
Abstract
For both hepatoblastomas and hepatocellular carcinoma, knowledge regarding distinct pathologic phenotypes has undergone considerable expansion, what has resulted in novel and more refined classifications of these tumors. These morphotypes are increasingly supplemented by cytogenetic and molecular findings, leading to characteristic signatures. It is expected that these findings will be the baseline for improved risk stratifications to be employed for novel treatment strategies.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Adenomatous Polyposis Coli
- Biliary Atresia
- Focal Nodular Hyperplasia
- Extramedullary Hematopoiesis
- Alagille Syndrome
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
9.1 Hepatoblastoma and Related Tumors
9.1.1 Introduction
Hepatoblastoma is a term proposed by Willis for all embryonal tumors containing hepatic epithelial parenchyma more or less resembling fetal or embryonal tissue (Willis 1962). Early reports reviewed by Ishak and Glunz (1967) employed several terms to denote hepatoblastoma, including primary hepatoma or carcinoma of the liver in infancy and childhood, mixed tumors of the liver, and hepatic embryonic tumors. As the prognosis of these lesions was dismal those days, it was not thought important to classify the tumors differently. Subsequently, this changed considerably, leading to the concept of hepatoblastoma that we have today (reviews: Dehner 1978; Weinberg and Finegold 1983; Stocker 2001; Zimmermann 2005; Meyers 2007; Finegold et al. 2008). Hepatoblastoma is predominantly a disorder of the liver in small children and infants (see the chapter on epidemiology), but may occur in adults up to the age of 80 years.
9.1.2 Classification of Hepatoblastoma
Willis (1962) had classified hepatoblastoma into three types: (1) embryonic hepatoma, containing only embryonic liver tissue; (2) mixed tumors containing both epithelial and mesenchymal components; and (3) rhabdomyoblastic mixed tumors. In the same period, Edmondson still had epithelial hepatoblastomas under the category primary carcinoma of the liver, and the other lesions were mixed tumors (Edmondson 1956). Later, a so-called “pure hepatoblastoma” was added to this classification, denoting mixed tumors without osteoid (Neimann et al. 1963), while the osteoid-containing variant was identified as a separate entity (Milman and Grayzel 1951); this is currently obsolete. A major breakthrough was the proposition by Ishak and Glunz, who divided the tumors into an epithelial type and a mixed epithelial mesenchymal type, further breaking down the epithelial tumors into those with fetal or embryonal cells (Ishak and Glunz 1967), a refined and improved modification of the classification previously advocated by Willis. The criteria for modern classifications of hepatoblastoma have recently been reviewed (Rowland 2002), and a more expanded classification of the hepatoblastoma tumor family has been proposed (Zimmermann 2005). Table 9.1 shows the hepatoblastoma classification currently employed by SIOPEL.
About 55% of hepatoblastomas are epithelial (30% fetal, 20% fetal-embryonal, 3% macrotrabecular, 2% small cell undifferentiated), and 45% are mixed epithelial and mesenchymal, but when all types are considered, around 85% contain both fetal and embryonal components in variable proportions. There is no relationship between the age of the child and the predominant cell type in hepatoblastoma.
9.1.3 Macroscopy
Hepatoblastomas are more commonly solitary than multifocal, mostly expanding masses of roughly spherical shape (Ishak and Glunz 1967). Focal calcifications may be seen macroscopically; radiologically, coarse calcifications are present in hepatoblastoma in 50% of the cases, but many of them are in fact mineralized foci of osteoid (Dachman et al. 1987). Some tumors present with prominent feeding arteries and enlarged or engorged veins. Such tumors are hypervascular multifocal hepatoblastomas at imaging and may be confounded with hemangioendothelioma (Ingram et al. 2000; Lu and Greer 2007).
The gross and histologic work-up of hepatoblastomas has been formulated in a College of American Pathologists protocol (Finegold et al. 2007). A detailed description of the gross features delivers informations that are important for the confirmation of staging and for prognostication. Several parameters of growth patterns have been shown to be predictors of failed conservative treatment (FCT), such as multifocality, portal vein involvement, hepatic vein involvment, and vena cava involvement (Von Schweinitz et al. 1994; Davies et al. 2004; D’Antiga et al. 2007). Gross examination must include the precise assessment of radicality, what is sometimes difficult owing to the complexity of resection surfaces, especially in the fixed state of specimens. For the evaluation of surgical resection margins (Dicken et al. 2004) and the assessment of microscopic residual disease, it is recommended that surgeons and pathologists find a way to identify critical margin areas and the vascular and biliary trees, for example, by use of colored sutures or inking. In newborns with congenital hepatoblastoma, macroscopy must include the examination of the placenta, because hepatoblastoma can metastasize to this organ (Robinson and Bolande 1985; Doss et al. 1998).
9.1.4 Histopathology of Hepatoblastomas
Based on the histomorphologic features that had led to the current classifications, the specific features of the types and subtypes of hepatoblastoma have been worked out (Edmondson 1956; Ishak and Glunz 1967; Lack et al. 1982; Weinberg and Finegold 1983; Dehner and Manivel 1988; Haas et al. 1989; Conran et al. 1992). In the following overview, the types and subtypes follow the SIOPEL classification.
9.1.4.1 Fetal Hepatoblastoma: The Differentiated Phenotype with a Favorable Histology
The morphology of fetal-type cells has already been specified in detail in 1967 (Ishak and Glunz 1967). They are smaller than normal adult hepatocytes but larger than normal fetal hepatocytes, with a well-defined outline, and present a cytoplasm that varies from eosinophil and is slightly granular to clear (Fig. 9.1a and b), resulting in a characteristic dark-and-clear cell pattern, or a pattern of light and dark lobules. The pale or clear aspect of the cytoplasm is sometimes striking (Fig. 9.1d) and is caused by accumulation of glycogen and lipids. Marked lipid accumulation induces vacuolization of the cell, commonly seen after chemotherapy. Bile production can be noted, sometimes with dense bile deposits in canalicular-like structures. The nuclei are round to slightly ovoid, with a rather fine chromatin structure and one small acidophilic nucleolus. Fetal cells are arranged in cords, nests, or nodules, without a prominent stroma (except around feeding vessels or in certain tumor variants with a lobular growth pattern and hamartoma-like features) and with a rather poorly developed network of reticular tissue (Ishak and Glunz 1967). In compact areas, sinusoids may be absent, resulting in a cellular mosaic pattern (Haas et al. 1989). Extramedullary hematopoiesis is a typical feature (Fig. 9.1c), whereby erythroblasts predominate, but megakaryocytes are also in evidence. These foci are intrasinusoidal and are not seen in the adjacent normal liver tissue.

(a) Fetal hepatoblastoma. The cells form solid formations and nodules, and exhibit a clear cytoplasm (hematoxylin and eosin stain). (b) In this fetal hepatoblastoma, both clear and dark cells are noted (hematoxylin and eosin stain). (c) Several hematopoietic clusters are seen in this fetal hepatoblastoma (hematoxylin and eosin stain). (d) Clear cells of a fetal hepatoblastoma at higher magnification (hematoxylin and eosin stain)
In the well-differentiated variety of fetal hepatoblastoma, there is little mitotic activity (two or less than two mitotic figures per ten high-power [×40 objective] fields; pure fetal histology). A subset of fetal hepatoblastoma shows significant mitotic activity (>2 mitotic figures in ten high-power fields). This variant is called mitotically active fetal or crowded fetal hepatoblastoma, because cytoplasmic glycogen storage is less and hence the proportion of a sample occupied by nuclei is increased.
9.1.4.2 Embryonal Histology: A Common Partner of Fetal Tissue Components
The embryonal pattern almost always occurs in combination with fetal components (purely embryonal hepatoblastomas are exceptional observations Borman et al. 1961). Embryonal hepatoblastoma cells are less differentiated than fetal cells, poorly cohesive, elongated, and have a high nuclear cytoplasmic ratio and a sparse, compact basophilic or amphophilic, poorly outlined cytoplasm (Fig. 9.2a). Bile production is not seen. The nuclei are oval rather than round and show a coarser chromatin, with prominent parachromatin and one enlarged amphophilic or acidophilic nucleolus. Mitoses are more frequent than in the fetal subtype. Transitions between fetal cells and embryonal cells are common, and sometimes embryonal tissue is seen at the periphery of otherwise fetal tissue areas (Fig. 9.2b). The growth pattern of embryonal areas is complex and includes solid sheets or plates of variable thickness, incomplete or complete tubuloacinar profiles (Fig. 9.2c), and rosette-like configurations (pseudorosettes) resembling primitive bile ducts of the embryonic liver prior to the sixth week of gestation (Gonzalez-Crussi et al. 1982; Fig. 9.2d). Micropapillary structures may occur and sometimes form a dominant pattern. The vascular network differs from that of fetal-type tumors, in that a fine capillary network and larger vascular channels are present, sometimes forming dilated channels with incomplete endothelial lining (vascular lakes lined by tumor cells; Ishak and Glunz 1967) or pelioid areas. In contrast to fetal tumors, extramedullary hematopoiesis is very rarely observed in embryonal areas. Embryonal tumor tissue exhibits necroses and apoptotic bodies, most marked in chemotherapy-treated tumors.

(a) Embryonal components of a mixed epithelial hepatoblastoma (hematoxylin and eosin stain). (b) In this embryonal/fetal hepatoblastoma, embryonal components are located to the periphery of fetal parts (hematoxylin and eosin stain). (c) Embryonal area of a hepatoblastoma with acinar formations (hematoxylin and eosin stain). (d) In this embryonal area, immature tubular structures are present (hematoxylin and eosin stain)
9.1.4.3 Macrotrabecular Hepatoblastoma: A Distinct Growth Pattern
Gonzalez Crussi and coworkers have described hepatoblastoma areas having a macrotrabecular growth pattern in otherwise typical tumors. Macrotrabecules are 10–20 or more cells thick, resulting in a typical pattern (Fig. 9.3a and b). The cells in the macrotrabecular parts may be fetal, embryonal, or indistinguishable from those of adult-type hepatocellular carcinoma (Gonzalez-Crussi et al. 1982). The macrotrabecular phenotype occurs as a pure form, but may also be mixed with other histologies. It has been proposed that a hepatoblastoma with only an isolated macrotrabecular focus should not be classified as a macrotrabecular subtype (Conran et al. 1992), but the cut-off criteria have not yet been defined.

