
Abstract
Immunotoxicology is the study of undesired modulation of the immune system by extrinsic factors. Toxicological assessments have demonstrated that the immune system is a target following exposure to a diverse group of xenobiotics including ultraviolet radiation, chemical pollutants, therapeutics, and recreational drugs. There is a well-established cause and effect relationship between suppression of the immune response and reduced resistance to infections and certain types of neoplasia. In humans, mild-to-moderate suppression of the immune response is linked to reduced resistance to common community-acquired infections, whereas opportunistic infections, which are very rare in the general population, are common in individuals with severe suppression. Xenobiotic exposure may also result in unintended stimulation of immune function. Although a cause and effect relationship between unintended stimulation of the immune response and adverse consequences has yet to be established, evidence does suggest that hypersensitivity, autoimmunity, and pathological inflammation may be exacerbated in susceptible populations exposed to certain xenobiotics. Xenobiotics can act as allergens and elicit hypersensitivity responses, or they can modulate hypersensitivity responses to other allergens such as pollen or dust mite by acting as adjuvants, enhancing the development or expression of hypersensitivity. Allergic contact dermatitis, allergic rhinitis, and asthma are the most commonly encountered types of hypersensitivity reactions resulting from chemical exposure. The immunologic effectors and mechanisms involved in autoimmune reactions are the same as those associated with responses to foreign antigens; however, the reactions are directed against the host’s own cells. Thus, chemicals that induce immune suppression, nonspecific immunostimulation, or hypersensitivity may also impact autoimmunity. Risk assessment for immunotoxicity should be performed using the same approaches and principles for other noncancer effects. However, since xenobiotics may have effects on more than one aspect of immune function, immunotoxicity data should be evaluated separately for evidence of suppression, stimulation, hypersensitivity, and autoimmunity.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Allergic contact dermatitis
- Autoimmunity
- Benchmark dose
- Buehler test
- Guinea pig maximization test
- Hypersensitivity
- Human repeated insult patch test
- Immunomodulation
- Immunosuppression
- Immunostimulation
- Local lymph node assay
- Respiratory sensitization
- Skin sensitization
- TCDD
- Risk assessment
- Weight of evidence
Introduction
Immunotoxicology is the study of undesired modulation of the immune system by extrinsic factors, thus potentially increasing the risk of infectious or neoplastic diseases, allergy/asthma, or autoimmune diseases. Toxicological assessments have demonstrated that the immune system is a target organ following exposure to environmental agents such as ultraviolet radiation, chemical pollutants, therapeutics, and recreational drugs (Table 1). Immunotoxicity testing guidelines have been established to assess potential immunosuppression [1–3], allergic skin sensitization [4–6], and autoimmunity [7], although the most abundant and reliable data available to risk assessors pertain to suppression and hypersensitivity. After providing a brief overview of the immune system, this chapter is organized around sections that review the basic concepts of immunotoxicology for the four major types of chemical-related immune effects (suppression, enhancement, hypersensitivity, and autoimmunity) and their application to risk assessment.
The Immune System and Immune Function
The primary function of the immune system is to destroy or neutralize infectious agents and certain tumor cells. Immune cells are located throughout the body, either in distinct organs, including the spleen, thymus, and lymph nodes, or in diffuse accumulations of lymphoid and myeloid cells associated with the mucosal surfaces of the urogenital and GI tracts, skin, and lung. These tissues are the primary sites for the immune system to detect entering pathogens and exogenous proteins. Immune responses driven by proteins or carbohydrates (antigens) unique to a particular pathogen or cell are referred to as antigen-specific or adaptive, whereas responses to genetic material or structural components that are similar in a wide range of organisms (e.g., viral double-stranded RNA or components of bacterial cell walls) are referred to as nonspecific or innate.
Innate immune responses are able to provide a rapid defense against pathogenic organisms because cells and molecules that mediate innate immunity do not require antigen recognition or cell division/maturation. Effector cells include macrophages (phagocytosis and killing of pathogens, antigen processing, production of proinflammatory mediators), dendritic cells (antigen processing and presentation to lymphocytes, cytokine production), natural killer (NK) cells (killing of certain tumor cells and pathogens), and polymorphonuclear (PMN) leukocytes (phagocytosis and killing of bacteria). Soluble mediators participate in innate responses as well; components of the complement cascade lyse cells and augment phagocytosis of bacteria and various cytokines upregulate inflammation. However, cells and soluble mediators of the innate response also contribute to the development of adaptive immune responses via activation and regulation of other lymphocyte subpopulations.
The adaptive immune response entails recognition of foreign antigens presented to cell surface receptors on lymphocytes by innate antigen processors, followed by gene transcription, production of growth factors, clonal expansion of antigen-specific lymphocytes, activation of effector mechanisms that are ultimately directed against infectious agents or neoplastic cells, and the generation of long-lived memory cells. The response is target specific but develops slowly, peaking after 5–10 days. Lymphocytes are the principal cellular effectors of adaptive responses, classified first by their tissue of origin, and then by function or maturational state. Bone marrow-derived lymphocyte progenitors which mature in specific regions of the bone marrow, spleen, and lymph nodes are referred to as B lymphocytes or B cells. These B cells secrete antibodies, which are the soluble mediators responsible for humoral immunity. Cell-mediated immunity, on the other hand, is coordinated by the other major class of lymphocytes, T cells or T lymphocytes. T cells are derived from the common lymphocyte progenitor population that migrate to and mature in the thymus. T lymphocyte subpopulations include cells that assist in and amplify other immune responses (T helper cells; TH), downregulate other immune responses (T suppressor cells; TS), or destroy infected or neoplastic cells (cytotoxic T cells; TC). TH cells produce cytokines that regulate immune function and can be further subdivided into subpopulations which assist other T cells (TH1) or which stimulate and perpetuate antibody responses (TH2). TH2 cytokine production predominates in newborns, and the release of these cytokines is associated with increased allergy and asthma.
Antigen-naïve circulating B cells encounter antigen in lymph nodes or tissue-associated lymphoid tissues via membrane-bound immunoglobulin (Ig) molecules that act as antigen receptors. Receptor cross-linking initiates a signal transduction cascade that, with the appropriate stimulus from TH2 cytokines, leads to activation, clonal expansion and differentiation into antibody-secreting plasma cells, and a small population of long-lived memory cells. The five classes of antibodies, IgM, IgG, IgE, IgA, and IgD, are distinguished by the characteristics of their heavy chain polypeptides, and each class has distinct expression patterns and functional properties. IgM and IgG antibodies are frequently measured in studies of immune function for potential suppression, while increased total and antigen-specific IgE may indicate allergic hypersensitivity responses. Although IgA has a central role in mucosal immunity, neither IgA nor IgD is routinely evaluated.
Unintended Suppression or Enhancement of Immune Function
Suppression and stimulation of immune function are conceptually and biologically distinct, although when either occurs following xenobiotic exposure, the outcome is generally considered adverse. Unintended immunosuppression is universally recognized as adverse because immunosuppressed individuals are more susceptible to infections and certain types of malignancies. As a result, testing and regulatory strategies were designed to detect and characterize suppression of the immune response. In contrast, stimulated immune function is usually perceived as beneficial, as exemplified by the use of adjuvants to improve responses to vaccination. However, enhancement and suppression are not mutually exclusive events, particularly when the underlying mode of immunotoxicity is modulation of regulatory cytokines that drive either inflammatory or antibody responses to antigens. In this situation, the apparent outcome of hazard identification testing (change vs. no change) will depend on which immune functions are evaluated. Thus, failure to consider unintended stimulation of immune function as potentially adverse may result in disregarding data that indicate exposure-related effects with the potential to impact inflammation and disease.
Suppression
Studies dating from the 1970s conclusively established that exposure to diverse chemical classes adversely affects immunocompetence (Table 1) [1–3, 8, 9]. In humans, mild-to-moderate suppression reduces efficacy of vaccines and increases the likelihood of common community-acquired infections, but does not affect the incidence of opportunistic infections that typically accompany severe forms of immunodeficiency diseases [10, 11]. Maternal exposure to compounds such as polychlorinated biphenyls (PCBs) during the period of fetal immune system development may result in persistent immunosuppression in the offspring, at doses that do not affect adult immune function [1, 12].
Hazard Identification and Assessment of Immunosuppression
Clinical and Epidemiological Data
Functional assessment of the human immune system is technically simple and may be accomplished by monitoring the primary antibody response to vaccination or delayed-type hypersensitivity response to naturally occurring antigens in adults or infants [13, 14]. A description of biomarkers useful in the evaluation of immunity in epidemiological studies is provided in Report of the Bilthoven Symposium: Advancement of Epidemiological Studies in Assessing the Human Health Effects of Immunotoxic Agents in the Environment and the Workplace [15]. Although biomarkers are available, it is important to note that detection of mild-to-moderate immunodeficiency may be difficult in humans using common assays (e.g., leukocyte counts, immunoglobulin levels) because the range of “normal” is large. Likewise, identifying a small exposure-related change in infectious disease incidence is also difficult [16, 17]. Researchers have linked low-to-moderate degrees of immunosuppression and increased infectious disease incidences with chronic psychological factors including separation and divorce, caregiving for Alzheimer’s patients, and bereavement [11].
Chronic stress has been linked to reactivation of latent viruses, such as CMV, HSV-1, or EBV, as measured either by reoccurrence of symptoms or by elevations in specific antibody titer [18–24]. Elevated antiviral antibody titer signals viral reactivation and replication and precedes disease onset, although only about 20% of those with elevated titers actually develop clinical disease. The immunosuppressive effects of stress have been confirmed in a controlled infectious challenge study in humans [25].
Animal Data
Animal immunotoxicity data are generated as the result of regulatory mandate or by research conducted by government, academic, and commercial laboratories and may include relatively insensitive markers (lymphoid organ weights, blood cell counts), functional assessment (e.g., the response to immunization), or, rarely, assessment of resistance to infection or tumor challenge (i.e., host resistance assays). Host resistance is typically the standard against which other assays are judged because altered resistance is a biologically plausible effect with clear relevance for potential adverse effects in humans. However, challenge agents in host resistance assays should be chosen to explore or confirm a known functional defect, not used to screen for effects. Nonlethal resistance models, in which numbers of tumor foci, viral titers, or bacterial counts in target tissues are assessed, provide greater sensitivity than mortality because the data provide a quantitative assessment of the host response to challenge and are a better reflection of protective immunity in the organism. Furthermore, the biological relevance of death as an endpoint is questionable when most or all of the immunocompetent controls do not survive because the virulence or number of the challenge agents simply overwhelms the initial response to infection, killing the host before a protective response can be mounted.
If host resistance data are not available, potential effects on host resistance must be inferred from other immune endpoints. The predictive power of commonly used assays for altered host resistance assays varies although concordance rates may reach 100% when data from multiple assays are combined [16, 17]. Common endpoints and methods used to assess immunosuppression are described in the International Programme on Chemical Safety (IPCS) Environmental Health Criteria 180: Principles and Methods for assessing Direct Immunotoxicity with Exposure to Chemicals [26]. The document also provides context and information that may assist in the interpretation of immunosuppression data for risk assessment. The Risk Assessment section of this chapter includes additional discussion of important issues to be considered when evaluating immunotoxicity data.
Stimulation
Deliberate stimulation of the immune system is a routine and mostly beneficial clinical procedure. Chemical and/or biological adjuvants are routinely included in vaccines to increase and prolong the immune response and to improve the response to weak antigens; successful induction of protective immunity is often dependent on their activity. However, failure to control the intensity and duration of normally protective immune responses, as a consequence of inherited defects, an ongoing disease process, or chemical exposure, is a well-documented cause of immune-mediated tissue damage.
