Quinn M, Kirk N. Uterosacral nerve fibre proliferation in parous endometriosis. J Obstet Gynaecol. 2004;24:189–90.
Berkley KJ, Dmitrieva N, Curtis KS, Papka RE. Innervation of ectopic endometrium in a rat model of endometriosis. Proc Natl Acad Sci U S A. 2004;101:11094–8.
Tokushige N, Markham R, Russell P, Fraser IS. Nerve fibres in peritoneal endometriosis. Hum Reprod. 2006;21:3001–7.
Tokushige N, Markham R, Russell P, Fraser IS. Effect of progestogens and combined oral contraceptives on nerve fibers in peritoneal endometriosis. Fertil Steril. 2009;92:1234–9.
Mechsner S, Schwarz J, Thode J, Loddenkemper C, Salomon DS, Ebert AD. Growth-associated protein 43-positive sensory nerve fibers accompanied by immature vessels are located in or near peritoneal endometriotic lesions. Fertil Steril. 2007;88:581–7.
Mechsner S, Kaiser A, Kopf A, Gericke C, Ebert A, Bartley J. A pilot study to evaluate the clinical relevance of endometriosis-associated nerve fibers in peritoneal endometriotic lesions. Fertil Steril. 2009;92:1856–61.
Alvarez P, Chen X, Hendrich J, Irwin JC, Green PG, Giudice LC, Levine JD. Ectopic uterine tissue as a chronic pain generator. Neuroscience. 2012;225:269–82.
Koninckx PR, Meuleman C, Demeyere S, Lesaffre E, Cornillie FJ. Suggestive evidence that pelvic endometriosis is a progressive disease, whereas deeply infiltrating endometriosis is associated with pelvic pain. Fertil Steril. 1991;55:759–65.
Anaf V, Chapron C, El Nakadi I, De Moor V, Simonart T, Noël JC. Pain, mast cells, and nerves in peritoneal, ovarian, and deep infiltrating endometriosis. Fertil Steril. 2006;86:1336–43.
Vercellini P, Somigliana E, Viganò P, Abbiati A, Daguati R, Crosignani PG. Endometriosis: current and future medical therapies. Best Pract Res Clin Obstet Gynaecol. 2008;22:275–306.
Wang G, Tokushige N, Markham R, Fraser IS. Rich innervation of deep infiltrating endometriosis. Hum Reprod. 2009;24:827–34.
Ebert AD, Bartley J, David M. Aromatase inhibitors and cyclooxygenase-2 (COX-2) inhibitors in endometriosis: new questions–old answers? Eur J Obstet Gynecol Reprod Biol. 2005;122:144–50.
Bergqvist A, Nejaty H, Froysa B, Bruse C, Carlberg M, Sjoblom P, Söder O. Production of interleukins 1beta, 6 and 8 and tumor necrosis factor alpha in separated and cultured endometrial and endometriotic stromal and epithelial cells. Gynecol Obstet Invest. 2000;50:1–6.
Anaf V, Simon P, El Nakadi I, Fayt I, Simonart T, Buxant F, Noel JC. Hyperalgesia, nerve infiltration and nerve growth factor expression in deep adenomyotic nodules, peritoneal and ovarian endometriosis. Hum Reprod. 2002;17:1895–900.
Tingåker BK, Irestedt L. Changes in uterine innervation in pregnancy and during labour. Curr Opin Anaesthesiol. 2010;23:300–3.
Caterina MJ, Julius D. The vanilloid receptor: a molecular gateway to the pain pathway. Annu Rev Neurosci. 2001;24:487–517.
Barde YA. The nerve growth factor family. Prog Growth Factor Res. 1990;2:237–48.
Hohn A, Leibrock J, Bailey K, Barde YA. Identification and characterization of a novel member of the nerve growth factor/brain-derived neurotrophic factor family. Nature. 1990;344:339–41.
Hallbook F, Ibanez CF, Persson H. Evolutionary studies of the nerve growth factor family reveal a novel member abundantly expressed in Xenopus ovary. Neuron. 1991;6:845–58.
Mantyh PW, Koltzenburg M, Mendell LM, Tive L, Shelton DL. Antagonism of nerve growth factor-TrkA signaling and the relief of pain. Anesthesiology. 2011;115:189–204.
Meakin SO, Shooter EM. The nerve growth factor family of receptors. Trends Neurosci. 1992;15:323–31.
Borghese B, Vaiman D, Mondon F, Mbaye M, Anaf V, Noël JC, de Ziegler D, Chapron C. Neurotrophins and pain in endometriosis. Gynecol Obstet Fertil. 2010;38:442–6.
