
Abstract
The outer membrane (OM) is the front line of leptospiral interactions with their environment and the mammalian host. Unlike most invasive spirochetes, pathogenic leptospires must be able to survive in both free-living and host-adapted states. As organisms move from one set of environmental conditions to another, the OM must cope with a series of conflicting challenges. For example, the OM must be porous enough to allow nutrient uptake, yet robust enough to defend the cell against noxious substances. In the host, the OM presents a surface decorated with adhesins and receptors for attaching to, and acquiring, desirable host molecules such as the complement regulator, Factor H. On the other hand, the OM must enable leptospires to evade detection by the host’s immune system on their way from sites of invasion through the bloodstream to the protected niche of the proximal tubule. The picture that is emerging of the leptospiral OM is that, while it shares many of the characteristics of the OMs of spirochetes and Gram-negative bacteria, it is also unique and different in ways that make it of general interest to microbiologists. For example, unlike most other pathogenic spirochetes, the leptospiral OM is rich in lipopolysaccharide (LPS). Leptospiral LPS is similar to that of Gram-negative bacteria but has a number of unique structural features that may explain why it is not recognized by the LPS-specific Toll-like receptor 4 of humans. As in other spirochetes, lipoproteins are major components of the leptospiral OM, though their roles are poorly understood. The functions of transmembrane outer membrane proteins (OMPs) in many cases are better understood, thanks to homologies with their Gram-negative counterparts and the emergence of improved genetic techniques. This chapter will review recent discoveries involving the leptospiral OM and its role in leptospiral physiology and pathogenesis.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Peripheral Membrane Protein
- Pathogenic Leptospire
- Outer Membrane Lipoprotein
- Surface Lipoprotein
- Lipoprotein Signal Peptide
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Lipopolysaccharide (LPS)
LPS is a major component of the leptospiral OM and its polysaccharides dominate the leptospiral surface. The degree to which LPS is exposed on the leptospiral surface is reflected in the abundance of electron-dense particles on the surface of L. interrogans after incubation with a gold-labeled anti-LPS monoclonal antibody (Fig. 1). Agglutination occurs within minutes in the presence of small concentrations of LPS-specific antibodies. Monoclonal antibodies to LPS mediate macrophage opsonization (Farrelly et al. 1987) and protect animals against challenge with pathogenic leptospires (Jost et al. 1989). LPS-specific immune responses are the basis for the sterilizing immunity elicited by whole cell vaccines (Midwinter et al. 1994). Given the sensitivity of leptospires to LPS-specific antibodies, it is not surprising that there is tremendous selective pressure to undergo genetic changes leading to O-antigen variation. Hundreds of leptospiral serovars have been defined, based on differential reactivity with antibodies or antisera in the microscopic agglutination test (MAT). The simple addition of LPS-antiserum to a leptospiral culture can result in the growth of escape mutants with altered LPS.

Leptospira interrogans coated with gold-labeled anti-LPS monoclonal antibodies. The number of electron-dense particles reflects the level of LPS exposure on the leptospiral surface
Despite its accessibility, LPS is by no means a liability for these organisms. Expression of intact LPS appears to be essential for leptospiral survival both inside and outside the mammalian host. This conclusion is based in part on the finding that the rfb locus encoding the enzymes responsible for LPS biosynthesis was relatively spared of insertions in a study of random transposon mutagenesis (Murray et al. 2009a), suggesting that most LPS mutants are nonviable for growth in culture. The rare mutants that did survive transposon insertion into the LPS locus were attenuated for virulence and were rapidly cleared after challenge (Murray et al. 2010). Interestingly, the LPS expressed by one of these LPS mutants, M1352, had little or no change in its molecular mass, suggesting that even subtle changes in LPS can result in a loss of virulence. Mutant M1352 was effective as a live attenuated vaccine, stimulating both homologous and heterologous immunity in the hamster model of leptospirosis (Srikram et al. 2011). Differential detection of organisms in the liver and kidney using serovar-specific monoclonal antibodies suggests that the O-antigen side chains of leptospiral LPS are not static and may undergo antigenic changes during infection (Nally et al. 2005a).
1.1 LPS Structure and Biosynthesis
As in Gram-negative bacteria, leptospiral LPS consists of three components: lipid A , the core, and polysaccharide. The L. interrogans genome contains homologs of all the genes required for lipid A biosynthesis (Ren et al. 2003). The structure of leptospiral lipid A has now been fully elucidated and found to contain both similarities with, and striking differences from, typical forms of lipid A (Que-Gewirth et al. 2004). The first key difference is that L. interrogans converts the usual GlcNAc (N-acetylglucosamine) disaccharide backbone of lipid A to GlcNAc3 N, so that each of the two sugars has two amino groups instead of one. Consequently, there are four amide-linked fatty acids in the L. interrogans lipid A instead of two. This is unusual, but has been observed in some environmental bacteria. In addition, the leptospiral fatty acids in leptospiral lipid A differ in length from those typically found in Gram-negative lipid A and some are unsaturated. An even more unusual aspect of L. interrogans lipid A involves the phosphate residue. E. coli lipid A has two phosphates, one on each end of the disaccharide, whereas leptospiral lipid A has a single phosphate, and that single phosphate is methylated. Methylated phosphates are extremely unusual in biology and have not been previously observed in lipid A.
1.2 Innate Immunity: TLR4 and TLR2
The structural differences between LPS of E. coli and Leptospira are of great interest because of their differential recognition by TLR4, the Toll-like receptor involved in the innate immune response to LPS. While human TLR 4 reacts with E. coli LPS at extremely low concentrations, it is unable to interact with leptospiral LPS (Werts et al. 2001). Failure of human TLR4 to recognize leptospiral LPS may be one reason why humans are accidental hosts in whom leptospirosis occasionally causes overwhelming, lethal infections. In contrast, murine TLR4 is able to recognize leptospiral LPS (Nahori et al. 2005) and mice are natural, reservoir hosts for pathogenic leptospires. This idea is consistent with the observation that while mice with intact Toll-like receptors are resistant to leptospiral infection, young (but not adult) C3H/HeJ mice lacking TLR4 are susceptible to lethal infection with L. interrogans (Viriyakosol et al. 2006). Surprisingly, leptospiral LPS is recognized by both human and murine TLR2, the Toll-like receptor primarily involved in lipoprotein recognition. The importance of both TLR2 and TLR4 receptors in mice was highlighted by the finding that only when both of these receptors were knocked out did adult C57BL/6 J mice develop lethal infections after leptospiral challenge (Nahori et al. 2005). Murine TLR4 and TLR2 appear to recognize different leptospiral LPS components: TLR4 recognizes leptospiral lipid A while TLR2 recognizes the polysaccharide or 2-keto-3-deoxyoctonoic acid (KDO) portion of leptospiral LPS (Nahori et al. 2005; Werts 2010).
1.3 LPS Assembly and Transport
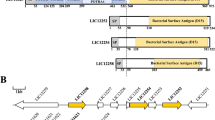
Many of the genes involved in LPS export to the OM are present in leptospiral genomes, suggesting that the processes are similar to those in typical Gram-negative bacteria. A number of excellent reviews on the subject of LPS assembly and transport have recently been published (Ruiz et al. 2009; Sperandeo et al. 2009). The lipid A and core components of LPS are assembled on the cytoplasmic surface of the inner membrane. These rough LPS molecules (lacking the O-antigen) are transported to the periplasmic leaflet of the inner membrane by the ABC transporter, MsbA. It has not yet been determined which of the many L. interrogans ABC transporters is MsbA. O-antigen is assembled via the Wzy-dependent pathway in which polysaccharides are synthesized on the cytoplasmic surface of the inner membrane, followed by transport across the inner membrane by the Wzx flippase (LIC12135), where they are ligated to rough LPS by Wzy O-antigen ligase (LIC11753). After polysaccharide has been added to the LPS core, full-length (smooth) LPS is transported across the periplasm by LptA to the LPS assembly site on the OM formed by LptD (aka OstA, LIC11458) and LptE (11007). LptD is a porin-like molecule and appears to be involved in translocating LPS to the OM surface. Cryo-electron tomography has shown that the thickness of the L. interrogans LPS layer, and presumably the length of its polysaccharide, is 50 % greater than that of L. biflexa (Fig. 2), which again illustrates the importance of LPS for virulence (Raddi et al. 2012).

Cryo-electron tomography of L. interrogans versus L. biflexa. The thickness of the LPS layer of L. interrogans is 9.2 nm versus 6.0 nm for L. biflexa. The increased thickness of the L. interrogans LPS layer is probably important for virulence. Reproduced from Raddi et al. (2012)
2 Outer Membrane Proteins (OMPs)
2.1 General Considerations
In recent years, much has been learned about the identity, expression, and functions of OMPs. The picture of the OM that has emerged (Fig. 3) is the result of improved methods for determining whether proteins are located in the OM and on its surface. A number of cell fractionation methods have been developed, including Triton X-114 fractionation (Haake et al. 1991; Zuerner et al. 1991), isolation of OM vesicles by sucrose density gradient fractionation (Haake and Matsunaga 2002; Nally et al. 2005b), and membrane fractionation (Matsunaga et al. 2002). Of particular importance are methods to identify surface-exposed OMPs. Multiple assays should be applied, including surface and subsurface controls, before concluding whether a particular protein is surface exposed. The most accurate methods include surface immunoprecipitation (Haake et al. 1991), surface biotinylation (Cullen et al. 2003), surface proteolysis (Pinne and Haake 2009), and surface immunofluorescence (Pinne and Haake 2011).

Membrane architecture of L. interrogans. The outer membrane contains LPS, lipoproteins such as LipL21, Loa22, and LigA, and transmembrane proteins such as FecA, OmpL1, TolC, and possibly Loa22. Peptidoglycan and the periplasmic flagella PF are located in the periplasm
Particularly useful has been the application of matrix-assisted laser desorption/ionization time of flight (MALDI-TOF) to identification of surface-exposed (Cullen et al. 2005) and OM-associated proteins (Cullen et al. 2002; Nally et al. 2005b). More is now known about the absolute level of expression of leptospiral proteins than in almost any other bacterial species, thanks to the proteome-wide application of MALDI-TOF to identify and quantify leptospiral proteins (Malmström et al. 2009). Absolute quantification was achieved by inclusion of isotope-labeled reference peptides in leptospiral samples. DNA microarrays have been used to examine the response of leptospiral transcript levels to environmental signals including temperature upshift (Lo et al. 2006), osmolarity (Matsunaga et al. 2007a), iron levels (Lo et al. 2010), serum (Patarakul et al. 2010), and macrophage-derived cells (Xue et al. 2010). Proteomic methods have also been used to examine thermoregulation (Lo et al. 2009) and posttranslational modification of OMPs (Cao et al. 2010; Eshghi et al. 2012). Proteome arrays have been used to identify fibronectin-binding OMPs (Pinne et al. 2012) and seroreactive OMPs (Lessa-Aquino et al. 2013).