Adverse effects associated with stimulation of the immune system by a xenobiotic include inappropriate stimulation or skewing of normally protective immune responses, direct allergenicity of the xenobiotic, induction or worsening of autoimmune disease, and nonspecific inflammation. Inappropriate stimulation of responses to infectious agents that are normally protective may increase inflammation, resulting in excess tissue damage or the potential of exposing cryptic host antigens (self-antigens the host normally does not encounter), one possible pathway to autoimmune disease. Skewing of the immune response to favor either inflammation or allergy can also occur; some studies have linked such changes with an increased risk of developing allergies and reduced resistance to certain infectious agents. Hypersensitivity and autoimmune disease are covered elsewhere in this review. Inflammation is a normal component of tissue injury associated with toxicant exposure. This type of injury is generally considered in the context of specific organ systems and will not be discussed as part of this chapter. This section will examine the evidence that unintended stimulation of either the innate or adaptive immune responses should be considered an adverse effect and taken into account in a risk assessment.
Hazard Identification and Assessment of Immunostimulation
Regulatory acknowledgement of unintended immune system stimulation as an adverse effect is limited. However, the US Food and Drug Administration (FDA) Immunotoxicity Testing Guidance [27] and the FDA Guidance for Industry on Immunotoxicology Evaluation of Investigational New Drugs [28] both list unintentional immunostimulation as a category of immunotoxicity with potential adverse effects. The FDA [27] considers adverse immunostimulation to include unintentional immunogenicity and adjuvant activity and warns that “nonspecific enhancement of the immune response that might be interpreted as a beneficial effect may result in suppression of specific immunity against a particular infection.” The FDA also states that unintended stimulation may result in autoimmunity, hypersensitivity, and chronic inflammation.
Clinical and Epidemiological Data
Clinical data suggest that moderate stimulation of the immune system by adjuvant is not associated with immune-mediated disease in the general population, although adverse effects have been reported in a few individuals with preexisting autoimmune disease. For example, the use of “immunostimulating” herbal supplements has been temporally associated with flare-up of pemphigus vulgaris in two patients and with the onset and later flare-up of dermatomyositis in another [29]. Nevertheless, human studies suggest that routine vaccination against influenza and pneumonia is safe and effective in patients with various systemic autoimmune diseases, in spite of previously expressed concerns that immune system activation by components of the vaccine may activate or worsen systemic autoimmune disease [30–32].
In contrast, there are also several examples in which unintentional stimulation of human immune function by chemical exposure is known to cause adverse effects. Occupational exposure to silica stimulates innate immune system cells and is associated with development of human autoimmune disease [33]. Likewise, unintended upregulation of human antibody production by mercury is suggested by a study in which removal of dental amalgam from individuals with autoimmune thyroiditis and mercury hypersensitivity was determined to reduce autoantibodies to thyroglobulin and thyroid peroxidase [34].
Disruption of immune system balance by suppression of certain immune functions could theoretically lead to enhancement of others. In utero exposure to cigarette smoke has been strongly associated with increased risk of developing asthma [35, 36], yet animal data demonstrated that such exposures also increased susceptibility to tumor challenge via persistent suppression of cytotoxic T cell activity [37]. Exposure to PCBs was associated with less shortness of breath and wheeze [38], whereas immune suppression and increased infection were observed in similarly exposed populations [39, 40]. Given the immunomodulatory properties of some hormones, chemical disruption of endocrine system may also have adverse effects on immune function and balance in the immune system.
In aggregate, the literature suggests that inadvertent stimulation of the immune response may have adverse effects. In many cases, genotype appears to be a significant risk factor for the development of adverse effects, suggesting that sensitive individuals, rather than the general population, are at greater risk.
Animal Data
Immunotoxicity studies have consistently determined that exposure to various classes of xenobiotics and certain drugs is associated with increased or enhanced immune function, particularly the T cell-dependent antibody response (TDAR). In many cases, these same compounds were determined to hasten disease onset or enhance disease severity in genetically susceptible models. However, fewer reports describing the risk posed by unintended immunostimulation in nonsusceptible animals are available.
Unlike immune suppression, well-characterized methods to assess immunostimulation in rodent models are not widely available. However, a number of studies have shown that respiratory exposure to air pollutants (NO2, ozone, residual oil fly ash, and diesel exhaust) enhances both allergic sensitization to common allergens such as dust mite antigens and enhances respiratory responses to allergen challenge [41–43]. In addition, oxidant gases (e.g., ozone) have been reported to enhance immune pathology associated with influenza infection [44]. Other routes of exposure may produce similar effects; dioxin [45, 46] and ultraviolet radiation [47] decrease host resistance to influenza infection apparently by enhancing inflammation rather than suppressing immune defenses.
As noted above, the TDAR is widely used to detect suppression of immune function, and unexpected enhancement of the antibody response is also associated with adverse effects, particularly in susceptible subpopulations. For example, lead (Pb) exposure exacerbates autoimmune disease in a genetically prone mouse model of lupus, but does not induce disease in resistant strains of mice [48]. Rodgers et al. [49] reported that a single high but noncholinergic dose of the insecticide malathion increased the number of spleen cells of C57Bl/6 mice producing IgM antibody to sheep red blood cells, in the absence of changes in spleen weight or cellularity or cholinergic activation. A later study [50] determined that oral exposure to malathion (33–300 mg/kg/week, beginning at 6 weeks of age) accelerated the onset of autoimmune disease and increased autoantibody production in autoimmune-prone mice (MRL-lpr; a model of spontaneous systemic lupus erythematosus or SLE), but not in congenic-resistant mice (MRL +/+). Although high concordance between suppression of the IgM response and susceptibility to infectious disease has been demonstrated, it is unknown whether similar concordance exists between increased IgM responses and disease. The same is true for other functional endpoints (e.g., delayed-type hypersensitivity, NK cell activity).
Suppression and Enhancement Conclusions
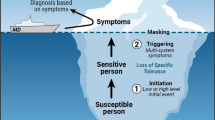
There is a well-established cause and effect relationship between suppression of the immune response and reduced resistance to infections and certain types of neoplasia. In humans, mild-to-moderate suppression of the immune response is linked to reduced resistance to common community-acquired infections, whereas opportunistic infections, which are very rare in the general population, are common in individuals with severe suppression. Quantitative functional data (e.g., responses to novel antigens) are generally predictive of resistance to infectious agents and tumor cells and provide insight into the potential consequences of suppression. Host resistance assays confirm that exposure increases the risk of infection, although models that rely on mortality as an endpoint may be misleading, if the dose or virulence of the challenge agent is not chosen with care. In contrast, a cause and effect relationship between unintended stimulation of the immune response, observed when testing for reduced function, and adverse consequences has yet to be established. Nevertheless, associative data suggest that hypersensitivity, autoimmunity, and pathological inflammation may be exacerbated in susceptible populations. As such, elevated responses in experimental groups of animals, immunized during tiered testing for immunotoxicity, should not be ignored, as it is a clear indication that modulation of the immune system has occurred. The final decision on how to interpret the data is a policy decision; the regulatory mandate, and the ability of the risk assessor to ask for additional testing, will determine how, or if, additional testing will be pursued when unintended stimulation is detected.
Hypersensitivity
Hypersensitivity (allergy) is defined as excessive humoral or cellular immune responses to an otherwise innocuous antigen, which can lead to tissue damage. Xenobiotics can pose a risk of allergic disease in one of two ways: they can act as allergens and elicit hypersensitivity responses, or they can modulate hypersensitivity responses to other allergens such as pollen or dust mite antigens by acting as adjuvants, enhancing the development or expression of hypersensitivity responses. This discussion will focus on chemical allergens. Chemicals that act as allergens include certain proteins that can by themselves induce an immune response and low molecular weight chemicals (known as haptens). Haptens are physically too small to induce a specific immune response but are chemically reactive and covalently bind to larger molecules, usually a protein, to form chemical-protein moieties of sufficient size to induce an immune response that is then hapten specific. For more details on the mechanisms underlying hypersensitivity reactions in general and allergic contact dermatitis (ACD) and respiratory allergies in particular, see Selgrade et al. [51].
ACD and hypersensitivity responses in the respiratory tract, including allergic rhinitis and asthma, are the most commonly encountered types of allergy resulting from occupational, consumer, and environmental exposures to chemicals [52]. In addition, certain drugs as well as certain food components are associated with systemic anaphylactic reactions [53]. Methods for hazard identification are very limited in the latter case; hence, this review will focus on ACD, for which there are widely accepted test guidelines [54], and hypersensitivity responses in the respiratory tract. Evidence for risk to the respiratory tract is largely based on epidemiology although experimental animal models have been used in research.
Hypersensitivity reactions to chemicals pose some particularly challenging problems for quantitative risk assessment because they develop in two separate stages: (1) induction and (2) elicitation. Induction (sensitization) is the first step and requires a sufficient single or cumulative exposure dose of the sensitizing agent to induce immune responses (i.e., development of antigen-specific antibodies or T cells). Obvious symptoms are generally not observed at this stage. Elicitation occurs in sensitized individuals upon subsequent exposure to the antigen and results in adverse responses that include inflammation. The dose responses for these two stages are different although not entirely independent [55–57]. Usually, the dose required for sensitization is higher than that required for elicitation. However, when the induction dose is high, the dose required to elicit a response is lower than that required when the induction dose is low. In practice, it is sometimes difficult to determine the point at which sensitization ends and elicitation begins. For this reason, risk assessments for hypersensitivity have been largely limited to hazard identification in which some chemicals are labeled as sensitizers, but without any indication of potency. However, significant progress has been made recently, particularly with ACD, in the development of dose–response relationships and thresholds. Various in vivo approaches to qualitative and quantitative risk assessment will be reviewed in this chapter. Because of animal welfare concerns and requirements imposed by the chemical legislation in Europe (e.g., 7th Amendment of the Cosmetics Directive and REACH), there is an increasing emphasis on the development of in vitro methods for hazard identification and potency characterization. This topic will be covered in another chapter of this book (see chapter Chemical Sensitization and Allergotoxicology, authored by Van Den Heuvel et al.).
Hazard Identification and Assessment of Skin Sensitization
The most familiar skin-sensitizing agent is poison ivy. However, ACD is one of the most commonly occurring occupational skin diseases [58], and fragrances, cosmetics, personal care products, reactive dyes, metals, preservatives, and pesticides all have the potential to cause skin sensitization. To insure consumer product and workplace safety, test guidelines have been developed and refined for the identification of chemicals that have the potential to induce hypersensitivity responses in the skin.
Induction
Clinical and Epidemiological Data
Experimental sensitization studies with human subjects have been used to provide dose–response information on the induction of ACD. These studies involve repeated application of the chemical to the skin under a patch, followed by a rest period and then a challenge exposure. Methods vary with respect to subject number, skin site, type and number of induction patches, patch application time, time between induction and challenge, and challenge duration. In all cases, enhancement of the skin response after challenge over that seen during early induction exposures has been the criterion by which induction of contact allergy is measured. The test most typically conducted is the human repeated insult patch test (HRIPT) [59]. A similar test, the human maximization test (HMT) found in earlier papers, differs from the HRIPT in that the patch application is to irritated skin. For this reason, HMT is not in common practice today although results from earlier studies may be available to risk assessors [60]. The readout for use in dermal sensitization risk assessments is usually incidence of sensitization. When HRIPTs are done using several different induction doses, a dose–response curve (induction incidence vs. dose expressed as amount of chemical applied per area of skin) and effect levels can be derived (i.e., the no-observed-effect-level or NOEL and the lowest-observed-effect-level or LOEL).