(a) Macrotrabecular hepatoblastoma at low magnification. Note the resemblance to hepatocellular carcinoma (hematoxylin and eosin stain). (b) The large cell plates (macrotrabecules) are clearly seen in this tumor (Hematoxylin and eosin stain). (c) Macrotrabecular hepatoblastoma consisting of embryonal and fetal cells (MT-2; hematoxylin and eosin stain). (d) Macrotrabecular hepatoblastoma consisting of hepatocyte-like cells (MT-1; hematoxylin and eosin stain)
Previously, macrotrabecular hepatoblastoma was characterized by a repetitive arrangement of fetal or embryonal cells in macrotrabecules (Haas et al. 1989), but these authors noted that occasionally the tumor cell size may resemble the cells of hepatocellular carcinoma. It has therefore been proposed to divide macrotrabecular hepatoblastomas into two categories (Zimmermann 2005): MT-1 composed of hepatocyte/HCC-like cells (Fig. 9.3d), and MT-2 composed of fetal and/or embryonal cells (Fig. 9.3c).
With regard to the biology of macrotrabecular hepatoblastomas, only two therapy studies have specifically referred to this variant. In the first study of 168 patients with hepatoblastoma, 18 patients had macrotrabecular tumors, and their estimated 24-month survival probability was 50% in comparison with 92%, 63%, and 0% for the purely fetal, embryonal, and small cell undifferentiated histologies, respectively (Haas et al. 1989). In a later second investigation that also identified the macrotrabecular variant, the histology of any type or subtype of hepatoblastoma did not have a significant prognostic effect (Conran et al. 1992). In summary, there are presently too few observations on this subtype to be certain whether it is prognostically unfavorable or not, but it is believed that the MT1 phenotype is a high-risk histology.
9.1.4.4 Undifferentiated Epithelial Hepatoblastomas
This rare subgroup of wholly epithelial hepatoblastomas is characterized by poorly differentiated or undifferentiated (anaplastic) cells cytologically resembling those of other “blue cellular tumors.” So far, undifferentiated hepatoblastoma is defined as an aggressive (high-risk) neoplasm composed of small cells, mostly with a diffuse growth pattern (hepatoblastoma, small cell undifferentiated; HB-SCUD). However, this group of tumors is heterogeneous and contains, apart from HB-SCUD, neoplasms with focal expression of anaplasia, tumors with undifferentiated cells of intermediate or large cells rather than small cells, and a subset of lesions, which seem to be related to malignant rhabdoid tumors.
9.1.4.4.1 Small Cell Undifferentiated Hepatoblastoma (HB-SCUD)
This neoplasm was originally termed, “anaplastic type,” and described as a lesion having small cells resembling those of neuroblastoma (Kasai and Watanabe 1970). However, small undifferentiated round and spindle-shaped cells have been described in hepatoblastoma earlier (Misugi et al. 1967). Haas et al. proposed to replace “anaplasia” by “small cell undifferentiated” (SCUD) (Haas et al. 1989). The small cell cytologic and histologic patterns have since been reported several times, but mostly in small numbers only (Sinniah et al. 1974; Lack et al. 1982; Weinberg and Finegold 1983; Haas et al. 1989; Gonzalez-Crussi 1991; Hansen et al. 1992; Stocker 1994). When the small cell feature is present in a significant proportion of a hepatoblastoma (75%) or as the sole cell type, this tumor subtype is typically found in infants younger than 1 year. The SCUD phenotype can be combined with any other hepatoblastoma types and subtypes. HB-SCUD is a rapidly growing and highly aggressive subtype of hepatoblastoma, with an estimated 2-year patient survival rate probably not exceeding 0% (Haas et al. 1989). Few studies have, however, systematically analyzed the clinical behavior of HB-SCUD, also related to the rarity of this lesion. In a study of completely resected hepatoblastoma, the 38% recurrence rate in tumors with a SCUD histology compared unfavorably with the overall estimated event-free survival rate of 91% for the entire group (Ortega et al. 2000). HB-SCUD is, therefore, one of the phenotypes in the hepatoblastoma family of tumors with an “unfavorable histology.”
Histologically, the typical growth pattern of HB-SCUD is diffuse (Fig. 9.4a) or spotty (Fig. 9.4b), with small cells being ovoid, stellate, or spindle-shaped with slight eosinophilia or amphophilia of the poorly developed cytoplasm. The nuclei exhibit a dense chromatin and variably prominent nucleoli (Fig. 9.4c). In silver stains, delicate reticulin fibers surround small to large clusters or sheets of tumor cells, without a distinct pattern.The stroma is usually scanty or lacking. The tumor tends to invade the adjacent liver substance, often with a prominent intravascular growth (Fig. 9.4d), and the neoplastic tissue may engulf preexisting bile ducts and ductules (Haas et al. 1989; Fig. 9.4e). Necrosis may be extensive (Fig. 9.4f). Immunohistochemically, the tumor cells are reactive for vimentin, pankeratin, and cytokeratins 8 (Fig. 9.5a). The proliferative activity is sometimes markedly increased (Fig. 9.5b).Typically, the tumor cells do not usually stain for AFP (with few exceptions; Abenoza et al. 1987), and in fact part of tumors with normal or only slightly elevated serum AFP and aggressive biology are HB-SCUD (De Ioris et al. 2008). Rare variants of HB-SCUD exhibit a myxoid stroma with large amounts of glycosaminoglycans (Fig. 9.6a and b), sometimes with mucoid microcysts (“mucoid anaplastic hepatoblastoma”; Joshi et al. 1984).

(a) Small cell undifferentiated hepatoblastoma, diffuse growth pattern (hematoxylin and eosin stain). (b) Small cell undifferentiated hepatoblastoma, spotty growth pattern (ematoxylin and eosin stain). (c) Cellular phenotype of small cell undifferentiated hepatoblastoma (hematoxylin and eosin stain). (d) Vascular invasion of small cell undifferentiated hepatoblastoma (hematoxylin and eosin stain). (e) In this small cell undifferentiated hepatoblastoma, tumor cells have infiltrated the tissue surrounding a bile duct (hematoxylin and eosin stain). (f) This tumor has undergone partial necrosis, visualized as an eosinophilic mass with ghost cells (lower half of figure) and a zone of damaged cells (hematoxylin and eosin stain)

(a) Cytokeratin 8 expression in cells of small cell undifferentiated hepatoblastoma (brown reaction product; CK8 immunostain). (b) Small cell undifferentiated hepatoblastomas may show high proliferative activity (proliferation marker/Ki-67 immunostain).

(a) Myxoid variant of small cell undifferentiated hepatoblastoma. Note the prominent myxoid (whitish-blue) intercellular matrix (hematoxylin and eosin stain). (b) The myxoid matrix is rich in glycosaminoglycans (maxtrix in blue, tumor cells in red; alcaline Alcian blue stain)
The SCUD phenotype may be focal and associated with fetal- and/or embryonal-type tumor cells; this phenomenon has previously been noted in descriptions of hepatoblastomas (Gonzalez-Crussi et al. 1982). Even incomplete, that is, nondiffuse expression of the SCUD phenotype has been shown to be unfavorable (Douglass et al. 1993; Haas et al. 2001). In a CCG report of 33 patients with Stage I hepatoblastoma, 3/18 patients whose completely resected hepatoblastomas were believed to be of pure fetal histology later developed pulmonary metastases. The histology review showed that all three tumors contained scattered microscopic foci of SCUD histology (Feusner et al. 1993; Haas et al. 2001).
9.1.4.4.2 Other Phenotypes of Undifferentiated Hepatoblastoma
Rare cases of undifferentiated hepatoblastomas do not express a small cell phenotype, but are composed of intermediate-sized cells or even large cells (hepatoblastoma, intermediate cell undifferentiated, HB-ICUD; hepatoblastoma, large cell undifferentiated, HB-LCUD; Zimmermann 2005). Medium-sized cells in undifferentiated hepatoblastoma have previously been noted (Lack et al. 1982) and earlier observations of large cells occurring in hepatoblastomas have been described (Weinberg and Finegold 1983). Large cell features associated with aggressive course are also recognized for other blastomas, including large cell medulloblastoma and large cell neuroblastoma. Some large cell hepatoblastomas are CD99-positive, in the absence of any other PNET features (Zimmermann 2005), but CD99 positivity has also been found in other hepatoblastomas (Ramsay et al. 2008).
9.1.4.4.3 Undifferentiated Hepatoblastoma with Rhabdoid Features and Malignant Rhabdoid Tumor
At least part of HB-SCUD seems to have a relationship to malignant rhabdoid tumors (MRT), which also occur in the liver (Parham et al. 1994; Scheimberg et al. 1996; Garcés-Inigo et al. 2009). MRT is histologically characterized by sheets of large polygonal cells with abundant cytoplasm and vesicular nuclei with a central prominent nucleolus. Part of the cells discloses the feature of so-called rhabdoid cells (Fig. 9.7a). The rhabdoid cell, which is a hallmark of MRT (although not present in each tumor), has an eosinophilic or amphophilic cytoplasm containing a spheroid perinuclear inclusion body that consists of intermediate filament whorls, and which is immunoreactive for both, epithelial and mesenchymal markers (mainly cytokeratins 8 and 18, and vimentin; Fig. 9.7b). A large subset, if not the majority, of MRTs are characterized by a recurrent deletion of region 11.2 of the long arm of chromosome 22 (22q11.2) and show truncating frameshift or nonsense mutations of INI1, immunohistochemically detectable by the loss of BAF47 reactivity.

(a) Malignant rhabdoid tumor of the liver. Some of the cells display so-called rhabdoid features (hematoxylin and eosin stain). (b) Part of the rhabdoid cells show excentric intermediate filament staining (vimentin immunostain). (c) Small cell undifferentiated hepatoblastoma with rhabdoid features. Note that the nuclei of a normal bile duct (center) stain for INI1, whereas the tumor cells do not (BAF47 immunostain)
In the liver of infants and small children, polyphenotypic tumors may develop, which are difficult to allocate to either MRT or undifferentiated hepatoblastoma, because they may share features with both tumor types (Wagner et al. 2007; Russo and Biegel 2009). This constellation has been confirmed in a recent study of 11 patients with hepatoblastomas showing normal or minimally increased serum AFP and a SCUD histology. Ten of these patients died of disease progression, and immunostaining revealed that tumors from six of six patients tested were INI1 negative, suggesting that at least some HB-SCUD cases may actually represent a form of MRTs (Trobaugh-Lotrario et al. 2009). This subset of INI1-negative SCUD tumors may be termed, rhabdoid-like tumors or HB-SCUD with rhabdoid features (Fig. 9.7c). Apart from their diagnostic and clinical relevance, these observations may provide clues for a deeper understanding of pathogenic pathways involved in undifferentiated hepatoblastoma. This issue is further discussed in Chap. 3.
9.1.4.5 Grading of Epithelial Types of Hepatoblastoma
So far, grading of hepatoblastomas or at least of their epithelial components is based on the concept of favorable versus unfavorable histology and still requires more studies. Hepatoblastomas with favorable histology are, in principle, purely fetal, well-differentiated neoplasms with minimal recognizable mitotic activity (see above). The typical unfavorable histology comprises a small cell undifferentiated phenotype with or without associated rhabdoid features, and the malignant rhabdoid tumor itself. The remaining tumor types and subtypes are, provisionally, classified as having “less favorable histopathologies,” that is, somewhere in between favorable and unfavorable. Probably, the deeper analysis of large studies will clarify this situation.
9.1.4.6 Mixed Epithelial and Mesenchymal Hepatoblastomas
This type of hepatoblastoma was described in detail in 1967 by use of the terms, epithelial and mesenchymal type or mixed hepatoblastoma (19 out of 35 analyzed hepatoblastomas; Ishak and Glunz 1967). This form of hepatoblastoma is characterized by a complex mixture of epithelial lineages and an immature-looking mesenchyme making part of the tumor itself rather than representing a stromal reaction. Later it was recognized that some of these tumors may contain heterologous components resembling those found in a teratoma. This is the reason why new classifications distinguish mixed hepatoblastomas with or without teratoid features.
9.1.4.6.1 Mixed Epithelial and Mesenchymal Hepatoblastoma Without Teratoid Features
This is the more common variant. Typically, the epithelial component is either fetal or mixed fetal-embryonal. In addition to the connective tissue following the vascular tree or forming septa there are areas of a primitive, sometimes hypercellular mesenchyme intimately admixed with the tumor epithelia. The cells forming this mesenchyme are spindle-shaped (sometimes fibroblastoid) or stellate, with delicate processes and elongated, inconspicuous nuclei. The cytoplasm is scanty. The extracellular matrix contains reticulin fibers and sometimes a myxoid alcianophilic substance. Foci of osteoid with or without mineralization are often seen (Fig. 9.8a). The cells within this tumor osteoid are indistinguishable from osteoblasts, but also stain for epithelial markers in addition to vimentin (cytokeratins). Bone formation may occur in metastases of hepatoblastomas, sometimes indistinguishable from osteosarcoma (Weinberg and Finegold 1983). Cartilage tissue seems to occur in mixed hepatoblastoma (review: Pang 1961), but true (hyaline) cartilaginous tissue is probably not common; it was not noted by Ishak and Glunz (1967) and was observed in the large SIOPEL pathology review only once. Mixed tumors may contain foci of squamous epithelia with formation of concentric pearls. They can express keratohylaine granules, sometimes with marked keratinization and a foreign body reaction with giant cells (Ishak and Glunz 1967).