Browne AS, Yu J, Huang RP, Francisco AM, Sidell N, Taylor RN. Proteomic identification of neurotrophins in the eutopic endometrium of women with endometriosis. Fertil Steril. 2012;98:713–9.
Barcena de Arellano ML, Arnold J, Lang H, Vercellino GF, Chiantera V, Schneider A, Mechsner S. Evidence of neurotrophic events due to peritoneal endometriotic lesions. Cytokine. 2013;62:253–61.
Levi-Montalcini R, Skaper SD, Dal Toso R, Petrelli L, Leon A. Nerve growth factor: from neurotrophin to neurokine. Trends Neurosci. 1996;19:514–20.
Hunter DD, Myers AC, Undem BJ. Nerve growth factor-induced phenotypic switch in guinea pig airway sensory neurons. Am J Respir Crit Care Med. 2000;161:1985–90.
McMahon SB, Bennett DL, Priestley JV, Shelton DL. The biological effects of endogenous nerve growth factor on adult sensory neurons revealed by a trkA-IgG fusion molecule. Nat Med. 1995;1:774–80.
Bennett DL, Koltzenburg M, Priestley JV, Shelton DL, McMahon SB. Endogenous nerve growth factor regulates the sensitivity of nociceptors in the adult rat. Eur J Neurosci. 1998;10:1282–91.
Emanueli C, Salis MB, Pinna A, Graiani G, Manni L, Madeddu P. Nerve growth factor promotes angiogenesis and arteriogenesis in ischemic hindlimbs. Circulation. 2002;106:2257–62.
Cantarella G, Lempereur L, Presta M, Ribatti D, Lombardo G, Lazarovici P, Zappalà G, Pafumi C, et al. Nerve growth factor-endothelial cell interaction leads to angiogenesis in vitro and in vivo. FASEB J. 2002;16:1307–9.
Kraemer R, Nguyen H, March KL, Hempstead B. NGF activates similar intracellular signaling pathways in vascular smooth muscle cells as PDGF-BB but elicits different biological responses. Arterioscler Thromb Vasc Biol. 1999;19:1041–50.
Calza L, Giardino L, Giuliani A, Aloe L, Levi-Montalcini R. Nerve growth factor control of neuronal expression of angiogenic and vasoactive factors. Proc Natl Acad Sci U S A. 2001;98:4160–5.
Hull MA, Thomson JL, Hawkey CJ. Expression of cyclooxygenase 1 and 2 by human gastric endothelial cells. Gut. 1999;45:529–36.
Lambiase A, Bracci-Laudiero L, Bonini S, Starace G, D'Elios MM, De Carli M, De Carli M, Aloe L. Human CD4+ T cell clones produce and release nerve growth factor and express high-affinity nerve growth factor receptors. J Allergy Clin Immunol. 1997;100:408–14.
Nilsson G, Forsberg-Nilsson K, Xiang Z, Hallbook F, Nilsson K, Metcalfe DD. Human mast cells express functional TrkA and are a source of nerve growth factor. Eur J Immunol. 1997;27:2295–301.
Kobayashi H, Gleich GJ, Butterfield JH, Kita H. Human eosinophils produce neurotrophins and secrete nerve growth factor on immunologic stimuli. Blood. 2002;99:2214–20.
Barouch R, Kazimirsky G, Appel E, Brodie C. Nerve growth factor regulates TNF-alpha production in mouse macrophages via MAP kinase activation. J Leukoc Biol. 2001;69:1019–26.
Frossard N, Freund V, Advenier C. Nerve growth factor and its receptors in asthma and inflammation. Eur J Pharmacol. 2004;500:453–65.
Healy DL, Rogers PA, Hii L, Wingfield M. Angiogenesis: a new theory for endometriosis. Hum Reprod Update. 1998;4:736–40.
Kim SH, Choi YM, Chae HD, Kim KR, Kim CH, Kang BM. Increased expression of endoglin in the eutopic endometrium of women with endometriosis. Fertil Steril. 2001;76:918–22.
Singh M, Meyer EM, Simpkins JW. The effect of ovariectomy and estradiol replacement on brain-derived neurotrophic factor messenger ribonucleic acid expression in cortical and hippocampal brain regions of female Sprague–Dawley rats. Endocrinology. 1995;136:2320–4.
Simpkins JW, Green PS, Gridley KE, Singh M, de Fiebre NC, Rajakumar G. Role of estrogen replacement therapy in memory enhancement and the prevention of neuronal loss associated with Alzheimer's disease. Am J Med. 1997;103:19S–25S.