Today, for ethical reasons (i.e., the risk of sensitizing a previously naïve individual), skin sensitization hazard is usually assessed using an animal model (see below) such as the mouse local lymph node assay (LLNA), guinea pig maximization test, or guinea pig Buehler test. The HRIPT is sometimes employed as a confirmatory assay to substantiate the lack of sensitization at an exposure level which was identified as a NOEL in an animal model or which was derived as a likely NOEL from quantitative structure–activity relationships [61]. Epidemiological data documenting sensitization, usually from occupational cohorts, may also be available. The prevalence of acute contact dermatitis in a population exposed to a particular substance may be used in hazard identification and may also provide dose–response information, NOEL, LOEL, and benchmark dose (BMD), if adequate exposure data, expressed as skin area dose, are available.
Animal Data
Several testing strategies using laboratory animals are available to evaluate the potential for chemicals to induce contact sensitization. Historically, guinea pig models have been used. These methods rely on the induction of sensitization in animals and then the subjective evaluation of erythema and edema following test article challenge at a naïve site. Two of the most commonly used methods are the Buehler test [62] and the guinea pig maximization test [63]. In recent years, the mouse LLNA [64] has undergone extensive development and validation and was peer reviewed by the Interagency Coordinating Committee on the Validation of Alternative Methods [65, 66]. This assay evaluates only the induction phase of contact hypersensitivity and uses quantitation of 3H-thymidine incorporation into proliferating draining lymph node cells (clonal expansion) to evaluate the sensitization potential of chemicals. The LLNA has a number of advantages over the guinea pig assay including the use of a quantitative endpoint based on the relevant mode of action, acquisition of dose–response data, reduction in the number of animals used, lack of interference when testing colored materials, and a reduction in animal distress and the time required to conduct a study. By evaluating the dose response, this assay provides data that can be used to calculate an EC3 (the chemical exposure concentration required to induce a threefold increase in stimulation over control), which can then be used to rank the potency of the chemical.
A number of studies that compared human NOELs and BMDs with LLNA thresholds (EC3 values) indicate that area doses are directly comparable between mice and humans [67–69]. Therefore, the LLNA EC3 value has been suggested as a surrogate NOEL in quantitative risk assessment. In contrast, the traditional guinea pig tests are not as amenable to relative potency/quantitative assessments. Recently, guinea pig protocols have been modified in order to generate such data [70–72], but these protocols have not yet been validated. Hence, for de novo investigations, the LLNA is the animal test method best suited for quantitative estimations of skin-sensitizing potency [73].
Elicitation
Clinical and Epidemiological Data
The elicitation dose–response relationship for ACD can be determined in human studies using subjects that already have an established allergy. However, these data are likely to be rare because diagnostic tests frequently employ a single, relatively high concentration in a patch test [74] because the goal is to identify the sensitizer rather than determine the NOEL/LOEL concentration for elicitation. The concentration of the sensitizer (in a suitable vehicle such as petroleum jelly) can easily be varied in a patch test to determine a minimum elicitation threshold (MET), e.g., inducing a response in 10% of the subjects tested (MET10). Alternatively, the repeated open application test (ROAT) [75] can be employed in which formulations with different concentrations of the sensitizer, as well as a control formulation without the sensitizer, are employed and a NOEL or BMD determined. There is good correlation between the results from the patch test and the ROAT [76]. As noted earlier, the induction and elicitation dose responses are not entirely independent, and there is not a clear demarcation between induction and elicitation. Exposure can continue to contribute to induction even as elicited responses are occurring, and therefore, subsequent elicitations could occur at lower doses. Hence, it is best to determine elicitation thresholds in well-established rather than newly allergic subjects because it is assumed that induction reaches a maximum (levels off) at some point.
Animal Data
As discussed above for clinical endpoints, elicitation thresholds in newly sensitized animals depend on the frequency and dose used for sensitization. Elicitation NOELs are usually not determined in animal studies, because long-term exposure protocols that might address this issue have not been developed.
Hazard Identification and Assessment of Respiratory Sensitization
Although various types of immune-mediated injury can occur in the lung as a result of chemical exposure [52], those that cause rhinitis and asthma via IgE- (sometimes referred to as atopy) and TH2-cell-mediated responses are of particular concern. The incidence of atopic disease and asthma in the USA increased dramatically between 1980 and 1995 and remains a concern today [77], particularly in children. A similar trend also appeared in other westernized nations [78]. Also, work-related asthma is the most commonly reported occupational lung condition [79]. Allergic asthma can result from exposure to proteins (frequently enzymes) or from exposure to low molecular weight chemicals that, as with contact sensitizers, must be chemically linked with a protein carrier in order to sensitize the immune system (see Table 2 for example compounds).
Currently, there are no universally accepted animal models for human risk of respiratory sensitization that permit the determination of the dose–response relationship or relative potency of enzymes or low molecular weight chemicals for allergen-specific antibodies or symptoms of allergy via the inhalation route. Given the importance of IgE for many cases of respiratory sensitization, the generation of total IgE or antigen-specific IgE or cytophilic (usually predominantly IgE) serum antibodies is often used as a biomarker of disease to obtain threshold or benchmark value in both animal and human studies. Although the presence of IgE antibody does not equate to disease, it does increase the risk for development of allergy symptoms, but there are no absolute relationships between the levels of IgE antibody and symptoms. In the case of detergent enzymes, more intense exposures have been associated with symptoms, while less intense exposures have been associated with production of allergen-specific antibodies [83]. Both the occupational and guinea pig data indicate that there are thresholds for the induction of antibodies and for the elicitation of symptoms [84].
Whereas there is no doubt that IgE antibody plays an important role in respiratory allergy to proteins, there is some debate about the relevance of IgE antibody for the development of occupational asthma in response to chemical allergens. Although chemical respiratory allergens induce specific IgE in some symptomatic subjects, about half of subjects with allergy and asthma associated with diisocyanates do not exhibit this response and have late-onset (possibly IgG- or cell-mediated) responses in the absence of an immediate response [85–87]. Similarly, a small portion of subjects with asthma induced by acid anhydrides also exhibit late-onset responses (associated with IgG) in the absence of immediate responses [88, 89]. Hence, some portion of disease could be missed using IgE exclusively as the biomarker.
Induction
Clinical and Epidemiological Data
Dose–response data on the induction of respiratory sensitization may be obtained from epidemiologic studies that are either designed as a prospective study (with new workers, or when a new compound is introduced in the workplace or consumer market) or as a retrospective study (often involving experimental measurement of reconstituted exposure scenarios). There are several examples, both for protein and low molecular weight chemicals, where no observed effect concentration (NOEC) or benchmark concentrations have been derived (based on respiratory symptoms and/or cytophilic antibody responses assessed by skin prick or serum testing) (see Table 2) [80, 81, 90–92]. However, sufficient data for determination of the dose response for sensitization are available only for a very limited number of allergens.
Animal Data
Guinea pig and mouse models have been used to study both protein and low molecular weight chemical sensitizers. For protein allergens, such as detergent enzymes, animal dander, and biotechnology products, induction of cytophilic antibodies in experimental animals (IgE in mouse and rat and IgG1 in guinea pigs) following intratracheal or involuntary aspiration exposure has been used successfully for both hazard identification and to develop dose–response data. The best example comes from the detergent industry [84]. In these studies, guinea pigs and more recently mice have been used to establish the relative potency for respiratory sensitization of different enzymes based on the dose response. The endpoint measured was enzyme-specific, serum, cytophilic antibodies (IgE and IgG1). Subtilisin was chosen as the reference allergen because the American Conference of Government and Industrial Hygienists developed a threshold limit value in the workplace for subtilisin A of 60 ng protein/m3 based on historic human data, and the industry itself had an occupational exposure guideline of 15 ng protein/m3 that prevented induction of occupational asthma in their workforce. By comparing new enzymes to this reference allergen, it was possible to use animal serum antigen-specific cytophilic antibody levels to set safe exposure concentrations for humans [93].
Dose responses and thresholds for both immune and respiratory endpoints characteristic of asthma were also demonstrated in mice exposed by involuntary aspiration to mold extracts. Differences in potency between microbial extracts were demonstrated [82, 94, 95]. However, because these exposures included all the mold proteins extracted, it is not possible to draw conclusions about administered dose of specific allergens. In the case of low molecular weight compounds, dose–response data and no effect levels have been demonstrated for toluene diisocyanate (TDI) exposures using a guinea pig model and cytophilic antibody as the endpoint. Exposure to higher TDI concentrations resulted in both a greater percentage of animals producing antibodies and higher antibody titers. Pulmonary sensitivity, assessed by bronchial provocation challenge with TDI conjugated to protein, also demonstrated no effect at the same exposure level [96]. Although NOELs or benchmark concentrations can be derived from inhalation studies in experimental animal such as those described above, there are significant uncertainties that result from the lack of standardized guidelines. The length of the daily exposure, number of exposure days, and the challenge concentration and endpoints assessed are all variables that can influence results. Also, both the detergent matrix and components of the microbial extracts may have adjuvant effects that can facilitate induction [84, 97]. Thus, experimental exposures are best conducted using a matrix that closely mimics environmental exposures.
As the studies cited here indicate, it is commonly assumed that allergic sensitization of the respiratory tract results from inhalation exposure to the allergen. Recently, animal studies have suggested that sensitization, at least to low molecular weight chemicals, can occur following dermal exposure and that both immunologic and respiratory responses characteristic of asthma have been observed [98–100]. The possibility that dermal exposure might be an alternative route of sensitization or that more than one route of exposure could contribute to the sensitization complicates efforts to develop quantitative assessments.
Elicitation
Clinical and Epidemiological Data
Human data derived from epidemiological studies or experimental/diagnostic provocation tests can be used to develop dose–response relationships and thresholds for the elicitation of respiratory allergy. However, in practice, data are available for only a handful of low molecular weight chemical and protein exposures (Table 2). Occupational studies have reported thresholds for organic acid anhydrides and for isocyanates [80, 101]. For high molecular weight antigens, thresholds have been reported for wheat flour, natural rubber latex, western red cedar, and rat allergens [102]. For both proteins and low molecular weight chemicals, the prevalence of sensitized workers experiencing symptoms, the frequency of symptoms, and their severity (e.g., measured as force expiratory volume in 1 s — FEV1) were correlated with the mean exposure concentrations of antigen in air [80]. Although data are not available on the relationship between the induction and elicitation dose responses for respiratory sensitization, it is likely that as with skin sensitization the two are interdependent, and therefore, it would be best to determine elicitation thresholds in well-established rather than newly allergic subjects.
Animal Data
Because elicitation thresholds in newly sensitized animals likely depend on the frequency, dose, and route used for sensitization and because there are no standardized animal models for respiratory allergy, it is difficult to interpret elicitation dose responses in the few studies where they are described. Most data on elicitation after inhalation exposure obtained in animal models of respiratory sensitization were obtained after a single or a few induction exposures, rather than after long-established respiratory sensitization, and induction was often done by injection [103, 104] or dermal application [105–107] of the test substance. Also, in some cases, animals sensitized to low molecular weight chemicals were challenged with chemical-protein adducts, rather than free chemical.
Hypersensitivity Conclusions
In conclusion, data exist that can be used to assess the risk of inducing or triggering (eliciting) allergic responses following exposure to both protein and low molecular weight (hapten) chemicals. For ACD, standardized test methods and a large body of data have been developed. For respiratory sensitization, standardized tests do not exist, and the database is much smaller. In both cases, there are considerable uncertainties surrounding the relationship between induction and elicitation doses, the timing of induction and elicitation exposures (both duration and the interval between), the contribution of the exposure matrix to the response, and the role that genetic susceptibility plays (particularly in the case of respiratory responses to proteins).