2.2 Lipoprotein OMPs
Bacterial lipoproteins are proteins that have been posttranslationally modified by fatty acids (i.e., lipids) at a cysteine residue. This cysteine becomes the amino-terminal residue after the signal peptide has been removed by lipoprotein signal peptidase. Because the fatty acids of lipoproteins are extremely hydrophobic, they become embedded into membrane lipid bilayers and provide an anchor for lipoproteins to be tightly associated with the membrane. Treatments such as salt and urea that remove peripheral membrane proteins from membranes will not remove lipoproteins. This demonstrates that even though lipoproteins are generally not transmembrane proteins (Loa22 appears to be an exception), lipoproteins remain tightly associated with membranes, even after treatment with reagents that remove peripheral membrane proteins (Matsunaga et al. 2002). In contrast to the hydrophobicity of fatty acids, the protein components of most lipoproteins are typically hydrophilic and relatively soluble in aqueous buffers when expressed as recombinant proteins without their signal peptide. As such, the protein components of lipoproteins project out from membranes and decorate their surfaces. The first bacterial lipoprotein to be described was the Murein (or Braun’s) lipoprotein, which is integrated into the inner leaflet of the E. coli OM by lipids at its amino terminus and covalently attached to peptidoglycan (PG) at its carboxy-terminal lysine. Murein lipoprotein is a major OM protein of E. coli and serves as an important structural role in maintaining cellular integrity by providing a link between the OM and the PG cell wall. The OM-PG linkage is so important that E. coli has a number of other proteins that play similar roles, including OmpA and Pal (peptidoglycan-associated lipoprotein). Leptospires also have a number of OmpA-related proteins, such as Loa22, that are presumed to play similar OM-anchoring roles.
2.3 Lipoprotein Lipidation and Export
The steps involved in lipoprotein lipidation and export are shown in Fig. 3. Proteins with amino-terminal signal peptides, including lipoproteins, are exported across the inner membrane by the Sec translocase complex. Orthologs of all essential components of the Sec translocase complex are present in Leptospira. Upon reaching the periplasm, lipoproteins of Gram-negative bacteria are processed by a series of three enzymes that remove the signal peptide and modify the new N-terminal cysteine with fatty acids. Each of these three lipoprotein processing enzymes is also present in Leptospira (Haake 2000; Nascimento et al. 2004). The first of these enzymes is lipoprotein diacylglyceryl transferase (Lgt), which attaches a diacyl group containing two fatty acids to the sulfhydryl residue of cysteine via a thioester linkage. Because Lgt is the first enzyme in the series, its active site is presumably responsible for identifying the “lipobox ,” which distinguishes the signal peptides of lipoproteins from those of other exported proteins. Lipoprotein signal peptidase (Lsp) is the second enzyme in the series, and is responsible for removing the signal peptide, so that cysteine becomes the N-terminal amino acid of the mature lipoprotein. The third enzyme in the series is lipoprotein N-acyl transferase (Lnt), which adds a third and final fatty acid to the now available amino residue of cysteine via an amide bond. Interestingly, most Gram-positive bacteria lack Lnt and their lipoproteins are usually diacylated rather than triacylated (Kovacs-Simon et al. 2011). Perhaps Gram-negative bacteria triacylate their lipoproteins to strengthen the connection between the OM lipid bilayer and lipoproteins involved cell wall anchoring.
Experimental verification of lipidation is important when examining how lipoproteins interact with leptospiral membranes and the host innate immune system. Several methods are available. A commonly used method is to add radiolabeled palmitate to growth medium to demonstrate incorporation of label into the protein, which can be purified by immunoprecipitation. [14C]palmitate labeled at each of its carbons is the preferred form of palmitate because the higher specific activity results in a much shorter time to identification of bands by autoradiography. It should be noted that spirochetes, including leptospires, can digest fatty acids to two-carbon fragments which are incorporated into amino acid biosynthetic pathways. In this way, [14C]palmitate could potentially label any protein. For this reason, it is desirable to take advantage of the acid-labile linkage of the palmitate to the lipoprotein by demonstrating that the label can be removed from the lipoprotein by treatment of the electrophoresis gel containing the protein with acetic acid: immunoblots would remain positive, while the autoradiogram would become negative. Historically, globomycin has been used to inhibit lipidation of lipoproteins. However, it should be noted that globomycin selectively inhibits lipoprotein signal peptidase, the second enzyme in the series, so proteins could still become acylated through the previous step mediated by Lgt. Indirect evidence for lipidation can be obtained by Triton X-114 detergent fractionation. This detergent is similar to Triton X-100 except that Triton X-114 has a shorter polyethylene side chain, giving Triton X-114 a much lower cloud point. As a result, Triton X-114 solutions that occur in a single phase at 4 °C partition into two phases upon warming to 37 °C: a heavier, detergent-rich “hydrophobic” phase and a lighter, detergent-poor “hydrophilic” phase. Lipoproteins extracted by treatment of bacteria with Triton X-114 on ice should partition into the hydrophobic phase. The combination of sequence analysis plus behavior in Triton X-114 is an argument, albeit indirect, for lipidation.
Significant progress has been made in predicting which leptospiral genes encode lipoproteins. Lipoprotein signal peptides differ from other signal peptides in that they contain a “lipobox” sequence near the carboxy-terminal region of the signal peptide. In E. coli, the lipobox sequence is typically Leu-Leu-X-Y-Cys. There is relatively little variation in E. coli lipobox sequences and substitutions that do occur are with conservative amino acids: X is typically Ala, but can also be Thr or Ser, while Y is typically Gly, but can also be Ala. Based on sequences from experimentally verified lipoproteins, spirochete lipobox sequences are much more variable than those of E. coli. As a consequence, the Psort lipoprotein prediction program, based on lipoprotein sequences of E. coli and related bacteria has low (17–33 %) sensitivity for spirochetal lipoproteins. A later algorithm, LipoP, that utilized hidden Markov model statistical methods had significantly greater (50–81 %) sensitivity, but also higher (8–30 %) false positivity. To address these problems, we developed a spirochete-specific lipoprotein algorithm called “SpLip” based on sequences from 28 experimentally verified spirochetal lipoproteins. The SpLip algorithm is a hybrid approach of supplementing weight matrix scoring with rules including exclusion of charged amino acids from the lipobox (Setubal et al. 2006).
The inclusion in SpLip of rules based on sequences of experimentally verified lipoproteins reduces false positive hits, but these rules require further validation. For example, one of the SpLip rules is that the only allowed amino acids at the −1 position are Ala, Gly, Ser, Thr, Asn, Gln, or Cys. Now that a large number of leptospiral genomes have been sequenced, researchers at the J. Craig Venter Institute have discovered that newly sequenced homologs of known lipoproteins may have additional amino acids at the −1 position (Daniel Haft, personal communication). This genome sequence analysis will provide much greater confidence regarding the plasticity of amino acids at positions within the lipobox. It is unclear why spirochetal lipobox sequences are so much more variable than those of E. coli. One possible explanation for this difference is that E. coli growth rates are so much faster than spirochete growth rates. As a result, E. coli enzymes, including those involved in lipoprotein processing, must have much higher rates of catalysis than spirochetal enzymes. Higher catalytic rates may require higher substrate fidelity to maintain enzymatic efficiency.
After lipidation occurs, lipoproteins either remain in the outer leaflet of the inner membrane (IM) or undergo trafficking to one or more of four other possible destinations. From inside to outside these are: the inner leaflet of the OM, the outer leaflet of the OM, as a peripheral OM protein, and secretion beyond the cell. In E coli, lipoproteins destined for the OM are recognized by the IM ABC transporter-like sortase complex LolCDE (Yakushi et al. 2000) and then presented to periplasmic lipoprotein-binding chaperone LolA (Yokota et al. 1999) for transport to the OM lipoprotein receptor LolB (Yokota et al. 1999). The L. interrogans serovar Copenhageni genome appears to contain multiple homologs of a LolADE subset (LolA-1 and -2, LolD-1 and -2 and LolE-1, -2, and -3) (Fig. 4) (Nascimento et al. 2004). A LolC homolog is missing, as is LolB. LolB homologs so far have only been detected in β- and γ-Proteobacteria, and LolC function might be provided by one of the LolE homologs. While it remains to be determined which of the spirochetal genes are indeed functional Lol orthologs or functionally diverse paralogs, it appears likely that the Lol pathway is involved in shuttling lipoproteins from the IM to the OM.

Leptospiral lipoprotein export. Lipoproteins are exported via the Sec pathway (Step 1) from the cytoplasm to the periplasmic leaflet of the inner membrane PLIM where they are lipidated (Step 2). After lipidation, export to the periplasmic leaflet of the outer membrane PLOM occurs via the Lol pathway (Step 3a). Export to the surface leaflet of the outer membrane SLOM could occur either by the Type II Secretion System (T2SS, Step 3b) or by a lipoprotein flippase
Even less is known about how lipoproteins travel to the leptospiral surface and beyond. However, it would not be surprising if leptospiral lipoproteins follow the model established for B. burgdorferi lipoprotein secretion. In this model, the targeting information of surface lipoproteins was found to be located in the intrinsically disordered N-terminal tether peptides, with sorting signals differing from those in other well-characterized diderm model systems (Kumru et al. 2010, 2011; Schulze and Zückert 2006; Schulze et al. 2010). A surface lipoprotein’s periplasmic conformation (or lack thereof) was found to determine its ability to cross the OM, and crossing could be initiated by a disordered C-terminus after insertion of the protein to the periplasmic leaflet of the OM (Chen and Zückert 2011; Schulze et al. 2010). Shown in the context of cell envelope biogenesis (Fig. 4), our evolving working model of the leptospiral lipoprotein transport pathway consists of LolCDE and LolA orthologs being mainly involved in periplasmic sorting, while surface lipoproteins use two additional, so far uncharacterized modules to facilitate translocation, or “flipping,” of surface lipoproteins through the OM: (i) a periplasmic surface lipoprotein “holding” chaperone functioning like the chaperones guiding transmembrane proteins (TMPs) to the OM (Bos et al. 2007), and (ii) an OM lipoprotein translocase complex functioning similarly to lipid flippases (Pomorski and Menon 2006) (Fig. 4).
Alternative routes to the leptospiral surface are possible. Sec-dependent or -independent bacterial protein secretion pathways in Leptospira are limited to a type 2 secretion system, which might be involved in lipoprotein secretion tracking the Klebsiella model (d’Enfert et al. 1987; Sauvonnet and Pugsley 1996) (Fig. 4), a twin-arginine translocation (Tat) system, which may provide for export of folded proteins from the cytoplasm to the periplasm (Lee et al. 2006), and a type I secretion system (discussed in Sect. 2.14).
