
Abstract
The variations in nutrients (molar ratios) and chlorophyll-a in the Jiaozhou Bay were examined before and after a diatom bloom in a period that lasted from November 2003 to March 2004. Negative relationships between nutrient concentrations, Si /P, Si /N ratios, and chlorophyll-a content were found during the bloom , which reflected the relationship between nutrient concentrations, phytoplankton biomass , and growth. Large increase in nutrient concentrations, particularly SiO3–Si after the late autumn, is one of major reasons inducing the diatom bloom in winter, and the bloom was finally controlled due to SiO3–Si depletion by phytoplankton . The bloom was mainly controlled by SiO3–Si . Before 1998, relatively low level of SiO3–Si kept ecological balance of eutrophication waters in the Jiaozhou Bay . In recent years, however, human activities have increased SiO3–Si concentration, which is likely one of the primary causes for the increased diatom blooms in the Jiaozhou Bay . Therefore, it is necessary to control SiO3–Si concentration in the Jiaozhou Bay.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
In the past few decades, coastal marine phytoplankton blooms have increased dramatically in frequency, intensity, and geographic distribution (Smayda 1990; Hallegraeff 1993; Hodgkiss and Ho 1997; Paerl 1997; Zhou et al. 2003, 2008). It has been suggested that nutrient -impacted coastal and offshore waters are experiencing an epidemic of harmful phytoplankton blooms (Smayda 1990). However, as Hodgkiss and Ho (1997) showed, it is still not possible to conclude the extent to which the increase in blooms in coastal waters can be attributed to the increase in nutrient levels, since so many other factors are involved. Obviously, the occurrence and increase of phytoplankton blooms are related to the changes in nutrient ratios (Hodgkiss and Chan 1987; Smayda 1989; Hodgkiss and Ho 1997; Flynn 2002; Domingues et al. 2005). Diatom blooms are common in temperate aquatic systems, and their dynamics are often controlled by nutrients, especially Si (Conley and Malone 1992). As a result of increased human activities , N and P loads have considerably increased, especially N. In contrast to N and P, Si concentration may be reduced due to hydrologic changes in a watershed (Correll et al. 2000; Humborg et al. 2000). The situation has seriously affected the N:P:Si ratio and resulted in lower Si :N and higher N:P ratios in lotic systems, and it has been argued that nutrient limitation of phytoplankton growth in aquatic systems is moving toward a higher incidence of P and Si limitation (Turner et al. 2003). Similar change has been also observed in the Jiaozhou Bay (Shen 2001; Shen et al. 2006).
The study on long-term changes in nutrient structure in the Jiaozhou Bay showed that since the 1960s, due to the influences of human activities , N and P concentrations have significantly increased, while Si concentration has remained at a relatively stable or lower level due to damming of waterways, which has resulted in a change in nutrient limitation of phytoplankton growth from N and P to Si (Shen 2001). Si limitation appears favorable for controlling the ecological equilibrium of the Jiaozhou Bay (Shen et al. 2006). Ecological conditions were reasonably stable up until 1998, when accelerating eutrophication became more evident in the Jiaozhou Bay (Yao et al. 2007). In recent years, due to rapid economic developments, including city construction that has been increasing at 15% per year since 1997 with corresponding increases in cement consumption in Qingdao City. The waste entering the bay by surface runoff has increased the Si concentration in the seawater, and the occurrence of bloom increased dramatically in the Jiaozhou Bay . For example, blooms of Skeletonema costatum and Eucampia zodiacus occurred in July 1998 and June 1999, respectively, in the north-eastern part of the Jiaozhou Bay (Huo et al. 2001; Zhang et al. 2002). Subsequently, the blooms caused by Coscinodiscus asteromphanus and Lauderia annulata were found by our investigations in June 2003 and February 2004, respectively, and the latter was found in winter for the first time.