Autoimmunity
Autoimmunity and autoimmune diseases result from immune responses against self-molecules. The immunologic effectors and mechanisms involved in autoimmune reactions are the same as those associated with responses to foreign antigens, including activation of the innate and adaptive immune systems, production of inflammatory mediators, and activation of T lymphocytes or the generation of antibodies. However, in the case of autoimmunity, the response is directed to self-antigens. Thus, chemicals that induce immune suppression, nonspecific immunostimulation, or hypersensitivity may also impact autoimmunity. In many instances, the events that initiate the immune response to self are unknown although specific gene polymorphisms, gender-related hormones, and exposures to certain therapeutic drugs, bacteria, and viruses have been shown to be associated with the induction, development, or exacerbation of autoimmunity. In some cases, the causal link between bacterial or viral infection and autoimmunity has been fairly well established. Many peptide fragments of microbial agents are homologous with host proteins, and the induction of an immune response to these antigens can result in cross-reactivity with self-antigens and the induction of autoimmunity. A number of chemicals and therapeutic agents have been identified as potential triggers for autoimmunity and have been suggested to both induce onset and modulate disease severity [7, 108]. While it is believed that a genetic predisposition to self-reactivity exists in all individuals with autoimmune diseases, differing susceptibility factors may govern the timing or specific disease an individual develops. As with other multifactorial diseases, such as cancer, it is suggested that both genetic and environmental factors interact to determine disease outcome and progression; however, we have little knowledge with regard to whether they result in cumulative and sequential changes or are the sequelae of mixtures of exposures.
Autoimmune disorders can affect virtually any site in the body and present as a spectrum of diseases ranging from organ specific, in which antibodies and T cells react to self-antigens localized in a specific tissue, to systemic, characterized by reactivity against a specific antigen or antigens present in various tissues. Organ-specific autoimmune diseases are typically characterized by cell-mediated immune responses directly affected by autoreactive CD8+ (cytotoxic) TC cells or indirectly via release of proinflammatory cytokines and other soluble mediators by activated CD4+ TH cells and macrophages. In contrast, systemic autoimmune diseases are generally characterized by specific autoantibodies, which can cause injury via activation of complement, blocking or stimulating cell surface receptors, or by aggregation into immune complexes that activate nonspecific inflammatory responses.
Hazard Identification and Assessment of Autoimmunity
Clinical and Epidemiological Data
Recent estimates suggest that 3–5% of the general population suffers from autoimmune diseases, and there is epidemiological evidence that the prevalence of certain autoimmune diseases is increasing in industrialized countries [7]. It has also been suggested that a number of common health problems, such as atherosclerosis, inflammatory bowel disease, and aspects of male and female infertility, may have an autoimmune component. The incidence of some autoimmune diseases has a clear gender bias, indicating a potential role for estrogens, androgens, and/or gonadotropins. Women have a significantly higher risk of developing an autoimmune disease than men, and in a majority of the most common autoimmune diseases (thyroiditis, scleroderma, systemic lupus erythematosus (SLE), multiple sclerosis (MS), rheumatoid arthritis (RA)), a female predominance is observed. However, for some autoimmune diseases, such as ankylosing spondylitis and adult-onset diabetes, there appears to be a higher risk among men. Lifestyle factors such as diet, smoking, recreational drug use, exposure to ultraviolet radiation, and environmental chemicals have all been implicated in the pathogenesis of autoimmune diseases [7, 109].
Familial aggregation and laboratory animal studies suggest a strong association between genetics and most autoimmune diseases. Genetic defects that lead to primary immunodeficiencies (PID) have identified critical steps in the process of establishing tolerance and immune regulation that are associated with clinical manifestations of autoimmunity [110, 111]. The two most common antibody deficiencies, selective IgA deficiency and common variable immunodeficiency, are associated with self-reactivity to a broad group of target tissues, and clinical manifestations of autoimmunity may appear in as many as 35% of individuals with the PID. The fact that not all individuals with a particular PID develop any or the same manifestations of autoimmunity is further support for the influence of environmental factors on the development and progression of these diseases. Susceptibility genes identified in PID have shown the importance of mutations in proteins associated with somatic recombination of T and B cell surface receptors, Fas-mediated T cell apoptosis, negative selection in the thymus, the development and activation of regulatory T cells, and the production of complement components [112]. Specific alleles within the major histocompatibility complex (MHC) gene region, functionally polymorphic genes encoding Fc and immunoinhibitory receptors such as CTL-A4, have been implicated as genetic factors in determining disease susceptibility, pathogenesis, and the course of many autoimmune diseases.
Occupational epidemiology studies often provide the best opportunity for identifying chemical-induced modulation of the immune system in human populations, as exposure levels tend to be higher than those found outside the workplace. Work-related exposures to compounds such as crystalline silica, heavy metals, and solvents have been associated with a number of systemic autoimmune diseases. Individuals with high-level exposures to silica-containing mineral dusts have been shown to demonstrate elevated risk for a number of autoimmune diseases, including RA, scleroderma ANCA-related vasculitis, and SLE. Exposures to tobacco smoke and iron particles have been shown to increase disease incidence and exacerbate the severity of symptoms in workers exposed to silica, stressing the need to identify potentially hazardous coexposures to accurately assess the risk for development of disease.
Vinyl chloride has been linked to the development of a scleroderma-like disease characterized by skin thickening, Raynaud’s phenomenon (discoloration of the extremities due to peripheral vasoconstriction), acroosteolysis (shortening of the terminal digital phalanges due to bone resorption), and pulmonary involvement. The linkage between vinyl chloride and autoimmunity stimulated research into associations between systemic autoimmune diseases and other solvents (e.g., trichloroethylene, trichloroethane, and xylenes), predominantly in occupational settings. An increased risk for systemic sclerosis was reported in several studies, but the risk is not consistent for all systemic autoimmune diseases [7].
As we better understand the consequences of immune dysregulation, there is increasing suspicion that early life exposures may lead to increased risk for autoimmune diseases later in life. A number of health concerns have been raised with regard to the children of women who received diethylstilbestrol (DES) during pregnancy to prevent preterm delivery or pregnancy loss. As part of the follow-up to a large multicenter epidemiologic study that examined the incidence of cancer and other diseases in DES-exposed and unexposed cohorts, Noller et al. [113] examined the self-reported prevalence of autoimmune diseases in 1,711 exposed women and 922 controls. The overall frequency of autoimmune diseases was significantly elevated in exposed women when compared with the control group (28.6 vs. 16.3 per thousand, p=0.02). A number of additional studies have suggested that the offspring of DES-treated women exhibit a variety of immune system perturbations, including enhanced T cell proliferation and elevated NK cell activity, that could contribute to immune dysregulation [114–116] and an elevated risk for autoimmune disease.
Miller et al. [117] have proposed a structured set of criteria to define environmentally associated autoimmune diseases in the human population. The five primary elements of these criteria are temporal plausibility; exclusion of other causative agents; dechallenge (resolution or improvement of the condition after removal of the agent); rechallenge (recurrence or worsening of the condition after reexposure to the agent); and biological plausibility. Identification of analogous cases, nearly identical cases, and evidence for a dose–response effect are also considered as supportive of a proposed association. The proposed tiered approach provides a framework upon which to assess the level of evidence for associations between exposures to exogenous agents and autoimmune diseases. For example, the development of eosinophilia–myalgia syndrome (EMS) and toxic oil syndrome (TOS), autoimmune disorders similar to diffuse fasciitis with eosinophilia and systemic sclerosis, has been associated with the ingestion of impure L-tryptophan containing dietary supplements and the consumption of contaminated rapeseed oil produced by a particular refinery [118, 119]. A number of studies suggested that the degree of illness correlated with the amount and frequency of intake [120–122], suggesting a potential dose–response relationship. These are two of the rare instances in which there is epidemiologic evidence for a temporal association between a specific environmental exposure and the onset of autoimmunity. However, there is often a long latency period between exposure and the development of disease, and for many compounds, the weight of evidence from human studies remains only suggestive.
Animal Data
Several investigators have examined immunologic effects in inbred and autoimmune prone mouse strains following prenatal or perinatal exposure [123]. The prototypical immunotoxicant 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) has been shown to induce thymic atrophy, alter thymocyte maturation and expression of MHC molecules, and increase the number of extrathymic autoreactive T cells [124–126], suggesting that the compound may promote autoimmunity. In utero exposure to TCDD has been reported to alter the time to disease onset in mice prone to autoimmune disease [127–129]. In C57Bl/6 mice, a strain not known for the development of autoimmune disease, gestational exposure to TCDD altered T cell populations in the spleen and thymus [130]. Increased immune complex and complement component 3 (C3) deposition in the glomeruli and elevated titers of autoantibodies indicate that developmental exposure to TCDD may have resulted in an increased risk for the development of autoimmunity in these mice [130].
Animal models of autoimmunity have been used to explore both molecular mechanisms and therapeutic interventions for a variety of autoimmune diseases [131]. However, there are currently no validated models to assess or identify chemicals that induce or exacerbate autoimmune diseases. The popliteal lymph node assay (PLNA), which measures nonspecific stimulation and proliferation in the lymph nodes draining chemically exposed tissues, has been shown to be a useful tool for screening for immunostimulating compounds [132]. In some situations, animal models of autoimmunity may utilize immunization with purified self-antigens, often in the presence of adjuvants, to elicit autoimmune responses. Rodents that are genetically predisposed to develop autoimmune disease, such as the lupus-prone MRL (MRL-lpr) mouse and the nonobese diabetic (NOD) mouse model for insulin-dependent diabetes, have been used to elucidate the role of specific genetic loci in the disease process. As our understanding of the genes that are associated with predisposition for specific autoimmune diseases in the human population has increased, animal models with similar genetic defects have been used to evaluate the potential for xenobiotics to modulate autoimmunity in genetically susceptible individuals. Consistent with what is believed to occur in humans, in some of these genetically prone models, autoimmunity is induced only following exposure to chemical or biological agents.
Autoimmunity Conclusions
Although it is believed that a genetic predisposition to self-reactivity exists in all individuals with autoimmune diseases, the lack of concordance between identical twins (9–30% depending on the disease) suggests that both genetic and environmental factors interact to determine the specific disease an individual develops, and disease outcome and progression. Limited evidence from occupational and epidemiologic studies suggests associations between chemical exposures and autoimmune diseases. However, only rarely have specific cause and effect relationships between chemical exposures and autoimmunity been established. Animal models to evaluate the impact of chemicals on induction and exacerbation of autoimmunity are largely disease specific, and the lack of widely accepted screening tools has hampered our ability to predict for these types of immune perturbations and potential impact on human health.
Risk Assessment
Risk assessment for immunotoxicity should be performed using the same approaches and principles for other noncancer effects. The basic method for performing human health risk assessment for environmental exposures is a well-defined process constituting problem formulation and four evaluation steps: hazard identification, hazard characterization (or dose–response assessment), exposure assessment, and risk characterization [133–135]. Problem formulation is a planning activity in which the risk assessor defines the goals, scope, and focus of the risk assessment including factors such as at-risk populations (workers, children, etc.), exposure routes, temporal scope options (acute, chronic, etc.), and endpoints evaluated (i.e., comprehensive or a targeted assessment like immunotoxicity). The first evaluation step for developing a risk assessment, hazard identification, is the process of collecting and analyzing the full range of human and animal data for a given compound and drawing conclusions on whether or not it causes adverse effects. Dose–response assessment is the second evaluation step, with the goal of characterizing the relationship between dose and the incidence of an adverse effect. The next step, exposure assessment, details the route, nature, and extent of human contact with the compound. Risk characterization is the final step in the risk assessment process in which the scientific evidence of adverse effects is integrated with the relevant exposure scenarios to provide a summary of the risk to human health including the assumptions and uncertainties involved in the evaluation.