2.4 LipL32, the Major Outer Membrane Lipoprotein
As is strikingly apparent in protein stains of whole bacteria fractionated by SDS-PAGE, LipL32 is the most abundant protein in pathogenic Leptospira spp. Localization of LipL32 to the OM was demonstrated by Triton X-114 fractionation (Haake et al. 2000) and isolation of leptospiral OM vesicles by sucrose density gradient ultracentrifugation (Haake and Matsunaga 2002; Nally et al. 2005b). One of the early challenges encountered in these studies was that solubilization with Triton X-114 in the presence of EDTA results in degradation of LipL32 and other leptospiral membrane proteins by endogenous protease(s) when the detergent extract is warmed from 4 to 37 °C. This challenge was overcome by Zuerner et al. (1991) who found that addition of calcium prior to warming the extract prevented LipL32 degradation. The relationship between LipL32 and calcium was further elucidated by the LipL32 crystal structure, which revealed an acidic pocket formed in part by an extraordinary region of the LipL32 sequence in which seven out of eight amino acids are aspartates (Vivian et al. 2009). Co-crystalization of LipL32 with calcium showed that two of these aspartates are involved in calcium ion coordination (Hauk et al. 2009). When the aspartates in the calcium-binding pocket were mutated to alanines, denaturation of LipL32 in response to heat was similar with or without calcium. This elegant study used circular dichroism and tryptophan fluorescence to show that calcium helps LipL32 resist thermal denaturation (Hauk et al. 2012).
The abundance of LipL32 contributed greatly to its unfortunate misidentification as a surface lipoprotein. The first studies to claim LipL32 surface localization involved surface biotinylation experiments (Cullen et al. 2003, 2005). This technique involves addition of the biotinylation reagent sulfo-NHS-LC-biotin, which is considered to be “membrane impermeable” if membranes are intact but is a small enough molecule to penetrate through damaged membranes. This issue is problematic for spirochetes for which the OM is fragile and subject to disruption if organisms are not handled carefully. In this context, it is worth noting that in the second of these biotinylation studies, the cytoplasmic protein GroEL and the periplasmic protein FlaB1 were also found to be biotinylated (Cullen et al. 2005). Surface immunoelectron microscopy studies with LipL32 antibodies showed increased labeling of leptospiral cells compared to control antibodies. However, the number of gold particles (10.8 particles per cell) was far below what would have been expected for such an abundant protein. Seemingly confirmatory whole cell ELISA studies added to the confusion. The LipL32 surface protein dogma was recently overturned when more careful surface immunofluorescence studies were performed, including a number of controls including antisera to positive and negative control antigens and comparisons of intact and methanol fixed organisms (Pinne and Haake 2013). Studies were performed in parallel on intact and fixed organisms and fluorescence microscopy images were obtained using identical exposure times to ensure that they were truly comparable. LipL32 immunofluorescence of intact organisms was mostly negative, but occasionally showed irregular staining patterns, particularly if organisms were disrupted by shear force. However, the homogeneous staining observed with methanol fixed organisms did not occur with intact organisms. These immunofluorescence studies were supplemented with surface proteolysis studies showing that treatment with Proteinase K could digest LipL32 only if organisms were disrupted. Treatment of organisms with Proteinase K had previously been shown to be a reliable method for identifying surface proteins (Pinne and Haake 2009). In conclusion, LipL32 appears to be located in the periplasmic leaflet of the OM, a location shared by LipL36 (Shang et al. 1996). Based on Triton X-114 fractionation, other subsurface lipoproteins including LipL31 (Haake and Matsunaga 2002) and LruB (Verma et al. 2005) appear to be restricted to the periplasmic leaflet of the inner membrane. Interestingly, despite its location, LruA (also known as LipL71) modulates interactions with mammalian apolipoprotein A-I (Zhang et al. 2013).
If LipL32 is not on the leptospiral surface to any significant extent, what is its function? This is an important question given that pathogenic leptospires devote such a large amount of their protein-synthetic resources to expression of LipL32. Based on the Triton X-114 and OM vesicle evidence that LipL32 is an OM protein, it would be logical to conclude that the protein is located in the inner leaflet of the OM. Considering the known size of a LipL32 molecule (29 Å × 50 Å) as determined crystallographically (Vivian et al. 2009), the average length (10 µM) and diameter (0.1 µM) of leptospiral cells, and the quantitation of 38,000 copies of LipL32 molecules per cell (Malmström et al. 2009), it can be estimated that LipL32 occupies an extraordinary 20 % of the leptospiral OM inner surface. Perhaps LipL32 serves some structural role, for example, in OM stabilization? One possible function is as a calcium sink. Calcium is well-known to be important for membrane integrity generally and chelation of divalent cations with EDTA is essential for release of the OM from leptospiral cells (Haake et al. 1991; Haake and Matsunaga 2002; Nally et al. 2005b). However, LipL32 does not appear to be essential for OM integrity, given that a Himar transposon mutant of L. interrogans serovar Manilae lacking LipL32 had normal morphology and growth rate compared to the wild type (Murray et al. 2009b).
Aside from serving as a large calcium sink for leptospiral cells, the function of LipL32 is not understood. There is strong evidence that LipL32 is expressed during infection, given that there is intense staining by immunohistochemistry for LipL32 in the kidneys of infected animals (Haake et al. 2000) and that LipL32 is one of the most dominant seroreactive antigens recognized during acute and convalescent leptospirosis (Lessa-Aquino et al. 2013). On the other hand, LipL32 is not essential for infection given that the lipL32 transposon mutant was able to cause acute, lethal infections in hamsters and chronic infections in rats that were indistinguishable from those caused by the wild-type organism (Murray et al. 2009b). Nevertheless, given the large amount of LipL32 expressed by pathogenic leptospires, this protein has the potential to play a critical role in stimulating the host inflammatory response during infection. Purified, native (and therefore lipidated) LipL32 stimulates an innate immune response through TLR 2 (Werts et al. 2001). Inflammation in the kidney, a major target organ during leptospirosis, is manifested by interstitial nephritis. LipL32 induces interstitial nephritis in kidney proximal tubule cells (Yang et al. 2002) and the inflammation induced by LipL32 is mediated by TLR 2 (Yang et al. 2006). For reasons that remain obscure, LipL32 is one of the most highly conserved leptospiral OMPs among pathogenic leptospires, suggesting that it might be a favorable vaccine target for induction of cross-protective immunity. However, results obtained by immunization with a large variety of different LipL32 constructs remain largely negative or at best indeterminate, which may be related in part to its subsurface location. Readers interested in more information on this subject and other aspects of LipL32 are referred to the excellent, recent review by Murray (2013).
2.5 Loa22 and Other OmpA-Like Proteins
The second most abundant OM protein is Loa22 (Malmström et al. 2009). While there remains some uncertainty as to whether Loa22 is a lipoprotein, it is covered here because of experimental evidence of lipidation: Expression of Loa22 in E. coli resulted in labeling with [3H]palmitate (Koizumi and Watanabe 2003). This result is somewhat surprising because of the unusual Loa22 lipobox: SFTLC. As mentioned above, virtually all amino acids found in the −1 position relative to cysteine have been relatively small amino acids, and we are unaware of any documentned examples of a large hydrophobic amino acid like leucine in that location. For this reason, Loa22 is not predicted to be a lipoprotein by the SpLip algorithm. However, it is predicted to be a lipoprotein by the LipoP algorithm. While the [3H]palmitate-labeling data should be considered more convincing than the bioinformatic data, they would have been more conclusive if the experiment also had been performed in L. interrogans and if the label had been shown to be acid labile.
Lipoprotein or not, Loa22 represents a conundrum because it is both surface exposed and binds peptidoglycan via a carboxy-terminal OmpA domain. OmpA domains are peptidoglycan binding domains found in proteins that, like OmpA, link membranes to the cell wall situated beneath the OM. In the case of Loa22, the OmpA domain begins at amino acid 111 and occupies more than half the protein. There are strong immunofluorescence data showing that Loa22 is surface exposed (Ristow et al. 2007). One possible explanation for these data is that Loa22, like E. coli murein lipoprotein, exists in both peptidoglycan-bound and -free forms. The peptidoglycan-free form of murein lipoprotein has been found to be surface exposed (Cowles et al. 2011). The second explanation is that in the 90 amino acid segment between the signal peptide and the OmpA domain, Loa22 crosses the OM at least once. This 90 amino acid segment is hydrophilic and lacks the amphipathic beta sheets typically found in transmembrane OM proteins. Instead, as shown in Fig. 5a, there is an alpha-helical stretch with a strongly hydrophobic region on one face of the helix. This suggests that Loa22 is similar to the E. coli OM lipoprotein Wza, which forms large channels for export of the high molecular weight capsular polysaccharides. Wza forms octamers (Fig. 5b) in which the hydrophobic faces of the Wza monomers interact with the hydrophobic interior of the OM, while the hydrophilic faces form the walls of the channel (Dong et al. 2006). Although the role of Loa22 in the OM remains uncertain, that role appears to be essential for virulence; a Himar transposon mutant lacking Loa22 expression was unable to cause lethal infections in hamsters and guinea pigs, although it was able to cause bacteremia and renal colonization (Ristow et al. 2007). It is interesting to note that a homolog of the loa22 gene with 56 % sequence identity is present in L. biflexa, indicating that just because a gene is present or not in leptospiral saprophytes does not predict whether it is likely to be required for virulence in leptospiral pathogens.

Loa22 as an alpha-helical transmembrane OM protein. Panel a shows a helical wheel for the putative alpha-helical transmembrane domain of Loa22. The collection of nonpolar residues on one face indicates that the transmembrane helix could be amphipathic. Panel b shows the monomeric and octameric forms of Wza, which serves as a model for how Loa22 crosses the OM. Reproduced from Dong et al. (2006)
Loa22 belongs to a family of seven leptospiral OmpA -like proteins. The other members of the family differ from Loa22 in multiple ways. They do not appear to be lipoproteins, they tend to be much larger proteins, and appear to be more typical transmembrane OM proteins along the lines of the E. coli version of OmpA. For example, LIC10050 has a signal peptidase 1 cleavage site and is predicted to be a 78-kD protein with 22 beta-sheet transmembrane segments. However, in all cases, leptospiral OmpA-like proteins are probably important in linking the OM to the peptidoglycan cell wall.
2.6 Outer Membrane Lipoprotein LipL41
LipL41 is the third most abundant OM lipoprotein (Malmström et al. 2009). Levels of lipL41 transcript (Matsui et al. 2012) and LipL41 protein (Cullen et al. 2002; Nally et al. 2001b) are remarkably unaffected by temperature, osmolarity, and other environmental factors. The stability of LipL41 expression is useful as a control when studying the effects of growth conditions on the expression of other genes and proteins. For example, LipL41 antiserum is frequently included in immunoblots to compare the loading of bacteria per lane (Matsunaga et al. 2013). Although it is treated as one, it would be incorrect to call lipL41 a “housekeeping gene” until more is known about its function. Although too preliminary to be conclusive, a clue to the function of LipL41 is that it was identified as a potential hemin-binding protein in hemin-agarose affinity chromatography (Asuthkar et al. 2007). King et al. (2013) were unable to confirm hemin-binding activity. However, a subsequent study documented a submicromolar hemin-LipL41 dissociation constant and identified amino acids involved in hemin binding (Lin et al. 2013). Interestingly, the same study found that LipL41 forms a supramolecular assembly consisting of 36 molecules (Lin et al. 2013).