(a) Osteoid formation in mixed epithelial and mesenchymal hepatoblastoma (hematoxylin and eosin stain). (b) In this mixed epithelial and mesenchymal hepatoblastoma, cells in epithelium and in osteoid contain melanin (brown), a feature of the teratoid variant (hematoxylin and eosin stain). (c) Melanin formation at higher magnification (hematoxylin and eosin stain). (d) Tumor necrosis in melanotic teratoid hepatoblastoma causes accumulation of melanin in macrophages (phagocytosis of melanosomes; hematoxylin and eosin stain)
9.1.4.6.2 Mixed Epithelial and Mesenchymal Hepatoblastoma with Teratoid Features
These are mixed hepatoblastomas, which reveal multiple lines of cell and tissue differentiation in addition to immature mesenchyme, myoid cell lineages, and osteoid tissue. These so-called teratoid lines include intestinal or mucinous epithelium, melanin-containing cells, endocrine elements, immature striated muscle cells, and glioneural tissue (Watanabe et al. 1975; Manivel et al. 1986; Abenoza et al. 1987; Conran et al. 1992; Kim et al. 2001). It is noteworthy that the first pediatric osteoid-containing liver tumor was described in 1898 under the term, teratoma hepatis (Misick 1898), but the term, teratoid hepatoblastoma, was coined in 1986 (Manivel et al. 1986) and is now frequently replaced by the term proposed by the SIOPEL classification, that is, mixed epithelial and mesenchymal hepatoblastoma with teratoid features. This subtype of hepatoblastoma has no connection with teratomas, which are germ cell tumors, but there are very rare instances where teratoid hepatoblastoma occurs together with true hepatic teratoma (Conrad et al. 1993) or with yolk sac tumor (Cross and Variend 1992).
Teratoid hepatoblastomas exhibit the same epithelial components as other hepatoblastomas and they often also contain osteoid, sometimes in an excessive manner (Schlecht et al. 1996). “Teratoid” epithelia come as mucinous or goblet cell formations resembling intestinal or bronchial linings and neuroepithelium. Smooth and striated muscle cells may occur, but are rare. Glioneural components are characterized by a fibrillary glial matrix immunostaining for glial fibrillary acidic protein. Well-differentiated ganglionic cells may be seen. Melanin-containing cells are sometimes prominent (melanotic hepatoblastoma; Fig. 9.8b and c); they contain densely packed melanosomes and are immunoreactive for the marker, HMB45. Melanin-containing granules are also present in neoplastic epithelial cells and are sometimes observed in osteoblast-like cells located within osteoid (Fig. 9.8b) and in macrophages (Fig. 9.8d). Endocrine/neuroendocrine differentiation can occur in teratoid hepatoblastoma (Ruck and Kaiserling 1993). In contrast to other hepatoblastoma types and subtypes, teratoid components are negative for glypican 3 expression (Zynger et al. 2008). Neuroendocrine components and melanin-containing cells seem to be more resistant to chemotherapy than epithelial lineages (Forouhar et al. 1984), but it is not known whether this has an impact for prognosis.
9.1.5 Immunohistochemistry of Hepatoblastomas
The immunohistochemical assessment of hepatoblastomas is a somewhat problematic issue, because these neoplasms display variable immunophenotypes, can express antigens seen in other pediatric malignancies, and hence do not possess a distinct immunohistochemical profile (Ramsay et al. 2008).
Hepatoblastomas express the cytokeratins of hepatocyte lineages, that is, cytokeratins 8 and 18 (Abenoza et al. 1987; Van Eyken et al. 1990; Ramsay et al. 2008). Also the osteoblastoid/osteocytoid cells located within osteoid of mixed hepatoblastomas express cytokeratins 7, 8, and 18 (Van Eyken et al. 1990). The higher differentiation status of fetal-type cells, with production of a canalicular domain, is shown by positivity for polyclonal CEA (Fasano et al. 1998), and by a difference of claudin expression in comparison with embryonal-type tissue (Halasz et al. 2006). Alpha-fetoprotein (AFP) is long known to be expressed in about half of hepatoblastomas (Abenoza et al. 1987; Ramsay et al. 2008), but is consistently lacking in HB-SCUD. In a systemtic study, AFP was detected in about half of the cases of hepatoblastoma (Ramsay et al. 2008).
As outlined elsewhere, hepatoblastomas and related tumors exhibit abnormalities of the Wnt/beta-catenin signaling pathway (Buendia 2002; Yamaoka et al. 2006; Lopez-Terrada et al. 2009a; Fig. 9.9a–c). Subsequent to mutations in the beta-catenin (CTNNB1) gene, beta-catenin bypassing the proteasomal degradation pathway is translocated to the nucleus, where it can be detected by immunohistochemistry, mainly in less differentiated cells (Wei et al. 2000; Yamaoka et al. 2006), a phenomenon which is an important prognostic marker in hepatoblastoma (Park et al. 2001) and is associated with overexpression of cyclin D1 and fibronectin and poorly differentiated histology in hepatoblastoma (Takayasu et al. 2001).

(a) Fetal hepatoblastoma with membranous expression of beta-catenin (in red; beta-catenin immunostain). (b) This hepatoblastoma in part shows cytoplasmic beta-catenin expression (beta-catenin immunostain). (c) In addition to cytoplasmic expression, this tumor displays nuclear reactivity for beta-catenin, indicating beta-catenin gene mutation (beta-catenin immunostain)
Hepatoblastomas can express glypican 3 (GPC3; mutated in the Simpson-Golabi-Behmel tissue overgrowth syndrome), one of the six known members of a heparin sulfate proteoglycan anchored to the cell membrane and detected in hepatic stem cells and being one of the most overexpressed genes in hepatoblastoma by microarray analysis (Luo et al. 2006). In a study of 65 hepatoblastomas, all cases had cytoplasmic immunoreactivity for GPC3 with greater than 90% of cases showing strong and diffuse positivity. GPC3 was present in epithelial lineages (including the small cell undifferentiated subtype), but not in mesenchymal or teratoid components (Zynger et al. 2008).
A subset of hepatoblastomas expresses human choriogonadotropins (hCG) in the tumor cells, clinically causing virilization and precocious puberty (Behrle et al. 1963; review: Nakagawara et al. 1982). Expression of hCG may be associated with concomitant secretion of AFP (Nakagawara et al. 1985). These two markers showed a discordant behavior of the plasma levels during chemotherapy and radiotherapy in two patients (Hung et al. 1963; Braunstein et al. 1972) and a concordant behavior in one (Kumar et al. 1978). Interestingly, one study uncovered that all out of seven hCG-producing hepatoblastomas showed hCG expression in multinucleated syncytiotrophoblast-like giant cells accompanied by round and clear cells with squamous metaplasia, suggesting a choriocarcinoma lineage (Watanabe et al. 1987). Another hormone that may exceptionally be produced by hepatoblastoma is renin, associated with hyperreninemia and hypertension (Moritake et al. 2000).
9.1.6 Growth Patterns, Proliferation, and Differentiation Characteristics in Hepatoblastomas
Hepatoblastomas show various growth patterns. One which is crucial for outcome is angioinvasion that may extensively involve tumor-associated vessels (Fig. 9.10a), small vessels in portal tracks (Fig. 9.10b), and larger veins, including the portal vein (Fig. 9.10c) and branches thereof (Fig. 9.10d). Apart from components of the invasive machinery, these growth patterns require distinct proliferation features. There is a significant association between the histologic type, DNA content, and proliferation, in that fetal tissue is diploid, embryonal tissue is aneuploid, and the proliferative index is higher in embryonal cells than in fetal cells (Rugge et al. 1998; Zerbini et al. 1998; Tsai et al. 2009). The proliferation is commonly lowest in the fetal phenotype (Rugge et al. 1998; Tsai et al. 2009).The proliferative activity of hepatoblastomas has been found to be lower in low stage tumors than in stages III and IV, and was higher in metastases than in primary tumors (Ara et al. 1997).

(a) This hepatoblastoma exhibits massive angioinvasion with formation of tumor plugs (hematoxylin and eosin stain). (b) Hepatoblastoma angioinvasion in small portal tract vessels (hematoxylin and eosin stain). (c) Invasion of the portal vein, with formation of a tumor thrombus. The wall of the vein (pink) is seen to the left and above the center (hematoxylin and eosin stain). (d) Obturation of a small portal vein branch by hepatoblastoma (hematoxylin and eosin stain)
Among cell cycle regulators, cyclins and cyclin-dependent kinase inhibitors (CDKNs) are in part deregulated in hepatoblastomas (Gray et al. 2000). Cyclin D1 acts as a switch at the G1-S checkpoint, and a polymorphism of codon 242 of the cyclin D1 gene affects the age of onset of hepatoblastoma (Pakakasama et al. 2004). It has been found that CDKN2A, CDKN2B, and CDKN2C genes are structurally unmodified in these tumors, whereas CDKN2A, normally silenced in the liver, and CDKN2C are expressed in hepatoblastoma, and cyclin D exhibits a shift in expression (Iolascon et al. 1998). Expression of the cyclin-dependent kinase inhibitor, p27(KIP1) is generally decreased in more aggressive tumors and this has also been found to play a role in hepatoblastoma. P27(KIP1) is not mutated in hepatoblastoma but shows increased transcriptional activity (Hartmann et al. 2000). Well-differentiated low-proliferative fetal tumors markedly express p27, embryonal patterns show a variable expression (less in proliferative areas), and most small cell hepatoblastomas do not express p27 (Brotto and Finegold 2002). Continuous growth of hepatoblastomas is also mediated by high expression of spindle checkpoint kinases, specifically Polo-like kinase 1 (PLK1), being a poor-prognosis indicator (Yamada et al. 2004). The Wnt/beta-catenin signaling pathway, which plays a role in the pathogenesis of hepatoblastoma, affects proliferation and growth, in that stabilized beta-catenin promotes hepatocyte proliferation and inhibits TNFalpha-induced apoptosis (Shang et al. 2004), and is associated with overexpression of cyclin D1 in hepatoblastomas (Takayasu et al. 2001).
Growth regulation of hepatoblastomas involves the insulin-like growth factor-II (IGF2) signaling pathway (Li et al. 1995; Rainier et al. 1995; Yun et al. 1998; Tomizawa and Saisho 2006). IGF2 is a maternally imprinted gene and encodes a fetal peptide hormone that regulates cell proliferation, differentiation, and cell migration. IGF2 acts via binding to the type 1 IGF tyrosine kinase receptor (IGF-1R). IGF2 is expressed in hepatoblastomas and this expression is inversely correlated with the degree of differentiation, lacking in fetal-type cells and being high in embryonal-type cells (Akmal et al. 1995).The allelic expression of IGF2 is regulated by the methylation status of a distinct site (CTCF) in the H19 gene differentially methylated region (DMR) that represents the parental origin of the IGF2 allele: in normal tissues, the maternal allele is unmethylated, whereas the paternal CTCF site is methylated. The maternally expressed H19 gene belongs to an imprinted cluster on chromosome 11p15 and encodes a noncoding mRNA, which controls the expression of the neighboring, paternally transcribed IGF2 gene. Hepatoblastomas show monoallelic expression of H19 (Ross et al. 2000). Loss of imprinting of IGF2 in hepatoblastomas correlates with hypermethylation of the H19 region (Honda et al. 2008), and there is a high frequency of inactivation of H19 in sporadic hepatoblastomas (Fukuzawa et al. 1999). In hepatoblastoma, the IGF2/IGF-IR pathway is interacting with PLAG1, a developmentally regulated zinc finger transcription factor, which positively regulates IGF2 (Van Dyck et al. 2007), and which is overexpressed in hepatoblastomas (Zatkova et al. 2004).
9.1.7 Chemotherapy Effects in Hepatoblastomas
Hepatoblastomas treated by chemotherapy undergo complex changes that may mimic or obscure viable tumor persistence, rendering interpretation of post-chemotherapy resection specimens sometimes difficult (Lowichik et al. 2000).
The main chemotherapy-induced tumor changes comprise necrosis, apoptosis, and inflammatory/immune reactions directed against decaying tumor, fibrosis, vascular changes, and cellular alterations of residual (viable) tumor. Macroscopically, treated tumors appear contracted and nodular, more sharply delineated than native lesions (Fig. 9.11a). Histologically, most of the former tumor usually consists of a fibrous tissue. Often these fibrotic areas are present in the form of nodular hypocellular structures containing blood vessels and old hemorrhage (Fig. 9.11b). It is assumed that the shape of these structures reflects the previous vascular tree of the tumor. Chemotherapy-induced necrosis presents as an eosinophilic and slightly granular mass (Fig. 9.11c), which is sharply demarcated from the adjacent liver by granulation and fibrous tissue containing macrophages and lymphocytes, and sometimes foreign body giant cells. Focal calcifications/mineralizations may be seen.In a study of 17 hepatoblastomas treated with preoperative chemotherapy, there was no obvious correlation between the extent of necrosis and the number of courses of chemotherapy (Saxena et al. 1993). In some cases, necroses contain clusters of epithelial-like cells that are very difficult to identify in regard to their nature, so that the question as to residual viable tumor should be answered with great caution. Chemotherapy induces marked vascular changes, characterized by thickening of the vessel wall and vascular stenosis (Fig. 9.11d). Fetal hepatoblastoma may undergo marked steatosis subsequent to chemotherapy (Fig. 9.11e). Therapy-associated cellular and nuclear anomalies of viable tumor may render the classification of the residual tumor impossible (Fig. 9.11f). A notable feature in tumors treated with chemotherapy is the extensive presence of osteoid (Fig. 9.11g). In a comparative study, osteoid was present in 36% of untreated cases, occupying less than 5% of the surface area, compared with 82% in the treated group (Saxena et al. 1993). Whether this phenomenon reflects a distinct effect of chemotherapy (Heifetz et al. 1997) or is caused by other factors, including sampling effects, has not yet been clarified. Keratinizing squamous epithelia are a typical chemotherapy effect (Fig. 9.11h).