Hazard Identification
Hazard identification from immunotoxicity data should result in weight of evidence (WOE) conclusions based on the available human and animal data, both positive and negative, for a given chemical. A chemical may have effects on more than one aspect of immunotoxicology, and therefore, the database should be evaluated for each type of immunotoxicity (suppression, stimulation, hypersensitivity, and autoimmunity) separately. Data from human exposures (e.g., occupational exposure studies) are preferred because fewer assumptions are required to determine the relative risk of immunotoxicity for the general population from human data than from animal data. The human data, however, are often incomplete and lack adequate exposure information or predictive immune measures. Therefore, animal data often serve as the basis for quantitative analysis to drive the risk assessment. The use of animal data for immunotoxicity risk assessment is supported by the general consistency between human evidence of immunotoxicity from clinical and epidemiological studies and the experimental animal data [2, 3, 9].
The WOE conclusions are developed on the basis of the human and experimental evidence, dose–response relationship, strength of the association, consistency of association, temporal association, coherence, specificity, analogy, and biological plausibility of the full dataset for immunotoxicity [136, 137]. Data are evaluated within the same or similar assays, as well as across divergent measures of the immune system and across multiple species. The WOE conclusions are strengthened by consistency (particularly across species, sex, or related endpoints), concordance, and breadth (range of effects) of the evidence for immunotoxicity.
Key Factors in Evaluating Immunotoxicity Across Studies
Factors such as stress, gender, species, and age may affect immunity and therefore may influence the results of immunotoxicity studies. The results themselves also must be evaluated in terms of biological significance of the observed effects. The following list discusses important considerations for evaluating data from each study for evidence of immunotoxicity.
Adversity or Biological Significance
To determine biological significance of animal data, one must consider the degree of change in a given measure of immunity that constitutes an adverse or biologically significant effect. The conservative approach is that any statistically significant effect should be considered meaningful with larger effects receiving greater weight, provided the quality of the animal data is sufficient. This approach is based on the assumption that a linear relationship exists between loss of immune responsiveness and increased risk of developing disease. A linear relationship is consistent with our understanding of immunological processes and is supported both by animal [17] and human studies [138] in which changes in immune tests correlated progressively with increased incidence of disease over a broad range.
Mode of Action
Information on the mode of action can be used to evaluate human relevance and to help predict the types of adverse effect that might be expected to occur as well as the persistence of effects. For example, alterations in B cell function, but not T cell function, would be expected to affect resistance to intracellular pathogens, but not viral infections. Alterations in stem cells would likely have long-term effects, as compared to alterations limited to secondary lymphoid organs such as the spleen or lymph node.
Stress
It is important to control for the influence of stress on the immune system in experimental design because of the strong immunomodulatory effects of stress. If immunotoxicity is observed at dose levels which do not induce overt toxicity, the test chemical can be considered immunotoxic independent of whether it occurs via a direct effect on the immune system or an indirect effect, such as induction of a stress response. In lieu of clear evidence of general toxicity, stress-induced immunotoxicity can be determined by testing in adrenalectomized animals.
Gender Considerations
Qualitative and quantitative gender-dependent differences in baseline levels of immune function are well known in humans and laboratory animals and in part have been linked to relative levels and response to sex steroids. If data are available for both sexes, the most sensitive gender can be used for the risk assessment. For chemicals with known effects on the endocrine system, data from both sexes are preferred.
Species and Strain Considerations
In cases where species or strain differences in immunotoxicity are demonstrated, data on toxicokinetics and the mode(s) of action can help select the best animal model for immunotoxicity in humans. In the absence of data to inform selection of the most appropriate animal model for a given chemical, the most sensitive species is utilized.
Age at Initial Exposure
The persistence of effects in adult animals has not been systematically evaluated; however, the expectation is that immune function returns to normal as immunotoxicants are cleared unless the chemical permanently damages sources of progenitor cells. On the other hand, developmental exposure has been shown to have lasting effects, persisting for weeks or months [1, 139]. Developmental immunotoxicity may be manifested qualitatively (i.e., affecting the developing immune system without affecting the adult) or quantitatively (i.e., affecting the developing immune system at lower doses). It is also suspected that the elderly may experience greater effects than young adults exposed to immunotoxic chemicals, although whether this is due to a general decline in immune responses or remodeling and dysregulation in specific cell populations remains to be determined [140].
Weighing the Evidence (WOE) to Draw Immunotoxicity Hazard Identification Conclusions
For each area of immunotoxicity (suppression, stimulation, hypersensitivity, or autoimmunity), the strength of the evidence varies by the methods used with some types of data presenting clear evidence of immunotoxicity, while the value of other data ranges from those that provide some support for immunotoxicity to equivocal.
Human Data
Epidemiological studies demonstrating an association between chemical exposure and disease burden are considered strong data, particularly if quantitative exposure information is available. Controlled clinical studies with quantitative evaluation of immune function can also provide clear evidence of immunotoxicity such as skin prick test data demonstrating hypersensitivity, autoantibodies indicating autoimmunity, or alterations in antibody responses to vaccination to support suppression or stimulation. In vivo functional assays to naturally occurring antigens are preferred because they test an intact immune system rather than a subset of isolated cells. However, they are not frequently performed, because in vivo testing procedures require injection of antigen into human subjects. The observational and in vitro assays most commonly used in human immunotoxicity studies (i.e., enumeration of immune system components and lymphocyte proliferation) are less invasive, but these endpoints are considered poor predictors of immunotoxicity. These less predictive immune measures can be used to support more definitive animal and human data, to determine biological plausibility, and to consider potential mechanisms when developing the WOE conclusions for immunotoxicity.
Animal Data to Evaluate Suppression and Stimulation
The strongest support for chemical-mediated immunosuppression from animal data is provided by data demonstrating suppression in a host resistance assay. Chemical-related reduction in a functional immune assay also represents clear evidence of immunosuppression. Observational assays (lymphocyte phenotyping, cytokines, complement, etc.), hematology, histopathology, and organ weight data may support biological plausibility or mode of action, but these data are not generally considered to be reliable predictors of immunosuppression. Immune enhancement may be detected in assays designed to test for immunosuppression, so the database should be evaluated for stimulation after evaluating the data for suppression. Although stimulation of endpoints such as increased antibody synthesis may not necessarily constitute sufficient evidence of adversity, it should raise concern that susceptible populations may be adversely affected.
Animal Data to Evaluate Hypersensitivity
Although animal data could be used to address the risk for induction or elicitation of hypersensitivity, in practice, preventing the induction of sensitization would preclude the need to evaluate elicitation. As a result, and due to the relationship between the induction and elicitation dose, elicitation thresholds are generally not determined from animal studies. The LLNA, guinea pig maximization test, and Buehler test all present strong evidence for induction of dermal hypersensitivity or skin sensitization from animal models. Increased antigen-specific cytophilic antibodies currently represent the best measure of induction of respiratory sensitization, although there are no universally accepted animal models for respiratory sensitization. Observational assays may be useful to support biological plausibility or mode of action.
Animal Data to Evaluate Autoimmunity
Chemical-related increases in disease incidence and/or disease progression in autoimmune prone animal models are considered strong evidence of increased risk of autoimmune disease. Increased levels of autoantibodies or positive results in lymph node proliferation assays also present some evidence of autoimmunity. Observational assays, hematology, histopathology, and organ weight data are equivocal evidence but can be used to support biological plausibility or mode of action of more predictive measures of autoimmunity.
WOE Conclusions
The WOE summary for immunotoxicity hazard identification should include conclusions on the evidence for suppression, enhancement, hypersensitivity, and autoimmunity. These conclusions should describe the database in terms of consistency and biological plausibility including strengths, weakness, uncertainties, and data gaps. Modulation of the same assay in multiple species or multiple functional assays that are biologically related increases the strength of the data indicating immunotoxicity. Just as positive data on a range of assays strengthen the WOE for immunotoxicity, negative data on a range of more predictive assay such as immune function data increase confidence to support lack of immunotoxicity.
Dose–Response
Dose–response assessments can be performed for each form of immunotoxicity (suppression, unintended stimulation, hypersensitivity, and autoimmunity) supported by the database for a given chemical or simply for the lowest effect level as this would be protective of other endpoints. If dose–response evaluations are performed for multiple endpoints, factors other than the lowest effect level (e.g., the steepness of the dose–response curve and the relative severity of health effects) can be considered at later stages of the risk assessment. A dose–response relationship is a necessary criterion in demonstrating chemical immunotoxicity. The dose–response functions for immunotoxicity are generally assumed to be nonlinear and to demonstrate a threshold dose below which effects on immunity would not be expected, consistent with these assumptions for other noncancer endpoints. In addition to identifying the shape of the dose–response curve and the effective dose range for immunotoxicity, factors such as exposure (route, timing, and duration), toxicokinetics, and other issues that might affect comparisons with human exposure scenarios are identified and discussed as part of the dose–response evaluation [141, 142].
The results from dose–response analyses can be used in various ways depending on the specific goals of the risk assessment: establishment of a health-based guidance value, estimation of the margin of exposure, or quantitative estimation of the magnitude of the risk at the level of human exposure. In each case, the critical effect(s) is selected, and data from these endpoints are used to calculate the guidance values. The critical effects for immunotoxicity data should generally represent the most sensitive adverse immune endpoint(s) from the most appropriate species. Health-based guidance values can be calculated from the critical effect by dividing the dose associated with no adverse effects (the no-observed-adverse-effect-level—NOAEL, or the mathematically determined benchmark dose—BMD) by a measure of the uncertainty in the assessment (the uncertainty factors or UFs) using approaches that are generally similar across agencies [142–144].
Guidance values for suppression, unintended stimulation, and autoimmunity are calculated using all of the standard UFs (intraspecies, interspecies, database) as well as subchronic to chronic and LOAEL (lowest-observed-adverse-effect-level) to NOAEL as determined by the scope and the data used for the critical effect. Risk assessment of autoimmunity from animal data illustrates issues presented for risk assessment based on animal data that model the most sensitive subpopulation of humans. Reduction in the intraspecies UF may be considered when human risk of autoimmunity is estimated from autoimmune prone animals when an identified genetic defect allows them to be considered specific models of susceptible humans rather than the general population. UFs applied to hypersensitivity also present a special case wherein the use of an additional matrix and time and use factor has been proposed when expected human exposure differs from the experimental situation [67].
Exposure Assessment
Exposure assessment is the process of comparing the exposure levels associated with reference values obtained in the dose–response assessment to known or estimated human exposure in order to help quantify the risk to a population. Specific guidelines on exposure assessment [145, 146] and guidance specific to assessing exposure of children [147] have been published separately and will not be discussed here. The importance of route of exposure and the adequacy of route-specific animal data on immunotoxicity for a given chemical deserve particular attention for exposure assessment and are discussed below.