The lipL41 gene is located immediately upstream of a smaller gene, with which it is co-transcribed. For this reason, the smaller gene has been designated lep for lipL41 expression partner (King et al. 2013). Even though lipL41 transcript levels were unaffected in a lep transposon mutant, LipL41 levels were greatly reduced. Because Lep expression appeared to be required for stable expression of LipL41, perhaps by acting as a chaperone, researchers examined whether Lep bound to LipL41. Lep molecules were found to bind to LipL41 molecules at a molar ratio of 2:1 (King et al. 2013). Neither a lipL41 nor a lep mutant was attenuated for virulence in hamsters. Interestingly, Lep was not detected by whole organism MALDI-TOF (Malmström et al. 2009), indicating that Lep is only required in small amounts transiently during export of LipL41 to the OM.
2.7 The Lig Family of OM Lipoproteins
The Lig family of OM lipoproteins was discovered by screening L. kirschneri and L. interrogans expression libraries with convalescent human leptospirosis sera. This approach identified GroEL, and DnaK, and LipL41, and three novel genes encoding a series of bacterial immunoglobulin (Ig)-like domains. The proteins encoded by these novel genes were designated as Leptospiral Ig-like proteins LigA, LigB, and LigC (Matsunaga et al. 2003). The Lig proteins consist of a lipoprotein signal peptide followed by a series of 12–13 Ig-like domains and, in the case of LigB and LigC, a large carboxy-terminal domain. The region upstream of ligA and ligB, as well as the first six Ig-like domains of LigA and LigB, are virtually identical, indicating that the ligA gene resulted from a partial gene duplication event. This event likely occurred relatively late in leptospiral evolution, as ligA is found only found in stains of L. kirschneri and L. interrogans (McBride et al. 2009). In contrast, LigB is found in all pathogenic Leptospira species. LigC is also widely distributed but is a pseudogene or absent in some strains. Sequence comparison revealed a surprising degree of mosaicism, indicating genetic rearrangements involving ligB gene fragments of L. interrogans and L. kirschneri (McBride et al. 2009). OMP mosaicism can confer a survival advantage in the face of antigenic pressure.
Temperature and osmolarity are key environmental signals that control the expression of the Lig proteins. In the process of examining the interaction of leptospires with cells in tissue culture, Matsunaga et al. (2005) observed that the addition of EMEM tissue culture medium to leptospiral culture medium induced LigA and LigB expression and caused a substantial increase in released LigA. Sodium chloride was primarily responsible for these effects. All other EMEM components, including iron, bicarbonate, and oxygen concentrations, had no effect on Lig expression. As shown in Fig. 6, addition of sodium chloride, potassium chloride, or sodium sulfate to leptospiral medium (EMJH) to the level of osmolarity found in the mammalian host (~300 mOsm/L) induced expression of both cell-associated LigA and LigB, and release of LigA into the culture supernatant. Osmolarity affects both lig transcript and Lig protein levels (Matsunaga et al. 2007b). In addition to its effects on Lig protein expression, osmolarity increases the transcription of the leptospiral sphingomyelinase, Sph2 (Matsunaga et al. 2007b), the putative adhesin, LipL53 (Oliveira et al. 2010), and a number leptospiral lipoproteins and OMPs (Matsunaga et al. 2007a). These results suggest that leptospires upregulate a defined set of OMPs when they encounter mammalian host tissues and sense an increase in osmolarity. The sensory transduction proteins involved in osmoregulation have not yet been defined.

Induction of Lig expression by osmolarity. Expression of LigA and LigB is strongly induced by addition of salt to Ellinghausen-McCullough-Johnson-Harris EMJH medium. LigA is found in both the cellular c and supernatant s fractions. A variety of salts are effective, indicating that induction of Lig expression is mediated by osmolarity rather than any particular salt component
More recently, it was discovered that expression of the lig genes is also regulated by temperature. The long 175 nucleotide 5′ untranslated region is predicted to contain secondary structure that includes and obscures the ribosome binding site and start codon, preventing binding to the ribosome and initiation of translation (Fig. 7). Toeprint experiments showed binding of ribosomes to the lig transcript was poor unless most of the left stem of predicted structure 2 (Fig. 7) was removed. In E. coli, a lig′-′bgaB translational fusion transcribed from a heterologous promoter was regulated by temperature, demonstrating the ability of the lig sequences to exert posttranscriptional control by temperature. Mutations on the left or right stem of structure 2 partially relieved inhibition of β-galactosidase expression; inhibition was regained when the mutations were combined to restore base pairing, providing evidence that base-paired RNA is a component of the inhibitory element. These results are consistent with a model in which structure 2 functions as a thermolabile “thermometer,” transacting factors may also have a dominant role in melting the inhibitory stem.

Secondary structure of the 5′ untranslated region of the lig genes. The mRNA of the lig genes has an unusually long 5′ untranslated region which is predicted to form two stem-loop structures. Structure 2 obscures the ribosome binding site (SD) and start codon and must be unfolded for translation to occur
The upregulation of LigA and LigB by osmolarity and temperature suggests that these proteins are expressed early during mammalian host infection and may be involved in critical bacterial-host interactions. Various lines of evidence support these conclusions. Patients with leptospirosis have a strong antibody response to the Lig Ig-like repeat domains, suggesting that recombinant Lig repeats would be useful serodiagnostic antigens, confirming that Lig proteins are expressed during infection (Croda et al. 2007). Lig proteins are expressed on the leptospiral surface based on immunoelectron microscopy (Matsunaga et al. 2003) and LigA is released from leptospiral cells (Matsunaga et al. 2005). Osmotic induction of Lig expression resulted in L. interrogans becoming more “sticky,” with increased adherence to several different extracellular matrix proteins, including fibronectin, fibrinogen, and collagens I and IV (Choy et al. 2007). Heterologous expression of LigA and LigB in L. biflexa increased adherence to eukaryotic cells and fibronectin (Figueira et al. 2011). We advocate this “gain of function” approach when studying potential leptospiral adhesins as a way to evaluate the significance of protein-protein interaction assays. LigB binds more avidly to fibronectin and fibrinogen than LigA and the LigB binding activity was localized to 3 of the 12 LigB Ig-like domains; domains 9–11 were both necessary and sufficient to reproduce the binding activity of LigB (Choy et al. 2011). A remarkable aspect of these studies is the range of different proteins to which LigB is able to bind with high avidity. LigB not only binds to complement components and the complement regulatory protein, Factor H, but also inhibits complement activity (Castiblanco-Valencia et al. 2012; Choy 2012). These results suggest that a role of LigB is to coat the leptospiral surface with a variety of circulating host proteins and protect leptospires from host defense mechanisms.
Leptospiral vaccines are discussed in Chapter by Ben Adler, this volume, while their use in humans and animals is described in Chapters by D.A. Haake and P.N. Levett and by W.A. Ellis, this volume. Nevertheless, it should be mentioned in this context that when L. interrogans sv Copenhageni is the challenge strain, immunization of hamsters with LigA converts a lethal infection into sublethal kidney colonization. The initial studies showed that the unique part of LigA (Ig-like domains 7–13) was most effective as a vaccine (Silva et al. 2007). Subsequent studies localized the Ig-like domains involved in immunoprotection (Coutinho et al. 2011). There was an absolute requirement for LigA domains 11 and 12. However, these two domains were not sufficient for immunoprotection; a third, flanking domain (either domain 10 or 13) was needed. This requirement for three contiguous Ig-like domains near the carboxy-terminal end of the molecule is highly reminiscent of the finding that LigB domains 9–11 are required for binding activity (see previous paragraph). LigA immunization is effective not only when injected subcutaneously as a purified, recombinant protein, but also when expressed in a lipidated form in E. coli that is administered orally (Lourdault et al. 2014). Some important caveats are in order. LigA does not provide sterilizing immunity and because the immunoprotective region of the LigA molecule is subject to variation (McBride et al. 2009), cross-protective immunity may be limited. Additionally, no homologous protection was elicited following immunization of hamsters with LigA from L. interrogans serovars Manilae (Deveson Lucas et al. 2011) or Canicola (N. Bomchil, personal communication). An important goal of future studies is to understand why LigA appears to protect against challenge by some serovars but not others.
2.8 More Outer Membrane Lipoproteins
As summarized in Table 1, quantitative MALDI-TOF data reveal that after LipL32, Loa22, and LipL41, the next most abundant OM lipoproteins are LipL36, LipL21, and LipL46 (Malmström et al. 2009). Although LipL36 is an OM protein, it is not surface exposed, being restricted to the inner leaflet of the OM (Haake et al. 1998). Based on serological evidence (Haake et al. 1998), immunohistochemistry data (Barnett et al. 1999) and downregulation of LipL36 expression at physiologic osmolarity (Matsunaga et al. 2007a, b), LipL36 appears to be expressed only when leptospires are outside the mammalian host. In contrast to LipL36, LipL21 and LipL46 are both surface-exposed and expressed during infection (Cullen et al. 2003, Matsunaga et al. 2006). While not quite as abundant as originally thought, LipL21 is highly expressed during infection based on immunoblot analysis of organisms harvested from infected guinea pigs (Nally et al. 2007) and immunohistochemistry of liver from infected hamsters (Eshghi et al. 2009). LipL46 can also be detected immunohistochemically in a variety of organs during infection (Matsunaga et al. 2006).
A fundamental difference between leptospiral saprophytes and pathogens is that saprophytes are serum sensitive while pathogens are serum resistant. A common serum resistance mechanism shared by many bacterial pathogens is binding the complement regulators factor H and factor H protein-1. Using a ligand blot approach, L. interrogans was found to have two factor H-binding proteins with molecular masses of 25- and 50-kDa. The 25-kDa factor H-binding protein was initialy referred to as LfhA (leptospiral factor H-binding protein) (Verma et al. 2006). A subsequent study identified the same protein as a laminin binding adhesin and applied the designation Lsa24 (leptospiral surface adhesin 24-kD) (Barbosa et al. 2006). Structural analysis revealed that LfhA/Lsa24 was a member of a family of six leptospiral adhesins that share structural similarities with endostatin (Stevenson et al. 2007). For this reason, LfhA/Lsa24 was renamed LenA. In addition to binding Factor H, LenA was subsequently found to bind plasminogen (Verma et al. 2010). Binding of LenA to plasminogen facilitated conversion to plasmin, which in turn degraded fibrinogen, suggesting a role for LenA in penetration through, and/or escape from, fibrin clots. Several other leptospiral OMPs have also been implicated in plasminogen binding and activation (Fernandes et al. 2012; Vieira et al. 2012).
2.9 Transmembrane Outer Membrane Proteins
Transmembrane OMPs are defined as integral OM proteins that contain strands that traverse the lipid bilayer of the OM. Such proteins can be visualized by freeze-fracture electron microscopy (FFEM), a technique that separates the two leaflets of membranes, exposing transmembrane OMPs as studs in a sea of lipid. When applied to spirochetes, FFEM revealed that pathogenic spirochetes, including leptospires, have transmembrane OMPs in far fewer numbers than typical Gram-negative bacteria (Haake et al. 1991; Radolf et al. 1989; Walker et al. 1991). Transmembrane OMPs are essential for OM-containing bacteria because of their unique ability to form pores or channels that allow bacteria to acquire nutrients and to export toxins and waste products. For researchers interested in bacterial surface antigens, transmembrane OMPs are of great interest because their surface-exposed loops represent potential targets of a protective immune response.