(a) Macroscopic features of resected hepatoblastoma post-chemotherapy (cut surface). The tumor appears contracted, consists of yellowish nodules, and reveals white areas of fibrosis/scarring. (b) Nodular areas of fibrosis and perifocal liver atrophy after chemotherapy (hematoxylin and eosin stain). (c) Hepatoblastoma with post-chemotherapy necrosis (hematoxylin and eosin stain). (d) Chemotherapy-induced vascular change with marked thickening of the vessel wall and vascular stenosis (center; hematoxylin and eosin stain). (e) Marked fatty change of fetal hepatoblastoma after chemotherapy (hematoxylin and eosin stain). (f) Prominent cellular and nuclear atypia in hepatoblastoma after chemotherapy (hematoxylin and eosin stain). (g) Post-chemotherapy hepatoblastoma rich in osteoid. The nature of the small epithelial focus to the left of the red osteoid (residual tumor vs. atrophic liver) is difficult to assess (hematoxylin and eosin stain). (h) Several foci of squamous epithelium after chemotherapy of hepatoblastoma (hematoxylin and eosin stain)
Several post-chemotherapy histological features apparently having an impact on outcome have been described in hepatoblastoma. They comprise vascular invasion in the tumor capsule (risk factor for subsequent metastatic disease), necrosis greater than 75% (favorable prognostic indicator), and increased proliferative activity in residual tumor (poor prognostic indicator). No reproducible effects were found for marked osteoid production, fibroblastic proliferation around necrosis, and intimal thickening, occlusion or hyalinosis of blood vessels (review: Lowichik et al. 2000).
9.1.8 Cholangioblastic Hepatoblastoma (Hepatoblastomas with Cholangioblastic Features) and “Ductal Plate Tumors”
A small subset of hepatoblastomas exhibits, mostly at the periphery of the otherwise typical hepatoblast formations, cytokeratin 19-positive bile duct cells and even duct-like profiles in a focal distribution pattern (Fig. 9.12a) (Zimmermann 2002; Libbrecht et al. 2003). As these biliary epithelial cells are remote from preexisting ductular and ductal cells of the host liver, but rather constitute part of the tumor itself, these lesions have been proposed to be termed, hepatoblastoma with cholangioblastic features or cholangioblastic hepatoblastoma (Zimmermann 2002, 2005). The cholangioblastic features may not be recognizable with ease in conventional sections, but immunostaining for cholangiocyte lineage markers will uncover the cells of interest (Fig. 9.12b). In other situations, organoid tumors reveal numerous and small nodules consisting of immature hepatoid cells, encircled by a thin rim of biliary cells and sometimes with slits resulting in a double layer of cholangiocytes mimicking an abnormal ductal plate (so-called “ductal plate tumor”; Gornicka et al. 2001; Zimmermann 2002; Fig. 9.12c). So far, cholangioblastic features have predominantly been detected in fetal-type and embryonal-hepatoblastoma, but the proportion of tumors exhibting these features and the prognostic impact of this change have not been elucidated so far.

(a) Hepatoblastoma with cholangioblastic features. Note the bile duct-like profiles in close association with hepatoblastoma cells (hematoxylin and eosin stain). (b) In this cholangioblastic hepatoblastoma, numerous cells are reactive for a bile duct cell marker (cytokeratin 19 immunostain). (c) Ductal plate tumor consists of small hepatoid nodules (“liverlets”) associated with cholangiocellular profiles mimicking components of a ductal plate (the latter in red; cytokeratin 19 immunostain)
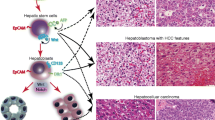
9.1.9 Transitional Liver Cell Tumor (TLCT)
Transitional liver cell tumor (TLCT) is a recently described malignant liver cell neoplasm that chiefly occurs in older children and young adolescents (Prokurat et al. 2002). TLCTs have a rather characteristic clinical presentation, histopathology, immunohistochemistry, and treatment response. The tumors are highly aggressive and usually present as large neoplasms associated with high or very high serum AFP levels. Most of the lesions reported so far were initially diagnosed as hepatoblastoma in needle biopsies, and the histology was later reviewed owing to the very unfavorable outcome after chemotherapy designed for hepatoblastoma. Histologically, TLCT have a rather complex pattern, with hepatoblastoma-like cells, cells resembling those of HCC, and intermediate cell forms (Fig. 9.13a). Multinucleated giant cells are a typical feature (Fig. 9.13b). The cellular features of larger cells vary from cholangiocyte-like elements (Fig. 9.13c) to immature hepatoid cells (Fig. 9.13d). A biliary phenotype is visualized by epithelial membrane antigen (EMA) staining (Fig. 9.13e). Part of the tumors express beta-catenin, with a mixed nuclear and cytoplasmic expression pattern (Fig. 9.13f). The term, transitional, has been proposed based on the hypothesis that the relevant tumor cell might be located between a hepatoblast and a hepatocyte, but this has to be analyzed in greater depth in the future, and also with molecular methods.

(a) Transitional liver cell tumor consisting of hepatoblastoma-like cells and larger cells (hematoxylin and eosin stain). (b) Transitional liver cell tumors frequently show multinucleated giant cells (hematoxylin and eosin stain). (c) This transitional liver cell tumor exhibits cholangiocyte-like cells (hematoxylin and eosin stain). (d) Transitional liver cell tumor with large and poorly differentiated hepatoid cells (hematoxylin and eosin stain). (e) The cholangiocyte lineage in this transitional liver cell tumor is visualized by focal positivity for epithelial membrane antigen (reactivity in red; EMA immunostain). (f) Nuclear and cytoplasmic expression of beta-catenin in transitional liver cell tumor (reactivity in red; beta-catenin immunostain)
9.1.10 Tumors Possibly Related to the Hepatoblastoma Tumor Family
Within pathology reviews of large international prospective studies on pediatric liver tumors, most of the neoplasms are classifiable, but a minority of the lesions will not fit into known categories and may, therefore, be classified as NOS (not otherwise specified) until a precise nosological assignment is possible. In addition, there are novel tumor entities that seem to share certain features with hepatoblastomas, although the exact relation between the lesions is not yet known. According to this author’s view, one of these tumors may be nested stromal epithelial tumor of the liver (Heerema-McKenney et al. 2005; Meir et al. 2009; Rod et al. 2009), also termed desmoplastic nested spindle cell tumor of the liver (Hill et al. 2005). This is a hepatic neoplasm in infants and older children and in young adolescents, associated with Cushing syndrome in some of the patients and with a variable course, one patient so far showing recurrence and extrahepatic metastasis (Brodsky et al. 2008). Histologically, nests of epithelial-like cells surrounded by spindle cells showing calcifications and osteoid are a hallmark (Fig. 9.14a and b). The epithelial cells express cytokeratin 8 (Fig. 9.14c), but not a hepatocyte marker, Hep Par1 (Fig. 9.14d). Interestingly, the nested cells exhibit nuclear and cytoplasmic positivity for beta-catenin (own observations; Fig. 9.14e).The spindle cells are positive for vimentin and alpha-smooth muscle actin (myofibroblasts; Fig. 9.14f), suggesting abnormal epithelial-mesenchymal transition and resembling the morphology of a deranged liver bud. The epithelial nest cells express ACTH (Heerema-McKenney et al. 2005; Rod et al. 2009) and corticotropin-releasing hormone (Rod et al. 2009), suggesting an ectopic ACTH syndrome (EAS; Rod et al. 2009).


(a) Epithelial cell clusters surrounded by spindle cells in nested stromal epithelial tumor of the liver (hematoxylin and eosin stain). (b) The nested structures may contain psammomatous calcifications and osteoid (hematoxylin and eosin stain). (c) Epithelia of the nested structures are positive for cytokeratin (reactivity in brown; cytokeratin 8 immunostain). (d) Normal hepatic parenchymal cells express the hepatocyte marker, Hep Par 1 (brown; left bottom corner), In contrast, epithelia of nested structures (to the right and top) are negative (Hep Par 1 immunostain). (e) Beta-catenin reactivity (in part nuclear) in nested stromal epithelial tumor (beta-catenin immunostain). (f) The spindle cells surrounding the epithelial clusters are positive for alpha-smooth muscle actin (brown; alphaSMA immunostain)
9.2 Pediatric Hepatocellular Carcinoma
9.2.1 Definition and Epidemiology
Hepatocellular carcinoma (HCC) is a primary malignant tumor of the liver derived from hepatocytes. HCC accounts for approximately 21% of all malignant liver tumors diagnosed in children (Weinberg and Finegold 1983; Stocker 2001), but for only a minority of pediatric solid tumors, as less than 1% of HCCs are diagnosed in patients younger than 20 years of age (Carriaga and Henson 1995). So far, it is not yet known whether pediatric HCC is the same or a disease different from adult-type HCC (Czauderna 2002). HCC is more often diagnosed in males, and mostly in children older than 10 years of age, representing the majority (87%) of malignant liver tumors diagnosed in adolescents (LaBrecque 1996; Darbari et al. 2003; SEER 2006). However, typical HCC can also occur in young children, including infants. Clinical presentation typically includes hepatomegaly and a palpable abdominal mass, often associated with abdominal pain, anorexia, abnormal liver enzymes, and AFP elevation, commonly used as a tumor marker (Ishak and Glunz 1967; Lack et al. 1983).
9.2.2 Etiology
Incidence of HCC is higher in children living in endemic hepatitis B regions (Africa and South-East Asia) (Bellani and Massimino 1993; Moore and Hesseling 1997), commonly acquired perinatally, and where incidence rates have significantly decreased due to the implementation of immunization programs (Montesano 2002; Chang 2003). Other common etiological factors associated with adult HCC, such as underlying liver disease and cirrhosis, HCV infection, chronic alcohol abuse, and exposure to aflatoxin B1, are not relevant etiologic factors for pediatric HCC. Constitutional genetic and metabolic abnormalities are more often associated with HCC diagnosed in children from countries with low HBV endemic rates (Table 9.2). HCC was reported in approximately 18% of children with hereditary tyrosinemia type 1 (fumarylacetoacetate hydrolase deficiency) (Weinberg et al. 1976; Demers et al. 2003) before therapy was available. Glycogen storage diseases, particularly type 1a, are also associated with the development of hepatic tumors in children, including HCC (Coire et al. 1987; Bianchi 1993; Siciliano et al. 2000).
Familial adenomatous polyposis (FAP) caused by germline mutation of the adenomatous polyposis coli (APC) gene is typically associated with the development of hepatoblastoma in children, and has also been implicated in the pathogenesis of HCC and fibrolamellar carcinoma (Kingston et al. 1982; Giardello et al. 1996), suggesting that APC mutations may confer a general predisposition to tumorigenesis in the liver (Thomas et al. 2003). APC and beta-catenin (CTNNB1) mutations have been identified in benign precursor lesions, hepatoblastomas, and HCC (Cieply et al. 2009), and the resulting canonical Wnt pathway constitutional activation is now considerd a common oncogenic pathway in liver tumors (Zucman-Rossi et al. 2006).
HCC and cholangiocarcinomas have been observed in patients with familial cholestatic syndromes, including Alagille syndrome (Kaufman et al. 1987; Rabinovitz et al. 1989), and extrahepatic biliary atresia with both HCC and HB in children (Taat et al. 2004). HCC has also been described in cirrhotic livers of children following parenteral nutrition (Vileisis et al. 1982). Finally, pediatric liver tumors, including HCC have been reported in association with neurofibromatosis, ataxia-telangiectasia (Ettinger and Freeman 1979; Weinstein et al. 1985; Geoffroy-Perez et al. 2001; Ucar et al. 2005) and in patients with Fanconi’s anemia treated with anabolic steroids, with tumor regression observed with steroids withdrawal (Abbondanzo et al. 1986; Touraine et al. 1993).
9.2.3 Pathology of Adult-Type Hepatocellular Carcinoma
9.2.3.1 Gross Presentation
Macroscopic appearance of HCC depends on the presence of underlying liver disease, size of the tumor, and intrahepatic vascular spread. The presence of cirrhosis is much less common in pediatric than in adult HCC patients. HCC can occur as a solitary, circumscribed mass in one lobe of the liver, more frequently the right, but more often involves both lobes of the liver. Tumor lesions growing without underlying cirrhosis tend to be large, nonencapsulated and exhibit an infiltrative growth pattern, with intrahepatic metastases and numerous tumor nodules (Fig. 9.15). The cut surface is usually soft (with exception of the fibrolamellar variant) and bile-stained, different from hepatoblastoma, with areas of necrosis and hemorrhage. Four main gross growth patterns are recognized: expanding (“pushing”) lesions; pedunculated (“hanging”) lesions; invading lesions; and multifocal lesions (review: Zimmermann 2000). Vascular spread is common and the portal veins, hepatic veins or vena cava may be involved; however, intrahepatic metastases are usually through the portal veins (Ishak and Goodman 1999). Invasion of the biliary ducts is not common, but may cause biliary obstruction. Extrahepatic metastatic spread, often to the lungs and rarely to the brain, occurs via the hepatic veins (Katzenstein et al. 2002).