It is generally accepted that the circulating concentration of immunotoxicant or immunotoxic metabolite(s) determines the level of immunotoxicity, and equal concentrations of immunotoxicant in the blood following different routes of exposure should result in similar levels of systemic immunotoxicity. However, to avoid complications of distinct local toxicity, animal data on immunotoxicity should match the most likely route of human exposure for a given chemical [26, 148]. Unlike most toxicological endpoints, for immunotoxicology, the route of exposure is not simply a local increased concentration or uptake issue. There are local, partially independent portions of the immune system associated with each of the major routes of exposure: inhalation, dermal, and oral. Therefore, route of exposure is potentially more important for immunotoxicity endpoints, and since local immunotoxicological effects may occur independent of systemic immunity, a separate evaluation of local immune responses may be appropriate. For example, chemical immunotoxicity may suppress immune function at the site of exposure, such as inhalation-dependent suppression of the resident macrophage populations of the lung, without affecting immune function of macrophages elsewhere in the body [149].
Risk Characterization
Risk characterization for immunotoxicity is a synthesis of estimates of exposure levels and health risks that should be developed as for other forms of toxicity. During the summary and integration portion of the risk assessment process, the hazard identification, quantitative dose–response assessment, and exposure assessment are combined along with a critical appraisal of the toxicity information. The critical evaluation contains a review of the overall quality of the assessment including a thorough appraisal of the strengths, weaknesses, and uncertainties in the assessment as well as an evaluation of confidence in the conclusions. Risk characterization also includes a section describing risk in terms of the nature and extent of harm. Additionally, to the extent permitted by available data, risk characterization indicates how risk varies with exposure and provides information to help risk managers evaluate a range of options.
Risk Assessment Conclusions
Human health risk assessments for immunotoxicity from chemical exposure are developed in four evaluation steps: hazard identification, hazard characterization (or dose–response assessment), exposure assessment, and risk characterization. The hazard identification conclusions are developed using a weight of evidence approach based on the available human and animal data for a given chemical. The database should be evaluated separately for each type of immunotoxicity (suppression, stimulation, hypersensitivity, and autoimmunity). Dose–response relationships for immunotoxicity are expected to display a threshold dose below which effects are not observed. The possibility of route-specific, local immune responses should be considered during hazard identification and exposure assessment. Risk assessments for immunotoxicity should result in a risk characterization that summarizes the evidence for immunotoxicity risk and describes the uncertainties in the evaluation.
Conclusions
Chemical modulation of the immune system may result in suppression, enhancement, hypersensitivity, or autoimmunity. Immunosuppression associated with chemical exposure is universally regarded as adverse because immunosuppressed individuals are at greater risk for infectious or neoplastic diseases. There are well-established testing guidelines to screen and evaluate xenobiotics for the potential to cause immunosuppression. The consequences of unintended stimulation of the immune system are less clear. Nevertheless, stimulation should be considered to be potentially adverse as it may exacerbate inflammation, hypersensitivity, or autoimmunity in susceptible populations. The most common hypersensitivity reactions to chemicals can be split into dermal effects (ACD) and respiratory effects (allergic rhinitis and asthma). There are well-established screening and testing guidelines for ACD; however, there are no standardized tests to evaluate potential respiratory sensitizers. As a result, assessment of respiratory sensitizers involves a greater reliance on human epidemiologic studies. For autoimmunity, there are a number of disease-specific animal models to evaluate xenobiotics for their potential to induce or exacerbate autoimmune diseases. However, as is the case for respiratory hypersensitivity, animal models are not available to more broadly screen chemicals for potential autoimmunity. Risk assessment for each type of immunotoxicity (suppression, stimulation, hypersensitivity, and autoimmunity) can be performed using the same general procedures as for other noncancer effects.
References
Luebke RW, Chen D, Dietert R, Yang Y, King M, Luster M (2006) The comparative immunotoxicity of five selected compounds following developmental or adult exposure. J Toxicol Environ Health B Crit Rev 9:1–26
Descotes J (2003) From clinical to human toxicology: linking animal research and risk assessment in man. Toxicol Lett 140–141:3–10
Vos JG, Van Loveren H (1998) Experimental studies on immunosuppression: how do they predict for man? Toxicology 129:13–26
van Loveren H, Cockshott A, Gebel T, Gundert-Remy U, de Jong WH, Matheson J, McGarry H, Musset L, Selgrade MK, Vickers C (2008) Skin sensitization in chemical risk assessment: report of a WHO/IPCS international workshop focusing on dose-response assessment. Regul Toxicol Pharmacol 50:155–199
Gerberick FG, Ryan CA, Dearman RJ, Kimber I (2007) Local lymph node assay (LLNA) for detection of sensitization capacity of chemicals. Methods 41:54–60
Basketter DA, Evans P, Fielder RJ, Gerberick GF, Dearman RJ, Kimber I (2002) Local lymph node assay―validation, conduct and use in practice. Food Chem Toxicol 40:593–598
WHO (2006) Principles and methods for assessing autoimmunity associated with exposure to chemicals, IPCS environmental health criteria 236. International programme on chemical safety. World Health Organization, Geneva, Switzerland
Selgrade MK (2007) Immunotoxicity: the risk is real. Toxicol Sci 100:328–332
Koller LD (2001) A perspective on the progression of immunotoxicology. Toxicology 160:105–110
Luebke RW, Parks C, Luster MI (2004) Suppression of immune function and susceptibility to infections in humans: association of immune function with clinical disease. J Immunotoxicol 1:15–24
Luster MI, Blanciforti LM, Germolec DR, Parks C, Kashon M, Luebke R (2004) Associating changes in the immune system with clinical diseases for interpretation in risk assessment. In: Maines M, Costa L, Reed D, Hodgson E (eds) Current protocols in toxicology. Wiley, New York, NY
Dietert RR (2008) Developmental immunotoxicology (DIT): windows of vulnerability, immune dysfunction and safety assessment. J Immunotoxicol 5:401–412
Van Loveren H, Van Amsterdam JG, Vandebriel RJ, Kimman TG, Rumke HC, Steerenberg PS, Vos JG (2001) Vaccine-induced antibody responses as parameters of the influence of endogenous and environmental factors. Environ Health Perspect 109:757–764
Gans H, DeHovitz R, Forghani B, Beeler J, Maldonado Y, Arvin AM (2003) Measles and mumps vaccination as a model to investigate the developing immune system: passive and active immunity during the first year of life. Vaccine 21:3398–3405
Van Loveren H, Germolec D, Koren H, Luster M, Nolan C, Repetto R, Smith E, Vos JG, Vogt R (1999) Report of the Bilthoven symposium: advancement of epidemiological studies in assessing the human health effects of immunotoxic agents in the environment and the workplace. Biomarkers 4:135–157
Luster MI, Portier C, Pait DG, White KL Jr, Gennings C, Munson AE, Rosenthal GJ (1992) Risk assessment in immunotoxicology. I. Sensitivity and predictability of immune tests. Fundam Appl Toxicol 18:200–210
Luster MI, Portier C, Pait DG, Rosenthal GJ, Germolec DR, Corsini E, Blaylock BL, Pollock P, Kouchi Y, Craig W, White KL, Munson AE, Comment CE (1993) Risk assessment in immunotoxicology. II. Relationships between immune and host resistance tests. Fundam Appl Toxicol 21:71–82
Yang EV, Glaser R (2000) Stress-induced immunomodulation: impact on immune defenses against infectious disease. Biomed Pharmacother 54:245–250
Glaser R, Pearson GR, Bonneau RH, Esterling BA, Atkinson C, Kiecolt-Glaser JK (1993) Stress and the memory T-cell response to the Epstein–Barr virus in healthy medical students. Health Psychol 12:435–442
Glaser R, Rice J, Sheridan J, Fertel R, Stout J, Speicher C, Pinsky D, Kotur M, Post A, Beck M (1987) Stress-related immune suppression: health implications. Brain Behav Immun 1:7–20
Esterling BA, Antoni MH, Kumar M, Schneiderman N (1993) Defensiveness, trait anxiety, and Epstein–Barr viral capsid antigen antibody titers in healthy college students. Health Psychol 12:132–139
Cohen S (1995) Psychological stress and susceptibility to upper respiratory infections. Am J Respir Crit Care Med 152:S53–S58
Biondi M, Zannino LG (1997) Psychological stress, neuroimmunomodulation, and susceptibility to infectious diseases in animals and man: a review. Psychother Psychosom 66:3–26
Kasl SV, Evans AS, Niederman JC (1979) Psychosocial risk factors in the developmental of infectious mononucleosis. Psychosom Med 41:445–466
Cohen S, Tyrrell DA, Smith AP (1991) Psychological stress and susceptibility to the common cold. N Engl J Med 325:606–612
WHO (1996) Principles and methods for assessing direct immunotoxicity associated with exposure to chemicals, IPCS environmental health criteria 180. International programme on chemical safety. World Health Organization, Geneva, Switzerland
U.S. FDA (1999) Guidance for industry: immunotoxicity testing guidance. Report. Center for Devices and Radiological Health, Rockville, MD
U.S. FDA (2002) Guidance for industry: immunotoxicity evaluation of investigational new drugs. Report. Center for Drug Evaluation and Research, Rockville, MD
Lee AN, Werth VP (2004) Activation of autoimmunity following use of immunostimulatory herbal supplements. Arch Dermatol 140:723–727
Holvast B, Huckriede A, Kallenberg CG, Bijl M (2007) Influenza vaccination in systemic lupus erythematosus: safe and protective? Autoimmun Rev 6:300–305
Elkayam O, Ablin J, Caspi D (2007) Safety and efficacy of vaccination against Streptococcus pneumonia in patients with rheumatic diseases. Autoimmun Rev 6:312–314
Gluck T, Muller-Ladner U (2008) Vaccination in patients with chronic rheumatic or autoimmune diseases. Clin Infect Dis 46:1459–1465
Parks CG, Conrad K, Cooper GS (1999) Occupational exposure to crystalline silica and autoimmune disease. Environ Health Perspect 107(Suppl 5):793–802
Sterzl I, Prochazkova J, Hrda P, Matucha P, Bartova J, Stejskal V (2006) Removal of dental amalgam decreases anti-TPO and anti-Tg autoantibodies in patients with autoimmune thyroiditis. Neuro Endocrinol Lett 27(Suppl 1):25–30