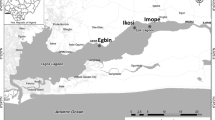
In this section, four observation stations associated with the bloom were set up in the Jiaozhou Bay (Fig. 1) and surface seawater samples were collected every ten days from November 2003 to March 2004, for determining phosphate (PO4–P) , silicate (SiO3–Si ), nitrate (NO3–N) , nitrite (NO2–N) , ammonium (NH4–N), and chlorophyll-a (Chl-a) . Dissolved inorganic nitrogen (DIN) was calculated as the sum of NO3–N, NO2–N, and NH4–N. Seawater transparence was observed using a transparent scale. Phytoplankton samples were collected vertically from the bottom to surface layer using a phytoplankton net (37 cm diameter, 140 cm length, and 77 meshes). We use the data observed during five months before and after the bloom , as well as the data from investigations since 1991, the variations of nutrient concentrations and molar ratios, phytoplankton cell abundance, and Chl-a content are discussed; the relation of Si supply and diatom bloom are studied; and the causes of diatom bloom increasing in recent years are explored.

Survey stations in the Jiaozhou Bay
1 Nutrient Concentrations and Molar Ratios
The average concentrations and molar ratios of nutrients in the surface layer water during investigations are listed in Table 1. Particularly high nutrient concentrations were found at station A5 situated in the north-eastern part of the bay with a weak water exchange (Zhao et al. 2002), which is near the main industrial and residential areas of Qingdao City. Very high DIN /PO4–P (N/P) ratio and higher SiO3–Si /PO4–P (Si /P) ratio were also found at station A5, as a result of very high DIN and higher SiO3–Si concentrations. Station C3 is situated in the middle part of the bay with relatively low nutrient concentrations and ratios. Stations A1 and A3 are located in the marine aquaculture areas and were influenced by waste from agricultural nonpoint sources along the coast. Moreover, station A1 was also influenced by the discharge of the Dagu River water. The concentrations and ratios of nutrient at stations A1 and A3 were relatively high and between stations A5 and C3. The SiO3–Si /DIN (Si /N) ratio was low in the four stations due to high concentrations of DIN and relatively low concentrations of SiO3–Si .
2 Variations in Nutrient Concentrations and Ratios During Ten-Day Period
Variations in nutrient concentrations in the surface water at stations A1, A3, A5, and C3 during ten-day period from November 2003 to March 2004 (Fig. 2) showed that the maximum changes in DIN , PO4–P , SiO3–Si concentrations were at station A5 and the minimum changes were at station C3. DIN concentrations reached very high values of 67.3, 160.3 μmol L−1 for stations A3 and A5 on January 8, 13.2 μmol L−1 for station C3 on January 17, and 44.6 μmol L−1 for station A1 on January 30. After attaining high values, DIN concentrations decreased to minimum values of 7.8, 34.0, 6.4 μmol L−1, respectively, at stations A3, A5, and C3, and 35.9 μmol L−1 at station A1 on February 9. They then increased again. PO4–P concentrations generally showed a downward trend and declined to the lowest values of 0.33, 0.36, 0.26 μmol L−1 at stations A1, A3, and A5, respectively, and 0.27 μmol L−1 at station C3 on February 18 and then recovered rapidly. The change of SiO3–Si concentration was significant. The maximum values of 12.2 and 24.9 μmol L−1 at stations A1 and A3, respectively, occurred on January 8, and the maximum of 97.7 μmol L−1 at station A5 and the second highest value of 8.1 μmol L−1 at station C3 occurred on December 21. After the maximum values, the concentrations of Si decreased rapidly to the lowest values of 3.2, 1.2, 0.49, and 0.38 μmol L−1 at stations A1, A3, A5, and C3, respectively, on February 9 or 18.

Ten-day period variations in nutrients and Chl-a contents: filled circle—Chl-a; filled square—DIN ; filled diamond—PO4–P ; filled triangle—SiO3-Si
Variations in nutrient molar ratios in the surface water at stations A1, A3, A5, and C3 during ten-day period (Fig. 3) were similar to those of nutrients. The maximum changes in N/P, Si /P, Si /N ratios were also found at station A5, and the minimum changes were at station C3. The highest or second highest N/P ratios occurred at stations A1, A3, and A5 on February 9 or January 30 and then descended obviously. The Si /P ratios reached the highest values of 19.1, 17.2, 43.0, and 11.8 at stations A1, A3, A5, and C3 on January 8, December 30, December 10, and January 17, respectively. After the highest, the Si /P ratios decreased rapidly to the lowest values of 3.4, 1.8, 1.6 for stations A1, A3, and A5, respectively, on February 18 and 1.3 for station C3 on February 9. They then increased again. Small changes were found in Si /N ratios. After experiencing the highest values, they decreased to the lowest values of 0.09, 0.06, 0.03, and 0.05 at stations A1, A3, A5, and C3, respectively, on February 18 or 28 and then increased gradually.