Hepatocellular carcinoma in a 16 year-old with multiple tumor nodules, necrotic foci, and vascular invasion
9.2.3.2 Histology of Adult-Type Hepatocellular Carcinoma
Microscopically, HCC diagnosed in children are overall similar to those in adults (Ishak and Goodman 1999; Farhi et al. 1983; Lack et al. 1983). Tumor cells variably resemble hepatocytes depending on the degree of differentiation. The nuclear/cytoplasmic ratio is elevated, and nuclei are usually prominent, irregular, and hyperchromatic. Cytoplasm is usually eosinophilic and granular, but may also appear clear, contain glycogen, fat, Mallory bodies, or ground-glass-like inclusions. Bile canaliculi are present in approximately half of the tumors. The most common architectural pattern in well and moderately well-differentiated HCC is the trabecular (plate-like) pattern, with tumor cells growing in cords separated by sinusoid-like blood spaces (Fig. 9.16a and b). A pseudoglandular and acinar pattern is commonly found admixed with the trabecular pattern. Cells can also be arranged as sheets without sinusoids (solid pattern).

(a) Moderately differentiated HCC in a patient with hereditary Tyrosinemia. (b) Poorly differentiated HCC in a 7 year-old, arising in a non-cirrhotic liver. Tumor cells grow in cords and macrotrabecular array, forming pseudoacini and tubules focally. Note the abundant mitoses. (c) Glypican staining of the same tumor showing variable cytoplasmic positivity. (d) β-catenin staining demonstrating strong cytoplasmic positivity in the tumor and membranous staining in the surrounding compressed liver
According to histological grade, HCC can be classified into well-differentiated, moderately differentiated, poorly differentiated, and undifferentiated types (Hirohashi and Ishak 2000). Edmondson and Steiner (Edmondson and Steiner 1954) proposed a scale of I to IV with increasing nuclear irregularity, hyperchromatism, and nuclear/cytoplasmic ratio, associated with decreasing differentiation. Grading can also be done by nuclear features alone or in combination with microvascular invasion (Nzeako et al. 1995; Ishak and Goodman 1999). Reporting histological grade has been recommended for adult HCC resection specimens; however, the association between grade and prognosis and its clinical relevance is not entirely clear (Lai et al. 1979; Dabbs and Geisinger 2004).
Immunohistochemical stains used in combination with histomorphology, even though not helpful in most cases to distinguish HCC from hepatoblastoma or benign lesions, may be useful to differentiate HCC from other tumor types, including cholangiocarcinoma. HepPar-1 (Hepatocyte Paraffin 1), although not completely specific for hepatocytes, shows characteristic cytoplasmic granular staining in approximately 90% of HCCs. Identification of a canalicular pattern using carcinoembryotic antigen (polyclonal CEA) is also useful to differentiate HCC from other malignancies. Alpha-fetoprotein (AFP) is usually elevated in the serum of hepatoblastoma and HCC patients, and often focally present in the tumor cells. Hepatocytes express cytokeratins 8 and 18, while biliary epithelium expresses CK7, CK19, CK8, and CK18, but aberrant cytokeratin expression in HCC and cholangiocarcinoma tumor cells limits the use of these antibodies for tumor classification (Wu et al. 1996). CD34 positive immunostaining of the sinusoids can be helpful to distinguish well differentiated HCC from regenerative nodules, but not adenomas (Gouysse et al. 2004; Varma and Cohen 2004). Glypican 3 is a novel serum and histochemical marker for hepatocellular carcinoma identified by expression profiling. Glypican 3 (GPC3), a heparan sulfate proteoglycan, is expressed at a markedly elevated level in hepatocellular carcinoma and a promising marker for hepatocellular carcinoma in routine histological examination, and as a potential target in monoclonal antibody-based hepatocellular carcinoma therapy (Fig. 9.16c) (Capurro et al. 2003; Yamauchi et al. 2005). Beta-catenin immunostaining shows strong cytoplasmic positivity in HCC (Fig. 9.16d).
9.2.4 Fibrolamellar Hepatocellular Carcinoma
The fibrolamellar variant of HCC constitutes a distinctive clinical and histological variant of HCC that occurs almost exclusively in adolescents and young adults, accounting for almost a third of HCCs diagnosed in patients below 20 years of age (Farhi et al. 1983; Lack et al. 1983; Haas et al. 1989; El-Serag et al. 2003). The incidence of fibrolamellar HCC is similar in males and females, and characteristically presents in patients without cirrhosis, hepatitis, metabolic, or other underlying chronic liver disease. These tumors are usually slow growing, present with minimal elevation of or normal serum AFP, and are often resectable or curable by transplantation, with an overall 5 year survival that exceeds 55% in most series (Soreide et al. 1986; Pinna et al. 1997). However, and although originally thought to carry a better prognosis, this is likely due to the absence of associated cirrhosis (Kakar et al. 2005). A recent study of the Pediatric Intergroup Hepatoma Protocol INT-0098 demonstrated, unlike previous reports, that children with fibrolamellar HCC do not have a favorable prognosis and do not respond any differently to current therapeutic regimens than patients with typical HCC at similar stage (Katzenstein et al. 2003).
On gross examination they are well circumscribed, firm masses with characteristic radiating fibrous septae, resembling focal nodular hyperplasia (Fig. 9.17a). Their characteristic microscopic appearance consists of cords and nests of large neoplastic hepatocytes with granular oncocytic cytoplasm, separated by dense hyalinized collagen bands (Fig. 9.17b). Some of the cells contain so-called pale bodies.