Gold DR, Burge HA, Carey V, Milton DK, Platts-Mills T, Weiss ST (1999) Predictors of repeated wheeze in the first year of life: the relative roles of cockroach, birth weight, acute lower respiratory illness, and maternal smoking. Am J Respir Crit Care Med 160:227–236
Jaakkola JJ, Gissler M (2004) Maternal smoking in pregnancy, fetal development, and childhood asthma. Am J Public Health 94:136–140
Ng SP, Silverstone AE, Lai ZW, Zelikoff JT (2006) Effects of prenatal exposure to cigarette smoke on offspring tumor susceptibility and associated immune mechanisms. Toxicol Sci 89:135–144
Weisglas-Kuperus N, Patandin S, Berbers GA, Sas TC, Mulder PG, Sauer PJ, Hooijkaas H (2000) Immunologic effects of background exposure to polychlorinated biphenyls and dioxins in Dutch preschool children. Environ Health Perspect 108:1203–1207
Heilmann C, Grandjean P, Weihe P, Nielsen F, Budtz-Jorgensen E (2006) Reduced antibody responses to vaccinations in children exposed to polychlorinated biphenyls. PLoS Med 3:1352–1359
Dallaire F, Dewailly E, Vezina C, Muckle G, Weber JP, Bruneau S, Ayotte P (2006) Effect of prenatal exposure to polychlorinated biphenyls on incidence of acute respiratory infections in preschool Inuit children. Environ Health Perspect 114:1301–1305
Gilmour MI, Selgrade MJK, Lambert AL (2000) Enhanced allergic sensitization in animals exposed to particulate air pollution. Inhal Toxicol 12(Suppl 3):373–380
Steerenberg PA, Withagen CE, van Dalen WJ, Dormans JA, Heisterkamp SH, van Loveren H, Cassee FR (2005) Dose dependency of adjuvant activity of particulate matter from five European sites in three seasons in an ovalbumin-mouse model. Inhal Toxicol 17:133–145
Gilmour MI, Jaakkola MS, London SJ, Nel AE, Rogers CA (2006) How exposure to environmental tobacco smoke, outdoor air pollutants, and increased pollen burdens influences the incidence of asthma. Environ Health Perspect 114:627–633
Selgrade MK, Illing JW, Starnes DM, Stead AG, Menache MG, Stevens MA (1988) Evaluation of effects of ozone exposure on influenza infection in mice using several indicators of susceptibility. Fundam Appl Toxicol 11:169–180
Teske S, Bohn AA, Hogaboam JP, Lawrence BP (2008) Aryl hydrocarbon receptor targets pathways extrinsic to bone marrow cells to enhance neutrophil recruitment during influenza virus infection. Toxicol Sci 102:89–99
Luebke RW, Copeland CB, Bishop LR, Daniels MJ, Gilmour MI (2002) Mortality in dioxin-exposed mice infected with influenza: mitochondrial toxicity (reye’s-like syndrome) versus enhanced inflammation as the mode of action. Toxicol Sci 69:109–116
Ryan LK, Copeland LR, Daniels MJ, Costa ER, Selgrade MJ (2002) Proinflammatory and Th1 cytokine alterations following ultraviolet radiation enhancement of disease due to influenza infection in mice. Toxicol Sci 67:88–97
Hudson CA, Cao L, Kasten-Jolly J, Kirkwood JN, Lawrence DA (2003) Susceptibility of lupus-prone NZM mouse strains to lead exacerbation of systemic lupus erythematosus symptoms. J Toxicol Environ Health A 66:895–918
Rodgers KE, Imamura T, Devens BH (1986) Organophosphorus pesticide immunotoxicity: effects of O,O,S-trimethyl phosphorothioate on cellular and humoral immune response systems. Immunopharmacology 12:193–202
Rodgers KE (1997) Effects of oral administration of malathion on the course of disease in MRL-lpr mice. J Autoimmun 10:367–373
Selgrade MJK, Germolec DR, Luebke RW, Smialowicz RJ, Ward MD, Bowman CB (2008) Immunotoxicology. In: Smart RC, Hodgson E (eds) Molecular and biochemical toxicology. Wiley, Hoboken, NJ, pp 765–803
Kirchner DB (2002) The spectrum of allergic disease in the chemical industry. Int Arch Occup Environ Health 75(Suppl):S107–112
Treudler R, Kozovska Y, Simon JC (2008) Severe immediate type hypersensitivity reactions in 105 German adults: when to diagnose anaphylaxis. J Investig Allergol Clin Immunol 18:52–58
WHO (1999) Principles and methods for assessing allergic hypersensitization associated with exposure to chemicals, IPCS environmental health criteria 212. International programme on chemical safety. World Health Organization, Geneva, Switzerland
Friedmann PS, Moss C, Shuster S, Simpson JM (1983) Quantitative relationships between sensitizing dose of DNCB and reactivity in normal subjects. Clin Exp Immunol 53:709–715
Scott AE, Kashon ML, Yucesoy B, Luster MI, Tinkle SS (2002) Insights into the quantitative relationship between sensitization and challenge for allergic contact dermatitis reactions. Toxicol Appl Pharmacol 183:66–70
Friedmann PS (2007) The relationships between exposure dose and response in induction and elicitation of contact hypersensitivity in humans. Br J Dermatol 157:1093–1102
Andersen KE (2003) Occupational issues of allergic contact dermatitis. Int Arch Occup Environ Health 76:347–350
McNamee PM, Api AM, Basketter DA, Frank Gerberick G, Gilpin DA, Hall BM, Jowsey I, Robinson MK (2008) A review of critical factors in the conduct and interpretation of the human repeat insult patch test. Regul Toxicol Pharmacol 52:24–34
Kligman AM (1966) The identification of contact allergens by human assay. 3. The maximization test: a procedure for screening and rating contact sensitizers. J Invest Dermatol 47:393–409
Politano VT, Api AM (2008) The research institute for fragrance materials’ human repeated insult patch test protocol. Regul Toxicol Pharmacol 52:35–38
Buehler EV (1965) Delayed contact hypersensitivity in the guinea pig. Arch Dermatol 91:171–177
Magnusson B, Kligman AM (1969) The identification of contact allergens by animal assay. The guinea pig maximization test. J Invest Dermatol 52:268–276
Kimber I, Dearman RJ, Basketter DA, Ryan CA, Gerberick GF (2002) The local lymph node assay: past, present and future. Contact Dermatits 47:315–328
Dean JH, Twerdok LE, Tice RR, Sailstad DM, Hattan DG, Stokes WS (2001) ICCVAM evaluation of the murine local lymph node assay. Conclusions and recommendations of an independent scientific peer review panel. Regul Toxicol Pharmacol 34:258–273
Api AM, Basketter DA, Cadby PA, Cano MF, Ellis G, Gerberick GF, Griem P, McNamee PM, Ryan CA, Safford R (2008) Dermal sensitization quantitative risk assessment (QRA) for fragrance ingredients. Regul Toxicol Pharmacol 52:3–23
Griem P, Goebel C, Scheffler H (2003) Proposal for a risk assessment methodology for skin sensitization based on sensitization potency data. Regul Toxicol Pharmacol 38:269–290
Schneider K, Akkan Z (2004) Quantitative relationship between the local lymph node assay and human skin sensitization assays. Regul Toxicol Pharmacol 39:245–255
Anderson C, Hehr A, Robbins R, Hasan R, Athar M, Mukhtar H, Elmets CA (1995) Metabolic requirements for induction of contact hypersensitivity to immunotoxic polyaromatic hydrocarbons. J Immunol 155:3530–3537
van Och FM, Vandebriel RJ, Prinsen MK, De Jong WH, Slob W, van Loveren H (2001) Comparison of dose-responses of contact allergens using the guinea pig maximization test and the local lymph node assay. Toxicology 167:207–215
Yamano T, Shimizu M, Noda T (2005) Quantitative comparison of the results obtained by the multiple-dose guinea pig maximization test and the non-radioactive murine local lymph-node assay for various biocides. Toxicology 211:165–175
Kimber I, Basketter DA, Berthold K, Butler M, Garrigue JL, Lea L, Newsome C, Roggeband R, Steiling W, Stropp G, Waterman S, Wiemann C (2001) Skin sensitization testing in potency and risk assessment. Toxicol Sci 59:198–208
Devos SA, Van Der Valk PG (2002) Epicutaneous patch testing. Eur J Dermatol 12:506–513
Nakada T, Hostynek JJ, Maibach HI (2000) Use tests: ROAT (repeated open application test)/PUT (provocative use test): an overview. Contact Dermatits 43:1–3
Fischer LA, Voelund A, Andersen KE, Menne T, Johansen JD (2009) The dose-response relationship between the patch test and ROAT and the potential use for regulatory purposes. Contact Dermatits 61:201–208
Moorman JE, Rudd RA, Johnson CA, King M, Minor P, Bailey C, Scalia MR, Akinbami LJ (2007) National surveillance for asthma―United States, 1980-2004. MMWR Surveill Summ 56:1–54
Sennhauser FH, Braun-Fahrlander C, Wildhaber JH (2005) The burden of asthma in children: a European perspective. Paediatr Respir Rev 6:2–7
Petsonk EL (2002) Work-related asthma and implications for the general public. Environ Health Perspect 110(Suppl 4):569–572
Arts JH, Mommers C, de Heer C (2006) Dose-response relationships and threshold levels in skin and respiratory allergy. Crit Rev Toxicol 36:219–251
Merget R, Kulzer R, Dierkes-Globisch A, Breitstadt R, Gebler A, Kniffka A, Artelt S, Koenig HP, Alt F, Vormberg R, Baur X, Schultze-Werninghaus G (2000) Exposure-effect relationship of platinum salt allergy in a catalyst production plant: conclusions from a 5-year prospective cohort study. J Allergy Clin Immunol 105:364–370
Chung YJ, Copeland LB, Doerfler DL, Ward MD (2010) The relative allergenicity of Stachybotrys chartarum compared to house dust mite extracts in a mouse model. Inhal Toxicol 22:460–468
Sarlo K, Kirchner DB (2002) Occupational asthma and allergy in the detergent industry: new developments. Curr Opin Allergy Clin Immunol 2:97–101
Schweigert MK, Mackenzie DP, Sarlo K (2000) Occupational asthma and allergy associated with the use of enzymes in the detergent industry―a review of the epidemiology, toxicology and methods of prevention. Clin Exp Allergy 30:1511–1518
Bernstein DI, Cartier A, Cote J, Malo JL, Boulet LP, Wanner M, Milot J, L’Archeveque J, Trudeau C, Lummus Z (2002) Diisocyanate antigen-stimulated monocyte chemoattractant protein-1 synthesis has greater test efficiency than specific antibodies for identification of diisocyanate asthma. Am J Respir Crit Care Med 166:445–450
Bernstein JA (1996) Overview of diisocyanate occupational asthma. Toxicology 111:181–189
Cartier A, Grammer L, Malo JL, Lagier F, Ghezzo H, Harris K, Patterson R (1989) Specific serum antibodies against isocyanates: association with occupational asthma. J Allergy Clin Immunol 84:507–514
Grammer LC, Shaughnessy MA, Henderson J, Zeiss CR, Kavich DE, Collins MJ, Pecis KM, Kenamore BD (1993) A clinical and immunologic study of workers with trimellitic-anhydride-induced immunologic lung disease after transfer to low exposure jobs. Am Rev Respir Dis 148:54–57
Grammer L, Shaughnessy M, Kenamore B (1998) Utility of antibody in identifying individuals who have or will develop anhydride-induced respiratory disease. Chest 114:1199–1202
Baur X (2003) Are we closer to developing threshold limit values for allergens in the workplace? Ann Allergy Asthma Immunol 90:11–18
Mapp CE, Boschetto P, Maestrelli P, Fabbri LM (2005) Occupational asthma. Am J Respir Crit Care Med 172:280–305
Heederik D, Houba R (2001) An exploratory quantitative risk assessment for high molecular weight sensitizers: wheat flour. Ann Occup Hyg 45:175–185
Sarlo K (1994) Human health risk assessment: focus on enzymes. In: Cahn A (ed) Proceedings of the 3rd World conference on detergents. American Oil Chemists Society Press, Chicago, IL, pp 54–57
Chung YJ, Coates NH, Viana ME, Copeland L, Vesper SJ, Selgrade MK, Ward MD (2005) Dose-dependent allergic responses to an extract of Penicillium chrysogenum in BALB/c mice. Toxicology 209:77–89