Transmembrane OMPs have an amino-terminal signal peptide, which facilitates their secretion across the inner membrane to the periplasm by the Sec translocase complex. After removal of the signal peptide by signal peptidase I, transmembrane OMPs are shuttled across the periplasm to the OM by the chaperone SurA (Sklar et al. 2007). LIC12922 of L. interrogans serovar Copenhageni has been identified by X-ray crystallography to have both the parvulin and peptide-binding domains of SurA (Giuseppe et al. 2011). The peptide-binding domain allows SurA to keep transmembrane OMPs in an unfolded form until they are delivered to the OMP assembly complex, which consists of the transmembrane OMP, BamA, and several accessory lipoproteins. L. interrogans has a BamA homologue with four POTRA domains that are involved in the folding, assembly and insertion of transmembrane OMPs in the OM (Tommassen 2007).
2.10 Discovery of the Porin OmpL1
OmpL1 was one of the first porins to be described in a spirochete, preceded only by the 36.5 kD porin of Spirochaeta aurantia (Kropinski et al. 1987). The discovery of OmpL1 resulted from experiments aimed at identifying surface-exposed OMPs. Using a technique called “surface immunoprecipitation,” antibodies raised to whole L. kirschneri bacteria were added to intact bacteria followed by gentle washing to remove unbound antibodies. The antibody-antigen complexes were solubilized using Triton X-100 detergent and then purified using Protein A beads. In addition to LPS, the surface immunoprecipitate was found to contain three proteins with molecular masses of 33-, 41-, and 44-kD (Haake et al. 1991). The amount of the 33-kD protein was increased in a highly passaged strain of L. kirchneri, correlating with the density of transmembrane particles visualized by FFEM. Isolation of the gene encoding the 33-kD protein revealed a series of porin-like transmembrane segments (see next section), and henceforth the protein was called OmpL1 (Haake et al. 1993). The other two proteins were subsequently identified as LipL41 and LipL46. Confirming its role in the leptospiral OM, OmpL1 was later found to have several other properties typical of porins, including: 1. Heat-modifiable electrophoretic mobility; 2. Cross-linkable trimers; and 3. The ability to form channels in lipid bilayers (Shang et al. 1995).
Bacterial porins are of great interest because of their surface exposure and potential to serve as targets of a protective immune response. Like most porins, OmpL1 is hydrophobic and requires detergent for solubilization. Recombinant OmpL1 expressed in E. coli with a His6 tag can be purified by nickel chromatography under denaturing conditions. Unfortunately, this denatured form of OmpL1 proved to be ineffective as a vaccine (unpublished results). However, when hamsters were immunized with OmpL1 expressed in E. coli as a membrane protein, this resulted in partial protection from lethal and sublethal infection, particularly when combined with a lipidated form of LipL41 (Haake et al. 1999). The ompL1 gene is present and moderately well conserved (~90 % deduced amino acid sequence identity) across a broad range of pathogenic Leptospira species. Interestingly, comparison of sequences from a number of Leptospira strains revealed that 20 % of strains carried mosaic ompL1 genes composed of segments with multiple leptospiral ancestries arising from horizontal DNA transfer and genetic recombination (Haake et al. 2004). These sequence variations, of course, could limit cross-protection from an OmpL1-based vaccine. Other leptospiral genes that have been found to undergo mosaicism include ligA and ligB (McBride et al. 2009).
2.11 Beta-Barrel Structure of Transmembrane OMPs
As mentioned in the previous section, OmpL1 has a series of transmembrane segments characteristic of channel-forming porins. The transmembrane segments of a number of OMPs from a variety of Gram-negative bacteria have been determined by X-ray crystallography to have a beta-sheet conformation, such that the orientation of amino acid side chains is 180° opposite of those of adjacent amino acids. This allows the side chains of alternating amino acids to interface with the lipid bilayer or with the aqueous pore of the channel. As these transmembrane segments thread their way back and forth across the lipid bilayer, they form the walls of a cylinder or barrel, and such proteins are called “beta barrels.” The beta-sheet conformation in these transmembrane strands is the basis for transmembrane OMP prediction programs such as TMBB-PRED (Bagos et al. 2004) and TMBETA-NET (Gromiha and Suwa 2005).
Screening of the L. interrogans serovar Copenhageni genome for OMPs by querying the TMBB-PRED webserver revealed 84 genes that met the relatively stringent cutoff score of ≤2.965. As a positive control, the TMBB-PRED algorithm gave OmpL1 a score of 2.900, the sixth best score of any leptospiral protein. A useful feature of TMBB-PRED is that the output includes a plot of the probability of transmembrane membrane beta-strands. As shown in Fig. 8, the TonB-dependent receptor, HbpA received a score of 2.939 and was predicted to have 22 transmembrane beta strands. Using homology-based annotation and sequence-based criteria (signal peptide + ≤3 alpha helices + ≥6 transmembrane beta strands) a list of 184 possible transmembrane OMPs was derived (Pinne et al. 2012). These putative transmembrane OMPs and 177 predicted lipoproteins were expressed by in vitro transcription/translation to construct an OMP proteome array to screen for adherence to fibronectin. 14 novel leptospiral fibronectin-binding proteins were identified, including Lsa66, a previously identified OmpA-like adhesin (Oliveira et al. 2011). Adherence function was confirmed by expression of proteins in L. biflexa, conferring dramatically increased fibronectin-binding activity on this surrogate host.

Topology of TonB-dependent receptor HbpA. Hemin-binding protein A (HbpA, LIC20151) is predicted to have a PLUG domain and a TonB-Dependent Receptor TBDR domain. The PLUG domain sits inside the beta-barrel formed by the TBDR domain, reproduced from Oke et al. (2004). The beta-barrel structure is predicted using the TMBB-PRED algorithm
2.12 Experimental Validation of Transmembrane OMPs
A new paradigm has emerged for experimental confirmation of transmembrane OMPs. Originally, Triton X-114 detergent extraction and phase partitioning was thought to be a more or less definitive test for localization of leptospiral proteins (Haake et al. 1991). OMPs were expected to be found, in whole or in part, in the Triton X-114 detergent phase, while cytoplasmic and inner membrane proteins remained in the protoplasmic cylinder fraction and periplasmic proteins fractionated to the aqueous phase. Although many OM components, including LPS and many OMPs, were found in the Triton X-114 detergent phase, it is now clear that a number of transmembrane OMPs do not behave as expected in this detergent (Pinne and Haake 2009).
We now advocate a multistep strategy for defining transmembrane OMPs. The first step is sequence analysis. The sequence of transmembrane OMPs should begin with a signal peptide and signal peptidase I cleavage site but lack a lipobox. The sequence of the mature protein should contain multiple beta-sheet transmembrane segments (predicted using an algorithm such as TMBB-PRED) and should not contain a hydrophobic, membrane-spanning alpha helix. Of course, OMPs such as Loa22 with alpha-helical transmembrane domains are an exception to this rule. The second step is to test whether the protein is an integral membrane protein by treating total leptospiral membranes with reagents, such as high salt, urea, or sodium bicarbonate, that remove membrane-associated proteins (Matsunaga et al. 2002; Pinne and Haake 2009). The third step is to test for surface exposure. Conclusions should not be based on a single method. Several complementary methods are available: surface immunofluorescence, surface proteolysis, and surface biotinylation. In each of these methods, it is essential to include controls. In the case of surface immunofluorescence, control experiments with preimmune sera to show that antibody binding to the leptospiral surface is a result of immunization with the protein of interest must be included. In negative control experiments, it is important to counterstain the slide with DAPI (4′,6-diamidino-2-phenylindole) to show that organisms are present. OmpL1 as a positive control for surface exposure and the endoflagellar protein FlaA1 as a subsurface control must be included. Relatively abundant periplasmic proteins such as FlaA1 are preferred as subsurface controls because these would more readily become surface exposed as a result of OM disruption than cytoplasmic proteins such as GroEL. Information about obtaining antisera for surface and subsurface control antigens is available on our website: http://id-ucla.org/sharing.php.
Using this strategy, four novel leptospiral transmembrane OMPs were defined: OmpL36, OmpL37, OmpL47, and OmpL54 (Pinne and Haake 2009). Each of these four proteins was found to have a signal peptide and signal peptidase I cleavage site and at least 6 membrane-spanning beta-strands. Although OmpL36 and OmpL37 were partially removed from total membrane fractions by sodium bicarbonate, none was removed by high salt or urea. All four proteins were found to be surface exposed by surface immunofluorescence, surface proteolysis, and surface biotinylation except for OmpL36, which was not digested by the highest concentration of proteinase K. It should be noted that OmpL47 (also known as Q8F8Q0) had previously been identified by surface biotinylation as a component of the leptospiral “surfaceome” (Cullen et al. 2005) and is annotated as a glycosyl hydrolase. The behavior of these proteins in Triton X-114 cell fractionation experiments was surprising in that only OmpL54 was found in the Triton X-114 detergent phase. OmpL36 was not extractable with Triton X-114 and was found entirely in the protoplasmic cylinder fraction, which is consistent with the subsequent finding that this protein is a flagellar component (Wunder et al. 2013). While OmpL37 and OmpL47 were partially or completely extracted with Triton X-114, these proteins fractionated into the aqueous phase rather than the detergent phase. These results suggest that localization by Triton X-114 fractionation alone may be unreliable for some types of proteins, especially transmembrane OMPs.
2.13 OMPs Involved in Import Pathways
Pathogenic and saprophytic leptospires appear to have a full complement of TonB-dependent receptors (TB-DRs). TB-DRs are beta-barrel OMPs that function as high affinity receptors and channels for uptake of substrates such as vitamin B12 (cobalamin), iron, and other heavy metals. Uptake is energy- dependent and requires interactions between TB-DRs in the OM and TonB in the IM. L. interrogans has 12 genes encoding TB-DRs and 3 genes encoding TonB. Thanks to the elegant work of Picardeau and colleagues on TB-DRs of L. biflexa, the function of several leptospiral TB-DRs is now known (Louvel et al. 2006). For example, the L. biflexa mutant lacking gene LEPBIa2760 was unable to grow on the siderophore desferrioximine as a source of iron, thereby indicating that this gene encodes the siderophore uptake receptor CirA. Because many TB-DRs are highly conserved across leptospiral species, this information is relevant to pathogenic leptospires. The amino acid sequence of LEPBIa2760 is 77 % identical with that of LIC11694. Likewise, LEPBIa1883 and LIC10714 encode the Fe3+-dicitrate receptor FecA. As shown in Fig. 9, both LIC11694 (CirA) and LIC10714 (FecA) have paralogs that presumably perform similar, if not redundant, functions. LIC20151 has been shown to bind hemin, and represents a third TB-DR class. Three additional TB-DR classes remain to be characterized, but presumably are involved in uptake of vitamin B12, copper, or nickel (Schauer et al. 2008). Leptospires also have OM proteins involved in TonB-independent import pathways, such as FadL (LIC12524), the long-chain fatty acid transporter.