Sohrabji F, Miranda RC, Toran-Allerand CD. Identification of a putative estrogen response element in the gene encoding brain-derived neurotrophic factor. Proc Natl Acad Sci U S A. 1995;92:11110–4.
Gibbs RB. Treatment with estrogen and progesterone affects relative levels of brain-derived neurotrophic factor mRNA and protein in different regions of the adult rat brain. Brain Res. 1999;844:20–7.
Krizsan-Agbas D, Pedchenko T, Hasan W, Smith PG. Oestrogen regulates sympathetic neurite outgrowth by modulating brain derived neurotrophic factor synthesis and release by the rodent uterus. Eur J Neurosci. 2003;18:2760–8.
Singer CA, Figueroa-Masot XA, Batchelor RH, Dorsa DM. The mitogen-activated protein kinase pathway mediates estrogen neuroprotection after glutamate toxicity in primary cortical neurons. J Neurosci. 1999;19:2455–63.
Patrone C, Andersson S, Korhonen L, Lindholm D. Estrogen receptor-dependent regulation of sensory neuron survival in developing dorsal root ganglion. Proc Natl Acad Sci U S A. 1999;96:10905–10.
Wilson ME, Dubal DB, Wise PM. Estradiol protects against injury-induced cell death in cortical explant cultures: a role for estrogen receptors. Brain Res. 2000;873:235–42.
Bjorling DE, Beckman M, Clayton MK, Wang ZY. Modulation of nerve growth factor in peripheral organs by estrogen and progesterone. Neuroscience. 2002;110:155–67.
Li B, Sheng X, Song M, Zhang H, Weng J, Zhang M, Hu X, Zhou J, Xu M, Weng Q, Watanabe G, Taya K. Expression of nerve growth factor and its receptors TrkA and p75 in the uterus of wild female ground squirrel (Citellus dauricus Brandt). Gen Comp Endocrinol. 2012;1(176):62–9.
Shi Z, Arai KY, Jin W, Weng Q, Watanabe G, Suzuki AK, Taya K. Expression of nerve growth factor and its receptors NTRK1 and TNFRSF1B is regulated by estrogen and progesterone in the uteri of golden hamsters. Biol Reprod. 2006;74:850–6.
Pinto FM, Armesto CP, Magraner J, Trujillo M, Martin JD, Candenas ML. Tachykinin receptor and neutral endopeptidase gene expression in the rat uterus: characterization and regulation in response to ovarian steroid treatment. Endocrinology. 1999;140(6):2526–32.
Sumino H, Ichikawa S, Kanda T, Sakamaki T, Nakamura T, Sato K, Kobayashi I, Nagai R. Hormone replacement therapy in postmenopausal women with essential hypertension increases circulating plasma levels of bradykinin. Am J Hypertens. 1999;12(1O Pt 1):1044–7.
Papka RE, Storey-Workley M. Estrogen receptor-alpha and -beta coexist in a subpopulation of sensory neurons of female rat dorsal root ganglia. Neurosci Lett. 2002;319:71–4.
Schaible HG, Grubb BD. Afferent and spinal mechanisms of joint pain. Pain. 1993;55:5–54.
Toyomoto M, Ohta M, Okumura K, Yano H, Matsumoto K, Inoue S, Hayashi K, Ikeda K. Prostaglandins are powerful inducers of NGF and BDNF production in mouse astrocyte cultures. FEBS Lett. 2004;562:211–5.
Kanda N, Koike S, Watanabe S. Prostaglandin E2 enhances neurotrophin-4 production via EP3 receptor in human keratinocytes. J Pharmacol Exp Ther. 2005;315:796–804.
Ota H, Igarashi S, Sasaki M, Tanaka T. Distribution of cyclooxygenase-2 in eutopic and ectopic endometrium in endometriosis and adenomyosis. Hum Reprod. 2001;16:561–6.
Bartley J, Mechsner S, Beutler C, Halis G, Lange J, Ebert AD. COX-2-expression in extragenital endometriosis lesions as a novel therapeutical approach? Zentralbl Gynakol. 2003;125:252–5.
Fagotti A, Ferrandina G, Fanfani F, Legge F, Lauriola L, Gessi M, Castelli P, Barbieri F, et al. Analysis of cyclooxygenase-2 (COX-2) expression in different sites of endometriosis and correlation with clinico-pathological parameters. Hum Reprod. 2004;19:393–7.
Buchweitz O, Staebler A, Wulfing P, Hauzman E, Greb R, Kiesel L. COX-2 overexpression in peritoneal lesions is correlated with nonmenstrual chronic pelvic pain. Eur J Obstet Gynecol Reprod Biol. 2006;124:216–21.