(a) Well circumscribed, sclerotic appearing fibrolamellar carcinoma, with characteristic central radiating scar. (b) Fibrolamellar carcinoma with characteristic dense fibrous stroma separating cords and nests of neoplastic hepatocytes with abundant eosinophilic cytoplasm
Foci of conventional HCC and of adjacent areas of focal nodular hyperplasia have been described associated with this lesion (Saul et al. 1987; Berman et al. 1988). Staining for CK7, CEA, fibrinogen, and copper is commonly found in fibrolamellar HCC (Lefkowitch et al. 1983).
9.2.5 Differential Diagnoses
The most common differential diagnosis of pediatric HCC is hepatoblastoma and rarely other metastatic lesions, including carcinomas in older children. Cytologic atypia may be variable in pediatric HCC, as in adult HCC, but cells are in general larger than in hepatoblastoma, bile producing, with more prominent cytologic atypia and nuclear pleomorphism, and often abnormal mitoses present. Other characteristic histologic features of hepatoblastoma, such as coexistence of histologic patters, extramedullary hematopoiesis, and mesenchymal components, are not usually seen in HCC.
Hepatic adenomas occur in older children, and may be challenging to differentiate from other lesions, including well-differentiated HCC, particularly in small biopsies. Adenomas arise from non-cirrhotic livers and may be multiple, such as those seen in patients with glycogen storage disorders. In resection specimens a pushing border and, occasionally a capsule, may be identified surrounding sheets and cords of hepatocytes larger and usually paler than surrounding hepatocytes, and commonly containing cytoplasmic glycogen or fat. Immunohistochemical staining for proliferation markers such as Ki67 or endothelial markers (CD34) may be diagnostically useful (Libbrecht et al. 2001; Gouysse et al. 2004). Hepatic adenomas may sometimes be difficult to differentiate from focal nodular hyperplasia, particularly in small biopsies.
Fibrolamellar HCC can be clinically and radiographically difficult to differentiate from focal nodular hyperplasia, due to its indolent course and typical central scar. However, in most cases microscopic diagnosis can be easily made by its distinctive histologic features.
Lastly, it is worth mentioning that patients with both hepatoblastoma and HCC have been simultaneously described, and simultaneous presence of adenomas and HCC is not uncommon, particularly in patients with glycogen storage disorders (Parker et al. 1981; Coire et al. 1987).
9.2.6 New Knowledge on Pathogenic/Molecular Pathways
The molecular genetic alterations associated with the multistep process of hepatocarcinogenesis have been extensively studied (Bannasch 1996; Buendia 2000; Thorgeirsson and Grisham 2002, review: Zimmermann 2006; Aravalli et al. 2008). However, little knowledge is available regarding the biology of pediatric HCC, a different and heterogeneous group of tumors often occurring without underlying liver disease, and not associated with common HCC etiological factors seen in adults, with the exception of HBV infection. Hepatitis B virus is a hepatotropic DNA hepadnavirus associated with a lifetime risk of HCC for children infected at birth, of approximately 50%. Integration of HBV in the host genome has been proposed as a possible oncogenic mechanism for a proportion of patients (Matsubara and Tokino 1990; Wang et al. 1990), possibly through chromosomal instability (Bréchot 2004).
Genetic studies of mostly adult HCC have demonstrated multiple chromosomal abnormalities, predominantly losses, in contrast to HB with few characteristic chromosomal changes, commonly trisomies (Wong et al. 2000). Increased chromosomal instability has been reported in tumors associated with hepatitis B virus infection. Most common chromosomal losses are on chromosomes 17p, 13q, 9p, 6q, and 16p. LOH is common in adult HCC and frequently involves tumor suppressor genes, with deletions reported most commonly at 8p (48%), 17p (45%), 4q (38%), 1p (33%), 13q, 16q, 6q, 16p, 1q, and 9p with a frequency higher than 20% (Buendia 2002; Tornillo et al. 2002).
Alterations common to HCC and hepatoblastoma include gain of chromosomes 1q, 8q, and 17q, and loss of 4q. Another important common feature shared by the two tumor types is the frequent activation of Wnt/beta-catenin signaling by stabilizing mutations of beta-catenin (Buendia 2002) (Fig. 9.1e). Recent application of array CGH technology (Lopez-Terrada et al. 2009b) has been proposed as a genomic profiling tool applicable to surgical specimens, and useful for the differential diagnosis of HCC versus hepatoblastoma.
Genetic analysis of fibrolamellar HCC demonstrated fewer chromosomal abnormalities compared with those reported in literature for conventional hepatocellular carcinoma, with most common abnormalities on chromosomes 7 and 8. Fibrolamellar carcinomas with chromosomal changes appear to behave more aggressively than cases with normal karyotypes (Wilkens et al. 2000; Kakar et al. 2009). Epigenetic studies of a series of fibrolamellar HCCs demonstrated that genomic instability is rare in this variant when compared with viral-associated hepatocellular carcinomas (Vivekanandan et al. 2009). Signaling pathway analysis has demonstrated overexpression of genes in the RAS, MAPK, PIK3, and xenobiotic degradation pathways in this group of HCCs (Kannangai et al. 2007).
In recent years, numerous studies have aimed at identifying critical signaling pathways involved in hepatocarcinogenesis, and particularly hepatitis-associated HCC. Some of the most relevant, aberrantly activated pathways include the P53 pathway, mitogen-activated protein kinase (MAPK), Wnt/beta-catenin, epidermal growth factor (EGF), and transforming growth factor-beta (TGFbeta) pathways (Anders et al. 2003; Lee and Thorgeirsson 2005). Several gene expression profiling studies have specifically addressed differences between clinical HCC subtypes and searched for biomarkers that could serve as prognostic predictors, or therapeutic targets (Lau et al. 2000; Graveel et al. 2001; Okabe et al. 2001; Shirota et al. 2001; Delpuech et al. 2002; Lee and Thorgeirsson 2004). A recent study identified differentially expressed genes in HCC versus hepatoblastoma, and between tumors and adjacent liver (Luo et al. 2006). Other high throughput technologies (proteomics, metabolomics, micro RNA profiling) have also been applied recently to study the biology of HCC (Blanc et al. 2005; Li et al. 2005; Yang et al. 2008).
9.2.7 Conclusion
Unfortunately, most of these studies are aimed at investigating adult HCC, and it is not clear how much of what we have learned from these applies to those diagnosed in pediatric patients. Pediatric HCC is a rare, heterogeneous disease, and very little is understood today regarding how genetic predisposition, metabolic disorders, cholestasis, or even potential exposures during infancy and childhood, participate in the carcinogenesis of these rare group of tumors.
The diagnosis and clinical management of pediatric HCC still represents a tremendous challenge. Only by collaborative efforts and by incorporating new biologic parameters and traditional diagnostic algorithms, similar to other pediatric neoplasms, will it be possible to improve diagnosis, clinical stratification, and sucessfull treatment of pediatric HCC patients.
References
Abbondanzo SL, Manz HJ, Klappenbach RS et al (1986) Hepatocellular carcinoma in an 11-year-old girl with Fanconi’s anemia. Report of a case and review of the literature. Am J Pediatr Hematol Oncol 8:334–337
Abenoza P, Manivel JC, Wick MR et al (1987) Hepatoblastoma: an immunohistochemical and ultrastructural study. Hum Pathol 18:1025–1035
Akmal SN, Yun K, MacLay J et al (1995) Insulin-like growth factor 2 and insulin-like factor binding protein 2 expression in hepatoblastoma. Hum Pathol 26:846–851
Anders RA, Yerian LM, Tretiakova M et al (2003) cDNA microarray analysis of macroregenerative and dysplastic nodules in end-stage hepatitis C virus-induced cirrhosis. Am J Pathol 162:991–1000
Ara T, Fukuzawa M, Oue T et al (1997) Immunohistochemical assessment of the MIB-1 labeling index in human hepatoblastoma and its prognostic relevance. J Pediatr Surg 32:1690–1694
Aravalli RN, Steer CJ, Cressman EN et al (2008) Molecular mechanisms of hepatocellular carcinoma. Hepatology 48:2047–2063
Bannasch P (1996) Pathogenesis of hepatocellular carcinoma: sequential cellular, molecular, and metabolic changes. Prog Liver Dis 14:161–197
Behrle FC, Mantz FA, Olson RL et al (1963) Virilization accompanying hepatoblastoma. Pediatrics 32:265–271
Bellani FF, Massimino M (1993) Liver tumors in childhood: epidemiology and clinics. J Surg Oncol Suppl 3:119–121
Berman MA, Burnham JA, Sheahan DG et al (1988) Fibrolamellar carcinoma of the liver: an immunohistochemical study of nineteen cases and a review of the literature. Hum Pathol 19:784–794
Bianchi L (1993) Glycogen storage disease I and hepatocellular tumours. Eur J Pediatr 152(Suppl 1):S63–S70
Blanc JF, Lalanne C, Plomion C et al (2005) Proteomic analysis of differently expressed proteins in hepatocellular carcinoma developed in patients with chronic viral hepatitis C. Proteomics 5:3778–3789
Borman JB, Harbott AJ, Morris D (1961) Hepatic lobectomy in infancy for hepatoblastoma. Br J Surg 49:11–13
Braunstein GD, Bridson WE, Glass A et al (1972) In vivo and in vitro production of human chorionic gonadotropin and alpha-fetoprotein by a virilizing hepatoblastoma. J Clin Endocrinol 35:857–862
Bréchot C (2004) Pathogenesis of hepatitis B virus-related hepatocellular carcinoma: old and new paradigms. Gastroenterology 127(Suppl 1):S56–S61
Brodsky SV, Sandoval C, Sharma N et al (2008) Recurrent nested stromal epithelial tumor of the liver with extrahepatic metastasis: case report and review of literature. Pediatr Dev Pathol 11:469–473
Brotto M, Finegold MJ (2002) Distinct patterns of p27/KIP1 gene expression in hepatoblastoma and prognostic implications with correlation before and after chemotherapy. Hum Pathol 33:198–205
Buendia MA (2000) Genetics of hepatocellular carcinoma. Semin Cancer Biol 10:185–200
Buendia MA (2002) Genetic alterations in hepatoblastoma and hepatocellular carcinoma: common and distinctive aspects. Med Pediatr Oncol 39:530–535
Capurro M, Wanless IR, Sherman M et al (2003) Glypican-3: a novel serum and histochemical marker for hepatocelular carcinoma. Gastroenterology 125:89–97
Carriaga MT, Henson DE (1995) Liver, gallbladder, exrahepatic bile ducts, and pancreas. Cancer 75(Suppl):171–190
Chandra CR, Stocker JT (1992) The liver, gallbladder and biliary tract. In: Stocker JT (ed) Pediatric pathology. JB Lippincott, Philadelphia
Chang MH (2003) Decreasing incidence of hepatocellular carcinoma among children following universal hepatitis B immunization. Liver Int 23:309–314
Cieply B, Zeng G, Proverbs-Singh T et al (2009) Unique phenotype of hepatocellular cancers with exon-3 mutations in beta-catenin gene. Hepatology 49:821–831
Coire CI, Qizilbash AH, Castelli MF et al (1987) Hepatic adenomata in type Ia glycogen storage disease. Arch Pathol Lab Med 111:166–169
Conrad RJ, Gribbin D, Walker NI et al (1993) Combined cystic teratoma and hepatoblastoma of the liver. Probable divergent differentiation of an uncommitted hepatic precursor cell. Cancer 72:2910–2913
Conran RM, Hitchcock CL, Waclawiw MA et al (1992) Hepatoblastoma: the prognostic significance of histologic type. Pediatr Pathol 12:167–183
Cross SS, Variend S (1992) Combined hepatoblastoma and yolk sac tumor of the liver. Cancer 69:1323–1326
Czauderna P (2002) Adult type vs. childhood hepatocellular carcinoma – are they the same of different lesions? Med Pediatr Oncol 39:519–523
Dabbs DJ, Geisinger KR (2004) Recommendations for the reporting of tissues removed as part of the surgical treatment of malignant liver tumors. Hum Pathol 35:1315–1323
Dachman AH, Pakter RL, Ros PR et al (1987) Hepatoblastoma: radiologic-pathologic correlation in 50 cases. Radiology 164:15–19
D’Antiga L, Vallortigara F, Cillo U et al (2007) Features predicting unresectability in hepatoblastoma. Cancer 110:1050–1057
Darbari A, Sabin KM, Shapiro CN et al (2003) Epidemiology of primary hepatic malignancies in U.S. children. Hepatology 38:560–566
Davies JQ, de la Hall PM, Kaschula RO et al (2004) Hepatoblastoma – evolution of management and outcome and significance of histology of the resected tumor. A 31-year experience with 40 cases. J Pediatr Surg 39:1321–1327
Dehner LP (1978) Hepatic tumors in the pediatric age group: a distinctive clinicopathologic spectrum. Perspect Pediatr Pathol 4:217–268