Relatedness tree for leptospiral TonB-dependent receptors TBDRs. The L. interrogans serovar Copenhageni strain L1-130 genome is predicted to contain 9 TBDR genes involved in uptake of vitamin B12, iron and other metals. The functions of the three TBDR genes that have been elucidated are shown
2.14 OMPs Involved in Export Pathways (TolC and GspD)
Leptospires have at least two different OMP-mediated export pathways: Type 1 secretion involving TolC and Type 2 secretion involving GspD. Type 1 secretion is Sec-independent, meaning that substrates can be exported directly from the cytoplasm. In the case of proteins (e.g., hemolysins), this means that a signal peptide is not required. Type I secretion can also be involved in efflux of drugs or toxins, such as heavy metals. TolC is the OMP component of the Type 1 secretory apparatus and forms a beta barrel channel in the OM and spans the periplasm to the IM where it engages with a translocase to form a contiguous passage from the cytoplasm to the exterior of the cell. L. interrogans encodes seven TolC homologs, presumably to accommodate different types of translocases and substrates. One of these TolC proteins, LIC12575, is expressed at high levels in cultivated cells. Type 2 secretion is Sec-dependent, meaning that proteins exported via this pathway must have a signal peptide and be secreted first to the periplasm before exiting the cell. As discussed above, Type 2 secretion represents a potential pathway for lipoprotein export in Leptospira species, as has been demonstrated in Klebsiella (d’Enfert et al. 1987; Sauvonnet and Pugsley 1996). Possible substrates include potential lipoproteins LigA and Sph2, which are released from L. interrogans in response to elevated osmolarity and/or temperature (Matsunaga et al. 2005, 2007b).
2.15 LipL45 and Related Peripheral Membrane Proteins
LipL45 was first identified as a protein, designated Qlp42, whose expression was upregulated when L. interrogans cultures were shifted from 30° to 37 °C (Nally et al. 2001a). Subsequent studies revealed that Qlp42 was initially expressed as a 45-kD lipoprotein, the carboxy-terminal portion of which was removed to become a 31-kD peripheral membrane protein, designated P31LipL45 (Matsunaga et al. 2002). Peripheral membrane proteins are membrane-associated proteins that are not integrated into the lipid bilayer and can be removed by treating membranes with a variety of reagents such as high salt, urea, or sodium bicarbonate. The latter two reagents removed P31LipL45 from L. interrogans membranes, but had no effect on LipL41. Interestingly, in addition to upregulation of expression at higher temperatures, P31LipL45 was dramatically increased in stationary phase cultures of L. interrogans. The function, membrane location(s), and surface exposure of P31LipL45 remain to be determined. Genome sequencing has revealed that LipL45 belongs to a large family of leptospiral proteins; L. interrogans has 11 LipL45-related genes, most of which are predicted to be lipoproteins. Although LipL45 itself is the most highly expressed member of the family in cultivated cells (Malmström et al. 2009), two other family members are expressed at comparable levels, which probably explains why P31LipL45 appears as a doublet in many strains of pathogenic leptospires (Matsunaga et al. 2002).
References
Asuthkar S, Velineni S, Stadimann J, Altmann F, Sritharan M (2007) Expression and characterization of an iron-regulated hemin-binding protein, HbpA, from Leptospira interrogans serovar Lai. Infect Immun 75:4582–4591
Bagos PG, Liakopoulos TD, Spyropoulos IC, Hamodrakas SJ (2004) PRED-TMBB: a web server for predicting the topology of beta-barrel outer membrane proteins. Nucleic Acids Res 32:W400–W404
Barbosa AS, Abreu PA, Neves FO, Atzingen MV, Watanabe MM, Vieira ML, Morais ZM, Vasconcellos SA, Nascimento AL (2006) A newly identified leptospiral adhesin mediates attachment to laminin. Infect Immun 74:6356–6364
Barnett JK, Barnett D, Bolin CA, Summers TA, Wagar EA, Cheville NF, Hartskeerl RA, Haake DA (1999) Expression and distribution of leptospiral outer membrane components during renal infection of hamsters. Infect Immun 67:853–861
Bos MP, Robert V, Tommassen J (2007) Biogenesis of the gram-negative bacterial outer membrane. Annu Rev Microbiol 61:191–214
Cao XJ, Dai J, Xu H, Nie S, Chang X, Hu BY, Sheng QH, Wang LS, Ning ZB, Li YX, Guo XK, Zhao GP, Zeng R (2010) High-coverage proteome analysis reveals the first insight of protein modification systems in the pathogenic spirochete Leptospira interrogans. Cell Res 20:197–210
Castiblanco-Valencia MM, Fraga TR, Silva LB, Monaris D, Abreu PA, Strobel S, Jozsi M, Isaac L, Barbosa AS (2012) Leptospiral immunoglobulin-like proteins interact with human complement regulators factor H, FHL-1, FHR-1, and C4BP. J Infect Dis 205:995–1004
Chen S, Zückert WR (2011) Probing the Borrelia burgdorferi surface lipoprotein secretion pathway using a conditionally folding protein domain. J Bacteriol 193:6724–6732
Choy HA (2012) Multiple activities of LigB potentiate virulence of Leptospira interrogans: inhibition of alternative and classical pathways of complement. PLoS ONE 7:e41566
Choy HA, Kelley MM, Chen TL, Moller AK, Matsunaga J, Haake DA (2007) Physiological osmotic induction of Leptospira interrogans adhesion: LigA and LigB bind extracellular matrix proteins and fibrinogen. Infect Immun 75:2441–2450
Choy HA, Kelley MM, Croda J, Matsunaga J, Babbitt JT, Ko AI, Picardeau M, Haake DA (2011) The multifunctional LigB adhesin binds homeostatic proteins with potential roles in cutaneous infection by pathogenic Leptospira interrogans. PLoS ONE 6:e16879
Coutinho ML, Choy HA, Haake D (2011) A LigA three-domain region protects hamsters from lethal infection by Leptospira interrogans. PLoS Neg Trop Dis 5:e1422
Cowles CE, Li Y, Semmelhack MF, Cristea IM, Silhavy TJ (2011) The free and bound forms of Lpp occupy distinct subcellular locations in Escherichia coli. Mol Microbiol 79:1168–1181
Croda J, Ramos JG, Matsunaga J, Queiroz A, Homma A, Riley LW, Haake DA, Reis MG, Ko AI (2007) Leptospira immunoglobulin-like proteins as a serodiagnostic marker for acute leptospirosis. J Clin Microbiol 45:1528–1534
Cullen PA, Cordwell SJ, Bulach DM, Haake DA, Adler B (2002) Global analysis of outer membrane proteins from Leptospira interrogans serovar Lai. Infect Immun 70:2311–2318
Cullen PA, Haake DA, Bulach DM, Zuerner RL, Adler B (2003) LipL21 is a novel surface-exposed lipoprotein of pathogenic Leptospira species. Infect Immun 71:2414–2421
Cullen PA, Xu X, Matsunaga J, Sanchez Y, Ko AI, Haake DA, Adler B (2005) Surfaceome of Leptospira spp. Infect Immun 73:4853–4863
d’Enfert C, Ryter A, Pugsley AP (1987) Cloning and expression in Escherichia coli of the Klebsiella pneumoniae genes for production, surface localization and secretion of the lipoprotein pullulanase. EMBO J 6:3531–3538
Deveson Lucas DS, Cullen PA, Lo M, Srikram A, Sermswan RW, Adler B (2011) Recombinant LipL32 and LigA from Leptospira are unable to stimulate protective immunity against leptospirosis in the hamster model. Vaccine 29:3413–3418
Dong C, Beis K, Nesper J, Brunkan-LaMontagne AL, Clarke BR, Whitfield C, Naismith JH (2006) Wza the translocon for E. coli capsular polysaccharides defines a new class of membrane protein. Nature 444:226–229
Eshghi A, Cullen PA, Cowen L, Zuerner RL, Cameron CE (2009) Global proteome analysis of Leptospira interrogans. J Proteome Res 8:4564–4578
Eshghi A, Pinne M, Haake DA, Zuerner RL, Frank A, Cameron CE (2012) Methylation and in vivo expression of the surface-exposed Leptospira interrogans outer membrane protein OmpL32. Microbiol 158:622–635
Farrelly HE, Adler B, Faine S (1987) Opsonic monoclonal antibodies against lipopolysaccharide antigens of Leptospira interrogans serovar hardjo. J Med Microbiol 23:1–7
Fernandes LG, Vieira ML, Kirchgatter K, Alves IJ, de Morais ZM, Vasconcellos SA, Romero EC, Nascimento AL (2012) OmpL1 is an extracellular matrix- and plasminogen-interacting protein of Leptospira spp. Infect Immun 80:3679–3692
Figueira CP, Croda J, Choy HA, Haake DA, Reis MG, Ko AI, Picardeau M (2011) Heterologous expression of pathogen-specific genes ligA and ligB in the saprophyte Leptospira biflexa confers enhanced adhesion to cultured cells and extracellular matrix components. BMC Microbiol 11:129
Giuseppe PO, Von Atzingen M, Nascimento AL, Zanchin NI, Guimaraes BG (2011) The crystal structure of the leptospiral hypothetical protein LIC12922 reveals homology with the periplasmic chaperone SurA. J Struct Biol 173:312–322
Gromiha MM, Suwa M (2005) A simple statistical method for discriminating outer membrane proteins with better accuracy. Bioinformatics 21:961–968
Haake DA (2000) Spirochaetal lipoproteins and pathogenesis. Microbiol 146:1491–1504
Haake DA, Matsunaga J (2002) Characterization of the leptospiral outer membrane and description of three novel leptospiral membrane proteins. Infect Immun 70:4936–4945
Haake DA, Walker EM, Blanco DR, Bolin CA, Miller MN, Lovett MA (1991) Changes in the surface of Leptospira interrogans serovar grippotyphosa during in vitro cultivation. Infect Immun 59:1131–1140
Haake DA, Champion CI, Martinich C, Shang ES, Blanco DR, Miller JN, Lovett MA (1993) Molecular cloning and sequence analysis of the gene encoding OmpL1, a transmembrane outer membrane protein of pathogenic Leptospira spp. J Bacteriol 175:4225–4234
Haake DA, Martinich C, Summers TA, Shang ES, Pruetz JD, McCoy AM, Mazel MK, Bolin CA (1998) Characterization of leptospiral outer membrane lipoprotein LipL36: downregulation associated with late-log-phase growth and mammalian infection. Infect Immun 66:1579–1587