Ward MD, Chung YJ, Copeland LB, Doerfler DL (2010) A comparison of the allergic responses induced by Penicillium chrysogenum and house dust mite extracts in a mouse model. Indoor Air 20:380–391
Karol MH (1983) Concentration-dependent immunologic response to toluene diisocyanate (TDI) following inhalation exposure. Toxicol Appl Pharmacol 68:229–241
Instanes C, Ward MD, Hetland G (2006) The fungal biopesticide Metarhizium anisopliae has an adjuvant effect on the allergic response to ovalbumin in mice. Toxicol Lett 161:219–225
Farraj AK, Boykin E, Haykal-Coates N, Gavett SH, Doerfler D, Selgrade M (2007) Th2 cytokines in skin draining lymph nodes and serum IgE do not predict airway hypersensitivity to intranasal isocyanate exposure in mice. Toxicol Sci 100:99–108
Klink KJ, Meade BJ (2003) Dermal exposure to 3-amino-5-mercapto-1,2,4-triazole (AMT) induces sensitization and airway hyperreactivity in BALB/c mice. Toxicol Sci 75:89–98
Sailstad DM, Ward MD, Boykin EH, Selgrade MK (2003) A murine model for low molecular weight chemicals: differentiation of respiratory sensitizers (TMA) from contact sensitizers (DNFB). Toxicology 194:147–161
Yokota K, Johyama Y, Yamaguchi K, Takeshita T, Morimoto K (1999) Exposure-response relationships in rhinitis and conjunctivitis caused by methyltetrahydrophthalic anhydride. Int Arch Occup Environ Health 72:14–18
Baur X, Chen Z, Liebers V (1998) Exposure-response relationships of occupational inhalative allergens. Clin Exp Allergy 28:537–544
Pauluhn J, Mohr U (1994) Assessment of respiratory hypersensitivity in guinea-pigs sensitized to diphenylmethane-4,4′-diisocyanate (MDI) and challenged with MDI, acetylcholine or MDI-albumin conjugate. Toxicology 92:53–74
Botham PA, Rattray NJ, Woodcock DR, Walsh ST, Hext PM (1989) The induction of respiratory allergy in guinea-pigs following intradermal injection of trimellitic anhydride: a comparison with the response to 2,4-dinitrochlorobenzene. Toxicol Lett 47:25–39
Arts JH, Kuper CF, Spoor SM, Bloksma N (1998) Airway morphology and function of rats following dermal sensitization and respiratory challenge with low molecular weight chemicals. Toxicol Appl Pharmacol 152:66–76
Zhang XD, Fedan JS, Lewis DM, Siegel PD (2004) Asthmalike biphasic airway responses in Brown Norway rats sensitized by dermal exposure to dry trimellitic anhydride powder. J Allergy Clin Immunol 113:320–326
Arts JH, Muijser H, Appel MJ, Frieke Kuper C, Bessems JG, Woutersen RA (2004) Subacute (28-day) toxicity of furfural in Fischer 344 rats: a comparison of the oral and inhalation route. Food Chem Toxicol 42:1389–1399
Pollard KM, Hultman P, Kono DH (2010) Toxicology of autoimmune diseases. Chem Res Toxicol 23:455–466
Heindel J, Cooper GS, Germolec D, Selgrade M (1999) Linking environmental agents to autoimmune diseases. Environ Health Perspect 107(Suppl 5):659–813
Carneiro-Sampaio M, Coutinho A (2007) Tolerance and autoimmunity: lessons at the bedside of primary immunodeficiencies. Adv Immunol 95:51–82
Torgerson TR (2008) Immune dysregulation in primary immunodeficiency disorders. Immunol Allergy Clin North Am 28:315–327
Rose NR, Mackay IR (2006) The autoimmune diseases. Elsevier, St. Louis, MO
Noller KL, Blair PB, O’Brien PC, Melton LJ 3rd, Offord JR, Kaufman RH, Colton T (1988) Increased occurrence of autoimmune disease among women exposed in utero to diethylstilbestrol. Fertil Steril 49:1080–1082
Ford CD, Johnson GH, Smith WG (1983) Natural killer cells in in utero diethylstilbesterol-exposed patients. Gynecol Oncol 16:400–404
Ways SC, Mortola JF, Zvaifler NJ, Weiss RJ, Yen SS (1987) Alterations in immune responsiveness in women exposed to diethylstilbestrol in utero. Fertil Steril 48:193–197
Burke L, Segall-Blank M, Lorenzo C, Dynesius-Trentham R, Trentham D, Mortola JF (2001) Altered immune response in adult women exposed to diethylstilbestrol in utero. Am J Obstet Gynecol 185:78–81
Miller FW, Hess EV, Clauw DJ, Hertzman PA, Pincus T, Silver RM, Mayes MD, Varga J, Medsger TA Jr, Love LA (2000) Approaches for identifying and defining environmentally associated rheumatic disorders. Arthritis Rheum 43:243–249
Patterson R, Germolec D (2005) Toxic oil syndrome: review of immune aspects of the disease. J Immunotoxicol 2:51–58
Kaufman LD, Krupp LB (1995) Eosinophilia-myalgia syndrome, toxic-oil syndrome, and diffuse fasciitis with eosinophilia. Curr Opin Rheumatol 7:560–567
Back EE, Henning KJ, Kallenbach LR, Brix KA, Gunn RA, Melius JM (1993) Risk factors for developing eosinophilia myalgia syndrome among L-tryptophan users in New York. J Rheumatol 20:666–672
Kamb ML, Murphy JJ, Jones JL, Caston JC, Nederlof K, Horney LF, Swygert LA, Falk H, Kilbourne EM (1992) Eosinophilia-myalgia syndrome in l-tryptophan-exposed patients. JAMA 267:77–82
Tabuenca JM (1981) Toxic-allergic syndrome caused by ingestion of rapeseed oil denatured with aniline. Lancet 2:567–568
Holladay SD (1999) Prenatal immunotoxicant exposure and postnatal autoimmune disease. Environ Health Perspect 107(Suppl 5):687–691
Holladay SD, Lindstrom P, Blaylock BL, Comment CE, Germolec DR, Heindell JJ, Luster MI (1991) Perinatal thymocyte antigen expression and postnatal immune development altered by gestational exposure to tetrachlorodibenzo-p-dioxin (TCDD). Teratology 44:385–393
Blaylock BL, Holladay SD, Comment CE, Heindel JJ, Luster MI (1992) Exposure to tetrachlorodibenzo-p-dioxin (TCDD) alters fetal thymocyte maturation. Toxicol Appl Pharmacol 112:207–213
Silverstone AE, Frazier DE, Fiore NC, Soults JA, Gasiewicz TA (1994) Dexamethasone, β-estradiol, and 2,3,7,8-tetrachlorodibenzo-p-dioxin elicit thymic atrophy through different cellular targets. Toxicol Appl Pharmacol 126:248–259
Mustafa A, Holladay SD, Goff M, Witonsky S, Kerr R, Weinstein DA, Karpuzoglu-Belgin E, Gogal RM Jr (2009) Developmental exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin alters postnatal T cell phenotypes and T cell function and exacerbates autoimmune lupus in 24-week-old SNF1 mice. Birth Defects Res A Clin Mol Teratol 85:828–836
Silverstone AE, Gavalchin J, Gasiewicz TA (1998) TCDD, DES, and estradiol potentiate a lupus-like autoimmune nephritis in NZB × SWR (SNF1) mice. Toxicologist 42:403
Smith DA, Germolec DR (2000) Developmental exposure to TCDD and mercuric chloride in autoimmune-prone MRL/lpr mice. Toxicologist 54:8
Mustafa A, Holladay SD, Goff M, Witonsky SG, Kerr R, Reilly CM, Sponenberg DP, Gogal RM Jr (2008) An enhanced postnatal autoimmune profile in 24 week-old C57BL/6 mice developmentally exposed to TCDD. Toxicol Appl Pharmacol 232:51–59
Germolec DR (2005) Autoimmune diseases, animal models. In: Vohr HW (ed) Encyclopedic reference of immunotoxicology. Springer, Berlin, Germany, pp 75–79
Pieters R, Ezendam J, Bleumink R, Bol M, Nierkens S (2002) Predictive testing for autoimmunity. Toxicol Lett 127:83–91
NRC (1983) Risk assessment in the federal government: managing the process, Committee on the institutional means for assessment of risks to public health, Commission on life sciences. National Research Council, Washington DC, pp 1–192
NRC (1994) Science and judgment in risk assessment, Committee on risk assessment of hazardous air pollutants, Board on environmental studies and toxicology, Commission on life sciences. National Research Council, Washington DC, pp 1–672
WHO (1999) Principles for the assessment of risks to human health from exposure to chemicals, IPCS environmental health criteria 210. International programme on chemical safety. World Health Organization, Geneva, Switzerland
Hill AB (1965) The environment and disease: association or causation? Proc R Soc Med 58:295–300
Weed DL (2005) Weight of evidence: a review of concept and methods. Risk Analysis 25:1545–1557
Luster MI, Germolec DR, Parks C, Blanciforti L, Kashon M, Luebke RW (2005) Associating changes in the immune system with clinical diseases for interpretation in risk assessment. In: Tryphonas H, Fournier M, Blakley BR, Smits JE, Brousseau P (eds) Investigative immunotoxicology. Taylor & Francis, New York, NY, pp 165–182
Dietert RR, Dietert JM (2007) Early-life immune insult and developmental immunotoxicity (DIT)-associated diseases: potential of herbal- and fungal-derived medicinals. Curr Med Chem 14:1075–1085
Huang H, Patel D, Manton K (2005) The immune system in aging: roles of cytokines, T cells and NK cells. Front Biosci 10:192–215
U.S. EPA (2002) A review of the reference dose and reference concentration processes. Report. Risk Assessment Forum, Washington, DC, pp 1–192
WHO (2009) Principles for modelling dose-response for the risk assessment of chemicals, IPCS environmental health criteria 239. International programme on chemical safety. World Health Organization, Geneva, Switzerland
U.S. EPA (1995) Use of the benchmark dose approach in health risk assessment. Report. Risk Assessment Forum, Washington DC, pp 1–93
U.S. EPA (2000) Benchmark dose technical guidance document. External review draft. Report. Risk Assessment Forum, Washington DC, pp 1–96
WHO (2000) Human exposure assessment, IPCS environmental health criteria 214. International programme on chemical safety. World Health Organization, Geneva, Switzerland
U.S. EPA (1992) Guidelines for exposure assessment. Report. Risk Assessment Forum, Washington DC, pp 1–126
U.S. EPA (2005) Guidance on selecting age groups for monitoring and assessing childhood exposures to environmental contaminants. Report. Risk Assessment Forum, Washington DC, pp 1–50
U.S. EPA (1998) Health effects test guidelines: OPPTS 870.7800 immunotoxicity. Report. Office of Prevention, Pesticides and Toxic Substances, Washington DC
Selgrade MK, Gilmour MI (2006) Immunotoxicology of inhaled compounds―assessing risks of local immune suppression and hypersensitivity. J Toxicol Environ Health A 69:827–844
Acknowledgments
This research was supported by the Intramural Research program of the National Institutes of Health, National Institute of Environmental Health Sciences. The findings and conclusions in this report represent the opinions of the authors and do not represent, and should not be construed to represent, statements, opinions, or conclusions of the National Toxicology Program, NIEHS, NIH, the Environmental Protection Agency, or the United States Government. This chapter has been reviewed by the Environmental Protection Agency’s Office of Research and Development and approved for publication. We thank Dr. Andrew Hotchkiss and Dr. David Lehmann for their thoughtful review of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer Basel AG
About this chapter
Cite this chapter
Rooney, A.A., Luebke, R.W., Selgrade, M.K., Germolec, D.R. (2012). Immunotoxicology and Its Application in Risk Assessment. In: Luch, A. (eds) Molecular, Clinical and Environmental Toxicology. Experientia Supplementum, vol 101. Springer, Basel. https://doi.org/10.1007/978-3-7643-8340-4_9
Download citation
DOI: https://doi.org/10.1007/978-3-7643-8340-4_9
Published:
Publisher Name: Springer, Basel
Print ISBN: 978-3-7643-8339-8
Online ISBN: 978-3-7643-8340-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)