Dehner LP, Manivel JC (1988) Hepatoblastoma: an analysis of the relationship between morphologic subtypes and prognosis. Am J Pediatr Hematol Oncol 10:301–307
De Ioris M, Brugieres L, Zimmermann A et al (2008) Hepatoblastoma with a low serum alpha-fetoprotein level at diagnosis: the SIOPEL group experience. Eur J Cancer 44:545–550
Delpuech O, Trabut JB, Carnot F et al (2002) Identification, using cDNA macroarray analysis, of distinct gene expression profiles associated with pathological and virological features of hepatocellular carcinoma. Oncogene 21:2926–2937
Demers SI, Russo P, Lettre F et al (2003) Frequent mutation reversion inversely correlates with clinical severity in a genetic liver disease, hereditary tyrosinemia. Hum Pathol 34:1313–1320
Dicken BJ, Bigam DL, Lees GM (2004) Association between surgical margins and long-term outcome in advanced hepatoblastoma. J Pediatr Surg 39:721–725
Doss BJ, Vicari J, Jacques SM (1998) Placental involvement in congenital hepatoblastoma. Pediatr Dev Pathol 1:538–542
Douglass EC, Reynolds M, Finegold M et al (1993) Cisplatin, vincristine, and fluorouracil therapy for hepatoblastoma: a Pediatric Oncology Group study. J Clin Oncol 11:96–99
Edmondson HA, Steiner PE (1954) Primary carcinoma of the liver: a study of 100 cases among 48,900 necropsies. Cancer 7:462–503
Edmondson HA (1956) Differential diagnosis of tumors and tumor-like lesions of liver in infancy and childhood. AMA J Dis Child 91:168–186
El-Serag HB, Davila JA, Petersen NJ et al (2003) The continuing increase in the incidence of hepatocellular carcinoma in the United States: an update. Ann Intern Med 139:817–823
Ettinger LJ, Freeman AI (1979) Hepatoma in a child with neurofibromatosis. Am J Dis Child 133:528–531
Farhi DC, Shikes RH, Murani PJ et al (1983) Hepatocellular carcinoma in young people. Cancer 52:1516–1525
Fasano M, Theise ND, Nalesnik M et al (1998) Immunohistochemical evaluation of hepatoblastomas with use of hepatocyte-specific marker, hepatocyte paraffin 1, and the polyclonal anti-carcinoembryonic antigen. Mod Pathol 11:934–938
Feusner JH, Krailo MD, Haas JE et al (1993) Treatment of pulmonary metastases of initial stage I hepatoblastoma in childhood. Report from the Childrens Cancer Group. Cancer 71:859–864
Finegold MJ, Lopez-Terrada DH, Bowen J et al (2007) Protocol for the examination of specimens from pediatric patients with hepatoblastoma. Arch Pathol Lab Med 131:520–529
Finegold MJ, Egler RA, Goss JA et al (2008) Liver tumors: pediatric population. Liver Trans 14:1545–1556
Forouhar FA, Quinn JJ, Cooke R et al (1984) The effect of chemotherapy on hepatoblastoma. Arch Pathol Lab Med 108:311–314
Fukuzawa R, Umezawa A, Ochi K et al (1999) High frequency of inactivation of the imprinted H19 gene in “sporadic” hepatoblastoma. Int J Cancer 82:490–497
Garcés-Inigo EF, Leung R, Sebire NJ et al (2009) Extrarenal rhabdoid tumours outside the central nervous system in infancy. Pediatr Radiol 39:817–822
Geoffroy-Perez B, Janin N, Ossian K et al (2001) Cancer risk in heterozygotes for ataxia-telangiectasia. Int J Cancer 93:288–293
Giardello FM, Petersen GM, Brensinger JD et al (1996) Hepatoblastoma and APC gene mutation in familial adenomatous polyposis. Gut 39:867–869
Gonzalez-Crussi F, Upton MP, Maurer HS (1982) Hepatoblastoma. Attempt at characterization of histologic subtypes. Am J Surg Pathol 6:599–612
Gonzalez-Crussi F (1991) Case 1 Undifferentiated small cell (“anaplastic”) hepatoblastoma. Pediatr Pathol 11:155–162
Gornicka B, Ziarkiewicz-Wroblewska B, Michalowicz B et al (2001) Immature hepatic tumor of bimodal differentiation in a young adult patient: a novel lesion expressing β-catenin and mimicking a distinct phase of hepatogenesis. J Hepatol 34:955–961
Gouysse G, Franchon S, Hervieu V et al (2004) Endothelial cell differentiation in hepatocellular adenomas: implications for histopathological diagnosis. J Hepatol 41:259–266
Graveel CR, Jatkoe T, Madore SJ et al (2001) Expression profiling and identification of npvel genes in hepatocellular carcinomas. Oncogene 20:2704–2712
Gray SG, Hartmann W, Eriksson T et al (2000) Expression of genes involved with cell cycle control, cell growth and chromatin modification are altered in hepatoblastomas. Int J Mol Med 6:161–169
Haas JE, Muczynski KA, Krailo M et al (1989) Histopathology and prognosis in childhood hepatoblastoma and hepatocarcinoma. Cancer 64:1082–1095
Haas JE, Feusner JH, Finegold MJ (2001) Small cell undifferentiated histology in hepatoblastoma may be unfavorable. Cancer 92:3130–3134
Halasz J, Holczbauer A, Paska C et al (2006) Claudin-1 and claudin-2 differentiate fetal from embryonal components in human hepatoblastoma. Hum Pathol 37:555–561
Hansen K, Bagtas J, Mark HF et al (1992) Undifferentiated small cell hepatoblastoma with a unique chromosomal translocation: a case report. Pediatr Pathol 12:457–462
Hartmann W, Waha A, Koch A et al (2000) p57(KIP2) is not mutated in hepatoblastoma but shows increased transcriptional activity in a comparative analysis of the three imprinted genes p57(KIP2), IGF2, and H19. Am J Pathol 157:1393–1403
Heerema-McKenney A, Leuschner I, Smith N et al (2005) Nested stromal epithelial tumor of the liver: six cases of a distinctive pediatric neoplasm with frequent calcifications and association with Cushing syndrome. Am J Surg Pathol 29:10–20
Heifetz SA, French M, Correa M et al (1997) Hepatoblastoma: the Indiana experience with preoperative chemotherapy for inoperable tumors; clinicopathological considerations. Pediatr Pathol Lab Med 17:857–874
Hill DA, Swanson PE, Anderson K et al (2005) Desmoplastic nested spindle cell tumor of liver: report of four cases of a proposed new entity. Am J Surg Pathol 29:1–9
Hirohashi S, Ishak KG (2000) Tumours of the liver and intrahepatic bile ducts. In: Hamilton SR, Aaltonen LA (eds) Pathology and genetics of tumours of the digestive system. IARC Press, Lyon
Honda S, Arai Y, Haruta M et al (2008) Loss of imprinting of IGF2 correlates with hypermethylation of teh H19 differentially methylated region in hepatoblastoma. Br J Cancer 99:1891–1899
Http://seer.cancer.gov Surveillance Epidemiology and End Results Web site
Hung W, Blizzard RM, Migeon CJ et al (1963) Precocious puberty in a boy with hepatoma and circulating gonadotropin. J Pediatr 63:895–903
Ingram DJ, Yerushalmi B, Connell J et al (2000) Hepatoblastoma in a neonate: a hypervascular presentation mimicking hemangioendothelioma. Pediatr Radiol 30:794–797
Iolascon A, Giordani L, Moretti A et al (1998) Analysis of CDKN2A, CDKN2B, CDKN2C, and cyclin Ds gene status in hepatoblastoma. Hepatology 27:989–995
Ishak KG, Glunz PR (1967) Hepatoblastoma and hepatocarcinoma in infancy and childhood. Cancer 20:396–422
Ishak KG, Goodman ZD (1999) Hepatocellular carcinoma. In: Ishak KG, Goodman ZD, Stocker JT (eds) Tumors of the liver and intrahepatic bile ducts. Armed Forces Institute of Pathology, Washington
Joshi VV, Kaur P, Ryan B et al (1984) Mucoid anaplastic hepatoblastoma. A case report. Cancer 54:2035–2039
Kakar S, Burgart LJ, Batts KP et al (2005) Clinicopathologic features and survival in fibrolamellar carcinoma: comparison with conventional hepatocellular carcinoma with and without cirrhosis. Mod Pathol 18:1417–1423
Kakar S, Chen X, Ho C et al (2009) Chromosomal changes in fibrolamellar hepatocellular carcinoma detected by array comparative genomic hybridization. Mod Pathol 22:134–141
Kannangai R, Vivekanandan P, Martinez-Murillo F et al (2007) Fibrolamellar carcinomas show overexpression of genes in the RAS, MAPK, PIK3, and xenobiotic degradation pathways. Hum Pathol 38:639–644
Kasai M, Watanabe I (1970) Histologic classification of liver-cell carcinoma in infancy and childhood and its clinical evaluation. Cancer 25:552–563
Katzenstein HM, Krailo MD, Malogolowkin MH et al (2002) Hepatocellular carcinoma in children and adolescents: results from the Pediatric Oncology Group and the Children’s Cancer Group intergroup study. J Clin Oncol 20:2789–2797
Katzenstein HM, Krailo MD, Malogolowkin MH et al (2003) Fibrolamellar hepatocellular carcinoma in children and adolescents. Cancer 97:2006–2012
Kaufman SS, Wood RP, Shaw BW et al (1987) Hepatocarcinoma in a child with the Alagille syndrome. Am J Dis Child 141:698–700
Kim L, Park YN, Kim SE et al (2001) Teratoid hepatoblastoma: multidirectional differentiation of stem cell of the liver. Yonsei Med J 42:431–435
Kingston JE, Draper GJ, Mann JR et al (1982) Hepatoblastoma and polyposis coli. Lancet 1:457
Kumar EV, Kumar L, Pathak IC et al (1978) Clinical, hormonal and ultrastructure studies of a virilizing hepatoblastoma. Acta Paediatr Scand 67:389–392
LaBrecque D (1996) Neoplasia of the liver. Liver and biliary disease. W Wilkens, Baltimore, pp 391–438
Lack EE, Neave C, Vawter GF (1982) Hepatoblastoma. A clinical and pathologic study of 54 cases. Am J Surg Pathol 6:693–705
Lack EE, Neave C, Vawter GF et al (1983) Hepatocellular carcinoma. Review of 32 cases in childhood and adolescence. Cancer 52:1510–1515
Lai CL, Wu PC, Lam KC et al (1979) Histologic prognostic indicators in hepatocellular carcinoma. Cancer 44:1677–1683
Lau WY, Lai PB, Leung MF et al (2000) Differential gene expression of hepatocellular carcinoma using cDNA microarray analysis. Oncol Res 12:59–69
Lee JS, Thorgeirsson SS (2004) Genome-scale profiling of gene expression in hepatocellular carcinoma: classification, survival prediction, and identification of therapeutic targets. Gastroenterology 127(Suppl 1):S51–S55
Lee JS, Thorgeirsson SS (2005) Genetic profiling of human hepatocellular carcinoma. Semin Liver Dis 25:125–132
Lefkowitch JH, Muschel R, Price JB et al (1983) Copper and copper-binding protein in fibrolamellar liver cell carcinoma. Cancer 51:97–100
Li X, Adam G, Cui H et al (1995) Expression, promoter usage and parental imprinting status of insulin-like growth factor II (IGF2) in human hepatoblastoma: uncoupling of IGF2 and H19 imprinting. Oncogene 11:221–229
Li C, Tan YX, Zhou H et al (2005) Proteomic analysis of hepatitis B virus-associated hepatocellular carcinoma: identification of potential tumors markers. Proteomics 5:1125–1139
Libbrecht L, De Vos R, Cassiman D et al (2001) Hepatic progenitor cells in hepatocellular adenomas. Am J Surg Pathol 25:1388–1396
Libbrecht L, Desmet V, Roskams T (2003) Stages of normal and aberrant intrahepatic bile duct development in a mixed hepatoblastoma. Histopathology 42:618–620
Lopez-Terrada D, Gunaratne PH, Adesina AA et al (2009a) Histologic subtypes of hepatoblastoma are characterized by differential canonical Wnt and Notch pathway activation in DLK+ precursors. Hum Pathol 40:783–794
Lopez-Terrada D, Patel K et al (2009b) Characteristic chromosomal copy number changes are associated with subtypes of hepatoblastoma and HCC identified by array-CGH analysis. Society of Pediatric Pathology, Boston
Lowichik A, Zhou H, Pysher TJ et al (2000) Therapy associated changes in childhood tumors. Adv Anat Pathol 7:341–359
Lu M, Greer ML (2007) Hypervascular multifocal hepatoblastoma: dynamic gadolinium-enhanced MRI findings indistinguishable from infantile hemangio-endothelioma. Pediatr Radiol 37:587–591
Luo JH, Ren B, Keryanov S et al (2006) Transcriptomic and genomic analysis of human hepatocellular carcinomas and hepatoblastomas. Hepatology 44:1012–1024
Manes JL, Kissane JM, Valdes AJ et al (1977) Congenital hepatic fibrosis, liver cell carcinoma and adult polycystic kidneys. Cancer 39:2619–2623
Manivel C, Wick MR, Abenoza P et al (1986) Teratoid hepatoblastoma. The nosologic dilemma of solid embryonic neoplasms of childhood. Cancer 57:2168–2174
Matsubara K, Tokino T (1990) Integration of hepatitis B virus DNA and its implications for hepatocarcinogenesis. Mol Biol Med 7:243–260
Meir K, Maly A, Doviner V et al (2009) Nested (ossifying) stromal epithelial tumor of the liver: case report. Pediatr Dev Pathol 12:233–236
Meyers RL (2007) Tumors of the liver in children. Surg Oncol 16:195–203
Milman DH, Grayzel DM (1951) Mixed tumor of the liver; report of a case with a review of the literature. AMA Am J Dis Child 81:408–420
Misick OS (1898) A case of teratoma hepatis. J Pathol Bact 5:128–137
Misugi K, Okajima H, Misugi N et al (1967) Classification of primary malignant tumors of liver in infancy and childhood. Cancer 20:1760–1771
Montesano R (2002) Hepatitis B immunization and hepatocellular carcinoma: the Gambia Hepatitis Intervention Study. J Med Virol 67:444–446
Moore SW, Hesseling PB (1997) Hepatocellular carcinoma in children. Pediatr Surg Int 12:266–270
Moritake H, Taketomi A, Kamimura S et al (2000) Renin-producing hepatoblastoma. J Pediatr Hematol Oncol 22:78–80
Nakagawara A, Ikeda K, Hayashida Y et al (1982) Immunocytochemical identification of human chorionic gonadotropin- and alpha-fetoprotein-producing cells hepatoblastoma associated with precocious puberty. Virchows Arch A Pathol Anat Histopathol 398:45–51