Haake DA, Mazel MK, McCoy AM, Milward F, Chao G, Matsunaga J, Wagar EA (1999) Leptospiral outer membrane proteins OmpL1 and LipL41 exhibit synergistic immunoprotection. Infect Immun 67:6572–6582
Haake DA, Chao G, Zuerner RL, Barnett JK, Barnett D, Mazel M, Matsunaga J, Levett PN, Bolin CA (2000) The leptospiral major outer membrane protein LipL32 is a lipoprotein expressed during mammalian infection. Infect Immun 68:2276–2285
Haake DA, Suchard MA, Kelley MM, Dundoo M, Alt DP, Zuerner RL (2004) Molecular evolution and mosaicism of leptospiral outer membrane proteins involves horizontal DNA transfer. J Bacteriol 186:2818–2828
Hauk P, Guzzo CR, Roman Ramos H, Ho PL, Farah CS (2009) Structure and calcium-binding activity of LipL32, the major surface antigen of pathogenic Leptospira sp. J Mol Biol 390:722–736
Hauk P, Barbosa AS, Ho PL, Farah CS (2012) Calcium binding to leptospira outer membrane antigen LipL32 is not necessary for its interaction with plasma fibronectin, collagen type IV, and plasminogen. J Biol Chem 287:4826–4834
Jost BH, Adler B, Faine S (1989) Experimental immunisation of hamsters with lipopolysaccharide antigens of Leptospira interrogans. J Med Microbiol 29:115–120
King AM, Bartpho T, Sermswan RW, Bulach DM, Eshghi A, Picardeau M, Adler B, Murray GL (2013) Leptospiral outer membrane protein Lipl41 is not essential for acute leptospirosis but requires a small chaperone protein, Lep, for stable expression. Infect Immun 81:2768–2776
Koizumi N, Watanabe H (2003) Molecular cloning and characterization of a novel leptospiral lipoprotein with OmpA domain. FEMS Microbiol Lett 226:215–219
Kovacs-Simon A, Titball RW, Michell SL (2011) Lipoproteins of bacterial pathogens. Infect Immun 79:548–561
Kropinski AM, Parr TR Jr, Angus BL, Hancock RE, Ghiorse WC, Greenberg EP (1987) Isolation of the outer membrane and characterization of the major outer membrane protein from Spirochaeta aurantia. J Bacteriol 169:172–179
Kumru OS, Schulze RJ, Slusser JG, Zückert WR (2010) Development and validation of a FACS-based lipoprotein localization screen in the Lyme disease spirochete Borrelia burgdorferi. BMC Microbiol 10:277
Kumru OS, Schulze RJ, Rodnin MV, Ladokhin AS, Zuckert WR (2011) Surface localization determinants of Borrelia OspC/Vsp family lipoproteins. J Bacteriol 193:2814–2825
Lee PA, Tullman-Ercek D, Georgiou G (2006) The bacterial twin-arginine translocation pathway. Annu Rev Microbiol 60:373–395
Lessa-Aquino C, Borges Rodrigues C, Pablo J, Sasaki R, Jasinskas A, Liang L, Wunder EA Jr, Ribeiro GS, Vigil A, Galler R, Molina D, Liang X, Reis MG, Ko AI, Medeiros MA, Felgner PL (2013) Identification of seroreactive proteins of Leptospira interrogans serovar Copenhageni using a high-density protein microarray approach. PLoS Negl Trop Dis 7:e2499
Lin MH, Chang YC, Hsiao CD, Huang SH, Wang MS, Ko YC, Yang CW, Sun YJ (2013) LipL41, a hemin binding protein from Leptospira santarosai serovar Shermani. PLoS ONE 8:e83246
Lo M, Bulach DM, Powell DR, Haake DA, Matsunaga J, Paustian ML, Zuerner RL, Adler B (2006) Effects of temperature on gene expression patterns in Leptospira interrogans serovar Lai as assessed by whole-genome microarrays. Infect Immun 74:5848–5859
Lo M, Cordwell SJ, Bulach DM, Adler B (2009) Comparative transcriptional and translational analysis of leptospiral outer membrane protein expression in response to temperature. PLoS Negl Trop Dis 3:e560
Lo M, Murray GL, Khoo CA, Haake DA, Zuerner RL, Adler B (2010) Transcriptional response of Leptospira interrogans to iron limitation and characterization of a PerR homolog. Infect Immun 78:4850–4859
Lourdault K, Wang LC, Vieira A, Matsunaga J, Melo R, Lewis MS, Haake DA, Gomes-Solecki M (2014) Oral immunization with E. coli expressing a lipidated form of LigA protects hamsters against challenge with Leptospira interrogans serovar Copenhageni. Infect Immun doi:10.1128/IAI.01533-13
Louvel H, Bommezzadri S, Zidane N, Boursaux-Eude C, Creno S, Magnier A, Rouy Z, Médigue C, Saint Girons I, Bouchier C, Picardeau M (2006) Comparative and functional genomic analyses of iron transport and regulation in Leptospira spp. J Bacteriol 188:7893–7904
Malmström J, Beck M, Schmidt A, Lange V, Deutsch EW, Aebersold R (2009) Proteome-wide cellular protein concentrations of the human pathogen Leptospira interrogans. Nature 460:762–766
Matsui M, Soupé ME, Becam J, Goarant C (2012) Differential in vivo gene expression of major Leptospira proteins in resistant or susceptible animal models. Appl Environ Microbiol 78:6372–6376
Matsunaga J, Young TA, Barnet JK, Barnett D, Bolin CA, Haake DA (2002) Novel 45-kilodalton leptospiral protein that is processed to a 31-kilodalton growth-phase-regulated peripheral membrane protein. Infect Immun 70:323–334
Matsunaga J, Barocchi MA, Croda J, Young TA, Sanchez Y, Siqueira I, Bolin CA, Reis MG, Riley LW, Haake DA, Ko AI (2003) Pathogenic Leptospira species express surface-exposed proteins belonging to the bacterial immunoglobulin superfamily. Mol Microbiol 49:929–945
Matsunaga J, Sanchez Y, Xu X, Haake DA (2005) Osmolarity, a key environmental signal controlling expression of leptospiral proteins LigA and LigB and the extracellular release of LigA. Infect Immun 73:70–78
Matsunaga J, Wernied K, Zuerner R, Frank A, Haake, DA (2006) LipL46 is a novel, surface-exposed lipoprotein expressed during leptospiral dissemination in the mammalian host. Microbiol 152, 3777–3786
Matsunaga J, Lo M, Bulach DM, Zuerner RL, Adler B, Haake DA (2007a) Response of Leptospira interrogans to physiologic osmolarity: relevance in signaling the environment-to-host transition. Infect Immun 75:2864–2874
Matsunaga J, Medeiros MA, Sanchez Y, Werneid KF, Ko AI (2007b) Osmotic regulation of expression of two extracellular matrix-binding proteins and a haemolysin of Leptospira interrogans: differential effects on LigA and Sph2 extracellular release. Microbiol 153:3390–3398
Matsunaga J, Schlax PJ, Haake DA (2013) Role for cis-acting RNA sequences in the temperature-dependent expression of the multiadhesive Lig proteins in Leptospira interrogans. J Bacteriol 195:5092–5101
McBride AJ, Cerqueira GM, Suchard MA, Moreira AN, Zuerner RL, Reis MG, Haake DA, Ko AI, Dellagostin OA (2009) Genetic diversity of the leptospiral immunoglobulin-like (Lig) genes in pathogenic Leptospira spp. Infect Genet Evol 9:196–205
Midwinter AC, Vinh T, Faine S, Adler B (1994) Characterization of an antigenic oligosaccharide from Leptospira interrogans serovar pomona and its role in immunity. Infect Immun 62:5477–5482
Murray GL (2013) The lipoprotein LipL32, an enigma of leptospiral biology. Vet Microbiol 162:305–314
Murray GL, Morel V, Cerqueira GM, Croda J, Srikram A, Henry R, Ko AI, Dellagostin OA, Bulach DM, Sermswan RW, Adler B, Picardeau M (2009a) Genome-wide transposon mutagenesis in pathogenic Leptospira species. Infect Immun 77:810–816
Murray GL, Srikram A, Hoke DE, Wunder EA Jr, Henry R, Lo M, Zhang K, Sermswan RW, Ko AI, Adler B (2009b) Major surface protein LipL32 is not required for either acute or chronic infection with Leptospira interrogans. Infect Immun 77:952–958
Murray GL, Srikram A, Henry R, Hartskeerl RA, Sermswan RW, Adler B (2010) Mutations affecting Leptospira interrogans lipopolysaccharide attenuate virulence. Mol Microbiol 78:701–709
Nahori MA, Fournie-Amazouz E, Que-Gewirth NS, Balloy V, Chignard M, Raetz CR, Saint Girons I, Werts C (2005) Differential TLR recognition of leptospiral lipid A and lipopolysaccharide in murine and human cells. J Immunol 175:6022–6031
Nally JE, Artiushin S, Timoney JF (2001a) Molecular characterization of thermoinduced immunogenic proteins Q1p42 and Hsp15 of Leptospira interrogans. Infect Immun 69:7616–7624
Nally JE, Timoney JF, Stevenson B (2001b) Temperature-regulated protein synthesis by Leptospira interrogans. Infect Immun 69:400–404
Nally JE, Chow E, Fishbein MC, Blanco DR, Lovett MA (2005a) Changes in lipopolysaccharide O antigen distinguish acute versus chronic Leptospira interrogans infections. Infect Immun 73:3251–3260
Nally JE, Whitelegge JP, Aguilera R, Pereira MM, Blanco DR, Lovett MA (2005b) Purification and proteomic analysis of outer membrane vesicles from a clinical isolate of Leptospira interrogans serovar Copenhageni. Proteomics 5:144–152
Nally JE, Whitelegge JP, Bassilian S, Blanco DR, Lovett MA (2007) Characterization of the outer membrane proteome of Leptospira interrogans expressed during acute lethal infection. Infect Immun 75:766–773
Nascimento AL, Ko AI, Martins EA, Monteiro-Vitorello CB, Ho PL, Haake DA, Verjovski-Almeida S, Hartskeerl RA, Marques MV, Oliveira MC, Menck CF, Leite LC, Carrer H, Coutinho LL, Degrave WM, Dellagostin OA, El-Dorry H, Ferro ES, Ferro MI, Furlan LR, Gamberini M, Giglioti EA, Góes-Neto A, Goldman GH, Goldman MH, Harakava R, Jerônimo SM, Junqueira-de-Azevedo IL, Kimura ET, Kuramae EE, Lemos EG, Lemos MV, Marino CL, Nunes LR, de Oliveira RC, Pereira GG, Reis MS, Schriefer A, Siqueira WJ, Sommer P, Tsai SM, Simpson AJ, Ferro JA, Camargo LE, Kitajima JP, Setubal JC, Van Sluys MA (2004) Comparative genomics of two Leptospira interrogans serovars reveals novel insights into physiology and pathogenesis. J Bacteriol 186:2164–2172
Oke M, Sarra R, Ghirlando R, Farnaud S, Gorringe AR, Evans RW, Buchanan SK (2004) The plug domain of a neisserial TonB-dependent transporter retains structural integrity in the absence of its transmembrane beta-barrel. FEBS lett 564:294–300
Oliveira TR, Longhi MT, Goncales AP, de Morais ZM, Vasconcellos SA, Nascimento AL (2010) LipL53, a temperature regulated protein from Leptospira interrogans that binds to extracellular matrix molecules. Microbes Infect 12:207–217