Ten-day period variations in nutrient ratios and Chl-a content: filled circle—Chl-a; filled square—N/P; filled diamondt—Si /P; filled triangle—Si /N
3 Variation in Phytoplankton Cell Abundance and Biomass During Ten-Day Period
Variations in average cell abundances of phytoplankton at the four stations during ten-day period (Fig. 4) showed that a minimum value of 0.153 × 103 cells L−1 was on December 21, after which the cell abundance increased quickly and reached a maximum of 493 × 103 cells L−1 on February 9 and then descended quickly again. Among the four stations, the maximum change in phytoplankton cell abundance occurred at station A5 and the maximum value of phytoplankton cell abundance reached 640 × 103 cells L−1 on February 9. Phytoplankton dominant species were L. annulata, S. costatum, Rhizosolenia delicatula, Chaetoceros debilis, Asterionella kariana, etc., suggesting that this bloom was mainly composed of the smaller cell species of L. annulata. At the peak of the bloom , the cell abundances of L. annulata reached 30.7 × 103, 330.7 × 103, 541.3 × 103, and 75.7 × 103 cells L−1 and accounted for 35.4, 29.2, 84.6, and 37.1% of total cell abundances at stations A1, A3, A5, and C3, respectively. The ten-day period changes in Chl-a content (Fig. 2) were consistent with those in cell abundances of phytoplankton ; the most notable event occurred at stations A3 and A5. After December 21, Chl-a contents increased gradually and the highest Chl-a content of 39.03 μg L−1 appeared at station A5 and the lowest that of 1.96 μg L−1 was at station A1 at the peak of the bloom , suggesting that this bloom mainly occurred at station A5.

Ten-day period variations in phytoplankton cell abundance: filled circle—average cell abundances for four stations; filled square—station A5
4 Relationships Between Nutrient Concentrations and Phytoplankton Biomass
A comparison of nutrient concentrations and Chl-a content at the four stations during the course of this study, Chl-a content was very high for (8.03 ± 10.46) μg L−1 at station A5, following was (2.68 ± 2.71) μg L−1 at station A3. The lowest Chl-a content was at station A1 for (0.96 ± 0.63) μg L−1, which was consistent with the changes in cell abundances of phytoplankton , suggesting that phytoplankton growth was mainly controlled by nutrients. However, the phytoplankton growth was not only dependent on nutrient concentrations but also on other environment factors (Reynolds 1989; Hodgkiss and Ho 1997; Domingues et al. 2005). Cloern (1987) showed that the variability of phytoplankton biomass in estuaries can be related to the changes in light availability. When comparing environment conditions at station C3 with other stations, the differences in water temperature and salinity were insignificant (Table 1). Light availability (seawater transparence) was better at station C3 ((2.1 ± 1.0) m) than other stations ((1.3 ± 0.6) m, (1.6 ± 0.6) m, and (1.5 ± 0.4) m at stations A1, A3, and A5, respectively), which was favorable for phytoplankton photosynthesis.