Chang SY, Ho YS. Immunohistochemical analysis of insulin-like growth factor I, insulin-like growth factor I receptor and insulin-like growth factor II in endometriotic tissue and endometrium. Acta Obstet Gynecol Scand. 1997;76:112–7.
Kimpinski K, Mearow K. Neurite growth promotion by nerve growth factor and insulin-like growth factor-1 in cultured adult sensory neurons: role of phosphoinositide 3-kinase and mitogen activated protein kinase. J Neurosci Res. 2001;63:486–99.
Jones DM, Tucker BA, Rahimtula M, Mearow KM. The synergistic effects of NGF and IGF-1 on neurite growth in adult sensory neurons: convergence on the PI 3-kinase signaling pathway. J Neurochem. 2003;86:1116–28.
Ghahary A, Murphy LJ. Uterine insulin-like growth factor-I receptors: regulation by estrogen and variation throughout the estrous cycle. Endocrinology. 1989;125(2):597–604.
Wilson ME. Effects of estradiol and exogenous insulin-like growth factor I (IGF-I) on the IGF-I axis during growth hormone inhibition and antagonism. J Clin Endocrinol Metab. 1998;83:4013–21.
Watanabe H, Kanzaki H, Narukawa S, Inoue T, Katsuragawa H, Kaneko Y, Mori T. Bcl-2 and Fas expression in eutopic and ectopic human endometrium during the menstrual cycle in relation to endometrial cell apoptosis. Am J Obstet Gynecol. 1997;176:360–8.
Merry DE, Korsmeyer SJ. Bcl-2 gene family in the nervous system. Annu Rev Neurosci. 1997;20:245–67.
Hilton M, Middleton G, Davies AM. Bcl-2 influences axonal growth rate in embryonic sensory neurons. Curr Biol. 1997;7:798–800.
Fernandez AM. Gonzalez de la Vega AG, Planas B, Torres-Aleman I. Neuroprotective actions of peripherally administered insulin-like growth factor I in the injured olivo-cerebellar pathway. Eur J Neurosci. 1999;11:2019–30.
Cardona-Gomez GP, Mendez P, DonCarlos LL, Azcoitia I, Garcia-Segura LM. Interactions of estrogens and insulin-like growth factor-I in the brain: implications for neuroprotection. Brain Res Brain Res Rev. 2001;37:320–34.
Nilsen J, Brinton RD. Impact of progestins on estrogen-induced neuroprotection: synergy by progesterone and 19-norprogesterone and antagonism by medroxyprogesterone acetate. Endocrinology. 2002;143:205–12.
Singh M. Mechanisms of progesterone-induced neuroprotection. Ann N Y Acad Sci. 2005;1052:145–51.
Sugawara J, Fukaya T, Murakami T, Yoshida H, Yajima A. Increased secretion of hepatocyte growth factor by eutopic endometrial stromal cells in women with endometriosis. Fertil Steril. 1997;68:468–72.
Maina F, Hilton MC, Ponzetto C, Davies AM, Klein R. Met receptor signaling is required for sensory nerve development and HGF promotes axonal growth and survival of sensory neurons. Genes Dev. 1997;11:3341–50.
Khan KN, Masuzaki H, Fujishita A, Kitajima M, Sekine I, Matsuyama T, Ishimaru T. Estrogen and progesterone receptor expression in macrophages and regulation of hepatocyte growth factor by ovarian steroids in women with endometriosis. Hum Reprod. 2005;20:2004–13.
Ota H, Igarashi S, Hatazawa J, Tanaka T. Distribution of heat shock proteins in eutopic and ectopic endometrium in endometriosis and adenomyosis. Fertil Steril. 1997;68:23–8.
Plumier JC, Hopkins DA, Robertson HA, Currie RW. Constitutive expression of the 27-kDa heat shock protein (Hsp27) in sensory and motor neurons of the rat nervous system. J Comp Neurol. 1997;384:409–28.
Costigan M, Mannion RJ, Kendall G, Lewis SE, Campagna JA, Coggeshall RE, Meridith-Middleton J, Tate S, et al. Heat shock protein 27: developmental regulation and expression after peripheral nerve injury. J Neurosci. 1998;18:5891–900.
Lewis SE, Mannion RJ, White FA, Coggeshall RE, Beggs S, Costigan M, Martin JL, Dillmann WH, et al. A role for HSP27 in sensory neuron survival. J Neurosci. 1999;19:8945–53.
Tang PZ, Gannon MJ, Andrew A, Miller D. Evidence for oestrogenic regulation of heat shock protein expression in human endometrium and steroid-responsive cell lines. Eur J Endocrinol. 1995;133:598–605.