Oliveira R, de Morais ZM, Goncales AP, Romero EC, Vasconcellos SA, Nascimento AL (2011) Characterization of novel OmpA-like protein of Leptospira interrogans that binds extracellular matrix molecules and plasminogen. PLoS ONE 6:e21962
Patarakul K, Lo M, Adler B (2010) Global transcriptomic response of Leptospira interrogans serovar Copenhageni upon exposure to serum. BMC Microbiol 10:31
Pinne M, Haake DA (2009) A comprehensive approach to identification of surface-exposed, outer membrane-spanning proteins of Leptospira interrogans. PLoS ONE 4:e6071
Pinne M, Haake DA (2011) Detection of leptospiral surface-exposed proteins via immunofluorescence. J Vis Exp 53:2805
Pinne M, Haake DA (2013) LipL32 is a subsurface lipoprotein of Leptospira interrogans: presentation of new data and reevaluation of previous studies. PLoS ONE 8:e51025
Pinne M, Matsunaga J, Haake DA (2012) A novel approach to identification of leptospiral ligand-binding proteins: an outer-membrane protein microarray. J Bacteriol 194:6074–6087
Pomorski T, Menon AK (2006) Lipid flippases and their biological functions. Cell Mol Life Sci 63:2908–2921
Que-Gewirth NS, Riberio AA, Kalb SR, Cotter RJ, Bulach DM, Adler B, Saint Girons I, Werts C, Raetz CR (2004) A methylated phosphate group and four amide-linked acyl chains in Leptospira interrogans Lipid A. J Biol Chem 279:25420–25429
Raddi G, Morado DR, Yan J, Haake DA, Yang XF, Liu J (2012) Three-dimensional structures of pathogenic and saprophytic Leptospira species revealed by cryo-electron tomography. J Bacteriol 194:1299–1306
Radolf JD, Norgard MV, Schulz WW (1989) Outer membrane ultrastructure explains the limited antigenicity of virulent Treponema pallidum. Proc Natl Acad Sci USA 86:2051–2055
Ren SX, Fu G, Jiang XG, Zeng R, Miao YG, Xu H, Zhang YX, Xiong H, Lu G, Lu LF, Jiang HQ, Jia J, Tu YF, Jiang JX, Gu WY, Zhang YQ, Cai Z, Sheng HH, Yin HF, Zhang Y, Zhu GF, Wan M, Huang HL, Qian Z, Wang SY, Ma W, Yao ZJ, Shen Y, Qiang BQ, Xia QC, Guo XK, Danchin A, Saint Girons I, Somerville RL, Wen YM, Shi MH, Chen Z, Xu JG, Zhao GP (2003) Unique physiological and pathogenic features of Leptospira interrogans revealed by whole-genome sequencing. Nature 422:888–893
Ristow P, Bourhy P, da Cruz McBride FW, Figueira CP, Huerre M, Ave P, Saint Girons I, Ko AI, Picardeau M (2007) The OmpA-like protein Loa22 is essential for leptospiral virulence. PLoS Pathog 3:e97
Ruiz N, Kahne D, Silhavy TJ (2009) Transport of lipopolysaccharide across the cell envelope: the long road of discovery. Nat Rev Microbiol 7:677–683
Sauvonnet N, Pugsley AP (1996) Identification of two regions of Klebsiella oxytoca pullulanase that together are capable of promoting beta-lactamase secretion by the general secretory pathway. Mol Microbiol 22:1–7
Schauer K, Rodionov DA, de Reuse H (2008) New substrates for TonB-dependent transport: do we only see the ‘tip of the iceberg’? Trends Biochem Sci 33:330–338
Schulze RJ, Zuckert WR (2006) Borrelia burgdorferi lipoproteins are secreted to the outer surface by default. Mol Microbiol 59:1473–1484
Schulze RJ, Chen S, Kumru OS, Zückert WR (2010) Translocation of Borrelia burgdorferi surface lipoprotein OspA through the outer membrane requires an unfolded conformation and can initiate at the C-terminus. Mol Microbiol 76:1266–1278
Setubal JC, Reis M, Matsunaga J, Haake DA (2006) Lipoprotein computational prediction in spirochaetal genomes. Microbiol 152:113–121
Shang ES, Exner MM, Summers TA, Martinich C, Champion CI, Hancock RE, Haake DA (1995) The rare outer membrane protein, OmpL1, of pathogenic Leptospira species is a heat-modifiable porin. Infect Immun 63:3174–3181
Shang ES, Summers TA, Haake DA (1996) Molecular cloning and sequence analysis of the gene encoding LipL41, a surface-exposed lipoprotein of pathogenic Leptospira species. Infect Immun 64:2322–2330
Silva EF, Medeiros MA, McBride AJ, Matsunaga J, Esteves GS, Ramos JG, Santos CS, Croda J, Homma A, Dellagostin OA, Haake DA, Reis MG, Ko AI (2007) The terminal portion of leptospiral immunoglobulin-like protein LigA confers protective immunity against lethal infection in the hamster model of leptospirosis. Vaccine 25:6277–6286
Sklar JG, Wu T, Kahne D, Silhavy TJ (2007) Defining the roles of the periplasmic chaperones SurA, Skp, and DegP in Escherichia coli. Genes Develop 21:2473–2484
Sperandeo P, Deho G, Polissi A (2009) The lipopolysaccharide transport system of Gram-negative bacteria. Biochim Biophys Acta 1791:594–602
Srikram A, Zhang K, Bartpho T, Lo M, Hoke DE, Sermswan RW, Adler B, Murray GL (2011) Cross-protective immunity against leptospirosis elicited by a live, attenuated lipopolysaccharide mutant. J Infect Dis 203:870–879
Stevenson B, Choy HA, Pinne M, Rotondi ML, Miller MC, Demoll E, Kraiczy P, Cooley AE, Creamer TP, Suchard MA, Brissette CA, Verma A, Haake DA (2007) Leptospira interrogans endostatin-like outer membrane proteins bind host fibronectin, laminin and regulators of complement. PLoS ONE 2:e1188
Tommassen J (2007) Getting into and through the outer membrane. Science 317:903–904
Verma A, Artiushin S, Matsunaga J, Haake DA, Timoney JF (2005) LruA and LruB, novel lipoproteins of pathogenic Leptospira interrogans associated with equine recurrent uveitis. Infect Immun 73:7259–7266
Verma A, Hellwage J, Artiushin S, Zipfel PF, Kraiczy P, Timoney JF, Stevenson B (2006) LfhA, a novel factor H-binding protein of Leptospira interrogans. Infect Immun 74:2659–2666
Verma A, Brissette CA, Bowman AA, Shah ST, Zipfel PF, Stevenson B (2010) Leptospiral endostatin-like protein A (LenA) is a bacterial cell-surface receptor for human plasminogen. Infect Immun 78:2053–2059
Vieira ML, Atzingen MV, Oliveira R, Mendes RS, Domingos RF, Vasconcellos S, Nascimento AL (2012) Plasminogen binding proteins and plasmin generation on the surface of Leptospira spp.: the contribution to the bacteria-host interactions. J Biomed Biotechnol 2012:758513
Viriyakosol S, Matthias MA, Swancutt MA, Kirkland TN, Vinetz JM (2006) Toll-like receptor 4 protects against lethal Leptospira interrogans serovar icterohaemorrhagiae infection and contributes to in vivo control of leptospiral burden. Infect Immun 74:887–895
Vivian JP, Beddoe T, McAlister AD, Wilce MC, Zaker-Tabrizi L, Troy S, Byres E, Hoke DE, Cullen PA, Lo M, Murray GL, Adler B, Rossjohn J (2009) Crystal structure of LipL32, the most abundant surface protein of pathogenic Leptospira spp. J Mol Biol 387:1229–1238
Walker EM, Borenstein LA, Blanco DR, Miller JN, Lovett MA (1991) Analysis of outer membrane ultrastructure of pathogenic Treponema and Borrelia species by freeze-fracture electron microscopy. J Bacteriol 173:5585–5588
Werts C (2010) Leptospirosis: a Toll road from B lymphocytes. Chang Gung Med J 3:591–601
Werts C, Tapping RI, Mathison JC, Chuang TH, Kravchenko V, Saint Girons I, Haake DA, Godowski PJ, Hayashi F, Ozinsky A, Underhill DM, Kirschning CJ, Wagner H, Aderem A, Tobias PS, Ulevitch RJ (2001) Leptospiral endotoxin activates cells via a TLR2-dependent mechanism. Nature Immunol 2:346–352
Wunder EA Jr, Figueira CP, Benaroudj N, Hu B, Tong BA, Trajtenberg F, Liu J, Reis MG, Charon N, Buschiazzo A, Picardeau M, Ko AI (2013) Leptospira Fcp1 is a key protein in determining the coiled morphology of purified flagella and conferring translational motility and virulence for the spirochete. In: 8th Scientific Meeting of the International Leptospirosis Society, Fukuoka, Japan
Xue F, Dong H, Wu J, Wu Z, Hu W, Sun A, Troxell B, Yang XF, Yan J (2010) Transcriptional responses of Leptospira interrogans to host innate immunity: significant changes in metabolism, oxygen tolerance, and outer membrane. PLoS Negl Trop Dis 4:e857
Yakushi T, Masuda K, Narita S, Matsuyama S, Tokuda H (2000) A new ABC transporter mediating the detachment of lipid-modified proteins from membranes. Nat Cell Biol 2:212–218
Yang CW, Wu MS, Pan MJ, Hsieh WJ, Vandewalle A, Huang CC (2002) The Leptospira outer membrane protein LipL32 induces tubulointerstitial nephritis-mediated gene expression in mouse proximal tubule cells. J Am Soc Nephrol 13:2037–2045
Yang CW, Hung CC, Wu MS, Tian YC, Chang CT, Pan MJ, Vandewalle A (2006) Toll-like receptor 2 mediates early inflammation by leptospiral outer membrane proteins in proximal tubule cells. Kidney Int 69:815–822
Yokota N, Kuroda T, Matsuyama S, Tokuda H (1999) Characterization of the LolA-LolB system as the general lipoprotein localization mechanism of Escherichia coli. J Biol Chem 274:30995–30999
Zhang K, Murray GL, Seemann T, Srikram A, Bartpho T, Sermswan RW, Adler B, Hoke DE (2013) Leptospiral LruA is required for virulence and modulates an interaction with mammalian apolipoprotein AI. Infect Immun 81:3872–3879
Zuerner RL, Knudtson W, Bolin CA, Trueba G (1991) Characterization of outer membrane and secreted proteins of Leptospira interrogans serovar pomona. Microb Pathog 10:311–322
Acknowledgments
The authors are extremely grateful to Dr. James Matsunaga for his helpful comments on regulation of Lig expression. Current work in Dr. Haake’s laboratory is supported by NIH Grant R01 AI034431 and a VA Merit Award. Current work in Dr. Zückert’s laboratory is supported by NIH Grant P30 GM103326 and a University of Kansas Medical Center Research Institute Lied Basic Science Pilot Grant.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Haake, D.A., Zückert, W.R. (2015). The Leptospiral Outer Membrane. In: Adler, B. (eds) Leptospira and Leptospirosis. Current Topics in Microbiology and Immunology, vol 387. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-662-45059-8_8
Download citation
DOI: https://doi.org/10.1007/978-3-662-45059-8_8
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-662-45058-1
Online ISBN: 978-3-662-45059-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)