Chl-a content at station A5 with high concentration of nutrients always kept a relative high level, even though after the peak of the bloom (Fig. 2). Pearson correlative analyses between nutrient concentrations and Chl-a content at every station before and after the bloom show that the most notable negative correlations were found at station A5, following station C3 (Table 2), which reflected the nutrient uptake and release associated with phytoplankton growth and death. The nutrients in the Jiaozhou Bay come mainly from terrestrial sources (Shen 2001; Wang et al. 2006; Sun et al. 2011b). Due to a few rainwater and wastes from agriculture and marine aquaculture in winter, the outer-source nutrients come mainly from industrial waste and residential sewage (including construction wastewater) from urban districts. Station A5 is near river mouths discharged waste (sewage); nutrients discharged in a steady stream were a main cause of maintaining high Chl-a content there. Investigations showed that the maximum SiO3–Si concentration was found in the Licun River mouth with 119.8 μmol L−1 (Yao and Shen 2006), which is possibly relative to a mass of silicious construction wastewater entering. In addition, the regeneration and accumulation of nutrients in late autumn was also an important source deriving from plankton death and decomposition. Continuous access to nutrients is essential for maintaining phytoplankton growth. Nutrient availability has frequently outweighed other factors as being the easiest way to initiate a bloom (Liu et al. 2005). As a result of negative relationships between SiO3–Si concentration and Chl-a content, the clearly or more clearly negative relationships between the ratios of Si /P, Si /N, and Chl-a content were found at stations A3, A5, and C3 (Table 2), suggesting that phytoplankton growth was related to the ratios of limiting nutrients (Shen et al. 2006). However, poor relations between N/P ratio and Chl-a content were found at those stations, due to N/P ratio much higher than Redfield value (16) (Redfield et al. 1963).
5 Bloom and Si Supply
Bloom is characterized by high values of Chl-a and phytoplankton biomass (Liu et al. 2005). As mentioned above, station A5 was the major area of this bloom . When SiO3–Si and PO4–P concentrations reached the maximum values (December 21), phytoplankton cell abundance and Chl-a content began to increase. Particularly after January 8, phytoplankton bred rapidly, phytoplankton cell abundance increased from 5.38 × 103 to 640 × 103 cells L−1 (Fig. 4), and Chl-a content increased from 0.56 to 39.03 μg L−1 (Fig. 2) (February 9). Similar variations also occurred at stations A1, A3, and C3. Paerl (1997) showed that particular importance in nutrient -sensitive water is the rates of new N and other nutrient supply-fundamental determinants of bloom development, maintenance, and proliferation. In the Jiaozhou Bay , phytoplankton community composition is mainly diatom , accounting for more than 99% (Sun et al. 2011a). Because of N excess, PO4–P plenty, and SiO3–Si relative insufficient in the Jiaozhou Bay , continuous supply of Si was the determinant function for occurrence and development of this bloom . Along with the increasing in phytoplankton quantity, the nutrient concentrations descended quickly to the lowest values of 34.0, 0.27, and 0.49 μmol L−1 for DIN , PO4–P , and SiO3–Si respectively at the peak of the bloom . Very high N/P ratio of 130.8, lower Si /P ratio of 4.6, and very low Si /N ratio of 0.04 were found at the bloom peak, and after which, they decreased to the lowest values of 42.8, 1.6, and 0.03 for N/P, Si /P, and Si /N ratios respectively, suggesting that the changes in nutrient ratios were behind in the nutrient concentrations. Above data show that at the peak of the bloom , only SiO3–Si concentration decreased to below the threshold value of diatom growth (2.0 μmol L−1) while DIN and PO4–P concentrations were clearly higher than their threshold values (1.0 and 0.10 μmol L−1, respectively) (Brown and Button 1979; Perry and Eppley 1981; Goldman and Glibert 1983; Nelson and Brzezinski 1990). Among various molar ratios of nutrients, N/P ratio was far higher than Redfield value (16) for the mean ratio of nutrients contained in marine diatom , only Si /P and Si /N ratios related to SiO3–Si were much lower than Redfield values (16 and 1, respectively) (Redfield et al. 1963; Brzezinski 1985). The differences in Si /P and Si /N ratios were about one order of magnitude larger around the bloom . Large increase in nutrients, especially SiO3–Si after the late autumn, led to rapid increase of diatom , which is one of major reasons arousing the bloom in winter. After which, due to SiO3–Si was nearly exhausted by phytoplankton and very low Si /P and Si /N ratios, the bloom was finally controlled. The finding is somewhat similar to the nutrient -enriched Chesapeake Bay, where the magnitude of the spring phytoplankton bloom was controlled by SiO3–Si (Conley and Malone 1992). The changes in nutrients and the ratios around the bloom peak showed that the occurrence and end of the bloom were not only controlled by nutrients, but also well related to the changes in nutrient ratios. Obviously, as Smayda (1989) put it, other factors will be involved in regulating bloom events, but nutrient ratios can be considered to be of major importance. Phytoplankton bloom dynamics rely on the synergistic interactions of favorable physical, chemical, and biotic conditions (Paerl 1988). Continuous supply of Si was the determinant function for occurrence and development of this bloom ; however, the bloom was not happening when highest SiO3–Si concentration. In addition to plenty of nutrients supply, suitable temperature and salinity , weather and sea conditions also play an important role in the bloom formation. At the peak of the bloom on February 9, seawater was maroon. Weather was smoke and fog, small stormy waves, and the wind speed of 2.5 m/s which were favorable to gathering phytoplankton .