Wing LY, Chuang PC, Wu MH, Chen HM, Tsai SJ. Expression and mitogenic effect of fibroblast growth factor-9 in human endometriotic implant is regulated by aberrant production of estrogen. J Clin Endocrinol Metab. 2003;88:5547–54.
Kanda T, Iwasaki T, Nakamura S, Kurokawa T, Ikeda K, Mizusawa H. Self-secretion of fibroblast growth factor-9 supports basal forebrain cholinergic neurons in an autocrine/paracrine manner. Brain Res. 2000;876:22–30.
Novella-Maestre E, Herraiz S, Vila-Vives JM, Carda C, Ruiz-Sauri A, Pellicer A. Effect of antiangiogenic treatment on peritoneal endometriosis-associated nerve fibers. Fertil Steril. 2012;98:1209–17.
Ferrara N, Davis-Smyth T. The biology of vascular endothelial growth factor. Endocr Rev. 1997;18:4–25.
Machado DE, Abrao MS, Berardo PT, Takiya CM, Nasciutti LE. Vascular density and distribution of vascular endothelial growth factor (VEGF) and its receptor VEGFR-2 (Flk-1) are significantly higher in patients with deeply infiltrating endometriosis affecting the rectum. Fertil Steril. 2008;90:148–55.
Pupo-Nogueira A, de Oliveira RM, Petta CA, Podgaec S, Dias Jr JA, Abrao MS. Vascular endothelial growth factor concentrations in the serum and peritoneal fluid of women with endometriosis. Int J Gynaecol Obstet. 2007;99:33–7.
Sondell M, Sundler F, Kanje M. Vascular endothelial growth factor is a neurotrophic factor which stimulates axonal outgrowth through the flk-1 receptor. Eur J Neurosci. 2000;12:4243–54.
Sondell M, Lundborg G, Kanje M. Vascular endothelial growth factor has neurotrophic activity and stimulates axonal outgrowth, enhancing cell survival and Schwann cell proliferation in the peripheral nervous system. J Neurosci. 1999;19:5731–40.
Hobson MI, Green CJ, Terenghi G. VEGF enhances intraneural angiogenesis and improves nerve regeneration after axotomy. J Anat. 2000;197(Pt 4):591–605.
Barcz E, Rózewska ES, Kaminski P, Demkow U, Bobrowska K, Marianowski L. Angiogenic activity and IL-8 concentrations in peritoneal fluid and sera in endometriosis. Int J Gynaecol Obstet. 2002;79:229–35.
Khan KN, Masuzaki H, Fujishita A, Kitajima M, Hiraki K, Miura S, Sekine I, Ishimaru T. Peritoneal fluid and serum levels of hepatocyte growth factor may predict the activity of endometriosis. Acta Obstet Gynecol Scand. 2006;85:458–66.
Matsuzaki S, Murakami T, Uehara S, Yokomizo R, Noda T, Kimura Y, Okamura K. Erythropoietin concentrations are elevated in the peritoneal fluid of women with endometriosis. Hum Reprod. 2001;16:945–8.
Suzumori N, Zhao XX, Suzumori K. Elevated angiogenin levels in the peritoneal fluid of women with endometriosis correlate with the extent of the disorder. Fertil Steril. 2004;82:93–6.
Kats R, Collette T, Metz CN, Akoum A. Marked elevation of macrophage migration inhibitory factor in the peritoneal fluid of women with endometriosis. Fertil Steril. 2002;78:69–76.
Szamatowicz J, Laudański P, Tomaszewska I, Szamatowicz M. Chemokine growth-regulated-alpha: a possible role in the pathogenesis of endometriosis. Gynecol Endocrinol. 2002;16:137–41.
Maas JW, Groothuis PG, Dunselman GA, de Goeij AF, Struijker-Boudier HA, Evers JL. Development of endometriosis-like lesions after transplantation of human endometrial fragments onto the chick embryo chorioallantoic membrane. Hum Reprod. 2001;16:627–31.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Japan
About this chapter
Cite this chapter
Tokushige, N. (2014). Role of Nerve Fibres in Endometriosis. In: Harada, T. (eds) Endometriosis. Springer, Tokyo. https://doi.org/10.1007/978-4-431-54421-0_13
Download citation
DOI: https://doi.org/10.1007/978-4-431-54421-0_13
Published:
Publisher Name: Springer, Tokyo
Print ISBN: 978-4-431-54420-3
Online ISBN: 978-4-431-54421-0
eBook Packages: MedicineMedicine (R0)