Nakagawara A, Ikeda K, Tsuneyoshi M et al (1985) Hepatoblastoma producing both alpha-fetoprotein and human chorionic gonadotropin. Clinicopathologic analysis of four cases and a review of the literature. Cancer 56:1636–1642
Neimann N, Rauber G, Manciaux N (1963) Les tumeurs malignes du foie chez l’enfant. Rev Int Hepat 13:81–129
Nzeako UC, Goodman ZD, Ishak KG (1995) Comparison of tumor pathology with duration of survival of North American patients with hepatocellular carcinoma. Cancer 76:579–588
Okabe H, Satoh S, Kato T et al (2001) Genome-wide analysis of gene expression in human hepatocellular carcinomas using cDNA microarray: identification of genes involved in viral carcinogenesis and tumor progression. Cancer Res 61:2129–2137
Ortega JA, Douglass EC, Feusner JH et al (2000) Randomized comparison of cisplatin/vincristine/fluorouracil and cisplatin/continuous infusion doxorubicin for treatment of paediatric hepatoblastoma: a report from the Children’s Cancer Group and the Paediatric Oncology Group. J Clin Oncol 18:2665–2675
Pakakasama S, Chen TT, Frawley W et al (2004) CCND1 polymorphism and age on onset of hepatoblastoma. Oncogene 23:4789–4792
Pang SC (1961) Bony and cartilaginous hepatoblastoma. J Pathol Bacteriol 82:273–280
Parham DM, Weeks DA, Beckwith JB (1994) The clinicopathologic spectrum of putative extrarenal rhabdoid tumors. An analysis of 42 cases studied with immunohistochemistry or electron microscopy. Am J Surg Pathol 18:1010–1029
Park WS, Oh RR, Park JY et al (2001) Nuclear localization of beta-catenin is an important prognostic factor in hepatoblastoma. J Pathol 193:483–490
Parker P, Burr I, Slonim A et al (1981) Regression of hepatic adenomas in type Ia glycogen storage disease with dietary therapy. Gastroenterology 81:2129–2137
Pinna AD, Iwatsuki S, Lee RG et al (1997) Treatment of fibrolamellar hepatoma with subtotal hepatectomy or transplantation. Hepatology 26:877–883
Prokurat A, Kluge P, Kosciesza A et al (2002) Transitional liver cell tumors (TLCT) in older children and adolescents: a novel group of aggressive hepatic tumors expressing beta-catenin. Med Pediatr Oncol 39:510–518
Rabinovitz M, Imperial JC, Schade RR et al (1989) Hepatocellular carcinoma in Alagille’s syndrome: a family study. J Pediatr Gastroenterol Nutr 8:26–30
Rainier S, Dobry CJ, Feinberg AP (1995) Loss of imprinting in hepatoblastoma. Cancer Res 55:1836–1838
Ramsay AD, Bates AW, Williams S et al (2008) Variable antigen expression in hepatoblastomas. Appl Immunohistochem Mol Morphol 16:140–147
Robinson HB, Bolande RP (1985) Case 3. Fetal hepatoblastoma with placental metastases. Pediatr Pathol 4:163–167
Rod A, Voicu M, Chiche L et al (2009) Cushing’s syndrome associated with a nested stromal epithelial tumor of the liver: hormonal, immunohistochemical, and molecular studies. Eur J Endocrinol 161:805–810
Ross JA, Radloff GA, Davies SM (2000) H19 and IGF-2 allele-specific expression in hepatoblastoma. Br J Cancer 82:753–756
Rowland JM (2002) Hepatoblastoma: assessment of criteria for histologic classification. Med Pediatr Oncol 39:478–483
Ruck P, Kaiserling E (1993) Melanin-containing hepatoblastoma with endocrine differentiation. An immunohistochemical and ultrastructural study. Cancer 72:361–368
Rugge M, Sonego F, Pollice L et al (1998) Hepatoblastoma: DNA nuclear content, proliferation indices, and pathology. Liver 18:128–133
Russo P, Biegel JA (2009) SMARCB1/INI1 alterations and hepatoblastoma: another extrarenal rhabdoid tumor revealed? Pediatr Blood Cancer 52:312–313
Saul SH, Titelbaum DS, Gansler TS et al (1987) The fibrolamellar variant of hepatocellular carcinoma. Its association with focal nodular hyperplasia. Cancer 60:3049–3055
Saxena R, Leake JL, Shafford EA et al (1993) Chemotherapy effects on hepatoblastoma. A histological study. Am J Surg Pathol 17:1266–1271
Scheimberg I, Cullinane C, Kelsey A et al (1996) Primary hepatic malignant tumor with rhabdoid features. A histological, immunocytochemical, and electron microscopic study of four cases and a review of the literature. Am J Surg Pathol 20:1394–1400
Schlecht I, Riebel T, Henze G (1996) Unusually intensively ossified hepatoblastoma (teratoid type) in a 1-year-old girl (in German). Rofo 165:201–203
Shang XZ, Zhu H, Lin K et al (2004) Stabilized beta-catenin promotes hepatocyte proliferation and inhibits TNFalpha-induced apoptosis. Lab Invest 84:332–341
Shirota Y, Kaneko S, Honda M et al (2001) Identification of differentially expressed genes in hepatocellular carcinoma with cDNA microarrays. Hepatology 33:832–840
Siciliano M, De Candia E, Ballarin S et al (2000) Hepatocellular carcinoma complicating liver cirrhosis in type IIIa glycogen storage disease. J Clin Gastroenterol 31:80–82
Simson IW (1982) Membranous obstruction of the inferior vena cava and hepatocellular carcinoma in South Africa. Gastroenterology 82:171–178
Sinniah D, Campbell PE, Colebatch JH (1974) Primary hepatic cancer in childhood. Arch Dis Child 49:496
Soreide O, Czerniak A, Bradpiece H et al (1986) Characterstics of fibrolamellar hepatocellular carcinoma. A study of nine cases and a review of the literature. Am J Surg 151:518–523
Stocker JT (1994) Hepatoblastoma. Semin Diagn Pathol 11:136–143
Stocker JT (2001) Hepatic tumors in children. Clin Liver Dis 5:259–281
Taat F, Bosman DK, Aaronson DC (2004) Hepatoblastoma in a girl with biliary atrasia: coincidence or co-incidence. Pediatr Blood Cancer 43:603–605
Takayasu H, Horie H, Hiyama E et al (2001) Frequent deletions and mutations of the beta-catenin gene are associated with overexpression of cyclin D1 and fibronectin and poorly differentiated histology in childhood hepatoblastoma. Clin Cancer Res 7:901–908
Thomas D, Pritchard J, Davidson R et al (2003) Familia hepatoblastoma and APC gene mutations: renewed call for molecular research. Eur J Cancer 39:2200–2204
Thorgeirsson SS, Grisham JW (2002) Molecular pathogenesis of human hepatocellular carcinoma. Nat Genet 31:339–346
Tomizawa M, Saisho H (2006) Signaling pathway of insulin-like growth factor-II as a target of molecular therapy for hepatoblastoma. Wordl J Gastroenterol 12:6531–6535
Tornillo L, Carafa V, Sauter G et al (2002) Chromosomal alterations in hepatocellular nodules by comparative genomic hybridization: high-grade dysplasic nodules represent early stages of hepatocellular carcinoma. Lab Invest 82:547–553
Touraine RL, Bertrand Y, Foray P et al (1993) Hepatic tumours during androgen therapy in Fanconi anemia. Eur J Pediatr 152:691–693
Trobaugh-Lotrario AD, Tomlinson GE, Finegold MJ et al (2009) Small cell undifferentiated variant of hepatoblastoma: adverse clinical and molecular features similar to rhabdoid tumors. Pediatr Blood Cancer 52:328–334
Tsai HW, Tsai HH, Kuo FY et al (2009) Computerized analyses of morphology and proliferative activity differentiate hepatoblastoma from paediatric hepatocellular carcinoma. Histopathology 54:328–336
Ucar C, Caliskan U, Troy H et al (2005) Hepatoblastoma in a child with neurofibromatosis type 1. Pediatr Blood Cancer 49:357–359
Van Dyck F, Declercq J, Braem CV et al (2007) PLAG1, the prototype of the PLAG gene family: versatility in tumour development. Int J Oncol 30:765–774
Van Eyken P, Sciot R, Callea F et al (1990) A cytokeratin-immunohistochemical study of hepatoblastoma. Hum Pathol 21:302–308
Varma V, Cohen C (2004) Immunohistochemical and molecular markers in the diagnosis of hepatocellular carcinoma. Adv Anat Pathol 11:239–249
Vileisis RA, Sorensen K, Gonzalez-Crussi F et al (1982) Liver malignancy after parenteral nutrition. J Pediatr 100:88–90
Vivekanandan P, Thomas D, Torbenson M et al (2009) Methylation regulates hepatitis B viral protein expression. J Infect Dis 199:1286–1291
Von Schweinitz D, Wischmeyer P, Leuschner I et al (1994) Clinico-pathological criteria with prognostic relevance in hepatoblastoma. Eur J Cancer 30A:1052–1058
Wagner LM, Garrett JK, Ballard ET et al (2007) Malignant rhabdoid tumor mimicking hepatoblastoma: a case report and literature review. Pediatr Dev Pathol 10:409–415
Wang J, Chenivesse X, Henglein B et al (1990) Hepatitis B virus integration in a cyclin A gene in a hepatocellular carcinoma. Nature 343:555–557
Watanabe I, Kasai M, Watanuki T et al (1975) Malignant mixed tumor of the liver: report of a case and a review of the literature. Acta Hepatogastroenterol (Stuttg) 22:158–164
Watanabe I, Yamaguchi M, Kasai M (1987) Histologic characteristics of gonadotropin-producing hepatoblastoma: a survey of seven cases from Japan. J Pediatr Surg 22:406–411
Wei Y, Fabre M, Branchereau S et al (2000) Activation of beta-catenin in epithelial and mesenchymal hepatoblastomas. Oncogene 19:498–504
Weinberg AG, Mize CE, Worthen HG et al (1976) The occurrence of hepatoma in the chronic form of hereditary tyrosinemia. J Pediatr 88:434–438
Weinberg AG, Finegold MJ (1983) Primary hepatic tumors of childhood. Hum Pathol 14:512–537
Weinstein S, Scottolini AG, Loo SY et al (1985) Ataxia telangiectasia with hepatocellular carcinoma in a 15-year-old girl and studies of her kindred. Arch Pathol Lab Med 109:1000–1004
Wilkens L, Bredt M, Flemming P et al (2000) Cytogenetic aberrations in primary and recurrent fibrolamellar hepatocellular carcinoma detected by comparative genomic hybridization. Am J Clin Pathol 114:867–874
Willis RA (1962) The Pathology of tumours of children. Charles C. Thomas, Springfield
Wong N, Lai P, Pang E et al (2000) A comprehensive karyotypic study on human hepatocellular carcinoma by spectral karyotyping. Hepatology 32:1060–1068
Wu PC, Fang JW, Lau VK et al (1996) Classification of hepatocellular carcinoma according to hepatocellular and biliary differentiation markers. Clinical and biological implications. Am J Pathol 149:1167–1175
Yamada S, Ohira M, Horie H et al (2004) Expression profiling and differential screening between hepatoblastomas and the corresponding normal livers: identification of high expression of the PLK1 oncogene as poor-prognosis indicator of hepatoblastomas. Oncogene 23:5901–5911
Yamaoka H, Ohtsu K, Sueda T et al (2006) Diagnostic and prognostic impact of beta-catenin alterations in pediatric liver tumors. Oncol Rep 15:551–556
Yamauchi N, Watanabe A, Hishinuma M et al (2005) The glypican 3 oncofetal protein is a promising diagnostic marker for hepatocellular carcinoma. Mod Pathol 18:1591–1598
Yang J, Zhou F, Xu T et al (2008) Analysis of sequence variations in 59 microRNAs in hepatocellular carcinoma. Mutat Res 638:205–209
Yun K, Jinno Y, Sohda T et al (1998) Promoter-specific insulin-like growth factor 2 gene imprinting in human fetal liver and hepatoblastoma. J Pathol 185:91–98
Zatkova A, Rouillard JM, Hartmann W et al (2004) Amplification and overexpression of the IGF2 regulator PLAG1 in hepatoblastoma. Genes Chromosomes Cancer 39:126–137
Zerbini MC, Sredni ST, Grier H et al (1998) Primary malignant epithelial tumors of the liver in children: a study of DNA content and oncogene expression. Pediatr Dev Pathol 1:270–280
Zimmermann A (2000) Tumors of the liver – pathologic aspects. In: Blumgart LH (ed) Surgery of the liver, biliary tract and pancreas, vol II, 4th edn. Saunders Elsevier, Philadelphia, pp 1085–1130, Section 13, Chapter 69
Zimmermann A (2002) Hepatoblastoma with cholangioblastic features (‘cholangioblastic hepatoblastoma’) and other liver tumors with bimodal differentiation in young patients. Med Pediatr Oncol 39:487–491
Zimmermann A (2005) The emerging family of hepatoblastoma tumours: from ontogenesis to oncogenesis. Eur J Cancer 41:1503–1514
Zimmermann A (2006) Malignant epithelial liver tumors in children, adolescents and adults: molecular pathways of cancerogenesis, growth, and spread. In: Lee F (ed) Liver cancer: new research. Nova Biomedical Books, New York, pp 1–41, Chapter 1
Zucman-Rossi J, Jeannot E, Nhieu JT et al (2006) Genotype-phenotype correlation in hepatocellular adenoma: new classification and relationship with HCC. Hepatology 43:515–524
Zynger DL, Gupta A, Luan C et al (2008) Expression of glypican 3 in hepatoblastoma: an immunohistochemical study of 65 cases. Hum Pathol 39:224–230
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer Berlin Heidelberg
About this chapter
Cite this chapter
Zimmermann, A., Lopez-Terrada, D. (2011). Pathology of Pediatric Liver Tumors. In: Zimmermann, A., Perilongo, G. (eds) Pediatric Liver Tumors. Pediatric Oncology. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-14504-9_9
Download citation
DOI: https://doi.org/10.1007/978-3-642-14504-9_9
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-14503-2
Online ISBN: 978-3-642-14504-9
eBook Packages: MedicineMedicine (R0)