According to the quarter investigations from May 1991 to May 2004, the nutrient concentrations tended to increase in the Jiaozhou Bay , especially after 1999. To take station A5 as example (Figs. 5 and 6), the changes in DIN , PO4–P , and SiO3–Si concentrations were between 7.1 and 44.4 μmol L−1, 0.08 and 1.8 μmol L−1, below the detection limit , and 6.1 μmol L−1, respectively, before 1999 (1990s). Among these three nutrients, SiO3–Si concentration was the lowest. The probability of SiO3–Si concentration less than 2.0 μmol L−1 (threshold value of diatom growth) accounting for all investigation data was as high as 50%, while that of PO4–P concentration less than the threshold value (0.1 μmol L−1) was only 6%, and all DIN concentrations were greater than the threshold value (1 μmol L−1). During 1999 and 2004 (2000s), DIN , PO4–P, SiO3–Si concentrations were between 7.4 and 123.7 μmol L−1, 0.16 and 2.0 μmol L−1, below the detection limit , and 45.4 μmol L−1, respectively. Comparing with 1990s, their average values increased 0.99 times for DIN , 0.25 times for PO4–P , and 3.2 times for SiO3–Si , and the increasing of SiO3–Si was the most obvious. The variations in nutrients at station A5 were consistent with the Jiaozhou Bay (Sun et al. 2011b). SiO3–Si is mainly from river transport . According to the data from Qingdao Observatory, annual average precipitation was 643.7 mm for the 1990s and 684.2 mm for the 2000s. The increase in precipitation means the increase in SiO3–Si into the Jiaozhou Bay brought by precipitation surface runoff . However, it possibly shows that less increase in precipitation is insignificant for SiO3–Si increase from the 1990s to the 2000s. The increasing of SiO3–Si is possibly relative to rapid economic developments, especially city construction in Qingdao since the late 1990s. To take floor space of buildings completed as example, floor space of buildings completed per annum were 203 × 104–259 × 104 m2 during 1995–1998, 474 × 104 m2 in 1999, and 635 × 104 m2 in 2004, suggesting that a lot of construction wastewater entering the bay increased Si concentration in the seawater. The changes in nutrient supplies are often accompanied by alterations in nutrient ratios (Yin et al. 2001). The molar ratios of nutrients also increased with the increase of nutrient concentrations (mainly N and Si ). In the 1990s, the changes in N/P, Si /P, and Si /N ratios were between 15.9 and 88.4, 0 and 12.2, 0 and 0.51, respectively, and Si /P and Si /N ratios were much lower than Redfield value (16 and 1, respectively) (Redfield et al. 1963; Brzezinski, 1985). In the 2000s, N/P, Si /P, and Si /N ratios were between 22.4 and 118.1, 0 and 37.8, 0 and 0.57, respectively, and increased 0.35, 0.88, and 0.19 times, respectively, as compared with 1990s. Relative to the nutrient concentrations increased significantly, temperature and salinity in the surface seawater had no obvious changes (Fig. 7). The survey found that the season averages of temperature and salinity were (15.72 ± 7.75) ℃ and 31.34 ± 0.95 in the 1990s, and (14.32 ± 8.42) ℃ and 31.38 ± 0.87 in the 2000s, respectively. Higher temperature in the 1990s is associated with the data of a lack of winter. Many nutrient enrichment experiments have shown that marine phytoplankton blooms are often nutrient limited (Hodgkiss and Ho 1997). Although eutrophication in the Jiaozhou Bay was more serious before 1998, there were few bloom events and the bay was in relatively stable ecological condition. Low SiO3–Si concentration and Si /P, Si /N ratios limited phytoplankton growth, especially in winter (Shen 2001) when phytoplankton reproduced frequently (Wu et al. 2004). In recent years, the nutrient concentrations showed increasing trends in the Jiaozhou Bay , especially Si limitation was alleviated compared to the 1990s due to the increasing in SiO3–Si concentration, which supplied material base for phytoplankton growth (Sun et al. 2011b). The study showed that from the 1990s to the 2000s, net phytoplankton cell abundance increased from (2.74 ± 3.17) × 103 cells L−1 to (13.04 ± 27.08) × 103 cells L−1, increased almost one order of magnitude (Fig. 7). After 1998, the bloom event significantly increased with the increase in nutrient concentrations, molar ratios, and phytoplankton number. According to the report of Wu et al. (2005), before the 1990s, there was not bloom event report in the Jiaozhou Bay , in the late 1990s, the rapid development of bloom , almost occurred every year, among them, two bloom events were recorded in 1999, 2003, and 2004. So the nutrient loading has an obvious relationship with increased blooms (Hallegraeff 1993). For example, in a bloom of E. zodiacus occurred in the north-eastern part of the Jiaozhou Bay (Zhang et al. 2002), PO4–P and SiO3–Si concentrations decreased from 2.1 and 19.2 μmol L−1 to 0.40 and 0.92 μmol L−1, respectively. Only when SiO3–Si concentration decreased to below threshold value of diatom growth (2.0 μmol L−1), the bloom was finally controlled. This is very similar to the development and end of the bloom found by us in winter in this section.

Years variations in nutrient concentrations at station A5 from 1991 to 2004. Dotted line—averaged value; F—February; Ma—March; Ap—April; M—May; J—June; A—August; S—September; N—November; D—December

Years variations in nutrient ratios at station A5 from 1991 to 2004. Dotted line—averaged value; F—February; Ma—March; Ap—April; M—May; J—June; A—August; S—September; N—November; D—December

Years variations in temperature , salinity , and phytoplankton cell abundance at station A5 from 1991 to 2004. Dotted line—averaged value; F—February; Ma—March; Ap—April; M—May; J—June; A—August; S—September; N—November; D—December
Here, we can suggest that SiO3–Si kept ecological balance of eutrophication waters in the Jiaozhou Bay where Si is relatively insufficient. Once the SiO3–Si concentration increases significantly, the equilibrium between nutrients and phytoplankton will be destructed and lead to rapid increase of diatom and arouse blooms when other conditions (such as temperature , water current state) are quite suitable. It is likely one of the primary causes for the increased blooms in recent years in the Jiaozhou Bay . In this sense, SiO3–Si limitation is favorable for keeping ecological equilibrium of the Jiaozhou Bay (Shen et al. 2006). From a practical perspective, regulating nutrient supply is often the only feasible management approach to bloom control (Paerl 1997). Hence, we suggest that from now on, the increase of SiO3–Si concentration should be controlled in the Jiaozhou Bay .
References
Brown, E. J., & Button, D. K. (1979). Phosphate-limited growth kinetics of Selanastrum capricornatum (Chlorophyceae). Journal of Phycology, 15, 305–311.
Brzezinski, M. A. (1985). The Si:C:N ratio of marine diatoms: Interspecific variability and the effect of some environmental variables. Journal of Phycology, 21, 347–357.
Cloern, J. E. (1987). Turbidity as a control on phytoplankton biomass and productivity in estuaries. Continental Shelf Research, 7, 1367–1381.
Conley, D. J., & Malone, T. C. (1992). Annual cycle of dissolved silicate in Chesapeake Bay: Implications for the production and fate of phytoplankton biomass. Marine Ecology Progress Series, 81, 121–128.
Correll, D. L., Jordan, T. E., & Weller, D. E. (2000). Beaver pond biogeochemical effects in the Maryland Coastal Plain. Biogeochemistry, 49, 217–239.
Domingues, R. B., Barbosa, A., & Galvao, H. (2005). Nutrients, light and phytoplankton succession in a temperate estuary (the Guadiana, south-western Iberia). Estuarine, Coastal and Shelf Science, 64, 249–260.
Flynn, K. J. (2002). How critical is the critical N:P ratio? Journal of Phycology, 38, 961–970.
Goldman, J. C., & Glibert, P. M. (1983). Kinetics of inorganic nitrogen uptake by phytoplankton. In E. J. Carpenter & D. G. Capone (Eds.), Nitrogen in marine environments (pp. 233–274). New York: Academic press.
Hallegraeff, G. M. (1993). A review of harmful algal blooms and their apparent global increase. Phycologia, 32, 79–99.
Hodgkiss, I. J., & Chan, B. S. S. (1987). Phytoplankton dynamics in Tolo Harbour. Asian Marine Biology, 4, 103–112.
Hodgkiss, I. J., & Ho, K. C. (1997). Are changes in N:P ratios in coastal waters the key to increased red tide blooms? Hydrobiologia, 352, 141–147.
Humborg, C., Conley, D. J., Rahm, L., Wulff, F., Cociasu, A., & Ittekkot, V. (2000). Silicon retention in river basins: Far-reaching effects on biogeochemistry and aquatic food webs in coastal marine environments. Ambio, 29, 45–50.
Huo, W. Y., Yu, Z. M., Zou, J. Z., Song, X. X., & Hao, J. H. (2001). Outbreak of Skeletonema costatum red-tide and its relations to environmental factors in Jiaozhou Bay. Oceanologia et Limnologia Sinica, 32, 311–318. (in Chinese with English abstract).
Liu, D. Y., Sun, J., Zou, J. Z., & Zhang, J. (2005). Phytoplankton succession during a red tide of Skeletonema costatum in Jiaozhou Bay of China. Marine Pollution Bulletin, 50, 91–94.
Nelson, D. M., & Brzezinski, A. (1990). Kinetics of silicate acid uptake by natural diatom assemblages in two Gulf & Stream warm-core rings. Marine Ecology Progress Series, 62, 283–292.
Paerl, H. W. (1988). Nuisance phytoplankton blooms in coastal, estuarine and inland waters. Limnology and Oceanography, 33, 823–847.
Paerl, H. W. (1997). Coastal eutrophication and harmful algal blooms: Importance of atmospheric deposition and groundwater as “new” nitrogen and other nutrient sources. Limnology and Oceanography, 42, 1154–1165.
Perry, M. J., & Eppley, R. W. (1981). Phosphate uptake by phytoplankton in the central North Pacific Ocean. Deep-Sea Research, 28, 39–49.
Redfield, A. C., Ketchum, B. H., & Richards, F. A. (1963). The influence of organisms on the composition of seawater. In M. N. Hill (Ed.), The Sea (Vol. 2, pp. 26–77)., the composition of seawater New York: Wiley.
Reynolds, C. S. (1989). Physical determinants of phytoplankton succession. In U. Sommer (Ed.), Plankton Ecology (pp. 9–56). Berlin Heidelberg: Springer-Verlag.
Shen, Z. L. (2001). Historical changes in nutrient structure and its influences on phytoplankton composition in Jiaozhou Bay. Estuarine, Coastal and Shelf Science, 52, 211–224.
Shen, Z. L., Liu, Q., Wu, Y. L., & Yao, Y. (2006). Nutrient structure of seawater and ecological responses in Jiaozhou Bay, China. Estuarine, Coastal and Shelf Science, 69, 299–307.
Smayda, T. J. (1989). Primary production and the global epidemic of phytoplankton blooms in the sea: A linkage? In E. M. Cosper, V. M. Bricelj, & E. J. Carpenter (Eds.), Novel phytoplankton blooms (pp. 449–483). Berlin, Heidelberg, New York: Springer Verlag.
Smayda, T. J. (1990). Novel and nuisance phytoplankton blooms in the sea: Evidence for a global epidemic. In E. Graneli, B. Sundstrom, R. Edler, & D. M. Anderson (Eds.), Toxic Marine Phytoplankton (pp. 29–40). New York: Elsevier Science.
Sun, X. X., Sun, S., Wu, Y. L., Zhang, Y. S., & Zheng, S. (2011a). Long-term changes of phytoplankton community structure in the Jiaozhou Bay. Oceanologia et Limnologia Sinica, 42, 639–646. (in Chinese with English abstract).
Sun, X. X., Sun, S., Zhao, Z. X., & Shen, Z. L. (2011b). Long-term changes in nutrient concentration and structure in the Jiaozhou Bay. Oceanologia et Limnologia Sinica, 42, 662–669. (in Chinese with English abstract).
Turner, R. E., Rabalais, N. N., Justic, D., & Dortch, Q. (2003). Future aquatic nutrient limitations. Marine Pollution Bulletin, 46, 1032–1034.
Wang, X. L., Li, K. Q., & Shi, X. Y. (2006). Marine environmental capacity of pollutants in Jiaozhou Bay (pp. 1–22). Beijing: Science Press.
Wu, Y. L., Sun, S., Zhang, Y. S., & Zhang, F. (2004). Quantitative study on long-term variation of phytoplankton in Jiaozhou Bay. Oceanologia et Limnologia Sinica, 35, 518–523. (in Chinese with English abstract).
Wu, Y. L., Sun, S., & Zhang, Y. S. (2005). Long-term change of environment and it’s influence on phytoplankton community structure in Jiaozhou Bay. Oceanologia et Limnologia Sinica, 36, 487–498. (in Chinese with English abstract).
Yao, Y., & Shen, Z. L. (2006). Characteristic of nutrient and its structure in Jaozhou Bay. Oceanologia et Limnologia Sinica, Supplement, 339–347. (in Chinese with English abstract).
Yao, Y., Zheng, S. Q., & Shen, Z. L. (2007). Study on the mechanism of eutrophication in the Jiaozhou Bay. Marine Science Bulletin, 26(4), 91–98. (in Chinese with English abstract).
Yin, K. D., Qian, P. Y., Wu, M. C. S., Chen, J. C., Huang, L., Song, X., et al. (2001). Shift from P to N limitation of phytoplankton growth across the Pearl river estuarine plume during summer. Marine Ecology Progress Series, 221, 17–28.
Zhang, Y. S., Wu, Y. L., Zou, J. Z., Yu, Z. M., & Pu, X. M. (2002). A red tide caused by diatom Eucampia zodiacus in the Jiaozhou Bay. Oceanologia et Limnologia Sinica, 33, 55–61. (in Chinese with English abstract).
Zhao, L., Wei, H., & Zhao, J. Z. (2002). Numerical study on water exchange in Jiaozttou Bay. Oceanologia et Limnologia Sinica, 33, 23–29. (in Chinese with English abstract).
Zhou, M. J., Yan, T., & Zou, J. Z. (2003). Preliminary analysis of the characteristics of red tide areas in Changjiang River estuary and its adjacent sea. Chinese Journal of Applied Ecology, 14, 1031–1038. (in Chinese with English abstract).
Zhou, M. J., Shen, Z. L., & Yu, R. C. (2008). Response of a coastal phytoplankton community to the increased of nutrient input from the Changjiang (Yangtze) River. Continental Shelf Research, 28, 1483–1489.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer-Verlag GmbH Germany, part of Springer Nature
About this chapter

Cite this chapter
Shen, Z., Yao, Y., Wu, Y. (2020). Silica Supply and Diatom Blooms in Jiaozhou Bay. In: Shen, Z. (eds) Studies of the Biogeochemistry of Typical Estuaries and Bays in China. Springer Earth System Sciences. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-662-58169-8_13
Download citation
DOI: https://doi.org/10.1007/978-3-662-58169-8_13
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-662-58167-4
Online ISBN: 978-3-662-58169-8
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)